chapter 7: in vitro and in vivo supramolecular biomembrane
TRANSCRIPT

Chapter 7
In vitro and In vivo supramolecular biomembrane engineering using a lipidated coiled-coil motif
A simple and generic tool for the efficient formation of coiled coils at the surfaces of
cells and zebrafish embryos is presented here. A pair of complementary coiled-coil-
forming peptides, denoted E and K, were synthesized and conjugated to a cholesterol
anchor. The resulting lipidated peptides entered into CHO cell membranes
spontaneously in vitro and into the skin of zebrafish embryos in vivo. Addition of the
acetylated complementary peptide resulted in efficient coiled coil formation at the
surface of cells and zebrafish embryos. As the coiled coil unit acts as a molecular
velcro, this supramolecular tool enables the modification of biomembranes with a
wide range of molecular constructs. As a case study, the docking of liposomes to cell
membranes and zebrafish skin was investigated. Liposomes bearing the
complementary peptide efficiently docked at these biological interfaces through coiled
coil formation. Importantly, this is the first time that in vivo membrane modification
has been achieved. It is envisaged that this tool will aid the study of membrane
processes by modulating the chemistry of these bio-interfaces. Furthermore,
applications in the area of pharmacology and drug delivery such as high-throughput
screening of drugs are expected.
This work is published: H. Zope, F. Versluis, A. Ordas, J. Voskuhl, H. P. Spaink and
A. Kros, Angew. Chem. Int. Ed. Manuscript accepted for publication.

Chapter 7
Introduction
The molecular building blocks available in biological systems are able to self-
assemble into well-defined structures in an extremely controlled manner. These
structures need to be flexible and adaptive to the environment to enable them to carry
out their function in a regulated manner. Therefore, nature uses multiple weak
interactions (e.g. hydrogen bonding and van der Waals interactions) to act as the glue
to hold these structures together. When many weak interactions cooperatively
combine, relatively stable entities are produced, which retain the ability to ‘respond’
to external stimuli such as fluctuations in ion concentration, pH and temperature. For
many years, nature has been a source of inspiration for supramolecular chemistry.1-4
Here, scientists design relatively simple molecules which assemble into functional
materials with well-defined properties, typically following the bottom up approach.5-8
Recent progress has resulted in molecular systems which are responsive to multiple
stimuli and are therefore highly controlled, emulating nature ever more closely. A
relatively new branch of development is the application of supramolecular constructs
in an in vitro and in vivo environment, to directly study and influence biological
processes in live cells.
Introduction of supramolecular systems into a biological environment allows for the
placement of chemically tailored systems into, for example, cell membranes. This
enables the modification or regulation of cellular behavior through artificial signals.9
From a supramolecular chemistry point of view, two types of approach can be pursued
to introduce chemical species into a cell membrane: 1) specific binding of guest
molecules to membrane anchored biomolecules such as native proteins10 or 2) non-
specific labeling of membranes with the aid of hydrophobic and electrostatic
interactions or through a chemical cross linker.11-13 Lipidated peptides are particularly
good candidates for application in biological systems as their aggregation behavior
can be controlled by carefully balancing the hydrophobicity of the anchor and the
hydrophilicity of the cargo,14 which aids the incorporation of lipidated peptides into
membranes. By employing a coiled coil motif as the peptide segment, a highly
specific recognition system can be introduced into live cells. The coiled coil motif
acts as “molecular Velcro” and can thus be used to link distinct molecular constructs
together.15-20 An example of specific labeling of proteins through coiled coil formation
was recently supplied by Matsuzaki et al.21 Surface modification through non-specific
160

In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
binding of polymers to cell membranes has also been studied, for example by Ijiro et
al.22 Lipid grafted polymers adhered to cell membranes and could potentially act as a
scaffold to which a wide range of functional moieties could be attached, thereby
intervening in the chemistry of the cell. Furthermore, cationic graft copolymers have
also been shown to interact electrostatically with cell membranes, resulting in
chemically altered cell membranes.23 Whereas all these examples illustrate that in
vitro membrane functionalization is a highly rewarding strategy, examples of efficient
in vivo strategies are currently still lacking. Therefore, it is our goal to transiently
modify lipid membranes through specific supramolecular interactions in an in vitro
and in vivo environment. This will enable studies of membrane related processes by
modulating the chemistry at these interfaces. For this purpose, a pair of
complementary coiled coil forming lipidated peptides E and K,15, 24, 25 was used to
specifically introduce a non-covalent and bio-orthogonal recognition motif to
influence the chemistry of biological membranes (Scheme 1).
Scheme 1. Schematic representations of A) the lipidated E and K peptides, a CPE
decorated liposome and the fluorescently labelled E peptide, B) anchoring of CPK
into CHO cell membranes and subsequent coiled coil formation with fluorescently
labeled E or liposomes decorated with E peptides and C) anchoring of CPK into the
161

