chapter 5shodhganga.inflibnet.ac.in/bitstream/10603/36846/11/16 chapter-5.pdf · chapter-5 integral...
TRANSCRIPT

CHAPTER-5
In Vivo Hepatoprotective Activity
of Nigella sativa L. Seed in
Various Germination Phases
“One thing I have learned in a long life: that all our science,
measured against reality, is primitive and childlike -- and yet
it is the most precious thing we have.”
~Albert Einstein

CHAPTER-5
Integral University, Lucknow 166
5.1 Rationale
Largest and complex organ in the body is liver which deals multiple and
diverse functions to safeguard internal environment of the body. Liver is involved in
synthesizing number of various plasma proteins such as albumin, fibrinogen and the
intermediate metabolism of protein, fat and carbohydrates. It is also involved in
synthesis of clotting factors and also produces a number of enzyme, formation and
excretion of bile. It acts as a storage depot for proteins, glycogen, various vitamins
and metals. It also has a role in the regulation of blood volume by transferring the
blood from portal to systemic circulation and its reticule endothelial system and
participates in immune mechanism. It plays a central role in detoxification and
excretion of many endogenous and exogenous compounds. Hence, any injury to it or
impairment of its function has grave implication for the health of the affected person.
Liver diseases are affecting humans of all ages and one of the major causes of
morbidity and mortality in public. About 20,000 deaths occur every year due to liver
disorder. Hepatocellular carcinoma is one of the ten most common tumors in the
world with over 2,50,000 new cases each year (Gupta and Misra, 2006). According
to WHO estimates, globally 170 million people are chronically infected with
hepatitis C alone and every year 3–4 millions are newly added into the list. Also,
there are more than 2 billion people infected by hepatitis B virus (Negi et al., 2008).
Every year about 18,000 people are reported to die due to liver cirrhosis caused by
hepatitis (Handa, 1991).
Usually liver fibrosis is initiated by hepatocyte damage and various biologic
factors such as hepatitis virus, bile duct obstruction, overload of cholesterol,
schistosomiasis or chemical factors such as CCl4 administration, alcohol intake etc.
were known to contribute to liver fibrosis. Hepatic fibrosis is a major feature of wide
range of chronic liver injuries including metabolic, viral, cholestatic and genetic
diseases. The failure of bile salt excretion in cholestatic leads to retention of
hydrophobic bile salts within the hepatocytes and causes apoptosis and/or necrosis

CHAPTER-5
Integral University, Lucknow 167
(Miyoshi et al., 1999). Oxidative stress has been implicated in the pathogenesis of
various liver diseases including alcoholic liver disease, nonalcoholic fatty liver
disease, and chronic hepatitis C (Seki et al., 2005; Kitase et al., 2005). The liver
diseases are classified as acute or chronic depending on the duration of the disease.
Acute viral hepatitis and drug reactions account for the majority of cases of acute
liver diseases. Chronic liver damage is a worldwide common pathology
characterized by inflammation and fibrosis that can lead to chronic hepatitis,
cirrhosis and cancer (Tessitore and Bollito, 2006; Kohle et al., 2008). Chronic
hepatitis or long term intoxification can severely injure hepatic cells.
Drug/chemical-mediated hepatic injury is the common sign of drug toxicity
(Lee, 2003) and accounts for greater than 50% of acute liver failure cases. Hepatic
damage is the largest obstacle to the development of drugs and is the major reason
for withdrawal of drugs from the market (Cullen and Miller, 2006). Drug-induced
liver disease can be predictable (high incidence and dose-related) or unpredictable
(low incidence and may or may not be dose-related). Conventional drugs used in the
treatment of liver diseases are sometimes inadequate and can have serious adverse
effects. Herbal medicines are in great demand in the developed as well as developing
countries for primary healthcare because of their wide biological and medicinal
activities, higher safety margins and lesser costs (Chattopadhyay et al., 2007).
Presently a few hepatoprotective drugs and that too from natural sources are
available for the treatment of liver disorders. Hence, people are looking at the
traditional systems of medicine for remedies to hepatic disorders.
The present study is aimed to evaluate the hepatoprotective and antioxidant
activity of methanolic extract of the Nigella sativa L. from different germination
stages of seed against CCl4-induced hepatotoxicity in rats as hepatotoxin to prove its
claim in folklore practice against liver disorders with no side effects.

CHAPTER-5
Integral University, Lucknow 168
5.2 REVIEW OF LITRARURE
5.2.1 Hepatotoxicity (causes and pattern of liver damage)
As the major drug metabolizing and detoxifying organ in the body, the liver
is subjected to potential damage from an enormous array of
pharmaceutical and environmental chemicals. More than 900 drugs have been
implicated in causing liver injury (Keeffe et al., 2004) and it is the most common
reason for a drug to be withdrawn from the market. Chemicals often cause
subclinical injury to liver which manifests only as abnormal liver enzyme tests.
Drug-induced liver injury is responsible for 5% of all hospital admissions and 50%
of all acute liver failures (Ostapowicz et al., 2002). Liver is susceptible to injury
from drugs and other substances. 75% of blood coming to the liver arrives directly
from gastrointestinal organs and then spleen via portal veins which bring drugs and
xenobiotics in near-undiluted form. There are many mechanisms behind liver
damage, many chemicals damage mitochondria and its dysfunction releases
excessive amount of oxidants which in turn injured hepatic cells. Activation of some
enzymes in the cytochrome P-450 system such as CYP2E1 also leads to oxidative
stress (Jaeschke H et al., 2002). Injury to hepatocyte and bile duct cells lead to
accumulation of bile acid inside liver that promotes further liver damage (Patel T et
al., 1998).
Drugs/chemicals produce a wide variety of clinical and pathological hepatic
injury. Biochemical markers (e.g. alanine transferase, alkaline phosphatase and
bilirubin) are often used to indicate liver damage. Liver injury is defined as a rise in
either ALT level more than three times of upper limit of normal, ALP level more
than twice upper limit of normal or total bilirubin level more than twice upper limit
of normal when associated with increased ALT or ALP (Mumoli N et al., 2006).
Three important characteristics of the P450 system have role in drug induced
toxicity (Figure 5.1).

