chapter 10 - post-transcriptional gene control
TRANSCRIPT

Chapter 10 - Post-transcriptional Gene Control

Chapter 10 - Post-transcriptional Gene Control
10.1 Processing of Eukaryotic Pre-mRNA
10.2 Regulation of Pre-mRNA Processing
10.3 Transport of mRNA Across the Nuclear Envelope
10.4 Cytoplasmic Mechanisms of Post-transcriptional Control

Post-transcriptional Gene Control
10.1 Processing of Eukaryotic Pre-mRNA
• Pre-mRNA is capped, polyadenylated, spliced, and associated with RNPs in the nucleus before export to the cytoplasm.
ribonucleoprotein

Overview of RNA processing and post-transcriptional gene control.
• Step 1 – Use of alternative exons during pre-mRNA splicing• Step 2 – Use of alternative poly(A) sites. • Step 3 –
• Properly processed mRNAs – exported to the cytoplasm
• Improperly processed mRNAs –• blocked from export to the cytoplasm• degraded the exosome complex containing
multiple ribonucleases• Step 4 – Translation initiation factors – bind to the 5′ cap
cooperatively with poly(A)-binding protein I bound to the poly(A) tail and initiate translation
• Step 5 – mRNA degraded in cytoplasmic P bodies –translational repression
• Deadenylated and decapped by enzymes• Degraded by cytoplasmic exosomes• Control of mRNA degradation rate – regulates mRNA
abundance and amount of protein translated• Step 6 – mRNAs synthesized without long poly(A) tails –
translation regulated by controlled synthesis of a long poly(A) tail by a cytoplasmic poly(A) polymerase
• Step 7 – Translation regulation by other mechanisms –• miRNA (~22-nucleotide RNAs) – inhibit translation of
mRNAs to which they hybridize, usually in the 3′ untranslated region
• Step 8a – tRNAs – transcribed by Pol III and processed in the nucleus
• Step 8b – rRNAs – transcribed by Pol I – processed in the nucleolus
• Step 9 – Regions of precursors cleaved from the mature RNAs – degraded by nuclear exosomes

• Step 1: • Nascent RNA (β-globin RNA) 5′ end capped with 7-methylguanylate (shortly after RNA polymerase II initiates
transcription at the first nucleotide of the first exon of a gene)• Pol II transcription – terminates at any one of multiple termination sites downstream from the poly(A) site in
final exon • Step 2: Cleavage enzyme cleaves primary transcript at the poly(A) site.• Step 3: Polyadenylation enzyme adds a string of adenosine (A) residues (~250 A residues in mammals, ~150 in
insects, and ~100 in yeast). • Step 4:
• Short primary transcripts with few introns – splicing follows cleavage and polyadenylation (shown)• Long transcripts with multiple introns – introns spliced out of the nascent RNA during transcription
Overview of mRNA processing in eukaryotes
Why cleavage?

• Cellular RNAs and precursors.

Splicing process
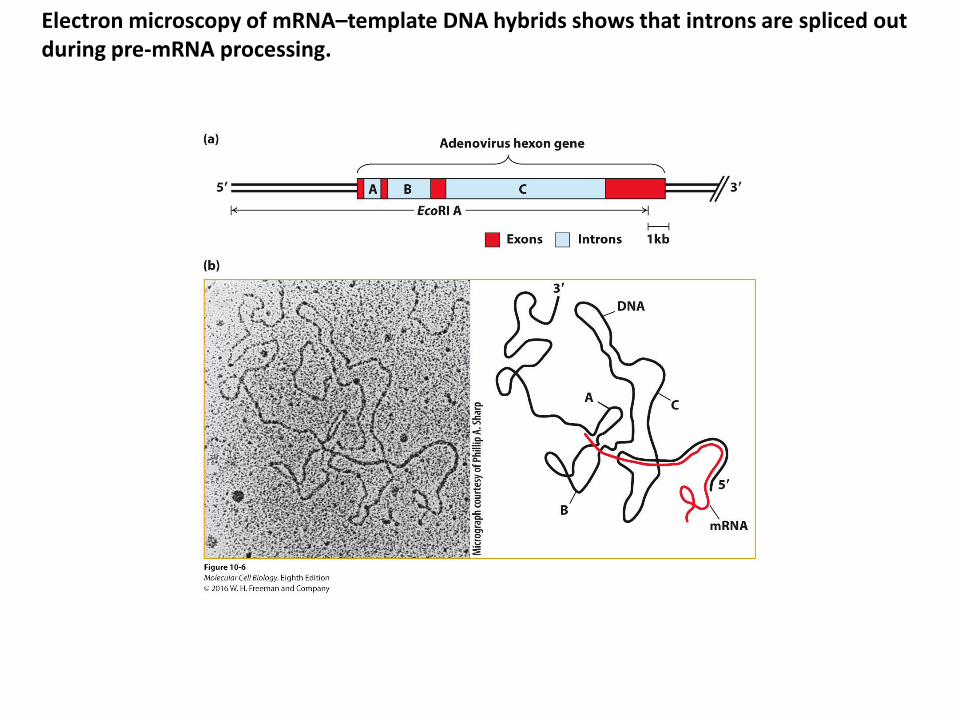
Electron microscopy of mRNA–template DNA hybrids shows that introns are spliced out during pre-mRNA processing.

