calcium distribution in islets of langerhans: a study of calcium
TRANSCRIPT

J. Cell Sci. 19, 395-409 (i97S) 395Printed in Great Britain
CALCIUM DISTRIBUTION IN ISLETS OF
LANGERHANS: A STUDY OF CALCIUM
CONCENTRATIONS AND OF CALCIUM
ACCUMULATION IN B CELL ORGANELLES
S. L. HOWELL, W. MONTAGUE AND MARGARET TYHURSTBiochemistry Laboratory, School of Biological Sciences, University of Sussex,Fainter, Brighton, Sussex, England
SUMMARY
Calcium concentrations of various pancreatic B cell organelles have been determined byX-ray microanalysis of areas of frozen sections of unfixed rat islets of Langerhans. Highestconcentrations were detected in storage granules and in mitochondria, although calcium wasalso present in nuclei, in areas of endoplasmic reticulum and of cytoplasm.
Accumulation of 45Ca by isolated organelles has been studied in homogenates and isolatedsubcellular fractions of rat islets of Langerhans. In the presence of a permeant anion (oxalateor phosphate), accumulation of 4tCa into mitochondria and microsomes was strongly stimulatedby ATP. This net uptake was diminished during incubation of homogenates or of a mito-chondria-plus storage granule-rich fraction in the presence of cyclic AMP, dibutyryl cyclicGMP, 2:4-dinitrophenol or of ruthenium red. Investigations of the characteristics of 45Caaccumulation by homogenates prepared from storage granule-depleted islets showed nodifferences from those of normal islets, suggesting that the granules do not represent an im-portant labile pool of calcium. With the exception of cyclic AMP and cyclic GMP none of theinsulin secretagogues tested (glucose, leucine, arginine, adrenalin, noradrenalin, theophylline,glibenclamide) altered calcium accumulation by islet homogenates.
On the basis of absolute calcium levels and of 46Ca uptake studies it is concluded that islet Bcells contain a readily exchangeable mitochondrial calcium pool, and an endoplasmic reticulumpool containing a lower concentration of calcium which is also readily exchangeable. Thestorage granules, despite their high calcium content, do not appear to constitute a labile pool.It seems likely that the labile mitochondria and endoplasmic reticulum pools play a predominantrole in the regulation of cytoplasmic free calcium levels, which may in turn be important in theregulation of rates of insulin secretion.
INTRODUCTION
There is evidence to suggest that calcium may be the link between stimulation andsecretion in a variety of endocrine tissues including the pancreatic B cell, and thatintracellular calcium concentrations may play an important role in the regulation ofrates of insulin secretion (Malaisse, 1973). Extensive studies have been made of theinflux and efflux of 45Ca in isolated islets of Langerhans during stimulation of insulinsecretion in response to a variety of agents, as reviewed by Malaisse (1973). However,there have so far been few investigations of the localization of calcium within B cellorganelles or of factors which may alter its distribution. The problems involved,both in preparing subcellular fractions which contain the various B cell organelles in

396 S. L. Howell, W. Montague and M. Tyhurst
purified form and in avoiding diffusion of calcium out of and between fractions duringthe separation procedure, make it very difficult to determine the exact subcellulardistribution either of labelled calcium or of total cellular calcium amongst thevarious organelles.
We have attempted to resolve some of these problems by determining the distribu-tion of total calcium in frozen sections of unfixed islets by X-ray microanalysis. Inaddition this paper presents results of a study of the accumulation of ^Ca by parti-culate components of homogenates and subcellular fractions of rat islets of Langer-hans, and of its regulation by nucleotides, insulin secretagogues and other agents.A brief report of some aspects of this work has already appeared (Howell & Montague,1975)-
MATERIALS AND METHODS
Reagents46CaCl2 of specific radioactivity 28 /tCi//tg, 3i//tg Ca/ml, was obtained from the Radio-
chemical Centre, Amersham, Bucks. ATP, cyclic AMP, dibutyryl cyclic AMP, cyclic GMP,and dibutyryl cyclic GMP, were obtained from Boehringer Corp., Uxbridge, Middlesex, U.K.ADP, AMP, adenosine, ruthenium red and dinitrophenol were obtained from Sigma Chemicals,Kingston-upon-Thames, U.K. Other reagents were of 'Analar' grade or of the purest gradewhich was commercially available. Millipore membranes were obtained from the MilliporeCorp., London, N.W.10, U.K.
Methods
Tissue preparation. Islets of Langerhans were isolated by collagenase digestion of pancreatictissue taken from female rats (200-240 g) (Howell & Taylor, 1966), using a bicarbonate-buffered medium (Gey & Gey, 1936) containing 5-5 nun glucose and 2 mM CaCl2- Finalrinsing and separation of the islets was performed in medium from which calcium was omitted.The islets were collected in a Kontes all-glass homogenizer, the medium was removed and theislets were homogenized in 0-25 M sucrose buffered with 10 mM Tris-HCl (pH 72). In someexperiments subcellular fractions were obtained from the homogenate by centrifugation at4 °C as follows: 600 g for 5 min to give a pellet of nuclei + debris; the supernatant was centri-fuged at 24000 g for 10 min to give a pellet of mitochondria + storage granules and this super-natant was in turn centrifuged at 105000 g for 60 min to give a microsomal pellet and finalsupernatant (Howell, Fink & Lacy, 1969). All pellets were resuspended in 0-25 M sucrose—10 mM Tris-HCl (pH 72) and assayed simultaneously at the end of the centrifugation.
Assay of 16Ca accumulation. Accumulation of 45Ca by homogenates or isolated subcellularfractions was estimated by incubation of 5o-/tl aliquots of homogenate or subcellular fractionsresuspended in sucrose (025 M), with 50 /tl of a solution which was prepared to give finalconcentrations of 0125 M sucrose, 35 mM KC1, 10 mM Tris-HCl, 4 mM MgCl2, 4 mM KjHPO4,10 mM sodium succinate, 05 mM 3-isobutyl-i-methylxanthine, 20 /IM CaCla and 4tCaCla(5 /tCi/ml) pH 72. Nucleotides and other agents were added at the concentrations indicated inindividual experiments.