CHAPTER-5
Integral University, Lucknow 169
a. Direct toxicity
b. Hepatic conversion of xenobiotics to an active toxin
c. The immune mechanism usually by a drug or a metabolite acting as hepatic to
convert cellular protein into an immunogen.
Figure 5.1: Drug and toxin induced hepatic injury.
Exposure to various organic compounds including a number of
environmental pollutants and drugs can cause cellular damages through metabolic
activation of those compounds to highly reactive substances such as reactive oxygen
species (ROS). Drugs or toxins that have a pharmacological hepatotoxicity are those
that have predictable dose-response curves (higher concentrations cause more liver
damage) and well characterized mechanisms of toxicity such as directly damaging
liver tissue or blocking a metabolic process. Idiosyncratic injury occurs without
warning when agents cause non-predictable hepatotoxicity in susceptible individuals
which is not related to dose and has a variable latency period (Zimmerman, 1978).

CHAPTER-5
Integral University, Lucknow 170
Idiosyncratic hepatotoxicity has led to the withdrawal of several drugs from market
even after rigorous clinical testing as part of the FDA approval process.
5.2.2 Carbon tetrachloride: A most commonly used hepatotoxin
Chemical toxicity comprises an important source of reactive oxygen species
(ROS), which may occur through processes such as inhibition of
mitochondrial electron transport chain and subsequent accumulation of
intermediates inactivation of antioxidant enzymes and deletion of radical
scavengers (Mates, 2000). Carbon tetrachloride (CCl4) is a well known hepatotoxin
and exposure to this chemical is known to induce oxidative stress and causes liver
injury by the formation of free radicals (Sil et al., 2007). Carbon tetrachloride is
one of the most commonly used hepatotoxin in the experimental study of liver
disease (Johnston et al., 1998).
Carbon tetrachloride is used in the synthesis of chlorinated organic
compounds, including chlorofluorocarbon refrigerants. It is also used as an
agricultural fumigant and as a solvent in the production of semiconductors, in the
processing of fats, oils and rubber and in laboratory applications (Lewis, 1993;
Kauppinen et al., 1998). CCl4 continues to provide an important service today as a
model substance to elucidate the mechanisms of action of hepatotoxic effects such as
fatty degeneration, fibrosis, hepatocellular death, and carcinogenicity. CCl4 has been
extensively used to study liver injury induced by ROS in the mouse model, which is
closely analogous to hepatotoxicity in humans.
CHAPTER-5
Integral University, Lucknow 171
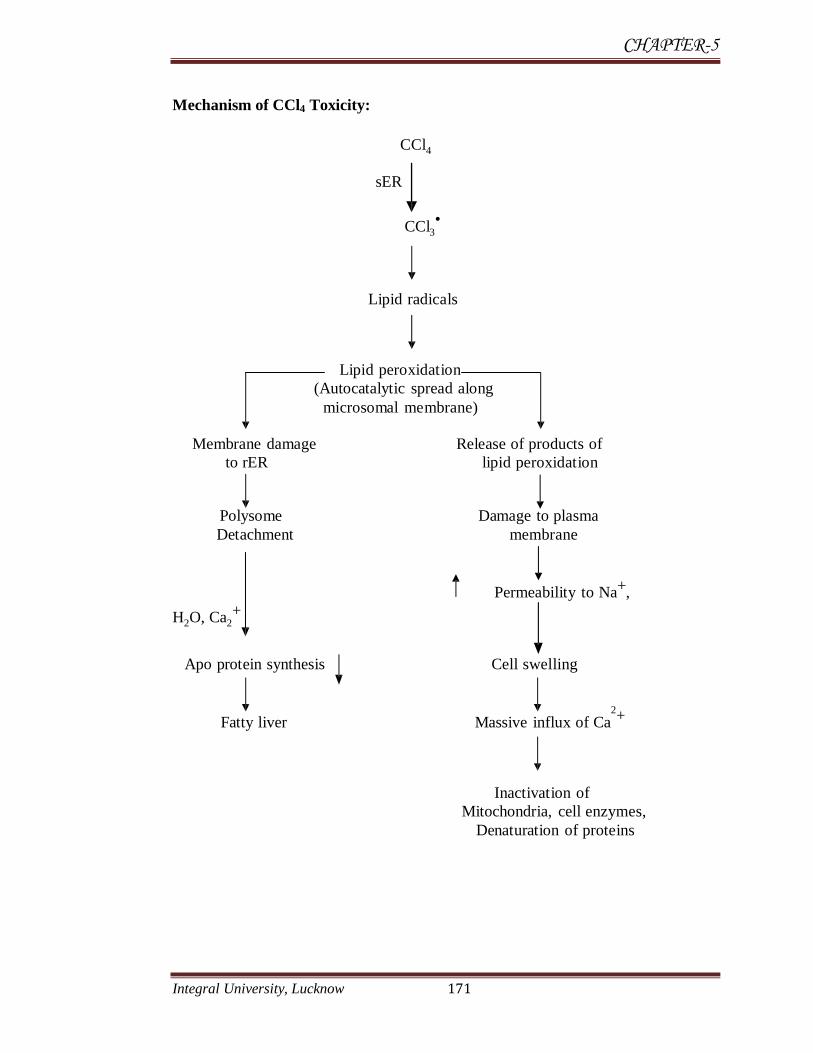
Mechanism of CCl4 Toxicity:
CCl4
sER
CCl3•
Lipid radicals Lipid peroxidation
(Autocatalytic spread along
microsomal membrane)
Membrane damage Release of products of
to rER lipid peroxidation Polysome Damage to plasma
Detachment membrane
Permeability to Na+,
H2O, Ca2+
Apo protein synthesis Cell swelling
Fatty liver Massive influx of Ca2+
Inactivation of
Mitochondria, cell enzymes,
Denaturation of proteins