Chapter 7
skin of a zebrafish embryo and subsequent coiled coil formation with fluorescently
labeled E or liposomes decorated with peptide E.
Here, a generic supramolecular tool which allows us to rapidly and efficiently form
coiled coil motifs at the surface of biological membranes was developed. This is of
interest as a wide range of molecular constructs can be introduced to the surface of the
cell through a specific coiled coil formation. Coiled coil forming peptides E
[(EIAALEK)3] and K [(KIAALKE)3] 26-29 were first covalently conjugated to a
PEG12 spacer. Subsequently, a cholesterol moiety was coupled to the pegylated
peptides yielding CPE and CPK (Scheme 1A).30 The cholesterol moiety allows for the
immediate insertion of the lipidated peptides into membranes through hydrophobic
interactions and the PEG12 moiety was incorporated to aid efficient molecular
recognition between the peptide segments E and K. Recently, it was shown that upon
the addition of micellar solutions of either CPE or CPK to plain liposomes, the
lipidated peptides spontaneously inserted into liposomal membranes.30 In the present
study, CHO cell membranes and the skin of zebrafish embryos were modified with
coiled coil forming peptides by the addition of a micellar solution of CPE or CPK
resulting in immediate incorporation of these amphiphiles into the membranes.
Subsequently, the complementary peptide was added, resulting in coiled coil
formation. This system thus enables the docking of a wide variety of molecular
constructs through coiled coil formation (Scheme 1B and C).
Results and Discussion
In order to establish coiled coil formation at the surfaces of Chinese hamster ovarian
(CHO) cells, it was first confirmed that the cholesterol anchor entered into the
membranes of the cells. After the addition of a 5 µM solution of fluorescein labeled
CPE and incubation for a few minutes, confocal microscopy was used to image the
treated cells. As expected, the cells showed highly fluorescent membranes, indicating
that CPE was efficiently incorporated into the cell membrane (Figure 1). Surprisingly,
some internalization of CPE was observed. A similar effect was observed by Irvine et
al., who used a diacyllipid (C18) and cholesterol to anchor the oligonucleotides into
cell membranes.31 These lipidated oligonucleotides are able to anchor on the cell
membrane in 30 min whereas lipidated peptides could anchor in less than 5 min. One
explanation for this is that as oligonucleotides are more anionic as compared to the
162

In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
peptides used in this study, electrostatic repulsion might delay incorporation into the
membrane. It was also found that incorporating a PEG24 spacer between the
oligonucleotide and lipid segments drastically reduced the insertion capability; a
PEG12 spacer did not hinder membrane insertion. Future work in our group will focus
on these issues in order to have a better understanding of peptide amphiphile
membrane insertion.
Figure 1. Confocal images of CHO cells. The cells were treated with the nuclear stain
Hoechst for 30 min followed by incubation with fluorescently labeled CPE at 37 °C.
Images were obtained A) 0 min and B) 10 min after the addition of fluorescent CPE (5
µM). The inset shows a magnified image, revealing some internalization of the
lipopeptide. The scale bar represents 25 µm.
Subsequently, it was investigated whether the peptides at the CHO cell surface were
accessible and capable of forming coiled coils with the complementary peptide. For
this, cells were treated with a CPK solution of a final concentration of 5 μM, which is
the standard concentration for the lipidated peptides used throughout this study. A 5
minute incubation time was followed by washing with HBSS buffer to remove any
unbound CPK. Next, a solution containing 2.5 µM carboxyfluorescein labeled peptide
E was added to the cells and incubated for 10 minutes. Three more washes with HBSS
were performed to again remove any non-bound peptide E. Confocal imaging of the
CHO cells revealed that the fluorescent signal was located at the cell membranes
(Figure 2). In a control experiment the addition of CPK was omitted and as a result
the cells showed no fluorescence (Appendix, Figure S1). These observations strongly
suggest that the coiled coil formation between the membrane bound peptide E and
peptide K is efficient. Additionally, the opposite route could also be taken, i.e.,
163

Chapter 7
addition of CPE first, followed by treatment with fluorescent K peptides, which also
yielded specific coiled coil formation, as observed by the fluorescence located at the
membranes (Appendix, Figure S2). These results show that both CPK and CPE can
be used to functionalize the CHO cell membrane. In contrast, the protein engineering
approach only led to coiled coil formation when the engineered membrane protein
contained peptide E, but not the other way around. 32 Furthermore, our system
potentially allows for the modification of cell membranes of various cell lines without
the aid of laborious protein engineering protocols. Additionally, our supramolecular
method is fast and the necessary peptides can be synthesized within one day, as
opposed to the protein engineering approach, where the incorporation of the modified
receptor could take significantly more time.
Figure 2. (A, B, C) Confocal microscopy images (A: bright field, B: fluorescence and
C: overlay) indicating coiled coil formation between E and K at the surface of CHO
cells. First, a 5 µM solution of CPK in HBSS was added to the cells, 5 min incubation
time. Subsequently, fluorescently labeled (carboxyfluorescein) E peptides (2.5 µM)
were added and incubated for 5 minutes. Imaging took place after three washes with
HBSS. Experiments were performed at 37 °C. The scale bar represents 50 µm. (D, E,
F) Confocal microscopy images (D: bright field, E: fluorescence and F: overlay)
indicating docking of liposomes at the surface of CHO cells. First, a 5 µM solution of
CPK in HBSS was added to the cells, 5 min incubation time. Subsequently, CPE
decorated liposomes (0.25 mM total lipid concentration and 1 mol% CPE) with 1
mol% DHPE-TR were added to the cells. All the experiments were performed at 37
°C. The scale bar represents 25 µm.
164