After 20 min incubation at 23 °C the particulate-bound calcium was separated from thecalcium by filtration through Millipore membranes (450 nm, HAWP or 100 nm, VCWP),each membrane being rinsed 3 times with 025 M sucrose in 10 mM Tris-HCl (pH 7-2) toremove unbound radioactivity. Radioactivity remaining on the filters was determined in aliquid scintillation spectrometer (Beckman LS 233) using Instagel (Packard Instruments) asscintillant.
Cryo-ultramicrotomy technique. Islets were incubated at 37 °C until immediately beforeimmersion in liquid nitrogen. The procedures for freezing of individual islets and for sectioningwere as described previously (Howell & Tyhurst, 1974), except that no trough liquid was used;

Calcium distribution in B cell organelles 397
sections were transferred directly from the dry knife surface to (Formvar + carbon)-coatednickel grids located on the knife holder or on the knife itself, by means of an eyelash probe.Both specimen and knife temperatures were maintained at —80 °C. After collection of sectionsthe grids were transferred to a Petri dish in the bottom of the Cryokit chamber and allowed toreach room temperature over a period of 3 h. Dririte was placed in the chamber in order tomaintain a dry atmosphere. The sections were stored in a dry atmosphere at room temperaturepiior to analysis.
X-ray microanalysis. Sections were analysed in an AEI EMMA-4 analytical electron micro-scope using crystal spectrometers. The minimum probe size used was 02 /im and the micro-scope was operated at 60 kV. Count rates were assessed for 100 s at the peak for calcium andthen for 100 s with the settings displaced to one side of the peak. A white count was recordedto give an assessment of the continuum, radiation in the area analysed and to allow for correctionfor variable thickness of the specimens, and the count rate recorded was assessed according tothe formula c = (P — b)/(W — Wb) where P is the peak counts obtained, b the counts obtainedfrom the same area with the detector offset from the peak, W is the white count in the samearea and Wb is the white count obtained from an area away from the specimens, in this casethe Formvar film. In order to obtain a valid sample, sections were prepared from severaldifferent islets on separate occasions.
Protein determination. Protein concentrations were determined by the method of Lowry,Rosebrough, Farr & Randall (1951) using crystalline bovine albumin as standard. Calciumaccumulation was expressed as nmol Ca accumulated per mg protein per 20-min incubationperiod; mean values ± S.E.M. are shown throughout.
Electron microscopy. After incubation in the conditions described above, subcellular fractionswere repelleted by centrifugation before fixation with 3 % glutaraldehyde, postfixation withOsO4, dehydration and embedding by standard procedures. Sections were examined in anAEI EM 6B electron microscope.
RESULTS
Calcium distribution in B cells determined by X-ray microanalysis
The following organelles, which are identifiable in frozen sections of unfixed islets(Howell & Tyhurst, 1974), and are of a size (0-2 /im or greater) which can be locatedwith the probe diameter available on the EMMA-4 instrument, were analysed for
Table 1. Mass ratio of calcium in various B cell organelles determined byX-ray microanalysis
Organelle
MitochondriaStorage granulesNucleiRough-surfaced endoplasmic reticulumOther cytoplasmic areas
ioJ x Massratio
57 ± 1164 ± 1435 ± 741 ± 83° ± 8
No. ofobservations
1816
1557
Results are given as means + S.E.M. of the number of observations shown.
their calcium content: nuclei, mitochondria, storage granules, areas of rough-surfacedendoplasmic reticulum, and areas of cytoplasmic ground substrate. The results shownin Table 1 are presented as the mass ratio of calcium found in each organelle; thismethod of expression includes a factor (the white count) which corrects for possibledifferences in mass (including section thickness) between the various areas which are
398 »S. L. Howell, W. Montague and M. Tyhurst
analysed. All of the areas examined contain detectable concentrations of calcium, thevalues falling into 2 groups: those areas of relatively low concentration (mass ratio30-41) which include the nuclei, endoplasmic reticulum and areas of cytoplasmicground substance, and a second group of significantly (P < 0-05) higher concentra-tion (mass ratio 57—64) which comprise the mitochondria and storage granules. Themean concentration of calcium in the granules appears to be greater than that in themitochondria although the differences were not statistically significant; similarly,differences in mass ratio between nuclei, rough-surfaced endoplasmic reticulum andcytoplasmic ground substance were not significant.
accumulation by homogenates and subcellular fractionsSome of the characteristics of calcium accumulation by particulate components
of islet homogenates have been described in an earlier communication (Howell &Montague, 1975). Briefly, optimal binding occurred at an initial pH of the incubation
20 r
wO1-Q.
00
•5
10
u
Oxalate
Phosphate
2 5 10
[Anion], mM
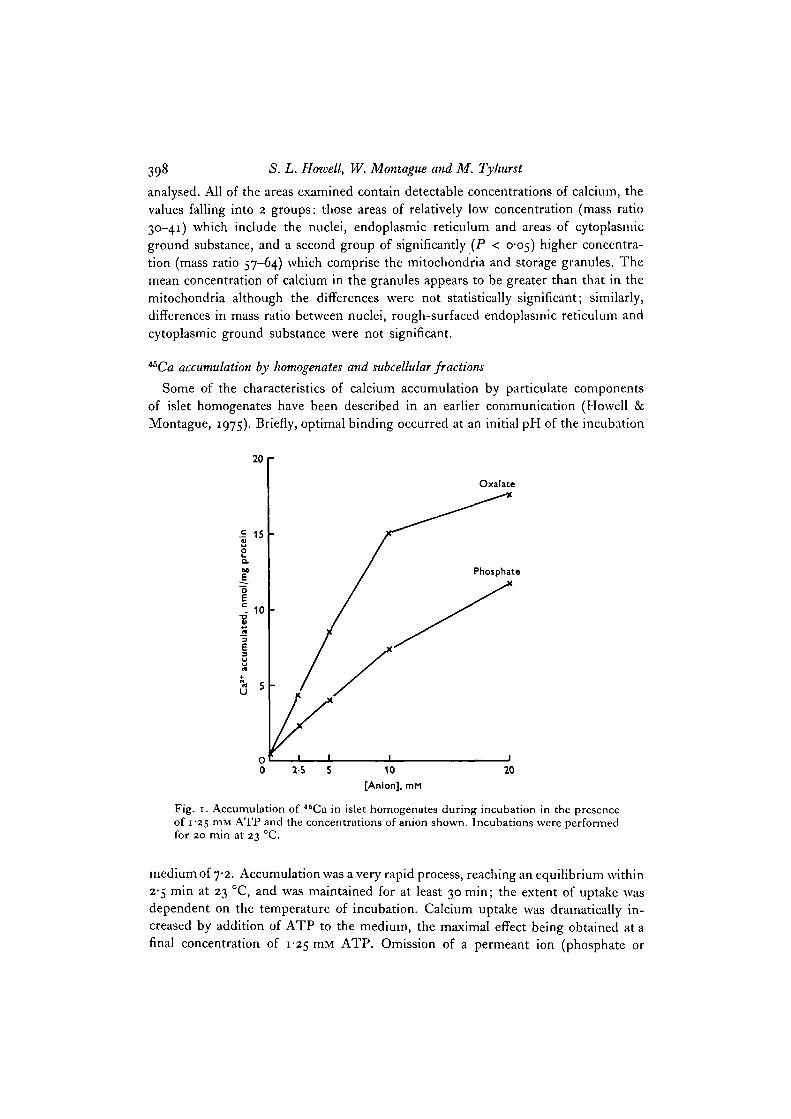
Fig. 1. Accumulation of 46Ca in islet homogenates during incubation in the presenceof 1-25 mM ATP and the concentrations of anion shown. Incubations were performedfor 20 min at 23 °C.