CHAPTER-5
Integral University, Lucknow 172
The hepatotoxic effect of CCl4 are largely due to its active metabolites,
CCl3 (trichloro methyl radicals) CCl3•-
(tri chloro methyl per oxy radicals) is
most reactive species and causes damage to biological macromolecules by
combining with them there by causing covalent modification and setting the
chain reactions of lipid peroxidation and ultimately cell death (Benedetti et al.,
1987). Among various mechanism involved in hepatotoxic effect of carbon
tetrachloride, one is oxidative damage through free radicals generation (Deleve et
al., 1995). The hepatotoxicity of CCl4 is attributed to the formation of
trichloromethyl and trichloromethyl peroxyl radicals, initiating lipid peroxidation
and resulting in fibrosis and cell necrosis (Kadiiska et al., 2000).
At the molecular level CCl4 activates tumor necrosis factor TNF-alpha, nitric
oxide (NO), transforming growth factors, TGF-alpha and TGF-beta in the cell
processes that appear to direct the cell primarily toward self destruction or fibrosis.
TNF-alpha pushes toward apoptosis whereas the TGFs appear to direct toward
fibrosis. CCl4 intoxication also leads to hypomethylation of cellular components; in
the case of RNA the outcome is thought to be inhibition of protein synthesis, in the
case of phospholipids it plays a role in the inhibition of lipoprotein secretion (Weber
et al., 2003). The over production of ROS and therefore oxidative stress can be
initiated by a variety of factors including exposure to xenobiotics, such as
acetaminophen and carbon tetrachloride (Ronsein et al., 2005). Of concern here is
the adverse effect of carbon tetrachloride to the liver, having visualized the
prominent functions of the liver for survival. According to Liu et al., (1995),
ingestion of carbon tetrachloride can lead to marked hepatotoxicity.
5.2.3 Free radicals and oxidative damage in liver
A free radical can be defined as any species which is capable of
independent existence and contains one or more unpaired electrons (Halliwell and
Gutteridge, 1989). In excess, free radicals can damage cell membranes and cause
cell necrosis; they can cause damage to all macromolecules, lipids, proteins

CHAPTER-5
Integral University, Lucknow 173
mitochondrial and nuclear DNA molecules of the cells causing inflammation or
lesion on various organs (Beckman et al., 1990; Halliwell and Gutteridge, 1984). It
is increasingly being realized that majority of disorders/diseases is mainly due to
the imbalance between pro-oxidant and antioxidant homeostatic phenomenon in the
body. Pro-oxidants conditions either dominate due to increase in the generation of
free radicals and/or their inadequate quenching or scavenging in the body (Tiwari,
2001). The reactive oxygen species play an important role related to the degenerative
or pathological processes of various serious diseases such as aging (Burns et al.,
2001), cancer, coronary heart disease, Alzheimer’s disease (Smith et al., 1996; Diaz
et al., 1997), neurodegenerative disorders, atherosclerosis, cataracts and
inflammation (Aruoma, 1998).
ROS such as superoxide radical, hydroxyl radical, peroxyl radical and nitric
oxide radical attack biological molecules such as lipids, proteins, enzymes, DNA
and RNA. This may be lead to cell or tissue injury associated with aging,
atherosclerosis, carcinogenesis (Keli Chen et al., 2005) and development of
chronic diseases related to the cardio and cerebrovascular systems. More
than one hundred disorders in humans including atherosclerosis, arthritis, ischemia,
reperfusion injury of many tissues, central nervous system injury, gastritis, cancer
and AIDS associate with reactive oxygen species (Cook and Samman, 1996).
ROS play an important role in fibrogenesis throughout increasing platelet-
derived growth factor. Most hepatocellular carcinomas occur in cirrhotic livers and
the common mechanism for hepatocarcinogenesis is chronic inflammation associated
with severe oxidative stress; other risk factors are dietary aflatoxin B1 consumption,
cigarette smoking and heavy drinking. Ischemia–reperfusion injury affects directly
on hepatocyte viability particularly during transplantation and hepatic surgery.
Ischemia activates Kupffer cells which are the main source of ROS during the
reperfusion period (Pablo Muriel, 2009). Hepatocellular carcinoma is one of the most
malignant and frequent worldwide spreading diseases. It is the third most common
cause of cancer deaths (Pisani et al., 1999; Parkin et al., 2005). Most Hepatocellular

CHAPTER-5
Integral University, Lucknow 174
carcinoma occurs in cirrhotic livers and the common mechanism for
hepatocarcinogenesis is chronic inflammation associated with severe oxidative stress
(Seitz et al., 2006). There is a large body of evidence indicating that ROS play a
pathogenesis role in carcinogenesis in liver (Marx et al., 2004).
5.2.4 Antioxidant
Almost all organisms are well protected against free radical damage by anti-
oxidative enzymes such as superoxide dismutase (SOD) and catalase (CAT) or
chemical compounds such as α-tocopherol, ascorbic acid, carotenoids, polyphenol
compounds and glutathione. When the mechanism of antioxidant protection
becomes unbalanced by factors such as ageing, deterioration of physiological
functions may occur resulting in diseases and accelerated ageing. However,
antioxidant supplements or antioxidant containing foods may be used to help
the human body to reduce oxidative damage (Mau et al., 2001; Gulcin et al.,
2002).
All living organisms have endogenous defense systems against oxidative
damage such as lipid peroxidation DNA damage (Lee et al., 2005) and inhibition
of cell communication due to reactive oxygen species (ROS). Antioxidants protect
against chemotherapy toxicity and local toxic effects of tumors or surrounding
tissues. The protection of cells against damage from oxygen and its metabolites can
be accomplished through enzymatic and non-enzymatic means. Superoxide
dismutase, catalase and glutathione peroxides (GPx) are considered to be the
primary antioxidant enzymes, since they are involved in the direct elimination of
reactive oxygen species. Glutathione-S-transferase (GST), glutathione reductase
(GR) and glucose- 6-phosphate dehydrogenase (G6PD) are secondary antioxidant
enzymes which help in the detoxification of reactive oxygen species by
decreasing peroxide levels by GST or by maintaining a steady supply of metabolic
intermediates like glutathione as by GR and NADPH by, G6PD for the primary
antioxidant enzymes. The non-enzymatic small molecular antioxidants include

