cellentryofarginine-richpeptidesisindependent … · cellentryofarginine-richpeptidesisindependent...
TRANSCRIPT

Cell Entry of Arginine-rich Peptides Is Independentof Endocytosis*□S
Received for publication, July 21, 2008, and in revised form, November 11, 2008 Published, JBC Papers in Press, December 1, 2008, DOI 10.1074/jbc.M805550200
Gohar Ter-Avetisyan‡, Gisela Tunnemann‡, Danny Nowak‡, Matthias Nitschke§, Andreas Herrmann§, Marek Drab¶,and M. Cristina Cardoso‡�1
From the ‡Max Delbrueck Center for Molecular Medicine, D-13125 Berlin, the §Department of Biology/Biophysics, HumboldtUniversity Berlin, D-10115 Berlin, the ¶Department of Molecular Biology, Max Planck Institute for Infection Biology, D-10117 Berlin,and the �Department of Biology, Technische Universitat Darmstadt, D-64287 Darmstadt, Germany
Arginine-rich peptides are a subclass of cell-penetrating pep-tides that are taken up by living cells and can be detected freelydiffusing inside the cytoplasm and nucleoplasm. This phenom-enon has been attributed to either an endocytic mode of uptakeand a subsequent release from vesicles or to direct membranepenetration (transduction). To distinguish between both possi-bilities, we have blocked endocytic pathways suggested to beinvolved in uptake of cell-penetrating peptides. We have thenmonitored by confocal microscopy the uptake and distributionof the cell-penetrating transactivator of transcription (TAT)peptide into living mammalian cells over time. To prevent sideeffects of chemical inhibitors, we used genetically engineeredcells as well as different temperature. We found that a knock-down of clathrin-mediated endocytosis and a knock-out ofcaveolin-mediated endocytosis did not affect the ability of TATto enter cells. In addition, the TAT peptide showed the sameintracellular distribution throughout the cytoplasmandnucleusas in control cells. Even incubation of cells at 4 °C did not abro-gate TAT uptake nor change its intracellular distribution. Wetherefore conclude that this distribution results from TAT pep-tide that directly penetrated (transduced) the plasma mem-brane. The formation of nonselective pores is unlikely, becausesimultaneously added fluorophores were not taken up togetherwith the TAT peptide. In summary, although the frequency andkinetics of TAT transduction varied between cell types, it wasindependent of endocytosis.
Thediscovery that the transactivator of transcription (TAT)2protein of human immunodeficiency virus type 1 was able to
traverse cellular membranes and subsequently affected genetranscription (1, 2) led to the emergence of a new research fieldon cell-penetrating peptides (CPPs), also known as proteintransduction domains (PTDs) or membrane transduction pep-tides (3). CPPs opened up the possibility to effectively delivercell-impermeable hydrophilic compounds into living cells. Thecargos reported to be shuttled to intracellular compartmentsinclude drugs (4), fluorophores (5), peptides (6–8), nucleicacids (9), proteins (10–12), nanoparticles (13), and liposomes(14, 15). The exact mechanism of cellular entry of CPPsremained unknown, but it was thought to be receptor-, energy-,and temperature-independent. In 2003 this unique mode ofuptake was refuted as a methodological artifact, and endocyto-sis was suggested as the main pathway of cellular uptake ofCPPs in live cells (16, 17). Arginine-rich peptides (RRPs) werenot only historically the first (TAT) (1, 2) type of CPPsdescribed, but they combined high uptake ability with moder-ate toxicity (18). Some groups observed a nonendocytic inter-nalization pathway (8, 18–21), whereas others assigned CPPuptake to endocytic pathways, as CPPs were internalized andstored inside the vesicles. Endocytosis is broadly subdividedinto phagocytosis and pinocytosis. Whereas phagocytosis isrestricted to specialized cells like macrophages and leukocytes,pinocytosis occurs in all eukaryotic (or mammalian) cell typesthrough at least four different endocytic pathways (22). Threeof them have been implicated as routes for internalizationdepending on the CPP sequence and cargo of the CPPs as fol-lows: clathrin-mediated endocytosis (23), caveolae-mediatedendocytosis (24, 25), macropinocytosis (26–28), as well as theinvolvement of more than one endocytic pathway (16, 19).However, most of the studies so far utilized chemical inhibitorsto characterize the contribution of a distinct endocytic pathwayand could not exclude inhibitor-associated side effects.RRPs such as TAT linked to high molecular weight cargos
(e.g. proteins) were taken up into cells solely by endocytosis.When conjugated to lowmolecularweight cargos (e.g.peptides)however, in addition to be present in vesicles, they could befound freely diffusing in the cytoplasm and the nuclear com-partment (8). However, a consensus regarding the latter uptakemode has not been reached. Our translocation studies of oli-goarginines and oligolysines of various chain lengths and con-centrations into living cells demonstrated the coexistence oftwo uptake modes (8, 18). Whereas a subset of the intracellularpeptidewas found inside cytoplasmic vesicles (Fig. 1A), some ofthe peptide displayed a rather homogeneous distribution
* This work was supported by grants from the Deutsche Forschungsgemein-schaft and the Volkswagen Foundation (to M. C. C.). The costs of publica-tion of this article were defrayed in part by the payment of page charges.This article must therefore be hereby marked “advertisement” in accord-ance with 18 U.S.C. Section 1734 solely to indicate this fact.Author’s Choice—Final version full access.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Figs. S1–S5 and Movies 1–10.