medium of 7-2. Accumulation was a very rapid process, reaching an equilibrium within2-5 min at 23 °C, and was maintained for at least 30 min; the extent of uptake wasdependent on the temperature of incubation. Calcium uptake was dramatically in-creased by addition of ATP to the medium, the maximal effect being obtained at afinal concentration of 1-25 mM ATP. Omission of a permeant ion (phosphate or
Calcium distribution in B cell organelles
oxalate) reduced the binding by homogenates to very low levels, either in the absenceor presence of ATP; phosphate or oxalate were satisfactory but oxalate was moreeffective in this respect than was phosphate (Fig. 1). Heating of the homogenate to80 °C for 3 min before incubation completely abolished the calcium accumulation.
5-0 r
.£ 40
30
i:§ 20o
i c 10
r+i
Homogenate Nuclei Mitochondria Microtomes Supernatant+granulej
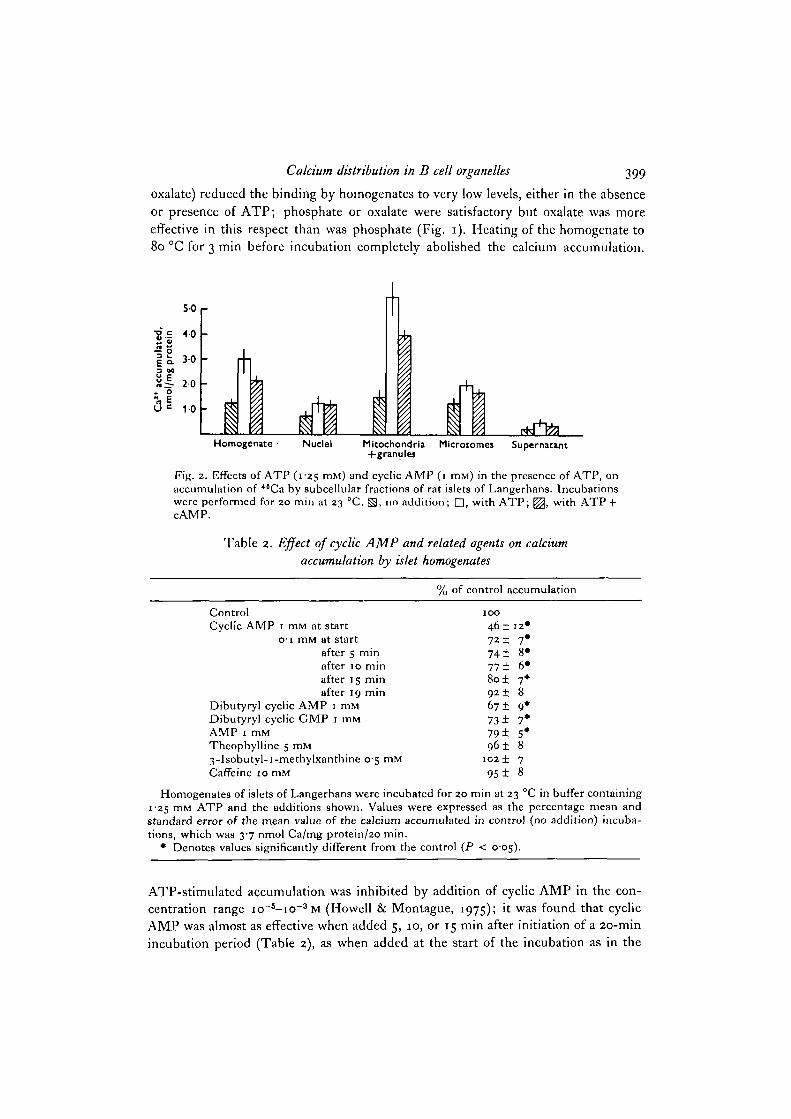
Fig. 2. Effects of ATP (125 ITIM) and cyclic AMP (1 mM) in the presence of ATP, onaccumulation of 48Ca by subcellular fractions of rat islets of Langerhans. Incubationswere performed for 20 min at 23 °C. ^ , no addition; Q, with ATP; ^ , with ATP +cAMP.
Table 2. Effect of cyclic AMP and related agents on calciumaccumulation by islet homogenates
% of control accumulation
ControlCyclic AMP i mM at start
o-i mM at startafter $ minafter io minafter 15 minafter 19 min
Dibutyryl cyclic AMP 1 mMDibutyryl cyclic GMP 1 mMAMP 1 mMTheophylline 5 mM3-Isobutyl-i-methylxanthine 0-5 mMCaffeine 10 mM
1 0 0
46 ± 12*72 ± 7*74 ± 8*77 ± 6*80 ± 7*92 ± 867 ± 9*73 ± 7*79 ± 5*96 ± 8
102 ± 795 ± 8
Homogenates of islets of Langerhans were incubated for 20 min at 23 CC in buffer containing1 25 mM ATP and the additions shown. Values were expressed as the percentage mean andstandard error of the mean value of the calcium accumulated in control (no addition) incuba-tions, which was 37 nmol Ca/mg protein/20 min.