Consensus sequences around splice sites in vertebrate pre-mRNAs.
• Intron splice site invariant bases (flanking bases indicated – found at frequencies higher than expected for a random distribution):
• 5′ GU• 3′ AG • Branch-point adenosine – usually 20–50 bases from the 3′ splice site
• Polypyrimidine tract near the 3′ end of the intron – found in most introns• Central region:
• 40 bases–50 kilobases• Only 30–40 nucleotides at each end of an intron are necessary for splicing to
occur at normal rates.

transesterification reactions result in the splicing of exons in pre-mRNA.
• Exon splicing – two sequential transesterification reactions [Arrows indicate where activated hydroxyl oxygens react with phosphorus atoms]:
• Reaction 1 – intron 5′ phosphorus-exon one 3′ oxygen ester bond –exchanged for an intron 5′ phosphorus ester bond with the branch-point A residue 2′ oxygen
• Reaction 2 – the exon two 5′ phosphorus – intron 3′ oxygen ester bond – exchanged for an exon two 5′ phosphorus ester bond with the 3′ oxygen of exon one
• joins the two exons• releases intron as a lariat
structure
How??
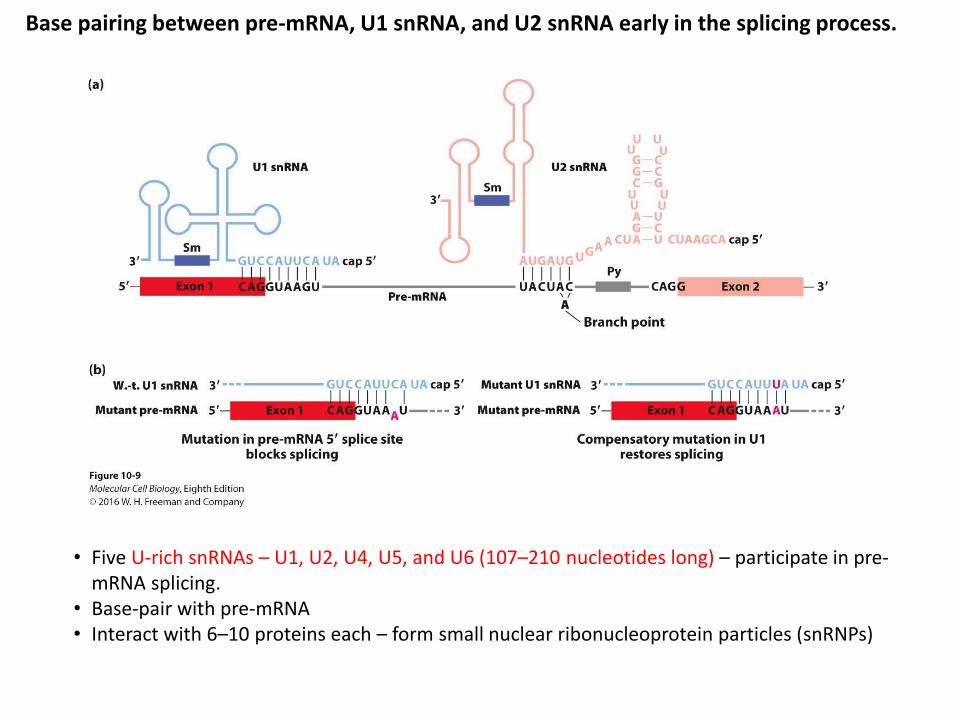
Base pairing between pre-mRNA, U1 snRNA, and U2 snRNA early in the splicing process.
• Five U-rich snRNAs – U1, U2, U4, U5, and U6 (107–210 nucleotides long) – participate in pre-mRNA splicing.
• Base-pair with pre-mRNA• Interact with 6–10 proteins each – form small nuclear ribonucleoprotein particles (snRNPs)