In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
These initial studies demonstrated that coiled coils can be formed at the surface of
CHO cells by simply incubating a cholesterol peptide with the CHO cells and
subsequently adding the complementary peptide. This system enables the docking of
various molecules and nanoparticles to the cells. It was recently demonstrated that 100
nm liposomes can be immobilized at interfaces mediated by coiled coil formation
between peptides E and K.33 Therefore it was of interest to study whether liposome
docking is also possible at a biological interface. Instead of adding a fluorescent
peptide to the pre-treated cells, the complementary peptide was incorporated into
liposomes (Figure 2). After membrane functionalization with CPK, fluorescent
liposomes, containing 1 mol% DOPE-LR and 1 mol% CPE were incubated with the
cells for 5-10 minutes. After 3 washes, confocal images of the cells were obtained
(Figure 2). The images show increased fluorescence at the cell membranes.
Furthermore, with the CPE liposomes by themselves, i.e. without the presence of
peptide K at the cell membranes, no docking to the cells was observed (Appendix,
Figure S3). This shows that large supramolecular assemblies can be docked onto
membranes in an in vitro environment, using the orthogonal non-covalent coiled coil
motif. These findings enable future applications such as biophysical studies or
pharmaceutical research, for example adjuvant modified cells can be injected inside
animals, which might lead to cell therapy applications.34
Preclinical studies are a required step in translating innovations from a laboratory
setting to the clinic. Recent restrictions on animal testing have increased the demand
for alternative animal models. Therefore, zebrafish have become an attractive model
organism in recent years as they can be utilized as a tool for medical research bridging
the gap between cell-based assays and mammalian models.35 Due to its low-cost of
husbandry, high reproductive capability, optical transparency of the embryos, and the
ease of experimental manipulation, zebrafish might be a valuable tool in the
development of novel drug delivery systems for human disease therapy.36
Consequently, it was examined whether our coiled coil based system could also
function in an in vivo zebrafish embryo environment. To the best of our knowledge,
surface modification of zebrafish has not been studied before and therefore it was
investigated whether our surface engineering system is functional in this in vivo
environment (Scheme 1C). As the epidermis of zebrafish embryo consists only of
165

Chapter 7
living cells, unlike terrestrial vertebrates in which dead, keratinized cells are present,
it is possible to functionalize the whole organism.
One day-old zebrafish embryos were pre-treated with a solution of CPK in egg water.
Next, fluorescently labeled peptide E was added and incubated with the zebrafish
embryos for 30 minutes, followed by 3 washes with egg water prior to imaging.
Fluorescent stereomicroscope imaging revealed that the skin of the embryos was
highly fluorescent (Figure 3A-C). This is a strong indication that efficient coiled coil
formation was accomplished in vivo. Furthermore, the control experiment in which
the embryos were not pre-treated with CPK showed no significant fluorescence
(Appendix, Figure S4). This strongly indicates that CPK efficiently decorated the
skin of the zebrafish and that the peptide was available for coiled coil formation with
its complementary peptide. Similar to in vitro experiments, functionalizing zebrafish
embryos with CPE followed by incubation with fluorescently labeled K peptide was
also attempted. Surprisingly, this alternative route did not yield efficient coiled coil
formation on the skin of the zebrafish embryos, which is in contrast to the cell-based
study. Fish are known to have a different lipid composition than terrestrial vertebrates,
as they contain high levels of polyunsaturated fatty acids.37, 38 Presumably, this lipid
composition favors the positively charged K peptide over the negatively charged E
peptide. However, further studies are necessary to elucidate the difference in the
behavior of the peptides at the surface of CHO cells versus zebrafish embryos.
Figure 3. (A, B, C) Coiled coil formation between CPK and fluorescently labelled E
peptides on the skin of zebrafish embryos, as indicated by fluorescent
stereomicroscope images of 1 day-old zebrafish embryos. The embryos were pre-
treated with 5µM CPK in egg water, followed by incubation with 2.5 µM
complementary fluorescent peptide E for 30 minutes at 34°C (A: bright field, B:
166

