cell-surface processing of pro-adamts9 … · koo et al, adamts9 processing 3/06 1 cell-surface...
TRANSCRIPT

Koo et al, ADAMTS9 processing 3/06
1
CELL-SURFACE PROCESSING OF PRO-ADAMTS9 BY FURINBon-Hun Koo, Jean-Michel Longpré‡, Robert P.T. Somerville, J. Preston
Alexander*, Richard Leduc‡ and Suneel S. Apte+
Dept. of Biomedical Engineering and Orthopaedic Research Center, Lerner Research Institute,Cleveland Clinic Foundation, 9500 Euclid Avenue, Cleveland OH 44195, ‡Department of
Pharmacology, Faculty of Medicine and Health Sciences, Université de Sherbrooke, Sherbrooke,Québec, J1H 5N4, Canada, *Triple Point Biologics, Portland, Oregon
Running title: Furin processing of pro-ADAMTS9Address correspondence to: Suneel S. Apte, M.B.B.S., D.Phil., Dept. of BiomedicalEngineering, Lerner Research Institute, Cleveland Clinic Foundation (ND20), 9500 EuclidAvenue, Cleveland OH 44195, USA. Telephone: 216 445 3278; Fax: 216 444 9198; E-mail:[email protected]
Processing of polypeptide precursors byproprotein convertases (PCs) such as furintypically occurs within the trans-Golginetwork. Here, we show in a variety of celltypes that the propeptide of ADAMTS9 isnot excised intracellularly. Pulse-chaseanalysis in HEK293F cells indicated that theintact zymogen was secreted to the cell-surface and was subsequently processedthere before release into the medium. Theprocessing occurred via a furin-dependentmechanism as shown using PC inhibitors,lack of processing in furin-deficient cells,and rescue by furin in these cells. Moreover,down-regulation of furin by siRNA reducedADAMTS9 processing in HEK293F cells.PC5A could also process proADAMTS9,but similarly to furin, processed forms wereabsent intracellularly. Cell-surface, furin-dependent processing of proADAMTS9creates a precedent for extracellularmaturation of endogenously producedsecreted proproteins. It also indicates theexistence of a variety of mechanisms forprocessing of ADAMTS proteases.
Zinc metalloendopeptidases, like mostproteases, are synthesized as zymogens, andthe amino-terminal propeptide is usuallyexcised. Propeptide excision usually leads toenzymatic activation, an important regulatoryevent, and it can occur intracellularly, at thecell surface, or extracellularly through avariety of proteolytic mechanisms. In one such
mechanism, the propeptide is proteolyticallyexcised by serine proteases of the mammaliansubtilisin-like proprotein convertase (PC)family (1-5). This mechanism is used by someMMPs (6,7), many ADAMs (8-10) and allADAMTS proteases studied thus far (11,12).In these proteases, removal of the propeptideappears to be mediated by the most widelydistributed PC, furin, and occurs within theconstitutive secretory pathway, specifically inthe trans-Golgi network (TGN) (7-9,13,14).
Furin is the best studied of the sevenPCs implicated in proprotein processing withinthe constitutive secretory pathway and it ispresent in virtually all cells (1,15). It is a type Itransmembrane protein that is itselfsynthesized as a zymogen that undergoesautocatalytic intramolecular activation (16).Furin cleaves on the carboxyl side of aconsensus recognition site that is rich in basicresidues (e.g. Arg-Xaa-Arg/Lys-Arg↓)(2,4,5,17). Most furin resides in the TGN, butsome is present at the plasma membrane, andshuttles between the cell surface and the TGN(18-20). Furin is also shed from cells and maybe functional in the extracellular space (21).Microbial toxins such as the anthrax protectiveantigen and diphtheria toxin are processed bycell surface furin, with important implicationsfor their toxicity (20,22). However, thephysiological role of cell-surface or secretedfurin in processing endogenous cellularproducts has remained elusive.
http://www.jbc.org/cgi/doi/10.1074/jbc.M511083200The latest version is at JBC Papers in Press. Published on March 14, 2006 as Manuscript M511083200
Copyright 2006 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
2
The ADAMTS proteases are a familyof 19 secreted enzymes, of which some havecritical physiological functions and have beenimplicated in inherited human disorders,namely Ehlers-Danlos syndrome type VIIC(ADAMTS2), Weill-Marchesani syndrome(ADAMTS10) and inheritedthrombocytopenic purpura (ADAMTS13) (23-26). Over-expression of ADAMTS5 andADAMTS4 is implicated in proteolytic loss ofaggrecan, a major cartilage component, inarthritis (27). ADAMTS proteases (exceptADAMTS13, the von Willebrand factor-processing protease, whose propeptidecontains 41 amino acid residues) aresynthesized with propeptides of around 220-240 a.a, which are larger than the MMP andADAM propeptides. Almost all ADAMTSpropeptides have consensus sites for theattachment of N-linked oligosaccharide (Asn-Xaa-Thr/Ser, where Xaa is any amino acidexcept Pro). Consensus recognition sequencesfor PCs are present at the junction of thepropeptide and catalytic domain, andADAMTS proteases may have additional,more N-terminal PC processing sites withintheir propeptides. Previous studies ofADAMTS1, ADAMTS4 and ADAMTS7suggested, that like ADAMs and the furin-processed MMPs, ADAMTS proteases wereprocessed in the TGN by PCs, although somecell surface processing of ADAMTS7 was alsoobserved (11-14,28).
ADAMTS9 is one of only twoADAMTS proteases that is highly conservedduring evolution (29-31). Its mRNA is widelyexpressed (29,30), especially duringembryonic development (32). ADAMTS9 isupregulated by inflammatory cytokines inchondrocytes (33). It may have a role inatherosclerosis and arthritis, since it canproteolytically process the proteoglycansversican and aggrecan (30) which areimportant constituents of the vessel wall, andcartilage, respectively. This potentialbiological significance of ADAMTS9
prompted an in-depth analysis of theprocessing of its propeptide. In the initialcharacterization of ADAMTS9, wedemonstrated that it could be processed atmore than one site (30). In continuing thesestudies, we have made the unexpectedobservation that ADAMTS9 processing isexclusively extracellular and occurs at the cellsurface in cells that express high levels offurin. These results establish a precedent forthe cell surface activation of precursorendogenous proteins by furin and are possiblyof broad biological relevance.
EXPERIMENTAL PROCEDURESExpression Plasmids and Site-DirectedMutagenesis Plasmids for expression offull-length ADAMTS9, or a truncated formcontaining the signal peptide, propeptide andcatalytic domain (Pro-Cat) with C-terminalmyc and His tags were previously described(30). Due to low expression levels of full-length ADAMTS9, we excised its openreading frame with the Myc-His tags as aNotI/PmeI fragment and then re-cloned thisinto pCEP4 (Invitrogen Corp, Carlsbad, CA)digested with Kpn I and blunt ended followedby Not I digestion. To insert a FLAG tagbetween Thr276 and Arg277 in the propeptide,site-directed mutagenesis (QuikChange kit,Stratagene, Cedar Creek, TX) was performedusing the forward primer 5′-AATAAGACGGACAACACAGACTACAA-GACGATGACGACAAGAGAGAAAAGAGGACCCAC-3′ (with FLAG encoding siteunderlined) and the reverse primer 5′ –GTGGGTCCTCTTTTCTCTCTTGTCGTCATCGTCTTTGTAGTCTGTGTTGTCCGTCTTAT-T – 3′ (FLAG encoding site underlined).