CHAPTER-5
Integral University, Lucknow 175
sulfhydryl compounds such as glutathione (GSH) and thiols, NADPH, ascorbate, α-
tocopherol etc., (Halliwell and Gutteridge 1984). Phytochemical components such
as polyphenols, ascorbic acid, and carotenoids also serve as antioxidants (Rice-
Evans et al., 1997). Oxygen is essential for aerobic life process. However, cells
under aerobic condition are threatened with the insult of reactive oxygen molecules
that are efficiently taken care of by the powerful antioxidant system in human body.
5.2.5 Hepatoprotective activities of Nigella sativa L.
A large number of medicinal plants have been tested and found to contain
active principles with curative properties against a variety of diseases (Lewis,
1977). Liver protective plants contain a variety of chemical constituents like
phenols, coumarins, lignans, essential oil, monoterpenes, carotinoids, glycosides,
flavanoids, organic acids, lipids, alkaloids and xanthenes. Recent experience has
shown that plant drugs are relatively non-toxic, safe and even free from serious
side effects (Momin, 1987).
Nigella sativa L. is one of the most revered medicinal seed in history. A large
number of in vitro and in vivo studies have been conducted on laboratory animals
and humans in order to investigate its pharmacological properties like
immunostimulation, anti-inflammatory, hypoglycemic, antihypertensive,
antiasthmatic, antimicrobial, antiparasitic, antioxidant as well as anticancer
properties (Randhawa and Alghamdi, 2002). Acute and chronic toxicity studies on
laboratory animals have reported that N. sativa, its oil and thymoquinone the most
widely studied active principle are quite safe particularly when given orally (Badary
et al., 1997; Mansour et al., 2001; Ali et al., 2003).
It has been reported that N. sativa oil possesses hepatoprotective effects in
some models of liver toxicity. However, it is N. sativa seed that is used in the
treatment of liver ailments in folk medicine rather than its oil. Clinical and
experimental investigations have shown that N. sativa has a protective effect against

CHAPTER-5
Integral University, Lucknow 176
oxidative damage in isolated rat hepatocytes (Daba et al., 1998). It was found that
the fixed oil of N. sativa has both antioxidant and anti-eicosanoid effects greater than
thymoquinone which is its active constituent (Houghton et al., 1995). Furthermore,
N. sativa has antioxidant activity by suppressing the chemiluminescence in
phagocytes (Haq et al., 1995).
Administration of oil extract of N. sativa to CCl4 intoxicated animals showed
significant hepatoprotective activity by restoring the hepatocellular activity. It has
been reported that thymoquinone (the active component of black cumin) presents in
high level in N. sativa possesses antioxidant properties (Hesham et al., 2002).
Thymoquinone prevents the formation of toxin stable complex by the combination of
CCl3O2 free radical and the glycolipid component of cell membrane and therefore
restores cellular architecture and prevent the leakage of its enzymes (Ali BH et al.,
2003). Significant hepatoprotective effects of N. sativa in carbon tetrachloride
(Mastour and Al-Ghamdi et al., 2003), D-galactosamine and turpentine oil- induced
liver damage (Subodh et al., 2011) were noted.
This was also reported that N. sativa has a significant hepatoprotective effect
in CCl4 intoxicated rabbits. N. sativa protects liver against fibrosis possibly through
immunomodulator and antioxidant activities. N. sativa seed extract reduced total
bilirubin, serum enzymes level in CCl4 treated animals and also prevent liver fibrosis
and cirrhosis, suggested that N. sativa protects liver against fibrosis possibly through
immunomodulator and antioxidant activities. (Turkdogan et al., 2003). N. sativa
seeds appeared to be safe and possibly protective against CCL4-induced
hepatotoxicity (Mastour and Al-Ghamdi, 2003).

CHAPTER-5
Integral University, Lucknow 177
5.3 MATERIALS AND METHODS
5.3.1 Collection of N. sativa seeds
Seeds of N. sativa were procured in September, 2010 from a herbal
shop in Lucknow, India and authenticated by a botanist at National Botanical
Research Institute, Lucknow. A voucher specimen of the seeds was kept in the
museum of the Department for future reference.
5.3.2 Germination of N. sativa seeds
Germination was done according to the method of Ahmad et al., (2010). Seed
lots used for different experiments showed germination capacities ranging from 80 to
98%. The seeds were surface sterilized with 0.1% HgCl2 for 3 min. They were rinsed
thoroughly with double distilled water and soaked in de-ionized water for 30 min.
For germination of seeds, they were placed on four folds of damp filter paper at 25°C
and incubated in dark till the initiation of sprouting after which they were placed at a
light intensity of 100 µmol m-2
s-1
(that was measured by LI-190SA quantum Sensor,
Li-COR Co., USA) and a 14/10 h (day/night) photoperiod till the complete plantlet
with two leaves were obtained. The complete germination took eleven days with
emergence of epicotyl, hypocotyl, roots and green leaves. Germination, defined as 1
mm radicle emergence, was followed for 11days. No contamination by
microorganisms was observed during this time period.
5.3.3 Harvest of germinated seeds
The germinated seeds of different days were harvested with sterilized forceps
and were kept on blotting sheet to remove excess water. The germinated seeds
collected for different experiments were used immediately for preparing extracts.

CHAPTER-5
Integral University, Lucknow 178
Seeds were considered to be germinated after the radicle emerged from the testa. All
the samples were stored at -80oC in a deep freezer until used further.
5.3.4 Preparation of distilled extracts
The samples of seed and germinated phases 5th, 7
th and 11
th day were shade-
dried and ground to a fine powder. The powder (20gm) was extracted with 200 ml
methanol solvent for 48 h in order to extract bioactive compounds using soxhlet
apparatus (AOAC method 1980). The extracts were filtered using Whatman filter
paper (No.1) and methanol was evaporated using rotary distillation apparatus to
obtained pure extract. Oily fraction of extracts was stored at 40C until use.
5.3.5 Animals
Male Wistar rats, weighing 150 - 200 g, were purchased from Central Drug
And Research Institute (CDRI), Lucknow, India and housed in a temperature
controlled room (22±2°C) with a 12 hour light-12 hour dark cycle and allowed free
access to a standard rat chow and filtered tap water for 7 days for acclimatization.
The study received the approval of the Institutional Animal Ethics Committee
(IAEC) of Era’s Lucknow Medical College & Hospital. Animals were cared for in
accordance with the internationally accepted principles for laboratory animal use and
care and the procedures followed were in accordance with the standards set forth in
the Guide for the Care and Use of Laboratory Animals (published by the National
Academy of Science, National Academy Press, Washington, D.C.). They were
housed under controlled conditions of temperature of 23±20C, relative humidity of
30–70% and 12 h light–12 h dark cycle. The animals were housed individually in
polypropylene cages containing sterile paddy husk (procured locally) as bedding
throughout the experiment. All animals were fed with sterile commercial pelleted rat
chow supplied by Hindustan Lever Ltd. (Mumbai, India) and had free access to
water. Animals were kept under fasting for overnight and weighed before the
experiment.