In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
fluorescence, C: overlay). (D, E, F) Docking of liposomes on the skin of zebrafish
embryos through coiled coil formation. Fluorescent stereomicroscope images of 2
day-old zebrafish embryos pre-treated with 5µM CPK in egg water followed by
incubation with 0.25 mM liposomes bearing 1 mol% CPE and 1 mol% DHPE-TR on
the membrane for 30 minutes at 34°C (A: bright field, B: fluorescence, C: overlay).
Embryos are shown in lateral view, anterior to the left.
As a final investigation, docking of liposomes, as previously demonstrated for the
CHO cells, could also be achieved in vivo, on the skin of 2 day-old zebrafish embryos
was studied. In each group, embryos were selected and these embryos were first
incubated with CPK in egg water for 30 min at 34 °C. After 30 min. embryos were
washed and then incubated with 1 mol % CPK decorated fluorescently labeled
liposomes for 30 min at 34 °C. Upon the addition of fluorescent E decorated
liposomes to zebrafish that were pre-treated with CPK, the skin lit up brightly,
revealing the docking of liposomes to the skin of the embryos (Figure 3D-F). Control
experiments in which no CPK was administered displayed no significant fluorescence,
showing that liposome docking can be attributed to the specific coiled coil
interactions between peptides E and K (Appendix, Figure S5). As zebrafish have
been intensively studied as a model for various infectious diseases,39, 40 this study
enables future studies in which infected zebrafish embryos are treated using
therapeutic liposomes controlled by supramolecular coiled coil interactions.
Liposomes contain both hydrophilic and hydrophobic regions which enables a wide
range of drug encapsulation possibilities, which could be controllably released into
the zebrafish embryos. Therefore, in future studies a variety of therapeutic substances
will be screened, to investigate their effect on a range of diseases.41
Conclusions
In conclusion, a generic tool to modify the surface of live cells and zebrafish embryos
in a fast and efficient manner was designed. By using a supramolecular approach,
employing an orthogonal coiled coil forming peptide pair, specific interactions
between peptides E and K lead to coiled coil formation at the membrane of CHO cells
and the skin of zebrafish embryos. This method can be used to dock a wide range of
materials at a high local concentration to CHO cells and zebrafish embryos, which
was demonstrated by liposome docking in vitro and in vivo. This generic method
167

Chapter 7
holds much promise as an elegant and fast tool in supramolecular biomembrane
engineering and drug delivery. It also enables biophysical studies of biological
processes occurring at membranes. In future work, the aim is to introduce specificity
into the system. This might be achieved by constructing mixed micelles containing
our lipidated peptides and lipidated aptamers. The aptamers can be designed to attach
to specific cell types. In this way, it might be viable to deliver the lipidated peptides to
a specific cell type. Subsequent addition of liposomes that are decorated with the
complementary peptide should give cell specific liposome docking.
168

In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
References 1. J. Voskuhl and B. J. Ravoo, Chem. Soc. Rev., 2009, 38, 495-505. 2. D. A. Uhlenheuer, D. Wasserberg, C. Haase, H. D. Nguyen, J. H. Schenkel, J.
Huskens, B. J. Ravoo, P. Jonkheijm and L. Brunsveld, Chem.-Eur. J., 2012, 18, 6788-6794.
3. R. V. Ulijn and A. M. Smith, Chem. Soc. Rev., 2008, 37, 664-675. 4. M. A. Kostiainen, O. Kasyutich, J. Cornelissen and R. J. M. Nolte, Nat.
Chem., 2010, 2, 394-399. 5. J. M. Lehn, Angew. Chem.-Int. Edit. Engl., 1990, 29, 1304-1319. 6. C. A. Mirkin, R. L. Letsinger, R. C. Mucic and J. J. Storhoff, Nature, 1996,
382, 607-609. 7. L. C. Palmer and S. I. Stupp, Acc. Chem. Res., 2008, 41, 1674-1684. 8. R. P. Sijbesma and E. W. Meijer, Current Opinion in Colloid & Interface
Science, 1999, 4, 24-32. 9. B. R. Peterson, Org. Biomol. Chem., 2005, 3, 3607-3612. 10. E. M. Sletten and C. R. Bertozzi, Angew. Chem.-Int. Edit., 2009, 48, 6974-
6998. 11. J. H. Felgner, R. Kumar, C. N. Sridhar, C. J. Wheeler, Y. J. Tsai, R. Border, P.
Ramsey, M. Martin and P. L. Felgner, J. Biol. Chem., 1994, 269, 2550-2561. 12. F. Borcard, A. Godinat, D. Staedler, H. C. Blanco, A. L. Dumont, C. Chapuis-
Bernasconi, C. Scaletta, L. A. Applegate, F. K. Juillerat, U. T. Gonzenbach, S. Gerber-Lemaire and L. Juillerat-Jeanneret, Bioconjugate Chem., 2011, 22, 1422-1432.
13. D. Dutta, A. Pulsipher, W. Luo, H. Mak and M. N. Yousaf, Bioconjugate Chem., 2011, 22, 2423-2433.
14. F. Versluis, H. R. Marsden and A. Kros, Chem. Soc. Rev., 2010, 39, 3434-3444.
15. A. L. Boyle and D. N. Woolfson, Chem. Soc. Rev., 2011, 40, 4295-4306. 16. H. Gradisar, S. Bozic, T. Doles, D. Vengust, I. Hafner-Bratkovic, A. Mertelj,
B. Webb, A. Sali, S. Klavzar and R. Jerala, Nature chemical biology, 2013, 9, 362-366.
17. B. Apostolovic, M. Danial and H. A. Klok, Chem. Soc. Rev., 2010, 39, 3541-3575.
18. H. R. Marsden, A. V. Korobko, E. N. M. van Leeuwen, E. M. Pouget, S. J. Veen, N. Sommerdijk and A. Kros, J. Am. Chem. Soc., 2008, 130, 9386-9393.
19. J. M. Mason and K. M. Arndt, ChemBioChem, 2004, 5, 170-176. 20. W. A. Petka, J. L. Harden, K. P. McGrath, D. Wirtz and D. A. Tirrell, Science,
1998, 281, 389-392. 21. Y. Yano and K. Matsuzaki, Biochim. Biophys. Acta-Biomembr., 2009, 1788,
2124-2131. 22. R. Kamitani, K. Niikura, T. Okajima, Y. Matsuo and K. Ijiro, ChemBioChem,
2009, 10, 230-233. 23. J. T. Wilson, V. R. Krishnamurthy, W. X. Cui, Z. Qu and E. L. Chaikof, J.
Am. Chem. Soc., 2009, 131, 18228-18229. 24. H. Robson Marsden, A. V. Korobko, T. Zheng, J. Voskuhl and A. Kros,
Biomaterials Science, 2013. 25. T. Zheng, J. Voskuhl, F. Versluis, H. R. Zope, I. Tomatsu, H. R. Marsden and
A. Kros, Chem. Commun., 2013, 49, 3649-3651. 26. J. R. Litowski and R. S. Hodges, J. Biol. Chem., 2002, 277, 37272-37279.
169