Plasmids for expression of full-lengthADAMTS1 or the propeptide and catalyticdomain of ADAMTS7 (ADAMTS7-Pro-Cat)were previously described (28,34). Antibodies and Immunoblotting Thepeptides RP1 and RP4 representing differentregions of the ADAMTS9 propeptide weresynthesized using Fmoc (N- (9-fluorenyl)
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
3
methoxycarbonyl) chemistry and conjugated toKLH. New Zealand White male rabbits (7-8pounds) were immunized with the conjugatesat biweekly intervals for 8 weeks. After aninitial injection in Freund’s complete adjuvant,subsequent injections were given inincomplete adjuvant. Antibody titer wasmeasured by enzyme-linked immunosorbentassay using free peptides. Affinity-purifiedantibodies were prepared using the respectiveimmobilized antigens. Anti-penta-Hismonoclonal antibody, anti-myc monoclonalantibody 9E10 and anti-FLAG M2 monoclonalantibody were obtained from commercialsources (Invitrogen Corp., and Sigma-AldrichChemical Co., St. Louis, MO). Anti-furinmonoclonal antibody was purchased fromAlexis Biochemicals (San Diego, USA).Immunoblotting was done using denaturingSDS-PAGE and electroblotting topolyvinylfluoridine membrane followed bydetection of the bound antibody usingenhanced chemiluminescense (AmershamBiosciences, Piscataway, NJ).Cell Culture, Transfection, And CellTreatmentsHEK293F cells stably transfected withADAMTS9 or Pro-Cat, COS-1 cells, LoVocells (ATCC No.; CCL-299), CHO-K1 cells,CHO RPE.40 (35) and rat chondrosarcomaRCS-LTC cells (36) were maintained asdescribed previously (30) or as per vendors’instructions. Transient transfections were doneusing Fugene 6 (Roche Diagnostics,Indianapolis, IN) as per the manufacturer’srecommendations. For inhibition of PCs, cellswere treated with the PC inhibitory peptidedec-Arg-Val-Lys-Arg-chloromethylketone(dec-RVKR-cmk, Calbiochem, La Jolla, CA)at different concentrations as indicated or withthe polypeptide α1-PDX (Portland variant of α1-antitrypsin) (37).Detection of Cell-Surface ProcessingBiotinylation of cells was done on ice toprevent labeling of intracellular proteins,essentially as previously described (28). As a
control to eliminate cell-surface proteins, cellswere treated with 0.05% trypsin/0.53mMethylenediamine-tetraacetic acid (EDTA) for20 min on ice, and trypsin was subsequentlyinactivated with 10% FBS supplementedmedium.
To examine the fate of cell surfacebiotinylated protein, biotinylated cells weretransferred to serum-free medium, and cellsand media were collected at sequential timepoints. Biotinylated proteins were capturedfrom samples using streptavidin-agarose(Sigma-Aldrich) and eluted by boiling inLaemmli sample buffer followed by SDS-PAGE and western blotting with appropriateantibodies.
Pro-Cat was purified from lysates ofstably-transfected cells using Pro-Bond resin(Invitrogen). For determination of processingof exogenously added Pro-Cat by cells,untransfected HEK293F cells were cultured toconfluence on 6 well plates and then in 293SFM-II medium (Invitrogen). Purified Pro-Cat(50 µg/well) was added to untransfectedHEK293F cells and the cells were culturedfurther over an 8 hr period. The medium wastaken for analysis at successive time points. Inparallel experiments purified Pro-Cat wasincubated with cell-free conditioned mediumfrom HEK293F cells to investigate whetherthe processing was cell-mediated or not. Thesamples were analyzed by western blottingwith anti-myc.Enzymatic Deglycosylation Removal of N-linked carbohydrate with peptide N-glycosidase F (PNGaseF, New EnglandBiolabs, Beverly, MA) was performed aspreviously described (30).Co-Transfection Of Pro-Cat And ProproteinConvertases Furin-deficient CHO RPE.40cells (35) and LoVo cells were transfectedwith Pro-Cat alone or together with plasmidsencoding the proprotein convertases furin,PACE4 and PC5A (kindly provided by Dr.Nabil Seidah). 48 hrs after transfection, cellswere incubated in 293 SFM-II medium for 24
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
4
hrs, and western blotting of the cell lysate andmedium was performed with anti-myc.Cell Surface Cross-linking AndImmunoprecipitation 48 hrs aftertransfection with the Pro-FLAG-Cat plasmid,HEK293F cells were washed four times withPBS and treated with the thiol-cleavable,membrane nonpermeable cross-linker 3, 3’-Dithiobis[sulfosuccinimidylpropionate](DTSSP, Pierce Endogen, Rockford, IL) tocross-link molecules at the cell surface. Thecells were lysed in lysis buffer (50 mM Tris-HCl, pH7.5, 5 mM EDTA, 150 mM NaCl, 1%Nonidet P-40, protease inhibitor cocktail(Roche Diagnostics) for 1 hr at 4°C andcentrifuged. The soluble portion of the lysatewas transferred to a fresh tube and incubatedovernight with anti-FLAG-agarose (Sigma-Aldrich) at 4°C with rotation. Anti-FLAG-agarose was pelleted by centrifugation at 1000rpm in a microcentrifuge and washed six timesin lysis buffer. The supernatant was discarded,and the bound protein was eluted with 0.1 Mglycine/HCl (pH 3.5) from the resin. Theeluted samples were analyzed by westernblotting with anti-furin monoclonal antibody(Alexis Biochemicals) or RP4 antibody.Metabolic Labeling And Pulse-ChaseAnalysis HEK-293F cells stably expressingPro-Cat were grown to confluence andincubated in Dulbecco’s modified Eagle’smedium (DMEM) without cysteine andmethionine (Sigma-Aldrich). After 24 h, themedium was replaced with cysteine-methionine free medium containingradioactive cysteine/methionine (50µCi/well in6-well plates) (Pulse medium) for 15 minusing EXPRE35S35S (Perkin-Elmer LifeSciences, Boston, MA). For pulse-chaseanalysis, the pulse medium was removed, andthe cells were washed 3 times on ice with PBScontaining 0.5mM MgCl2 and 0.5mM CaCl2,followed by incubation for 0, 15, 30, 60, 120min in DMEM containing 10% FBS (Chasemedium) at 37°C. At the end of the chaseperiod, cells were placed on ice, the medium
was collected, and the cells were washed 4times with PBS and extracted with lysis buffer.In parallel, samples were processed forremoval of cell surface proteins in these assaysby treatment with trypsin/EDTA for 20 min onice, and trypsin was subsequently inactivatedwith 10% FBS supplemented medium. Themedia and cell lysates were processed forimmunoprecipitation with anti-myc followedby fluorography as described previously (30).RNA Interference—HEK-293F cells stablyexpressing Pro-Cat were transfected with 20pmol Furin ShortCut siRNA Mix (NewEngland Biolabs) using Lipofectamine 2000(Invitrogen). After 48 hrs of incubationwithout antibiotics, the medium was changedto 293 SFM-II medium (Invitrogen) and cellswere incubated for a further 24 hrs. Theconditioned medium and cell lysate wereanalyzed by western blotting with anti-mycand anti-RP4. Cell surface biotinylation wasdone as above.
RESULTSThe Proteolytically Processed ADAMTS9Propeptide Is Present In The ConditionedMedium But Not In The Cell Lysate TheADAMTS9 propeptide (Fig. 1A,B) has 5consensus furin-recognition sequences (Arg-Xaa-Xaa-Arg). Arg-Lys-Asp-Arg33 and Arg-Thr-Arg-Arg 74 are near the signal peptidaseprocessing site, whereas the remaining threesites, (Arg-Glu-Lys-Arg 280, Arg-Thr-His-Arg283 and Arg-Thr-Lys-Arg 287) overlap, and areclustered near the junction of the propeptideand catalytic domain. We previously showedthat mutation of Arg74 and Arg287 abrogatedprocessing at these sites, whereas mutation ofArg280, which compromised two putative sites,was without effect (30). The propeptide alsocontains three sequence motifs for attachmentof N-linked oligosaccharides (Fig. 1B),whereas the catalytic domain has none.
