centre - university of toronto t-space · 2020. 4. 7. · table of contents list of abbreviations...
TRANSCRIPT

Dioision 0f Cardiova!X&r s~l?@~'y The Toronto Hospital
The Centre for Cardiov8sc\1k Research
A thesis submitted in conformity wîth the reqiiirements for the degee of Doctor of Pbilosophy.
Graduate Department of the hsühte of Medicai Scieneg University of Toronto
Copyright by Vivek Rao, MD (199%)

National Library Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services seivices bibliographiques
395 Wellington Street 395, rue Wellington OttawaON K 1 A W -ON K1AûN4 Canada canada
The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distribute or seil reproduire, prêter, distniibuer ou copies of this thesis in microform, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/nlm, de
reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts fkom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

UnivaSay of Tomnt0
ABSTRACT
BACKGROUND: ~ o v a S c u l a r disease remains an imporiant cause of morbidity aiad
mortaiity in North America. Surgical tnatment of cardiovasculaf disorciers has emeqed
over the past 40 yavs and is now a pnwm, enrctve therapy. Cardiop1egic arrest of the
heart enables surgeons to operate on a motionless field, but is associated with a degree of
perîopaative injury. Sensitive measures of myocardial mefabolism and hction meal a
deiayed recovery following surgery which may lead to postoperative morbidity and rnozfality.
These Senes of investigations were designeci to evaïuate the recovery of aembic
myocardial metabohm following cardioplegic arrest. W e believe that improuing the
transition from aMerobic to aerobic membolism may resuit in enhancd functional recovery
and impved patient outcornes following cardiac surgery. W e hypothesized that insulin
would stimuîaîe myocardial pynivate dehydrogenase activity and lead to improved metabolic
recovexy foiiowing s i m W ischemia and repemision.
MEEIODS: W e ernployed isoiated, cultured human v e n û i ~ cardiomyocytes to examine
the effects of ischemia and repenusion on myocardial metabolism. W e measured the degree
of cellular injury, intraceiluiar iactaîe accumulation, extraceliular lactate and pyruvate
release, intracellular high energy phosphate levek and the activity of rnitochondrial ppvate
dehydrogenase (PDH). We investigated the effect of preischemic exposure to insulin on the
recoveq of myocardial metabolism following ischemia and repemision.

RKSïETS: Prekhemic exposun to iasulin (10 RTn) was found to confer pmtection
q a h t ischemia, Iasulin was found to stimuiate riiitocbiondrial PDH dwhg stabjlizatïm lad
pnserved the activity of PDH folîowing iscbemia and npmision. Insuiin aposure reduœd
extmdular iaçtite rd- and impfoved the m o n of high en= phoqhîes. The
mechanism of insulin' effd on myocarraal PDH was fwnd to be mediated by Protein
Kmase CU stimulation of the PDH phoaphatlse &unit.
CONCLUSIONS: Preischemic exposure ta iasulia improved human cardiomyocyte tolerzil~lce
to ischemia and repemison. These inv- 0 prwide the nrst duect evidenœ that
insuiin can stimulate human myocardial pynrvate dehydrogenase and lcad to improved
metabolic recovery foîiowing ischemia. Novel -es of mya?udial protection can be
developed b exploit the beneficial metabolic e f f e of pynniate dehydrogenase stimulation.

This work would not be posi%Ie without the assbtauce of my many fnends and
coiteagues within the DiVison of CardiOvaScular Surgexy at the University of T010nto.
1 wouîd Mce to thanlr my mentor aab supaviaor, Dr. Richard D. WeiseI, for his
tirelessguldance,supportande~xwnagemcnt* IomforeMgraoefultohimfariatroducing
metothepl;ictiœofacademic~urg~andhopetojustifyhiscommittmmttaa~eintbe
years to corne.
I wouid iike to thank my Program Diredor in the Division of CardiOvaSculaf Surgery,
Dr. Christopher M. Feindel, f a his support and vaiuabie advice throughout my clinid and
research training. 1 am indebted to the members of my thesis cornmittee for their valuable
input and suggestions: Dr. D d d A.G. Mickie (Chical Biochemistry) and Dr. Brian H.
Robinson (Medical Genetics).
1 greatly appreaate the love and cornfort of my fàmily and especiaiiy my wife Zena
whose nurnerous sacrifices throughout my braining have aliowed me to pursue my goals.
Lastiy, I wouid like to thank the Heart and Shroke Foundation of Canada for
providing me with Research Fellowships in suppoa of this work.

Table of Contents
List of Abbreviations
ChuparOiu: KNOWLEDGE Tû DATE
1.1 Myocardial Protection for - surgery
1.1.1 In-ve Myocardiai Physiology 1.1 -2 Persistent Anaerobic Mefabolism and the
Recovery of Left Ventricuiar Functicm 1.1 -3 Aerobic versus Anaeaobic Glucose Melabolkm 1.1 -4 Stimitlating Glycolysis During Ischemia 1. i .5 ûptïmai Delivery of Cardioplegia 1.1.6 Stllnulating Myocairlial MetaboliSm with
Cardioplegic Additives
1.2 The Pyruvate Dehydrogenase (PDH) Cornplex
1.2.1 Structure and Function of the PDH Complex 1.2.2 Effect of khemia on PDH Activity 1.2.3 Irwlin and the Pynrvate Dehydrogenase Complex 1.2.4 nie Role of a Second Messenger
1 -3.1 Signal Transduction Pathways 1.3.2 The Discovery of Protein Kinase C 1.3.3 Isoform Specinc Pmperties of Protein Kiaase C 1 -3.4 Pharmaco10gic Moâiilation of Protein Kirme C

1.3.5 Protein giiiase C and Ischemic Reamditioning ... 35
chapter Ibo: INSULIN STXnIULATES MYOCARDIAL PYRUVATE IDEEYDROGENASE AND PROTECrS ISOLATED HUlMAN VENTRI- CARDIOMYOCYTES FROM SlMWATED ISCEKEMIA . .A6
2.1 Introduction . ..47
2.2 Methods . . -48
2.2.1 Human Ventriculaf Cardiomyocyte Culture . . -48 2.2.2 Simulated Ischemia and Repafusion . ..49 2.2.3 Expenmental ProtocoIs .. -50 2.2.4 Assesment of Cellular Injury . ..50 2.2.5 Biochemical Measuremerits ... 51 2.2.6 Statisticai Andysis . ..53
2.3.1 Assessrnerit of Cellular Injury 2.3.2 Insulin on PDH Activity 2.3.3 Insulin Effect on Intermediate Metabolites 2.3.4 In& Wect on Adenine Nucleotides
2.4 Conclusions . . .56
KINASE C-a DEPENDENT
Introduction
Methods
3.2.1 Insuiin Effect on Protein Kinase C 3.2.2 PDH Activity FoiIowing PKC Modulation 3.2.3 Statistical Andysis
Results
3.3.1 The Efféct of Insulùi on Protein Kinase C 3.3.2 PDH Activity Following PKC Modulation
Conclusions

chpar Fow: ADDITIONAL INVESTIGATIONS
4.2.1 Endotherial CeU Study 4.2.2 Porcine Model of Orthotopic Transplantation 4.2.3 Donor Operation 4.2.4 m e n t Opaation 4.2.5 Biochemical M-ts 4.2.6 Statisiical Analysis
4.3.1 Endothelial Cell Study 4.3.2 Porcine Study
QIoprer me: DISCUSSION
htroductim
Cell culture Model
5.2.1 Human Ventricuiar Cardiornyocytes 5.2.2 Sirnuiaieci Ischemia and Reprfbsion
Insulin Mediated Cardioprotection
5 -3.1 Reduction of Cellular Injury 5.3.2 Metabolic E f f ~ of Insuiin Exposure 5.3.3 Hemodynamic EffecfS of Insulin Exposure 5.3.4 Mechanism of the Insulin méct
Additional MectS of InsuIin Exposure
5.4.1 Insulin and Fatty Acid Metabolism 5.4.2 Insulin and the N a + Exchanger
Cell-specific Effeds of Insulin
Summary and ûrighd Contributions
Future Amis of Research

APPENDIX ONE APPENlDIX TWO

AdenOSiae AdaaoBinedipbphk Analysis of vPiPnce Adenosine triphosphate Adenosine tnphoaphatase Bovine Semm Aibumin -nazyarterybyppspgraftjng Calcium Ion Calcium Chlaride Calphosfin C Chdaythrine confaence Intervaf Creatine kinase MBfrsiictionofcreathekinase carbcm dioxide Creatine phosphate Counts per million Degms CeISius Change in absorbane 1.2-Diafylgiycerol Dichloroaawe DirnethyIsulphoxide DeoxyribonucIeic acid 1.2-DiOcfanoylglyœrOl "and others" Flavin adenine dinucleotide F I a . adenine dinucleotide (reduced) Fluoroscein isothi~cyanate Gram Guanosine triphosphate binding protein Hydrogen ion Hydrochioric acid N-[2-hydroxyethyI]piperaZine-N'-[2-etbanesulfonic] acid High @ofmance liquid chrornatography Sulphuric acid Hypoxanthine Immunoglobulin G hosine Inosïtol 1,4,5-triphosphate Insuiin receptor substare-1 Inteniaticmai Units Potassium ion Potassium chloride Potassium phosphate (monobasic) Potassium phosphate (dibasic)

kDa LAD m- M MARCgS Mo" lw% min mol mOsm mRNA
P Pa PKC PiMA FTCA RACKS rpm RNA SAS SEM SOD SPT
modalton L& anterior descending conniary arOay Müli- ( 1 0 Moles pr Litre Myristolstpri, ahninerich, C-kinase abstraie MagiiesiumTon Magnesium Chloride Minutes Mole (6.023 x 1 p particies) Mïlïïosmoles Messenger RNA Nano- (lm Nitrogen Sodium Ion Sodium bicarbonate Sodium bisulphite Sodium carbonate Sodium chloride Sodium hydroxide Disodium phosphate Sodium phosphate Dihydronicotinamide adenine dinucleotide (oxicii.lpn) Dihydrrmiwtinamide adenine dinucleotide (reduced) 2'-Gmeth yladenosine ûdds Ratio OXygen Percent (parts of a hundred) Phosphate b u f f ' saline Perchloric acid @Ci03 R(-)N6-@hmy1-2R-iSopf0~y1)-adenosine Phosphatidyl4,5-bisphosphate pi- (109 Negative logarïthm of hydrogen ion concentration Protein kinase C Phorbol 12-myristate 13-acetate Percutaneuus û'ansiuMnal coronary angiuplasty Receptor for activafed C-kinase subtype Revolutions per minute RibonucMc acid StatisticaI Analysis Systems Standard error of the mean Superoxide dismutase 8-psulphophenyl theaphylline M i c m (106)

Figure 2: The relationship betwrea myOcaLdial lirgte release, oxygeri extraction and acid release during rrpahisioa ancl the developmeat of postoperative low output syndrome @AIS) in 614 paticats uadagoing isolated ammary aaay bypass surgery. Patients who dtveloped LûS (11-36) had sigaincantly higher ladate release imrnedhtdy d c c~ossciamp removal (XCL OFF') and at fÏve miautes of repahision (5'). However, net myocanfial lactate Aease at ten minutes of repenusion (10') was not ciiffixent between groups. There were no diffefences in myocardial oxygen extracton or acid release at any t h e point. (Adopredfr<m 2kommici;is et ol.9
... Pape 41
Figure 3: Myocardial giucose me!aboiism. ... Page 42
Figure 4: The mitochondrial eiectron nansport (nspiratary) cchain. NADH is oxidized by complex 1 and coenzymeQ, a lipid soIuble quinone. Comp1cx II consis& of succiaatc C reduc&se and cœnzyme-Q nductase and transfers reduchg equilavents h m FADH, to comp1ex III (cytochrome c reductase). Cornplex III then transfers electrcms to complex IV. cytochrome c oxidase (COX). COX thai reacts directly with rno1eda.r oxygen to fomi hydroxyl ions (OH-) which react with the protans pmduced by the complex 1 reaction to f m water (',O). For each NADEI molede, three molecules of ADP are phosphorylatprl to form ATP, whüe two m01ecules of ATP are produoed for each molecuie of FADHZ.
... Page 43
Figure 5: The pyruvate dthydmgenase cumplex. The El mbunit acts to decarbaxyiate pyruvate to form CO, and thiamine pyrophosphate (TPP-EI).?~~ E2 subunit transfers the acetyl- group from TPP to a lipoy1 =factor and then subsequently to cofactor A, fomiing acetyl-CoA. The E3 subunit is a fiavin q u i M g enyzme which oniciizes the Lipoyl cofactor h m E2 by transfming elecirons to NAD+. The cornplex is regulated by two additionai enzymes: PDH kinase and PDH phosphaîase.
. . .Page 44

Figure 6: The insulin naptor compln. nie iaPulin rccepor cornplex is a fetrameric traasmembrane glycopotein corisistuig of two 120 kDa O and two 90 kDa B subdts. The two ar-sububits are linLtd by diSulphi& bonds, are entinly extradulat and amtain tbe insuün bmding sites. Each Bsubunit is ïinkd to an a-subunit by a disulphide bond, crosses the p h n a membrane and contains a turr,sine kinase domah in its intraceUuhr poition. The asockfion of insului protein with i reqtm lotimulates the tyrosine kinase activity of the Bsubimit and d t s in autopbobphorylation of the second BsubUIut, Autophosphorylaticm of the &subunit s t i m m the kinase advity of the feceptortowardsothasubstratepmteins, includingiiisulinreceptorsubsaatel w-1). ... Page 45
Figure 7: Representative photomiaograpbs of primary cultures of human pediatnc (A) and adult (B) venhricuîar cardiomyocytes. (2OX magnification, reprinted ficm Li et a P )
. . .Page 58
Figure 8: Schematic diagram of the quiprnerit required to simulate "ischemia" and reperfusion. 100% nitmgen gas (NJ is bubbled through two oxygen trapB prïur to flushing a seaïed plexigiass chamber. Four plates of cultured ceils can be placed in each chamber which is equipped with a cent& sampling dish to ensure the absence of oxygem and to monitor temperature. (Reprint& from Turniati et aP)
... Page 59
Figure 9: Representaîive photomimgraphs of hurnan ventriah cardiomyocytes foliowuig assesrnent for cellular viability using 0.4% tiypan blue dye. In control non-ischemic ceils (far left panel) no d u h r injury is visible. In contrast, the œlls in the far nght panel have undergone 90 minutes of ischemia and 30 minutes of repemision resulting in approximately 50% cellular injury.
. . .Page 60
Figure 10: Assessment of cellular injury foilowing ischemia and repenuSon. At an ambimt glucose conceritration of 5 mmoYL, thae was no sipnincant effkzt of insulin treatment. Howeveq exposure to 10 IU/L of insulia at an arnbient gïuoose concentration of 100 mmoYL reduced cellular injury with no fiirther protection demonstrated as the insulin concentration increased to 100 W.
. . .Page 61

FigureAl: Assessmentofdalari~uryfo~girdiemiaandreppfusi~~~. Eighercell injury was o b d as the gipcoae cmamtratiion incfeaSed @y tweway ANOVA, glirçcnre @cf F=6.48, p <0.0001). Tbe addition of 10 IUL of insuiin reduced d u i a r injury (anclin @kt F=U.26, p < 0.0001). Duncan's muitiple range test spccifird di&naaa betweai irisulin ami oon-însdh tnated p u p s at glruxwc ooncentratiioas of 50 and 1 0 mmollL.
...Page 62
Figure 12: Pymmte dehydrogemse (PDH) activity folîouhng thirty minutes of scposure to glucose and insulia (STABïUUTION); irimty minutes of ISCaEMIA and thirty minutes of REPERFUSION. h s u h expanne increasd PDH aictlvity at both levels of giucose @or to ischeda lad preventeû PDH Wvaticm diaing repemuicm. (Results wmpved to mm-isGhemic amtrolvaiues obtirineri at bgseline or h m d s nposed to equrvalent volumes of nornioxic PBS for equivdent the @mis.)
... Page 63
Figure 13: Pynivate dehydrogenase (PDH) activity following thirty minutes of srposure to mannitol (5 or 100 mmoVL) and insulin (O or 10 WL). Lnsulin resdted in simiiar PDH stllnuiation at both canœntrations of mannitol, indicating that the stimulatory e f fe of insuiin is indepaiderit of giucose.
... Page 64
Figure 14: Intraœlluiar lactate accumulation fouowing thirty minutes of stabibtion, ninety minutes of ischemia and thïrty minutes of repemiSion. Lactate extraction during stabiIïzaticm and repahision was highest in ceils exposed to 100 rnM and insulin. Howeva, insulin reduced iaciaîe accumiilation foUowing ischemia in cells exposai to a glucose concentration of 100 mM. Intracelluiar lactate increased in ail groups foiiowing ninety minutes of ischemia. (Results compared to non-ischemic wntrol va- obtained at baseline or h m c& erposed to equivalent volumes of normoxic PBS for quivalent tirne peaiods.) . . .Page 65

Figure 15: ExtraœUuiar ladate reiease into the supaaatuit wer each plate. Lactate rel*ise increased significantly with ischemia and remained elevakd durhg repafusion, suggestiag peEweflt aaaaobic metabolism. Insulin treabneat r e d d lactate dease at boîh glucose concentrations. (Results compered to non-ischemic control values obtained at beseline or h m cells exposed to eqiiivaent vohnes of normoxic PBS for eqiiivamt tirne periods.)
... Page 66
Figure 16: Iatracelfular adenosine tnphospbate (ATP) levels following -011, after niriety minutes of isdiemia and afkr thirty minutes of qerfkicm. ATP fell sïgnincantly in aU groups, but wpo ktta pserved in cells expoBed to 100
glwse and 10 Nn insulia. R e d t s apressed as a pacentage of colltrol values obtained at baseline or f b m d s exposai to equivaient volumes of normoxic PBS for equivalent the periods.
... Page 67
Figure 1 7 Intradluiar totai adenhenucleotide (TAN) Ieveis foilowing stabïhtion, aRg ninety minutes of ischemia and a f k thitty minutes of repafusion. TAN f a sigdcantly in ali groups, but was better pmserved in œUs exposed to high glucose and insulin. Mts expressed as a percentage of control values obtained at baseiïne or h m ceb exposed to equivalent volumes of nmoxic PBS for quivalent tirne periods.
... Page 68
Figure 18: In-situ fluorescent immunohistochemistry demonstrating the distn%ution of protein Ianase C using a 1:40 dilution of rabbit anti-human anti-pmtein kinase C IgG antibody. Panel A demonstrates gemdhed cytopbnic staining in œlis exposed to 5 mM gimse alone. Ceils exposed to 10 IW/L of insulin with 100 a glucose (panel B) dispiay a distribution of PKC staining to the sarcoIemnal membranes.
... Page 113
Figure 19: In-situ fluorescent imrnunohista:hemisûy demonstrating the distn'bution of protein kinase C using a 1 :40 dilution of rabbit anti-human anti-protein kinase C,, IgG antibody. Panel A demonstrates generaüzed cytaplasmic staullng in ceiis exposeü to 5 a glucose done. Cells exposed to the phorboI ester PMA (1 PM; panel B) or adaosine (50 FM, panel C) display a ndistribution of PKC to the perinuclear and sarcolemnd membrane. Ischemic preconditioning @anel D) d t s in a similar redistribution of PKC stainiag. (Reprinted h m Ikonomidis et aiP)
... Pape 114

Figure 20: Slot blot analysis d e m e g insuiin's ened on protein kinase C WC!) tmslocafion. Each lam is bloüed with U) pg of protein. The left paad shows no e&d of iasulin, PMA or adePosine on PKC* distributiou. The r&ht panel demonshates that insuiin, PMA and adenosiue cause a translocaticm of PKC-cu to the membrane fracfim. The adenOSU1e mediateci PgC-cr taasîocaîion was iihiaited by the suicinosine nceptor antagonist SPT.
...Pape 115
Figure 21: Measuriernent of total piobin kinase C (PKC) activity usllig an in vin0 phosphorylation assay. nie phosphorylation of a PKC sp&c peptide (epidermal growth fador mcepor, R K R . ) is mm*tsured ~01OrimetRcally as spectrophotornetric absorbanœ at 570 nm and comcted for protM cuntent. The p h h l estez PMA resuited in a sigmficant stimuiation of PKC activity compared to 5 glucose alone. A signincant interactive effect between insulin and giucose and total PKC activity was fomd (gluCose*illSUlin, F=S.l4, p-0.035 by nnANOVA). Insulin shulated PKC activity with a greata e f b t observed at an ambient giucose concentration of 100 mM.
...Page 116
Figure 22: The activity of mitochondrial ppvate dehydrogenase (PDH) activity following protein kinase C (PKC) madukition. The phorbol ester PMA stimulated PDH activity to a si* extent as high glucose and insulin. CeU exposed to high glucose and insului in the pfe~ence of the PKC antagonists calphosth C (CALC) and chelyrauine (CHEL) demonstrated PDH activities simiiar to control values (5 a glucose).
...Pape 117
Figure 23: Twenty-four hour survival foilowing ninety minutes of ischemia and repemision. Endotheliai d s (EC) displayed incnased sensitivity to ischemia compareci to cardiomyocytes (CM). Insulin treatment conferred protection to cardiomyocytes, but not to endothelial tek.
... Pape 166
Figure 24: Endothelial celi pynivate dehydrogenase (PDH) activity and extracellular lactate release foiiowing exposure to insulin. Insuiin stimuiated PDH activity and reduced extracellular lactate release.
... Page 167
Figure 25: Effect of continuous low flow donor blood pgfusion during three hours of hypothermie stmage. Measurements were obtained prbr to organ procurement (PRE); afta the initiai cafdjopllegic infusion ( P B ) ; after the completion of the lefi atrial (LA), ngbt aîrial (RA) and pulmonary arterial (PA) anastomoses; a m removai of the aortic crossclamp QCL) and every 15 minutes during reperfusion. Hearts in the BLûûD pemised group displayed persistent anaerobic metabolism with greater iactate and acid release during cardiuplegk arrest. There were no differences during the repemision perid.
... Page 162 XV

Figure 26: Effect of continuous lm flow donor blood pafusion during three hom of hypothermie storage. Perfiisicm of daior bLood (BLOOD) led to an impnmd recovery of left ventricular devdopeû prrssure compared to ncm-pesrwtd mm1 heam (CONT).
...Page 163
Figrire 27: Effect of conlinuous low flow dona blood pemision enhanced with insulin. Insulin treatment (lNS: 10 MIL) d t e d Bi an e a r k rec~very of aembic metabolism and improved the remvery of lefi vmtricular deweloped pmsure compared to blood alone (BLOOD) or non-pemised control hearts (CONT).
. . .Pape 164
Figure 28: U p p r PuneZ: Myocardial ladate flux during and afkr cardioplegic arrest. Patients who d v e d iasuün erihanced CafdiopIegia displayed lactate exîmction immediately a f k r crosscm removal compared to persistent lactaie release in the placebo grwp. Lowiier P d Left vmtricular fuaction was betm preserved in the insulin cardiople& group afta two hours of Rperfusim.
...Pape 165

CârdiovaScuIaf disease is the most prevaient chronic disease in North Amexka and the
leaduig cause of morbidity and rnOrfality.l3 Athemclen,tic coronary artay disease accounts
forwer6096 o f a l l d e a î h s d u e t o c a r d i a c ~ . '
Medical treatment remains the prefened theiapy for the initial management of angha
pectmkY UnfOTa1Rately, many patients continue to have symptoms despite maximai medical
therapy and are forced to severeiy restrict their aCtiYities of d d y living.
Surgeons once held the N e f that the heart would never be able to withstand a
surgicai insuit. The famous surgeon, Thdore BiIfroth, was once quoted " Those who
attempt to operate on the heart are doomed to fail and to lose the esteem of their
colieagues" .' Nevertheles, surgical repair of congeniîai and acquired heart disease contintml
to interest swgeons for many years. Ludwign Rehn is cndited with perfkmhg the first
cardiac operation in 1897 by repairing a stab wound to the heart with direct suture c ~ o ~ u f e . ~
Improving the blood supply to the heart proved to k fàr more dificuit as ail
techniques employed usually resulted in vascular fibrosis and loss of patency. Claude Beck,
at the Cleveland Qinic, developed methods to indirectly revascularize the heart by suturing
adjacent structures such as pericardium, pericardiai fat and omentum in an aüempt to develop
mUaW blood flow.' Arthur Vieberg, a Canadian surgeon at The Royal Victoria Hospital
in Montteal, first describeci in 1946 a kchnique of implanthg the left internai rnammary
(thora&) artexy (LITA) into a myocardial tunnel hopig to establish coiiateral circulaîion with
the left anterior descendkg coronary artery (LAD).8 The success of this operation was
quexied by many but was subsequently validated in 1960 by Mason Sones at the Cleveland
ciinic who performed se1ecfive coronary arteriography and àemonstrated patent collateral

2
communications betwem the LJTA and the LAD in two patients who were operated on 5 and
6 yars pior to aiigi~graphy.~
The fifit dued cofoaary artaial aaastomosis was actiially desaibed in an animal
modd by Alexis Carrel in 1910.n Using a cryopresewed camtid artery graft, Caml
pesfmed an anastomosis between the descendhg t h d c aorta and the left coronary artery.
Caml commenteci that it took him 5 minutes to perform the anastomosis, but that intractable
ventricuiar fibrillation developed after only thne minutes following interruption of the blood
flow to the coronary artery. He M e r commented that the continuous beating motion of
the heart made dissection difficult and that such an operation would have to take less than
three minutes in order to be sucasfd.
in 1955, Melrose descnbed the fïrst elective amst of the heart using a potassium rich
"cardioplegicn sol~tion.~' This solution provided a motionles, bloodless field for surgical
intervention. Unf~rtunately , the h . potassium concentration (240 mmol) resuïted in satere
cardiac injury.12 Reintroduction of a crystailoid solution with a lower potassium
concentration in the late 1970's enabled surgeons to electively arrest the heart with much less
ischemic injury than with intermittent aor&ic crossclamping (occlusion). "
The benefits of hypothennia were fbsî espoused in 1950 by Dr. Wilfred G. Bigelow
at the University of Toronto.'' In a series of classical experiments, Bigelow and wlleagues
were able to show that moderate hypothennia (25-28C) mis able to protect the heart h m
ischernic injury. This technique, with or without intermittent aortic crossclamping was
widely used d h g the early years of cardiac surgery.
In 1967, Favalm described the use of autogenous saphenous vehs to bypass stenotic
lesions of the coronary arteriesOu Coronary artery bypass grafong (CABG) is now the most

3
commody ~ m e d surgical procedure in North Ameria2 Several, large randomhd
cliinicaI trials have demonstrated the supaiority of CABG over medical treatment for
p1011ging Me and reducing symptoms in subsets of patients with athemsclerotic ainnary
severe triple v d a>raiary artery disease,' unstable an- and left ventricular
dysfiinction.' Since its original description by Favaioro, s e v d advances have been made to
reduœ the morbidity and mortality associated with CABG.Ia Most importa;lltly, advances in
myocardial ptectionWl9 have dowed surgeons to opadte on an increagIlgly high risk
patient population without a signifiant increase in paioperative complications.
Unfortunatey, several subgroups continue to be at increased risk for perkpedve
morbidity and mortality. These include fernales,= patients with poor Preoperative left
ventncular func t i~n~~ and those who prrsent for s u r g q with either unstable angîna or a
reoent myacardial infarcti~n.~~- Improved methods of @operative rnyocardial protection
are required to reduce the risks of surgery for these preCanow high risk patient populations.
Modifications to the delivery and composition of cardioplegic formulatjcms may lead
to improved myocardial. protection. Despite the success of contemporary coroaary bypass
surgery, sensitive indices of myocardial metabolhm and fwiction rweal delayed recovery
following cardioplegic The investigations descrii in this thesis w a e designeci
to investigate the me&bolic sequeiae of cardioplegic amst and to stimulate the recovery of
normal aaobic metabolism foliowing a pexiod of ischemia and npemision. Stimulating the
recovery of aerobic metabolism rnay lead to an eariier recmery of left ventricular functicm
and lead to improved surgical outcornes following coronary artery bypass surgefy.

the meîabolic respoases to simitlatPA cardiaplegic amat (iihemia) and qxdkireperfusion. 'Ibe use
of an isolatCui d culture model removes the possliIe Confouding &ects of d e r d types
and organ systems. In addition, the extracellular environment can be carefiilly co~ltroiied to
an extensive metabolic evafuation of isotatpri human cardiomyocytes subjec&d ta simutated
low fiow cardioplegic arrest and repemLpion. However, the in via0 redts of these
investigations require confinnation in a whole organ or in vivo model.