CHAPTER-5
Integral University, Lucknow 179
5.3.6 Study design
Hepatoprotective effects of N. sativa methanolic extracts of different
germination stages were studied against carbon tetrachloride (CCl4) induced
hepatotoxicity in Wistar rats. Dose of N. sativa extracts was selected as per study of
Mehta et al., (2009). Silymarin (50 mg/kg b.w.) was used as standard
hepatoprotective drug (Nayak et al., 2012, Palanivel et al., 2008).
5.3.6.1 Carbon tetrachloride induced hepatotoxicity
Carbon tetrachloride (CCl4) toxicity was induced according to the method of
Asif Mir et al., 2010. Acute CCl4 toxicity was induced by treating rats for 24 hrs by
oral administration of CCl4 mixed with olive oil as vehicle in 1:1 ratio (3 ml/kg
b.w.).
5.3.6.2 Hepatoprotective effects of N. sativa
To study hepatoprotective effects of N. sativa during different phases of its
germination in carbon tetrachloride induced (CCl4) induced hepatotoxicity, animals
were divided in following groups containing 6 rats each.
Group-I (control) Received single dose of distilled water orally from day 1 to day 7
followed by olive oil (1.5ml/kg) on day 7.
Group-II (CCL4) Received distilled water for six days followed by CCl4 with olive
oil (3 ml/kg) orally on day 7 (Oral administration of CCl4 mixed
with olive oil as vehicle in 1:1 ratio (3 ml/kg b.w.)
Group-III Received Silymarin (50 mg/kg) orally from day 1 to day 7 followed by
CCl4 with olive oil (3ml/kg) on day 7 after 2 hrs.
Group- IV (Seed extract) Received N. sativa seed extract 1 g/kg orally from day 1
to day 7 followed by CCl4 with olive oil (3ml/kg) on day 7 after 2 hrs.
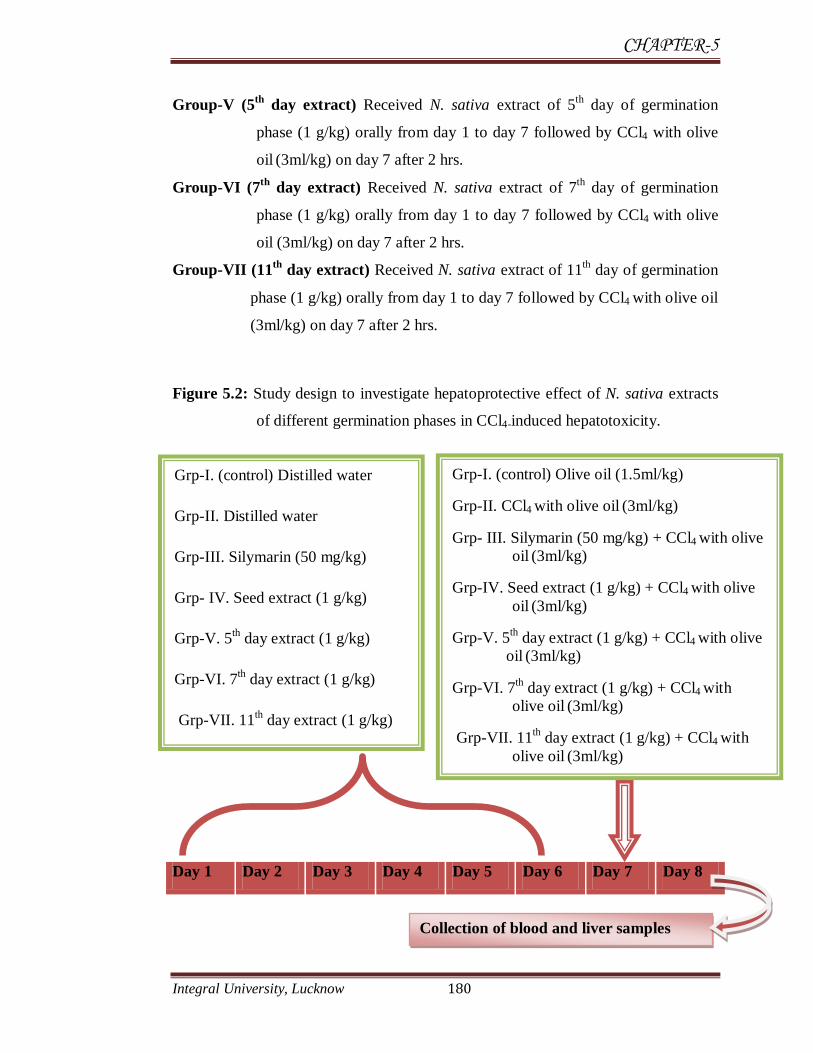
CHAPTER-5
Integral University, Lucknow 180
Group-V (5th
day extract) Received N. sativa extract of 5th
day of germination
phase (1 g/kg) orally from day 1 to day 7 followed by CCl4 with olive
oil (3ml/kg) on day 7 after 2 hrs.
Group-VI (7th
day extract) Received N. sativa extract of 7th
day of germination
phase (1 g/kg) orally from day 1 to day 7 followed by CCl4 with olive
oil (3ml/kg) on day 7 after 2 hrs.
Group-VII (11th
day extract) Received N. sativa extract of 11th day of germination
phase (1 g/kg) orally from day 1 to day 7 followed by CCl4 with olive oil
(3ml/kg) on day 7 after 2 hrs.
Figure 5.2: Study design to investigate hepatoprotective effect of N. sativa extracts
of different germination phases in CCl4-induced hepatotoxicity.
Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7 Day 8
Collection of blood and liver samples
Grp-I. (control) Distilled water
Grp-II. Distilled water
Grp-III. Silymarin (50 mg/kg)
Grp- IV. Seed extract (1 g/kg)
Grp-V. 5th day extract (1 g/kg)
Grp-VI. 7th day extract (1 g/kg)
Grp-VII. 11th day extract (1 g/kg)
Grp-I. (control) Olive oil (1.5ml/kg)
Grp-II. CCl4 with olive oil (3ml/kg)
Grp- III. Silymarin (50 mg/kg) + CCl4 with olive
oil (3ml/kg)
Grp-IV. Seed extract (1 g/kg) + CCl4 with olive
oil (3ml/kg)
Grp-V. 5th day extract (1 g/kg) + CCl4 with olive
oil (3ml/kg)
Grp-VI. 7th day extract (1 g/kg) + CCl4 with
olive oil (3ml/kg)
Grp-VII. 11th day extract (1 g/kg) + CCl4 with
olive oil (3ml/kg)

CHAPTER-5
Integral University, Lucknow 181
5.3.7 Blood sampling
Cardiac puncture (Diaphragmatic approach): General anesthesia was
administered and the animal placed on solid surface with its ventrum exposed
thexyphoid process was palpated at the caudal aspects of the animal sternum. A
notch was present on both side of this process. A 1.5O, 22 gauge needle attached to a
5 ml syringe was inserted into either notch and directed towards the heart as
determined by palpating the apex beat. Negative pressure was applied by placing
slight backward pull of the plunger, once it has been inserted beneath the skin.
Reflux of blood was apparent once the needle penetrated the heart.
Plate 5.1: Blood sampling by cardiac puncture and collection of liver by dissection.