Two non-overlapping syntheticpeptides from the propeptide (RP1 and RP4;sequences are not provided for proprietaryreasons of Triple Point Biologics) within the
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
5
propeptide were used for the production ofpolyclonal antisera. In HEK293F cells stablyexpressing full-length ADAMTS9 (Fig. 2A),anti-RP4 recognized the expected ~180 kDazymogen in the cell lysate. Smallerpolypeptides (37 kDa and a ~20-22 kDadoublet) were recognized in the conditionedmedium. These bands were not seen inuntransfected HEK293F cells (Fig. 2A). Whenthe medium was deglycosylated using PNGaseF, the 37 kDa band and the ~22 kDa doubletincreased in mobility and migrated at 28 and15 kDa respectively, demonstrating thepresence of N-linked carbohydrate (Fig. 2B),characteristic of propeptide fragments.Notably, the ~22 kDa doublet converted to a~15 kDa doublet, indicating two proteinspecies rather than glycoforms of the samefragment. When these experiments wereconducted using an ADAMTS9 active sitemutant (ADAMTS9 Glu381Ala; a widely usedmetalloprotease inactivating mutation (12,38)),no differences were seen (Fig. 2B); thus, theanti-RP4 reactive fragments were notautoproteolytic in origin. In western blottingwith anti-RP1 the medium showed a weaklyimmunoreactive 37 kDa band, but not the ~22kDa doublet (Fig. 2C), whereas PNGaseFtreatment clearly resolved the 28 kDa bandseen with RP4 and a 15 kDa doublet also seenwith RP4. The comparable results with RP4and RP1 as well as the presence ofglycosylation identified these fragments in themedium as originating from the propeptide.The RP1 peptide is adjacent to an N-linkagesite and the carbohydrate may mask reactivityof the ~22 kDa fragments without priorPNGase F treatment. Since anti-RP4 providedmore robust immunoreactivity, it was used insubsequent experiments.
Since the large size and extensiveglycosylation of full-length ADAMTS9 (30)precludes accurate resolution in gels, theabove studies were repeated using anexpression plasmid encoding the ADAMTS9signal peptide, propeptide and catalytic
domain (Pro-Cat) with a myc-His tag at the C-terminus (Fig. 2D). The results were identical,i.e., anti-RP4 detected the intact Pro-Catzymogen (55 kDa) only in the cell lysate, and37 kDa and 20-22 kDa fragments were foundonly in the conditioned medium (Fig. 2D).Similarly, anti-myc detected only theunprocessed 55 kDa Pro-Cat zymogen in thecell lysate, and only the processed catalyticdomain (29 kDa) in conditioned medium (Fig.2D). Incubation of conditioned medium withPNGase F enhanced the mobility of propeptidefragments to the same extent as with full-length ADAMTS9 (Fig. 2E). That thesefragments originated from the propeptide wasfurther verified by insertion of a FLAGepitope tag between Thr276 and Arg277, i.e.upstream of the furin processing site at Arg287.Western blotting with anti-FLAG providedsimilar results to that with anti-RP4, withdifferences (Fig. 2F) reflecting distinctlocations of the FLAG and RP4 epitopeswithin the propeptide. These resultsdemonstrate that the propeptide is part of theintact zymogen in HEK293F cells, but that it ispresent as a distinct, proteolytically derivedentity in their conditioned medium.
We conclude that the 37 kDa band seenwith all propeptide antibodies corresponds tothe complete propeptide. Whendeglycosylated, its size (28 kDa), iscompatible with processing at the most N-terminal and C-terminal PC processing sites,i.e., Leu34-Arg287 (28 kDa). The site at whichthe processing of the prodomain to the 20 and22 kDa RP4 reactive fragments occurs is notestablished, but it may represent cleavage atthe Arg-Arg209 -↓Ser. This site is not a typicalfurin processing site, but it has certainattributes that favor PC processing- such asHis at the P6 position and Ser at the P1′position.
We asked whether the conditionedmedium of cells endogenously expressingADAMTS9 or from other transfected celltypes had similar processing profiles. RCS-
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
6
LTC chondrosarcoma cells endogenouslyexpressing ADAMTS9 demonstrated an anti-RP4-reactive 22 kDa band in their conditionedmedium, but not in the cell lysate, where onlya 180 kDa band corresponding to the intactzymogen was detected (Fig. 3A). When COS-1 and CHO-K1 cell lines were transientlytransfected with Pro-Cat, anti-RP4 detectedonly the unprocessed zymogen in cell lysate,but both the secreted, unprocessed zymogenand proteolytically processed pro-domain inthe conditioned medium (Fig. 3B,C, leftpanels). Western blotting with anti-mycdetected the intact zymogen in cell lysate andthe 29 kDa processed catalytic domain in theconditioned medium (Fig. 3 B, C, rightpanels). Thus, in all the cell types examined,the ADAMTS9 propeptide was not processedwithin the cell. Varying amounts ofunprocessed Pro-Cat were detected inconditioned medium of transfected COS-1 andCHO-K1 cells (Fig. 3 B, C, left panels), butlittle in HEK293F cells (Fig. 2). Together, theresults suggest that the ADAMTS9 propeptideis intrinsically resistant to removal within thesecretory pathway, since it is not processedintracellularly regardless of the cell type.ADAMTS9 Processing Differs From That OfOther ADAMTS Proteases Previous studiesdemonstrated intracellular processing ofADAMTS1, ADAMTS4 and ADAMTS7(13,14,28,34). We directly comparedADAMTS9 processing with ADAMTS1 andADAMTS7 processing. ADAMTS1 andADAMTS9 Pro-Cat were transfected intoQBI-HEK 293A cells, followed by metaboliclabeling with radioactive amino acids andimmunoprecipitation with anti-ADAMTS1and anti-penta-His. Fluorography of the cell-and medium-derived proteins demonstratedADAMTS1 zymogen (110 kDa) and theprocessed enzyme (87 kDa) in the cell, butonly the unprocessed 55 kDa ADAMTS9 Pro-Cat in cells (Fig. 4A). In a second set ofexperiments, ADAMTS9-Pro-Cat wascompared to ADAMTS7-Pro-Cat. Cell lysate
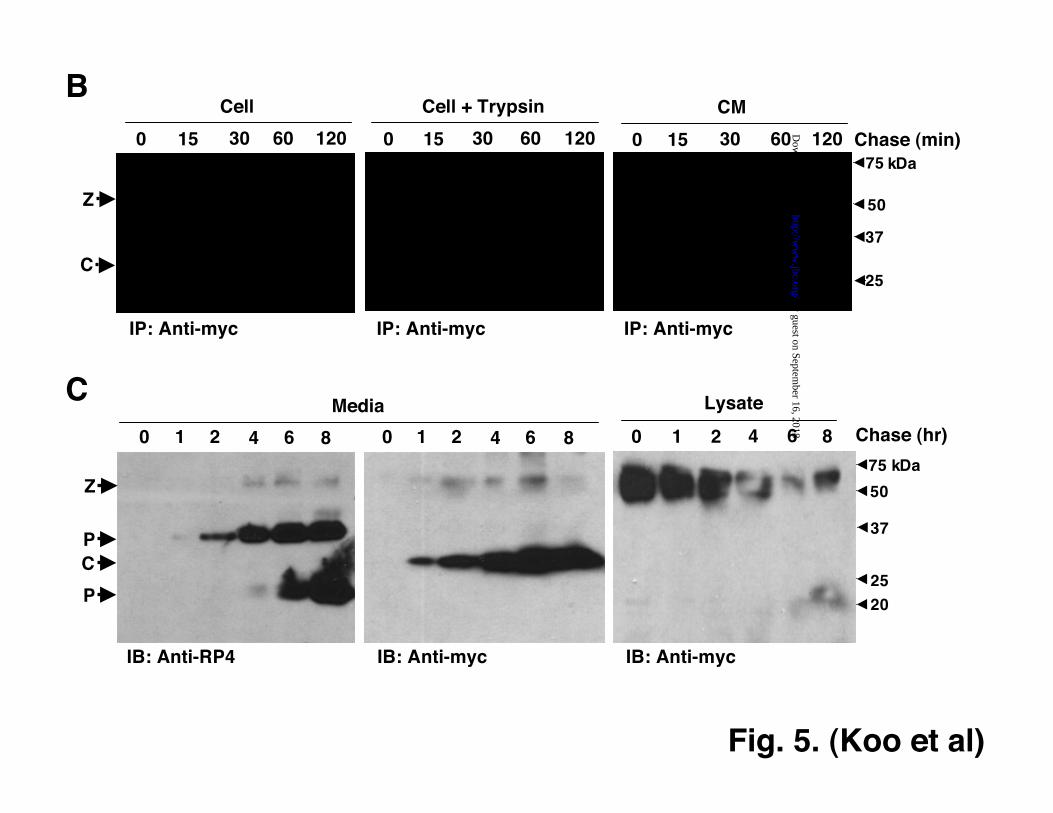
from ADAMTS7-Pro-Cat-transfectedHEK293F cells showed both the intact Pro-Catand the predicted furin-processed catalyticdomain, whereas ADAMTS9-Pro-Cattransfected cells showed only the unprocessedzymogen (Fig. 4B). These results demonstratethat differences in biosynthetic mechanismsexist in the ADAMTS family.ADAMTS9 Is Processed At The CellSurface The presence of only the intact pro-ADAMTS9 in cells and of the processedpropeptide exclusively in the conditionedmedium, suggested that propeptide processingoccured extracellularly, i.e. at the cell-surfaceor in the medium. We utilized a variety ofapproaches to examine this possibility further.First, cell surface proteins were labeled withbiotin. Biotinylated proteins were purifiedfrom detergent-extracted cells usingstreptavidin-agarose capture, and analyzed byimmunoblotting with anti-myc. As a control,cells were treated with trypsin to remove cellsurface proteins and proteoglycans. Intact Pro-Cat (55 kDa) was present at the surface ofHEK293F cells (Fig. 5A).