1 To whom correspondence should be addressed. Tel.: 49-6151-16-2377; Fax:49-6151-16-2375; E-mail: [email protected].
2 The abbreviations used are: TAT, transactivator of transcription; BHK, babyhamster kidney; cav, caveolin; CHC, clathrin heavy chain; CPP, cell-pene-trating peptide; DMEM, Dulbecco’s modified Eagle’s medium; FCS, fetalcalf serum; FITC, fluorescein isothiocyanate; HIV, human immunodefi-ciency virus; KO, knockout; LMW, low molecular weight; PTD, protein trans-duction domain; RRP, arginine-rich peptide; TAMRA, 5,6-carboxytetram-ethylrhodamine; TAT-bt-SAv, TAT-biotin-streptavidin complex; tet,tetracycline; Tf, transferrin; tTA, transcriptional activator; WT, wild type.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 284, NO. 6, pp. 3370 –3378, February 6, 2009Author’s Choice © 2009 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
3370 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 6 • FEBRUARY 6, 2009
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

throughout the cytoplasm and high accumulation inside thenucleolar compartment (Fig. 1, B and C). The latter is hence-forth termed transduction. It is still unclear whether transduc-tion reflects CPPs that enter living cells by a not yet definedmechanism and/or CPPs that are released from endo- or lyso-somes after endocytosis. To understand if both intracellularphenotypes are two distinct intermediate stages of the sameprocess or indicate different uptake routes, we monitored thecytoplasmic and nucleolar localization of RRPs upon inhibitionof endocytosis. In addition to TAT, we used the peptide PTD4,which is an artificial, more amphipathic CPP with a reducednumber of arginines and increased �-helical content comparedwith TAT (29). Most importantly, to suppress endocytic path-ways, we restricted ourselves to the usage of genetically induc-ible systems, knock-out (KO) cell cultures, or physicalmethods,thus avoiding any potential side effects of chemical inhibitors ofendocytosis.
EXPERIMENTAL PROCEDURES
Cells andCultureConditions—The following cells were used:BHK21 (C-13) fibroblasts (baby hamster kidney clone 13,American Type Culture Collection), BHK21-tTA/anti-clathrinheavy chain (CHC) cell line (30), cav-1-KO and cav-1-wtmouseendothelioma cell lines (31), C2C12mouse myoblasts (32), and3T3 mouse fibroblasts (Invitrogen). The cells were cultured inDulbecco’s modified Eagle’s medium (high glucose, withsodium pyruvate and L-glutamine) (PAA, Pasching, Austria)supplemented with 10 or 20% (the latter for C2C12 cells only)fetal calf serum (PAA, Pasching, Austria), 2 mM L-glutamine(Invitrogen), 50 �g/ml gentamicin (PAA, Pasching, Austria).For the cav-1-KO and cav-1 wild type (WT) endothelioma celllines, the growthmedia was additionally supplementedwith 1%nonessential amino acids (Sigma), 1% sodium pyruvate(Invitrogen), and 2 mM 2-mercaptoethanol diluted in phos-phate-buffered saline (Invitrogen). For the BHK21-tTA/anti-
CHC cell line, the following additives were added: 10% tetracy-cline-negative fetal calf serum (PAA, Pasching, Austria), 0.2mg/ml geneticin (G418, Invitrogen), 0.5 �g/ml puromycin(Sigma), 2 �g/ml tetracycline (Sigma). For the induction ofCHC antisense RNA expression, tetracycline was removedfrom the medium.Peptides, Proteins, and Fluorophores—Peptides 5,6-car-
boxytetramethylrhodamine (TAMRA)-TAT (GRKKRRQRRR)and TAMRA-PTD4 (YARAARQARA) (29) were synthesized asD-isomers and coupled directly to TAMRA at the N terminus(Peptide Specialty Laboratories GmbH, Heidelberg, Germany).Peptides and labelswere diluted inRPMI 1640mediumwithoutphenol red (PAA, Pasching, Austria) and applied at 10 �M finalconcentration to the cells. At lower concentrations, as we haveshown previously (8), only the endocytic mode of uptake wasdetected (supplemental Fig. S1). To monitor CHC expressionand function, transferrin (Tf) conjugated to Alexa Fluor 633(Invitrogen) was added as a marker for clathrin-dependentendocytosis. Nonreactive forms of the fluorophores FITC (flu-orescein isothiocyanate) and TAMRA were generated by incu-bation with Tris buffer (indicated by an asterisk) and used as asmall molecule to monitor the generation of pores during thetransduction event. To control for the complete inhibition ofendocytosis at 4 °C, the globular protein TAT-biotin-streptavi-din (TAT-bt-SAv) labeled with the fluorophore cyanine dyewas used as an additional marker for fluid-phase uptake (sup-plemental Fig. S2).Live Cell Uptake Experiments—For all experiments either
8-well �-slides or 35-mm �-dishes (Ibidi, Martinsried, Ger-many) were used. The cells were seeded onto the observationchambers and incubated overnight at 37 °C with 5% CO2. Thepeptides TAMRA-TAT and TAMRA-PTD4 (10 �M), the flu-orophores FITC* and TAMRA* (10 �M), and Tf (10 �g/ml)were diluted in RPMI 1640 medium (PAA, Pasching, Austria)without phenol red and used at the indicated concentrations.Special care was taken to ensure that the volume of the peptidesolution above the cells was comparable in the two differentobservation chambers and that the exchange against the appro-priate peptide (label, marker) dilutions was performed verygently. Immediately after addition of the peptide (label,marker)to the cells, time lapses over 60 min (with time intervals of oneimage per min) were recorded. The experimental settings forthe confocal microscope were identical for all experiments.For the uptake experiments at 4 °C a custom-built cooling
chamber was used. The height of this cooling chamber occupy-ing the cooling flow and the radius of the loophole at the bottomwere optimized to guarantee a constant temperature exchangebetween the 35-mm �-dish observation chamber and the coolwater flux. The temperature was regulated by a thermostat, andthe exact temperature of 4 °C inside themedium above the cellswas verified by measurements with a thermometer before andafter the 1-h time lapses.Microscopy, Image Acquisition, and Analysis—Confocal
optical sections were acquired with a Zeiss confocal laser scan-ning microscope, LSM510 Meta, mounted on an Axiovert 200M inverted microscope equipped with a live cell microscopeincubation cage (Okolab, Ottaviano, Italy) using a 63� plan-apochromat NA1.4 oil immersion, phase-contrast objective.
FIGURE 1. Schematic illustration (upper panel), corresponding represent-ative cell images, and relative proportions of the observed intracellulardistributions of TAT CPP in different cell types after 1 h of incubation.A, TAT peptide is present in the medium and in endocytic vesicles, but notfreely available inside the cytoplasm or the nuclear compartment. B, TAT pep-tide reached all intracellular compartments and accumulated, in addition, invesicles. C, TAT peptide is homogeneously distributed throughout the cyto-plasm and accumulated in the nucleolus. Scale bar, 10 �m.