Chapter 7
27. H. R. Marsden, N. A. Elbers, P. H. H. Bomans, N. Sommerdijk and A. Kros, Angew. Chem.-Int. Edit., 2009, 48, 2330-2333.
28. B. Apostolovic, S. P. E. Deacon, R. Duncan and H. A. Klok, Macromol. Rapid Commun., 2011, 32, 11-18.
29. M. B. Hansen, W. P. R. Verdurmen, E. H. P. Leunissen, I. Minten, J. C. M. van Hest, R. Brock and D. Lowik, ChemBioChem, 2011, 12, 2294-2297.
30. F. Versluis, J. Voskuhl, B. van Kolck, H. Zope, M. Bremmer, T. Albregtse and A. Kros, J. Am. Chem. Soc., 2013, 135, 8057-8062.
31. H. P. Liu, B. Kwong and D. J. Irvine, Angew. Chem.-Int. Edit., 2011, 50, 7052-7055.
32. Y. Yano, A. Yano, S. Oishi, Y. Sugimoto, G. Tsujimoto, N. Fujii and K. Matsuzaki, ACS Chem. Biol., 2008, 3, 341-345.
33. J. Voskuhl, C. Wendeln, F. Versluis, E. C. Fritz, O. Roling, H. Zope, C. Schulz, S. Rinnen, H. F. Arlinghaus, B. J. Ravoo and A. Kros, Angew. Chem.-Int. Edit., 2012, 51, 12616-12620.
34. M. T. Stephan, J. J. Moon, S. H. Um, A. Bershteyn and D. J. Irvine, Nat. Med., 2010, 16, 1035-U1135.
35. S. A. Brittijn, S. J. Duivesteijn, M. Belmamoune, L. F. M. Bertens, W. Bitter, J. D. De Bruijn, D. L. Champagne, E. Cuppen, G. Flik, C. M. Vandenbroucke-Grauls, R. A. J. Janssen, I. M. L. De Jong, E. R. De Kloet, A. Kros, A. H. Meijer, J. R. Metz, A. M. Van der Sar, M. J. M. Schaaf, S. Schulte-Merker, H. P. Spaink, P. P. Tak, F. J. Verbeek, M. J. Vervoordeldonk, F. J. Vonk, F. Witte, H. P. Yuan and M. K. Richardson, Int. J. Dev. Biol., 2009, 53, 835-850.
36. C. Chakraborty, C. H. Hsu, Z. H. Wen, C. S. Lin and G. Agoramoorthy, Curr. Drug Metab., 2009, 10, 116-124.
37. R. J. Henderson and D. R. Tocher, Prog. Lipid Res., 1987, 26, 281-347. 38. M. Holtta-Vuori, V. T. V. Salo, L. Nyberg, C. Brackmann, A. Enejder, P.
Panula and E. Ikonen, Biochem. J, 2010, 429, 235-242. 39. A. M. van der Sar, B. J. Appelmelk, C. Vandenbroucke-Grauls and W. Bitter,
Trends Microbiol., 2004, 12, 451-457. 40. G. J. Lieschke and P. D. Currie, Nat. Rev. Genet., 2007, 8, 353-367. 41. R. Carvalho, J. de Sonneville, O. W. Stockhammer, N. D. L. Savage, W. J.
Veneman, T. H. M. Ottenhoff, R. P. Dirks, A. H. Meijer and H. P. Spaink, PLoS One, 2011, 6.
170