CHAPTER-5
Integral University, Lucknow 182
5.3.8 Biochemical estimation
5.3.8.1 Liver enzymes
Estimation of serum levels of glutamate oxaloacetate transaminase (SGOT),
glutamate pyruvate transaminase (SGPT) and alkaline phosphatase (ALP). Total
bilirubin was measured by standard method using semi-auto analyzer.
5.3.8.2 Tissue Homogenate Preparation
10% (w/v) homogenate of rat liver was prepared with the aid of York’s
homogenizer fitted with Teflon plunger in 0.1 M phosphate buffer (pH 7.5). The
whole homogenate was first centrifuged at 2500xg for 10 minutes in a refrigerated
centrifuge. The pellet consisting of nuclear fraction and cell debris was discarded.
The supernatant was further centrifuged at 11,000xg for 15 min and mitochondrial
fraction was separated. The clear supernatant was further centrifuged at 100,000xg
for 90 min and the resulted supernatant was used for enzyme activities.
5.3.8.3 Lipid peroxide level (Ohkawa et al., 1979)
Procedure: 0.2 ml of each sub-cellular fraction having 3 to 10 mg protein,
was mixed with 1.0 ml of 20% acetic acid followed by the addition of 0.2 ml of 8%
aqueous SDS. The mixture was adjusted to pH 4 by addition of concentrated NaOH
solution if needed. After adjusting the pH of reaction mixture, 1.5 ml of 0.8% TBA
solution and sufficient amount of distilled water was also added to achieve the final
volume of 4 ml. The reaction mixture was incubated in a boiling water bath for
1hour. After cooling to room temperature, 3 ml of n-butanol was mixed. The reaction
mixture was centrifuged at 10,000 x g for 15 min. A clear butanol fraction obtained
from centrifugation was used for measuring the absorbance at 532 nm.
Calculation: An appropriate standard made up of malondialdehyde (MDA)
2.5 nmol was run simultaneously. Result was expressed in nmol MDA/gm.

CHAPTER-5
Integral University, Lucknow 183
5.3.8.4 Superoxide dismutase (McCord Fridovich, 1969)
Procedure: 2 ml of homogenate was dispended in centrifuge tube. The tube
was placed in a refrigerated centrifuge and spin at 10,000x g for 15 min. To the
supernatant for each sample, 313mg/ml ammonium sulphate was added to the final
concentration of 50%. The tube was shaken thoroughly and kept for 4 hours in cold
(40C). Thereafter, the tube was centrifuged at 14,000x g for 30 min at 4
0C. The
supernatant sample was dialyzed against cold tripled distilled water with 3 changes,
each change after 3 hrs interval. The content of dialysis bags were subsequently used
as enzyme source. Experimental tubes was contain 0.3 ml of 1.5mM
nitrobluetetrazolium (NBT), 0.2 ml of 0.93 mM phenazinemethosulphate, 1 ml of
20.4mM pyrophosphate buffer (pH 9.2), 1 ml triple distilled water and 0.2ml enzyme
source. The second setup reference tubes received the above reagents except the
enzyme source. The reaction was started simultaneously in two sets by the addition
of 0.1 ml 2.34mM NADH. After an interval of 90 sec, 1 ml glacial acetic acid was
added to each tube for checking the reaction then 0.2 ml enzyme source was added
for checking the reaction followed by addition of 0.2 ml enzyme source in the
reference tubes. The absorbance of these tubes was read at 560 nm in a
spectrophotometer against blank (NBT+PMS+Buffer+TDW).
Calculation: The unit of enzyme activity was defined as the amount of
enzyme required to inhibit the reduction in optical density of nitro blue tetrazolium
up to 50%, in 1 min at 560 nm under the assay condition. Results were expressed as
unit/mg protein
5.3.8.5 Catalase (Aebi, 1974)
Procedure: 3.0 ml of H2O2 phosphate buffer was pipette into the cuvette
followed by addition of required amount of tissue supernatant (cytosolic fraction) as
enzyme source and the content was mixed thoroughly. The decrease in absorbance at

CHAPTER-5
Integral University, Lucknow 184
240 nm recorded after every 30 sec for 3 min using UV spectrophotometer. Result
was expressed as unit/mg protein.
5.3.8.6 Reduced glutathione (Eliman et al., 1959)
Procedure: 1 ml liver (10%) homogenate was deprotenized by adding 1 ml
of 10% TCA and centrifuged at 6000x g for 5 min. 0.5 ml aliquots for supernatant
was mixed with 0.5 ml of double distilled water thereafter 2 ml of 0.4 M Tris buffer
and 0.1 ml DTNB were added to it with proper stirring. The absorbance was read at
412 nm within 5 min of the addition of DNTB.
Calculation: GSH in the sample was calculated using the standard curve and
results were expressed as µmol/g tissue.
5.3.9 Histopathological study
For histopathology half of liver tissue of each rat was stored in 10% formalin
solution for 48 hours or until processing. For block preparation, liver were processed
using a graded ethanol series and embedded in paraffin. 4 µm paraffin sections were
cut and stained with Haematoxylin and eosin. The study was conducted in
consultation with the trained pathologist under light microscope with 10x, 40x, and
100x magnifications.
Tissue fixation, embedding & sectioning
Tissue Fixation
The rats were sacrificed after the completion of the period of the dose and the
organs were collected in normal saline. The tissue was cut in small pieces
(L.Section/T.Section, approx 2.3 mm in size). The tissue was fixed in 10% formalin
for 24 hours and washed in running tap-water after that tissue was kept in distilled