• Denotes values significantly different from the control (P < 005).
ATP-stimulated accumulation was inhibited by addition of cyclic AMP in the con-centration range IO~6-IO~3M (Howell & Montague, 1975); it was found that cyclicAMP was almost as effective when added 5, 10, or 15 min after initiation of a 20-minincubation period (Table 2), as when added at the start of the incubation as in the

400 S. L. Howell, W. Montague and M. Tyhurst
previous experiments. Comparison of the abilities of impure subcellular fractions(nuclei + debris, mitochondria + granule or microsome-rich fraction, prepared bydifferential centrifugation) to accumulate Ca showed that the effect of cyclic AMP wasexerted principally on the fraction containing mitochondria + storage granules, therebeing much smaller effects on nuclear and microsomal fractions (Fig. 2).
Table 3. Effect of inhibitors on calcium accumulation by islet homogenates
% of control accumulation
Control (i-25 mM ATP)Ouabain o-i mMFluoride 10 mMRuthenium red 10 /IM
o-i mMOligomycin 2 fig/fd2:4-dinitrophenol 0-2 mM
1 0 0
95 ±135 ±60 ±49 ±71 ±57 ±
68#
1 0 *
' 4 *8*7#
Homogenates of islets of Langerhans were incubated for 20 min at 23 °C in buffer containing1-25 mM ATP and the additions shown. Values were expressed as the percentage mean andstandard error of the mean value of the calcium accumulated in control (no addition) incuba-tions, which was 3-3 nmol Ca/mg protein/20 min.
• Denotes values significantly different from the control (P < 0-05).
Other nucleotides were also examined for their effectiveness in diminishing calciumaccumulation. Dibutyryl cyclic AMP and dibutyryl cyclic GMP were less effectivethen cyclic AMP at the same concentration (1 mM). Similarly AMP was partiallyeffective when used at high concentrations (Table 2). The cyclic nucleotide phos-phodiesterase inhibitors, 3-isobutyl-i-methylxanthine, theophylline and caffeinedid not alter calcium accumulation when added to homogenates, and 3-isobutyl-i-methylxanthine was included routinely in the experiments utilizing cyclic nucleotides.
ATP-dependent uptake of calcium in homogenates was inhibited 45 % by o-1 mMdinitrophenol, an uncoupler of oxidative phosphorylation, or by the respiratory in-hibitor oligomycin (2/(g/ml). Accumulation was also inhibited by ruthenium redwhich is reported (Moore, 1971) to be a specific inhibitor of mitochondrial calciumuptake (Table 3). In subcellular fractionation experiments these agents were foundto be effective principally on the mitochondria + storage granule fraction (results notshown). Sodium fluoride, an inhibitor of ATPase activity, stimulated calcium uptakeby homogenates in similar conditions, while ouabain (an inhibitor of Na-K ATPase)had no significant effect on calcium accumulation.
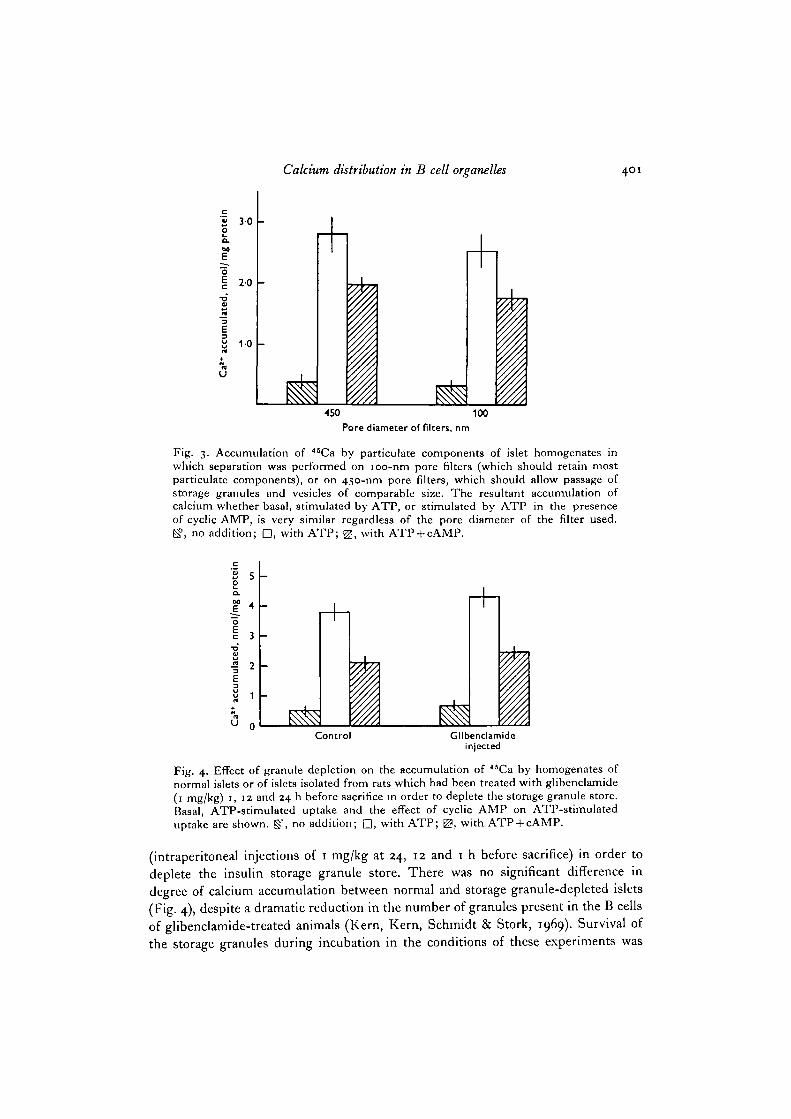
The role of the insulin storage granules in promoting calcium uptake was studiedin two ways. First, filters were used of pore size which would either permit thepassage of the granules (450 nm pore size) or retain them (100 nm pore size). In theseexperiments there was no significant difference in the basal or ATP-stimulated uptakeof calcium or in the effectiveness of cyclic AMP in inhibiting calcium uptake, by theorganelles retained by the two types of filter (Fig. 3). Secondly, calcium accumulationin homogenates of normal islets on 100-nm filters was compared with that in homo-genates of islets from rats which had previously been treated with glibenclamide
Calcium distribution in B cell organelles 4 0 1
S 30o
00
•5
20
"S
u
10
•450 100
Pore diameter of filters, nm
Fig. 3. Accumulation of 45Ca by paniculate components of islet homogenates inwhich separation was performed on 100-nm pore filters (which should retain mostparticulate components), or on 450-nm pore filters, which should allow passage ofstorage granules and vesicles of comparable size. The resultant accumulation ofcalcium whether basal, stimulated by ATP, or stimulated by ATP in the presenceof cyclic AMP, is very similar regardless of the pore diameter of the filter used.E?, no addition; D, with ATP; £?, with ATP + cAMP.