TAT Transduction Occurs in the Absence of Endocytosis
FEBRUARY 6, 2009 • VOLUME 284 • NUMBER 6 JOURNAL OF BIOLOGICAL CHEMISTRY 3371
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

The microscope incubation cage maintained a humidifiedatmosphere of 5% CO2 and 37 °C, which was used throughoutexcept for the low temperature experiments. For all settings themain beam splitter was HFT UV/488/543/633, and the specificparameters for the single fluorophores were as follows: FITCexcited at 488 nm, detectedwith a 500–530-nmbandpass filter;TAMRA-TAT and TAMRA-fluorophore excited at 543 nm,detected with a 565–615-nm bandpass filter, transferrin-AlexaFluor 633 excited with 633 nm, detected with a 650-nm longpass filter. Phase contrast images were recordedwith excitationat 488 nm and detection in the transmission channel. The laserpower for observation was 1% (488 nm, 25 milliwatts), 7% (543nm, 1 milliwatt), and 25% (633 nm, 5 milliwatts). Settings wereadjusted in a way that image pixels were not over- or underex-posed with the range indicator function in the Zeiss laser scan-ning microscopy software version 3.2. To ensure that weakintracellular fluorescence signals of the peptides were notmissed, a set of overexposed images was additionally collected.For the quantification of transduction, 100–150 cells per
transduction experiment were evaluated to obtain the percent-age of transduced cells (indicated by nucleolar appearance ofthe labeled peptide), and the kinetics of TAT transduction wasfurther characterized by the earliest time point when transduc-tion could be detected within a field of view (initiation time oftransduction).Western Blotting—ForWestern blot analysis of the cav-1-KO
andWT cells, half a million cells were counted, resuspended in100 �l of phosphate-buffered saline, and boiled in Laemmlisample buffer for 10 min, and cell lysates were analyzed bySDS-PAGE followed by blotting onto polyvinylidene difluoridemembranes. Signals were detected with the following primaryantibodies: rabbit anti-Cav-1 polyclonal antibody (LifeSpanBioSciences, Inc., Seattle,WA) andmouse anti-Cav-2 and anti-Cav-3 monoclonal antibodies (1:2000 dilution, BD Transduc-tion Laboratories). Anti-rabbit IgG-HRP (Sigma) and anti-mouse IgG-HRP (enhanced chemiluminescence, AmershamBiosciences) were used as secondary antibodies. Immunoreac-tive signalswere visualized using enhanced chemiluminescenceplus detection solution (Amersham Biosciences) and recordedusing a luminescence imager (Luminescent Image AnalyzerLAS-1000, FUJI Photo Film, Tokyo, Japan).
RESULTS
Because of their particularly high transduction ability, whichsolely depends on a minimal number of arginines, RRPs play aspecial role among CPPs (18, 33–35). To clarify the role ofendocytosis in the uptake mode of CPPs with low molecularweight (LMW) cargos into living cells, we investigated theoccurrence and extent of transduction of TAT and PTD4 asCPPs with high and low transduction frequency, respectively(8). The intracellular distribution of peptides in living cells wasanalyzed by laser scanning confocalmicroscopy. To unravel therelevance of endocytic routes for the uptake and intracellulardistribution of peptides, endocytic pathways were specificallyinhibited by genetic approaches orwere blocked in ensemble byincubation of cells at low temperature.Role of Clathrin-mediated Endocytosis in CPP Uptake—
Clathrin-dependent endocytosis represents a major endocytic
pathway. For example, transferrin is taken up exclusively by thisroute, and several enveloped viruses (36), such as equine arte-ritis virus (37), exploit this route (38). As an early step of thisroute, upon binding of an extracellular ligand to specific cell-surface receptors, clathrin together with other adapter proteinsbuilds an endocytic coat at the plasmamembrane (Fig. 2A). Thecoated membrane buds and pinches off to form a cargo-filledvesicle (Fig. 2A).Clathrin-dependent endocytic uptake of TAT has been
repeatedly reported as a possible mechanism for CPP entry (23,39, 40). To clarify the contribution of clathrin-dependent endo-cytosis in the uptake mode of arginine-rich CPPs, we used theBHK21-tTA/anti-CHC (30) cell line. This cell line expressesantisense CHC RNA under the control of a tetracycline-re-sponsive element (Fig. 2B). More specifically, the transcriptionactivator (tTA) is composed of the DNA binding domain oftetracycline repressor protein and a C-terminal activationdomain of VP16 (herpes simplex virus protein) that functionsas a strong transcription activator (41). The presence of tetra-cycline prevents binding of the transactivator tTA to the oper-ator sequence and thus transcription of antisense RNA. Inthe absence of tetracycline the transactivator tTA binds to itsoperator sequence and activates the transcription of antisenseRNA.As a consequence, the synthesis and functionality ofCHCprotein is significantly reduced, thereby suppressing clathrin-dependent endocytosis (30). The absence of tetracycline for 2days was reported to inhibit 90% of transferrin internalization,and the expression of theCHCproteinwas reduced to 10% over6 days in the absence of tetracycline (30). Therefore, to exploreto what extent transduction of TAT depends on clathrin-de-pendent endocytosis, uptake of TAMRA-tagged TAT andPTD4 was investigated in the presence (�tet) and absence(�tet) of tetracycline over 6 days. To simultaneously controlthe level of clathrin-dependent endocytosis, internalization ofAlexa Fluor 633-labeled transferrin was monitored (Fig. 2C).The control cells (�tet cells) are shown on the left panel of
Fig. 2C and supplemental Fig. S3. TAT was homogeneouslydistributed throughout the cytoplasm and accumulated in thenucleolus (Fig. 2C, arrowheads) and therefore displayed theuptake mode of transduction. In addition, the labeled peptidewas also present in cytoplasmic vesicles.PTD4 applied at the same concentration and monitored at
identical confocal microscope settings did not show any trans-duction-associated cytoplasmic localization comparable withTAT. However, we observed the presence of PTD4-containingvesicles (Fig. 2C, left side, lower panel). Fluorescent transferrinwas internalized at high rates and enriched in the trans-Golginetwork (42).Inhibition of the clathrin-dependent endocytic pathway in
�tet cells (Fig. 2C and supplemental Fig. S3A, right panel) wasverified by the suppression of uptake of transferrin. TAT dis-played the same intracellular distribution regarding the vesicu-lar uptake as well as transduction, indicated by similar intensi-ties inside the nucleolar compartment compared with thecontrol (�tet) cells (Fig. 2C and supplemental Fig. S3). Similarto the control cells, no diffuse intracellular occurrence wasobserved for PTD4, but in contrast to �tet cells the vesicularinternalization of PTD4 was almost completely abolished.