mRNA splicing
https://www.youtube.com/watch?v=FVuAwBGw_pQ
Two actions – cut and ligation

mRNP
• From their transcriptional birth to their degradation, cellular mRNAs are coated with proteins in messenger ribonucleoprotein (mRNP) complexes. The mRNP composition controls every aspect of the life of the mRNA, from pre-mRNA processing to mRNA localization, translation and turnover.

GU----A-----AG?How to recognize the exon and intron?

SR proteins
• SR proteins are a conserved family of proteins involved in RNA splicing. SR proteins are named because they contain a protein domain with long repeats of serine and arginine amino acid residues, whose standard abbreviations are "S" and "R" respectively.

Exon recognition through cooperative binding of SR proteins and splicing factors to pre-mRNA.
• Pre-mRNAs (humans):• Exons – avg ~150 bases• Introns – avg ~3500 bases – longest exceed
500 kb • Degenerate 5′ and 3′ splice site and branch
point sequences – multiple copies likely to occur randomly in long introns
• Additional sequence information is required to define the exons that should be spliced together in higher organism pre-mRNAs with long introns.
• SR proteins contribute to exon definition in long pre-mRNAs.
• SR proteins:• Interact with exonic enhancer sequences
(ESEs) within exons• Contain several RS protein-protein
interaction domains rich in arginine (R) and serine (S) residues
• Mediate cooperative binding of U1 snRNP to a true 5′ splice site and U2 snRNP to a branch point through a network of protein-protein interactions that span an exon

• Self-splicing introns –two types:• Group I introns – in nuclear rRNA genes of
protozoans• Group II introns –
• In protein-coding genes and some rRNA and tRNA genes in mitochondria and chloroplasts of plants and fungi
• Fold into a conserved, complex secondary structure containing numerous stem-loops –
Self-splicing introns…

Post-transcriptional Gene Control
10.2 Regulation of Pre-mRNA Processing
• Alternative promoters, primary transcript alternative splicing, and cleavage at different poly(A) site cleavages yield different mRNAs from the same gene in different cell types or at different developmental stages.
• RNA-binding proteins that bind to specific sequences near splice sites regulate alternative splicing.
• Rare RNA editing of mRNA sequences in the nucleus have important consequences by altering the amino acid encoded by an edited codon.

A cascade of regulated splicing controls sex determination in Drosophila embryos. • (a) Sex-lethal (Sxl) protein: intronic splicing silencer
• Present only in female embryos (no functional Sxl protein expressed in males)
• Binds to a premRNA sequence near the 3′ end of the intron between exon 2 and exon 3 –
• blocks U2AF and U2 snRNP association of with the adjacent 3′ splice site (used in males).
• female splicing (exon 2–4) –• deletes exon 3 (contains premature stop codon) • Sxl expressed
• male splicing (exon 2–3) –• mRNA contains exon 3 premature stop codon.• no functional Sxl protein expressed in males
• (b) Sxl blocks tra exon 1–2 splicing.• Only female embryos produce functional Tra protein.• Tra – promotes specific Dax splicing
• (c) Dsx (transcription factor) expression:• Females –
• cooperative binding of Tra protein and Rbp1 and Tra2 SR proteins activates exon 3–4 splicing and cleavage/polyadenylation(An) at the 3′ end of exon 4.
• expresses female Dsx isoform with exon 4-encoded sequence – transcription factor
• Males –• embryos lack functional Tra• SR proteins do not bind to exon 4• exon 3 is spliced to exon 5.• expresses male Dsx isoform with exon 5-encoded
sequence (transcription repressor)• Distinct female and male Dsx proteins:
• female Dsx isoform – activates genes with Dsxtranscription factor binding sites, including genes that induce development of female characteristics
• male Dsx protein – represses expression of the same target genes with Dsx binding sites
Series of different proteins are expressed!