Since biotinylation is very sensitiveand may reveal only a small fraction of thetotal secreted ADAMTS9, we undertookpulse-chase analysis of all newly synthesizedprotein and followed the labeled protein intothe medium. The proportion of cell-surfacePro-Cat at any point during the chase wasdetermined by digesting away cell-surfaceproteins with trypsin. Over the period of thechase, the total cellular levels of the 55 kDaPro-Cat zymogen diminished slightly, but thetrypsin-sensitive protein accounted for asignificant fraction of it 30 min after synthesisand almost all the cellular protein by 2 hr (Fig.5B, left and center panels). This observationwas complemented by analysis of theprocessed catalytic domain in conditionedmedium. It was detectable by 30 min afterbiosynthesis and increased steadily throughoutthe chase (Fig. 5B, right panel). No zymogenwas detected in the conditioned medium. This
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
7
suggested that the majority of synthesizedzymogen was located at the cell surface andthat it was processed there prior to release ofthe catalytic domain into the medium.
To confirm that this was indeed thefate of cell surface Pro-Cat, biotinylated cellswere transferred to fresh medium, and thebiotinylated proteins were chased at hourlyintervals over an 8 hr period in the conditionedmedium and cells. As early as 2 hrs into thechase, detectable levels of processed RP4-reactive propeptide (37 kDa) and myc-reactivecatalytic domain (29 kDa) were detected in theconditioned medium, with a plateau attained at6 hrs (Fig. 5C, left and center panelrespectively). Notably, very little intact Pro-Cat was detected in the medium. The levels ofcell-surface Pro-Cat gradually declined andwere minimal by 6 hrs (Fig. 5C, right panel).The 20-22 kDa fragments did not appear until6 hrs (Fig. 5C, left panel), suggesting theyfollow, and may be dependent on initial PCprocessing, and that the proteolytic activityresponsible for these fragments is within theconditioned medium.
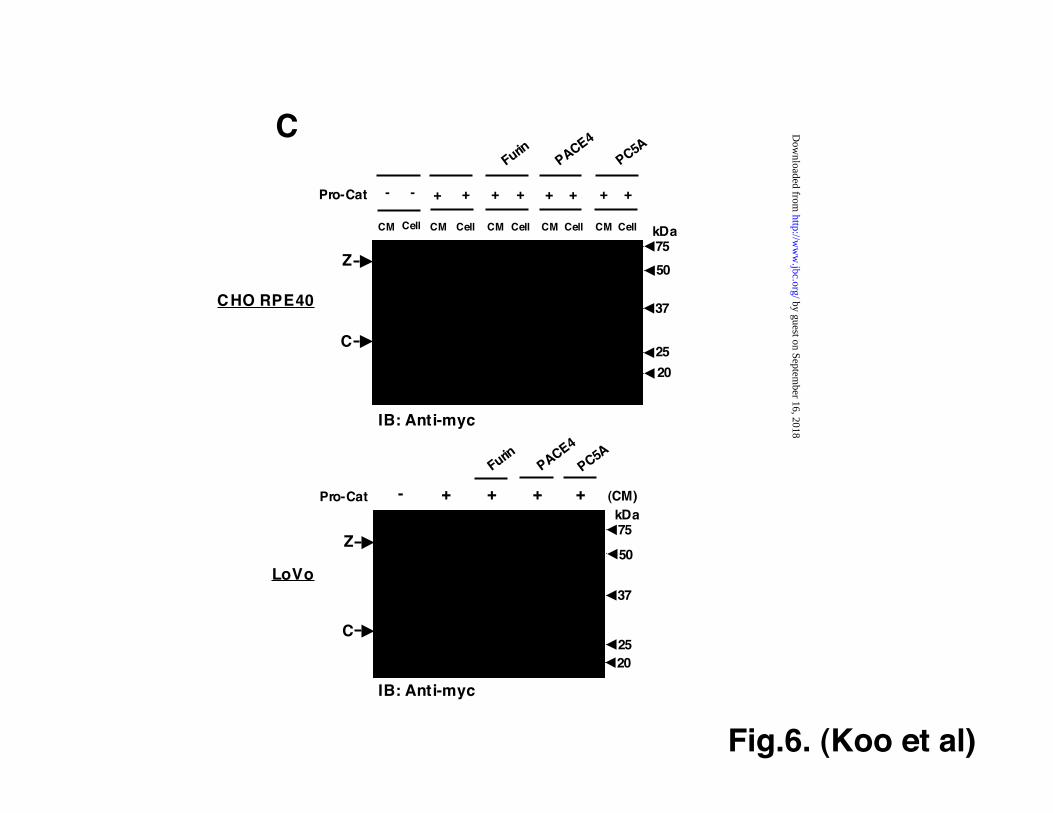
To ask whether exogenously addedPro-Cat was processed by the cell-surfaceproteolytic machinery, Pro-Cat purified fromdetergent-solubilized cell lysate using Ni-chelating resin was added to untransfectedHEK293F cells, and processing was evaluatedby western blotting of the medium. Noalteration was seen in conditioned mediumincubated in cell-free conditions, but anincrease in processing was observed after Pro-Cat was incubated with cells (Fig. 5D). Proprotein Convertases Mediate PropeptideProcessing Of ADAMTS9 Zymogen Wepreviously showed that furin processedADAMTS9 after Arg287 (30). To determine ifPCs have an exclusive role in ADAMTS9zymogen processing, cells were treated withincreasing concentrations of the membranesoluble PC inhibitor dec-RVKR-cmk.Processing of Pro-Cat was inhibited atconcentrations as low as 10 µM and was
essentially abolished by 50 µM (Fig. 6A). In asecond approach, HEK293F cells expressingPro-Cat were treated with increasing amountsof the furin inhibitor, α1-PDX (39), whichsubstantially reduced the processing of Pro-Cat in a 2 hr experiment (Fig. 6B). We nextinvestigated which widely-expressed PCsassociated with the constitutive secretorypathway could mediate processing. When Pro-Cat was expressed in CHO RPE.40 cells, theintact 55kDa zymogen was detectable in theconditioned medium and cell lysate and nofully processed catalytic domain was detectedin conditioned medium (Fig. 6C, upper panel).When furin was co-transfected with Pro-Cat,the 29 kDa processed catalytic domainappeared in the conditioned medium (Fig. 6C,upper panel). Transfection of PC5A (Fig. 6C,upper panel), and PC7 (data not shown) alsoled to appearance of the 29 kDa His-taggedcatalytic domain in the conditioned medium,but not in cell lysate. On the other hand,PACE4 did not produce the fully processedcatalytic domain (Fig. 6C, upper panel). Whensimilar experiments were conducted in LoVocells, the outcomes were comparable to thosein CHO RPE.40 cells (Fig. 6C lower panel).The ability of PACE4 to process Pro-Catefficiently to a 35 kDa myc-reactive fragmentargues for the presence of a PACE4susceptible site in the propeptide, possibly,Arg-Arg209↓Ser, but argues that PACE4 doesnot process ADAMTS9 following Arg287.Overall, the most efficient conversion was byfurin (Fig. 6C).Furin and ADAMTS9 are part of a complexat the cell surface Since all the aboveevidence suggested that ADAMTS9 wasprocessed at the cell-surface by furin, weasked whether furin was present at the surfaceof HEK293F cells. Using the samebiotinylation approach used to identifyADAMTS9 zymogen at the cell-surface, wecould also show the presence of furin at thecell-surface (Fig. 7A). In order to ask if furinand Pro-FLAG-Cat existed within a complex,
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
8
we first cross-linked cell-surface proteinsusing a reducible cross-linker andimmunoprecipitated cellular proteins usinganti-FLAG M2 agarose. Furin could be co-immunoprecipitated with FLAG-taggedADAMTS9 Pro-Cat suggesting its existence aspart of a cell-surface complex that alsocontains ADAMTS9 (Fig. 7B). However,efforts to immunoprecipitate furin andADAMTS9 Pro-Cat in solution or withoutcross-linking were unsuccessful (data notshown), suggesting that furin was not directlyresponsible for retention of Pro-Cat at the cellsurface. Furthermore, when furin was co-expressed with Pro-Cat in CHO RPE.40 cells,there was not an increase in cell surface Pro-Cat that would be expected if furin wasdirectly responsible for its sequestration.Instead, the cell-surface ADAMTS9 Pro-Catwas decreased when co-expressed with furin,suggesting that furin contributed directly toprocessing, but not sequestration at the cellsurface (Fig. 7C). Furin Is Essential For ADAMTS9Propeptide Processing At The Surface ofHEK293F cells Since the gain-of-functionexperiments that were done by transfectingfurin-deficient cells suggested that furin couldmost efficiently process the ADAMTS9propeptide, we asked whether it is responsiblefor the processing in HEK293F cells in a loss-of-function experiment. In order to testwhether reduction of furin levels in HEK293Fcells leads to reduction in cell surfaceprocessing, the cells were transfected withfurin siRNA, which significantly reduced totalcellular furin (Fig. 8A). In these cells, neitherthe total amount of ADAMTS9 Pro-Cat, northe total level of cell-surface Pro-Cat werealtered (Fig. 8B). However, in contrast to cellsthat were not siRNA treated, the conditionedmedium contained significant amount ofunprocessed zymogen (Fig. 8C).