6
1.1 MY0CARDIA.L PROTECTION M)B CORONARY BYPASS SURGERY
The first heart opaations were PafOrmed cm the beabing heart. The results of these
procedure~waenotdydependerrtonthetechnicalsucce~sofUieoperati~~~, butalsoonthe
ability of the surgeon to reduœ intnoperatve myocardial injury. The advent of
of surgery and repair both cornpiex amgenital and aquired kart discase.
Sinœ Melrose first described the use of a 'cardioplegica solution to electively amst
the heart," th= have beea many advances in both the composition and delivery of
cardioplegia. The ultimate goal of any peziopesative myoprotective stmtegy is to provide the
surgeon with a motiodess, bloodless field in which to @mm a technidLy pafect conniary
aoastomosis while at the same tirne, preventing any injury to the heart which rnay result in
metaboiic or fwicticmai abnonnalities in the postoperative @od. In order to meet the
myocardid nutrient tequirements during cardiac surgery, it is cnicial to understand the
physiology of the arrested h m .
Z~cu)per4tr*~e Myoc(Udial PIrySiology
The heart npresents less than 0.5% of body weight, yet it accounts for over 7% of
the body's resting oxygen consumption." Myocardial oxygen consumption (MVOa can be
readily caicuiated using the Fick equation if coronary blwd flow (CBF), arterial (C.03 and
coronary sinus (COQ) oxygen contents are knom (MVQ = CBF x CC&-C-OJ). Cardiac
muscle extracts much more oxygen in the normal state than other organs, and thus increased
myocardial oxygen consumption is primarily achieved by increases in coronary blood flow.
The left ventncle ansumes approximaîeIy 8 mL of 4 per 100 g of myocardium per minute
in a normal human subject at rest. Durkg potassium i n d d arrest, oxygen conswnption

of MVQ are: heart nite, stroke work (the ana within the pressure-volume loop, which
incorporates afterld) and inotropic state. The relationship between M V a and heart rate,
stroke work and inotrapic state is almost iinear?" MVO, is greater when a low stn,ke
volume is ejeded against high pnssures than when large m k e volumes are ejected agaïmt
low aortic pressures.
During cardiac surgery, myocardial oxygen consumption varies widely. The lowest
MVG ocnirs whm the heart is arrestsd. Maximum MVO, occurs shortly after weaning from
cardiopulm~nary bypass whm the heart is rrpaying the oxygm debt incumd during the aortic
cross clamp period. In a series of classical experiments, Buclrberg et al examined myocardial
oxygen consumption during d i f f m t amditions of myocardial activity; the empty beating
heart, the fibrillating hart and the arrested heart? Myocardial oxygen consumption was
greakst during normothermic (37°C) fibnuaton and least duMg hypothermie (22'C) arrest.
Hyperkaiemic arrest achieved a reduction in MVQ h m 5.6 f 1.95 mUlûûg/min to 1.1
+ 0.4 mL/lOOg/min. Hypothermia reduced MVO, to 2.9 f 0.9 mUlO@/min while the - . combination of potassium induced arrest and modesdte hypothermia reduced MVQ to 0.3 f
0.1 mVlûûg/min. (Figure 1)
Myocardial ischemia occurs when there is an imbalance of oxygen supply and
demand. This imbalance results in anaerobic myocardial metabolism. nie end-products of
anaerobic metabohm rapidty lead to acidosis, mitochondrial dysfunction and myocyte
The myocardium is remarkably adept at utilùing any amilable substr;ite for energy
production. -hydrates, fatty acids and amino acids can be utiüzed for the formafion of

8
aϔyl-CoA. In the notmal non-hhemic myocardium, fany acids are the predominant
substrate for enexgy prOaucti~n;~~-~ however, during and after ischemia myocardial glucose
metabolism is highiy upregulated.* Durhg ischania, anaerobic giycolysis predominatw and
results in the production of lactate and hydrogen ions. Unfortu~îeiy, despite apparently
adequate -cm, anaerobic myocardial meâabolism oui persist with the amtinued
producfion of iactatC and hydtogen ions." Teoh et al examincd perioperative rnyocudial fatty
acid metabolhm in 18 patients undergohg isoIated CABG? These investi- found that
"C-labelled pairnitate was extracted nOm the heart but was not oxiCfj7PR. In patients
reœiving pure crystaiïoid cardiople@ (n =7), there was no detedable fatty acid oxidation.
In the remaining 11 patients who received blood Cafdioplegia, htty acid oxidation was
minimal. Myocardial fatty acid accumulation without oxidation may be deleterious and
contribute to repemision injury. The inability of the hart to oxidhe exogenous fatty acîds
~eflects a delayed recovery of norrnal aerobic metabolism foiiowing cardiupfegic anest.
Persistent AMerobic MetaboliSm and 171e Recowry of Left Vemnrrr& Funcn'on
In a nxent study of 614 patients undergohg isoiated CABG, 523 patients (84%)
displayed net myocardial lactate release folIowing aortic aossclamp releaseY AAa five
minutes of repemision, JOO patients (64%) displayed pergstent anaerobic metabolkm and
continued to release lactate into the coronary sinus. Thirty six patients (5.8%) developed
postoperative low cardiac output syndrome (LDS)). The objective definition of low output
syndrome includes any patient who requires inotropic or in-c balloon pump support for
greata than thirty minutes to maintah adquate hemodynamics aftm ail b l d gas, electrolyte
and volume abnoRnalities are armded.* This definition requires the active intervention of
the attendipg surgeon or intensivist; therefore, the development of LOS represents deiayed

9
recovay of lef€ veatricular funciion and a faiure of periopaab:ve mya'ardial pmtection.
Figure 2 illustrates the relatiomhip betweai myocardial lactate release, oxygen
extracction and acid release and the developmmt of poBtopaative LOS. Myocarrlial kdate
reiwse at crossclamp ternoval was higher in patients who developed LOS (0.92I0.2 mmoi/L
vs 0.45&0.02, p<O.Ol). Similady, myocardial lirtitc release aRer five minutes of
repafusicm was higher in patients who develaped LOS (O.2Sf 0.07 mmoYL vs O.O6f 0.01
mmoyL, p<O.û!5). Myocardial oxygen extraction or acid release were not differept in
patients who developed postoperative LOS. Stepwise logistic ngressicm identifieci pasisteat
lactate release after five minutes of repemision to be the only predictor of postoperative low
output syndrome (odds ratio OR 5.85, 95% confidence interval CI 2.1-16.3).
In a diversifiai surgicai popiitation, poor preopemtive left ventricular function
(OR=5.7,95%CI 3.69.0), repeat operation (OR=4.4, 9596CI 3-3-59), urgent surgery for
unstabIe angina (OR=3.7, 95%CI 2.3-5.9), f e d e gender (OR=2.5, %%CI 2.0-3.2),
diabetes meIlitu (OR=1.59, 951CI 1.3-2.0), age>70 years (OR=1.5, 9546CI 1.1-1.8),
lefi main disease (OR=1.4,9596CI 1.1-1.8), preopefative myocardial &&on (OR=1.4,
95%CI 1.0-1.9) and triple vesseI corouary artery disease (OR=1.3, 9596CI 1.0-1.7) were
found to be the independerit predictors of postaperative low output syndrome.% None of
these clinical risk k t o r s predict the deve1opment of low output syndrome in the
homogeneous study populations often employed for clinical triais in myocardial protection.'
Although we found m y o d a l lactate release at five minutes of reperfusion to have poor
predictive capability (area under ROC =O.6328), it was the only signifiant ri& factor for the
development of postoperative low output syndrome. The poor predictive capability may be
due to the fgct that in some patients with adquate myocardial protection, postoperative LOS

10
may be a r&t of intaopaative technid problems. Ladate rdease during nprfhion may
be highiy piedictive of LOS in patients who had an uneventfiil intmqemtive course.
U n f i y , it is diffidt to distindistinguish between the effecrr of inrteqI1SltP- myocaFdial
protection and inhaoperative misadvmture as these two events may be highIy correiated.
Nevddess, the development of postopedve low output syndrome in a patient who had
an meventful in-e course is due in part to a Aelrryed fecovery of normaî aembic
myocardial metabolism. nierdore, f'acilitating the transition h m anaerobic to h i c
metabofim foilowing CaIrdiopIegic arrest should lead to an improved recovery of lefi
ventXi* function.
Aembic W. Rnaerobic Glucose MetaboliSm
Figure 3 iiiustxates the myocardial metabohm of gîucose. Aembic glu-
meotbolism involves the conversion of glucose to pyruvate which is then mverted into
-1-CoA by the pyruvate dehydrogenase (PDH) enzyme cornplex. Acetyl-CoA the0 mtap
mitochondria and is metabolised via the Krebs cycle into carbon dioxide and water. The net
result of giucose oxidation is the production of 36 moles of ATP for each mole of glucose.
Anaembic giucose metaboliSm involves the converSon of glucose to lactate. The initial
phosphorylation of glucose consumes 2 moles of ATP to form hctose-16-diphosphaîe
(FDP). The conversion of FDP to lactate and wakr involves the production of 4 moles of
ATP. nius, the net energy production of glycolysis is 2 moles of ATP for each mole of
glucose. Unfortunately , the metabolic end-products: dihydroniwtinaniide-aderiine
dinucleotide [NADHJ, lactate and hydnogen ions act to inhibit phosphofnictokinase and can
thereby inhibit furth- glycolysis. This inhibition cesults in continuhg energy consumption
M m the initial stages of glycolysis -out any energy production. Not only is anaembic

11
Iine&bolism a les efficient source of ATP production, but it is also associated with the
production of h y m e n ions. The resultant demaad intracellular pH has detrimental
collse~umces to membrane and mîtochondrial stability.- ImpaiRd fundon of the sodium-
potassium ATPase pump increases cell membrane permeabiiity to extraceiiuîar calcium
resuiting in an eldion of intracelluiar caicium. Impairment of the ATP-dependent calcium
pump resdts in deaeased calcium ion scqu*ltration and eveiitdy in organelle injury and
myofibriilar c~ntracbre.~*~'
The citrïc acïd (Kreb's) cycle utiüzes aœtyL-CoA to produce a nwnber of d u &
ad&e nucleutides including dihydronicotinamideadenlne dinucleotiide (NADH) and flavin
adenine dinucleotide (FADH3. These flavoproteias are then utilized by the eleztron transport
chah within the inner membrane (cristae) of the rnitochondrial wmplex for energy
production. Figure 4 summarizes the major complexes of the mitochondrial respiratory
chah. The niitochondrial respiratory chah consists of a series of coupled reactions involvhg
elecani transfers h m iron-contaïhg hemoproteins or cytochromes. In each step, Kon
fluchiates from the reduced ferrous (Fe?+) form to the oxidized f h c @@+) state.
NADH is oxicüzed by cornplex 1 and the electrons transferred to coenyme, a lipid
soluble quinone. Complex 2 utilizes FADH, produced fkom the conversion of succinate to
fumarate to reduœ cœnzyme-Q. C0e~zyme-Q thus transfas the electrons frorn complexes
1 and 2 to complex 3 (cybchrome C reductase). Complex 3 thai transfers reducing
quilavents to cumplex 4 (cytochrome c oxidase). Cytochrome c oxidase (COX) reaas
directiy with molecular oxygen to form hydmxyl ions (OH-) which react with the protons
produced by the complex 1 reaction to form water (m). For each NADH molede, three
molecules of ADP are phosphorylated to form ATP while two molecules of ATP are

l2
proâuced for each molecuie of FADK. Oadaton of one mole of glucose yields 10 moles
of NADH and 2 moles of FAD&. In ytnitiou, two moles of ATP are produced h m the
anvexsion of succinylCoA to Succinata T'us, each mole of gïucose provides the net en-
quivakat of 36 moles of ATP foUowing oxidatve phospharylaîion.
Glucose oxidation can be summarued by two redox reactions:
Tota oxidation of glucose to CO2 and &O yields 686 W m o l . Each mole of ATP
stores approxhakly 8 kcai of energy." Thus, anaerobic glycolysis recovers only 2% of the
potential energy stored in giucose compared to 4296 reooverecl by oxidative phosphorylation.
Therefm, rnaintaining normal aerobic rnetabolism duMg and a f k cardioplegic anest
maximues the efficiency of ATP production h m glycolysis. In arlniticm to the increased
efficiency of ATP production, oxidative phosphorylation may be " c o m p a r t m e n ~ "
intracellularly such that the eaergy stores are diahguished h m ATP produœd by anaerobic
glymly sis.
Weiss et al found that in i s o M rabbit hearts, ATP generated h m aerobic
rnetabolism was preferentially used for myocardial contractility while anaerobicaily pmduced
ATP was prirnarily utilized for intracellular repaire4 This difference is teIeoIogically sound
because the ischemic cell shouid ConSave energy for survival fwictions only. Weksler et al
also demonsûateâ a pivotal role of anaerobic metabolisni in the presemtion of membrane-
bound calcium c h a ~ e l s . ~ These resuits also expiain the slow recovery in hction seen after
cardioplegic arrest in acutely ischemic hearts. The initial periods of repemision clearly Save

13
to replenish cellular energy stores which are used primady for intradular repair. Once the
Sodi-Pollares was the first to use an inûavmous infusion of glucose and insulin to
treat the e l m i o g r a p b i c abnormalitiies of an acute myocardial inf.arCtioneu Seveaal
hvestigators have since attenipted to improve myocardial tolerance to ischemia by either
enhancing preoperative ai- storeda- or by stimiilating giywlysis during ischernia.-n
The d t s of these investigations appear to be amtradictory. The two kgest evaluatiioos
took phce over 20 years a g ~ . ~ * Mittra et al reported on a praspective series of 170 patients
who received either a glucoscinsuiin-potassium suppiement or placebo shortly after bang
admitted with a diagnosis of acute myocardial infarcton.n nie design of this trial was
quential in that the first 85 patients d v e d standard therapy while the remaining 85
patients received the GIK treatment. This study demonsbrated a significant reduction in
rnortaiity in patients who receieved GIK (11.7% vs. 28.296, p<O.05). The authons
athibuted the reduction in mortality to a decread p a l e n c e of pst-infarction anhythmiaas.
A subsequent prospective randomized study involving 840 patients and fimded by the British
Medical Research Council mncluâed that exogenous glucose-insulin-potassium (GIQ
solutions did not have a beneficial effed on mortaiity or morbidity foIIowing acute
myocardial infarctoa6" They found that the mortality in the G E p u p was 23.9% v e ~ s
25.3 96 in the control group @=O. 6). The stuây ' s authors specirlatEul that improvements in
the management of pst-infarcpon complications superceded any potential benefit of G M
treatment.

14
Majid and wUeagues prospectiveïy evaîuated the use of GIK solutions for the
treatment of mgestive heart A Statistically signincant, but clinicaiiy margiaal
benefit was obsaved in 6 patients who receieved GM soluti~ll~ compand to conml patients.
Of interest, in patients who only received a high glucose supplement no hemodynamîc benefit
was observeci. The authors concludcd that msulin treatment improved the ability of the
myocaniium to metabolize giucose.
Although stimulation of glycdysis wouid intuitively be beaeficial, rnany investigatms
reported a detrimental e H l t on both myocardiai viability and f w r ~ t i o n . ' ~ ~ H m et al
demonstrated that cardioplegic arrest with solutions containug high concentrations of g h ~
led to iarger infarcts and depressed functionai reco~ery.~ Sirnilar.1~. ûrita reported that
storage of isoiated rat cardiomyocytes in presgvaton solutions wntaining high glucose
concentrations reduced cellular viability." Both of these investigatm athibuted their findùigs
to an accumulation of me&bolic end-products, namely lactate and h y m e n ions, which led
to intracellular acidosis and cell death. In Hearse's study, isohkd rat hearts were given a
single cardioplepic infusion prior to pf01onged, global ischemia. In this model, i n d g
the glucose concentration was ddeterious and the addition of insulin exacerbateci the injury.
In c o n w , Steinberg and Doherty demonstrated a benefiaal e f f a of glucose and insulin
containhg solution^.^^ Th& model employed multiple infusions of cardioplegia which
prevented the accumulation of toxic metabolites and led to irnproved functional recovery.
Other investigators who have atternpted to stimulate glycolyis by providing gluwsehsuün
solutions during reperfusion have reported a beneficial effe~t=-~ Thus, it appear~ that
in order to achieve a beneficial effect of metabolic stimuiation, one must prevent the
accumulation of end-products by either providing intemittent or continuous pemision of the

myocardium.
QJM Ddïwy qf Gmtiopkgia
Cardioplegic amst was originaüy achkved by idhion of a hypothermie ( 1 0 , high
potassium (27 mEq/L) so1ution into the amtic r0a1' Surgeoas maintained cardioplegic anest
with intemitîent infisions of a low potassium (8 mEqn) solution evay 15 to 20 minutes
during the aortic mssciamp mod. Howevex, sevexai investigators demonstrated that ttiis
fonn of myocardial protection resulted in a delay of both me&bolic and functicmai
-W. 'ma Fnmes et al demonsbrated that mixing blood from the bypass circuit with tbis
"crystalloid" solution in a 2: 1 ratio improved the recovery of both metabolism and function
following CardiopIegic arrest." In a prospective, randornized trial these investigaîors showed
that blood cardioplegia enhanced aerobic myocardial metabolism during aortic aoss-
clamping, increased myocardial oxygen coasumption, reduced anaaobic lactate production
and presemed high energy phosphate stores. Blood cardioplegia improved both systolic and
diastolic fwiction foilowing surgery. Since the publication of that trial, ail surgeons at the
University of Toronto switched to blood cardiopiegia. A subsequetlt clinid trial in patients
undergohg urgent revasc-tion for unstable an* demonstrated that blood cardioplegia
nduced both morbidity and mortality following ~urgery.~ Most institutions now employ a
b1ood:crystalloid ratio of 4: 1 or greatcr in theh mutine cardiap1egic formulations.* Blood
cardiople@a has distinct advantages over crystalloid solutions in tams of oxygen d e l i ~ e r y , ~
buffering capacip and the abiiity to prevent ïnwersib1e ischemic i n j u ~ y . ~
Cardiopllegia is traditionaliy deIivered antegrade into the aortic mot, a technique still
empIoyed by many cardiac surgeons. The initial arresting dose of between 500 to 1OOO mL
of cardiopIegh is given immedxatdy after the aortic cross-clamp is applied. Although

16
cardiop1legia can be given contiauously in an antegrade fkhion, it usually d t s in flooding
of the operative fkid and may compromise the technical q d t y of the distaI anastomosis.
Therefixe, most surgeons continue to give "maiatenancem cardiaplegia in an intermitfeat
Won. Since the aortic mot is clamped and isoIaîed h m systemic pemisicm, intermptions
in cardiopllegic delivery resuit in myocardial ischemia, Furthmore, in patients with sevexe
proximal disease, coronary sten0~e3 =y d u c e cardioplegic delivery and produce
Wtmgde delivery of doplegia inm the coronary sinus was origirially desccibed
by Gott in 1957? This technique allows for continuous delivery of cardioplegia with less
fiooding than observed with contuiuous antegrade deiivexy. However, most surgeons h d
corniary anastomosis. Sinœ hypotfiermic eardioplegic amst was thought to aliow fm safe
interniptions of cafdioplegia for up to twenty minutes, rnost surgeons employed intermittent
antegrade m o n and avoided the use of coronary sinus cafheters.
As Buckberg demonstratcd, hypottieda does not reduce myocardiai oxygen
requirements much beyond the reduction achieved with hyperkalemic a r r e ~ t . ~ In addition,
hypothermic cardiop1egia results in delayed recovery of both myocardiai metabohm and
ventricular f u n c t i ~ n . ~ ~ " - ~ See et al demonsbrated a signifiant impairment of mitochondrial
structure and function in a canine mode1 of prolonged hypothermic storage." Rosenkranz a
ai hypothesized that the metabolic dysfiinction may be partially due to the washout of Krebs'
cycle intermediates such as giutamate and aspaaate and showed that a normothermic (3m
induction of glutamate and aspartate d c h e d cardioplegia improved metaboiic fecovezy in
energy depleted hear&s.- Teoh et al then demonstrated that a terminal infusion of warm

17
blood cardioplegia (a 'hot shot") k m e d h l y @or to crosscbp release resulted in a
prolangation of elecfrornechanil rarwt, improvement in eerobic metaboliSm and inaeased
diastolic cornphn~e.~ The bewficial e&d of the hot shot was thought to be due to eariy
temperature &pendent rnitochondrial rrspiration and ATP generation. Patients who reœived
a terimaal warm blood infusion had signincantly higher ATP and glycogm stores cornparrd
to patients who did not receive a tmninai 'hot shotu. Since this technique d t e d in a
prolongation of electromechanical arrest, the ATP produced was presurnably used for repair
of intracefluiar ischemic injury and -011 of depl@ energy stores.
Clinid data indicaîed that normothermic induction cornbinexi with normothermic
terminal blood infusion facilitaml early recovery of m y e metabolic function.
Lichtenstein extrapolaîed these findings and proposed that normothermic pemision throughout
the crossciamp period might be benefi~hl.~' However, the modest inæase in myocardial
normothermic rnyocardial ischemia. Thus, Saiemo and coUeagues renewed the interest in
retrograde cardioplegia by delivering warm blood cardoplegia continuously hto the wronary
sinus.D1 A prospective. randomiÿed trial in 1732 patients revealed that normothermic blood
CardiopIegia giveii either aategra.de or retrograde Sgnificantly reduced the incidence of
postope&ve low output syndrome, but Med to show any ciifferences in perioperative
myocardial infkrctim @y ECG criteria) or m~rta l i ty .~
Although continuous retrograde delivery of wann blood cardiaplegia appead
d i v e , many investigators found that this technique provideci inadequate protection to the
right ventricle and posterior interventricular septum.- Since this region of rnyocafdiwn
remained normothermic, ischemic injury was potentiaily exaœrbated. Maatsura et al

Therefm? in an attempt to opîimjze myocardial pemision surgeons began to employ a
technique of combined antegrade and retmgde d e l i ~ e r y . ~ - ~ ~ In addition, Hayashida and
colleagues htmduced the concept of 'tepida carüiopiegia (29C) which was a natumi
compromise between h y p o t h d c (Io@) and normothemnic (37°C) extremes." These
investigators found that tepid blood cardioplegia reduceâ the anaaobic production of ladate
seem with normothermic m o n , but also prevented the delay in functional recovery
observed with hypothermie cardi~plegia.~ The same investigators then demonstrated that
providing tepid blood perfusion in a combined antegraddretrograde fàshion optUnized both
the temperature and delivery of cardioplesia,'a'w
GLUCOSE-INSULIN-POTASSIUM
The impvements in the delivery of cardioplegic solutions was paraiieled by
investigations into s e v d cardioplegic additives. The use of giucose and insulin enhanced
solutions for cardiac surgery mis unfortunately evaluated in an era of hypothennic,
crystalloid cardiopllegia. The mults of these investigations were controversiai and were
d e s c n i eariïer (page 13). The benefits of tepid blood cardioplegia erihanced with insulin
and delivered in a marner to optimize myocardial pemision have not been adequately
asessed." In addition, the management of systemic body temperatures duruig
cardiopulmonary bypass has also changed with the. Most surgeons now allow body
tempadhnes to "driftw to approximate1y 30-32°C. Previously, body tempemues were

19
actively cooled to below 2&C. Kuntschea et ai foMd that systemic hypothennia signifïcantly
impaireci the hepatic and pancreatic responses to a dextrose 1oad.l In particular, they f o d
hypothdc CaTdiopuimonary bypass and pcrsisied hto the eariy postopaative @ad.
Diabetic patients undesgohg nm- cardiopulmonary bypass displayed a simüar
enhanced cardioplegia would most W y demonstrate a benefit in the modern era of cardïac
surgery employing relativdy normothennic myocardial and systemic pemision.
Further evidence to support the narmothermic use of insuiin cornes h m severai
reports of its efficacy in the treatment of Iow cardiac output syndrome folIowing surgery.lOH1O
Of interest, Svedjeholm dernonstrated that high dose insulin potentiated the effects of
dopamine in the early postoperative period?' ûther investigators have also reported both a
VaSOdiIafOryfP and positive inotropicxn effect foIiowing insulin administraton. These reports
a h g with cLinical anecdotes support the need for a formal prospective re-evatuation of
glucose-insulin solutions in cardiac surgefy.
GLUTAMATE- ASPARTATE
Teoh et al used a glutamate and aspartate enriched cardoplegic solution in patients
undergohg isolated coronafy bypass surgery.lu Although low-ri& elective patients did not
benefit, glutamate-aspartate provided a slight benefit to those patients undergohg urgent
surgery for unstable angina. Similarly, Rosenkranz et ai showed a baiefit of glutamate
enriched blood dopleg ia in high risk patients with cardiogeaic shock? These authors
hypothesized that patients who present for surgery shortly after an ischemic event have a
deplebion of Krebs' cycle intermediates such as giutamate and aspartate. They suggested that

phosphates. However, the myocyte i s remrkably adept at utiüziag any available substrate.
Thus, in the piresence of oxygai, a norrnally hindionhg mitochondrial system should be able
to meet the mergy requirements of the cdl. The mle of Krebs' cycle intermediates may k
W y understood. A clarificafion of the mitochondrial rnetabolic defecf~ caused by ischemia
and npemision may p d t the development of a more site-speafic compound aimed at
restoring norrnal aerobic metahlism.
COENZYME Q
Ubiquinone, altanrativdy hown as coaizyme Q,, is a iiaturally arwring compownt
of the mitochondrial nspiratory chain.lM As discussed previously (page Il) , coenyzme Q
functions to transfer electrons h m complexes 1 and II to cornplex III. Several reports have
suggested that the endogemous leveis of coenyme QI,, are reduced in a wide varïety of
cardiac disorders, including patients with coronaf~ artery disease undergobg
fevaSc-tiOn. 11~11s Chen et al randomized 22 patients W ~ O undment cor~imry bypass
surgery to d v e Q, (1 1 patients) or placebo (n = 1 l ) . I u Aithough there were statistidy
signifiant improvements in mitochondrial ultrastnicture observed in biopsy specimens, ttiere
were no appreciable hemodynamic benefits. The Iole of coenzyme Q , as a cardioplegic
additive thus remains unclear.
DICHLOROACETATE
The stirniilntoiy efféct ~f dichloroaicetate (DCA) on pyruvate dehydrogenase acfivity
was originally described by Wtehoue and Randle in 1973.ll9 As d d b e d earlier, PDH

21
ngulates the amvemïon of pynmite to acetyl-CoA. Thus, stimuIation of this eazyme
cornplex may lead to incmued substrate lnnls for the -sr cycle. Substrate drivetn
stimuiatkm of oxidaîive phospharylaticm may lead to improved toIerance and recovery nOm
ischemia. DCA has beai used as a cardioplegic additive with variable effect. Wahr et al
demonsûated that DCA imp~oved giucose oxidation and led to better functional recovery
f o U d g ischemia.la In an aâcütional report, tbe same authors found that myocardial aiagy
charge was better preserved in DCA treated hearts and that DCA resuited in a reducticm in
both myocardial lactate release and NADH accumuiati~n.'~ niese authors concluded that
DCA resdted in improved f i r n c t i d recovery folïOmng global ischemia by stimulating
PDH. In contrast, Mazer et ai found that DCA did not improve rnyocardial sysblic function
dcspite a stimulation of aerobic carbohydrate metaboii~rn.'~ Thus, a cornpoumi which can
mimic DCA's stimulatory e f f î on PDH and also lead to impved functional nmvery
would be a usefiil additive to çardioplegic formulations. In orda to iden- such a
cornpond, a more detailed understanding of the ppvate dehydrogenase cornplex is
required.
1.2. THE PYRWATE DEElYDROGENASE COMPIZX
Sm<cnuc Ond Ration of the PDR Cornplex
Figure 3 demonstrates that pyruvate is an intermediary product of glycolysis. In
humans, ppvate is metabolized via four major eqmatic pauiways. Lactate dehydrogenase
WH) conveits pynivate to iactic acid as the final step in anaembic giyoolysis. q.niVate
carboxylase converts pyruvate to oxaioacetate, an intermediary Krebs' cycle component. In
the liva, alanine aminotransferase (ALT) regulates the reversible conversion of pyruvate to
the amino acid alanine. Pyruvate is converted to acetyl-CoA by pyruvate dehydrogenase

22
Pm*
The pynnrate dehydrogemse complex is a large macnm~oleaie (8500 kDa) which was
fust described by Reed and coileauges h 1 % P This complex teSides in the inna
membrane of the mitochoIlclria and is composed of sevexai distinct sub-units. The El subimit
is a tebramer of 2 or and B mmponmts each. This subunit acts to decarboxy1aî.e pynivate to
form CQ and thiamine pyrophospbate CrpP-El). The E2 subunit transfers the acetyl- gmup
h m TPP to a lipoy1 cofactor and then subseqwtly to cofactor A, forming acetyl-CoA.
Protein X also functions as an acyltransfaase, but its unique role in the complex is unc1ear.
The E3 subunit is a flavin requiring enynae which o x i a the iipoyl cofaaar h m E2 by
tmsferring electrons to NAD+. The wmplex is regulated by two additional enzymes: PDH
kinase and PDH ph~sphatase.~~ The PDH complur is illustrated in Figure 5.
Pynivaîe dehydrogemue kinase cataiyzes the phospharyWon of Saine residues in at
least two positions on the E l a subunit, raidexhg the complex inactive.'* It is postulated
that phosphorylation of a third serine residue is irreversible and renders the complex
insensitive to PDH ph~sphatase.'~ PDH kinase is inhi'bited by high oon~trations of
p p a t e and ADP. DichiorOQcefate is thought to act synergistically with ADP to inhibit the
PDR k h s , but the exact mechaniSm remains unclear." The PDH lanase is stimulated by
the end-products of PDH metabofism, niimely NADH and acetyl-CoA. mus, when the
mitochondrial NADH/NAD and acetyI-CoA/CoA ratios are high, the kinase is -Y
s t i r n a . Ravindran et al demonstrated that PDH kinase is regdatai by the redox state of
a lipoyi domain on the E2 subunit[= Ushg isoiateû PDH complex fnnn bovine kidney, they
found that compkte eaymatic delipoylation of the E2 subunit prevented NADH and acetyl-
CoA medi;ated stimulation of PDH kinase. Their pmposed mode1 suggests that acetyïation

23
of the L2 lipoyl domain of the E2 subunit produces a OOnfORnafiOnaf chauge tbat mhanc*i
biDdiDg to the PDH kinase. Restmhg the U Qmam to an oxidized form prevents PDH
kinase biading and subsequently preveats thePDII cornplex h m becoming phosphorylatpd.
AIthough each El-a subunit in the El tetramer h identicai, neariy complete iirnr?tivaficm of
the PDH compkx occrus if even one subunit is phosphorylsteA.'*
Pynivate dehydrogenase phosphatase rewses the E l u phosphorylation and retrans
the complex to the active state. Calcium and rnagaesium are re~uired by this enzyme f a
maximal activity.lB LUey et al found in bovine heart mitochondria that insulin pref-My
stimulates the PDH phosphafase.lm Larner et al fomd that incubation of epididymai fat with
insulin increased the sensitivity of PDH phosphatase to magnesium.*' Similarly, Newman
and colleagues found that isolatPri PDH phosphatase muid k stirnulated by incubation with
an insulin 'mediatora.lP In laboratory eovircmments, the PDH phosphatase is readily
inhiiited by sodium fluoride.
ïk Effect of Ischemia on PDH A m .
Mochizuki, and Neely demOIlSfrafed in 1980 that the recovery of normal giuaxe
oxidation in isolat& rat hearts subjected to global ischemia was delayed for over 20 minutes
foilowing normothermic reperfus~n.~ In this study, and in a preyious study by Liedtke and
N e l l i ~ , ~ ~ the infusion of pynivate during repahision substantiaüy irnproved the recovery of
mechanical function. Neely postulated that high cytosolic and rnitochondrial leveis of W H
and açylCoA following ischemia rnay act to inhibit the mitochondrial PDH cornplex?
Concomitant infusion of pyruvak prevented the inhibition of PDH and led to impved
fwictional recovery foïiowing ischemia.

24
gobayashi and Neely inv- the e&a of short tenn isdiemia and repemisiOn
on PDH activity in isoiated rat hearts subjcded to tm minutes of global ischemhul T h e
authors found that in hearts pemised with 11 mmoVL of giucose abne, ten minutes of
ischemia did not depress the acfivity of PDH. Appfolgmately 80% of the complex remaineci
in the active form in both control and ischemic h m . However, foîlowing 2 minutes of
repafuson only 45% of the enzyme d e d in the active fona. The addition of 10
mmoYL of pyruvate to the qafimte pmented the iaactivatcm of PDH.
The authon found that ischemia resuited in a 260-fold increase in the mitochondrial
NADWNAD ratio and a 43 96 reduction in myocardial ATP stores. Repfwion with glucuse
alone restored the rnitochondrial NADWAD ratio to near normal lm& and ATP stores to
72% of non-ischemic controls within two minutes. Repedhion with glucose and pyruvate
resulted in a more rapid resbration of mitochondrial NADWNAD ratios with a simiîar
r e c ~ ~ e r y of ATP stores.
Righ pst-ischemic NADHINAD ratios would be to stimulate the PDH
khi& and result in inactivation of the complex. However, the low mitochondriai ATP lm&
rnay have beea insuffiCient to allow for phosphorylation of the E-la subunit. As the celfulat
ATP stores became resiored following npemision, PDH kinase regain& the ability to
phosphorylate and inactivate the complex. In addition, the high leveis of ADP following
ischemia may inhibit the PDH kinase alIowing PDH to remain in its active fonn during
i s c h d Again, as mitochondriai ATPfADP stores are restored during reperfusion this
inhibition of PDH 14Mse would be lost, uniess high ambient concentrations of ppvate are
maintained with exogenous administration. Aitematively, the intraceUular acidosis which
accompanies ischemia may lead to activation of the PDH phosphatase which has a pH

25
optimum betweea 6.7 and 7.1 in the pregare of miignesi~m.~
Anadditidstudy by VaryetaP alsoJhowtdthPPDHactivity~asstuniilntPAby
no fiow ischemia. In thar isolateci rat kart model, these &ors found that pxeishemic
pemision with insuiin (20 IUL) causcd a f k t k bxease in PDH activity following 30
minutes of ischemia. Unforhiaately, they did not assess PDH activity during repafusim.
Howevex, k i r resuits suggest that pre-ischemic intemation an affect PDH d M t y durhg
ischemia. To date, the d t s of pn-ischemic interventions on the pst-ischernic activity of
PDH are unknown.
Lewandowski and White performed important eqexhents in isolatPA rabbit hearts
to detemiine the relationship between PDH inactivation and pst-ischemic fuactional
recoveqm In their experimental protocol, they stimuiaîed post-ischemic PDH actiyity by
the addition of 5 mmoYL dichloroace&te (DCA) to the repemiSare. In normal non-ischemic
hearts, DCA did not affect left ventri& function despite a stimuiation of PDH activity.
However, in pst-ischernic h m the rate-pressure-produe i@~antly from
83ûû*1800 to 21300*2400 when PDH acbivity was stimuiated by DCA treatnient during
repemision. Postischemic hearts displayed reduced pyruvate oxidation compared to non-
ischernic conbrois. DCA stimulated hearts increased the oxidation of pyruvafe without an
attendant in- in the by-products of glycolysis. Therefore, these authors concludeci that
countezacting depmsed PDH activity in the postischemic rnyocafdium preveated contrade
dysfunction. Furthemore, the imprwed cardiac @ormance did not d t h m 7 nor
require, hcreased glycolysis. Restaing carbon flux thrwgh PDH alone was suffiCient to
Unprove mechanid work by post-ischemic hearts.