TAT Transduction Occurs in the Absence of Endocytosis
3372 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 6 • FEBRUARY 6, 2009
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from
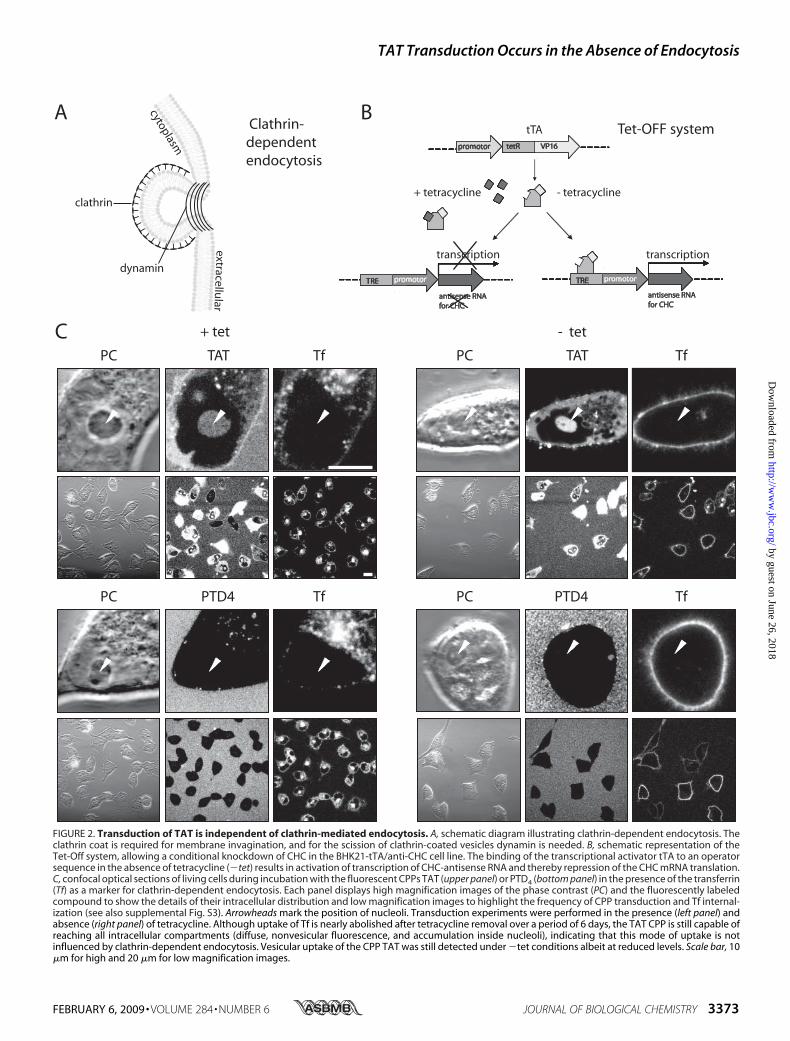
FIGURE 2. Transduction of TAT is independent of clathrin-mediated endocytosis. A, schematic diagram illustrating clathrin-dependent endocytosis. Theclathrin coat is required for membrane invagination, and for the scission of clathrin-coated vesicles dynamin is needed. B, schematic representation of theTet-Off system, allowing a conditional knockdown of CHC in the BHK21-tTA/anti-CHC cell line. The binding of the transcriptional activator tTA to an operatorsequence in the absence of tetracycline (�tet) results in activation of transcription of CHC-antisense RNA and thereby repression of the CHC mRNA translation.C, confocal optical sections of living cells during incubation with the fluorescent CPPs TAT (upper panel) or PTD4 (bottom panel) in the presence of the transferrin(Tf) as a marker for clathrin-dependent endocytosis. Each panel displays high magnification images of the phase contrast (PC) and the fluorescently labeledcompound to show the details of their intracellular distribution and low magnification images to highlight the frequency of CPP transduction and Tf internal-ization (see also supplemental Fig. S3). Arrowheads mark the position of nucleoli. Transduction experiments were performed in the presence (left panel) andabsence (right panel) of tetracycline. Although uptake of Tf is nearly abolished after tetracycline removal over a period of 6 days, the TAT CPP is still capable ofreaching all intracellular compartments (diffuse, nonvesicular fluorescence, and accumulation inside nucleoli), indicating that this mode of uptake is notinfluenced by clathrin-dependent endocytosis. Vesicular uptake of the CPP TAT was still detected under �tet conditions albeit at reduced levels. Scale bar, 10�m for high and 20 �m for low magnification images.