DISCUSSIONWe previously investigated PC-
processing of several ADAMTS zymogens(11,13,28,30,40). The absence of matureADAMTS9 in cell lysates that was noted in aprevious study reporting the complete primarystructure of ADAMTS9 (30) led us to carryout the present comprehensive analysis of itsmaturation. We examined ADAMTS9propeptide processing using variousapproaches, none of which provided anyevidence for intracellular processing. Bothgain-of-function, and loss-of-functionapproaches, supported the conclusion thatfurin processes the ADAMTS9 propeptide.Furthermore, the data suggest that the cell-surface is a major site of ADAMTS9propeptide processing in HEK293F cells orcells that express high levels of cell-surfaceprocessing activity. In contrast, cellsexpressing no furin activity (CHO RPE.40 orLoVo) or lower levels of furin than HEK293Fcells (e.g. COS-1 and CHO-K1) secretevarying amounts of unprocessedproADAMTS9. Whether proADAMTS9 isprocessed at the cell-surface or potentially inthe extracellular space, therefore appears todepend on the relative quantity of furin orother PCs present at these locations. AlthoughCHO RPE.40 and LoVo cells may expressother PCs, they processed ADAMTS9inefficiently. Indeed, the widespreadexpression of ADAMTS9 (32) is compatiblewith physiological processing by furin, sincethis is a ubiquitously distributed convertase.However, there is overlap of ADAMTS9expression (32) with PC5A during mousedevelopment (17) suggesting that processingby this enzyme may occur in vivo as it did inco-transfection experiments.
The data raised the issue of whether ornot furin might directly mediate cell surface-binding of ADAMTS9. We argue that it doesnot. When transfected into CHO.RPE 40 cells,furin does not increase the amount of cellsurface zymogen, as one might expect if it
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
9
contributed significantly to cell-surfacebinding, but decreases it, presumably becauseit processes ADAMTS9 and the processedforms are released. In addition, although wecould co-precipitate furin and ADAMTS9after chemical cross-linking of cell-surfacemolecules, we could not demonstrate aninteraction between them without cross-linking. This suggests the existence of cell-surface complex that contains furin,ADAMTS9, as well as other molecules.However, since unprocessed zymogen isultimately released into the medium in cellswith low levels of furin activity, the binding islikely to be of low affinity. Significantly,neither the cleaved propeptide nor theprocessed catalytic domain were ever detectedat the cell surface. This suggests that onceprocessed, by furin, the cell surfaceinteractions in which the Pro-Cat zymogenparticipates are abruptly disrupted. Thus,properties of both the propeptide and thecatalytic domain, or the junctional regionbetween them might be crucial in cell surfacebinding of this construct.We propose that cell-surface interactions ofADAMTS9 Pro-Cat could be one way oftargeting its proteolytic activity to thepericellular space. Although the function ofADAMTS9 is presently unknown, it is presentat the cell surface in cultured cells (30), and itshomologs are known to be involved in cellmigration (41,42), which typically requirescell-surface proteolysis (43). Thus, retention ofthe propeptide may be a mechanism forsequestration of ADAMTS9 at the cell surface,although the ADAMTS9 ancillary domain isalso likely to have a role in this context (30).
Traditionally, PCs have been shown tocleave their substrates intracellularly. This isparticularly true for furin, the best-knownmember of this protease family (1). Molecularshedding events that take place at the cellsurface are mostly attributable to PC-activatedcell surface proteases such as the ADAMs andMT-MMPs (44,45). Furin is known to exist at
the cell surface (19,46) and other PCs such asPACE4 and PC6/5A are known to be secretedand anchored in the extracellular matrix (47),and are therefore presumed to haveextracellular substrates. Protective antigen ofanthrax and diphtheria toxin are cleaved atconsensus furin cleavage sites on the exteriorof the cell, as well as in intracellular organelles(20,22). This process nevertheless differs fromcell-surface processing of endogenouslyproduced proproteins. Previously, biosyntheticanalysis of the extracellular matrix proteinfibrillin-1 had suggested that furin- processingof profibrillin-1 did not occur intracellularly,although the precise location of processingwas not identified (48). ADAMTS9, to ourknowledge, is thus the first cellular proteinshown to be selectively processed by furin atthe cell surface.
Why is the ADAMTS9 propeptide notexcised in the TGN? The reasons for this arepresently unknown and are the subject ofcontinuing investigation in our laboratory. It ispossible that the processing sites in thepropeptide may be conformationallyinaccessible to PCs intracellularly, or maskedby molecular chaperones. Once exported fromthe cell, the zymogen may undergo aconformational change or detach fromchaperones and enable furin processing.