26
McVeigh and Lopaschuk have s u g g d th& PDH acîiviîy is inhibiteci by fatty acid
fmm Qrculatirig fhe fatty acid me&boIism act to stimiilate_ the PDH kinase and thereby
"hibit giucose oxidation. In several reports, Lapaschuk has demonstrated that inhibithg
faüy acid metabolian by inhibition of d t i n e paimitoyltransfeaase (CPT) improves post-
Shortly afta the discovery of the pyruvate dehydrogenase complex in 1969," it was
show that insulin was capable of reguïating this enzyme. '&le in a review article published
in 1989, Weiland defined the characterisiic ffeatures of insulin's regdation of the pyxuvate
dehydrogenase complex in adipose tissue.'*
Insulùi activates PDH at physiological concentrations (K.: 10 mUL) Activation i s rqid and occurs in minutes. Insuiin activates via ppvate dehydrogenase phosphatas activation. Activation of the PDH phosphatase requires the presence of a metaboiizable sugar. Insulin does not change the rnitochondrial ATPIADP ratio. The activation pemists in mitochondria isoiated h m insulin-treated fat ce&. Cyclic AMP is nat responsible for the efféct of insulin on PDH. Insulin cannot be demonstrated to stimulate PDH in isolated fkt c d mitochondria, Insuiin activation of PDH has been dernonstrated in white and b r m adipcytes, fibroblasts, myocytes and lactating mammary giand, but not in liver*
Several of these statemmts require furth- discussion.
The dose-respcmse relationship betweai insulin and PDH bas beai extensively
evaîuated in adipose tissue and has km confirmed to'axw at physio1ogic concentrations (1-
100 mU/L,).161n However, in the sethg of acute ischemia, many investigators have
employed a much higher concentration of insulin (10-20 Un) in thar GIK fomuiatim.

27
The possible dmmguMhg eff8Ct of high iiwulin concentrations has not beai adeqiiately
daemhed. High doses of insulin may be rrquind to Jtimulate other protective &ectS, such
as reducing fatty acid oxidation. incfeasing glucose transport and reducing afterload.
Therefm, it is important t~ detamine if high doses of insulin continue to stimulate PDH
nie t h e course of the stimulatory &éct of insulin suggests Uiat it works on a post-
translational basis. That is, insulin does not require the synthesis of novd mRNA or proteins
to aext its action cm PDH activity. In seüings of impending ischemia, such as d o p l e g i c
arrest, it would be important to exert a stimulus which acts hmediately and persists during
the repemision phase. As mentioned previously, it is unclear if pm-ischemic insuiin treatment
GUI iesult in pst-ischemic PDH stimulation. Shuües on mitochondria isolated from fat d k
exposed to insulin suggest that the insuiin &ect can pasist for up to 30 minutes." Randle's
p u p have also examined a possible long-terni regulatory effect of PDH activity.ja
Conditions of chronic hypoxia, such as wngenital cyanotic heart disease, may be ameaable
to long-km PDH stimuIarion by pharmacoiogic additives. Recent work by Meranfe1#
mgge~ts thaî the aictivity of mitochondrial PDH and cytochrome oxidase is inhibited rapidly
foI.lowiag exposure to low partial pressures of oxygen @4 = 40 m d g ) .
Insulin has been demonstratsd to stimulate bah a metai dependent and independent
PDH phosphatase by Lanier's group."'' Lamer initially demonstrated that insulin acted by
reducing the PDH phosphatase's requirernerit for divalent cations (calcium and magnesium).
In a subsequent investigation of isolatpA bovine mitochondria, Lamer's group detected a
metal independent PDH phospiqtase which was not inhibited by okadaic acid, not stimulated
by spamine and not YnmunopreQpitated by antiphosphatase 2A anh'body.lD This novd

28
phoephatase demcmstrated a ttveefoId ina*ue in activity foliowing insului exposure.
Thcabilityofinsulin toregulatcthcPDHkhaseisiike1ydependmtuponchangesin
the mitochondrial ATPIADP and NADHINAD ratios. Howeva, Denton's group
demonstrated in isotateA white fat mitocbondria that iasuün treatmmt did not resuit in any
changes in ATP, ADP, NAD, NADH, acetyl-CoA or CoA which would result in inbrl'bitim
of PDH tjlia.Pe,.Ia In contrast, a shidy by Hughes a al demoastrated ùumzsed W4-
incofporatiion in mitochondria exposed to insuiin, suggesting incrrased activity of the Ianase
suôunitJB The authcm rationalized the apparent contradiction betweai increased PDB kinase
activity and the ovedi increase in PDH activity by claiming that insuiin acts to stimulate the
dephosphorylation of the cornplex which in tum leads to an inaeased turnover of the PDH
phosphoryhtiondephosphorylation cycle. If insuiin acts by inhibithg the kinase subunit,
then an ovedl decrase in tumova would have been expected.
Ihe Rolc of a Second Messenger
Much interest has centred on a possible second messetlger system responsible for
rndatbg insulin's effm on PDH."-1~'3'*14'a1s1*18 GottschaUc demonstirated in two sepiiiate
cell lines, rat embryonic fibroblasts and chinese hamster ovas, cells, that insulin' s stimulaMy
effect on PDH activity bypasses the insulin receptor tyrosine kinase? In œlls made
deficient of the insuiin receptor, PDH activity was StimulatPrl to a smilar extent as œlls
which overexpressed the insulin m t o r . However, Gottschalk employed transfection
vectors expressing normal human insului teceptors or receptors with inactivated tyrosine
kinase domains. Native insulin r e c e p a activity rnay have sti l i accounted for the stimukt~ry
e f f e on PDH activity. In ceb which overexpresseù the human insulin reœptor, saturation
of a second messenger system may have limiteci the stimulatory effect of insulin e n p o s ~ ~ ~

29
Discovery of a putative .insulin mediatar" bas thus iàr eluded investigators. Saltiel
proposed that these mediator molecules may be inosif01 phosphate glycans. One of these
phoephotidyl inosito1 glycans has bem s h m to d t in & lu^> synthesis of dpcvlgîymal
@AG).m DAG is a hown stimulator of praein kinase C, a cornmon second messeaga
presait in most œiî typeda S e and coUeagues Qnonstrated that insulin stimuiated
phoBphatidylinosito1 giycan hydrolysis with subsequent & mw syathesis of DAO and PKC
activaiion in BC3-HI myOCYfeS.m However, inhi'bition of G-protein activation by pertussis
toxin did not inhicbit insulin's ability to increase DAG concentrations and stimulate PKC.
These authm concludeâ that novel phosphatidylcholiae hydro1ysis was responsible for DAG
synthesis and subsequent PKC transidon and activation. Craven and DeRuberth aiso
demonstrated that exposure to high glucose amœntration results in & now synthesis of DAG
and incteased PKC activity.la
Benelli et al demonsûated in CUItured 2ajdela hepatoma tells that insulin activated
PDH via a PKCdependent pathway.la In this model, insulin was found to stimulate PDH
within five minutes of incubation and was maximal (70% increase over baseline) at 7.5
minutes. In the presence of the phorbol ester PMA (4fbphorbol 12B-myriState 13a-acetate),
PDH activity was increased within 3û seconds, achieved maximal activity (90% wer
basehe) at 5 minutes and was no longer detectable at ten minutes. In addition, incubation
with insulin and the PKC inhibitors staufospo~e and sphingosine wmpletely blocked the
stimulatory effect of insulin.
However, the d e of protein kinase C in the insulin signaUing pathway continues to
be c o n t r o v d . DowmguMion of PKC with chronic phorbol esta exposure does not
inhibit insqlin-induced glucose transport in B3CHl rny~cytes.'~ Similarly, KEp and Rarnlal

30
demOQlStrafedthatpn)teinlMaseCwasnatrequirrdfmkosetransportincultiaedslteleéal
muscle cells.la Stumpo and BiacWear demcmstrated that c-far expression in 3T3-L1
fibroblastswasina*isedafkinsutinaposun, eveniftheœiisweremadeprofeinkirÿise
deficiat.lm Both cfos and c-myc are eariy respaue peptides which have kai show11 to be
stimiitatPA by PKC.lQ The rde of protan kinase C in the insulin signalhg pathway
wntinues to be inMstigated predominantiy in adipose tisoue or non-cardiailc myocytesm.
Establishing the relationship between PKC and insulin in ischemic mya'ardial tissue is of
paramount importance in understanding the mechaniSm of the cardiop~otective e f h t of
insulin.
1.3. THE ROLE OF PROTEIN KINASE C
Signal Tzcurrducn'on P d m q s
Sutherland first proposed the model of a second messeriger system in 1972.1a In this
model, extracellular signais termed ligands either petrate the ceii membrane or bind to
membrane bound reqtors. Activatexi feceptOrs with or without an associated membraneœ
bound transducer (such as a G protein) trantranslate the 1igand-reçeptor binding signal into an
intracellular chemicai message. Most o b , this involves changing &ha the distribution or
the ancentration of " second messenger" compounds.
The insulin signalling pathway has been one of the most actively studied rnammalian
systems since Banting first described the hypogiyœmic effécts of this purified pr0tein.I" The
bovine and porcine hormones were sequenced by SangeP and cowmkers in 1955 and in
1963, Katsoyannis d d b e d the fgst chernid synthesis of purified insulin.In As M e r
information about the structure and function of human insulin became avaüable, many
investigaiors simultaneously concluded that insulin's inûaœiiuiar effezt involved the

Figure 6 ciescri- the struchin of the human iasulin LeceptoT. The insulin reaptor
cornpiex is a tetrameric transmembme giycoprntein consisbing of two 120 kDa and two
90 kDa B subunits.lp The two a-subunits are h k m i by disutphide bonds, are entirely
extraceiluiar and amtain the insuiin binding sites. Each &subuait is linlred to an a-subunit
by a disiilphide bond, crosses the piasma membrane and amtains a tyrosine kinase domain
in its intracellular portion. nie association of insulin protein with its ~~cep tor stimulates the
tyrosine kinase activity of the Bsubunit and d t s in autophosphorylation of the second &
subunit Autophosphorylation of the &subunit stimulates the kinase activity of the receptor
towards 0 t h substrate pk ins , including insulin receptor subsûate 1 @tS-l).m The insulin
receptm is capable of binding 1 or 2 molecules of insullli.Is The unoccupied a-subunit
exerts an inhibitory effed on the tyrosine kinase activity of its amespondkg Bs~bunit.
Proteolytic cleavage of the a-subunits relieves this inhibition and resuits in a stimWon of
kinase actiVity.lB
IRS-1 is a highly conserved Cytosolic protein with multiple serine/threonhe d d u e s .
Interestingly, IRS-1 contaias over 30 potential serine/threonine phosphoryîatim sites which
have homologies to protein kinase C. ln Prior to insuiin activation, IRS-1 is highly serine
phosphorylated and weakly threonine phosph~ryhted.'~ Foilowing insulin-reœptor bindùig,
there is an increase in both serine and Uinonine phosphorylation." Sinœ IRS-1 shares
homologous domains with a wide variety of intraceUular messagers, including PKC, it is
likely thaî insulin stimulation of PKC is associated with the phosphorylation of IRS-l.In
However, this association has not yet been demollsfrated.

Req tm mediattd hydrolysis of membrane-bound phosphatidyluiositol gïycans is now
realird to be a cornmon signal transduction mec)iiuii.(wn in mimy ceil types and organ
systerns-" S tnh and colleauges demOllSbafed that Mtol - l , 4 ,5 triphosphate p3) is reieased
into the cytaphm following hydrolysis of membrane bmmd phosphatidyl4,5-biphosphate
(PIPa and ùiduœs the mobibatiion of caicim fiom inbraicellular stores.1x 'Lbaeafter, Takai
repmted Uiat anothex product of PIP, hydrolysis, diacylglyœml, initïates the activation of a
s p a h h d protein Linase.lW This kinase had been previously ideiitified by the same group
in 1977'- to be protein kinase C and has since been show to be present in most cell types
and organ systems. '"MI
In its W v e f m , protein kinase C is widely dispased in the cytopIasm.
Stimulation of PKC resuits in its transiOcati011 to peMucIear and ceU membraneP
Translucaîion of PKC is associateci with binding to Receptor for Activated C-Kinase Subtype
(RACKS) which is present on cytoskeietal elements and the ceil membrane-lm Referatiai
binding to RACKS is thought to fiditate the isoforni-specifïc functions of PKC for dinerent
proteins." Although translocation of PKC from the cytosol to the plasma membrane is often
associated with stimitlation of the enzyme, it may not necessarily result in an increase in
kinase activity."
Isofom Speciic Pr~pem'es of Protein Kinase C
There have been numerous isofanns of protein kinase C identifd.'" AU represent
kndthreonine kinases with a similar molecular weight (68-83 kDa), however each may
play a different d e in the ceiluiar response to extenial ~tirnuli.'~ lbgoyevitch and
coUeagues descfibed the expression of numemus PKC isofoms in adult rat hart and

33
m e d that by fâr the most abundant was the eIsooform.m The otha isofarms preseat
in heart include the B, a, 8, eta and zeti isofann~.~*
NishUulrals Onginal description of the m e s of PKC suggested that it was
dependent. Based on these differe31t isofmm-speafic characterista, a ciassification system
for the PKC i s o f m is now commonly used in the 1iterature.l" The calcium dependent
isofarms or conventional P K ' s (cPKC) include the a, 8 and gamma subtypes and are
adivated by calcium, phosphatidylserine, dkylglycerol and phorbol esters such as PMA.
A s g ~ d group is comprised of four novei PKC's (nPKC), 6, E, eta ande. This group is
actiMtcd by phosphatidylsaine, diacy1glyerol and phorbol esters, but not calcium. A third
group includes two atypical PKC hforms (aPKC) zeia and lambda. These isoforms are
actiMted by phosphatidylserine, but not by diacyigiycerol, phorbol esters or calcium. The
zeta f m rnay exert a constant low level of activator-independent kinase activit~.'~ In
addition to their different responses to extemal and intanal stimuli. these isoforms may be
responsible for differential effects within the œil.In One manifestaiion of this property was
descnbed by Banerjee and colleagises who provideci evidenœ that a-adrenergic stimulation
d t e d in translocation of the a, 6, eta and zeta isofoms while the 8 and c isoforms were
unaffected.ta Stimulation with a DAG d o g u e resulted in a translocation of the a, 6, eta
and zeta isofoms with the r i s o f ~ ~ n translocafing to the perinuclear membrane. However,
a-adrmergic stimulation proîected against ischernic acidosis in a PlCCdependent manner
while exposure to the DAG analogue did not prevent acidosis. Thus, it appears that
inhibition of the E-isoform resuited in protection against acidosis, but this inhibition was
dependent upon stimulation of the a, 6, eîa and zeta isoforms.

whose &ty was stimuhd by the hyQolysis of membranebound phosphatidylinosito1
g l y c a n ~ . ~ ~ The naturally OcCuring by-product of PIP, hydroIysis, diaçylglycerol @AG),
is a potent stimiilatnr of p t e i n Linase C and has given rise to several synthetic analogues
such as l-oleoyl-2-~1glyycerol (OAG), l , 2 d i ~ o g i y c e r o I (DûG) and stearoy1-
arachidmoyl giyceml (S AG). la
Tumm pmmoting phab01 estas such as 4BphoroOl 12Brnyristate 13~-acetite
(PMA), have a structure which is very similar to that of DAG and also activafe PKC.'?
Howmr, uniike the rapidly metabolized diacylgiycerols, the phorbol esters are slowly
degradeci and can resdt in eventuai downregulaton of PKC activity with prolongeci
exposure. lalm
Inh'bition of PKC can be accomplished by a variety of biochemicai mechanisms.
Staurosporine and H-7 both inhibit PKC by intaference with the ATP-bùiding site of the
enyme.lW Unfortunately, these compounds iack specificity for PKC and can also interfere
with the ATP binding sites of other kinases such as protein kinase A, tyrosine Liaases and
calmoduiin-depaident kuiase. However, che1erythMe qmsents a PKC specific inhibitor
which aiso works by blodang the ATP binding site of the enzyme.'"
As mentioned previously, transiocafion of PKC is thought to muire stnictural
cytoskeletal elements such as micro tubule^.'^ Therefore, agents such as colchicine which
inhibits microtubule synthesis may be usai as a relatively non-spxiiic P K antagonist.
However, other cellular enzymps requiring cytoskeletal association may a h be inhibited by
colchicine. Interference with the lipid cofactor binding sites of PKC urn produce highly

In 1986, Miary, Jamings and Ramt published a landmark shuly in which they
reportcd that the magaitude of canine myocardial hhction ptoduced by a 40 minute
circumnex coroiiary artery occlusion was sipifkmtiy r e d d whm the myocardium at rislr
had previousiy been subjeded to four cycles of 5 minute wronary occlusion and 5 minutes
of qxxfhi~n.~* This phenornenon was termed "ischemic preconditioningW and may
represent the most potent form of endogrnous myocardial protection discovered to date.
Preconditioning has now been d & M in a wide variety of organs and multip1e
species.'" The preconditicming ~esponse ha9 been reported to occur in human
r n y o c a r d i ~ r n . ' ~ ~ ~ konomidis and coileauges have demonstrated that isolatPA cultures of
human ventricuiar cardiomyacytes can be protected from 90 minutes of prolonged ischemia
if they are first subjeaed to twenty minutes of brief ischemia followed by twenty minutes of
reperfusim. loner and coUeagues reviewed the d t s h m the TIMI 4 trial. IS In
patients who were admitted with a diagnosis of acute myocardial irkction, an anginal aîtack
in the preceduig 48 hours prior to admission led to a decreased incidence of in hospitai &th,
severe congestive heart Mure or shock. In addition, these patients exhibited las myocardial
damage as evidmced by CPK levels and eleçtrocardiographic abnodties.
Yelion and coïkagues reported the first experience of pfeconditioning for cardiac
~urgery.'~ In this report, brief aortic occlusion and tel- pior to prolonged aartic
crosscîamping led to irnpfoved preservation of high energy phosphates in atrial trabecuiae.
However, this fonn of myocardial protection is unlike1y to be adopted by many surgeons due

36
to the additid risk of multipIe aortic manipulatons and the poBsibüity of atheroscleriotic
embli." Eauciclating the mechaniSm of kichemk prrconditioning has ken the focus of
group in CofOziadO bave also demonsüated that PKC is required for ischemic preconditioning
of the rat heart.'n.mn Ikonomidis and coileagues have dernollstrafed that preconditicming
of isolated human ventricular cardiomyocytes is aiso dependent upon protan kinase C and
is mediated by the Aease of aden0sine.l
To date, a number of compounds have been demonstrated to rnimic the effect of
"ischemic" prec~~lditioning including adenosine,- bradykininm and a-adreaergic
agents. 'W.'" Each of these stimuli, in tum, bas been shown to be dependent upon PKC
stimiilati0n.'- The end efféctor of PKC stimulation is stül disputecl. However, besed upm
the many rne&hlic effeçts of premnditioning, it is reasonable to poshilate that PKC
stimulation may d t in a regdation of cellular metabofism.
Miyamae et al demonstrated in a porcine mode1 that ischemic preconditioning resdted
in a better preservation of rnyocardial ATP, phosphocreatine and intracellular pKn
However, this beneficial metaboiic eR't did not translate into protection against stunning.
Similarly, Ryzkledc and Kloner's group demonstrated in a canine mode1 of myocardial
stunning that ischemic preconditioning was unable to exert a beneficial functional e f f a m
Banerjeets group feported preiiminary data suggesting that ischemic preconditioning
may act to p e n t cytochrome induced œiiuiar ~x ida t ion .~ A report by van Wylm
indicated Wt ischemic pmconditioning aîteniiatprl the accumulation of intracelluiar iactate and

37
intedtial purine metabolites during wheqmt prolmged ischemhaO Wolfe and colleagua
providsa evideace that suggesfed that pncaditioning d t e d in pischemic giycogea
depletion which penteci subsequent intraceiiular acidosis during plonged i s c h d m
Hcwever, this d t is in contradiction to the results obtaïned h m Taegtmeyer's groiip who
demoiistrated that preischemic giycogen depletion did na improve functionai recovery a€kr
hypothdc ischemic amst in a rabbit h a u t m
U n f e y , the end-effectar mecbanism fm the ischemic preconditicming
phenornenon remains unknown. However, the attenuation of intraceUular acidosis with a
reduction in lactate accumulation and pRsenraton of myocardial ATP strongiy suggest that
preconditioning ex- a beneficial effect on myocardial metabolism. Due to the questionable
abiiity of ischemic preconditioning to faalitate the fecovery of left ventricuiar fiinction, it is
likely that the preservation in ATP emanates from an aMerobic source. As a coroiiary to
Weiss' theory on the funaionai c o m p a r t m e n ~ o n of ATP prod~ction,~ it appem that
anaerobic ATP presenration h m ischemic preconditioning is preferentially used for ceiluiar
homeostasis. The degree of aembic ATP production is probably insuffiCient to allow for
functional benefit. Therefm, metabolic stimulation of aerobic myocardial ATP synthesis
in combination with a preconditioning stimulus may be an ideal combination to pro-
against both myocardial infarcton and stunning.
1 4 SUMlMARY OF PROPOSED INVESTIGATIONS
Contemporary techniques of perioperative myocardial protection continue to d t in
aelayed recovery of normal aerobic rneotboli~rn.~ Pmnous studies evaiuating the effed of
giucose and insulin soIutions for penoperabve myocardial protection have been
~ntroversial.~ ui addition, these stuclies were performed in an ad of hypothennic

38
m y d and systmiic m o n which may have attmuated the benef~ciai e&cts of inSulni
treatment. Many gtuciies ernployed intravenous gIu00seinsulin-potassium sotutions and did
not examine the &8d of dind myocardial pemisicm with insulin-enhanced cardioplegia
delivemi at normothermic or tepid (29°C) temperatines.
Myocardial PDH ha3 bem demoIlSfrated to be inhibitecl during eady rrpafusion
foliowiag brief ischemia in isoIntprl nit h m r n The &ecfS of ischemia and repafusion on
hrmimr myocardial PDH remain undetermbed. Insulin has been shown to stimulate
mitochondrial PDH in adipose tiss~e,~'*'~ however the ability of insulia to stimulate
myOCOrdiQl PDH remains undetermineci.
The proposed investigations are designed to:
1. Evaiuate the effed of ischemia and npafusion on human myocardia pynivafe
dehydrogenase activity.
2. Evaiuate the effects of pre-ischernic insuiin exposure on pst-ischemic myocardial
3. Detennine the mechanisrn of the insulin effect on pst-ischernic myacardial
4. Evaiuate the diffefential effects of pischemic insulin exposure on hurnan ventricular
cardiomyocytes and human saphenou vein aidothehi celis.
We hypothesize that ischemia and repemision wiii inhibit the activity of myocardial
pyruvate dehydrogenase. We believe that pischemic exposure to insulin wiU preverit the
inhibition of PDH by a protein kinase C dependent mechanism. An improved transition from
anaerobic to aerobic metabolism foilowing prolongeci ischemia is believed to improve cellular
tolerance to ischemia. Furthemore, the metabolic effects of insuiin expomre are

To f'acilitate these investigations, we propose to employ cuitulres of isoIsitpA human
ventricufar carriiomyocytes and saphenous vein enQthelinl cells. The d t s of thest
isvestigations wiU provide important new informaficm &out the metabolic abnofmalities
associated with myocardial iscbemia In addition, the in vino evaluation of a simple, safe
rnetaboiic intervention would provide jdcaf icm fa UI viw investigations which uitimately
may lead to supgior myocardial toleranœ to ischemia and improved clinicai oufcofnes
following cardiac Surgery.

Figure 1: Myocardial oxygen wnsumption (MV03 in the empty beating, fibrillating, and a& heart at normothermia (37'C) and moderate hypothennia (22'C). M V Q is markedIy reduced following cardiopIegic*arrest with a further minor reduction associated with hypothefmia. (A@fredfrom Buckberg et al.')


GLUCOSE
2 ADP
LACTIC ACID
36 ATP
FATTY ACIDS
Figure 3: Myocard'i glucose metabolism.
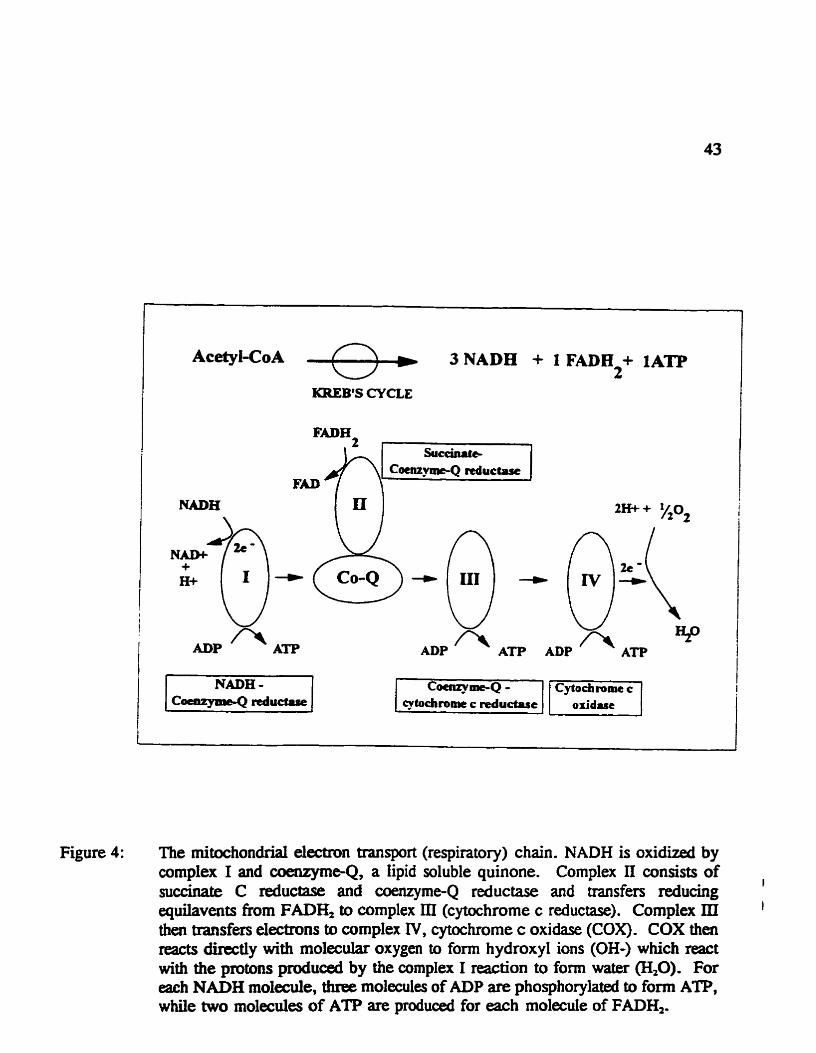
KREB'S CYCLE
FAD
+ H+ Nm3 ADP f i
Figure 4: The mitochondriai electron transport (respiratory) chain. NADH is oxidized by complex 1 and coenzyme-Q, a lipid soluble quinone. Complex II wnsists of
I succinate C reductase and coenzyme-Q reductase and transfers reducing quilavents fkom FADE& to complex III (cytochrome c reductase). Complex III I
then transfm electrons to complex IV, cytochrome c oxidase (COX). COX then reacts directiy with molecular oxygen ta form hydroxyl ions (OH-) which react with the protons produad by the complex 1 reaction to form water (H,O). For each NADH molecule, Uuee molecules of ADP are phosphorylated to form ATP, while two molecules of ATP are produced for each molecule of FADH,.

PDH PHOSPHA'IASE - PDH KINASE
Figure 5: The pyruvate dehydrogenase cornplex. The El subunit acts to decarboxylate pynivate to form CO, and thiamire pymphosphate (TFP-El). The E2 subunit transfers the acetyl- group h m TPP to a iipoyI cofhcto~ and then subsequentiy to cofactor A, f d g acetyl-CoA. The E3 subunit is a flavin re~uuing enyzme which oxidizes the lipoyl cofâctor h m E2 by t ransfdg electrons to NAD+. The complex is regulated by two additional enzymes: PDH kinase and PDH phosphatase.