TAT Transduction Occurs in the Absence of Endocytosis
FEBRUARY 6, 2009 • VOLUME 284 • NUMBER 6 JOURNAL OF BIOLOGICAL CHEMISTRY 3373
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

We confirmed that TAT transduction was observed in mostcells by acquiring low magnification images (Fig. 2C and sup-plemental Fig. S3). To control for any potential side effects oftetracycline, the parental BHK21 cells (43, 44) were incubatedwith the CPPs and transferrin in the presence and absence oftetracycline. However, no difference of peptide transductionand vesicle formation was observed (data not shown). There-fore, we conclude that clathrin-dependent endocytosis is notrequired for transduction of the RRP TAT fused to an LMWcargo.Role of Caveolin-mediated Endocytosis in CPP Uptake—Be-
sides the classical clathrin-mediated endocytic pathway, caveo-lae-mediated endocytosis is one of the main endocytic entryroutes into living cells (22, 45). For example, it is exploited bybacterial toxins and by simian virus 40 (36). Caveolae are flask-shaped, small (50–70 nmdiameter) invaginations in the plasmamembrane (Fig. 3A) that constitute membrane domainsenriched in cholesterol and sphingolipids, called lipid rafts (46).Caveolae are characterized by the presence of the integralmembrane protein caveolin-1 and are involved in the intracel-lular transport of lipid raft-associated molecules (47). Thispathway has been repeatedly reported as an uptake route forCPPs into the cells (24, 25). Former studies used fluorescentlylabeled �-subunit of cholera toxin as a marker to monitorcaveolar uptake. However, the pathway chosen by cholera toxinsubunit � depends on the cell type (48) and hence may not be afaithful indicator for a single internalization pathway. To spe-cifically inhibit only the caveolin-dependent route and to pre-vent the potential side effects caused by chemical inhibitors ofendocytosis, we made use of an endothelial heart cell line gen-erated from a knock-out (KO) mouse deficient for caveolin(cav-1) and the respective wild type (WT) cell line. As reportedpreviously (31), in the absence of caveolin-1, caveolin-2 proteinis degraded. This was corroborated by Western blot analysis(Fig. 3B). In contrast to WT cells, no cav-1 and cav-2 weredetected in the KO cells, and in addition, antibodies directedagainst muscle cell-specific cav-3 gave a much weaker signal inthe extract of KO cells compared with those from wild typecells.To elucidate if caveolae-dependent endocytosis plays a role
in the uptakemode of TATwith the LMWTAMRA,we appliedTAT (Fig. 3C, upper panel, and supplemental Fig. S4) and PTD4(Fig. 3C, middle panel) to the medium of wild type cells andcav-1-KO cells. To control whether the tagged fluorophoresupports peptide internalization, the uptake of TAMRA* aloneat the same concentration as the peptides was studied (Fig. 3C,bottom panel). For each experiment confocal optical sectionsare displayed at high and low magnification. Because of thelarge size of these cells, it was not possible to show a highernumber of cells per field and at the same time keep imagingparameters constant throughout all experiments. No differenceof TAT transduction between WT and cav-1-KO cells wasfound. TAT also became internalized by an endocytic route, asindicated by the punctated intracellular fluorescence. This sig-nalwas still present in the caveolin-deficient cells. In contrast toTAT, the PTD4 peptide entered the cells only by the endocyticmode and could not be detected freely inside the cyto- andnucleoplasm and the nucleolus (Fig. 3C, middle panel, arrow-
FIGURE 3. Transduction of TAT is independent of caveolin-mediatedendocytosis. A, schematic diagram illustrating structural features of flask-shaped caveolae, which are lined by caveolin. Caveolae-mediated endocyto-sis is driven by a coat made of the integral membrane proteins caveolin-1, -2,or -3. Dynamin is required for the scission of caveolae. B, Western blot analysisof the integral membrane proteins caveolin (cav) -1, -2, and -3 in WT andcaveolin 1 KO endothelioma cells. �-Tubulin is used to control for loading.C, confocal optical sections of living cells during incubation with the fluores-cent CPPs TAT (upper panel) or PTD4 (mid panel) and the fluorophore TAMRA*(bottom panel). Each panel displays images of the phase contrast (PC) and thepeptide or fluorophore fluorescence at high magnification to display theirintracellular distribution and at low magnification to highlight the frequencyof CPP transduction in WT and cav-1-KO cells (see also supplemental Fig. S4).Whereas the amphipathic control peptide PTD4 and the fluorophore TAMRA*were not transduced, the CPP TAT was homogeneously distributed in thecytoplasm, and it reached the nucleus where it accumulated inside the nucle-olar compartment (marked by arrowheads). Both CPPs displayed vesicularuptake in WT and cav-1-KO cells. Scale bar, 10 �m for high and 20 �m for lowmagnification images.
TAT Transduction Occurs in the Absence of Endocytosis
3374 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 6 • FEBRUARY 6, 2009
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

heads). TAMRA was excluded from the cytoplasm and intra-cellular compartments providing strong evidence that peptideuptake is not aided by the fluorophore. Based on these results,we conclude that caveolae-mediated endocytosis is notinvolved in the uptake mode of transduction of TAT conju-gated to an LMW cargo.Uptake of CPPs upon Shutting Off Endocytic Pathways—Be-
cause neither clathrin- nor caveolin-dependent endocytosisinhibits CPP uptake and in both cases we could still find TAT-containing vesicles concomitantly with freely available cyto-
plasmic TAT peptide, we tried nextto inhibit all endocytic pathwayssimultaneously.For this purpose, we followed the
internalization of TAT and PTD4 atlow temperature, 4 °C. Fig. 4Adepicts all potential endocyticuptake routes in cells that areexpected to be inhibited undersuch conditions. Previous reportsaddressing the uptake of arginine-rich CPPs with LMW cargos intoliving cells are inconsistent. Al-though some reports insisted on theinability of CPPs to penetrate cellsat low temperature and hence endo-cytosis would be required for inter-nalization (49), according to otherreports, CPP uptake is not inhibitedat 4 °C (14, 19, 20). Fig. 4B displaysthe experimental setup for assessingthe transduction ability of TAT andPTD4 in C2C12 mouse myoblasts(data not shown) and in BHK21hamster fibroblasts in the presence(37 °C) and absence (4 °C) of endo-cytosis by time-lapse confocalmicroscopy. To make sure thatpotential membrane lesions gener-ated by low temperature conditionswould not corrupt the transductionassay, the fluorophore FITC* wasapplied simultaneously with theTAMRA-labeled peptides to thecells. In case of severe membranedamage or pore formation inducedby the peptides, the 389-Da-sizedFITC* molecule should also bedetectable intracellularly. Ourobservations displayed as high andlow magnification images in Fig. 4Cand supplemental Fig. S5 reveal thateven at 4 °CTAT entered living cellsand distributed over the cytoplasmand nucleus, where it accumulatedinside nucleoli. At the same time thefluorophore FITC* did not gainaccess to intracellular compart-
ments indicating that the plasma membrane was not compro-mised. The complete obstruction of endocytosis was furtherconfirmed by the absence of fluorescently labeled vesicles at4 °C.Vesicleswere not observed for theCPPTATnor for PTD4.Furthermore, uptake of the globular TAT fusion protein TAT-bt-SAv that is restricted to endocytosis (8) was also blocked onthe level of the plasma membrane at 4 °C (supplemental Fig.S2). These results prove unambiguously that arginine-richCPPs like TAT are capable of reaching intracellular compart-ments of living cells by a mechanism that is independent of
FIGURE 4. Transduction of TAT is independent of endocytosis. A, schematic overview of the different path-ways of endocytic internalization that are suppressed at 4 °C: clathrin- and caveolin-dependent endocytosis,clathrin- and caveolin-independent endocytosis, and macropinocytosis. B, experimental strategy for transduc-tion experiments performed at 37 and 4 °C. C, confocal optical sections of living cells during incubation with thefluorescent CPPs TAT (upper panel) or PTD4 (lower panel) in the presence of the fluorophore FITC* as a smallmolecule marker to control for membrane pores or damage. Each panel displays high magnification images ofthe phase contrast (PC) and the fluorescently labeled peptide or fluorophore to show their uptake and intra-cellular distribution and low magnification images to highlight the frequency of transduction at 37 and 4 °C(see also supplemental Fig. S5). Arrowheads mark the position of nucleoli. The transduction experiments wereperformed in BHK21 cells kept at 37 and 4 °C. Although no intracellular vesicles were found at 4 °C, the trans-duction of TAT (nonvesicular, diffuse fluorescence with accumulation inside nucleoli) remained unchangedboth at 37 and 4 °C. In contrast, the amphipathic control peptide PTD4 and fluorophore FITC* were not trans-duced at 37 °C nor at 4 °C. Scale bars are 10 �m for high and 20 �m for low magnification images.