Although the processing mechanismsof all 19 ADAMTS proteases have yet to beelucidated, the observations reported here andin some other ADAMTS proteases, suggestthat the constitutive intracellular processingmodel is not universally applicable to thisfamily. Instead, ADAMTS proteases exhibit aspectrum of activation mechanisms thatincludes classical intracellular processing(ADAMTS1 and ADAMTS4) (13,14), cellsurface processing (ADAMTS9), acombination of intracellular and cell surfaceprocessing (ADAMTS7) (28) and possibly acombination of cell-surface and matrix-associated processing (ADAMTS10) (40). Infact, even in furin-rich HEK293F cells, a
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
10
significant proportion of ADAMTS10 appearsto be secreted as the unprocessed zymogen(40).
Beyond ADAMTS proteases, however,the broader significance of these results is thatthey offer fundamental insights onmechanisms of molecular maturation, andsuggest that cell-surface processing ofproproteins by furin and other PCs may bemore widely prevalent than hitherto suspected.
ACKNOWLEDGMENTS: This research wassupported by NIH award AR49930 to S. Apteand a grant from the Canadian Institutes ofHealth Research to R. Leduc. R. Leduc is aFonds de la Recherche en Sante du Quebec(FRSQ) Chercheur National. J-M. Longpre is arecipient of a PhD studentship of the FRSQ.The authors are grateful to Carl Blobel,Thomas Weimbs and Karen Colley for helpfuldiscussions.
REFERENCES
1. Thomas, G. (2002) Nat Rev Mol Cell Biol 3, 753-7662. Bergeron, F., Leduc, R., and Day, R. (2000) J Mol Endocrinol 24, 1-223. Zhou, A., Webb, G., Zhu, X., and Steiner, D. F. (1999) J Biol Chem 274, 20745-207484. Nakayama, K. (1997) Biochem J 327 ( Pt 3), 625-6355. Seidah, N. G., and Prat, A. (2002) Essays Biochem 38, 79-946. Pei, D., and Weiss, S. J. (1995) Nature 375, 244-2477. Yana, I., and Weiss, S. J. (2000) Mol Biol Cell 11, 2387-24018. Lum, L., Reid, M. S., and Blobel, C. P. (1998) J Biol Chem 273, 26236-262479. Cao, Y., Kang, Q., Zhao, Z., and Zolkiewska, A. (2002) J Biol Chem 277, 26403-2641110. Leonard, J. D., Lin, F., and Milla, M. E. (2005) Biochem J 387, 797-80511. Rodriguez-Manzaneque, J. C., Milchanowski, A. B., Dufour, E. K., Leduc, R., and
Iruela-Arispe, M. L. (2000) J Biol Chem 275, 33471-3347912. Kuno, K., Terashima, Y., and Matsushima, K. (1999) J Biol Chem 274, 18821-1882613. Longpre, J. M., and Leduc, R. (2004) J Biol Chem 279, 33237-3324514. Wang, P., Tortorella, M., England, K., Malfait, A. M., Thomas, G., Arner, E. C., and Pei,
D. (2004) J Biol Chem 279, 15434-1544015. Denault, J. B., and Leduc, R. (1996) FEBS Lett 379, 113-11616. Leduc, R., Molloy, S. S., Thorne, B. A., and Thomas, G. (1992) J Biol Chem 267, 14304-
1430817. Zheng, M., Seidah, N. G., and Pintar, J. E. (1997) Dev Biol 181, 268-28318. Mayer, G., Boileau, G., and Bendayan, M. (2003) J Cell Sci 116, 1763-177319. Mayer, G., Boileau, G., and Bendayan, M. (2004) J Histochem Cytochem 52, 567-57920. Klimpel, K. R., Molloy, S. S., Thomas, G., and Leppla, S. H. (1992) Proc Natl Acad Sci
U S A 89, 10277-1028121. Vidricaire, G., Denault, J. B., and Leduc, R. (1993) Biochem Biophys Res Commun 195,
1011-101822. Tsuneoka, M., Nakayama, K., Hatsuzawa, K., Komada, M., Kitamura, N., and Mekada,
E. (1993) J Biol Chem 268, 26461-2646523. Apte, S. S. (2004) Int J Biochem Cell Biol 36, 981-98524. Colige, A., Sieron, A. L., Li, S. W., Schwarze, U., Petty, E., Wertelecki, W., Wilcox, W.,
Krakow, D., Cohn, D. H., Reardon, W., Byers, P. H., Lapiere, C. M., Prockop, D. J., andNusgens, B. V. (1999) Am J Hum Genet 65, 308-317
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
11
25. Dagoneau, N., Benoist-Lasselin, C., Huber, C., Faivre, L., Megarbane, A., Alswaid, A.,Dollfus, H., Alembik, Y., Munnich, A., Legeai-Mallet, L., and Cormier-Daire, V. (2004)Am J Hum Genet 75, 801-806
26. Levy, G. G., Nichols, W. C., Lian, E. C., Foroud, T., McClintick, J. N., McGee, B. M.,Yang, A. Y., Siemieniak, D. R., Stark, K. R., Gruppo, R., Sarode, R., Shurin, S. B.,Chandrasekaran, V., Stabler, S. P., Sabio, H., Bouhassira, E. E., Upshaw, J. D., Jr.,Ginsburg, D., and Tsai, H. M. (2001) Nature 413, 488-494.
27. Arner, E. C. (2002) Curr Opin Pharmacol 2, 322-329.28. Somerville, R. P., Longpre, J. M., Apel, E. D., Lewis, R. M., Wang, L. W., Sanes, J. R.,
Leduc, R., and Apte, S. S. (2004) J Biol Chem 279, 35159-3517529. Llamazares, M., Cal, S., Quesada, V., and Lopez-Otin, C. (2003) J Biol Chem 278,
13382-1338930. Somerville, R. P., Longpre, J. M., Jungers, K. A., Engle, J. M., Ross, M., Evanko, S.,
Wight, T. N., Leduc, R., and Apte, S. S. (2003) J Biol Chem 278, 9503-951331. Huxley-Jones, J., Apte, S. S., Robertson, D. L., and Boot-Handford, R. P. (2005) Int J
Biochem Cell Biol32. Jungers, K. A., Le Goff, C., Somerville, R. P., and Apte, S. S. (2005) Gene Expr Patterns
5, 609-61733. Demircan, K., Hirohata, S., Nishida, K., Hatipoglu, O. F., Oohashi, T., Yonezawa, T.,
Apte, S. S., and Ninomiya, Y. (2005) Arthritis Rheum 52, 1451-146034. Rodriguez-Manzaneque, J. C., Milchanowski, A. B., Dufour, E. K., Leduc, R., and
Iruela-Arispe, M. L. (2000) J Biol Chem 275, 33471-33479.35. Spence, M. J., Sucic, J. F., Foley, B. T., and Moehring, T. J. (1995) Somat Cell Mol
Genet 21, 1-1836. Fernandes, R. J., Hirohata, S., Engle, J. M., Colige, A., Cohn, D. H., Eyre, D. R., and
Apte, S. S. (2001) J Biol Chem 276, 31502-31509.37. Dufour, E. K., Denault, J. B., Hopkins, P. C., and Leduc, R. (1998) FEBS Lett 426, 41-4638. Roghani, M., Becherer, J. D., Moss, M. L., Atherton, R. E., Erdjument-Bromage, H.,
Arribas, J., Blackburn, R. K., Weskamp, G., Tempst, P., and Blobel, C. P. (1999) J BiolChem 274, 3531-3540
39. Anderson, E. D., Thomas, L., Hayflick, J. S., and Thomas, G. (1993) J Biol Chem 268,24887-24891
40. Somerville, R. P., Jungers, K. A., and Apte, S. S. (2004) J Biol Chem 279, 51208-5121741. Blelloch, R., and Kimble, J. (1999) Nature 399, 586-59042. Rao, C., Foernzler, D., Loftus, S. K., Liu, S., McPherson, J. D., Jungers, K. A., Apte, S.