In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
Experimental section
Materials
The Fmoc-protected amino acids were purchased from Novabiochem. The Sieber
Amide resin was purchased from Agilent Technologies. Fmoc-NH-PEG12-COOH was
purchased from IRIS Biotech. DOPE, cholesterol and DOPC were obtained from
Avanti Polar Lipids. Cholesteryl hemisuccinate and 5,6-Carboxyfluorescein were
obtained from Sigma-Aldrich. 8 well plate Lab-tek was purchased from Thermo
Scientific, USA. DMEM media was obtained from PAA-the cell culture company. 3-
aminobenzoic acid (Tricaine) was purchased from Sigma-Aldrich.
General methods
Peptide synthesis
The cholesterol modified peptides were synthesized as described elsewhere in
chapter 4. Synthesis of fluorescently labeled peptides was performed on rink amide
resin. After initial Fmoc deprotection using 20% peperidine in DMF, Fmoc-Lys(boc)-
COOH was coupled, using 4 eq. of DIC and 4 eq. of HOBT, for 1 hour. Next, the side
chain Boc group was removed using TFA/ DCM (50:50 v/v) for 30 mins. The resin
was then neutralized using DIPEA/NMP (10:90 v/v) for 10 mins (3x). 5(6)-
Carboxyfluorescein (Sigma Adrich) was coupled using 4 eq. of DIC and 4 eq. of
HOBT dissolved in DMF and coupling was performed overnight. The resulting
fluorescently labeled resin was used to synthesize fluorescently labeled acetylated
peptide E and K along with fluorescently labeled CPE using standard Fmoc chemistry
procedure described earlier.30
Peptide and liposome sample preparation
Lipid (DOPC/DOPE/CH, molar ratios 2:1:1, total concentration 1 mM in CHCl3) and
peptide (50 µM in 1:1 CHCl3/MeOH) stock solutions were prepared. Peptide samples
were prepared by taking the appropriate amount of peptide stock, evaporating the
solvent over a stream of air, addition of HBSS and sonication for 1 min at 55°C.
Liposomes were prepared by using appropriate amounts of lipid and peptide stock
solutions evaporating the solvent over a stream of air, addition of HBSS and
sonication for 1 min at 55°C.
171