+ Phosphoryhtion of Intracellulrir Peptides
Figure 6: The insulin receptor wmplex. The insulin reœptor cornplex is a tetrameric transmembrane glycoprotein consisting of two 120 kDa a and two 90 kDa 8 subunits. The two a-subunits an linlred by disulphide bonds, are entirely extraceiiuiar and contain the insuiin binding sites. Each Bsubunit is linked to an a-subunit by a disuiphide bond, msses the plasma membrane and contains a tyrosine kinase domain in its intracelluiar portion. nie assoaation of insulin protein with its receptor stimuiates the tyrosine W d v i t y of the &subunit and resuits in autophosphorylation of the second Bsubwilt. Autuphosphorylation of the Bsubunit stllnuiates the kinase activity of the receptar towards other substrate proteins, including insuiin receptor substratel m-1)-

CHAPTER 'Iwo INSULIN STIlMULATlES MYOCARDIAL PYRUVATE DEKYDROGENASE
AND PROTECTS ISOLATED HUIMAN VENTRIcnn,aR CARDIOMYOCYTES FROM SIMULATED BCHElWIA

However, the effect of insuiin cm human myocardizl PIH remains mdetermineû.
Furthermore, the effed of ischemia and repafusion on the activity of human myocardial
PDH rernains unlaiown. Kobayashi and N d y m demanstrated that npahision following
brkf ischemia resuited in a 55% inhiiitim of rat myocardial PDH. This inhibition was
thought to be due to stimulation of PDH Iànase by accumutated NADH and acety1-CoA in
the presence of adequate intracelluiar ATP stores. LRwandowsLi and WhiteU provided
&dence in an isolaîed rabbit heart mode1 that the recovery of PDH activity dinectiy
correlated with the rrcovery of ventricular fiuiction. Therefore, attempts to stimulate PDH
acfivity and improve the transition from aMerobic to aembic metabolism foliowing ischemia
should improve œiluiar tolerance to ischemia and result in enhanaxi functional fecovery.
To investigaîe the abiüty of insulin to stimulate human myocardial pynivate
dehydrogenase, we employed a mode1 of simulateci ischemia and reperfiision using primary
culaires of isolated human right ventricular cardiomyocytes. This model has several
advantages for the investigation of cellular responses to ischemia and feperfusion. Firstly,
the effect of other ceii types such as endotheliai d s , fibroblasts, smmth muscle cek and
migating neutruphils are absent in this model. The effect of other organ systems such as the
liver, pancreas, brain and pezipheral vascular system are aiso absent. In addition, the
extraceiluiar environment can be carefully controiied to eliminaîe the effects of altemative
substmtes such as fatty acidsJ" and to inüwiuce metabolic interventions such as insulin or
glucose. The nature of the cuihired cardiomyocytes has been extensiveiy reviewed in

cardiomyocytes, these tells retain many charaderistics of àr vnio rnyocardium and provide
a useful model for the investigation of injury due to ischemia and repafusion. However, the
d t s re~uire confirmaton in a whoie organ or m I&O model.
2.2. METHODS
Hrcnion VmniaJor Ccvdiomyayte aclavc
Rimary cultures of human ventricular cardiomyocytes were established as previously
d e ~ c r i b e d . ~ ~ ~ ~ Brkfiy, 20 mg biopsies were obtained h m the nght ventricle of patients
undergohg caective repair of tetralogy of Faiiot (Courtesy of Dr. W.G. Wilüams .-
Hospital for Sick C h i l a , Toronto). The myocardial biapsies were washed in phosphaîe-
buffered saline without calcium (PBS: NaCI 136.9 mM, KCl2.7 mM, Na?Hp04 8.1 mM,
lZ&P04 1.5 mM). AAcr removing amnective tissue, the myocytes were separated using
aizymatic digestion with a mixture of 0.2% trypsin (Difco Laboratories; Detroit, MI) and
O. 1 96 collagenase (Worthington Biochernicai Corp. ; F d o l d , NJ) . The isolated d i s were
culturecl at 37T in 5% C 4 and 95% air in Imve's modified Dulbecco's medium (GIBCO
laboratories; Grand Island, NY) containing 10% fetal boWK senim, 100 U/mL pallcillin,
100 pg/mL streptornycin and 0.1 m M &mercaptoethanol. Purification was achiwed using
the dilution cloning technique. Enzymatically isoiated cells were seeded at a low density ( S e
100 d i s per 9 cm diameter culture dish). At this low density, individual cardiomyocytes
were distinguished morphologically by their rectangular shape and large 9ze (40x80 cm)
h m con taminating œlls such as fibroblasts and endothelid œlls. AU cells on the plate were
aUowed to divide and at 7 days cardiomyocyte, fibroblast and endothelial colonies formed.
Single cardiomyocyte colonies were thm transfernd usuig a Pasteur pipette to another cuiture

49
dish. Cell cuitures were uupeded daiiy and any conbmbatd cuiture dishes w a e discarded.
Culture purity gnater than 95% was afta the third cell passage with fi-t
monocIanal ant i i iy stainïng for human ventricular myosin heavy chain (Rougier BieTech
M.; Montreal, QUE). Ce& passage! 2 to 7 tims with a time h m primary cuiture of less
than 60 days were used for this shdy. Figure 7 ïiiustcates the phenotypic appearance of these
cardiomyocytes at 2ûûx magnincation.
SmuJcltcd Isahcma anà Repctjksbn
The cardiomyocytes were studied in PBS with rnagnesium and calcium WgC&
0.49rnM, CaCI, 0.69m.M). The addition of these divatent cations ensured th& PDH
phosphatase activity wouid not be inhibiteci (see page 23). In addition, œiis exposai to PBS
in the abserice of calcium lose their ability to adhere to culture dishes after approximately 30
minutes. Our in vin0 technique to simUfilfP. 'ischemia' and "repemisona has been
previously described in detail." Briefly, cells were stabikd for 30 minutes in 10 mL of
normoxic @O2= 150 mmHg) PBS. *Ischemiaw was simuLateci by placing the ceils into a
d e d plexigiass chamber fiushed with 100% nitrogen and exposing the celis to a low volume
(1.5 mL) of deoxygenated PM. Deoxygaiated PBS was prepared by degassing norrnoxic
PBS with 95% N, and 5 % CO, until the rneasured p Q reached O mmHg. 'RepemiSon" was
accomplished by exposure to 10 mL of normoxic PBS for 30 minutes. A srnail sarnple of
deoxygenated PBS (2 mL) was placed into a centre di& in the chamber to rnonitor
temperature and to con- the absence of oxygen at the end of the "ischemic" period. The
temperaaire was maintained at 37C throughout aU experimental pmtocols. The osmolarity
of the degassed PBS solutions was maintained between 280 and 320 mOsm/L using sodium
chloride or water as required. The pH of aU PBS solutions was mâintained between 7.35 and

50
7.45 with 1M hydrochloric acid or 1M Wum hydroxide as quired. Figure 8 is a schematic
mpmeatation of the equipmeat requind to simulate ischmlla and npezfkion. Nimgen gas
is bubbled through two oxygen trapa pria to flushing the seaied plexigiass chamber. A
singie port on the chamber is lefi opm to vent the c h b e r . The plaiglas cbambex is
phcd on an eiectric heating pad which raciiitaks the mainteriance of normothermic
te-
EqerUnental Pr0ta:oIs
During the minute stabïüzation period, ceils were exposai to varying
co~lcentratious of glucose and insulin. FoiIowing stabitiÿaton, each di plate was washed
with normoxic PBS without glucose or insulin prior to the ninety minute ischemic exposure.
Similarly, following ischemia d plates were washed and repemised for thi.rty minutes in
normoxic PBS wïthout any additives. Fo11owing a dose response analysis for both gîucose
and insulin cm cellular survival, we chose to cumpare four groups. A physiologie glu-
concentration of 5 m M was compared to a glucose concentration of 100 mM. The latter
concentration agproximates the glucose concentration in the cardioplegic solution currently
employed at The Toronto Hospital. nie effects of 10 IU/L of insulin was then evaiuated at
both leweIs of glucose. (Gmup I : 5mM gluc~se; G m q 2: 5mM glucose and 10 I U L insulin;
Grwp 3: 1 0 mM glucose; Group 4: 100 rnM glucose and 10 IüL insulin).
Assessntenr of C e Z I ~ l .
Cellular injury was assessed using nonanfluent cultures of cardiomyocytes
(approximately 337,000 œlls per 9 cm diameter culture dish) cuitured for 4 to 5 &YS aAa
the iast passage. Following the intervention of interest, ceil plates were incubated with 0.4%
trypan blue dye (Sigma Chernical Company; St. Louis, MO), dissolved in normal saline and

Mississauga, ON) at 2OX magnification. I n j d tells were unable to exclude the large
molecular weight dye and stained blue. The aumber of blue stained ce& was counted fiwi
five srandard locations on each plate and expres& as a perœntage of the total number of
cells. AU cuunts w a e p a f d by a single obsedver who was blinded to the intervention.
Fi- 9 displays nepresaitative microgrciphs (Zûûx mgnScaticm) asseshg cellular injury
foliowing normoxic stabili7ation and afta ninety minutes of ischemia and thirty minutes of
repemision.
BiOchemicd M m -
Connuent cuitures of cardiomyocytes (approximately 600,000 ceils per culture dish)
culaired for 5 to 10 days from the last passage w a e used for biochemical d y s i s . The
activity of pyruvate dehydrogenase (PDH) was rneasured afkr each intervention of interest
using the modifications describeci by Robinson et aim of the method of Sheu et al? Ce11
extracts were aliquoted into separate Eppendorf tubes containing PBS. Foilowing incubation
for 10 minutes, ceils were treated with a buffkr containkg 25 mM sodium fluoride and snap
fkozen in Iiquid nitmgen. Ceil extracts were then reacted with a '%-pyruvafe containhg
b a e r in an open Eppendorf insert and piaced in Sealeci containers containhg benu,thonium
hydroxide to trap "COz. The reaction was stopped with 10% trichloroacetic acid and "CO,
wllected for one hour. Following "C4 wilection, the 4 inserts were removed and
scintilfation fluid added to the exposed bnizothonium hydroxide. The co1Iected I4CQ was
then counted in a Bcounter. PDH activity was calcuiated a f k correction for protein content
and expressxi as nmol of pyruvate oxidized per mg of protein per minute.

52
In a dupliciaîe series of experimeats, hfmcdlular and exfmdlular lactate
accumulation was rneasured using an m c me&d (Stat-Pack rapid lactate test kit,
Behriag DiagnoiCS; La Job, CA). Briefly, lactate is converted to pyrwate with an
equimok reducfion of NAD to NAM. Pynivatc is trapped by conversion to L-alanine by
alanine aminotransferase. The amount of lactate in the sample is directly Praportional to the
amount of NAD r e d d to NADH. The concentration of NAD is ckrmined by meaJuDng
spectrophotornetric absorbante at 34û nm. htracellular and extracellular lactate
concentrations were thai corredcd for the DNA content of each plate.
In a separate Senes of experiments, extraœiluiar lactate and pynivate were measured
in tandem in the supernatant of each plate. Supeniatants were wiiected and treated with a
measured volume of 6% perchloric acid to inhibit emymaîic degradation. Lactate
concentrations were determineci as d e s c r i i above. Pyruvate concentrations were
determinexi using a lactate dehydrogenase (LDH) couplad remion. Briefly, ppvate content
was determined fluofornetncally by measuring the change in NADH as LDH converted
pynivate to lactate. The lactate/py~vate ratio (L/P d o ) was thai calciilatprl for each plate.
Lactate and pyruvate concentrations were coneded for the protein content of each plate using
the Bradford spectrophotometric dye &on with bovine semm albumin as the standard?
The concentration of hydrogen ion w] in the extraceilular fluid was determined by
mnverting the pH value measured using a b l d gas anaiyzer (1312 Blood Gas Manager,
Instmmmtation Laboratory; Milano, Italy) tu [H+ J by the formula= [R+] = antilog (-pH).
Confluait plates of cardiomyocytes were used to determine cellular adesine nucleotide
contents a f k the intervention of interest. The specimens were fiash frozen in liquid nitrogen
and thai fheaAried. Specimens were anal@ by high performance iiquid chromatography

53
with the modifications described by W U et P of the step gradient technique &elopcd
by Hull-Ryde et al.= Using thh me2had, we demmined myocardial ccmcmtrations of
ademshe triphosphate (ATP), aderioshe diphosphate (ADP) and adenosine monophosphate
(AMP). W e also measured creatine phosphte (8) and the metabolites adenosine (ADO),
inosine (INO), hypoxanthine and xanthine 0. Total adenine nucleutides wete
de&mhed as the sum of ATP, ADP and AMP. Total Qsradation producfs P P ) wexe
calcuîated as the sum of ADû, INO, HXN and XAN. Energy charge (EC) repre~ents the
utilizable high energy phosphate pool and was calculated as:
EC = ( A n +O.SADP)l(ATP+ ADP+ AMP).
The DNA in the cell ex- was recovered in 5% perchloric acid and quantitated
using a spectrophotomebnc, diphenylamine &or &on with calf thymus DNA as the
standard.= Hydrogen ion and adenine nucleotide values were then correctal for DNA content
from each plate.
Non-ischemic control &ornyocytes were subjected to similar protocols employing
nommxic PBS @4= 150 mrnHg) for equivalent the perïods as their ischernic counterparts.
Baseline biochemicai measurements were made after removing the culture media and washing
the cells with normoxic PBS. In a separate Sefies of experiments designed to * . a
glucose effect, cells were arposed to 5 or 100 mmoi/L of mannitol with or without insului.
StcuLtical
The SAS Statistical Package (SAS hstitute, Cary, NC) was employed for analysis of
a l i data. Raw data are expressed as the mean f standard exmr with eight plates per mup,
dess othenivise specifîed. Biochemical md-points are expressed as a percentage of the value
obtained h m non-ischemic anitrol plates exposed to an equiva2ent volume of normoxic PBS

54
for the equivaimt period of tirne. Analysis ofvPiimce (ANOVA) was used to s i a i u i t a n d y
compsnmcansbetwemgraips. Wbenstatisbc4U m . y signin*mt difféxences wexe found, they
were speafied by the method of least sigmficant differerirts. Statistïcaï signifïcance was
assumed at pCO.05.
23. RESULIS
A s s c S m e n t @ G ? ~ I *
The e f f e of insulin was asses& using two lewls of ambient glucose concentrations,
a physioIogic lwel of 5 mmol/L and a cardioplegic level of 100 mmoVL. At physiologie
giucose amcentrations, insulin fWed to produce a statistically signficant reduction in cellular
injury (F=0.27, p=0.84 by ANOVA). However, at an ambient glucose concentration of
100 mmoüL Iowa ceiiuiar injury was observexi foUowing exposure to 10 lU/L of humuiin
R (F-4.48, p=0.01 by ANOVA). No firrther protection could be demonstrated by
increesing the insulin concentration to 100 IUlL (Figure 10).
The effect of glucose was asessed with and without 10 IU/L of insulin. Higher ceii
injury was observed as the glucose concentration increased @y two-way ANOVA, glurse
@a F=6.48, p < 0.0001). The addition of 10 NL of insulin reduced cellular injury
(insuün @ect Ft24.26, p <0.0001). Thexe were no statisticaUy signifiant interactive
effects between gluwse concentration and insuiin treatment (gIucose*Ursulin Mecf F =2.05,
p =O.O8). Duncan's multiple range test specitied differences between insulin and non-insulin
treated groups at glucose concentrations of 50 and 100 mmoYL. (Figure 11)
l d n Effect on Pyruw~te Dehydrogenase Acti* .
Figure 12 demonsnates pynivate dehydrogenase activity foIIowing stabilhation,
ischemia and mpfkion. Insulin exposure increased PDH d v i t y at both l d s of giucose.

SS
High glucose amceatraticms a h incnased PDH activity compared to 5 rnmoVL glucose with
or without insuiin. PDH activity was inhibited foilowing ischemia in the gmups with 5
mmoi/L g i m , while those œlis expoBed to 100 moUL of giucose with or without insulia
retained PDH activities near control leveis. FoUowing npanision, only those tells acposed
to 100 mmoYL of glucose and insulin retained near nomai PDH activities while a i I other
groups demonstrated intiiited activity.
To ehmate . . a glucose eff-, cells wexe subjeded to 5 or 100 mmoYL of mannitol
with or without 10 IUlL of in&. Figure 13 illustrades the ability of insulin to stimulate
pyrwate dehydrogenase independent of glucose.
Inrulin Wea on Zntennediate Merabolites
Figure 14 displays the inhra,cellular lactate accumulation which was highest in tells
arposed to 100 rnmoi/L of glucose and insulin during stabilintim and m o n . However,
after 90 minutes of ischemia, intracellukir lacgte accumulation in œ U s exposed to 100
mmoIlL of glucose was lower when insului was added. Ischernia and repemiSion increased
intracellukir lactate in all groups.
Figure 15 iliustrates extracellular lactate release measured in the supernaDuit over each
plate. Ischernia resulted in a large increase in lactate release in al l groups. Extracellukir
lactate release femained eleMted foilowing repemision in di groups, suggesting persistent
anaerobic metabolism. hctate release was Iowa in the insulin treated ce&.
SimiiarIy, extracellular pyruvate concentrations were lower in the insulùi treated œlls.
The lactate/pyruvate ratios were higher in the cells exposed to high concentrations of glucose.
The raw biochemicai data wi@ rneasuremmts of iactate, pyruvate, pH and PDH activity
following each intervention are aven in Appendix 1.

56
l i ~ o n A d Q i i n c N U c l e 0 ~
Figure 16 illustrates the fidi in ATP in aJl groups foUowing ischemia. haaiceuular
ATP was best preserved in the group eqosed to 100 mmol/L glucose and insuiin comparsd
to the groups with 5 mmoYL giucose. Repemision resulted in only a s d increase in
innra~efluiar ATP with aiï four groups remainiag w d below non-ischemic amtrol values.
SimiMy, ischemia resuited in a signifiaint fdi in total adenine nuc lddes in aîi
groups (Figure 17); however, they were best presewed alls exposed to 100 rnmol/L ghcose
and insulin. Foliowing m o n , total adaiine nuc1eotides w a e higher in the 100 rnmoYL
glucose groups but remained below 50% of cuntml values. The raw biochemicai data with
measurements of intraceUuiar high energy phosphates and thev metabolites are given in
Appendix 2.
2.4. . CONCLUSIONS
This study examined the role of insulin and pyruvate dehyâmgenase activity in
primary cultures of human cardiomyocytes subjected to simulatprl ischemia and m o n .
W e have dernonshated that pre-ischemic exposure to insulin improves cellular tolerance to
Srnulateci "ischemia" and "repenUsionm in isolated human ventrïcular cardiomyocytes. The
activity of pyruvate dehydrogenase was shown to be inhibited foUowing ischemia and
remained depnssed foliowing thirty minutes of repemision. Insulin was shown to M y
stimulate pynivate dehydrogenase activity before ischemia and partially prevented the
inhibition of PDH activity during repemision. Insulin treatment also significantly reduced
intraceiiuiar lactate accumulation and extraœilular lactate release during ischemia.
Intraœllular high energy phosphate levels were better pfesemed in tells exposed to hi&
giucose and insulin.

57
The resuits of these inves@ticms suggest that insulia treatment can enhance PDH
surgery. Howeva, the mechaniSm by which insUün stimiilstris pyruvate dehydmgemse
actinty requim further investigation. Denniag the mchanism of insulin's effect on
myocardial PDH may aIlow for phmnacdogic enhancement of the sthuiatary response.

Figure 7: Representative photomicrographs of primary cultures of hurnan pediatric (A) and adult (B) ventricular cardiomyocytes. (200~ magnification, reprinted from Li et a1219)

'Ikp #1 h p #2 Humidifier 37'C 4'C 37' C
Figure 8: Schematic diagram of the equipment required to simulate "ischemia" and I
reperfllsion. 100% nitmgen gas (NJ is bubbled through two oxygen traps prior t
to flushing a d e d plexiglass chamber. Four plates of cultured ceUs can be placed in each chamber which is equipped with a central sarnpling dish to ensure the absence of oxygen and to monitor temperature. (Reprinted from Tumiati et alm)

Figure 9: Representative photomicrographs of human venfricular cardiomyocytes following assessrnent for cellular viability using 0.4% trypan blue dye. In control non- ischemic cells (far left panel) no cellular injury is visible. In contrat, the ceils in the far right panel have undergone 90 minutes of ischemia and 30 minutes of repemision resulting in approxirnately 50% cellular injury.

Figure 10: Assessrnent of cellular injury following ischemia and repemision. At an ambient I
glucose concentration of 5 mmoIL, there was no signifiant effect of insulin I treatment. However, exposure to 10 IU/L of insulin at an ambient glucose concentration of 100 mmol/L reduced cellular inj ury with no further protection demonstrateci as the insulin concentration increased to 100 TUIL.

GLUCOSE CONCENTRATION (mmoVL)
Figure 11: Assessrnent of cellular injury foUowing ischemia and reperfusion. Higher celi injury was observed as the glucose concentration increased @y tweway ANOVA, glucose qect F ~ 6 . 4 8 , p < 0.0001). The addition of 10 IUL of insulin reduced cellular injury (insulin effect F = 24.26, p < 0.000 1). Duncan ' s mu1 tip1e range test specified differences between insulin and non-insulin treated groups at glucose concentrations of 50 and 100 rnmoVL.

GLUCOSE CONCENTRATION (mrnoV1L) CONTROL
F i 2 : qmvate dehydrogenase (PDH) artiviry fofiûwing thirty minutes of a p u r e gIucose and insulin (STABILEATION); ninety minutes of ISCHEMIA and thirty minutes of REPERFUSION. Insulin exposure increased PDH activity at both levels of glucose @or to ischemia and prevented PDH inactivation during reperfusion. (Resuits compared to non-ischemic control values obtained at baseline or from ceiis exposed to equivalent volumes of normoxic PBS for equivalent time periods-)

'p<O.OS vs W NO INSULIN O INSULIN h.0 ~ N ~ U L W
MANNITOL CONCENTRATION (mmoVL)
Figure 13: Pynivate deh ydrogenase (PDH) ac tivity following thirty minutes of exposure to mannitol (5 or 100 mmol/L) and insulin (O or 10 IUIL). Insulin resulted in sirnilar PDH stimulation at both concentrations of mannitol, indicating that the stimulatory effect of insulin is independent of glucose.

w L
s + fi nos-
Y + I a * e -; 1;1 3 a J aol - W u ,: 2 REPER FUSION + m ; z i1
Q05 - * i
GLUCOSE CONCETJTRAnON (xnmoVL) CONTROL
Figure 14: IntraceUuiar lactate accumulation following thirty minutes of stabifi7iition, ninety minutes of ischemia and thirty minutes of reperfusion. Lactate extraction during stabihation and reperfusion was highest in celis exposed to 100 m M and insulin. However, insulin reduced lactate accumulation following ischemia in =Us exposed to a glucose concentration of 100 mM. Intracellular lactate increased in a l i groups following ninety minutes of ischemia. (Results compared to non-ischernic contrd values obtaùied at baseline or fkom cells exposed to quivalent volumes of
, normoxic PBS for equivalent time wods.)

S IO0 NON-ISCHEMIC GLUCOSE CONCENTRATION (mmoVL) CONTROL
Figure 15: ExtraceUuIar lactate release into the supernatant over each plate. Lactate release increased significantly with ischemia and remained elevated during reperfusion, suggesting persistent anaerobic metabolism. Insulin treatment reduced lactate reiease at both glucose concentrations. (Results wmpared to non-ischernic control values obtained at basehe or from c d s exposed to equivalent volumes of normoxic PBS for equivalent time penods.)

2 "1 F.li 44
REPERFUSION
5 IO0 NON-ISCHEM IC GLUCOSE CONCENTUTION (mmoYL) CONTROL
Figure 16: Intraceiiular adenosine triphosphate (ATP) levels following stabihation, after ninety minutes of ischemia and a f k thirty minutes of reperfusïon. ATP fell significantly in ai l groups, but was bew preserved in cells exposai to LOO a glucose and 10 IU/L insulin. (Results wmpared to non-ischemic anitrol values obtained at baseline or h m cells exposed to equivalent volumes of normoxic PBS for equivalent time periods.)

5 100 NON-ISCHEMIC
GLUCOSE CONCENTRATION (mmoVL) CONTROL
Figure 17: htraceiïular total adenine nudeotide U A . ) levels following stabiiization, aRer ninety minutes of ischemia and afkr thirty minutes of reperfiision. TAN fell significantly in ai l groups, but was better presewed in celis exposed to high glucose and insulin. (Results compared to non-ischemic wntrol values obtained at basehe or from ceils utposed to equivalent volumes of nonoxic PBS for equivalent time periods.)

INSULIN STIMULATES PDH PHOSPHATASE BY A PROTEiN KINASE C-a DElPENDENT

70
3.1. INTRODUCTION
We have demOIIStrafed that msulin nposiue pria to ischernia riesults in human
myocardial pyrwate dehydrogenase stimulation and partially prevents the inhibition of PDH
activity foUowing ischernia and repafuson. The pmsmt Series of investigations were
designed to derennine the mechaaiSm of the stimulatory e&ct of insulin,
Several investigatm have nported tbat irisuün's intraalluiar effects are mediatcd by
protein kinase C.U1alQsl*lm*l-m Howeva, both IClipHl and Blackshea? have shown that
insulùi mediated glucose transport in skieletal muscle das not require protein kinase C
activafion. In addition, Jarett and wIIeagues demonstrated that insulin 'mediatas' h m rat
skeletal muscle have diffefent effects on insulin-sensitive metaboiic pathways in isolated
adipoc~tes.'~ T'us, the mechanisms of insulin signalliag present in adipocytes, hepatocytes
or deletal muscle are like1y very diffefent h m the paîhways which exist in human
myocardium. Most investigators have employed isolated adipocytesin.J61*11"DD or
hepatocytes151a6 to explore the mechanisms of insuiin-mediated glucose transport or PDH
stimulation. U n f m I y , the intracellular effects of insulin on cardiac muscle are poorly
described.
The foliowing studies addressed two hypotheses:
1. Insulin exposure results in protein kinase C (PKC) activation in isolated hurnan
ventricular cardiornyocytes.
2. Insulin mediated stimulation of the pynivafe dehydrogenase cornplex is dependent
upon PKC activation.

To inv- these hypotheses, we eniphyed primary cultures of human vmtricuIat
cardiomyocytes as describeci in aiaptrr two. We assesai the ability of insului to stimuhte
protein kinase C by Ob-g the branslocatiion of PKC h m the cytosol to the perinuclear
and sarcollannal membranes. We d u a t d PKC translocafion by two methods: fiwrescent
in-situ immunohistochemistry b &mamate PK- translocation h m the cytosol to
the membrane; isoform specific quantification of PKC transIOcation using dot blot anatysis
and computerized densitometry. Since PKC translocation does not necessarily nsult in an
hcrease in the kinase activity" of the T e , we deve1oped an in virro assay to measure
the ability of PKC to phosphorylate an erizymeqxcific peptide (a &due of the epidermal
growth factor receptor, RKRT3LRRL).
To detamuie the effeds of PKC modulntion on myocardial pynivate dehydrogenase
activïty, we employed the phorbol ester PMA (4i3-phorbol 12Bmyristate 13a-acetate) as a
PKC agonist. In addition, we employed PKC antagonists which act by either inhibition of
the ATP binding site (CHEL: chelaythrine) or by interference of the lipid cofkctor bïnding
site of PKC ( C U : caiphostin C).
Dichioracetate was employed to inhibit the PDH kinase subunit and sodium fluonde
used to inhibit the PDH phosphatase subunit to M e r spenfy the mechanism of insulin's
stimulatory effect.
W e believe that determination of the intracellular mechanisms of insulin's effixt on
myocardiai ppvafe dehydrogemase may allow for augmentation of the stimulatory response
and lead to impmved cellular tolemce to ischemia,

Primary cultures of human ventricuiar cardiomyocytes were estabïished as previousiy
describeci in chapter two. Confluent culaires of cardiomyocytes (approximately 600,000 alls
pex dture di&) cuitured for 5 to 10 days from the last passage were employed for these
sbdies. InnJin qeéa on mm KIlrrrse C
Immunofluarescent staining ushg a monoclonal anisaisus antibody to protein kinase
C (PKC) was employed following exposure to insulin and/or high glucose in order to obsave
the patteni of intradlular PKC distniution. Foiiowing the intervention of interest, the
cardiomyocytes were fixeci in 100% methanol for 20 minutes then washed three times in
phosphate buffered saline. The œlis wexe thai s&ed with a 1:40 dilution of rabbit a&
human anti-protein kinase C IgG antiWy (Calbiochem Laboratones, La Jolla, CA) at
37°C for 45 minutes. The antibody was then removed with three 15 minute washings of PBS
in a gentle shaker at room temperaftire. The celis were then exposed to a 1:20 dilution of
fluoresah isothiocyanate-labelled goat anti-abbit IgG antibody (ICN Biochemicais,
MisSssauga, ONT) at 3743 for 45 minutes. Each plate was then Wasfied until background
staining in a control plate with no primary antibody was barely detectable. The plates were
then viewed and photographed under ultraviolet tght using an epinuorescent microscope with
a blue filter (Kikon Canada, Mississauga, ONT) at 200x magnincaîion. Positive coatmls
were obtained a h cells were exposecl to 10 nmollL of PMA (48-phorbol 12-myristate 13-
acetate), a PKC stimuiating phorbol ester, for Uiirty minutes. This concentration of PMA
was show to stimulate PDH activity in isoiated hepatoma tells.'" PMA was first dissolved
in dimethyIsulfoXde (DMSO) prior to dilution in PM. The final concentration of DMSO
was mainmed below 0.05% which does not produœ injury in human cardi~myocytes.~

73
In orda to detect isofonn spccinc transIocation of PKC. we pedOfmed dot blot
analyses on cytosoiic and membrane ~ 0 0 s . FoUowing the intervention of intaest, ceiis
were wahed, scraped and resuspended in 50 p L of 50 mmoYL TRIS-buffered saline. Cells
w e n thai sonicatcd and centrihiged at 14 000 rpm for 5 minutes. Foiiowing rem& of the
soluble cytosoiic supenÿitant Won, the plla was resuspeaded in 50 p L of TRIS-bufked
saline to yie1d the membrane enricheci fracton. Both fractons were then divideci inta equal
aliquots of 25 p L each. Che aliquot was employed for determination of protein concentration
and then the equivaient of 20 pg of protein for each sample was placed in the dot blot
apparatus. Foliowing protein transfer to nitroceUuiose, each blot was reacted with an isoform
specific a n t i i y for PKC-E. The same blot was then pmbed with an antiibody specifïc to
PKCu (AU antibodies courtesy of PIERCE Biotechnology; RocHord, IL.). Western blot
analysis using cherniiuminesœnt M o n demonstmted that each isoform-qecific antibody
was specinc for PKC with no evidence of non-specific background staining. Slot blots were
then iclwined using a commercially available soAware program (Molecular Images;
Mississauga, ONT) and each band assessed daisitometridy. For these studies, positive
controls were obtained foiiowing exposure to 10 nmoYL of PMA (as above) and aAer thirty
minute exposure to 50 and 100 pmoyL of adenosine. Adenosine has k e n show by
Ikonomidis et al to stimulate PKC translocation at these ~onœntrations.~ In addition, cells
w a e exposed to 50 pmoYL of ademosine in the -ce of the adenosine receptor antagonist
8-@suiphopfienyl)theophyiiine (SPT. courtesy of Research Biochemicals Intemalional.
Natick, MA). SPT is a non-selective adenosine receptor antagonist and has previously been
shown to inhibit the e f fm of adenosine at a concentration of 100 pmol/L." Both adenosine
and Sm weE &y dissolved in PBS to achieve the final desired concentration.

74
Pmain kinase C activity was measmeci using a modification of a pmiousfy report&
assay." Connuent CUItures of cardiomyocytes were expwed to the breatmmt of intexest for
thirîy minutes. nie tells were thca washed, saaped and resuspended in 50 pL of 50
mmoUt TRIS-buffered saline (150 mmoYL NaQ in 50% giycaol, pH 7.2). FoUowing
scmicaîion, 10 pi, of each ceiI extract was addeti to 15 pL of mon buffet for 60 minutes.
The r d 0 1 1 b s e r amsisted of quai concentratio~ls of a lissamine rhodamine Blabelled
peptide containhg a PKC-specifïc phosphorylatim site (epidennal growth factor receptor,
RKRTLRRL), an activating solution (Phosphatidyl-L-serine, lm*) and a b t d k
containing 10 mmollL ATP, 50 mmoi/L M g Q , 0.5 mmoVL CaCl2, 0.01 % Triton X-100
and 100 mmol/L Tris-(hydroxymethy1)-amino methane, pH 7.4. (AU reagmts h m PIERCE
Biotechn010gy; Rockford, IL.)
Foliowing incubation, the &on mixture was fiactionated through a DEAE-
sepharose column equilibrated with 20 rnmoYL HEPES @H 7.9 @4"c), 20% giycerol and
lmmollL EDTA. Phosphofylated peptide bound to the positively chargeci m1umn and was
then'eluted with a 2 m o n NaCl, H E P E S - b f l i saline solution. The absorbante of the
eluted fluid was then meanired using a spectmphotorneter (Beckmann Ltd; Fullerton, CA)
at 570 nm. Reaction baer that had not been exposed to any ceii extracts was also run
t h u g h the column and the eluate sewed as a negative control. CeU extracts exposed to 10
nmoUL PMA were employed as positive controls. Non-specific absorbante was adjusted
using the negative control and the finai absorbante correcfed for protein content and
expressed in relative units of absorbance/mg protein.