TAT Transduction Occurs in the Absence of Endocytosis
FEBRUARY 6, 2009 • VOLUME 284 • NUMBER 6 JOURNAL OF BIOLOGICAL CHEMISTRY 3375
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

endocytosis. However, the data also clearly show that aminimalnumber of arginines is crucial to permit transduction.Frequency and Initiation Time of TAT Transduction—To
quantify our results on the transduction ability in the absence ofa distinct pathway or all pathways of endocytosis, we evaluatedthe percentages of transduced cells after 60 min of incubation
with theCPPTATand the initiationtime of transduction (Fig. 5 andsupplemental movies S1–S10). Thecontrol cells (�tet, day 0) showed atransduction frequency of 83%.While over the first 3 days afterinduction of the conditional knock-down of CHC (�tet) the transduc-tion percentages were reduced to68%, they recovered back to 84%transduction frequencies by day 6,although the internalization offluorescently labeled transferrinwas reduced by 90%. The fre-quency of transduced cells wasidentical in cav-1-deficient andisogenic WT cells (54%), althoughin these cells it was lower than inthe BHK21-tTA/anti-CHC cells.Finally, the inhibition of all poten-tial pinocytic pathways at 4 °Cdetermined in BHK21 fibroblastsrevealed that the transduction ofthe TAT peptide is diminishedabout 12% in comparison withtransduction occurring in cellskept at 37 °C, but still 63% of thecells displayed nucleolar accumu-lation of TAT.In BHK21-tTA/anti-CHC cells,
the transduction mode of uptakeinitiated between 15 and 30 min ofpeptide addition after CHC knock-down as well as in control cells. Thekinetics of peptide uptake in cav-1-KO and wild type cells displayedan uptake initiation at 45 min afterapplication of TAT. The fastestuptake of TAT occurred in BHK21fibroblasts within 1 min of additionto the medium and independent oftemperature. A summary of theuptake kinetics ofTATpeptide in allconditions tested is presented inFig. 5.In conclusion, the frequency and
initiation time of TAT transductionwas unchanged within a given celltype independent of endocytosis.However, both parameters were celltype-specific, suggesting that themembrane composition influences
the velocity of transduction.
DISCUSSION
Despite the controversy and uncertainty regarding theuptake mechanism, the property of CPPs to deliver nonperme-ablemolecules into living cellsmakes them attractive vectors to
15min 30min 45min 60min
+ te
t- t
et,1
day
- tet
,2d
ays
- tet
,3d
ays
- tet
,4d
ays
- tet
,5d
ays
- tet
,6d
ays
cav-
1 KO
wt
at 4
°Cat
37°
C
0min
In
hib
itio
n o
f cla
thri
n-d
epen
den
t
en
do
cyto
sis
B
HK
21-t
TA/a
nti
-CH
C c
ells
Inh
ibit
ion
of c
aveo
lin-d
epen
den
t
e
nd
ocy
tosi
s
en
do
thel
ial c
ells
Inh
ibit
ion
of e
nd
ocy
tosi
s
B
HK
21 c
ells
15’
15’
30’
30’
30’
30’
30’
initiation time
45’
45’
1‘
1‘
% transduced cells standard deviation
±
83 ± 6
73,5 ± 5
68 ± 4
77 ± 3
81 ± 4
84 ± 5
72,5 ± 6
54 ± 3
54 ± 3
75 ± 4
63 ± 2
FIGURE 5. Kinetic and quantitative analysis of transduction. Assembly of confocal optical sections of livingcells with time intervals of 15 min derived from the 60-min time lapses (see supplemental movies S1–S10)displaying the kinetics of TAT uptake in the different cell lines. All cell lines showed an unchanged transductionbehavior in the control cells and the cells that were inhibited for a distinct or all pinocytic events. But theaverage entry time point of the TAT peptide is cell type-specific. The last column summarizes the transductionfrequencies of TAT after 60 min of incubation in cells, where distinct or all endocytic pathways were sup-pressed. Transduction frequency was scored by counting the percentage of cells showing the intracellularfreely available peptide (see Fig. 1, B and C). Scale bar, 10 �m.