S., Pavan, W. J., and Beier, D. R. (2003) Development 130, 4665-467243. Murphy, G., and Gavrilovic, J. (1999) Curr Opin Cell Biol 11, 614-62144. Schlondorff, J., and Blobel, C. P. (1999) J Cell Sci 112, 3603-361745. Werb, Z. (1997) Cell 91, 439-44246. Teuchert, M., Berghofer, S., Klenk, H. D., and Garten, W. (1999) J Biol Chem 274,
36781-3678947. Tsuji, A., Sakurai, K., Kiyokage, E., Yamazaki, T., Koide, S., Toida, K., Ishimura, K.,
and Matsuda, Y. (2003) Biochim Biophys Acta 1645, 95-10448. Milewicz, D. M., Grossfield, J., Cao, S. N., Kielty, C., Covitz, W., and Jewett, T. (1995)
J Clin Invest 95, 2373-2378
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
12
FIGURE LEGENDS
Figure 1. Schematic representation and modular structure of preproADAMTS9. A. Thevarious modules illustrated are: S, signal peptide; Pro, propeptide; Cat, catalytic domain; Dis,disintegrin-like module; TSR, thrombospondin type 1 repeat; CRD, cysteine-rich peptide; Gon-1,the C-terminal module homologous to GON-1. L1 and L2 are linkers between TSR arrays. Thefull-length enzyme has 1935 a.a. B. Sequence of the signal peptide (bold), propeptide and startof the catalytic domain (bold). Putative N-linked glycosylation sites and consensus furin-processing sites are shown.
Figure 2. Characterization of propeptide antibodies and absence of intracellular processingin cells transfected with various ADAMTS9 plasmids. The zymogen (Z), processedpropeptide fragments (P) and processed catalytic domain (C) are indicated. A-C. Western blotanalysis of transfected HEK293F cell and conditioned medium (CM) expressing full-lengthADAMTS9 or ADAMTS9-Glu381Ala (E>A). Antibodies used are shown under eachimmunoblot. D. Western blot analysis of HEK293F cells and conditioned medium (CM)expressing ADAMTS9 Pro-Cat using anti-RP4 (left panel) or anti-myc (right panel). Thestructure of Pro-Cat is illustrated above the gels E. Effect of PNGaseF on migration of anti-RP4-reactive bands from CM. F. Western blot analysis of CM, treated or untreated with PNGaseFfrom HEK293F cells expressing Pro-FLAG-Cat. The structure of this construct is shown abovethe gel.
Figure 3. Absence of intracellular processing of ADAMTS9 in cell types other thanHEK293F. The zymogen (Z), processed propeptide (P) and processed catalytic domain (C) areindicated. A. Western blotting of untransfected RCS-LTC cells with anti-RP4. A 22 kDapropeptide fragment is seen only in conditioned medium (CM). In cells, only the intact 180 kDazymogen is detected. B. Analysis of Pro-Cat-transfected COS-1 cells and their conditionedmedium (CM). Antibodies used are shown below each blot. C. Analysis of Pro-Cat-transfectedCHO-K1 cells and their conditioned medium (CM). Antibodies used are shown below each blot.
Figure 4. ADAMTS9 is processed differently from ADAMTS1 and ADAMTS7. A.HEK293F cells transiently transfected with ADAMTS1 or ADAMTS9 Pro-Cat were incubatedwith [35S]Met/Cys for 3 hrs, followed by immunoprecipitation with anti-ADAMTS1 or anti-penta-His, SDS-PAGE and fluorography. Note the presence of both the intact zymogen (Z) andmature ADAMTS1 (M) in the cell lysate, whereas only the ADAMTS9 Pro-Cat zymogen (Z) ispresent in cells and only the catalytic domain (C) in conditioned medium (CM). The 55 kDaADAMTS1 species (D) represent products of C-terminal proteolysis. B. ADAMTS9 Pro-Cat andADAMTS7 Pro-Cat were transfected into HEK293F cells followed by immunoblotting of cellsand conditioned medium (CM) with anti-myc. Note that the activated ADAMTS9 catalyticdomain (C) is present exclusively in the conditioned medium, whereas the ADAMTS7 catalyticdomain (C) is also visible in the cell lysate. An ADAMTS7 processing intermediate (I) is alsoseen in the cell lysate.
Figure 5. ADAMTS9 Pro-Cat is processed at the cell surface. A. HEK293F cells with stableexpression of Pro-Cat were biotinylated as described. Biotinylated proteins were affinity-purifiedusing streptavidin-agarose and analyzed by immunoblotting (IB) with anti-myc antibodies. The
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
13
biotinylated protein disappears on trypsin-EDTA treatment of cells, indicating its location at thecell surface. B. Pro-Cat was metabolically labeled and chased over time as described inExperimental Procedures. Fluorograms of anti-myc precipitated protein from total cell lysate(left panel), lysates of cells treated with trypsin (middle panel) and of conditioned medium (rightpanel) are shown. C. Fate of cell surface biotinylated Pro-Cat in stably transfected HEK293Fcells. Cells were biotinylated for 30 min., washed, returned to fresh medium, and the biotinylatedproteins in the medium and cell lysate were captured with streptavidin-agarose. Capturedproteins were analyzed by western blotting at hourly intervals up to 8 hrs after biotinylation withthe antibodies shown below each immunoblot (IB). D. Pro-Cat was purified from cell lysate oftransfected HEK293F cells as described in Experimental Procedures and incubated withuntransfected HEK293F cells or their cell-free medium for varying periods of time as shownfollowed by western blotting with anti-myc.
Figure 6. PC-dependent processing of proADAMTS9. The zymogen (Z), processedpropeptide fragments (P) and processed catalytic domain (C) are indicated. A. Dose dependentinhibition of Pro-Cat processing by the PC inhibitor dec-RVKR-cmk. Cells were treated with 0-100 nM inhibitor and conditioned medium (CM) was analyzed by western blotting using anti-RP4 or anti-myc. Note decreasing pro-domain and catalytic domain fragments with increasingconcentration of inhibitor. The precise identity of the molecular species migrating between 40-50kDa is unknown, but presumably represents processing intermediates. B. HEK293F cells stablytransfected with Pro-Cat were incubated for 2 hrs with increasing concentrations of α1-PDX asshown. Immunoblotting of conditioned medium showed a dose-dependent reduction of zymogenprocessing. C. Analysis of Pro-Cat processing by furin, PACE4 and PC5A in CHO RPE.40 cells.Pro-Cat was transfected alone or co-transfected with furin or other PCs in furin-deficient CHORPE.40 cells (top panel) or LoVo cells (bottom panel). Western blotting was done with anti-myc. Note that when seen in the CM, the zymogen is consistently 1-3 kDa larger than the mainband in cell lysate, indicating the addition of complex terminal carbohydrate structures. Noticesignificant processing of the zymogen in CM from the furin and PC5A, but not PACE4transfected cells.
Figure 7. Furin and ADAMTS9 Pro-Cat are part of a complex at the cell surface. A.HEK293F cells with stable expression of Pro-Cat were biotinylated as described in Materials andMethods. Biotinylated proteins were affinity-purified using streptavidin-agarose and analyzed byimmunoblotting (IB) with anti-furin. The biotinylated furin disappears on trypsin treatmentindicating its location at the cell surface. B. Furin and Pro-Cat associate at the cell surface ofHEK293F cells. Cells transiently expressing Pro-FLAG-Cat were immunoprecipitated with anti-FLAG after treatment with a reducible, cell-impermeable cross-linking agent andimmunoblotting was done using an anti-furin monoclonal antibody. Immunoprecipitated proteinwas not seen in cells treated with trypsin-EDTA prior to lysis. The bottom panel shows amountsof Pro-Cat in the immunoprecipitated protein of trypsinized or untrypsinized cells. C.Conditioned medium (upper panel) and cell lysate of CHO RPE.40 cells expressing Pro-Cat withor without furin were analyzed by cell surface biotinylation (upper panel) or western blottingwith anti-myc. Co-transfection with furin leads to decreased cell-surface zymogen. The lowerpanel shows comparative levels of Pro-Cat.