75
P p w m Dchydmgenase Aahiity Followùrg PKC Morhhtion
The activity of pyruvaYe dehydrogenase (PDH) was measimd a b each intervention
of interest using the modincafions desaibtd by Robinson et a P of the method of Shai et
aim as desc r i i in chapter m.
To as~ess the mle of PKC stimulation on PDH activity, ceils w a e exposed to 10
mi/L of PMA for tbirty minutes. Bendli and colieagues bave prwioudy shown in isolateci
hepatoma ceils that PMA results in PDH activafion within t a minutes of exp~sure.~'
Cardiomyocytes were then exposed to 100 mmoUL of glucose and 10 IU/L of insulin in the
presence of the PKC inhicbitors chelayllirine (CHEL: 1 firnoVL; Calbiochem Biochemicals,
La Joiia, CA) and calphosth C (CALC: 2 0 nmoyL; Calbiochem Biochernicals, La Jolla,
CA). As descnbed in chapter one, CHEL inhibits the ATP binding site of the m e .
CaEL is isolated h m the leaves and stems of Macleaya cordatu and ~~ and has
been shown to inhibit PKC at a concentration of 500 nrnoVL.IPr CALC is ~ecovered h m
Cladospon'm ~Iodoporioides and inhibits PKC by intrrfkzhg with the lipid conldor binding
site.'" CALC was found to inbiit PMA induced cardiomyocyte preconditioning at a dose
of 200 nmoVL." We therefore employed a CEEL dose which was approximately two thes
the published effective inhibitory dose (Calbiochem) and a CALC dose which was previously
shown in our lab to inhibit the functional properties of PKC.*
An additional expairnent involved cardiornyocyte exposure to 100 mmoYL of glucose
and 10 Iü/L insulin in the presence of the PDH phosphatase inhibitoi sodium fluoride (NaR
25 rnmol/L, Sigma Chemicals). Insulin has beai s h o w to stimulate the phosphaîase subunit
of the PDH complex in isolated bwine heart mitoch~ndria.'~ Ceil extracts were also
expoBed to the PDH kinase inhibitor dichicmacetate (DCA: 5 mmoyL) in the presence of

100 mmoi/L gîucuse and 10 NL insului.
. *
The SAS Statistical Package (SAS Institue, Cary, NC) was employed for analysis of
a l i daot Raw &ta are expressed as the mean f standard aror with eight plates per group,
uniess othemk specified. Anaiysis of variance (ANOVA) was used to simultaneously
compare means betweea groups. When statisticaily signifiant ciifferences were found, they
were specifîed by DUNCAN'S muttiple range test. Statisticat signifïcance was assumed at
p<o.05.
3.3. RESULTS
Ine Effetz of Z d n on ProteUr Kinase C
Fluorescent in-süu immunohistochemistry demonshated haî thirty minute exposure
to insulin resutted in a translocation of PKC h m the cytoplasm to the membrane
(Figure 18). A more modest trandocafion was observed in c a s exposed to 100 mmoi/L
glucose &ne. Figure 19 displays a simiiar immunohistochernicai study in cells exposed to
the phorbol ester PMA (1 pnolL) and adenosine (50 pmol/L), both of which have been
shown to stimulate PKC translOcafion.m In panel D, PKC translocation is obsemd in
cardiom yocytes which have undergone ischernic preconditioning .
Figure 20 displays a representative dot blot analysis which shows an isofm specinc
translocation of PKC in c a s exposed to insulin. Densitometric analyses mealed no e f fd
on PKC-é distributions with either insulin, the phorbol ester PMA (10 nmolL), 50 or 100
p m o n of adenosine or with the non-specifïc adenosine receptor antagonist Sm. However,
there was a marked cytosolic to membrane translocation of P K C a in ceUs exposed to insulin,
PMA and adenosine. This translocation was inhibiteci with SPT. Digitalized densitometry

77
revealed the foïlowhg membrane:cytosolic ratio5 for each group: 5 mM Glucose 1.43; 5 mM
Gluase and insiiün 7.75; 100 mM giucose 5.44; 100 mM giucose and insuiin 3.%; PMA
4.93; 50 pM adenosine 6.4; 50 adenosine and 1 0 f l SPT 2.27.
Figure 21 illustrates the effwt of insulin, glucose and PMA (10 nmoYL) cm total PKC
activity. A signincant interactive &kt was fouad betweea glucose and insulin with respect
to total PKC actmty (glucaree*innJ @kt, F=5.14, p=0.035). Insulin stimiilatprf PKC
activity with a higher effax seen at an arnbient glucose conceritration of 100 mmoYL. PMA
was a potent stimulaior of PKC activity wiîh an efféct almost double h a î of high glucose and
insulùi (O.58f 0.05 vs 0.29I0.04, p<O.OS by DUNCAN'S multiple range test).
P y m e Dehydmgenase Activity F o l M g Pn,eiin Kitcase C Modulan'on
Figure 22 displays the activity of rnitochonckhî ppvate dehydrogenase (PDH)
following exposure to 10 nmoi/L PMA, 200 nmoYL CALC, 1 pmoK CHEL and 25
mmoYL NaF. Cells exposed to CALC, CHEL and NaF were also exposed to 100 rnmoVL
glucose and 10 IUL insulin. PMA stirndated PDH activity to a level si& to that of 100
mmoi/L @ucose and insuiin while C U , CHEL and NaF prevented PDH stimulation.
3.4. CONCLUSIONS
In these Sefies of experiments we have deiemllned that insulin exposure results in
translocation of PKC-o h m the cytosd to the membrane. Translocation of PKC-a resuited
in an inaease in the kinase activity of the enzyme. Pharmacologie stimiilsticm of PKC with
the phorbol ester PMA resulted in a stimulation of myocardial PDH activity to a levd simüar
to that achiwed with 100 mmol/L glucose and insulin. Cells exposed to 100 mrnoVL glucose
and insulin in the presence of the PKC inhibitors C U or CEEL did not exhibit an
increase in PDH activity. Similarly, ce& exposeû to high giucose and insulin in the presence

78
of the PDH phosphatase inhibitor sodium flwride did not display a stimulation of PDH
activity.
niese studies demonsûate that in isoiated human ventricular cardiomyocytes, insuün-
mediated stllnitlation of pynivate dehydrogeriase activity is &pendent upun protein kinase C-
a stimufiition of the phosphatase subunit of the PDH complex.

Figure 18: In-siru fluorescent imrnunohistochernistry demonstrating the distribution of protein kinase C using a 1:40 dilution of rabbit anti-human anti-protein kinase C- IgG antibody. The left panel demonstrates generalizad cytoplasmic st&g in ceils exposeû to 5 mmoUL glucose alone. Ceils exposed to 10 IU/L of insulin with 100 mmoVL glucose (right panel) display a redistribution of PKC staining to the sarw1ernna.i membranes.

Figure 19: In-situ fluorescent immunohistochernistry demonstrating the distribution of protein kinase C using a 1:40 dilution of rabbit anti-human anti-protein kinase Cc- IgG antibody. Panel A dernonstrates generalized cytoplasmic staining in cells exposed to 5 m M glucose alone. Cells exposed to the phorbol ester PMA (1 &; panel B) or adenosine (50 pM, panel C) display a redistribution of PKC to the perinuclear and sarcolemnal membrane. Ischemic preconditioning (panel D) results in a similar redistribution of PKC staining. (Reprinted fkom Ikonomidis et aizod)

10 nM PMA
50 uM Adenosine
100 uM Adenosine
50 uM Adenosine + SPT
-71IpI-'
PKC - E PKC -
Figure 20: Slot blot analysis demonstrating insulin's effect on protein kinase C (PKC) transiocation. Each lane is blotted with 20 pg of protein. The left panel shows no effect of insulin, PMA or adenosine on PKC-e distribution. The right panel demonstrates that insulin, PMA and adenosine cause a translocation of PKC-at to the membrane fraction. The adenosine mediated PKC-û! translocation was inhibited by the adenosine receptor antagonist SPT.

"p4.05 different from 5 1
Figure 21: Measurement of total protein kinase C (PKC) activity using an NI vitro phosphorylation assay. The phosphorylation of a PKC specific peptide (epidennd growth factor receptor, RKRTLRRL) is measured colorimetrically as spectrophotomemc absorbame at 570 nm and corrected for protein content. The phorbol ester PMA resulted in a significant stimulation of PKC activity compared to 5 a glucose alone. A significant interactive effect between insulin and glucose and total PKC activity was found (glucose*imulin, F-5.14, p=0.035 by rm-ANOVA). Insulin stimulated PKC activity with a greater effect observed at an arnbient glucose concentration of 100 mM.
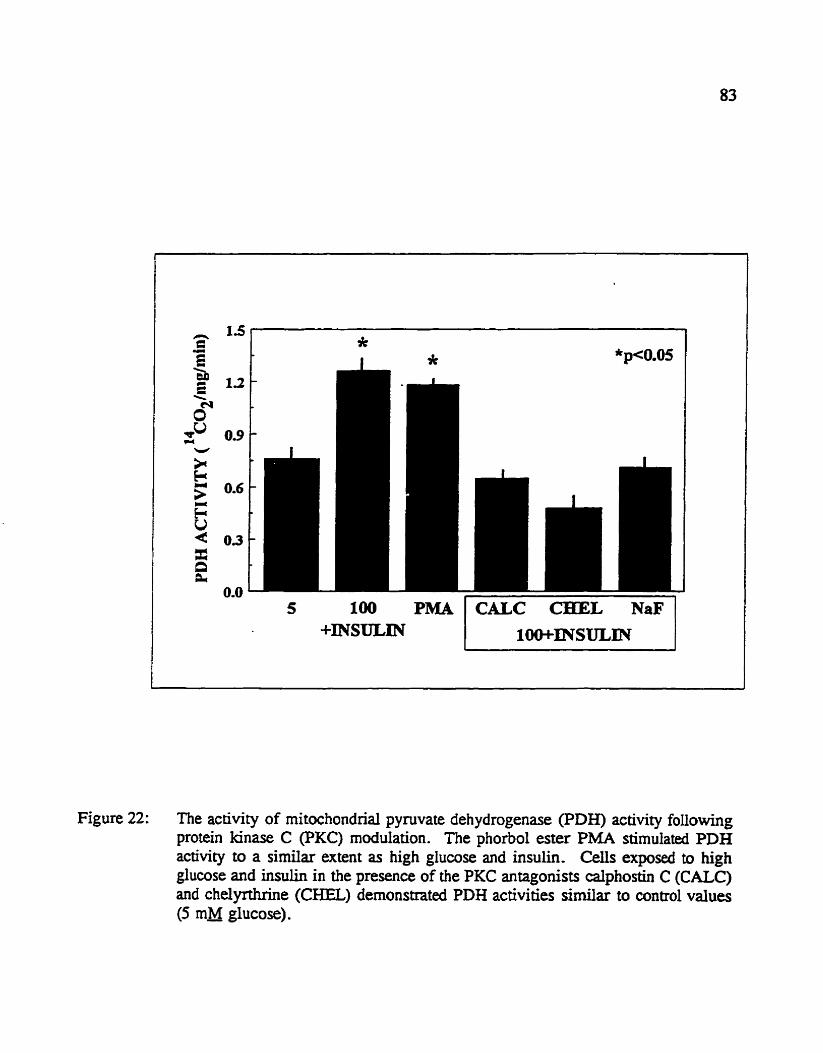
100 PMA -INSULIN
CALC L NaF 1Oo+rNsULm
Figure 22: The activity of rnitochondnal pyruvate dehydrogenase (PDH) activity following protein kinase C (PKC) modulation. The phorbol ester PMA stimulated PDH activity to a similar extent as high glucose and insulin. Cells exposed to high glucose and insulin in the presence of the PKC antagonists calphostin C (CALC) and chelyrthnne (CHEL) demonstrateci PDH activities similar to control valws (5 rnM glucose).

CBAPTIERFOUR
ADDITIONAL INVESTIGATIONS

85
4.1. INTRODUCTION
In chapm two and k, insulin was found to stimulate ppvate dehydrogenase in
isolatecl human veatticuiar &omyOCyfeS. Although vmtricuiaf dysfiuiction is clearly due
to cardiomyocyte injury, there is inæasing &darce that endothelial cell dysfuoction
contributes signifïcantly to ischernic npxfkion injury.-* EndothW ceils produce a
potent Msoactive compound now idmtified as nitric mide." Although nitnc oxide itsdf is
a negative inotrop,- preservatian of endothelisl fimction has been show to imprwe
ventricular function f0Uomng ~ o p 1 e g i c alTest."
The effect of metabolic intervention on endothelisl celi tolerance to ischemia is
unknown, The effect of altering the aembic to anaerobic ATP ratio in nonumtractile œk,
such as endothelium, is also unknown. Ischemic injury involves a c o m p l i W interaction
between coronafy endothelial œ i l s and cardiomyocytes. Howwer, in order to distinguish
differeritial responses between these two ceii types, an isoiated cell culture mode1 may be
preferred.
For example, Shirai and coiieagues demonstrakd that preconditioning was ineffective
in protectuig endothelial cells h m ischemic injury but was extrernely protective in
cardiomyocytes."' To detennine the effects of insulin exposue on endothehi ceii
metabolism we employed isolated cultures of human saphenous vein endothelial cells. These
cells have previously been show to respund in a si* manna to coronary and aortic
endothelium.-
W e origindy hypothesized that stimiilation of myocardial PDH by insulin would
improve the fecoveq of aerobic metabolism and lead to be$ter huictional recovery fohvhg
cafdioplegic arrest, Since the isolaîed cardiomyocytes employed in these sîudie~ w m

86
quiescent, we were unabIe to perfarm hmdicmai analyses in chapters two and three. We
thdore developed a porcine model of orthoOopic cardiac ttansplantation to evaluat~, the use
of insuiin-enhanced doplegia on the recovery of myoardial fùnction foilowing four houn
of hypothermie cardiop1egic arrest.
4.2. MmHODS
lih&hew Gdl snuty
Human saphmous vein endothelid tells were cuitureû as previously describeci.-
Briefly, segments of vein were tirst fiushed wiüi PBS and then an enzymatic solution
containhg 0.2% trypmn and 0.1 96 coiiapenase. The effluent was then collected and
incubated in medium-199 (GIBCO laboratores; Grand Island, NY) containing 20% f d
bovine senun, 100 UlmL penicillin and 100 IrglmL streptomycin. Endothelid œiis were
distinpuished M m fibroôlasts and smooth muscle celis by morphologid criteria. CeiI cuiture
pwity was greater than 95% and was confirmed after the third passage by
immunofiuorescenct staining with a monocIonal antibody against faicbr eight antigen. Cells
passaged 4 to 7 times and aged less Uian 75 &YS wexe used for this study.
Endothehl c d s were subjected to a similar protocol of ischemia and repemision as
descnibed in chapter two. Foliowing thirty minutes of reperfusion, ceUs were incubated for
24 hours in semm free medium. Non-viable cells lost th& adherence to the celi culture plate
and fimted M y into the supernatant. In mtrast, viable cells were firmly adherent and
required trypsinization in order to harvest. Each fraction (supernatant and trypsinized) was
then counted in a coulter counter which measured for particles above 10 microns in size.
chapter two.

We have pmiously d e s c n i a p r c k mode1 of orthotopic transplantation to
evaiuate the effects of l m fiow pemiSicm during hypothermie ( 4 0 storage.= We employed
a simplifed perfusion apparatus using donor blood harvesîed fiom the chest at the time of
orgao procurement to amtinuously p v i d e low flow (15 mllmin) pemiSion during three
hours of &mage. To evaluate the hemodynamic ened of insolin, 10 IU/L of hum& R was
addeci to the initial cardioplegic solution, to the donor b l d pafiisate and to the blood
cardiuplegic solution.
The nperimentai protoc01 was approved by our institutional aaimal care cornmittee
and was in accordance to the Gÿide for the Eore Md use of labormory Mimalr (NM
publiCCLtiOn no 862.3, r&ed 1985). Thuty two fernale Yorkshire pigs (50-6û kg) were used
to perform sixteen orthotopic cardiac transplants employing either continuous donor blood
pemi9on (BIXH)D, n=8) or WC storage in a crystalloid preservation solution
(CONTROL, n=8). In addition, 7 transplants were perf'ed employing wntinuous
pemison with donor blood supplemented with 10 IUL of humulin R (INSUUN). The size
mismatch between donor and recipient was les than 5% of body weight in all experiments.
Donor Operation
The animals were anesthetized with intrmuscular ketamine (30 mgkg) and
isoflurane, inhibaM and venfilated with 100% oxygen to maintain normocarbia, Following
stexnotomy, the heart and great vessels wexe exposexi. Umbilical tapes were placed around
the supCnor and inferior vena cava to p d t adjusbnent of cardiac preload by cavai swing.
Systemic anticoagulation was whieved with the intravenous injection of 10 000 units of
heparin.

A Millar niic~omanorneter was insated via the apa of the left vmtricle to permit on-
line measuremats of heart rate, lefi ventricular Sysblic aad diastolic pressures as well as the
maximum positive dezivative of the systolic pressme curm
Badine measuements of ali hemodyaamic pmme&m were made at a left ventriculaf
enddiastolic pressure (LVEDP) of 10 mmtIg and after bicavai saaring to achieve an LVEDP
of 2 mmFIg. A third measurement was made release of the b i c a a mares. Donor
hearts that failed to recover a left vmtricuiar developed pressure of 80 mmHg foliowing
release of the snares were discarded,
A pursestring suture was p l d in the asceadhg aorta to permit placement of a
cardioplegic cannula. Arterial and cozonafy sinus blood samples were obtained just prior to
aoaic crossclarnping and then one litre of a hypothennic (4Q, hyperkalemic crystalloid
solution (composition in m 1 L : Na+ 127, K+ 20, M e 6, Ci- 7, SO: 6, Pis-hydroxymethyl
aminomethane 0 4, Dextrose 135) was infused into the aortic mot to achieve
cardioplegic ;urest, Following cardioplegic infusion, the donor heart was extracted, placed
in a bag containhg 300mL of hypothermie cardioplegia and stored on ice. The d o p I e g i c
carmula and aortic crossclamp was left in place to permit perfùsion during storage.
In the BLOOD and INSULIN groups, donor blood was harvested from the chest
a f k organ extraction. Following Ntration for particdate matter and the addition of 10 000
units of heparin, the blocui was sfored in sîandard transfusion bags (lhzvenol, Baxta
Healtficare Corp, D d e i d , Ill.). in the INSULIN group, both the crystalloid cardiople@
and the donor blood p h s a t e were supplemented with 10 IU/L of h u m a R. Blood
perfusion was initiatexi within ten minutes of cardioplegic amst and was delivered at m m
temperature (2m) at a vertical height of 100 cm (to correspond to a perfusion pressure of

$9
60 mmsg) employing a standard intravmous bransfusicm apparatus ( F d ,
Heaithcare Corp). Blood pafusion was mhtained thn,ughout the storage period while the
donor kart was on iœ. A myocardial probe was insertcd into the apex of the left veatricle
in aii hearts to monitor temperaairit duriag m e . Coraiary flow rate was de&mhed as
the total volume of perfbak deliverd corrcted for storage tirne in minutes.
Recipient 0ptrmanon
Pnoperative sedation and anaesthesia was similar to the donor protocol. In addition,
a marginal ear vein was used for intravenous access and kept opai with a 50 mLlhour 5%
dexttose infusion. Continuous electrocardiographic monitoring was employed and a camtid
artery line was inserted to measure arterial pressure.
Foilowing stemotomy, the kart and gnat vesseis were exposeci. Umbrlid tapes
were placed around the superior and infaior vena cava. Systemic anticoagulation was
achieved by injacting heparin into the pump prime (10 000 U) in addition to an intrawnous
dose of 10 000 U. Ascending aortic and bicaval cannulation wae used to place the recipient
on cardiopulmonary bypass. Flow rates were adjusted to maintah systemic perfusion
pressures above 50 mmHg. No vasoactive medications were a d m i n i s ~ during
cardiopulmonary bypass. Systemic -sion was maintaineci at 37C
After the aorta was crossclamped, the rtxipient hem was extracted maintaining a cuff
of nght and lefi atrium. nie left hemiazygous vein was figatexl at its insertion into the
coronary sinus. The anastomotic rnargins were then inspected and trirnmed in pfeparation for
orthotopic transpiantation.
Donor blood pemigon of the allog& was stopped and an initial 350mL blood
cardioplegic dose infùsed in all groups at a flow rate of 100 W m i n . Cardiaplegic

90
probctioa consisted of a 2: 1 mixture of b1wd:aystalloid and was Uvered at 1VC foïïowing
the completion of each anasfornosis. Foliowing completion of the puimonary artaial
anastomosis, 350 rnL of blood cardioplegia was delive& at 37°C. Artaial and coronary
sinus blocxi samples were obtained at erh cardiop1egic infusion, at the time of cn>ssckmp
r e m d and e v q fiffeen minutes during r e p h i o n . If ventricular BrilIation ocaured
during repafusion, thne attempts wae made to defibrillate the heart. if unsuccesshil, 100
mg lidocaine was delivered intraveriously and defibnuation attempted once again.
At the compktion of forty-five minutes of m o n , hemodynamic mea~urements
were obtained. Preioad was adjusted by transient venous occlusion. FoIIowing completion
of aU measufements, the recipient was weaned off bypass if possible. If indicated, Ig
calcium chloride was given and if additionai inotropic support was required an isoproterenol
drip was esGrblished (4 mg/L at 100 mUmin). Thirty minutes foilowing discontinuafion of
cardiopulrn~~liiry bypass, decannulaîion was pexfbrmed and the anima sacrificeci by
intravenous potassium chloride injection. If the animal was not successfiil in maintainhg a
mean art& pressure above 60 mmHg for thirty minutes despite inotropic support, weaning
was deemed to be unsuccessful. Ventrïculat pacing was employed if necessary to mainiain
the heart rate over 80 beats per minute.
Biochemical Memr-
A r t . and coronary sinus blood sampIes were m y e d for the pamal pressure of
oxygen @Q3, carbon dioxide @CO& pH, hemoglobin concentration (Hb) and oxygen
saturation (SaW. ûxygen content (Q,Con) was calculated from the formula. @Con = 1.39
Hb*Sa02 + 0.003* p02. Blood samples for iactate determination were mixeci with a
measured volume of 6% perchloric acid. Lactate concentration mis measured in the protein-

91
fkee qematant with a mmmexcially available assay (R@d Lacrate Stit Pack, Caîbiochem-
Behring, Ia Jolla, W.)
Starisn'COIA~&
Statistid anaïysis was p a f d using the SAS statisticat software program (SAS
Institute; Cary, NC). Categorical data were anaiyzed by chi-squared or twetailed Fier's
exact test whexe appropMte. Continwws data are exprrssed as the meankstandard M m
and were analyzed by tweway Analysis of Variance (ANOVA) evaluating the main effects
of gmzp and rime as weU as the interactive (gmup*tac). Duncan's multiple range
Endothelid cells were highly susceptible to ischernic injury compared to human
ventri& cardiomyocytes. Following ninety minutes of ischemia and overnight incubation
in sexum-free medium, oniy 15 % of the endothehl ceus at risk remained compared io over
40% of the cardiomyocytes (Figure 23). Insulin treatment impmved cardiomyocyte survivai
but did not affect endothelial ceil tolerance to ischemia. However, insulin did stimulate
endothelial ceîi PDH activity and reduced extrace1lula.r Iactate release (Figure 24).
UnfortuMtey, this effect was ody obsaved prior to ischemia with no evidence of insuiin-
medïated stimuiation occiirring aRer ischemia and repemision. In addition, endothelid 4 s
did not increase extracellular lactate release in rrsponse to ischemia. In amtrast,
cardiomyocytes reacted to ischemia 6 t h an approximate two-fold incfea~e in iactate release
(Figure 15).

92
Po& shu€y
Figure 25 cietnonstrates the mefabOIÏc &cts of umthuous low fiow âcmor bLood
m o n dwing tkee hours of hypothermic storage. Dowr blood pafusim pamitted
limited myocardial metaboliSm and despite predomhmtïy anaerobic metaboïism led to a
signifiant improvemmt in functional recovery (Figure 26). The addition of insulia to the
pemisate and to the cardioplegic soluticms d f e d in simüar h a t e and acid rdease as in
the blood group. Howwer, ininsulin treated hearts displayed improved recovery of aerobic
metaboliSm which led to a m e r recovery of left ventncuiar function (Figure 27).
4.4. DISCUSSION
These additionai shidies provide evidence that inmlin treatment can affect endothem
c d metabolism. Exposure to insulin resufted in higher PDH activity and lower lactate
producfion. However, insulin treaûnent was unable to pmtect saphenous vein endothehi
celis from ischernic injury. It is possible that 90 minutes of ischemia was too severe an insult
for adotiiefial d i s and that insulin treatment rnay be protective against a less severe insult.
In the porcine model of orthotopic transplantation insulin treatment increased
myocardial oxygen extraction d d g organ implantation and resuited in irnpmed recovery
of left ventricuiar funaion. SUrpnsingly, there was no efféct of insului treatment on lactate
or acid release. This may be due to the fdd that hypothermic storage was employed which
may have a t t e n d any me&bolic stimulation. The metabolic différences observed occurred
foilowing the completion of the pulmonary arterial anastamosis at which the a i i hearts
received normothermic blood cardiopIegia. It is possible that maintahg normotherrnic
temperatures durhg storage rnay augment the metabolic eff- of insulin.

*p<0.05 vs CONTROL
*
Figure 23: Twenty-four hour sumival foilowing ninety minutes of ischemia and reperfusion. Endothelia1 œiis (EC) displayed increased sensitivity to ischemia comparrd to cardiomyocytes (CM). Insulin treatment conferred protection to cardiomyocytes, , but not to endothehl A s .
I

PDH ACTIVITY (nmollmg) O P P g i : a h 0 0 k b
O P g m g LACTATE RELEASE (mol/@

BLOOD CONTROL -e-
Figure 25: Effm of continuous low flow donor blood perfusion during three hours of hypothermie storage. Measurements were obtained pnor to organ procurement (PRE); after the initial cardioplegic infusion (PLEG); afkr the cornpletion of the left atriai (LA), right atrial (RA) and pulmonary artenal (PA) anastomoses; after , removal of the aortic crosscIamp (XCL) and every 15 minutes during reperfiision. Hearts in the BLOOD perfused group displayed persistent anaerobic metabolism with greater lactate and acid release during cardioplegic amest. There were no diffmces d u ~ g the reperfusion period.

V
2mmHg 10mmHg LVEDP 2 m m H g 10mmHg PRE POST
Figure 26: EEect of conthuous low flow donor blood pemision during three hours of , hypothemic storage. Perfûsion of donor blood (BLOOD) led to an improved recovery of left ventricular developed pressure compared to non-pemised control hearts (CONT) .

OXYGEN EXTRACXION (mUdL) LV DEVELOPED PRESSURE (mmHg)

-FIVE
DISCUSSION

99
5.1. INTRODUCTION
CardiOvaScular disease continues to be the mosf signincant source of morbidity and
morhlity in Canada.' Although many patients with angina pedoris are appropriately
managed with medical therapy, connary artery bypasp sufgety (CABG) remains a proven and
effdve treatment for athemsc1erOfic commry artay disme- Severai large, prospective
randomized trials have demonshated the supaiority of CABG wer medical treatmmt in
patients with lefi main disease,'6 were triple vessel coronary artery disease,' uris&b1e
anginat7 and left ventncular d~sfiuiction.~
Coronary artery bypass grafting was initiatly assoaated with signifiant morbidity and
mor&lity.ls However, advances in surgical techniques and perioperative myocardial
protection have led to impved surgical outcornes despite an increasingiy high ri& patient
pop~lation.'~~~~ Unfortunately , several subgroups of patients continue to be at high risL for
perioperative complications. These include patients with poor preopaative lefi ventricular
fiiricti~n~~ and those who require urgent revaSculafizaticm for unstable angina? Further
improvements in periaperative myocardial protection are r e q d to irnpmve the resuîts of
surgery in these precariously high risk patients.
Postoperative low cardiac output syndrome has been objedvely defineci as the need
for inotropic or intraaortic balloon pump support for longer than thirty minutes in the
intensive are unit UCU) despite o p W volume and electrolyte The development
of low output syndrome results in prolonged lengths of ICU and hospital stay and is
associated with a higher pfevalence of pexioperative myocardial infanction and operative
mortdity. Interventions designed to reduœ the incidence of postaperative low cardiac output
syndrome may have a subsîantid clinid and eccmomic impact on the delivery of essential

in a recent miew of 623 patiaits who wae enroIled in prospedive cliaical triais
assessiag periopaative myocardial rnetabolism, the development of low output syndrome was
found to be associated with persistent iinaerobic metaboliSm during early repahisi~n.~
The prevaience of low output syndrome was 13% in patients whose net myocardial lactate
release after five minutes of repafuscm was greater than 0.4 mm01 per Litre of blood
compared to 5 % in patients who released les than 0.4 mrnoLn of lactate @ < 0.M). In
patients who displayed net myocardial lactate extrocrion the prwalence of low output
syndrome was d y 3%. Therefore, improving the transition h m anaerobic to aaobic
metabolism foiiowing cardioplegic arrest may prevent the development of low output
syndrome.
Several clinid trials have demonsuami a delayed recovery of normal aerobic
myocardial metabolism foUowing cardioplegic a x r e ~ t . ~ ~ ~ * ~ ' - ~ Anaerobic glycolysis
produces only 2 moles of ATP for every mol of glucose utiiized compared to 36 moles of
ATP produced by oxidative phosphorylationen In addition, Weiss and colleagues
demonstrated that aaobic ATP prpduction is preferentially utilized for contractile functio~,~
whereas ATP produceci by aaaerobic giycolysis was utitized to maintain cellular integxity and
homeostasis. Thus, fàcilitating the reawery of normal oxidative phosphorylation following
cardioplegic arrest should resuit in impmed contrade function foiiowing surgery.
The pyruvate dehydrogetlslse (PDH) cornplex regulates the conversion of ppvate to
acetyl-CoA which in turn is utüued by the Krebs' cycle. When PDH is inhibited, pynivate
is converted to lactate by the enzyme lactate dehydrogenase WH). The latter reaction is
reversible and lactate can be exûacted by the heart and converted to pyruvate. Kobayashi

101
and Nedy demonstrated thaî the actîvity of rat myocardial PDH was significantly inhibifed
foUOwiag brief ischemia a d repafusi~n.~ Inhibition of myocardial PDH leads to persismt
of iasulin exposure on the activity of myocatdial pyruvate dehydrogenase was previously
unknown. Furthermore, the effects of pre-ischemic insulin treatmmt on the posr-ischemic
activity of pynivate dehydrogenase have not previousïy been defined.
W e hypothegzed that pischemic insulin treatment wouid prevmt the pst-ischemic
inhibition of PDH and faciltate the recovery of n o d myacardial m b i c metabolism. W e
empIoyed primary cuitures of isolated human venbricuiar cardiomyocytes to aramine the
metabolic response to Smulated cardiopIegic arrest (ischemia) and repemision. In addition,
we investigaied the role of protein khw C as a mediator of insulin's intracelluiar effects on
the myocardial pyruvate dehydrogemase cornplex.
Our investigations dernonstratexi that insulin was capable of stimulating human
myocardial PDH @or to ischemia and thatpre-ischemic insulin treatment partially prevented
the pst-ischemic inhibition of PDH. Furthermore, we found that the mechanism of the
beneficial effect of insuiin involved a PKC-a dependent stimulation of the PDH phosphatase
subunit. The results of these investigations suggest that pre-ischemic insulin treatment may
fiditate the recovery of normal aerobic metabolism foiiowing cardioplegic anteSf. We
beliewe that an earlier recovery of aerobic metaboihm may lead to impved myocardiai
function. Howwer, the in vifm results ob&ined h m these investigation require M e r

cOilfirmafion in a whole mgan or in viw model.
5.2. CZUCULTURE MODEL
Riaon Vaibicularcmdq.aytes
The cardiomyocytes employed in these studies have been exfensively evaiiiatpA in
prwious reports. 1*3739a The tells used in these studies w a e passaged 2-7 tirnes and were
cuitured for up to 60 days h m the tirne of piimary dture. These domyocytes recain
many characteristics of freshly isolaîecl cells, but have distinct differedlœs. These celis
becorne quiescent following enymatic digestion and passaging. Other investigators who have
successfully cultumi aduit cardiomyocytes have reported that the ceb lose theh ability to
con-- Despite an abundant suppIy of mitocfionciria and contractile proteins, the
sarcomeres becume disrupted d u ~ g division and do not reestablish their characteristic
functional format.
The cardiomyocytes in culhue are easily differentïated from other cell types.
EndothW œlis are oval shaped (15x20 cm) and fibroblasts are spincile-shaped (4x80 cm),
cornparrd to the redangular and much iarger cardiomyocytes (40x80 pm). in addition,
endothelid cells grow pooriy in the medium used for cardiomyocytes whereas fibroblasts
have a much fdster doubling time in culture and are easily ideritified as a spindle-shaped
contaminant.
The quiescait nature of the cardiomyocytes is W y a resuït of trypsin digestion whkh
causes a breakdom of myofibrillar mganbation. These œk may simulate the
cardioplegidy arrested heart encountaed duMg cardiac surgery. The œilular
concentrations of troponin 1, troponin T and the MB isoform of creatuie kinase are similar
to that seen in viw."' The ability to pmcondition these ceUs against prolonged ischemia is

membrane is similar to that fouad in the r n y o c a r d i ~ m . ~ ~ ~ ~ The activities of supaolnde
dismutase, catahe and glutathicme pemxidase are also simüar to those found in cardiac
biopsies.= In addition, giutathione peroxidase gene expression is simiiar to that seen in
whole h e a r t ~ . * ~
Therefm, despite th& quiescent state!, we believe that these œlls are phenotypidy
cardiomyocytes and provide a unique oppmtdty to d u a t e the cellular response to ischemia
and repafusion and the effects of metabolic interventions such as insuün. However, th&
inability to contract during 'nperfusion" may affect the interpretsition of our metabolic
resuits. The NI vivo contracting hem may s e e r h m a greater lactic acïdosis and a greater
depletion of high energy phosphates than seen in this study. However, a recmt clinid
h e w demonstrated that the myocardial be ls of high energy phosphates in the region
supplied by the left anterior descendhg artery fell by only 22% following cardioplegic arrest
and ten minutes of @sion." In contrast, the experiments described in chapter two
produced a greater than 50% fall in ATP and total a c i d e nucleotides.
SUnulared nI~chemia' and "Rep&sionn
Our technique of simuiated 'ischemia" and repemiSon has beai previousiy described
in detail? W e are able to produce a d e o x y g e d PBS sohtion with a m e a s d p4 of O
mmHg. This solution has a stable pH and can be storab in seaied containers for severai
weeks without an appreciable increase in oxygen tension. Exposure of this solution to a
normoc atmosphere results in a p Q of approximately 150 mmHg within 10 minutes. Brief

104
eqosms to normOlLia oess than 15 seconds) are required to adjust pH, osmolality a to
withdraw aliquots and do not resuit in incxieased oxygai tension.
Enposing cardiomyocytes to 90 minutes of "ischieniia" resuits in significant cellular
as assesed by tfypan blue uptake?" The amamt of cellular injury followiag
minutes of ischemia and thirty minutes of repafuscm is remarkably reproduci'ble and
avemges 45% of the cells at ri& Reducing the volume of solution mer the cclls h m 10
m . to 1.5 mL resulted in an accumulation of the products of anaerobic metabolisrn and a
marked reduction in the extràceluiar pH. Therefore, this modd is Smilar to the e f f e of
global ischemia on the heart. Unfornurately, the volume overlying the ceils rernains greater
thsn the solution to which the d i s are exposed during giobal ischemia and may actuaily
represent a form of low-flow ischernia analagous to limitai cardioplegic pemision.
We befieve that our mode1 qmsents low-flow ischemicr verms simple hypoxia due
to the effects of the volume reduction. Whai œils are exposed to 90 minutes of hyparicr
with IO nrL of deoxygenated PBS, the biochemicai abnoRnatities are much less severe and
celiular injury is only 25030% cumpared to approximately 45% with 1.5 mL of anoxic
PBS.' Therefore, the volume reduction over the culture dishes resuits in a greater ischemic
insult than occurs with simple hypoxia and reoxygenafion, likely due to the higher
concentrations of glycolytic metabolites.
In previous studies, ischernic injury was distinguished h m injury due to reperfusion
by assessing trypan blue exclusion immediately following the ischemic phase of the
experimedW- lschemia was found to injur approximate1y 30-3596 of the cek at ri&
with an additional 10% injury occurring following reperfùsion for thirty minutes. In these
Senes of expeximents, we did not assess injury foliowing ischemia alone nor did we assess

las
the effects of insulia exposure dimng isctimiia Heame dernonstrated that metaboiic
stimufstjon duriag ischemia was detrhmtd to the recovery of left ventricuiar functio~t.~
Thus, we chose to adrmrilster . . insulin prier to ischemia. To assess the role of insulin in
protecting agahst nperfusion injury, we nposed œ h to 5 or 100 mmoyL of glucose with
or without insuiin (10 IUL) during reperfirsion. In addition, we evafiratprl the ability of
insuiin to protect caniiomyocytes fiom exogmous in via0 &t injury. Insullli was found
to reduœ repemision injury and provide an antioxidant defmse. However, this protection,
whiie StatiSticaUy si-canî, was minimal compareci to the protection affimiexi by
conventional antioxidants such as Trolox or ascorbic acid." Tagami and colleagues have
demons&aed that insulin partially reversed the inhibition of antioxidant enzyme activities in
aortic endotheliai a î i s from diabetic rabbitS.= Our experiments were limited to the
assessment of glucose and pyruvate metabohm in cardiomyocytes subjected to simulated
ischemia and repemision. Further investigation is required to dua ie the ability of insulin
to stimulate native antioxidant defenses.
5.3. ' INSULIN MEDIATED CARDIOPROTECTION
Redudon of Cellular I i j . y
Pre-ischemic exposure to insului resdted in protection against injury foiiowing ninety
minutes of ischemia and thirty minutes of feperfusion. In cornparison to other interventions
such as ischemic preconditioning'" or adenosine pretteatment,' the magnitude of the
prokaive effect of insulin was small. Both preconditioning and adenosine reduced cellular
injury h m approximately 45% to 25%. whereas insulin only reduœd injury to
app~oYtimately 35%.