Chapter 7
CHO cell culturing and experiments
Chinese hamster Ovary (CHO) cells were cultivated in Dulbecco’s Modified Eagle’s
Medium (DMEM, PAA-The Cell Culture Company), supplemented with 10% fetal
calf serum (iron supplied), 2% of L-glutamine, 1% penicillin and 1% streptomycin.
The cells were cultured with 5% CO2 at 37 °C. The medium was refreshed every two
days and the cells were passaged by trypsinization at 70% confluence. For
fluorescence assays, the cells were transferred and grown on an 8-well slide (Lab-Tek,
Thermo scientific). One washing step with HBSS was performed. Subsequently, the 5
µM CPK solution was added and left to incubate for 5 minutes at 37 °C. Next, the
liquid was removed and a single washing step was performed. Afterwards, either
fluorescently labeled E peptide (2.5 µM) or 250 µM fluorescently labeled E decorated
liposomes were added to the cells and left to incubate for 5-10 minutes. Subsequently,
the microscope slide was transferred to a Leica SPE confocal microscope and images
were obtained within 1 h. In the CPE incorporation experiment, nuclei were stained
with 100 ng/mL Hoechst 33342 in DMEM for 30 min. 5 µM fluorescently labeled
CPE was added to the wells and cells were imaged over 10 min using an automated
Nikon TiE2000 confocal microscope.
Zebrafish embryo experiments
Zebrafish (Danio rerio) were handled in compliance with the local animal welfare
regulations and maintained according to standard protocols (http://ZFIN.org). After
harvesting embryos from single crosses they were grown at 28°C in egg water
(60μg/ml Instant Ocean sea salts). The stage of embryo was selected based on the auto
fluorescence levels. At one day post fertilization dechorionated embryos were
incubated with the peptides added to the egg water. The experimental group was pre-
treated in 5µM CPK for 30 minutes at 34°C. Next, CPK was replaced with 2.5 µM
complementary fluorescent E peptide and incubated for another 30 minutes at 34°C.
Embryos in the control group were exposed only to 2.5 µM complementary
fluorescent E peptide. Following fluorescent peptides were washed away three times
with egg water; embryos were anesthetized with 200 μg/mL buffered 3-aminobenzoic
acid (Tricaine) and imaged with a Leica M205FA stereo fluorescent microscope.
Docking of liposomes on the biological membranes was also studied using 2 day-old
zebrafish embryos as an in vivo system. 5 µM solution of CPK in egg water was
added to the embryos for 30 minutes incubation at 34°C. After subsequent washing,
172
In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
0.25 mM liposomes containing 1 mol% fluorescent DHPE-TR and 1 mol% CPE were
added to the embryos for another 30 minutes at 34°C. Washing and imaging was
performed as mentioned above.
Characterization of fluorescently labeled peptides using HRMS
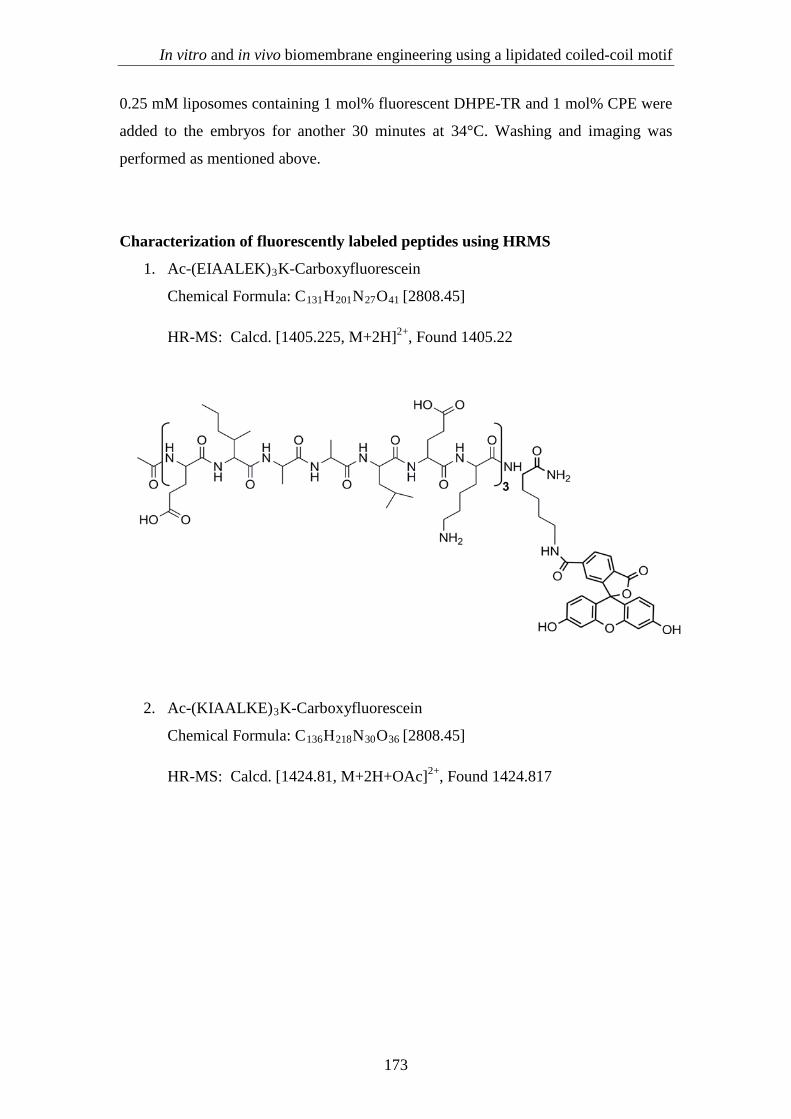
1. Ac-(EIAALEK)3K-Carboxyfluorescein
Chemical Formula: C131H201N27O41 [2808.45]
HR-MS: Calcd. [1405.225, M+2H]2+, Found 1405.22
2. Ac-(KIAALKE)3K-Carboxyfluorescein
Chemical Formula: C136H218N30O36 [2808.45]
HR-MS: Calcd. [1424.81, M+2H+OAc]2+, Found 1424.817
173
Chapter 7
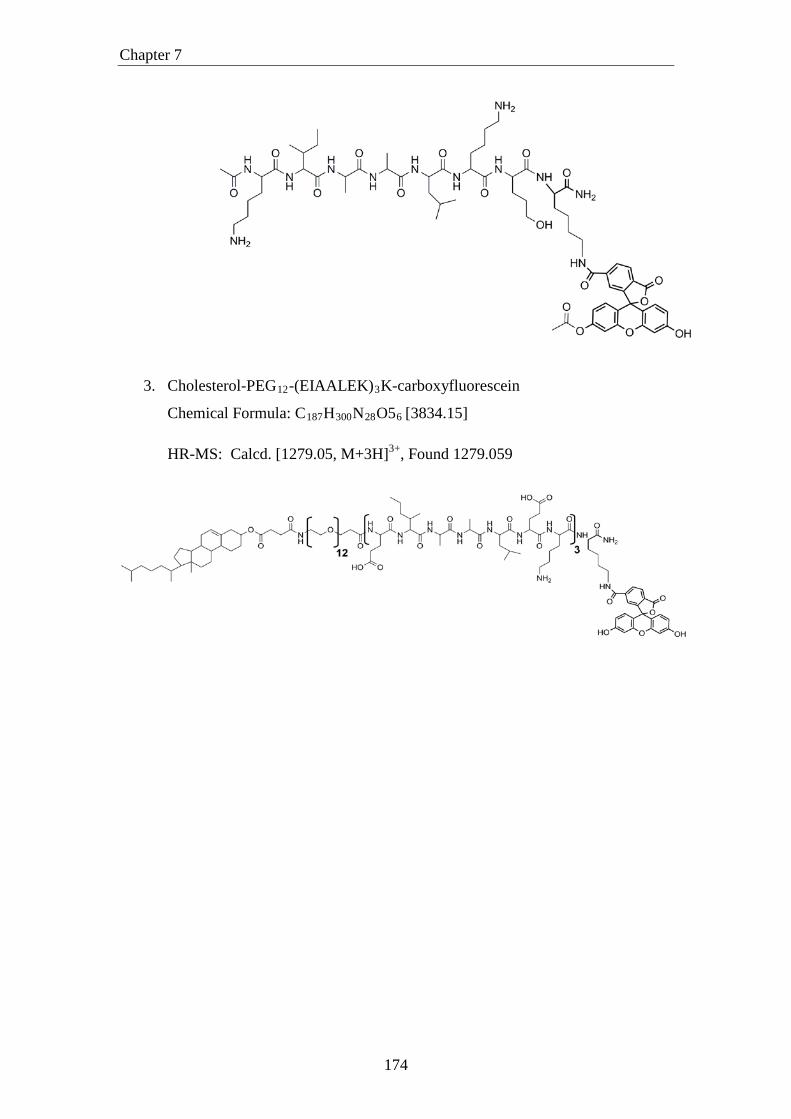
3. Cholesterol-PEG12-(EIAALEK)3K-carboxyfluorescein
Chemical Formula: C187H300N28O56 [3834.15]
HR-MS: Calcd. [1279.05, M+3H]3+, Found 1279.059
174