CHAPTER-5
Integral University, Lucknow 185
water for 10-15 minutes. Tissue was dehydrated in following graded series of
alcohol.
30% alcohol for 45 minutes.
50% alcohol for 45 minutes.
70% alcohol for 45 minutes.
90% alcohol for 45 minutes.
100% alcohol for 45 minutes.
Then, this tissue was transferred in absolute alcohol + xylene (1:1) for 35 minutes
and then after in pure xylene for 30 minutes.
Following chemicals are transferred in incubator at 60°C
Xylene + wax in 1:1 ratio for 45 minutes and pure wax for 45 minutes.
Tissue Embedding
The processed tissues were then transfer to the molten wax poured into the L-
block at 60°C, allowed to solidify for 10-15 minutes and removed from the L-block.
The casted wax blocks were left overnight at room-temperature to uniform
solidification. (Precautions were taken to minimize the appearance of air bubbles in
the blocks and to maintain the orientation of the tissue for transverse section (TS) or
longitudinal section (LS) while placing it into the molten wax in L-molds).
Section Cutting
With the help of microtome the section of the tissue has been cut (2-5 µm in
size). The thin section has been taken in clean slide coated with a mixture of egg
albumin + glycerol (1:1). Then it was unfolded with slight warm distilled water, then
heat fixed and the slides were kept for 12 hours.

CHAPTER-5
Integral University, Lucknow 186
Slide Staining
The tissue was immersed in xylene for 10 min.
Immersed in 100% alcohol for 10 min.
Immersed in 90% alcohol for 10 min.
Immersed in 70% alcohol for 10 min.
Immersed in 50% alcohol for 5-8 min.
Immersed in 30% alcohol for 5-8 min.
Immersed in distilled water for 5 min.
Immersed in Haematoxylin for 2-3 min.
Immersed in tap water for 10-15 min.
Immersed in acid water 3-5 min.
Immersed in distilled water for 5 min.
Immersed in 30% alcohol for 5-8 min.
Immersed in 50% alcohol for 5-8 min.
Immersed in 70% alcohol for 10 min.
Immersed in acid for 3-5 min.
Immersed in 90% alcohol for 15 min.
Immersed in 100% alcohol for 15 min.
Immersed in xylene for 5-10 min.
Then mounted on the slide with DPX.
Then left for 12 hours & seen under microscope.
CHAPTER-5
Integral University, Lucknow 187
5.3.10 Statistical analysis
Statistical significance was determined by One Way Analysis of Variance
(ANOVA) followed by Dunnet’s t-test to compare group means. The level of
significance was P < 0.001.
5.4 RESULTS AND DISCUSSION
5.4.1 Hepatoprotective effect of N. sativa extracts of different germination
phases against CCl4 induced hepatotoxicity.
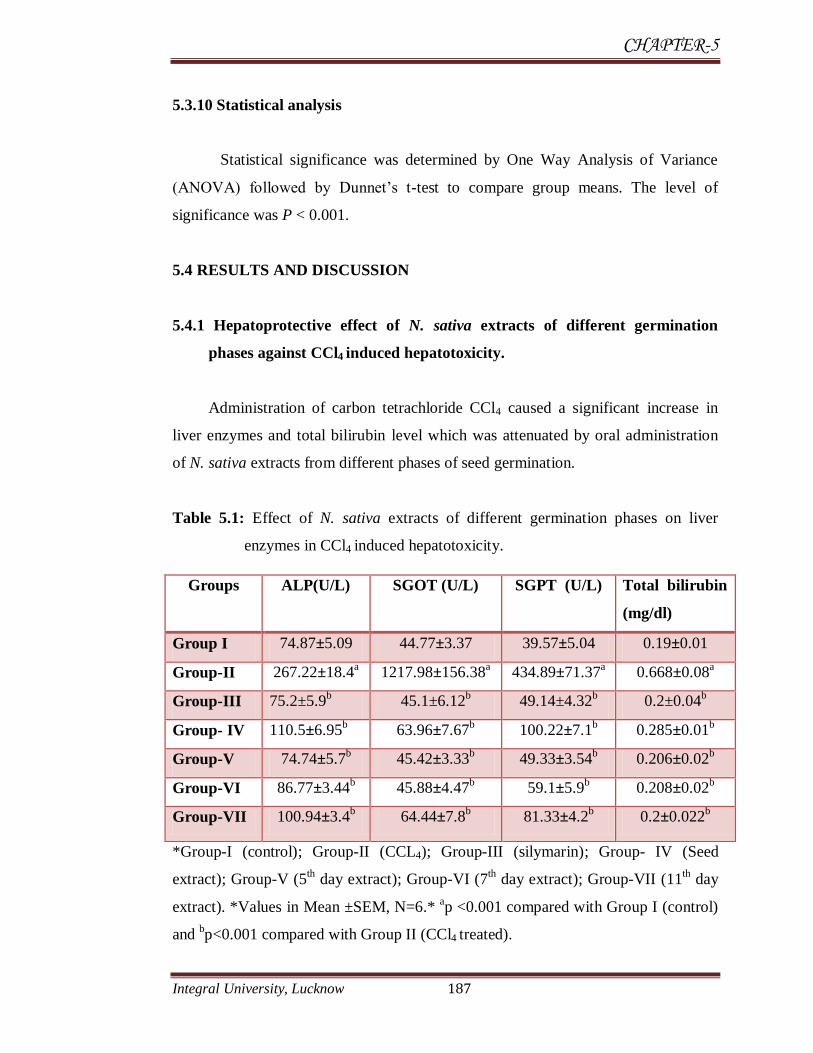
Administration of carbon tetrachloride CCl4 caused a significant increase in
liver enzymes and total bilirubin level which was attenuated by oral administration
of N. sativa extracts from different phases of seed germination.
Table 5.1: Effect of N. sativa extracts of different germination phases on liver
enzymes in CCl4 induced hepatotoxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract). *Values in Mean ±SEM, N=6.* ap <0.001 compared with Group I (control)
and bp<0.001 compared with Group II (CCl4 treated).
Groups ALP(U/L) SGOT (U/L) SGPT (U/L) Total bilirubin
(mg/dl)
Group I 74.87±5.09 44.77±3.37 39.57±5.04 0.19±0.01
Group-II 267.22±18.4a 1217.98±156.38
a 434.89±71.37
a 0.668±0.08
a
Group-III 75.2±5.9b 45.1±6.12
b 49.14±4.32
b 0.2±0.04
b
Group- IV 110.5±6.95b 63.96±7.67
b 100.22±7.1
b 0.285±0.01
b
Group-V 74.74±5.7b 45.42±3.33
b 49.33±3.54
b 0.206±0.02
b
Group-VI 86.77±3.44b 45.88±4.47
b 59.1±5.9
b 0.208±0.02
b
Group-VII 100.94±3.4b 64.44±7.8
b 81.33±4.2
b 0.2±0.022
b