106
The magniû.uk of the protective e&ct of ihsulin may have be!en limiteci by our modd
of low-fiow ischernia which was uscd to simuiate cardioplegic amst with iïmited pahuion.
An earlier study by Hearse in an isolated rat heart modei demonstrated that giycoIytic
stimiilatim following high gluccse expo~lire was &rhw&d and led to depressed riecovery
of fiindion foiiowing ischemia? Mannitol treatment aïme, as an osmotic wntrol, a b
fesulted in depnsd recovery compared to amtrol. Howevex, in& m e n t erocerhed
the injury in high glucose tmatexi hearts whiie insuiin crnenuaied the injury in the niannit01
treated hearts, suggesting that the injury sustained f b m high glucose was both metabolic and
osmotic in nature. Hearse's mode1 employed a single cardioplegic dose to achieve arrest and
then hearts wexe s u b j d to hypothermie global ischemùz for seventy minutes without any
cardioplegic pem>sion. In contrast, Doherty and mileagues perfonned experiments in a
simüar isolateci rat heart model with the important addition of inteminent cardïopIegic
infusions evay 15 minutes during two hours of hypothdc In Doherty's -y,
high glucose pemisates ditring doplegic arrest protected against ischernic injury and
resulted in superior recovery of function. W e believe that it is potentially detrimental to
atternpt metaboiic stimulation during low flow or global ischemia. Thus, we chose to
administer insulin prier to ischernia to prevent the irreversiible inactivation of the PDH
cornpiex during early repemision.
Several investigators have evafuated glucose-insulin so1utions in a wide vafiety of
animai rn~dels.~~~-~-~- Unforhwtey, these studies have yie1ded COIlflicting
resdts and the benefits of glucose andfor insulin in the setting of myocardial ischemia
remains controv&. Similarly, several clhicai studies evaluating the use of glucose-insulin
solutions have failed to cansistently demonstrate a protective effect.Y41-nsnx1m*1mf1~

doplegic arrest. In addition, îhese studies involved eittiea global ischemia or intermittent
CatdiopIegic pemisicm during the iscbemic paid. As Neely and Grotyohann d e m o n M ,
accumuiation of metabolic end-products such as lactate and acid can signifïcantly exacabate
ischemic injury."' Contemporary techniques of blood cafdioplegic protection offer continuous
norrnothermic or tepid ( 2 W ) penu9on combinai with the superior buffering qdities of
blood pemision comparexi to the crystdloid sofutions used by most in~estigators.~
Insulin and glucose enhanced cardioplegic solutions require a re-evaiuation with
cantemporasr techniques of blood cardiopkgk protection. In addition, the moleailar and
cellular effects of p i s c h e m i c insulin exposm nequire definition. Clarifcation of the effkct
of insulin on pst-ischemic myocardial metabolkm may af6-d an oppominity to
pharmacologically ahance and specify insului's actions while simultaneoudy preventing any
detrimental effects due to glycoIytic stimulation d u ~ g ischemia.
M ' f i e Effem of l d i n Epsure
Insulin treatment was found to d u c e extracellular lactate release, i n c ~ e a ~ e ktaîe
extraction and impmve the preservation of high energy phosphates following ischemia and
repemision. In addition. insulin was found to stimulate the activity of rnyocardial pyruvate
dehydrogenase and partiaüy prevented the inhibition of PDH activity following ischemia and
repemision. The magnitude of the effect of insulin on PDH activity appears small compared
to the large reduction in extracellular ladate release. However, the energy quivalent of

1041
oxidative phosphorylaticm of one mole of glueose would require the production of 18 moles
of laaate by glycolysis. Thus, a small increase in PDH activity can account for a large
nducticm in lactate production with quivalent eaergy production by the d.
Despite insuiin treatment, extraœUular laciate &ase remaided persistesitly el&
cornparrd to non-ischemic cuntrol cells. Similariy, high en- phosphate levels mmained
demonstrated that in conditions of zero or low flow ischemia, the accumiitation of lactate,
NADH and hydrogen d t s in signifcant myocardial damage mlated to the los of tissue
adeaine n~cleotides.~ Of interest, they found that the recweq of ventricular function
independent of the tissue adenine nucleotide levels, but was i n v d y related to the tissue
lactate content. niese findings aiso support our hypothesis that the ratio of aerobic to
Hearse's early study demonstrated that insulin aECeriuaîed the pst-ischemic depression
in left ventricular funcbion due to to p i s c h e m i c mannitol infusion.' However, insulin
aacerboted the depression in LV function when administered in cardioplegic s01utions with
high gïucose concenîrat%ms. Rearse aüributed this exacerbation to glycolytic stimulation
during global ischemia leading to an accumulation of toxic metabolites. These results have
been confïmmi by other investigat~rs.~ However, insulin treatment in the -ce of low
flow cardioplegic -sion may have beneficial hemodynamic effeds.
in a canine model, Lucchesi demonstrated a direct inotropic effect of insului
treatrnenP2 W e developed a porcine model of orthotopic transplantation ta wduate the
effm of low flow m o n during hypothennic (4C) storage? Using donor blood

109
hamsisd h m the chest at the thne of organ pmmmmnt, we found that continuous low
flow (15 &min) pafusicm d e g thrre hours of storage perrnitted Iimited myocardial
metabolism. (Figure 25) Despite predomjnantly ariaerobic me&bolism, low flow penusim
led to a signifiant impvement in f i m c t i d recmery. (Figure 26) Insului was alen addeci
to the initial cardiop1egic solution, to the donor b l d pemirrate and 0 the blood dap1egic
solution at a amtinuous concentration of 10 NIL. Insdin treatment Ynproved the recovefy
of aerobic metaboiism and led to a further recovery of left ventricular fùnction. (Figure 27)
required intraaortic M o o n pump (IABP) support foilowing coronary bypass surgery and
found an Unprovernent in cardiac fiuiction.'" Svedjehohn et al used a glucose, insulin and
giutamate solution for patients who couid not be weaned b m cardiopulm~nary bypass?
These authors identifiai sixteen patients who were unable to wean fmm bypass and gave
these patients subsequentiy required the IABP to be weaned h m bypass and al i three died
of irreversible cardiac injury. The authors concluded that administrah . .
'on of gIutamate-GI
sohtion may potentially reverse transient myocardial dysfiinction and avoid the use of
mechanical support. Unforhsnately, these studies were non-randomized and did not d u a i e
In a preliminary prospective randomized chicai trial, we evaliiatPA the effects of
insulin enhanced cardioplegia in patients undergoing isolated coronary bypass surgery." We
found that patients who received insulin enhanced cdioplegia (10 WL) displayed an
improved transition h m anaerobic to aerobic metaboiism. Cornpared to persistent lactate

110
feiease in the plaîebo p u p , patients in the ioJuliD gmup w a e found to extrad ladate
îmmxhtdy aAer crossclamp reïease. (Fm 28) The dif'fém~ces in lactate flux disQwed
a f k five minutes of repafusion; bowenr, at two hairs following crossclamp r e m d
insulin mrdiuplegia Ied to an improved rramry of left ventriah fiinction. Om previous
findings (Figure 2) also demonshrated that a transient mefaboiic derangement in the earfy
repahision peziod predicted the developmerrt of postoperative low cardiac output syndrome.
Therefare, the transient effect of insulin during early qexfbsion led to enhanced left
ventricular hction duMg a time @od at which patients are most at risk for suffeming
from Iow output syndrome. A iarper, muhicenter prosptive randomized trial is curzentiy
underway to d u a t e the ability of insulin enhanced cardioplegia to d u c e the incidence of
low cardiac output syndrome foUowing isolarPI1 coronary bypass surgexy.
Mechmbm of tire Znsuün mect
These studies demollstrated that insulin rnediated stimuiation of rnyocardial pyruvate
dehydrogenase led to improved metabolic mmvery foliowing ischernia and repfbsion. In
our cellular studies we found that this improved metaboIic status conferred protection against
cellular injury as assessed by trypan blue uptab. W e extrapokîed our cellular findings to
a porcine mode1 of prolonged hypothennic storage as weii as a clinid trial ewluating insulin
enhanaxi cardioplegia. In these in vivo studies, insulin resuited in rnetaboiic benefits which
translated into irnproved fiuictional recovery foliowing cardiopllegic arrest. UnforhinateIy,
in each study there was a delayed movefy of myocardial metaboliSm a n d h left ventricular
fwicîïon compared to pre-ischemic values. Exploiîing the mechanism of the insuiin e f f i
may allow for site-+c enhancement of the stimulatory eff- and further improvements
in both m-lic and fiinctional recovery.

Itis unclearifinsuîin m~prOteinLinaîcCadivityorifproteinkinaseC modulaus the
cellular q m s e to insulin. For example, Standaat and coiieagues demoastrated an
asSo&ion kianea insulin-mediated gbcOse tmqort ard imeased diacy1glyceroI l m l s in
BC3H-1 rnyocyte~.~~ The higha DAG 1evelS also led to activation of protein kinase C. The
sarne investigators also demonstrated a relationship between insulin-mediaîed giucose
transport and the DAGlPKC signalling pathway in rat Meta1 muscles.m However, Klip
demonstrated that insulin-mediated hexose transport in skeletal muscle was not dependent on
protein kinase C Insulin was also found to stimuiate both the N a + pimp
and the N a + antiporter by protein kinase C dependent pathways?" To M e r evaluate
the relationship between insulin and the protein kinase C Sgnalling pathway, Blackshear and
coileagues evaluated the phosphorylation of a major htraçellular substrate for PKC,
MARCKS (rnyristoylated, alanine-rich C hase s~bstrate).~ In Uiese elegant studies, the
authors employed fibrobiast ce11 lines that overexpressed normal human hsulin receptors.
These authors found that insulin activateci PKC to a minor extent in ceil lines that M y
overexpress insulin receptors, but cuncluded that this stimulatory effect yas iiiUely not of
physio1ogic importance. However, these studies did not incorporate models of hypoxia or
ischemia, Hypoxia has been shown to stimulate several insuiin-sensitive proteins such as the
GLUT4 glucose transporter.- In addition, hypoxic andlor ischemic stimuli have been
shown ta stimulate protein kinase C a~tivity.~'-

1x2
The stimiilatim of protein kinase C activity foUowing a brief ischemic insult bas bea
shown to induce aie ischemic pmcmdïtioning phenorne~~l.'- This efkct, Onginaiïy
d e s c r ï ï by Murry and cu11eagues in 1986,1m is the most powerful endogenous form of
myocardhi protection cüswvered to date. Several stimuli have been show to mimic the
preconditiothg &kt including but n d timited to a d e n ~ s i n e , ~ a-adrenergic
agents,ln- bradykini.n302 and endotbelin-1? In our cellular studies, we found that insulin
exposure resulted in a &indocation of protein kinase C-a from the cytopiasm to the
membrane (Figures 20 and 21). A similar translocaiion was observecl following a tweaty
minute ischernic preconditimhg stimulus and exposure to the phorbol ester PMA?
However, the in vitro kinase activity of PKC w a ~ found to be only miaimally stimulated
foiiowing insulin and high giucose expure. Ln contrast, both ischemic preconditioning and
PMA exposure d t e d in a substantial increase in kinase activity. Simüarly, compared to
either ischemic pfeconditioning or PMA exposure, insulin treatrnent resuited in only a minor
reduction in œUular injury. However, the ability of ischemic preconditioning to privent
myo*irdial dysfunction is q u e s t i o ~ b 1 e . ~ ~ In contrast, insulin has been demonstrated to
i m p e fiinctional recovery foliowing The differeaces may be due
to differential isofonn-specific translocation and stimulation of protein kinase C.
Banerjeets group dernonstrated in isolated rat heart that both transient ischemia and
phenylephrine treatment resuited in a translocation of PKCd and PKC-dm Interestingly,
these authors found that PKC-b branslocated to the sarcoIemnai membrane whereas PKC-r
was observed to enter the myocyte nucleü. These studies employed direct in-situ
immunofluorescent staining gmilar to our techniques employed in chapter three., however,
they were unable to detect a redistribution in the cr or B isofoms. in contras& we found that

dkmibuticm was observeci with the eiJofOrm. We cantirmed our h m u n o f l u ~ t and
dot blot analyses with an in vin0 asay to m*rcnue the kinase activity of PKC and found that
insulin d t e d in a stimulation of PKC activity. Uafortuaately, cornparrd to the results
obtained with either ischernic pmxmditiOnjII~ or PMA exposure, insulin treatment yielded
only a modest stimuiatim of PKC d v i t y . However, PMA -sure d t e d in a simiiar
stimiilstion of pyruvate dehydrogenase d v i t y as thaî observeci with high giucose and insuiin.
Therefore, it is W y that a Sngie PKC-isoform, probably PKC-CY, stimulated by both PMA
and insulin is respcmsiiIe for the stimulation of myocardial PDH. Supporthg this hypothesis
are the findings of M i e and Van Wylaiw who found that ischemic preconditioning
impmved the preservation of myocardial high energy phosphates and reduced myacardial
iacîate production. Additional isoform stimulation by PMA andfor ischemic preconditioning
may be responsible for the added protection not seen by insului hreatment alone.
Our results are in contrast to those obtained by Banerjee's group and is iikeIy due to
species-specific signalling pathways. Importantly , Banerjee's studies demonstrated that PKC
inhibition in the contml condition did not exacerbate ischemic injury. Furtheniore, PKC
translocation occurred rapidly within minutes a& the brief ischemic insuit. Thus, PKC must
reguiate a positive protective mechanism foiIowing Qther a brief ischemic or pharmacologie
stimulus and is capable of affording protection against subsequent prolonged ischemia.
The time course of the preconditioning e f f a is also important. Kloner's p u p
demonmatai in rats that repemision perïods for greater than one hour foilowing the
preconditioning stimulus abolished the protective effects. * However , a second " whdow"
of protection appears after approximately Wenty-four heurs.= DDumably, ably second

114
window e&d is a d t of altered gme ~ 0 0 in the pfec~aditioned myocardium.
Unfbrûmkiy, the chatzicteristics aad mechanisms of the sa'oad window e&d are vexy
pwriy undezstood. We f m d that insulin nposure for thirty minutes d k m û protedion
agaïmt ischemia and was also sufncieat to cause PDH and PKC stimulaiion. We did not
stimuiaiion by in& acts via a similnr mediam'am to ischemic preconditioning, it aui be
postuiated that protection against ischemia may be COIlfenied for as long as twenty four hours
afkr insulin treatment. If this if found to be true, then patients who present for
m e o n can be treated with insulin on the day @or to su~gery.
The relationship between PKC-a st imWon and PDH phosphatase stimulation
=mains w1cIear. In our studies, we found that insulin mediateci PDH stimulaîicm was
abolished by the PKC antagonists chelaythrine and calphostk C. Furthermore, exposure to
the phorbol ester PMA stimulated PDH activity to a si& extait as observeci following high
glucose and insulin treatment. Exposure to high glucose and insulin in the presence of the
PDH phosphatase inhibitor, sodium fluoride, aiso prevented PDH stimulation. T h , we
conclude that the effect of insului on myocardial PDH is dependent upon PKC mediated
stimulation of the PDH phosphatase. Lamer's group has also anived at a similar conclusion
in both bovine heart mitochondriaUD and adipose tissue.u1
The PDH phosphatase is comprised of two distinct subunits and is the most abundant
mitochondrial phosphoprotein ph~sphatase.'~ The catalytic activity is associated with the
srnalier subunit (50 kDa), while the larger subunit (97kDa) is a f1avoproki.n of unlaiown
fimction. nie phosphatase nequires magnesium and at saairating concentrations (10 mmol),
is capable of ten-fold stimulation by minomolar conœntratims of caici~rn.~ Binding studies

115
have demonstrated that the phosphatase binds to the E2 sabunit of the cornplex provided
calcium is pmmt? One caicium ion is reqwred far eech 147kDa unit of the phospm.
However, when the phosphatase binds to the E2 subunit, a second calcium binding site
becames appamt. It is betieved that the fint calcium bindirig site plays a pivotal d e as a
bridging ligand between the phosphatase and the E2 subunit while the second 'nding site
altefs the seasitivity of the the E2-p- mmpk to magnesiun, thefeby increasing
PDH phospharase activity with migomolar hcrements in caicium concentration^.^
The ability of insulin to Stimulate PDH phosphatase in permeabiihd mitochouciria
argued agahst the d e of an intennediary moiecule. However, when mitochomiria are
disnipted, the isoiated PDH phosphatase no longer remains sensitive to in~uiin.~' This
evidence, combined with the o b d o n s of a membrane bound phosphatidyl inositol glycan
menistm'56, suggests that a second messenger, k l y bond to the mitochondrial b e r
membrane is required to mediate the stimulatory e f f i of insulin on the PDH phosphatase.
W e hypothesize that PKCa stimWon resuits in the release or alteration of this
rnitochondrial membrane phospholipid which d t s in an hcrease in mitochondrial caicium
concentrations leaduig to enhanceci PDH phosphatase activity. This hypothesis would explain
why isolated mitochondria from insulin treated tissues retain stimdated PDH activity and
calcium sensitivityu' while completely extracted mitochondria do not demonstrate a sensitivity
to in~ulin.~ There is iïkely a cornpkx signalhg pathway between PKCo stimulation and
the modulation of the membrane bound phospholipid which may prevent dinct PDH
stimulation by PKC agonists in isolated mitochondria.

5.4. ADDITIONAL EFFEcTs OF INSuIm ExmmRE
The studies reported in this îksis fociucd on the efféct of insulin cm myocaiidial
pyrwate dehydmgenase. By utiliPag an iso- d dture model, we were able to m . elunuiate the effects of other cell types and organ systems. In addition, this model allows
for the careful c o ~ o l of extracellular conditions. Although useful for investigating a spedî~c
metabolic pathway, our model may not accuately represent in vbv conditions.
Insu& and Fmty AM MeÉQbOlism
Circulatingfreefattyacidslevelsa~animporiantfactorinischemiaandrepemisim.
Lopeschuk and colleauges have demonstrated that under aerobic amditions fatty aads are the
preferred substiate for energy production by the heart while during ischemia anaaobic
glycolysis pred~minates.'~ However, in an isolatPA rat heart model they found that pasistent
fany acid oxidatim during repemision inhibited glucose oxidation and depressed functicmai
fecovery. Stvnulating giucose oxidation during repafusion was found to irnpme mechanical
rrwvery. Similarly, using a camitine paimitoyhransfease 1 inhibitor to v e n t fatty acid
oxidation, these investigators demonstmed that glucose oxidation was improved leading to
enhanced recovesy of ventricular function."' In a cIinical trial in patients undergohg isolated
coronary bypass surgery, Teoh and colieagues demonstrated thaî "42 W e d pairnitate
extraction was increased during cardioplegic arrest, but that B-oxïdation of this fàtty acid was
inhibited.P Myocardial fàüy acid accumulstjon wïthout oxidation was found to be deleterious
and may reflect a derangement of normal myocardiat metabohm during repemision. Otha
agents employed to stimulate glucose oxidaîion during reperfùsion such as dichlofoacetafe
'- and molapneaio have resulted in improved functionai recovery foilowing ischemia.
Both of these agents act to stimulate mitochondrial ppvate dehydrogenase. Futha evidence

117
of suppressed gïtmse oxidation was pvided by Liedthe's group in a porcine mode1 of
regional ischemia and npafusion.' These investigators found that suppressing fatty acid
unda conditions of either physiologie or high ciratlating levels of fatty acids, glucose
oxidation during reperfusion was inhibited.
To date, th= have been no repas of direct measwements of PDH activity in
human rnyocardium subjected to ischemia and repemision. The in vivo e f f e of insulin on
hurnan rnyocardial PDH also r e d unknown. Thus, the beneficial hemodynamic effects
of insulin may by unreiated to a stimulation of PDH activity. Most investigators measure the
relative rates of 14-C lactate and tntiated water production as a surzogate assessrnent of PDH
activity?- However, Lewandowski and White perfonned direct measurements of PDH
acfivity in rabbit hearts using NMR spectrosoapy to assess "C ppvate oxidati~n.~~ These
authors fouad that dichloroacdate did not improve the function of normal hearts, but did
prevent functional impainnent in hearts that sUnered 10 minutes of global ischemia. PDH
acîinty was increased
in the hearts that received dichloroacefate and correhted directly with the recovery of pst-
Insuiin has bem show by many investigators to alter myocardial fn+ faay acid
me&bolism.- Early studies by Roger!? and McDanielm demonstrated that glucose-
insulin-potassium solutions effectively decreased circriliiting ke fatty acid Iwels and reduced
my<x#rdial fatty acid uptake by up to 75%. A recent study by Knuutim employed positron
emission tomography to evaluate myocardial giucose uptake in patients wïth coronary artay
disease. ?me investigators found that myocardial gïucose uptake was increased to a similar

118
extent by both insuiin and acipox, an antilipolytic drug which inhibits fatty acid synthesis.
However, myocardial giuawe uptake was not related to saum insulin leveis, but rather to
cirmlating fkee fâtty acid levels which were mdcdly duceci during insuiin admkkmicm.
T'us, an a d d i t i d beneficial e f h t of iasulin ûeaûnent may be to redwe free fktty
acid syathesis and b reduœ the accumulation of potentially toxic fatty acids in the
myocardium. Since our mode1 does not accomt for tfus efkct, the in MW response to msulia
treatmerit may be greater than the in vin0 response observed in these studies.
ZnnJul ami rhc Sodium-Hjdmgm Exdanger
The sodium-hydrogen antipt is a plasma membrane protein pce~ent in most ceil
types, but most notably in cardiomyocytes and renal tubular tells. During ischemia, this
protein exchanges potentiaiiy toxic hydmgen ions for sodium.m Unf-ly, this resuits
in an accumiIlation of inbracellular sodium which is removed by either the sodium-potassium
ATP dependent pump or the sodium4cium exchanger. Due to the low intradiuiar leveis
of AIT during ischemia, the sodium-potassium pump is ineffective and thus intraceuuiar
sodium is repiaced by dcium. Further increases in in trac el lu la^ calcium stores may occur
during reperfusion leading to mitochondrial swebg and ceU death, a phenornenon termesi
"The calcium p a r a d ~ x . " ~ ~ WMe the calcium paradox is not present in these studies due
to the presence of calcium in the phosphate buffered saline, the effects of caicium overlœd
due ta other pathologie mechanisms may exist.
Several investigators have suggested that inhibition of the Na-H antiport wodd reduce
the accumulation of intracellular sodium and thereby p e n t calcium-induced inju~y . -~
Several inhibitors of the Na-H+ pump inc1uding amiloride and HOE-642 have k e n show
to pmtect against both infarctm and stunning foiiowing is~hemia.-~'"

119
This membrane protan quires phospharylaticm to kanne activated. Xncerpi and
coileagues danonstrated in nit hepatocytes that insulia stimulated the Na-H antiport SlCtiYity
by a protan kinase C depndent rnecbism." Howmr, inhibiton of protein kinase C
aüemaks preamditioning and dœs not provide protection by blocking the Na-H+ a n t i m L
Furthermare, inhibition of the Na-H+ antiport pmte!cts against ischemic injury even in the
presenœ of PKC antagOIilSfS?
It appears counterintuitive thaî insulin stimulates the Na43 antiport while at the same
t h e deniRg protection against ischemia. This apparent contradiction rnay be zes01ved by
examinhg the inüaceliuiar high energy phosphate data obtained in chapter two. Insulin was
found to preserve ATP lewels follmwing ischemia It is poss'bte that insuiin provides sufficient
ATP presentation to allow for sodium-potassium exchange to occur via the ATP-dependent
pump. In this way, inûaceiiular sodium is removed without a quisite increase in calcium
concentrations. Opie's group arrived at si& mnclusions in an isolatprl rat heart
investigation? These authors found that lactate-pemised hearts displayed greater injury than
pynnrate pafused hearts despite a less severe acidosis. However, in the pyrwate perîûsed
hearts, greater ATP IeveIs were thought to aüow for adequate N a 4 exchange preventing
sodium-calcium exchange. In con-, the Na-K pump was thought to be inhibited in &aîe
pemised hearts due to low intraceiiular ATP lewels and thus inmiiuiar sodium was
exchanged for calcium, leading to ceii injury.
Sampson and coileagues also reported a d i n a stimulatnry effect of insulin on the Na-
K pump in skeletal m~scle.~' Of interest, this stimltlltory effect wwas also mediated by
protein kinase C. Thus, insulin may act to increase H+ efflux by stimulation of the Na-H+
antiport while at the same time preventing sodium (and calcium) accumulation by stirnWon

120
of the Ni& purnp. In these saies of e q e h m , we did net directy measme hydrogm
enlux but did notice a protective e 8 4 of insulti agaiiist extraœUuïar acidosis. Direct
mawnments of Na43 activity a d o r sodium-po-um uchange during ischemia and
repafusion would provide important dditioiial infbmmion about the intracelluiar e f f i of
insulin.
53. -c EFFECIS OFlNsuIm
Although insulin receptars are present on alrnost aU cell s d à œ s , différent organs and
tissues may have different respcmses to hormonal exposure. In partic&, the metabolic
response of endothelhl ceiis may be rnarlϟly different from that observed with ventncuiar
cardiomyocytes. For example, Piper's grwp demonstrated that coronary endothehi tells
rely predominantly on anaerobic glycolysis for energy producfion in anitnast to culturied rat
cardiomyocytes which continue to oxidize fatty acids.- Of note, endotheiial celi lactate
production remained constant even as the atmosphezic pO, dropped h m 100 mmHg to 3
mmHg." M o w 3 mmHg, iactate production increaseû invefsely as the pO, dropped to O. 1
m g . This 'Pasteur effect' is markedly diminished compareû to UI vivo cardiomyocytes
which begin to produœ large amounts of lactate at much higher oxygen tensions. Piper
suggests that coronary endothelial cells dispiay the 'Crabtree" effect. This phenornenon,
originaUy described in 1929 in tumour cells, refers to persistent anaerobic glycolysis despite
high oxygen ten~ions.~ Due to th& anatomic lucation, coroaary endothelial ceiis are W y
not affecfed by endogenous ïactate production which is released into the circukion uniike
cardiomyocytes which are exposed to the lactate and hydrogen ions which they p d u c e .
Since conniary endothW ceîi metabolism is highly resistant to the ambient oxygen tension,
it is unclear if a metaboiic intervention such as insulin would provide benefit.

121
Although vmtricular dysfiuicticm is ciearly due to cardiomyocyte injury, thae is
incieaSiLlg evibce that endothelia1 ceil dysfuncticm contributes signifïcantly to ischemic
npafusim injury- Endothelhl alls produce a potait vasoaceive compound now
idmtified as nitric oxide? Although nittic oxide itself is a negative isotrope,-
preservation of endotheIial fwiction has ken shown to imprwe veatricuiar function foilowing
Cardioplegic aIrest,"
Scharer and coUeagues demonstrated in hurnans that the vasculâr &kds of insulin
are mediated by nitric oxide release. If insuiin exposure can increase aidotheliaî QU nitric
oxi& production, it is possible that insulin may protect endothelid ds against ischemia.
To invdgate this hypothesis we perfôrmed additional expaiments employing primary
cultiaes of human saphenous vein endothelial ds.- Endothefial ceLls were found to be
highly susceptible to ischemic injury compared to hwnan ventricular cardiornyocytes.
FoIiowing ninety minutes of ischernia and ovemight incubation in serum-free medium, oniy
15% of the endothehl ceUs at ri& remaineci cornparrd to over 40% of the cardiomyocytes
(Figure 23). Insulin treatment improved cardiomyocyte survival but did not affect endothelial
ceil tolerance to ischernia, Howeva, insuiin did stimulate endothelial ceïi PDH activity and
reduced extraduiar lactate re1ease (Figure 24). It is therrfore possible that in& may
pmtect endothelial cells against a less severe ischemic insult.
5.6. RJMMARY OF INVESi'IGAATIONS AND ORIGINAL CONTRIBUTIONS
These investigations evalUiltPA the rnetabolic response of isolated human ventriadar
cardiomyocytes to simulated ischemia and w o n . Insuiin treatmmt was found to . stimulate aerobic metabiism and protect cardiomyocytes against ischemic injury. The nsuIts
of these investigations have yidded the follohg original contributions to the literature:

l22
Human myacardial pyruvate dehydrogaiarre is inhr'bited following simutnteA isc-
and repemisi011.
Insulin exposure results in a sîimWoa of human myocardial pynniate
dehydrog-.
The stimcilaiDly dfEa of insulin is mediated by increased PDH phosphatase activity
and is profeia kinase C s riepaiQent,
The stimiilatory efféct of insulin is not depaideat on ambient glucose concentrations.
FUTURE AREAS OF RESEARCH
These studies provide a rnetabolic ratide for administering insulin prkx to an
anticipated ischemic insult. Cardiopllegc amst is a unique situation in which ischemia cm
be anticipated and interventions such as insulin can be employed to irnprove myocardial
metabolic and fiinctional recovery. 'Ibo cluiical trials are cumntly evaluating the ability of
insului enhancexi cardioplegia to stimulate aerobic metabolism and impme venbricuiar
function following cardioplegic arrest. In the fint trial, insulin cardioplegia was found to
stimulate ktate extraction following aortic crossclamp removai. This impmved m v e r y of
aerobic metaboihm was fond to pnserve lefi ventncular fiuiction a k two hours of
repafusion (Figure 28). The second trial is a larger, multiceriter trial designed to assess the
ability of insulin cardioplegia to reduœ the incidence of perïoperative myocardial i.nfiu&on
and postopefative low output syndrome. The results of this trial will be available in
approxirnately two years.
Stimulation of aembic metabolism by other pharmacologie agents, such as
dicblatodcetafe, has been found to irnprove myocardial hinctional recovery following
is~hemia.'~~~-"'~ Dichloroacetate has not been e v a l d clinically as a cardiaplegic

123
additive. A d d i t i d clinical trials are requireû to as ses^ the beaefit of dichlmmcetak with
or without insulin.
Pafusing cardiac aiiograftp d m stmage with donor blood erihanced with insulia
was found to improve f u n c t i d t i ~ c o v q h11owing OZttlotopic transplantation. The
hemodynamic &ectS of insulin may be ben- in the &y pst-txansplant period if one
requires a positive inotmpe which a h red- systedc niscular r e s i s t a n ~ e . ' ~ ~ ~ ~ Further
investigations are required to evaluak the clinid baielits of insulin treatment either before,
duriag or affa cardiac transplantation.
The e f f e of insulin were demOIlStrated to be mediateci by PDH phosphatase
stimufation via a PKC-a dependent pathway. New phamiacologic agents designed
specificaily to enhance and augment this response may prove to be beneficial as dopllegic
additives. The additional molecuiar and biochemical eff- of insulin enposure, such as
sodium-hydrogen exhange and sodium-potassium exchange, require m e r investigation.
These investigations have provided additional evidence üiat stimulation of aerobic
metabolism following ischemia protects against cellular injury and may enhance functional
recovery. Additional methods of resuscitaîing the ischemic myocardium are required as the
proportion of high risk patients presenting for comnary bypass surgery inaeases.
Furthemore, enhancing myocardial tolerance to ischemia may aUow for an expanded use of
donor organs which are currently deerned unsuitable for ûansphntation. Cardiovasaikr
disease remains the predominant cause of m d t y in North Amedca. Coronary bypass
s u r g q is an important intervention for the treatment of atherosc1erotic heart disease. Novel
techniques of myocardial protection are required to reduce the risks of surgery in an
increasingly high risk patient population.