I 5Q.
I 4oI 3
UControl Gllbenclamlde
injected
Fig. 4. Effect of granule depletion on the accumulation of 46Ca by homogenates ofnormal islets or of islets isolated from rats which had been treated with glibenclamide(1 mg/kg) 1, 12 and 24 h before sacrifice in order to deplete the storage granule store.Basal, ATP-stimulated uptake and the effect of cyclic AMP on ATP-stimulateduptake are shown, g?, no addition; Q, with ATP; g£, with ATP + cAMP.
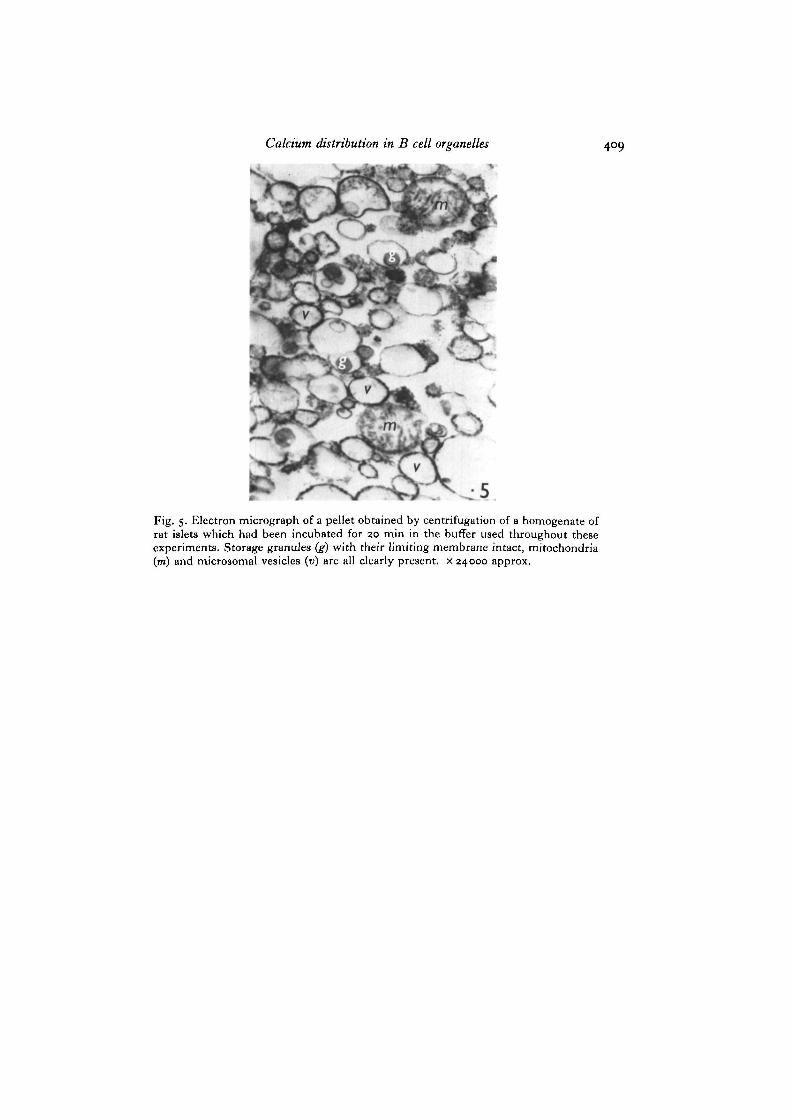
(intraperitoneal injections of 1 mg/kg at 24, 12 and 1 h before sacrifice) in order todeplete the insulin storage granule store. There was no significant difference indegree of calcium accumulation between normal and storage granule-depleted islets(Fig. 4), despite a dramatic reduction in the number of granules present in the B cellsof glibenclamide-treated animals (Kern, Kern, Schmidt & Stork, 1969). Survival ofthe storage granules during incubation in the conditions of these experiments was

402 S. L. Howell, W. Montague and M. Tyhurst
confirmed by electron-microscopic examination of the pellets at the end of theincubation period (Fig. 5).
The effects of various insulin secretagogues on calcium accumulation by islethomogenates were studied in a further series of experiments (Table 4). Glucose(20 mM) had a small but consistent effect in diminishing calcium accumulation byislet homogenates, but an exactly similar effect was shown by galactose at the sameconcentration. Leucine and arginine, both stimulators of insulin secretion, had no
Table 4. Effect of insulin secretagogues on ibCa accumulationby homogenates of rat islets of Langerhans
% of control accumulation
Control (1-25 mM ATP only) 100Glucose 20 mM 92 ± 5Galactose 20 mM 89 ± 7Glibenclamide 1 /tg//«l 102 ±5
10 /<g//*l 93 i 4100 /*g//fl 81 ± 7
Adrenalin 01 mM 96 ±8Noradrenalin 01 min 93 ±6L-Leucine 5 mM 105 ± 4L-Arginine HC1 5 mM 97 ± 6
Homogenates of islets of Langerhans were incubated for 20 min at 23 °C in buffer containing1-25 mM ATP and the additions shown. Values were expressed as the percentage mean andstandard error of the mean value of the calcium accumulated in control incubations which was36 nmol Ca/mg protein/20 min.
effect on uptake, while the inhibitors of secretion adrenalin and noradrenalin weresimilarly ineffective. Glibenclamide diminished calcium accumulation when addedat concentrations of 10 or 100/tg/ml, but was without significant effect at o-i ori-o/tg/ml (Table 4).