In vitro and in vivo biomembrane engineering using a lipidated coiled-coil motif
Appendix: supplementary information
Confocal images of CHO cells
A) B) C)
Figure S1. Confocal microscopy images (A: bright field, B: fluorescence and C:
overlay) indicating that peptide E by itself does not bind to the cell membranes. Here,
only fluorescently labeled (carboxyfluorescein) E peptides were added and incubated
for 5 minutes with the CHO cells. Imaging took place after three washes with HBSS.
Experiments performed at 37 °C. The scale bar represents 50 µm.
Figure S2. Confocal microscopy image of CHO cells, showing that the addition of
CPE first, followed by fluorescently labelled K resulted in efficient coiled coil
formation. First, 5 µM CPE (5 µM) was added to the cells for 10 min. Subsequent
addition of fluorescently labeled K peptides (2.5 µM). The image reveals that this
reverse route also yields fluorescence located at the cell membranes, indicating that
coiled coil formation occurred efficiently. All the experiments were performed at 37
°C. The scale bar represents 50 µm.
175
Chapter 7
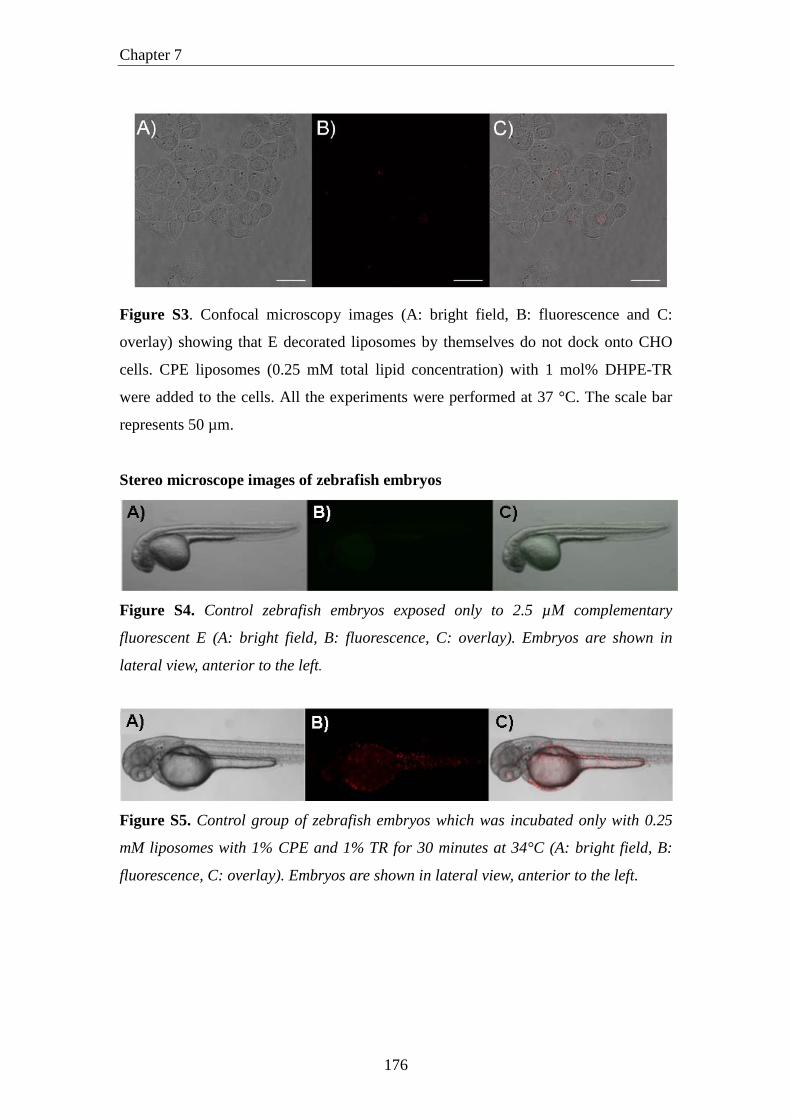
Figure S3. Confocal microscopy images (A: bright field, B: fluorescence and C:
overlay) showing that E decorated liposomes by themselves do not dock onto CHO
cells. CPE liposomes (0.25 mM total lipid concentration) with 1 mol% DHPE-TR
were added to the cells. All the experiments were performed at 37 °C. The scale bar
represents 50 µm.
Stereo microscope images of zebrafish embryos
Figure S4. Control zebrafish embryos exposed only to 2.5 µM complementary
fluorescent E (A: bright field, B: fluorescence, C: overlay). Embryos are shown in
lateral view, anterior to the left.
Figure S5. Control group of zebrafish embryos which was incubated only with 0.25
mM liposomes with 1% CPE and 1% TR for 30 minutes at 34°C (A: bright field, B:
fluorescence, C: overlay). Embryos are shown in lateral view, anterior to the left.
176