Model of splicing activation by Tra protein and the SR proteins Rbp1 and Tra2.
One of the SR proteins

Role of alternative splicing in the perception of sounds of different frequencies.
• Channel isoforms –• encoded by alternatively spliced mRNAs
produced from the same primary transcript
• alternative exons used at eight regions in the mRNA – 576 possible isoforms [Red numbers – regions where alternative splicing produces different isoforms]
• respond to different frequencies by opening at different Ca2+ concentrations
Ca2+-activated K+ channel
Frequencies…


Overview of RNA processing and post-transcriptional gene control.
•Next ?

Post-transcriptional Gene Control
10.3 Transport of mRNA Across the Nuclear Envelope
• An mRNP exporter ensures directional export by binding mRNPs in the nucleus, facilitating transport across the NPC, and releasing the mRNPs when mRNP adapter proteins are phosphorylated in the cytoplasm.
• The mRNP exporter binds most mRNAs cooperatively with SR proteins bound to exonic splicing enhancers and with REF associated with exon-junction complexes as well as with additional mRNP proteins.
• Pre-mRNAs still bound to spliceosomes are not exported, ensuring only mature mRNAs reach the cytoplasm.

Remodeling of mRNPs during nuclear export.
• Some mRNP proteins dissociate from nuclear mRNP complexes before export through an NPC.
• Some mRNPs remain associated – CBC (cap binding complex), NXF1/NXT1, and PABPN1 bound to the poly(A) tail –
• exported with the mRNP• dissociate from the mRNP
in the cytoplasm and are shuttled back into the nucleus through an NPC
• Translation initiation factor eIF4E replaces CBC bound to the 5′ cap.
• PABPC1 replaces PABPN1.

Nuclear pore complex (NPC)
https://www.youtube.com/watch?v=UyhqLpjicZg

Nuclear export
https://www.youtube.com/watch?v=9v-13EZWVk8


Formation of heterogeneous ribonucleoprotein particles (hnRNPs) and export of mRNPs from the nucleus.

Poly-adenylation

Model for cleavage and polyadenylation of pre-mRNAs in mammalian cells.
• Mechanism:• CPSF (Cleavage and polyadenylation specificity factor) –
binds to the upstream AAUAAA polyadenylation signal • CStF interacts with a downstream GU- or U-rich
sequence and with bound CPSF, forming a loop in the RNA.
• CFI and CFII binding – stabilize the complex• PAP (Poly(A) polymerase) – stimulates cleavage at a
poly(A) cleavage site (typically 15–30 nucleotides 3′ of the upstream polyadenylation signal)
• Cleavage factors – released• Downstream RNA cleavage product – rapidly degraded• PAP – adds ~12 A residues (from ATP) at a slow rate to
the 3′-hydroxyl group generated by the cleavage reaction
• PABPN1 (Nuclear poly(A)-binding protein) –• Binds to the initial short poly(A) tail• Accelerates the rate of addition by PAP• Signals PAP to stop adding As after 200–250 A
residues have been added

Post-transcriptional Gene Control
10.4 Cytoplasmic Mechanisms of Post-transcriptional Control
• Stability of most mRNAs is controlled by poly(A) taillength and binding of various proteins to 3’ UTR sequences.
• mRNA translation can be regulated by micro-RNAs and RNA interference by siRNAs and various degradation, cytoplasmic splicing, and polyadenylation mechanisms.
• Many mRNAs are transported to specific subcellular locations by sequence-specific RNA-binding proteins that bind 3′ UTR localization sequences.