TAT Transduction Occurs in the Absence of Endocytosis
3376 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 6 • FEBRUARY 6, 2009
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

be used in biological sciences as well as in medicine and bio-technology. Former studies used chemical inhibitors of endo-cytosis to assign the uptake of CPPs to a particular endocyticpathway. However, the potential side effects and lack of speci-ficity of such inhibitorsmake these studies difficult to interpret.Either the chemical compounds affect more than one specificpathway, like methyl-�-cyclodextrin that affects both lipid raft(26) and caveolin-coated vesicle formation (50) hence alsocaveolin-dependent endocytosis, or they have other side effectsthat may impact the import of RRPs, e.g. chlorpromazine wasshown to interface with a number of Ca2�-dependent signalingpathways (51) and to bind to dopamine receptors (19, 52). Thus,we have used genetically modified systems or physical methodsto clarify the role of endocytosis in the translocation of RRPs.The different phenotypes shown in Fig. 1 permit at least two
explanations for the occurrence of free RRPs inside the cyto-plasm and nucleoplasm. Either RRPs become endocytosed anda portion of the peptide stored in vesicles gets released into thecytoplasm or a second nonendocytic entry route that allowsRRPs to directly cross the plasma membrane has to beconsidered.Earlier reports associated uptake of RRPs with and without
attached cargos to distinct endocytic pathways (16, 23–28, 39,40, 49). However, our results unambiguously demonstrate thatthe transduction of TAT into living cells is not dependent onany endocytic or pinocytic events. We could exclude the path-way of clathrin-mediated endocytosis by a carefully controlledknockdown experiment. Also caveolin-mediated endocytosiswas not involved in TAT translocation, because caveolinknock-out cells showed an identical transduction frequency tothe wild type cells. Most importantly, TAT was not excludedfrom cells that were gently transferred to 4 °C, a state where allpotential endocytic pathways are inhibited.It is noteworthy to mention that the amount of TAT trans-
duced into a cell varies between cells within a single experi-ment, but its intracellular distribution does not. This phenom-enon was observed in every cell type and was independent ofwhich endocytic route was down-regulated. (Fig. 2–5 and sup-plemental Fig. S3–S5).Comparing the transduction frequencies of the specific cell
types, a variation from68 to 84% for the BHK21-tTA/anti-CHCcell line, over 63–75% in BHK21 cells, and to 54% in the mouseendothelioma cells can be found, suggesting that the particularmembrane composition of different cell types impacted ontransduction (Fig. 5). This was further corroborated by theobservation that the average initiation time of transductionwascell type-specific, but it did not change upon the inhibition ofendocytosis within a given cell type. All experiments were con-ducted in addition to PTD4 as a representative CPP (29) with areduced number of arginines compared with TAT. At the sameconcentration, this peptide did not gain access to the intracel-lular compartments in a freely diffusing form and was internal-ized exclusively by endocytosis. This could be explained by thefact that aminimal number of six arginines is required to permittransduction (18). Instead, PTD4 was predominantly internal-ized by the clathrin-dependent pathway. Although not capableof performing transduction, the CPP PTD4 was endocytosed
more efficiently than non-CPP compounds added to themedium, e.g. the fluorophore TAMRA* alone (Fig. 3C).Recentmechanistic studieswith artificialmembrane systems
described the formation of pores as a consequence of the inter-action with intermediate concentrations of the RRP TAT (53).However, the fluorophore FITC*, concomitantly applied to thecells together with the TAT peptide, remained outside of thecells, whereas TAT transduced selectively into the cells (Fig.4C), arguing against the formation of nonselective pores.In summary, our data indicate that TAT CPP internalization
is independent of endocytosis and occurs without disruption ofthe cell membrane. These properties and its high intracellularbioavailability make TAT CPP a very effective tool to deliversmall compounds into living cells.
Acknowledgments—We thank Kirsten Sandvig for the generous gift ofthe BHK-tTA/anti-CHC cell line. We are indebted to our colleaguesPetra Domaing for help in the cell culture, Robert Martin and JeffStear for fruitful discussions and support withmicroscopy, and Sebas-tian Haase for advice in the image analysis.
REFERENCES1. Frankel, A. D., and Pabo, C. O. (1988) Cell 55, 1189–11932. Green, M., and Loewenstein, P. M. (1988) Cell 55, 1179–11883. Dietz, G. P., and Bahr, M. (2007)Methods Mol. Biol. 399, 181–1984. Nori, A., Jensen, K. D., Tijerina, M., Kopeckova, P., and Kopecek, J. (2003)
Bioconjugate Chem. 14, 44–505. Mann, D. A., and Frankel, A. D. (1991) EMBO J. 10, 1733–17396. Shibagaki, N., and Udey, M. C. (2002) J. Immunol. 168, 2393–24017. Tunnemann,G., Karczewski, P., Haase, H., Cardoso,M.C., andMorano, I.
(2007) J. Mol. Med. 85, 1405–14128. Tunnemann, G., Martin, R. M., Haupt, S., Patsch, C., Edenhofer, F., and
Cardoso, M. C. (2006) FASEB J. 20, 1775–17849. Astriab-Fisher, A., Sergueev, D., Fisher, M., Shaw, B. R., and Juliano, R. L.
(2002) Pharmacol. Res. 19, 744–75410. Fawell, S., Seery, J., Daikh, Y., Moore, C., Chen, L. L., Pepinsky, B., and
Barsoum, J. (1994) Proc. Natl. Acad. Sci. U. S. A. 91, 664–66811. Nagahara, H., Vocero-Akbani, A. M., Snyder, E. L., Ho, A., Latham, D. G.,
Lissy, N. A., Becker-Hapak, M., Ezhevsky, S. A., and Dowdy, S. F. (1998)Nat. Med. 4, 1449–1452
12. Schwarze, S. R., Ho, A., Vocero-Akbani, A., and Dowdy, S. F. (1999) Sci-ence 285, 1569–1572
13. Lewin,M., Carlesso,N., Tung, C.H., Tang, X.W., Cory, D., Scadden,D. T.,and Weissleder, R. (2000) Nat. Biotechnol. 18, 410–414
14. Iwasa, A., Akita, H., Khalil, I., Kogure, K., Futaki, S., and Harashima, H.(2006) Biochim. Biophys. Acta 1758, 713–720
15. Torchilin, V. P., Rammohan, R., Weissig, V., and Levchenko, T. S. (2001)Proc. Natl. Acad. Sci. U. S. A. 98, 8786–8791
16. Drin, G., Cottin, S., Blanc, E., Rees, A. R., and Temsamani, J. (2003) J. Biol.Chem. 278, 31192–31201
17. Richard, J. P., Melikov, K., Vives, E., Ramos, C., Verbeure, B., Gait, M. J.,Chernomordik, L. V., and Lebleu, B. (2003) J. Biol. Chem. 278, 585–590
18. Tunnemann, G., Ter-Avetisyan, G., Martin, R. M., Stockl, M., Herrmann,A., and Cardoso, M. C. (2008) J. Pept. Sci. 14, 469–476
19. Duchardt, F., Fotin-Mleczek, M., Schwarz, H., Fischer, R., and Brock, R.(2007) Traffic 8, 848–866
20. Fretz, M. M., Penning, N. A., Al-Taei, S., Futaki, S., Takeuchi, T., Nakase,I., Storm, G., and Jones, A. T. (2007) Biochem. J. 403, 335–342
21. Ziegler, A., Nervi, P., Durrenberger, M., and Seelig, J. (2005) Biochemistry44, 138–148
22. Mayor, S., and Pagano, R. E. (2007) Nat. Rev. Mol. Cell Biol. 8, 603–61223. Richard, J. P., Melikov, K., Brooks, H., Prevot, P., Lebleu, B., and Cherno-
mordik, L. V. (2005) J. Biol. Chem. 280, 15300–1530624. Ferrari, A., Pellegrini, V., Arcangeli, C., Fittipaldi, A., Giacca, M., and Bel-
TAT Transduction Occurs in the Absence of Endocytosis
FEBRUARY 6, 2009 • VOLUME 284 • NUMBER 6 JOURNAL OF BIOLOGICAL CHEMISTRY 3377
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

tram, F. (2003)Mol. Ther. 8, 284–29425. Fittipaldi, A., Ferrari, A., Zoppe, M., Arcangeli, C., Pellegrini, V., Beltram,
F., and Giacca, M. (2003) J. Biol. Chem. 278, 34141–3414926. Kaplan, I. M., Wadia, J. S., and Dowdy, S. F. (2005) J. Controlled Release
102, 247–25327. Nakase, I., Tadokoro,A., Kawabata,N., Takeuchi, T., Katoh,H.,Hiramoto,
K., Negishi, M., Nomizu, M., Sugiura, Y., and Futaki, S. (2007) Biochemis-try 46, 492–501
28. Wadia, J. S., Stan, R. V., and Dowdy, S. F. (2004) Nat. Med. 10, 310–31529. Ho, A., Schwarze, S. R., Mermelstein, S. J., Waksman, G., and Dowdy, S. F.