-1.2 I
PRE XCL XCL ON OFF
LVEDP (mmng) :
Figure 28: Upper Panel: Myocardial lactate flux during and after cardioplegic mest. Patients 1
who received insulin enhanced cardioplegia displayed lactate extraction , immediately afkr crossclamp removal compared to persistent lactate! release in the placebo group. Lower Panel: Left ventricular function was better preserved in the insulin cardioplegia group after two hours of reperfusion.

Parisi A., KhUri S, Deupree RH, Sharma GVRK, Scott SM, Luchi RI: Medical mmpaied with surgical managemat of WlStable agiria: 5-year mortality and morbidity in the Vetaans Adannistrati - . 'cm Study. CïrculatiOn 1989; 80: 1176-1 189
Caüff RM, Harrell FE J, Lee KL, et aï: The evolution of medical and surgical therapy for corniary artery disease: A 15-year perspective. JMM 1989; 261: 2077-2086
Richadson RG: BiIlXoth and cardiac surgery. Lancet 1963; 1: 1323
Rehn L: On petrathg cardiac injuries and cardiac suturing. Ardi Min Chir 1987; 55: 315
Beck CS: The devdapmentof a new blood supply to the heart by operation. AM Swg 1935; 102: 805
Vineberg AM: The development of an anastomosis between the coronary vesstls and a transplantecl intanal rnammary artay. Cim M d Assoc 1 1946; 55: 1 17
Sones FM, Shirey EK: Cine coroaary arteriogzaphy. M d Concepts ~ O V O S C Dis 1962; 31: 735
Carrel A: On the experimentai surgery of the thoracic aorta and the heart. Ann Surg 1910; 52: 83
MeIrose DG, Dreyer B, BaitaIl HH, Baker JBE: Elective cardiac arrest. L.uncet 1955; 2: 21
Helmsworth JA, Kaplan S, Clark LC, et al: Myocardial injury assoQated with asystole induced with potassium citrate. ANI Surg 1959; 149: 200
Gay WA: Potassium induced cardiopIlegia. Ann ~ t c u : Swg 1975; 20: 95-100
Bigeiow WG, Lindsay WK, Greenwood WF: Hypothermia: its possible d e in cardiac ~ u r g e f ~ . ANL S W ~ 1950; 132: 849-866
Favaioro RG: Saphenous vBn aumgraft replacement of severe segmental comnary artery occlusion. AM Thorac Szug 1968; 5: 334

Myas WO, Gersh BJ, Fisher ID, et al: Mrdical vexsus eady surgical therapy in patients with triple v d dïsease and miid angïna psctoris: A CASS ngistry study of survival. Ami ~ ~ U P M C Slvg 1989; &O: 11761189
TaIram T, Peduzzi P, De&e KM, et al: SurvÏvai in subgroups of patients with ieft main annary diseesa' Vdaaas AdmmWmû . .
'on Cooperative Study of surgeq f a wronary arterial occlusive dis*ut. C&Z&&R 1982; 66: 14-22.
Christaich GT, Birnbaum PL, Weisel RD, d al: The changing pattern of coronary bypass surgery. CTrCJQtiOon 1989; 80: 151-161
Ikonomidis JS, Rao V, Wdsel RD, Hkyashida N, Shirai T: Myocardial protection f a oorcmary bypass surgeq: The Taront0 Hospital Perspective. Ann Zbroc Swg 1995; 60: 824-832.
Misare BD, Knilrenkamp IB, LevitPky S: Agtdependent seasitivity to unprotected cardiac ischemia: the senescent myocardium. 3 lhorac Caniovpcc Saqg 1992; 103: a 6 5
Christakis GT, Weisel RD, Fremes SE, et al: Coronary artay bypass surgery in patients with poor left ventncvlar fiuiction. J %rat Cordiovasc Surg 1992; 103: 1083-1092
Ctiristakis GT, Frwnes SE, Wusel RD, et al: Reducing the risk of urgent -OR for unstable angina: randomized clinid trial. J Varc Surg 1986; 3: 764-772
Teoh KR, Christdis GT, W-1 RD, et al: Increaseü risk of urgent revascularization. J %rat Cordiowsc Swg 1987; 93: 291-299
Weisel RD, Mickle DAG, Finkie CD, Tumiati LC, Madonik MM, Ivanov J: DeIayed myocardid me!taboIic recovefy after blood cardiopIegia. AM 27wtac Surg 1989; 48: 503-507
Yau TM, Ikonornidis JS, Wasel RD, et al: Which techniques of cardiaplegia m e n t ischemia? ANI lhOrac Swg 1993; 56: 1020-1028
Yau TM, Weisei RD, Mickie DAG, et al: Alternative techniques of cardioplegia. Grcularion 1992; 86(Suppl II): IUT-384
27. Best JB: Physiologicai Basis of Medical Practiœ. WzZiium and W r ' h 1985; 11: Baltimore.

Silvennan NA: Myocardial oxygen collsumption after reversible ischemia. J Cmd Surg 1994; 9(suppf): 4 6 5 4
Knilrenkamp IB, Sitvefman NA, Sorlie D, et al: -011 of postischemiç myocardial oXygen ritüizatim CCIiCUlatiOn 1986; 74(suppl III): Iïïl25-129
Opie LH, Owen P, Riemersrna RA: Relative rates of oxidation of glucose and fkee fatty aQds by ischaemic and non-ischaemic myocardium a f k coronary ligation. Eut J Cün Illyest 1973; 3: 419-435
Tedi KH, Mickle DAG, Weisel RD, a al: Decreased postopaative myocardial fatty acid oxidation. J Surg Res 1988; 44: 36-44
Twh KH, Mickle DAG, Weisel RD, et al: The effect of lactate infusion on myocardial meCabolhm and ventriculat function foiiowing ischemia and cardiap1egh C h J (=(vd 1990; 6: 38-46
MickIe DAG, del Nido PJ, W i n GJ, Harding RD: Exogenous substrate preference of the post-ischemic myocardium. Çcudiovasc Res 1986; 20: 256-263
Opie LH: Metabofic response during impending myocardial infarction: Relevance of studies of glucpse and fatty acid metaboiism in animais. Cit&on 1972; 45: 483-490
Ikonomidis JS, Ra0 V, Weisd RD, et al: Myocardial Iactate production during npemision predicts low output syndrome fououring CABG. C b J Cardi02 1995; 1 1 0 : 122E
Lawson CS: Does ischemic preconditioning occur in the hurnan heart? Cardiowc Res 1994; 28: 1461-1466
Ra0 V, Ivanov J, Weisel RD, Ikonomidis JS, Ch&akis GT, David TE: Predictors of low cardiac output syndrome a h coronary artery bypass. I Thorac CardiovQsc Swg 1996; 112: 38-51
Cross HR, Clarke K, Opie LH, Radda GK: 1s laictate-induced myocardial ischaemic injury mediated by decreased pH or increased myocardial lactate? J Mol Ce22 Cardi02 1995; 27: 13694381

N d y JR, Grotyobann LW: Role of glyco1ytic producfs in damage to ischemic myocardium. CCrc Rcs 1984; 55: 816424
Cross HR, Opie LEI, Radda Gg, Cfarke E. 1s a high giycogm content beneficial or detrimental to the ischemic rat hearî? CPrc k 1996; 78: 482-491
Weiss J, Hiltbrand B: Fundionai compartrnmfaticm of glycolytic vgsus oxidative metabolism in isolateA rabbit heart. J Clur Imirst 1985; 75: 436447
Weissier AM, Kruger FA, Baba N, Scarpell DG, Lüghfon RF, GaJli.more TK: Role of anagobic metaboliSm in the pnsavation of fundonal capacity and stnicanc of anoxic myocafdium. J Clur Invest 1968; 47: 403-416
Sodi-Pollares D, Testelli MD, Fieder BL, et al: EffectS of an inüavenous infusion of a potassium-glu~insulgi solution on the elecbKlcardi~graphic signs of myocardial infarcton. Am J Cardioz 1965; 5: 166181
Akagawa H, Tajiri T, Hoshino Y, et al: Questionable role of glucose-insulin in cardioplegic solution. J C o r d o ~ ~ ~ c Surg 1983; 24: 215-221
Apstein CS, Gravino FN, Haudenschiid CC: Determinants of a protective effect of glucose and insulin on the ischemic myocardium: Eff- cm contractile fiinction, diastolic cornpliance, metabolism and ultrastnichire during ischemia and reperfusion. C c r ~ R ~ s 1983; 52: SIS-26
Bormann BV, Scheld HH, Podniweit T, Boldt J, IEltig D, Hempelmann G: Enhancement of myocardial energy potentials in man by giucowinsulin treatmmt before and after ischemic heart arrest. J Cludiov~~~c Surg 1985; 26: 182-186
Haidex W, Benzer H, Schutz W, Wolner E: hprovement of cardiac preservation by preopemtive high insulin supply. J 27wm Cardiovasc Surg 1984; 88: 294-30
Iyengar SRK, Charrette ETP, Iyengar CKS, Wasan S: Myocardial glycogm in prevention of perioperative ischemic injury of the kart: A prelunuiary . m .
fePOft* G?Jz J Surg 1975; 19: 246-251
W e y DM, Ray IF, Myers WO, Sautter RD, Tewksbury DA: Importance of preopeaative myocardial giycogen lewels in human caidiac pfesefyation. 3 Thorac ~ o w c Surg 1979; 78: 678-687
Rousou JA, Engelman RM, Anisimowicz L, et al: Metabolic enhallcement of myocardial preservation during cardioplegic amst. 3 niomc ~ O Y Q T C Swg 1986; 91 : 270-276
Saleni0 TA, Chiong MA: CardiopIegic arrest in pigs: Efkts of glucose-containing solutions. J lbrac Cardioiurrc Sug 1980; 80: 929-933

Conti VR, Ica0 RL: Metabolic and fimdignaleff8dS of carbohydrate &strate with single dose and multiple dose dopiegia. Amr SUR 1983; 36: 320-327
Doherty NE, 'LiPocy IF, Geffin GA, O'Keefe DD, Titus JS, Daggett WM: Beoefits of glucose and oxygai in multidose cold eardiop1egia. J Thoryu: î2dbw.s~ Ssug 1992; 103: 219-229
Ebali FR, Weinberg EO, Gnce WN, Hmwitz GL, CS: Protective eEéct of increased giycolytic subsaabe against systoiic and diastoiic dysfiiacticm and increased conniary reSXs&nce from prolonged giobal uadeqmhïon and reperfusion in isolated rabbit hearts pemisrd with erythiocyte suspensions. Cix Rcs 1991; 68: -81
Fremes SE, Funikawa RD, Li R-K, Twniati LC, W& RD, Mickle DAG: In vitro assessrnent of the effects of giuaxe added to the University of Wisconsin solution on myocyte preservaton. CirculCLtiOn 1992; 86(suppl II): 1128941294
Fnmes SE, Guo LR, F\uukawa RD, Mickle DAG, Wekel RD: Cardiac storage with UW solution and glu-. A m InOruc Swg 1994; 58: 1368-1373
Guiibeau EJ, Moore LK, Viole AJ, et al: Effect of intermittent fisions of glucose-containing crystalioid cardiopkgic solutions on myocardial tissue lactic acid and recovery of amtractility. J ~ r a c CMüomc Surg 1984; 87: 920-929
Hearse DJ, Stewart DA, Braimbridge MV: Myocardial protection during ischemic cardiac arrest: Possible deleterious effects of glucose and mannitol in mronary infusates. J Inoroc Cardiow~sc Surg 1978; 76: 1623
Hewitt RL, Lulley DM, A b u n y GA, Drapanas T: Protective effect of myocardial glycogm OII cardiac fiuiction during anoxia. Surgery 1973; 773: 444-453
Hewitt RL, Loiiey DM, Adrouny GA, Drapanas T: Protedive effst of glycogem and glucose on the anoxic amsted hem. Surgery 1974; 75: 1-10
Lazar HL: Enhanced pnserVaton of acuteiy ischemic myocardium using giucoseinsulin-potassium solutions. J Coni Surg 1994; 9(suppl): 474-478
Liedtke Al, Hughes HC, Neely JR: E f f ' of excess giucose and insulin on glywlytic metaboiism duMg experïmmtal myocardial ischemia. Amer J Cord 1976; 38: 17-27
Lolley DM, He* RL, Drapanas T: Retmpemision of the hart with a solution of glucose, insulin and po@ssiurn during anoxic me&. J 2bruc Cardov~sc Surg 1973; 68: 364-370

Loiiey DM, Ray IF, Myers WO, Slaeldon G, Sautfer RD: RedUCfion of inmopemive myocardial hfhction by mains of exogenous maenhic substrate enhancement Piospective randomized sûxdy. ANI Ihom Sug 1978; 26: 515-524
MRC wodchg party cm the treatment of myocaidial infarcton: Poîasium, glucose and insulin treatment for acute myocardial idiuction. LO)DCU 1968; 286: 1355-1360
ûpie LH, Sruynell K, Owen P Effeds of glucose, iasulin and potassium infusion on tissue rnetaboiic changes within first honr of myocardial infarctioa in the babmn. amliaibn 1975; 52: 49-56
Onta H, Fukasawa M, HirooIca S, et al: Possible deleterious effects of glucose ai immature myocytes under hypothermie conditions. Amr &tac Surg 1994; 58: 1103-1 107
Owen P, du Toit EF, Opie LH: The optimal giucose concentraticm for intermitterit cardioplegia in isofatprl rat heart when addeci to St Thomas' Eospiîal carciioplegic soIution. J Thoroc Cmriiovtzsc Surg 1993; 105: 995-1006
Owen P, Dennis S, Opie LH: Glucose flux rate regulates onset of ischemic contracture in giobally und- rat hearts. arc Res 1990; 66: 344-354
Rao V, Cohen G, Weisel RD, et al: The use of glucose and insulin during h y p o t h d c and normothermic CABG. Amt N Y A d Sn' 1996; 793: 494-497
Steinberg JB, Dohezty NE, Munfakh NA, et al: ûxygenated cardioplegk The metaboiic and fwicîional eff#xs of giucose and insuiin. AM IhOrac Surg 1991 ; 5 1: 620429
von Oppell UO, du Toit EF, King LM, et al: St Thomas' Hospital cardiap1egic soIution: Beneficial effed of glucose and multidose reinfugons of cardioplegic ~0Iution. J ïbrm C ~ ~ O V ~ C Szug 1991; 102: 405-412
Wittnich C, Peniston CM, Sderno TA: The effects of glucoseumtaining solutions on ischemic myocardium. Ann C7hr Gynue 1987; 76: 77-83
Mittra B: Potassium, Glucose and insulin in matment of myocardial M o n . Lrutcet 1965; 443(2): 607609
Majid PA, Shanna B, Meeran MKM, Taylor SH: Insulin and glucose in the treatment of kart Mure. LMcet 1972; 2: 937-941
Fremes SE, Weisel RD, Christakis GT, et al: Myocardial metabolkm and ventncular fwiction foliowing mld potassium cardioplegia. J lhomc CanZiovasc Surg 1985; 89: 53 1-546

Welsei RD, Lipton 1, LyaU R, Baird RI: Cardiac metaboliSm and perfonaance following cold potassium cardiople@. û5lculmion 1978; 58: I217-226
Fnmes SE, ChristaLis GT, Weisl RD, et al: A ctinirrii trial of blood and uysfallloid cardioplegia J lhomc C 2 d b u s c Surg 1984; 88: 726741
Menasche P, Touchot B, Pradier F, et al: Simplified mdhod for delivering normothdc blood cardiop1egia. ANI mm Surg 1993; 55: 177-178
Yau TM, W e i d RD, Mickie DAG, et aï: Optimal delivay of blood cardiapllegia. arculan'ion 1991; &Ysuppl Iü): n][38&388
Buckkg GD: Integratcd myocardial management Background and Initial Application. J Cordioc Surgery 1995; 10: 68-89
Rosenfeldt FL: Cardioplegia: Blood or crystaUoid, Antegrade or retrograde, intermittent or orntinuous, warm or cold? Asio P m c J ï k ruc t2udhmc Surg 1994; 3(2): 71
Feindel CM, Tait GA, Wilson GJ: Multidose b l d versus crystalloid cardioplegia: cornpariscm by quantitative assessrnent of irrevdble myocanlial injury. J ZWmc Ckridiovasc Surg 1984; 87: 585-595
Gotî VL, G o d e z JL, Zuhdi MN, et al: Retrogxade m o n of the coronafy sinus for direct-vision aortic surgery. Surg GynecoZ Obsret 1957; 104: 319
Yau TM, Ikonomidis JS, Weisei RD, et al: Venûiculaf huiction afkr nmothefmic versus hypothermie cardioplegia. J 27wrac Movezsc Surg 1993; 105: 833-844
See YP, Wei& RD, Mickle DAG, et al: Prolonged hypothennic cardiac storage fa transplantation: the etfects on myocardial metabolism and mitochondrial functim. J &roc Cordioasc Swg 1992; 104: 817-824
Rosenkranz ER, Vinten-Johansen J, Buckberg GD, Okamoto F, Edward H, Bugyi H: Benefits of norrnothermic induction of b l d cafdioplegia in energy âepleted hearts, with maintenance of arrest with multidose, cold blood cardioplegic infusions. J Throc Cmdovasc Sutg 1982; 84: 667677
Rosenlaaoz ER, Bucldiag GD, Laks H, Mulder DG: Warm induction of d o p l e g i a with glutamate ainched blood in coronary patients with cardiogenic shock who are dependent on inotmpic drugs and intra-aortic balloon support. J W r 4 c CPmliovarc Surg 1983; 86: 507-518
Teoh KH, Chnstakis GT, Weise1 RD, et al: Accelerated myocardiat metabolic recovefy with terminai warm b l d cardioplegia (hot shot). J iIhortx ~ O W O S C Swg 1986; 91: 88&895

Lichtenstein SV, Ashe KA, El Dalati H, Cusimam, RJ, Panos A, Slu* AS: Wana Heari Surgery. J Ilu,mc Con3iowrc Sutg 1991; 101: 269-274
Salerno TA, Houck JI?, Bimou, CAM, et al: Retqmde contiauous warm biood cardioplegia: a new amcept E 111yocadiaî protection. Ann Thora Swg 1992; 51: 245-247
The Wann Hem Investigatm: Normothdc vemu hypothemiïc cardiac surgery: a randomhd triai in 1732 coronary bypass patients. Larrer 1994; 343: 559-563
Aronson S, Lee BK, Li- JR, et al: Asesment of retn,grade cardiople@ distribution using contrast echocardiograpby. Amr llromc Swg 1991; 52: 810-814
Crooke GA, Harris LEI, Grossi EA, et al: Biventricular of cold blood cardioplegic solution admimstered . . by differerit m g r a d e techniques. J Thorn Ckzrdiov~c Surg 1991; 102: 631637
QuintilIo C, VoQ P, Biiotta F, et al: Risk factors for incoqlete distribution of cafdioplegic solution during coronary artay grafting. J ~ m c Cardiovosc Swg 1995; 109: 439-447
Rudis E, Gates RN, Laks H, et al: Coronary sinus ostial occlusion during retrograde de1ivery of cardioplegic solution signincantly improves cardioplegic distribution and efficacy. J Thorac Cordovarc Swg 1995; 109: 941-947
Stirling MC,.McClanahan TB, Schott RJ, a al: Distribution of candioplegic solution infùsed antegradely and m g a d e l y in normal canine hearts. J T h o m Caniioyosc Swg 1989; 98: 10661076
Maatsum H, Lazar HL, Yang XM, et al: Detrimental effeds of interrupting wam blood cardioplegia during coronary revascularization. J Thorac Canliowc Surg 1993; 106: 357-361
IkonoMdis JS, Yau TM, Weisel RD, et al: Warm retrograde cardioplegk What happens when you tum it off? CM J CbrdioZ 1992; 8(B): 91B
Bhayana JN, Kalmbach T, Booth FV, et al: Combined antegrade/retrograde cardiop1egia for myocardial protection: a cîinicai trial. J Zhrac Crudiov(~sc Swg 1989; 98: 956960
I)rinkwater DC, Lalcs H, Buckberg GD: A new simplifieci method of optimiPng cardioplegic deliivery without right heart isolation: Antegrade/retrograde blood cardioplegia. J Thoruc Cardiovasc Surg 1990; 100: 5663
Hayashida N, Weisel RD, Shirai T, et al: Tepid antegrade and retrograde Csirdioplegh. ANI Thor~zc Swg 1995; 59: 723-729

Shirai T, Rao V, Weisel RD, et al. Anoegade and retqpde cardioplegkc Aitemate or simuîtaneous? 3 Inoroc Cmdiowrc Ssurg 19%; 112: 787-7%
Hayashida N, Tkcmomidis JS, Weisel RD, et al: The opbimal daplegic tempexatim. ANI %rut Swg 1994; 58: 961-971
Kuntschen FR, Galletti PM, Hahn C: Glucoseinsulin iateractims during cardiapulm~nary bypass. H y p o t h d v m N d - J Thoroc (=anüowmc Surg 1986; 91: 451459
SvedjehoIm R, Huljebtant 1, aalraason E, Va* 1: Glutamate and high dose gluminsulin-potassium (GIa) in the h*itment of severe cardiac Mure afte.r cardiac opefations. ANI 2hOruc Surg 1995; 59: S23430
Svmsçon S, Ekmth R, Nilsson F, Pontai J, Wiuiam-Olsson G: Insulia as a vasodilating agent in the first hour aRet cardiopuimonary bypass. S d J Thora C P n t i o ~ ~ c Surg 1989; 23: 139-143
CoIeman GM, Gradinac S, Taegtmeyer H, Sweeney M, F k e r H: Efflcacy of metabolic support with glucose-ïnsulin-potii.epjurn for left vmtncular pump mure afta aOTtOCOronary bypass surgexy. CfrCJlLtiOn 1989; 80(supplI): Dl-196
Gradinac S, Coleman GM, Taegtmeyer H, Sweeney MS, Frazier OH: Improved cardiac fwiction with glucose-insuiin-potassium after aoftocoronary bypass m g . AM lhorae Surg 1989; 48: 4-89
Svedjehoim R, Ekroth R, Joachimsson P, Tyden H: Highdose insulin impwes the efficacy of dopamine early aRer cardiac surgery. S c d J ZMr C o r d i o ~ ~ ~ ~ c Surg 1991; 25: 215-221
Lucchesi BR, Median M, men FJ: The positive inotropic action of insulin in the canine heart. Eur 3 Phormacol 1972; 18: 107-1 15
Teoh MI, Ivanov J, Weisel RD, et al: The surgical management of unstable angiaa: A cihical triai of warm induction giutamate aqmbk blood cardiopIegia for urgent revascularization. Ahtracts of the Amencan AssOCiCttion for Thora& Surgery 1990;
Crane FL, Hatefi Y, LRSm RL, et al: Isolation of a quinone from beef heart rnitochondria. Hochùn Biophysia Act0 1957; 25: 220-221
Chen Y, Lin Y-T, Wu S C : Effectiveness of coenzyme QI0 on myocardial

Okamoto F, Allen BS, BucLkrg GD, Leaf J, Wigyi H: Studies of amtroIled npafusion a f k ischemia, X: Repahisate composition: supplementai role of intravenous and intra#rn>nary cœqme QI0 in avoidug repesfusion damage. J lhomc Cmriiow~sc Surg 1986; 92: 573-582
tan ai^ J, Tominaga R, Y d h o s h i M, et al: Coenzyme Q10: the prophybctic aect cm low cardiac output fôliowhg ardiac vaive repiaœmeat. Ann Tlu,rac Siag 1982; 33: 145-151
Whitehouse S, Randle PJ: Activation ofpyruvate dehydrogemse in perfused rat hart by dichloroaœtate. WOdvm J 1973; 134: 65 1-653
Wahr JA, Childs KF, Bobg SF: DichlOrOacefate erihaaces myccardial functional and metabolic recovery following global ischernia. J Cardothorm*~ Vmc Anes 1994; 8: 192-197
Wahr JA, Olszanski D, Wds KF, Boihg SF: Dichloroaœtate enhanced myocardial fiinctional mmvery pst ischemia: ATi? and NADH recovery. J Surg Res 19%; 63: 22&224
h4azer CD, Cason BA, Stanley WC, et al: Dichloroace&te stimulata carbohydrate metabolism but does not improve systolic function in the pig heart. Am J Physiol 1995; 268: H879-885
LUm T, Petîit F, Reed L: a-Keto acid dehydrogenasae complexes X: Regulation of the activity of the pyruvate dehydrogenase cornplex h m beef kidney mitochmdria by phosphorylation and dephosphorylation. Plar N u l A d Sci USA 1969; 62: 234-24 1
Cooper RH, Rmde PJ, Denton RM: Stimulation of phosphorylation and inactivafion of pynivate dehydrogenase by physiologicai inhibitors of the pynivate dehydrogenase reaction. Nanue 1975; 257: 808-809 Robinson BH: Lactic Acidernia. In: Scriver CR, Beauùet AL, Sly WS, Vaiie D, eds. The Metabolic Basis of Inhdted Disease. New York, N'Y: McGraw-HiU Ress, 1989: 869-888
Sugden PJ, Rutson M, Kerbey AL, Randle PJ: Phosphorylatiun of additional sites cm pyruvate dehydrogenase inhibits its reactivation by pyruvate dehydrogenase phosphate phosphatase. Rochem J 1978; 169: 433-435
Pratt ML, Roche TE: Mechanism of pyruvate inhibition of kidney pynivate dehydrogenase kinase and synergistic inhibition by pynivate and ADP. J Bi01 Umn 1979; 254: 7191-7196

RaViiadran S, Radke GA, Gueat JR, Roche TE: Iipoyl domah-based mechanhm for the integrated f-k amtrol of the pynnratt dehydrogemae complejr by eahaacement of pynivate dehydrogeaase kinase acfiyity. J Biol Ciran 1996; 271: 653662
Denbn RM, Randle PJ, Martti BR: Stimularion by calciun ions of pyruvate ddiydrogume phosphatase. Iaodian I 1972; 128: 161-163
Lilley K, Zhang C, Vibr-Paiasi C, Laraa J, Huang L: Inailin mediator stimulation
Lama J, Huang LC, Slmilri S, et al: Insulin mediators and the conml of pynivate dehydrogena~e complex. Ann W A c d Sci 1989; 573: 297-305
Newmann JD, Armstrong JM, BoniStein J: Assay of insulin mediator activity with soluble pynnrate dehydrogenase phosphatase. hdoamCogy 1985; 116: 1912-1919
Mochinila S, Neely JR: Enefgy metabohm during reperfusiion following ischemia. J Physiol 1980; 76: 805-812
Liedtke AJ, Nellis SH: Effeds of buffered pynivate on regional cardiac function in moderate, short-term ischemia in swine heart. CYrc Res 1978; 43: 189.199
Kobayashi K, Neely JR: E f f ' of ischemia and repemision cm pynivate dehydrogenase acfivity in isolateri rat hearts. J Mol &C Gdiol 1983; 15: 359-367
Hucho F, R a d e DD, Roche TE, B ~ g e t t MW, Pelley W, Red U: Keto acid dehydrogenase complexes XW: Kinetic and regdatory properties of pyruvate dehydrogenase kinase and ppvate dehydmgenase phosphatase h m bovine kidney and hearî. Arch Biodrmr Bwphys 1972; 151: 328-340
Vary TC, Randle PJ: The effect of ischaemia on the activity of pyruvate dehydrogenase complex in rat heart. J Mol CeU Cmdd 1984; 16: 723-733
Lewandowski ED, White LT: Pynivate dehydrogenase influences postischemic heart function. Cixdatiion 1995; 91: 2Wl-2ûï9
Lopascfiuk GD, Spafford MA, Davies NJ, Waii SR: Glucose and pairnitate oxidation in isolated workhg rat hearts reperfûseû after a period of transient global ixhemia. Cïm Res 1990; 66: 56553
McVeigh JJ, Lopaschuk GD: Dichlomacetate stimutation of glucose oxidaîion improves recovery of ischemic rat hearts. Am J Physiol 1990; 259: H1079-1085

LopaJchuk GD, Wall SR, O k y PM, Davies NJ: Etoxomir, a carnitine paimibyltransfi 1 inhibitor, proteds h m h m fat& acid-induced ischemic injury indepeaderit of changes in long chain acy1carnitine. CCrc Rcs 1988; 63: 1036-1û43
Lopaschuk GD, SpafTord M: Response of isoW working hearts to fatty acids and carnitinepaimitoyltransferase I inhibition durîng duction of co~oxtary fIow in acutdy and chroaically âhbetic rats. CCrc Rcs 1989; 65: 378-387
Lqaschuk GD, McNd J, McVeigh H: Glucose orudation is stllnulatcd in repemised hearts with the carnitine paimitoyban&ase 1 inhrnitor, Ebxomir. Mol Ceü Biodicm 1989; 88: 175-179
Jungas RL: Effect of insulin on fatty acid synthesis h m pyruvate, hctate or aidogenous sounw in adipose tissue: ewidmce for the hormonal ngulation of pyruvate dehydrogemse. Endocn'mlogy 1970; 86: 13-1375
Denton RM, Coore HG, Martin BR, Randle PJ: Insulin activates pynmire dehydrogenase in rat epididymal adipose tissue. N ~ u e 1971; 231: 113-116
Wieland OH, Ummov T, Drexler P: Insulin, phospholipase and the activafion of the pyruvate dehydrogenase complex: an enigma. AM NY A d Sci 1989; 573: 274-284
Denton RM, Midgley P M , Rutter GA, Thomas AP, McConnack JG: Studies into the mechanism by which insulin activates pynivate dehydrogenase in adipose tissue. AM MAC& SCi 1989; 573: 285-296
Marchington DR, Kerbey AL, Randle PJ: Longer tem regulation of pyruvate dehydroge~se kinase in cul- rat cardiac rnyocyfes. Biochem J 1990; 267: 245-247
Mexante F, Rao V, Weisel RD, et al: Myocardial aerobic metaolism is irnpaued in cyanotic heart disease. Circulation 1996; 94: 1480
Hughes WA, Brownsey RW, Denton RM: Studies on the incofporation of 32-phosphate into pyruvate dehydrogenase in intact rat fàt cells. Effect of insulin. Biodien J 1980; 192: 469-481
Benelli C, Caron M, de Galle 33, Fouque F, Cherqui G, Clot J-P: Evidmce for a role of protein kinase C in the activation of the pyruvafe dehydrogenase complex by insuiin in Zajdela hepamma œiis. MetaboliSm 1994; 43(8): 1030-1034
Gottschalk WK: The pa@way mediating insulin's eff- on pyruvate dehydmgenase bypasses the insulin receptor tyrosine kinase. J Bio2 C9zem 1991 ; 266: 8814-8819