Pathways for degradation of eukaryotic mRNAs.
most common pathway
• Deadenylase complex shortens poly(A) tail to ≤20 A residues.
• Destabilizes PABPC1 binding• PABC1 loss weakens interactions between
the 5′ cap and translation initiation factors.
• Deadenylated mRNA:

P body
• Processing bodies (P-bodies) are distinct foci within the cytoplasm of the eukaryotic cell consisting of many enzymes involved in mRNA turnover. P-bodies have been observed in somatic cells originating from vertebrates and invertebrates, plants and yeast. To date, P-bodies have been demonstrated to play fundamental roles in general mRNA decay, nonsense-mediated mRNA decay, adenylate-uridylate-rich element mediated mRNA decay, and microRNA induced mRNA silencing.
1. decapping and degradation of unwanted mRNAs2. storing mRNA until needed for translation3. aiding in translational repression by miRNAs (related to siRNAs)

• (a) miRNAs:• Repress translation of target mRNAs• Hybridize imperfectly with target mRNAs • miRNA nucleotides 2–7 (seed sequence) – most critical for targeting it to a specific
mRNA 3’-UTR• (b) siRNA:
• Hybridizes perfectly with target mRNA• Causes cleavage of mRNA (position indicated by the red arrow), triggering its rapid
degradation

RISC
• The RNA-induced silencing complex, or RISC, is a multiprotein complex, specifically a ribonucleoprotein, which incorporates one strand of a single-stranded RNA (ssRNA) fragment, such as microRNA (miRNA), or double-stranded small interfering RNA (siRNA).[1] The single strand acts as a template for RISC to recognize complementary messenger RNA (mRNA) transcript. Once found, one of the proteins in RISC, called Argonaute, activates and cleaves the mRNA. This process is called RNA interference (RNAi) and it is found in many eukaryotes; it is a key process in gene silencing and defense against viral infections

Processing of miRNA
• miRNA transcription and processing:• RNA polymerase II transcribes primary miRNA transcripts (pri-
miRNA) – folds to form double strand region • Nuclear double-strand RNA–specific endoribonuclease Drosha
and double-strand RNA–binding protein DGCR8 (Pasha in Drosophila) bind pri-miRNA double strand regions
• Drosha cleaves the pri-miRNA – generates a ~70-nucleotide pre-miRNA
• Exportin 5 – nuclear transporter transports processed pri-miRNA to the cytoplasm
• Dicer in conjunction with the double-stranded RNA–binding protein TRBP (Loquacious in Drosophila) – processes pre-miRNA into a double-stranded miRNA with a two-base single-stranded 3′ end
• RISC complex –• binds one of the two strands. • incorporates mature miRNA into complex with Argonaute
proteins• mRNA translation inhibition:
• miRNA-RISC complexes associate with target mRNPs by base pairing between the Argonaute-bound mature miRNA and complementary regions in the 3′ UTRs of target mRNAs
• The more RISC complexes bound to the 3′ UTR of an mRNA, the greater the repression of translation
• RISC complex binding – causes bound mRNPs to associate with P bodies – mRNA degraded
• Alternative polyadenylation increases miRNA control options.

The function of miRNAs in limb development.
• Result – fundamental pattern maintained but abnormal limb development
• Conclusion – Dicer (miRNA) is required for morphogenesis but not patterning of the vertebrate limb

Polyadenylation and translation initiation

Model for control of cytoplasmic polyadenylation and translation initiation.
• Immature oocyte mRNAs containing the U-rich cytoplasmic polyadenylation element (CPE) initially have short poly(A) tails –translation repression:
• Hormonal stimulation of oocyte maturation:• Activates a protein kinase that phosphorylates
CPEB• Phosphorylated CPEB –
• releases Maskin• recruits CPSF (cleavage and polyadenylation
specificity factor) – binds the poly(A) site
• CPSF recruits the cytoplasmic poly(A) polymerase (PAP).
• PAP lengthens the poly(A) tail. • Longer poly(A) tail binds multiple copies of
cytoplasmic poly(A)-binding protein 1 (PABPC1).• PABC1 interacts with eIF4G – stabilizes interaction
of factors for ribosome recruitment and translation

Discussion with friends
• Find what is the nuclear localization sequence (NLS) of protein and its mechanism
• Search the Clustered regularly interspaced short palindromic repeats in the wikipedia site and explain the right figure. (https://en.wikipedia.org/wiki/CRISPR).
Transgenic monkey (2014, Nature)http://www.nature.com/news/first-monkeys-with-customized-mutations-born-1.14611

Discussion with friends
• What are the autosome and X chromosome?
• How does the male drosophila have no sex-lethal protein?
• Explain the right figure 2
Find this interesting paper!!!