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Koo et al, ADAMTS9 processing 3/06
14
Figure 8. Silencing of HEK293F furin expression suppresses ADAMTS9 Pro-Catprocessing. A. Furin siRNA leads to suppression of cellular furin. Western blot of cell lysatewas with anti-furin monoclonal antibody. B. Transfection with furin siRNA does not affect eitherthe total cellular levels of Pro-Cat or the amount located at the cell surface. The panel on the leftshows the total cellular Pro-Cat. The panel at right shows cell-surface Pro-Cat labeled bybiotinylation C. Expression of furin siRNA suppresses processing of ADAMTS9 Pro-Cat. Notethat cellsw ith suppressed furin levels generate mostly the 55kDa zymogen and little of theprocessed propeptide and catalytic domain.
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Pro Cat Dis TSR1 CRD SpacerTSR
Gon-1L1 L2N S
Fig. 1. (Koo et al)
A
MQFVSWATLLTLLVRDLAEMGSPDAAAAVRKDRLHPRQVKLLETLSEYEIVSPIRVNA
LGEPFPTNVHFKRTRRSINSATDPWPAFASSSSSSTSPQAHYRLSAFGQQFLFNLTAN
AGFIAPLFTVTLLGTPGVNQTKFYSEEEAELKHCFYKGYVNTNSEHTAVISLCSGMLGT
FRSHDGGYFIEPLQSMDEQEDEEEQNKPHIIYRRSAPQREPSTGRHACDTSEHKNRH
SKDKKKTRARKWGERINLAGDVAALNSGLATEAFSAYGNKTDNTREKRTHRRTKRFL
SYPRFVEVLVV-----
B
(Signal peptide)
(Catalytic domain) Putative furin cleavage site Putative N-glycosylation site
TSRTSR
58
289
232
175
116
33
287
74
280 283
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

25
37
5075
2015
- - + - +PNGaseF
B
IB: Anti-RP4
ACM Cell
IB: Anti-RP4
CADAMTS9 - + + + +
(E>A)
- - +PNGaseFADAMTS9 - + +
ADAMTS9CM Cell
- + +-250
25
37
50
75100150
20
25
37
5075
20
15
IB: Anti-RP1
Fig. 2. (Koo et al)
kDakDa kDaZ
P
P
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

E
IB: Anti-RP4
Pro CatN S Myc/
D
IB: Anti-RP4
- + - +PNGaseF
ADAMTS9+ + - -
+Pro-Cat
- +-CM Cell
Pro-CatCM Cell
- + +-
25
37
50
75
20
15
25
37
5075
20
15
IB: Anti-myc
25
37
50
75
20
CM Cell CM Cell- + +-
His
Fig. 2. (Koo et al)
kDa kDa kDa
Z
P
P
C
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FT276-FLAG-R277
IB: Anti-RP4 Anti-FLAG
25
37
20
15
- + - +PNGaseF+ + - +Pro-FLAG-Cat
-+
--
Pro CatN S Myc/His
Fig. 2. (Koo et al)
kDa
10
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

25
37
50
75
20
IB: Anti-RP4 IB: Anti-myc
CM CellPro-Cat
CM Cell- + +-
25
37
50
75
20
COS-1
25
37
50
75
20
CHO-K1
IB: Anti-RP4 IB: Anti-myc
BA
250
25
37
50
75100150
20
RCS-LTC
CM Cell
IB: Anti-RP4
25
37
50
75
20
C
CM Cell CM Cell- + +-
Pro-Cat - + +-CM Cell CM Cell
- + +-CM Cell CM Cell
Fig. 3. (Koo et al)
kDa kDa kDa
kDakDa
Z
P
Z
P
C
Z
PC
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

ADAMTS1
Cell CM
37
50
75100
2520
25
37
50
75
20
IP:
ADAMTS9-
Pro-Cat
Cell CM Cell CM
ADAMTS9-
Pro-Cat
Cell CM
ADAMTS7-
Pro-Cat
IB: Anti-myc
A B
Fig. 4. (Koo et al)
kDakDa
ZM
D Z
C
Z
C
I
Anti-ADAMTS1
Anti-penta-His
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Fig. 5. (Koo et al)
A
IB: anti-myc
Trypsin/EDTABiotinylation
Pro-Cat- -
+-++
- ++
Z
37
50
75
25
100
20
kDa by guest on Septem
ber 16, 2018http://w
ww
.jbc.org/D
ownloaded from
0 15 1206030 0 15 1206030 Chase (min)Cell Cell + Trypsin
0 15 1206030CM
25
37
50
75 kDa
B
IP: Anti-myc IP: Anti-myc IP: Anti-myc
Z
C
C
IB: Anti-RP4 IB: Anti-myc
Media Lysate0 6421 8
25
37
5075
20
kDa
P
PC
0 6421 80 6421 8
Z
IB: Anti-myc
Fig. 5. (Koo et al)
Chase (hr)
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

(hr)0 6421 8
Cell Cell-free
Z
C
IB: Anti-myc
0 6421 8
25
37
50
75
20
Fig. 5. (Koo et al)
kDa
D
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

A
0 1005025
10 (µM)
25
37
5075
20
IB: Anti-RP4
IB: Anti-myc
Dec-RVKR
0 1005025
10 (µM)
25
37
5075
20
Dec-RVKR
Fig.6. (Koo et al)
kDa
kDa
Z
P
P
C
Z
25
37
5075
20
kDa
(µg/ml)α1-PDX 0 210.5
25
37
5075
20
kDa
(µg/ml)α1-PDX 0 210.5
B
IB: Anti-RP4
IB: Anti-myc
P
P
C
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
C
CM
- - + + + + + + +
Furin PACE4PC5A
25
37
5075
Pro-Cat
Cell CellCellCellCell CMCMCMCM kDa
C
Z
20
+
Pro-Cat - + + ++Furin PACE4
PC5A
(CM)
25
37
50
75kDa
20
C
Z
CHO RPE40
LoVo
IB: Anti-myc
IB: Anti-myc
Fig.6. (Koo et al)
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

A
Trypsin/EDTA
IB: anti-furin
Biotinylation- -
+-++
Pro-Cat - ++
B
- +-- ++
Trypsin/EDTAPro-FLAG-Cat
IB: anti-furin
IB: anti-RP4
IP: anti-FLAG
Furin
IgGheavychain37
50
75
25
100
20
50
75
100
37
150
5075
kDa kDa
Pro-Cat
25
Furin
Fig.7. (Koo et al)
C
Pro-Cat- +
Biotinylation
IB: Anti-myc
Furin- + +
-
Z
37
50
75
2520
kDa
50
75Cell
CHO RPE.40
Z
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

50
75
100
150kDa
IB: anti-furin
Furin siRNA - +
Furin
CellA
37
50
75kDa
IB: anti-myc
Furin siRNA - +
Cell
Z
- +
BiotinylationB
Furin siRNA - + - +
Z
C25
37
50
75
20
CM
IB: anti-myc IB: anti-RP4
C
kDa
Fig.8. (Koo et al)
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Richard Leduc and Suneel S. ApteBon-Hun Koo, Jean-Michel Longpre, Robert P. T. Somerville, J. Preston Alexander,
Cell-surface processing of pro-adams9 by furin
published online March 14, 2006J. Biol. Chem.
10.1074/jbc.M511083200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from