DISCUSSION
Ca distribution revealed by X-ray microanalysis
The localization of ions within individual organelles is technically very difficultusing procedures which have hitherto been available. Thus, subcellular fractionationmethods allow scope for loss of material from individual sites by diffusion during thefractionation, as well as the possibility of cross-contamination between fractions andof spurious binding of ions to isolated organelles in broken cell preparations. Similarly,pyroantimonate precipitation procedures, originally devised for the localization ofsodium, have been used for the location of calcium since analysis of the electron-opaque precipitates has shown that they contain at least a proportion of calcium salts.However, it is only possible to infer calcium distribution with certainty by directanalysis of each deposit, since the relative proportions of sodium to calcium or other ionsmay vary considerably within apparently identical electron-opaque precipitates. Thistype of analysis of pyroantimonate precipitates was adopted successfully by Herman,

Calcium distribution in B cell organelles 403
Sato & Hales (3973), who showed that calcium-containing precipitates were depositedpredominantly on B granules and their membranes, on mitochondria, and on the cellmembrane. Incubation of the islets with high concentrations of glucose in order tostimulate insulin secretion increased the intensity of precipitate (presumably con-taining calcium) which was associated with storage granule membranes and withthe plasma membrane. In a similar study utilizing mouse islets and a pyroantimonateprocedure Schafer & Kloppel (3974) showed increased electron-opaque deposits inendoplasmic reticulum and mitochondria in resting (unstimulated) B cells, whileincreased precipitation was present on the plasma membrane and storage granulesacs in B cells which were stimulated to secrete insulin by hyperglycaemia. It was notpossible to compare the calcium content of the precipitates at any site in differentphases of secretory activity in order to confirm that the increased precipitation wasdue solely to calcium.
Direct estimation of calcium concentrations in frozen sections of unfixed tissueavoids these uncertainties and minimizes the chances of diffusion of ions by trans-ferring the tissue directly from a metabolically viable state during incubation invitro at 37 °C in bicarbonate-buffered salt solution to the temperature of liquidnitrogen within a few seconds (Howell & Tyhurst, 1974). Problems of cooling of thetissue are minimized by the very small size (diameter ~ 0-2 mm) of the islets whichprovide the starting material. Procedures of this type seem to provide the only possi-bility of obtaining accurate information about the calcium concentration in differentorganelles of individual cells (Appleton, 1974).
Regulation of calcium accumulation
It seemed likely from the data obtained by electron-microscope X-ray microanalysisthat several B cell organelles, including mitochondria, granules and endoplasmicreticulum, may play important roles in the storage of calcium in islets. However, ofall the subcellular components involved, only the storage granules can be separatedin a characterized and purified fraction (Howell et al. 3969). The major part of thismetabolic study was therefore performed using islet homogenates in which all theimportant subcellular components and the possibility of interactions between twoor more of them are retained; only in the later stages have attempts been made toidentify and separate the specific organelles which may be involved in mediating thevarious effects which were observed.
The system used in this study of calcium uptake by particulate components of islethomogenates involved the rapid separation of particulate-bound from free ^Ca onMillipore membranes, a method which has been successfully used in studies of cal-cium accumulation by mitochondria (Carafoli, Gamble & Lehninger, 1966), plasmamembranes from muscle (Sulakhe, Drummond & Ng, 3973) or liver (Schlatz &Marinetti, 3972), and sarcoplasmic reticulum fragments (McCollum, Besch, Entman& Schwartz, 1972). The properties of the system described under Results are ingeneral consistent with the known characteristics of calcium accumulation by mito-chondria or endoplasmic reticulum in various mammalian tissues.
Dinitrophenol (0-2 ITIM) would be expected to uncouple oxidative phosphorylation

404 S. L. Howell, W. Montague and M. Tyhurst
in islet homogenates and as a result to reduce mitochondrial calcium uptake. DNPproduced a reduction in net calcium accumulation of 43 % in these conditions, sug-gesting that this proportion of the total uptake by the homogenate was due to ATP-dependent mitochondrial calcium uptake. Similarly ruthenium red, reportedly aspecific inhibitor of mitochondrial calcium uptake (Moore, 1971), inhibited the ac-cumulation of isotope by over 60% when present at a concentration of icr4 M.Together these results indicate that 60 % of the total calcium accumulation by islethomogenates in the presence of phosphate is attributable to uptake by mitochondriaand that two thirds of this is ATP-dependent. The observed 8-fold stimulation of up-take in homogenates by ATP (Howell & Montague, 1975) suggests that one eighth ofthe maximal uptake in the presence of ATP may be a result of ATP-independentaccumulation (possibly by endoplasmic reticulum fragments or by mitochondria);the remaining 25-30 % of the ATP-stimulated uptake which is not accounted for isassumed to be due to ATP-driven accumulation by endoplasmic reticulum. The stimu-latory effect of fluoride is suggested to result from inhibition of phosphatase activity,with a resultant maintenance of the ATP level in the homogenate; by contrast, ouabainan inhibitor of Na+K+-dependent ATPase, was without significant effect on calciumaccumulation. Similarly 10 mM caffeine, which has been shown to cause releaseof calcium from sarcoplasmic reticulum through an effect which is not related tophosphodiesterase inhibition (Weber & Herz, 1968) and has been suggested to alterB cell calcium distribution in a similar way (Hedeskov & Capito, 1975), was withouteffect on calcium accumulation by islet homogenates (Table 1).