(2001) Cancer Res. 61, 474–47730. Iversen, T. G., Skretting, G., van Deurs, B., and Sandvig, K. (2003) Proc.
Natl. Acad. Sci. U. S. A. 100, 5175–518031. Drab, M., Verkade, P., Elger, M., Kasper, M., Lohn, M., Lauterbach, B.,
Menne, J., Lindschau, C., Mende, F., Luft, F. C., Schedl, A., Haller, H., andKurzchalia, T. V. (2001) Science 293, 2449–2452
32. Yaffe, D., and Saxel, O. (1977) Nature 270, 725–72733. Futaki, S., Suzuki, T., Ohashi, W., Yagami, T., Tanaka, S., Ueda, K., and
Sugiura, Y. (2001) J. Biol. Chem. 276, 5836–584034. Rothbard, J. B., Kreider, E., VanDeusen, C. L., Wright, L., Wylie, B. L., and
Wender, P. A. (2002) J. Med. Chem. 45, 3612–361835. Suzuki, T., Futaki, S., Niwa,M., Tanaka, S., Ueda, K., and Sugiura, Y. (2002)
J. Biol. Chem. 277, 2437–244336. Marsh, M., and Helenius, A. (2006) Cell 124, 729–74037. Nitschke, M., Korte, T., Tielesch, C., Ter-Avetisyan, G., Tunnemann, G.,
Cardoso,M. C., Veit,M., andHerrmann, A. (2008)Virology 377, 248–25438. Martin-Acebes, M. A., Gonzalez-Magaldi, M., Sandvig, K., Sobrino, F.,
and Armas-Portela, R. (2007) Virology 369, 105–11839. Kawamura, K. S., Sung, M., Bolewska-Pedyczak, E., and Gariepy, J. (2006)
Biochemistry 45, 1116–112740. Vendeville, A., Rayne, F., Bonhoure, A., Bettache, N., Montcourrier, P.,
and Beaumelle, B. (2004)Mol. Biol. Cell 15, 2347–236041. Gossen, M., and Bujard, H. (1992) Proc. Natl. Acad. Sci. U. S. A. 89,
5547–555142. Bennett, E.M., Lin, S. X., Towler,M. C.,Maxfield, F. R., and Brodsky, F.M.
(2001)Mol. Biol. Cell 12, 2790–279943. Feng, Y., Press, B., and Wandinger-Ness, A. (1995) J. Cell Biol. 131,
1435–145244. Press, B., Feng, Y., Hoflack, B., andWandinger-Ness, A. (1998) J. Cell Biol.
140, 1075–108945. Soldati, T., and Schliwa, M. (2006) Nat. Rev. Mol. Cell Biol. 7, 897–90846. Anderson, R. G. (1998) Annu. Rev. Biochem. 67, 199–22547. Simons, K., and Ikonen, E. (1997) Nature 387, 569–57248. Torgersen,M. L., Skretting, G., vanDeurs, B., and Sandvig, K. (2001) J. Cell
Sci. 114, 3737–374749. Zorko, M., and Langel, U. (2005) Adv. Drug Delivery Rev. 57, 529–54550. Saalik, P., Elmquist, A., Hansen,M., Padari, K., Saar, K., Viht, K., Langel, U.,
and Pooga, M. (2004) Bioconjugate Chem. 15, 1246–125351. Marshak, D. R., Lukas, T. J., andWatterson, D.M. (1985) Biochemistry 24,
144–15052. Seeman, P. (2002) Can. J. Psychiatry 47, 27–3853. Herce, H. D., and Garcia, A. E. (2007) Proc. Natl. Acad. Sci. U. S. A. 104,
20805–20810
TAT Transduction Occurs in the Absence of Endocytosis
3378 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 6 • FEBRUARY 6, 2009
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from

Herrmann, Marek Drab and M. Cristina CardosoGohar Ter-Avetisyan, Gisela Tünnemann, Danny Nowak, Matthias Nitschke, Andreas
Cell Entry of Arginine-rich Peptides Is Independent of Endocytosis
doi: 10.1074/jbc.M805550200 originally published online December 1, 20082009, 284:3370-3378.J. Biol. Chem.
10.1074/jbc.M805550200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2008/12/10/M805550200.DC1
http://www.jbc.org/content/284/6/3370.full.html#ref-list-1
This article cites 53 references, 21 of which can be accessed free at
by guest on June 26, 2018http://w
ww
.jbc.org/D
ownloaded from