Macaulay SL, Latkin RG: Phoirphoiipese C mimi- insulin action on pynivate dehyclrogemase and msulin mxhhr g d m by mt giucose transport or utüizatim. Ce- Signuüing 1990; 2: 9-19
Saltiel ARt Insulin generates an enzyme modulata from hepatic plasma membrane: regdation of denosine 3'J' manophosphate dkamwe, pynrvate dehydrogeriase and adenylate cyclase. Endomùurbgy 1981; 120: % 7 I n
Wtiel AR, Cuiream P: In search of a second messenger for insulin. Am J Physol 1988; 255: Cl-ClO
Saitiel AR: Insuh stimuiate!û diacyIglycer01 production d t s h m the hydroiysis of a novel phosphatidyl inosito1 glycan. J BioI Qian 198x 262: 11 1 6 1 121
Jamü L, Seah JR: Pyruvate dehydrogenase activaîion in adipocyte mitochondria by an insulin-generated mediator k m muscle. Science 1979; 206: 140'7-1408
Jarett L, Wong EHA, Macaulay SL, Smith JA: Insului mediators h m rat Metaï muscle have ciiffixent e f f i on insulin-sensitive pathways of intact adipocytes. Science 1985; 227: 533-535
Kasuga M, Karisscm FA, Kahn CR: Insulin stimuiates phosphorylat.011 of the 95000 dalton subunit of its own receptor. Wetlce 1982; 215: 185-187
Kasuga M. Fujita-Yamaguchi Y, Blithe DL, Kahn CR: Tyrosine speçinc protein kinase activity is associated with the p u i f i d insulin receptor. Proc Nul Acad Sci USA 1983; 80: 2137-2141
Ishinika T, Cooper DR, Farese RV: Insulin stimulates the translocation of protein kinase C in rat adipocytes. FPCF Letz 1989; 257: 337.340
Ishia>lca T, Hoffman J, Cooper DR, Watson J, P u s W D, Farese RV: Glucose-induced synthesis of diacylglyceml de novo 0 is with translocafion (activafiion) of protein kinase C. FEBS Len 1989; 249: 234-238
Rinaudo MT, Curto M, Bruno R, Marino C, Rosseüi V, Mostert M: Evidence of an insuiin genefafed ppvate dehydrogenase stirnuiating factor in rat brain plasna membranes. IN J Biochem 1987; 19: 909-913
Standaert ML, Musunuru K, Yarnada K, Cooper DR, Farese RV: Insulin stimuIaîed phosphatidylcholine h ydroly sis, diacy1glyceroYpmtein kinase C signalhg and hexose tansport in pertussis toxin-treated BC3H-1 myocytes. ~Ztular Signahg 1994; 6: 707-716
Craven PA, DeRubertis FR: Protein kinase C is activated in glomauli fm streptomtocin diabetic rats. J CIuz I n w t 1989; 83: 1667- 1675

Klîp A, R a W T: Rotein kinase C is net reqairtd for insulin stimuiation of hexose uptake in muscle tells in culture Bïuchem J 1987, 242: 131-136
Stumpo DJ, Blackshear PJ: h s u h and growth factot effécts on c-fos exprcssim in normal and protgn kinase Adicient 3T3-L1 fiibrobfasts and adipocytes. Prac N d A d Si3 USA 1986, 83: 9453-9457
Romero G, Larner J: Insulin mediators and the mechanism of insului action. Am, P h 1993; 24: 21-50
Sutherland EW: Studies on the mechaniSm of hormone action. SccenCe 1972; 177: 4 0 1 4 8
Banting FG, Best CH, CoEp JB, Campaii WR, Fletcher AA: Pancreatic extr;tcts in the treatment of dhbetes meIIitus. &z M d ASOC J 1922; 12: 141-146
Brown H, Sanger F, Kitai R: The stnictun of pig and sheep insuluis. BiOchem J 1955; 60: 556565
Katsoyannis PG, Tometsko A, Fuhida K: Insulin peptides K: The synthesis of the A-chain of insuiin and its combination with natural Bchain to generate insulin acfivity. I Am Chan Soc 1963; 85: 2863-2865
Myers MG, White MF: The new elements of insulin Sgaalling. Insuün receptor substrate-1 and proteins with SH2 domains. DWefes 1993; 42: 643-650
Sun XJ, Rothenberg P, Kahn CR, et al: Stnicture of the insuiin receptor subsoate IRS-1 defines a unique signal transduction protein. NMue 1991 ; 352: 73-77
White MF, Kahn CR: The insulin signailhg system. J Bi01 Chem 1994; 269(1): 1-4
Streb H, IMne RF, Berridge MJ, Shuiz 1: Release of Ca2+ h m a non-mitochondrial intraiceuuiar store in pancreatic achar cells by inositol 1,4,5-triphosphate. Nmre 1983; 306: 67-69
Takai Y, Kishimoto A, Kikkawa U, Mori T, Nxshinilra Y: Unsaturated diacylglycet01 as a possible messenger for the activation of calcium-activated, phosphoiipiddependent protein kinase system. BiOchem Biophys Res Clollzmul~ 1979; 91: 1218-1224
Takai Y, Kishimoto A, houe M, Nishipika Y: Studies on a cyclic nucleotide-independent protM kinase and its prwnyme in rnammalian tissue 1: Purifîcation and characterization of an active enzyme h m bovine cefebellum. J Bid - 1977; 252: 7603-7609

NrshiniLa Y: Studies and paspsctives of pIogin kinase C. Mence 1986; 233: 305-3 12
r i s i Y: The molecular hetemgeneity of protein kiriase C and implications for cellular regulation. N4nuc 1988; 334: 661-665
Nhhbka Y: Intracellular signaihg by hydrolysis of phospholipids and activation of pmteh kinase C. Sciaicc 1992; a8: 607414
Kraft AS, Andersan WB: P m 1 esten imease the amount of Ca2+, pimphoiipid-depaident protein icinase associatEd with plasma membrane. Niina 1983; 3û1: 621-623
Mochly-Rosen D, Khaner H, IapQ J, Smith BL: Intracelllular receptor for activaîed protein kinase C. J Bi02 C%em 1992; 266: 1486614868
Hug H, Sarre TF: Protein kinase C isoaizymes: divergence in signal transduction? &'ochem J 1993; 291: 329-343
Bogoyevitch MA, Parker PJ, Sugdm PH: Characterization of protein kinase C isotype expression in adult nit hem: Protein Linase C-E is a major isotype present and it is activated by phorbol esters, CpinephRne and aidothelin. Ch Res 1993; 72: 757-767
Ono Y, Fujii T, ûgita K, Kikkawa U, Igarashi K, Ni- Y: The structure, expression and propaties of additionai members of the protein kinase C family. J Biol Chem 1988; 263: 6927-2932
Mitchell MB, Meng X, Ao L, Brown JM, Harken AH, Banerjee A: Preconditionbg of isolateci rat heart is mediated by protein kinase C. arc Res 1995; 76: 73-8 1
Rehring TF. Fnese RS, Cleveland JC, et al: a-Adrenergic preseNafion of myocardial. pH during ischemia is PKC isoform dependent. J Surg Res 1996; 63: 324-327
Castagna M, Takai Y, Kaibuchi K, Sano K, Kikkawa U, Nishinika Y: Dkct activation of calcium-activafed, phospholipid- dependent protein kinase by tumour promoting phorbol esters. J Biol Chcm 1982; 257: 7847-7851
Stumpo DJ, Haupt DM, Blackshear PJ: Proteui kinase C isozyme distribution and d m - m o n in relation to insulin-stimulatecl c-fos induction. J Biol Qlem 1994; 269(33): 21 18621 190
Davis P, HïU C, Keech E, et al: Potent seiective inhibitors of protein kinase C. EEBS Lez? 1989; 259: 61-63
Herbert JM, Augereau JM, Maaand JP: Chelerythrhe is a potent and spedfîc inhibitor of protein kinase C. Biochmi Biophys Ra Commwz 1990; 172: 993-999

Liu Y, Ytrehus K, Downey M. Evidence that W w a t i o n of protein kinase C is a key mnt during ischemic pmccmditioning of rabbit myocardium. J Mol Cell Cardiof 1994; 26: 661-668
gobayashi E, Nakano H, Moriaato M, TanisinLi T: Calphosth C (UCN-1028C), a nwei miCrobial compound, is a highiy potent and Jpscific inhibitor of protein kinase C. Biochan BU,phys Res Canmun 1991; 176: 288293
BIWU RH, Miller FD, Memman RL, et ai: Inhiaition of protein kinase C by calphosth C is îight dependent. Biodunr Biophys & Ra 1991; 176: 28û-293
Murry CE, Jennings RB, Reimer KA: Reumditioning with ischemia: a delay of lethai œil injury in ischemic myocardium. Ci-on 1986; 74(5): 1124-1 136
ILDnomidis JS, Tumiati LC, Wasel RD, Mickie DAG, Li R-K: Preconditioning human ventricular cardiomyocytes with brief periods of simulaîe!d ischemia. C ~ ~ ~ ~ O Y ~ Q S C Res 1994; 28: 1285-1291
Kloner RA, Yelion DM: Does ischemic preconditioning amu in patients? J Amer COU Cordd 1994; 24: 1133-1 142
Y d o n DM, Alkhulaïfî AM, Pugsley WB: Preconditioning the human myocardium. LMcer 1993; 342: 276277
Abd-ElfiWah AS, Ding M, Wechder AS: Intemitterit aortic crossclamphg pments cumulative adenosine triphosphate depletion, venüicuiar fibrillation and dysfunction (stunning): is it preconditioning? J =rtx Cordiowc Surg 1995; 110: 328-329
Weisel RD, Hoy FBY, Baird RJ, et al: Improved myocardial protection during a pr01onged crossclamp period. Arm Thortx Surg 1983; 36: 664-674
Banejee A: Could the "dW preconditioning mechanwn please stand up? (or, a polyplanic platitude to preconditioning). Cmdiovasc Res 1994; 28: 1872- 1873
Cleveland JC, MeIdrum DR, Rowland RT, Sheridan BC, Banerjee A, Harken AH: The obligate mie of protein kinase C in mediating clinically d b l e c a d k preconditioning. Surgery 1996; 120: 345-352
Ikonomidis JS, Shirai T, WeiseI RD, et ai: Preconditioning cuitured human pediatric myocytes requires adenosine and protein kinase C. Am J Ahysiol 1997; 272: H1220-Hl230
Liu GS, Ganote CE, Downey IM: Activafion of adenosine receptors is nquired for protection fkom a direct opener of KATP channels in rabbit ventncular cardiomyocytes. CitCJLUion 1994; 90(suppI I): 1478

Liu GS, Thmton J, Van Winlde DM, et ai: Protedion egainst infarction affozded by pmmnditicming is mediatcd by Al receptors in rabbit heart. CTrcJatiOn 1991; 84: 350.356
Miyamae M, Fujiwara H, Eida M, et al: Precanditioning impmves enagy metaboliSm during qexfision but does not attenuate myocardial stmhg in porcine hearts. C~ICU&Z&II 1993; 88: 223234
Ovbe M, Ryzlderik K, Haie SL, Kloner RA: Recmditioning does not attenuate myocardial shmning. Ci- 1992; 85: 2247-2254
Caims CB, Wang J, Walther JM, Harken AH, Banerjee A: Ischemic precunditioning prevents cytochmme a43 induced cellular oxidation during cardiac ischeniia Circirhrion 19%; 94(8): 1-362
Van Wylm D: méct of ischemic preccmditioning on interstitial purine rnelzbolite and lactate accumulation during myocardial ischemia. CCrCtrlOtion 1994; 89: 2283-2289
Wolfe CL, Sievers RE, Vissaai FL, DomeJIy TT: Loss of myocardial protection after pfec~ndîtioning correlates with the tirne course of glycogen recovery within the preconditioneû segment. Circulan'on 1993; 87: 881-892
Lagerstrom CF, W a k r WE, Taegtmeyer H: Failure of giycogen depIeticm to improve lefi ventricular function of the rabbit heart after hypothamic ischernic arrest Cin: R ~ s 1988; 63: 81-86
Coore HG, Denton RM, Martin BR, Randle PJ: Reguiation of adipose tissue pyruvate dehydrogemse by insulin and otiier hormones. &'ochem J 1971; 125: 115-127
Mukherjee C, Jungas RL: Activation of pyruvate dehydrogenase in adipose tissue by insului. Evidence for an eff- of insulin on pyruvate dehydrogenase phosphatase. BiOchem J 1975; 148: 229-235
Saltiei AR, Fox JA, Sherline P, Cuatrecasas P: Insulin stimulates the release h m liver plasma membranes of a chexnical moddaîm of pyruvate dehydrogenase. Science 1986; 233: 967-972
Assimacopouios-Jeannet F, McCofmack JG, Prentki M, Jeanrenaud B, Denton RM: Paralle1 increases in rates of faay acid synthesis and in pynivate dehydrogemse acfivity in isolated rat hepatocytes i n c u W with insuiin. Bi& Biophys A m 1982; 717: 86-90
Li R-K, Mickle DAG, Weisel RD, et al: Human pediatric and adult ventncular cardiomyocytes in culture.; assessrnent of phenotypic changes with passaging. C ~ ~ O V ~ U C R~s 1996; 32: 362-373

Li R-K, Shaikh N, Weisel RD, W- WG, Mickle DAG: ûxyradical induced antioxidant and lipid changes in cuitwed human cardiomyacytes, Am J PhysioZ 1994; 266= H22U4-2211
Li R-K, Wasel RD, W m WG, Mickie DAG: Method of cuîbiring cardiomyocytes fnnn human pediatric ventricular myocardium. J ZEs aclr M& 1992; 14: 93-100
Mickie DAG, Li R-K, Weisel RD, Tumiati LC, W u T-W: Water soluble antioxidant @City against fke radical injury usiag dtured human venîricular myocytes and fï'broblasts and saphmous vein endotbeliat ceîis. J Mol Ccll Coniiol 11990; 22: 1297- 1304
'Iiimiati LC, Mickle DAG, Weisel RD, Williams WG, Li R-K: In vitro maiel to sûdy myocardial ischemic injury. J llss CWr M d 1994; 16: 1-9
Robinson BH, McKay N, Goodyer P, Lancaster G: Defective intramitochondrial NADH oxidation in skin fibroblasts fiom an infant with fatal neonatal lacticacidemia. Am J Hamt GenmLcs 1985; 37: 938-946
Sheu ICF, Hu CW, Utter MF: Pynivate dehydrogenase cornplex activity in nomial and deficient fibroblasts. J Clin I m t 198 1; 67: 1463-1471
Bradford MM, Anversa P, Fitzpatrick D, Argani S, Capasso JM: A rapid and sensitive method for the quantitation of rnimgram quantities of protein utilizing the principie of protein- dye binding Myocyte mitotic division in the aging mammalian rat heart. AnaI Biocfrem 1991; 69: 1159-1164
Hull-Ryde EA, Lewis WR, Veronee CD, Lowe JE: Simple step gradient elution of the major high-energy compounds and their metabolites in cardiac muscle using high- paf~nnance liquid chromatography. 3 Qlm-ogr 1986; 377: 165-174
Burton K: A study of the cunditions and the mechanisms of the diphenylamine feaction for the wlorimetric estimation of deoxyribonuc1eic acid. Biochem 3 1956; 62: 315-323
Blackshear PJ, Haupt DM, Shimpo DJ: Insulin activation of protein kinase C: A reassessrnent. J B i d Qrem 1991; 266: 1094M0952
hœqi S, Baldini P, Bducci V, 22i~eüi A, M y P: Modulation of the Na-H antiport by insulin: Interplay between protein kinase C, tyrosine kinase and protein phosphatases. J Cell Physiol 1994; 159: 2051212
Inoguchi Ty Xia P, K m b k i M, Higashi S, Feener EP, King GL: Insulin's effect on protein kinase C and dhcylgiycerol induced by diabetes and giucose in vascuiar tissues. Am J Physul 1994; 267: Eî69-379

Isbiaika T, Yamamoto M, Kajita K, et ai: Insuün stimuiates novei protein kinase C in rat adipocytes. Iliochem and Biophys Rcs Comnr 1992; 183(2): 814420
Sampson SR, Brudie C, dboim SV: Role of p t & kinase C in insulin activati,cm of the Na-K pump in dtured sLelegl muscle. Am I PVrysioI 1994; 266: C751€758
Wang JL,, Corbett JA, Marshall CA, M M ML. Glucose-induced insulin d o n h m p&ed Bcells: A role fin moditlaticm of Ca2+ M u x by CAMP and pmtan kinase Cdepeadent signal transduction pathways. J Biol Cha 1993; 268: 7785-779 1
Yu B, Standaert M, Amdd T, et al: Eff' of insulin on diacylgIyceroVprotein kinase C signaiiing and glucose transport in rat skeletal muscles in vivo and in vitro. Endoctr'nology 1992; 130: 3345-3355
Lefa AM, Tsao PS, Lefer DJ, Ma X: Role of endotheliai dysfiuiction in the pathogenesis of rrpemison injury a f k rnyocardial ischemia. FASEB JoMiol1991; 5: 2029-2034
Lefa DJ, Nalouiishi K. Johnston WE, Vinten-Johansen J: Antinwtrophil and myocardial protection actions of a novel nitric oxide donor afkr acute myocardial ischemia and repemision in dogs. on 1993; 88: 2337-2350
Mer DJ, NakaniShi K, Vinten-Johansen J: Endothelial and myocardial celï protection by a cysteine-containing nitric oxide donm after myocardial ischemia and reperfusim. J Cordovmc F ~ Z 1993; TZ(suppI7): SWS43
Li R-K. S W N, Weisel RD, Tumiati LC, W u T-W, Mickle DAG: Cuitured vascular endothelial c d suœptibility to extracellularly generated oxidant injriry. I Mol Ceü Crudiol 1992; 24: 595604
Nalranishi K, Piao ZQ, Vintm-Johansen J, et al: Blood cardioplegia enhanced with nitric oxide donor SPM-5185 counteracts popostischernic endothehi and ventricular dysfwrction. J niorac C"nnowc Surg 1995; lW(6): 1146-1 154
N M s h i K, Zhao ZQ, Vinten-Johansai J, Lewis JC, McGec DS, Hammon JW: Coronary artery endothelial dysfunction aRer ischemia, blood cardiopiegia and repemision. han Thorcu: Surg 1994; 58: 191-199
Richard V, Kaeffa N, Tron C, ThuiIlez C: Ischemic preçonditioning protects against coronafy endothelial dysfhction induced by ischemia and repemision. C i m o n 1994; 89: 1254-1261
Shirai T, Rao V, ~eisel*RD, Ikonomidis JS, et ai: Preconditioning human ventrîcular d o m y o c y t e s and endothelial cells. J Thoroc Crudiovrnc Surg 1997; In Press.

242. Tsao P, Aoki N, Lefer DJ, et al: Time course of endothelial dysfuncticm and myocardial injury during mya?udial ischemia and repafusion in the cat. CïtCU](ZtiOlé 1990; 82: 1402-1412
243. Palmer RMJ, Femige AG, Moncada S: Nihic oxide reïease accounts for biologic activity of endothekm derived relaxing factor. Nmwe 1987; 327: 525-526
244. Finkel MS, Oddis CV, Jacob TD, Walliiis SC, Hattler BG, Simmons RL: Negative inotmpic & i of cytokines on the heart mediated by nitric oxide. Science 1992; 257: 387-389
245. Kelly RA, Fkllingand JL, Smith TW: Nitric oxide and cardiac fiuicticm. Cnr Res 1996; 79: 363-380
246. Rao V, Feindel CM, W&l RD, Boylen P, Cohen G: Donor blood pemiSim improves myocardial recovefy folXowing cardiac transplantation. J H e a ~ LMg T~QILI~P- 1997; 16: 667-673
247. Moule SK, Denton RM. Multiple signaihg pathways involvedin the metabolic effm of insulin. Am I Cardo1 1997; 80: 41A49A
248. Lopaschuk GD. Alterations in fatty acid oxidation during repafuson of the heart after myocardial ischemia. Am J Cordi01 1997; 80: 1 lA-16A
249. Portman MA, Panos AL, Xia0 Y, et al. Influence of p H of cardiopiegic solutions on cellular energy metabolism and hydrogen ion flux during neonatal hypothermie circuiatory arrest and reperfixsion: a dynamic 31P nuclear magnetic resunance study in a pig model. J Tbrac Cardovasc Surg 1997; 114: 601-608
250. Taegtmeyer H, Gdwin GW, Doenst, Frazier OH. Substrate metabolkm as a determinant for postischemic fwictional recovery of the heart. Am J Canüol1997; 80: 3A-10A
251. Chnstakis GT, Weisel RD, Mickle DAG, et ai: Right venfricular function and metabolism. CircxdWon 1990; 82(suppl IV): lV332-340
252. Ikonomidis JS, Yau TM, Weisel RD, et al: Optimal flow rates for retrograde warm cadioplegia. J %rat Gndiov~sc Surg 1994; 107: 510-5 19
253. Teoh IM, Mickle DAG, Weisel RD, et al: Improving myocardial metabolic and funciional recovery after dopIegic arrest. J %rut &rdiovasc Swg 1988; 95: 788-798
254. Spahi R, Jacobsen SL, Skgmund B, Schwartz P, Piper HM: Substrate oxidaîion by adult cardiomyocytes in long-term primaxy culture. J Mol CeU &rdioZ 1989; 21: 175-185

Jacobsen SL, Pipa HM: Ceil cultures of aduit cardiomyocytes as models of the myocardium. J Mol CkZI CMüol1986; 18: 661-678
Marino TA, mdar S, W i s c m EC, et al: Pro1Xferating c d nuclear antigen in developing and addt rat cardiac muscle dis. C h Res 1991; 69: 1353-1360
Cowan DB, Weisel RD, Williams WG, Mckle DAG: The regdation of glutathme peroxiW gaie expression by oxygen tension in cultureci human cardiomyocytes. J Md CeU Conid 1992; 24: 423-433
Cowan DB, Weisel RD, Williams WG, Mickîe DAG: Identification of oxygen responsive elements in the S1-fiankhg region of the human glutathione peroxidase gaie. J Bi02 Chem 1993; 268: 26904-26910
v, ChristaLis GT, Weisel RD, et al: Does the internai thoracic artery graft delay the recovery of myocardial rneîabolism? Ann Thorm Surg 1996; 62: 1039-1044
Tagarni S, Kondo T, Yoshida K, Hirokawa J, Ohtsuka Y, Kawalmi Y: Enect of insulin on impaired antioxidant activities in aortic endothelial celis from diabetic rabbi&. Metaoollhmr 1992; 41(10): 1053-1058
Maroko PR, Libby P, Sobel BE, et al: Wect of glucose-insulin-potassium infusion on myocardia hfhrction following experimental coronary artery occlusion. Ckuhion 1972; 45: 116M173
Brodin LA, Dahlgren G, Ekestrom S, Sertergren G, Ohqvist G: Inauence of glucose-insulin-potassium on Ieft ventricuiar fundion duMg coronary artery bypass grafting. Sc& J 3hom Cardiovarc Surg 1993; 27: 27-34
McDaniel HG, Papapietro SE, Rogers WJ, et al: Glucose-insulin-potassium induced alterations in individual plasma kee fâîty acids in patients with acute myocardial infàrction. Am Heart J 1981; 102: 10-15
Rogers WJy Russell ROY McDaniel HG, Rackley CE: Acute effm of glucose-insulin-potassium infusion on myocardial substrates, coronary blood flow and oxygen consumption in man. Amer J Card 1977; 40: 421-427
Svedjeholm R, Hakanson E, Vanhanen 1: Rationale for metaboiic support with amino acids and glucose-insulin-potassium (GM) in cardiac surgery. ANI ThDrac Swg 1995; 59: SIS-S22
Svensson S, Baglin E, Ekmth R, Milocco 1, Nilsson F, WiUiam Olsson G: Haemodynamic effects of a single largedose of insulin in open heart surgery. Cardiovasc R a 1984; 18: 697-701

Sva1sson S, Svedjeholm R, Ehath R, et al: Tauma metabolism and the h m Uptake of substrabes and effécts of insulin e d y af?er cardiac opaations. J %MC Cbrdiovusc Surg 1990; 99: 1063-1073
Berti L, Mosthaf L, Kroder G, et al: Glucose induced translocation of protein kiriase C isoforms in rat-1 fibroblasts is paralleed by inhiiition of the insuiin recepfar tyrosine kinase. J Bwl Chem 1994; 269(5): 3381-3386
Cheng K, Lamer J: Intracdular mediatm of insulin adon. AM RA1 Physi021985; 47: 405-424
Facese RV, Standaezt ML, Francois AJ, et al: EffecfS of insulin and phorbol esters on subceiiular dimiution of protein idnase C isofonns in rat adipocytes. Baihem J 1992; 288(1): 3 19-323
Romen, G, Luttreii L, Rogol A, Zeller K, Hewlett E, Lanier J: Phosphatidylinosiîol-glycan anchors of membrane proteins: Potenhi pracursors of Insuiin mediators. Scietule 1988; 240: 509-51 1
Russ M, Ecke1 J: Insulin action on cardiac glucose transport: studies on the role of protein kinase C. Bi& et Biophys Acta 1995; 1265(1): 73-78
Sun DQ, Nguyen N, DeGrado TR, Schwaiger M, Brosius FC: Ischemia induces translocation of the insulin-responsive glucose tansporter GLUT4 to the plasma membrane of carciïac rnyocytes. Circuian'on 1994; 89: 793-798
Wheeler TJ: Translocation of glucose transporteres in response to anoxh in kart. J Biol Ckm 1988; 263(36): 19447-19454
Erikson TM, Velasco CE: Endothelin-l and rnyocardial pfeconditioning. Am HCMJ 1996; 132: 84-90
Li Y, Kloner RA: Cardioprotective effects of ischemic preconditioning oui be recaptured afta they are lost. J Am CoU ~ i 0 2 1994; 23: 470474
Teague WM, Pettit F, W u T-L, Srlbennan SR, Reed LJ: Purification and of pynivate dehydrogenase phosphatase h m bovine heart and kidney. Biocttemishy 1982; 21: 5585-5592
Nichols RJ, Denton RM: Towards the molecuiar basis for the reguiation of mitochondrial dehydmgenases by calaum ions. Mol Cell BiochQn 1995; 150: 203-212
Okuda K, Nohara R, Fujita M, Tardi N, Konishi J, Sasayama S: Improvement of myocardial ischemic dysfunction with d i c h l o d c acid: Experïmental study by repeated ischemia in dogs. J Cmdovasc Phm 1995; 26: 990-999

McConiiack JG, Barr RL, Wolff M, Lopaschuk GD: Ranolazine stimulates giucœe oxidation in normoxic, ischemic and repdiwd ischemic rat hearts. CZ'niulation 19%; 93: 135-142
Renstrom B. Nellis S, Liedtke Al: Me&bolic oxïdation of glucose during early myocardial repenuSon. Cire Rcr 1989; 65: 1094-1101
Stanley WC, Hemandez LA, Spires D, B ~ g a s J, Wallace S, McCormack JG: PyniMte debydrogenase d v i t y and malcmyl CoA levels in n o d and ischemic myocardium: Effkcts of dichlomœak. J Mol Ccll Crudioz 19%; 28: 905-914
Knuuti MJ, M M M, Yki-J&m, et al: The effm of insuiin and FFA on myaardial glucose upîake. J Md Cell Clrudioz 1995; 27: 135% 1367
Lazdunski M, Frelin C, Vigne P: The sodium-hydrogen exchange system in cardiai: cells: its biochemical and pharmacological properties and its d e in regulating intenial concentrations of sodium and intanal pH. J Mol C d CkdioI 1985; 17: 1 0 4 2
Alto LE, Dhalla NS: Myocarûiai cation contents d u ~ g induction of calcium paradox. Am J Physiol 1979; 237: H713-H719
Ikonomidis JS, Salerno TA, Wiünich C: Calcium and the heart: An essential partnership. CM 1 W o Z 1990; 60: 305-316
Mitchell MB, Winter CB, Banqee A, Harken AH: Inhibition of sarcoplasmic reticulurn calcium release reduces myocardiai stunning. J Surg Res 1993; 54: 41 1-417
Zimmerman ANE, Daems W, Hulsmann WC, Snijder J, Wisse E, Durrer R: Morphologid changes in heart muscle caused by successive perfusion with calcium-fxee and calcium-containing solutions (Catcium Paradox). Cardovusc Rcr 1967; 1: 201-209
Miura T, Ogawa T, Suzuki K, Goto M, Shimamoto K: Infant size limitation by a new Na+-H + exchange inhiitor, HOE 642: difference h m prrconditioning in the role of protein kinase C. JACC 1997; 29: 693-701
Myers ML, Karmayn M: Impmved cardiac function a f k prolonged hypothermie ischernia with the Na+/H+ exchange inhibitor HOE 694. ANI Zbruc Surg 1996; 61: 1400-1406
Tani M, Neely JR: Role of intracellular Na+ in CaS' overload and depfessed m v e r y of ventxiculaf function of repemised ischernic rat hearts. Possible involvement of Na+-H+ and Na+-Ca" exchange. CCrc Res 1989; 65: 1045-1056
Karmaqm M: Amiloride enhances postischemic ventncular recovery: possible role of Na-H exchange. Am 3 PhysoZ 1988; 255: H608-H615

293. Mertens S, Noii T, Spahr R, Krutâeldt A, Piper KM: Energetic response of coronary aidothelisl tells tû hypoxia, Anr J PhysioI1990; 258: H6899H694
294. Knrt;tfeldt A, Spahr R, Mertens S, Siegmund B, Pipa HM: Me&hIism of exogewus substrates by coronary endothelid cells in cuiture. J Mol Cen Cardi01 lm, 22: 1393-1404
295. Crabtree HG: Observation of the carbohydrate metaboliSm of tumours. Biochem J 1929; 23: 536-545
296. Schemr U, Randin D, Volienweider P, VoUenweider L, NiCod P: Nitric oxide release accounts for insulin's vascuiar effècts in hurnans. J Cün Zn= 1994; 9 4 251 1-2515

Appnâix 1: Extraccllulu Lactate and Ppvete Muclion
STABJWZATION Lacdnte (inoUmin/g)
&mate (moUnrln/g) Laclole/4)mwte Rudo
sxhocell&r pH PDH adv& (lvllourng)
ISCHEMIA hcdate (nwUmIn/g)
&mute (moUmtnlg) t O c ~ e / 9 ( ~ ~ u u i l ~ Ratio
&tmceUulor pH P M adv@ (moumg) +0.62 (.04)
REPBRFUSION tacdPh (nioVmldg) + 1.3 (0.4)
&m~ate (moUmln/g) +41 (6) & c ~ e / ~ m v o t e Ratio + 32 (5)
ajetmceluar pH 7.24 (0.04) PDH activity (nmol/mg) 1 + 1.01 (0.11)
Mean (SEM)
1.8 (0.2) 30 (6) 63 (6)
7.19 (0.05) m.96 (O. 13)
e1.2 (0.2) m 9 (2)
65 (9) 7.21 (0.04) + @+1.26 (O. 12)
CONTROL
0.4 (O. 1) 61 (5) 9 (1)
7.27 (0.03)
ontm ml plab r<sas#l atbudine (slcrbl&t&n)or iTta W ( i ~ ~ a ~ m t a ) or 150 (mperf'on) minutes of incubation in nonnoxic phoephate butked dine.

- - Oh^^^^ , P P P P r N N N N P O V w w w w w

IMAGE EVALUATION TEST TARGET (QA-3)
APPLIED 1 IMAGE. lnc 1653 East Main Street - -. - Rochester. NY 14609 USA -- -- - - Phone: 716/482-0300 -- -- - - Far: 716/208-5989
O 1993. Appüed image. Inc. All RWts Resefved