The finding of an effect of cyclic AMP on accumulation of ^Ca by islet-cell homo-genates is of some interest. Although the results reported here are qualitatively similarto those of Borle (1974), particularly with regard to the effect of cyclic AMP in re-leasing calcium which is already accumulated in mitochondria, there are quantitativedifferences both in the concentration of nucleotide which is required to increase effluxof calcium, and in the magnitude of the efflux. Thus in the purified liver, kidney andheart mitochondrial preparations used by Borle a concentration of 2 /tM cyclic AMPcould elicit a reduction of at least 60 % in the calcium content of mitochondria. By con-trast, in the present study a concentration of 1 mM cyclic AMP was required to pro-duce a 50 % reduction in calcium binding by islet homogenates. It seems likely thatthe reasons for this difference in sensitivity lie in the different preparations used. Thusin homogenates some residual phosphodiesterase activity may still be present, despitethe use of a potent inhibitor (3-isobutyl-i-methylxanthine) of this enzyme, so thata relatively high cyclic AMP concentration would be required to elicit its effects.Furthermore, the presence of other calcium-binding sites in the homogenate, someof which may not be sensitive to cyclic AMP or in which cyclic AMP may actuallystimulate calcium uptake as is the case in liver plasma membranes (Schlatz & Marin-etti, 1972) or sarcoplasmic reticulum fragments (Tada, Kiichberger, Repke & Katz,1974), would tend to diminish the effectiveness of cyclic AMP in lowering calciumconcentrations in a specific pool. The effects of cyclic AMP were also observed inmitochondria + storage granule pellets obtained from rat islets by differential centri-fugation: however, the use of a purified mitochondrial preparation from islets will be

Calcium distribution in B cell organelles 405
required to determine whether the islet mitochondrial calcium pool is affected bycyclic AMP in a similar way to that of liver, kidney and heart.
Malaisse, in studies of influx and efflux of ^Ca from isolated islets in a perfusionsystem, showed that cyclic AMP and agents which raise cyclic AMP levels can in-crease the efflux of calcium from B cells, even in the absence of extracellular calcium,and suggested that cyclic AMP might induce the release of calcium from an intra-cellular 'vacuolar calcium pool' (Malaisse, 1973). The present results seem consistentwith this interpretation. Furthermore, AMP which is effective (although less so thancyclic AMP) in reducing calcium accumulation by homogenates, has also been shownto stimulate insulin secretion (Iversen, 1970).
It has also been suggested that a variety of insulin secretagogues including glucoseand amino acids may affect insulin release by altering intracellular calcium levels.We have therefore investigated the possibility of direct effects of other insulin secre-tagogues on calcium accumulation by organelles in islet homogenates. Glucose had aconsistent small effect but this is not considered important for the regulation of secre-tion, since galactose, a sugar which does not stimulate insulin release, also had a smalleffect. The amino acids L-arginine and L-leucine were without effect at the concentra-tions tested; glibenclamide, an insulin-releasing sulphonylurea derivative, althougheffective at concentrations of 10 or 100/ig/ml, had no significant effect on calciumaccumulation when used at concentrations (o-i-i-o/ig/ml) at which it is effective ininducing secretion from intact cells. There is therefore no evidence that these agentsexert their effects on secretion directly by releasing calcium from an organelle boundto a cytosolic pool or by increasing calcium accumulation in organelles. They mayrather increase cytosolic calcium levels by preventing calcium efflux across theplasma membrane as suggested by Malaisse (1973), or perhaps by increasing thebinding of calcium to cytosolic proteins so that their efflux is effectively prevented, orby some completely separate mechanism.
The relative importance of the various organelles in calcium accumulation
Borle (1973) suggested that in liver cells the area of the plasma membrane (1133/im2) is far less than the area of inner mitochondrial membrane (29000 fim2), so thatpotentially the mitochondria have 30 times the area of the plasma membrane availablefor calcium transport, and are likely to play an important role in regulating intra-cellular calcium levels. In the B cell the situation is likely to be similar: the area ofplasma membrane (973 /ims: Dean, 1973) is comparable to that of a liver cell (1133/im2: Borle, 1973), and the number of mitochondria per B cell is almost identical(996: Dean, 1973 vs 1000 in a liver cell: Lehninger, 1964). Thus one might expect thesurface area of inner mitochondrial membrane to be similar to that calculated forliver also - about 29000/tm2 (Lehninger, 1964). In addition, the B cell endoplasmicreticulum, which also stores calcium, may contribute an area of membrane of 8517/im2, of which 6158/im2 is rough-surfaced endoplasmic reticulum (Dean, 1973).Finally there are in the mouse B cell ~ 13000 granules of mean diameter includingsacs of 0-289/im (Dean, 1973) which contribute an area of membrane of 3435 /im2.
Storage granules contain the highest calcium concentration of any of the organelles

406 S. L. Howell, W. Montague and M. Tyhurst
as determined by X-ray microanalysis. However, radioautographic experiments(Howell & Tyhurst, in preparation) failed to reveal the presence of label associatedwith the storage granules. Two possible explanations of this discrepancy seem pos-sible: either the granules do not accumulate labelled ^Ca in short-term incubationsof this type, or the teCa present in the granules is totally lost during fixation and de-hydration. However, separate experiments in which comparisons of calcium uptakeby subcellular particles were made after filtration through Millipore membranesshowed no additional uptake of calcium by the isolated organelles between islethomogenates which were filtered through 450-nm diameter filters (which shouldallow granules to pass through) and 100-nm filters (which should retain all granules);the mean diameter of the granule core, which is spherical, is 220 nm in the rat.Furthermore, comparison of homogenates of islets prepared from normal rats andfrom rats which had been pretreated with glibenclamide, a potent insulin secreta-gogue which produces marked degranulation of B cells, showed no difference in thecharacteristics of calcium handling between the 2 groups. These observations suggestthat despite the presence of high concentrations of calcium within the B granules,which had been demonstrated by X-ray microanalysis, they may play only a minorrole in the short-term regulation of cytosolic calcium levels.
Rough-surfaced endoplasmic reticulum contains a calcium concentration estimatedby microanalysis which is only half that of the storage granules, and seems to representa site of calcium exchange within the cell, as suggested by radioautographic graindistribution. This appears consistent with a regulatory role for calcium uptake byelements of the endoplasmic reticulum, which was also inferred from the presentstudies of ^Ca uptake by islet homogenates. In quantitative terms, however, theendoplasmic reticulum may not represent the major calcium store in B cells.
Other cytoplasmic areas and nuclei contained low calcium concentrations asdetermined by X-ray microanalysis, and showed no detectable 46Ca uptake by radio-autography. Nor is there evidence from ^Ca accumulation experiments with sub-cellular fractions in the present studies to suggest an important role of these areasin calcium regulation in the B cell.
B cell mitochondria also contain substantial concentrations of calcium, and incontrast to the granules can also be shown by electron-microscopic radioautographyto accumulate calcium-45 (Howell & Tyhurst, in preparation). Metabolic studieshave also shown the presence in islet homogenates of ATP-dependent 4BCa uptakeso that in islet B cells as in liver, kidney and heart (Borle, 1973, 1974), mitochondriamay represent major sites of calcium accumulation and storage. It seems likely thatthe mitochondrial calcium pool may predominate in the minute-by-minute regulationof B cell cytosolic calcium levels, since it contains a higher concentration of calciumthan does the endoplasmic reticulum and a far larger area of membrane which isavailable for calcium transport than any other organelle. Furthermore, it appears tobe subject to regulation by cyclic AMP, which is known to be involved in the regula-tion of insulin secretion, in a way which the other calcium stores are not. In thisrespect it seems likely to be the most important of the particulate calcium poolswhich we have so far been able to identify.

Calcium distribution in B cell organelles 407
We thank Dr T. C. Appleton of the Physiological Laboratory, Cambridge University for hisadvice and for facilities for performing X-ray microanalyses using the ART EMMA-4 analyticalelectron microscope in that department which was provided by the Wellcome Trust. Financialassistance from the Medical Research Council, British Diabetic Association and HoechstPharmaceuticals is also gratefully acknowledged. S.L.H. is a CIBA Fellow.
REFERENCES
APPLETON, T. C. (1974). A cryostat approach to ultrathin dry frozen sections for electronmicroscopy: a morphological and X-ray analytical study. J. Microscopy 100, 49-74.
BORLE, A. B. (1973). Calcium metabolism at the cellular level. Fedn Proc. Fedn Am. Socs exp.Biol. 32, 1944-1950.
BORLE, A. B. (1974). Cyclic AMP stimulation of calcium efflux from kidney, liver and heartmitochondria. J. Membrane Biol. 16, 221-236.
CARAFOLI, E., GAMBLE, R. L. & LEHNINGER, A. L. (1966). Rebounds and oscillations inrespiration-linked movements of Ca + + and H + in rat liver mitochondria. J. biol. Client. 241,2644-2652.
DEAN, P. M. (1973). Ultrastructural morphometry of the pancreatic B cell. Diabetologia 9,1 1 5 - 1 1 9 .
GEY, G. O. & GEY, M. K. (1936). The maintenance of human normal cells and tumour cells incontinuous culture. Am. J. Cancer 27, 45-76.
HEDESKOV, C. J. & CAPITO, K. (1975). The restoring effect of caffeine on the decreased sensi-tivity of the insulin secretory mechanism in mouse pancreatic islets during starvation.Hormone Metab. Res. 7, 1-5.
HERMAN, L., SATO, T. & HALES, C. N. (1973). The electron microscopic localization of cationsto pancreatic islets of Langerhans and their possible role in insulin secretion. J. Ultrastmct.Res. 42, 298-311.
HOWELL, S. L., FINK, C. J. & LACY, P. E. (1969). Isolation and properties of secretory granulesfrom rat islets of Langerhans. I. Isolation of secretory granule fraction. J. Cell Biol. 41,154—161.
HOWELL, S. L. & MONTAGUE, W. (1975). Regulation by nucleotides of "calcium uptake inhomogenates of rat islets of Langerhans. FEBS Letters, Amsterdam 52, 48-52.
HOWELL, S. L. & TAYLOR, K. W. (1966). Effect of glucose concentration on the incorporationof (3H)leucine into insulin in isolated rabbit islets of Langerhans. Biochim. biophys. Acta 130,
HOWELL, S. L. & TYHURST, M. (1974). Cryo-ultramicrotomy of rat islets of Langerhans. J. CellSri 15, 591-603.
IVERSEN, J. (1970). Secretion of insulin and glucagon from isolated perfused canine pancreasfollowing stimulation with cyclic AMP, glucagon and theophylline. Diabetologia 6, 673.
KERN, H. F., KERN, D., SCHMIDT, F. H. & STORK, K. (1969). Fine structure of the islets ofLangerhans in rats and rabbits after treatment with glibenclamide. Hormone Metab. Res. 1,Suppl. 1, 11-17.
LEHNINGER, A. L. (1964). In The Mitochondrion, p. 30. New York: Benjamin.LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (1951). Protein measure-
ments with the Folin phenol reagent. J. biol. Chem. 293, 265-275.MALAISSE, W. J. (1973). Insulin secretion: multifactorial regulation for a single process of
release. Diabetologia 9, 167-173.MCCOLLUM, W. B., BESCH, H. R., ENTMAN, M. L. & SCHWARTZ, A. (1972). Apparent initial
binding rate of calcium by canine cardiac relaxing system. Am. J. Physiol. 223, 608-614.MOORE, C. L. (1971). Specific inhibition of mitochondrial Ca++ transport by ruthenium red.
Biochem. biophys. Res. Comrnun. 42, 298-305.SCHXFER, H. J. & KL5PPEL, G. (1974). The significance of calcium in insulin secretion. Virchows
Archiv. patli. Anat. Physiol. 362, 231-245.SCHLATZ, L. & MARINETTI, G. V. (1972). Calcium binding to the rat liver plasma membrane.
Biochim. biophys. Acta 290, 70-83.SULAKHE, P. V., DRUMMOND, G. I. & Nc, D. C. (1973). Calcium binding by skeletal muscle
sarcolemma. J. biol. Chem. 248, 4150-4157.27 C E L 19

408 5. L. Howell, W. Montague and M. Tyhurst
TADA, M., KOCHBERGER, M. A., REPKE, D. I. & KATZ, A. M. L. (1974). Stimulation of calciumtransport in cardiac sarcoplasmic reticulum by cAMP dependent protein kinase. J. biol. Chem.249, 6174-6180.
WEBER, A. & HERZ, R. (1968). The relationship between caffeine contracture of intact muscleand the effect of caffeine on reticulum. J. gen. Pliysiol. 52, 750—772.
{Received 29 April 1975)
Calcium distribution in B cell organelles 409
Fig. 5. Electron micrograph of a pellet obtained by centrifugation of a homogenate ofrat islets which had been incubated for 20 min in the buffer used throughout theseexperiments. Storage granules (g) with their limiting membrane intact, mitochondria(m) and microsomal vesicles (v) are all clearly present, x 24000 approx.