CHAPTER-5
Integral University, Lucknow 188
5.4.1.1 ALP
Results of present study showed that level of ALP in control group was
74.87±5.09 U/L serum. There was a significant (P<0.001) increase in mean ALP
level in rats administered with CCl4 (3ml/kg) for 24 hrs (267.22±18.4 U/L serum).
Oral administration of N. sativa seed extract (non-germinated) and extracts of
different germination phases (5th day, 7
th day and 11
th day) ameliorated this increase
in ALP level. In all the four groups receiving N. sativa extracts rise in ALP level was
prevented (Table 5.1 and Figure 5.3). Among all extracts 5th
day germination extract
showed significant protective effect and reduced serum ALP level (74.74±5.7 U/L
serum) which was equal to control group (74.87±5.09 U/L serum) and the standard
drug silymarin (75.2±5.9 U/L serum). Extract of 7th day germination was on second
position (86.77±3.44 U/L serum) followed by 11th day extract (100.94±3.4 U/L
serum) and seed extract (110.5±6.95 U/L serum).
Figure 5.3: Effect of N. sativa extracts of different germination phases on ALP level
against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
0
50
100
150
200
250
300
Group I Group II Group III Group IV Group V Group VI Group VII
AL
P (
U/L
ser
um
)

CHAPTER-5
Integral University, Lucknow 189
5.4.1.2 SGOT
Table 5.1 and Figure 5.4 showed that administration of CCl4 (3ml/kg) for 24
hrs significantly (P<0.001) increase SGOT level as compared to control. The SGOT
value of CCl4 group (1217.98±156.38 U/L serum) was very high than control group
(44.77±3.37 U/L serum). Administration of N. sativa seed extract (non-germinated)
and extracts from different germination phases of seed (5th day, 7
th day and 11
th day)
ameliorated the increase in SGOT level due to CCl4 toxicity. Among all extracts 5th
day and 7th day germination extract showed significant protective effect against
CCl4-induced toxicity and reduced serum SGOT level (45.42±3.33 U/L serum and
45.88±4.47 U/L serum respectively) which was significantly (P<0.001) equal to
control group (44.77±3.37 U/L serum) as well as standard group (45.1±6.12 U/L
serum) followed by 11th
day extract (64.44±7.8 U/L serum) and seed extract
(63.96±7.67 U/L serum).
Figure 5.4: Effect of N. sativa extracts of different germination phases on SGOT
level against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
0
200
400
600
800
1000
1200
1400
1600
Group I Group II Group III Group IV Group V Group VI Group VII
SG
OT
(U
/L s
eru
m)
CHAPTER-5
Integral University, Lucknow 190
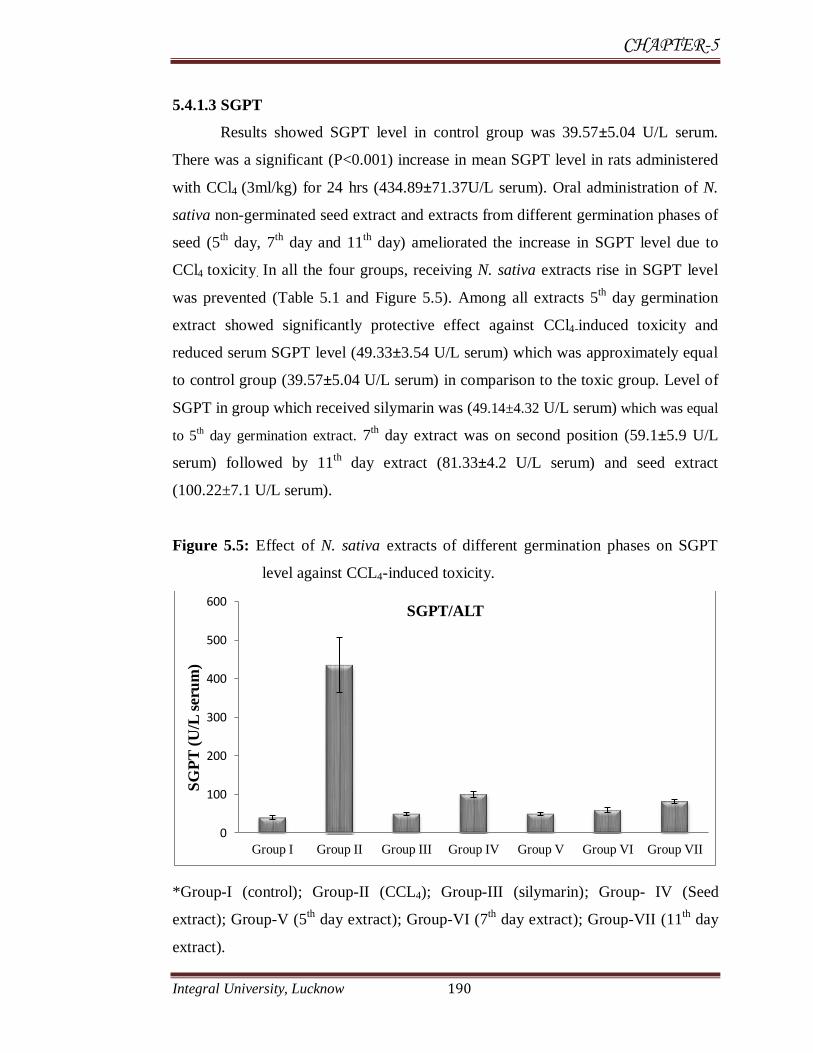
5.4.1.3 SGPT
Results showed SGPT level in control group was 39.57±5.04 U/L serum.
There was a significant (P<0.001) increase in mean SGPT level in rats administered
with CCl4 (3ml/kg) for 24 hrs (434.89±71.37U/L serum). Oral administration of N.
sativa non-germinated seed extract and extracts from different germination phases of
seed (5th day, 7
th day and 11
th day) ameliorated the increase in SGPT level due to
CCl4 toxicity. In all the four groups, receiving N. sativa extracts rise in SGPT level
was prevented (Table 5.1 and Figure 5.5). Among all extracts 5th day germination
extract showed significantly protective effect against CCl4-induced toxicity and
reduced serum SGPT level (49.33±3.54 U/L serum) which was approximately equal
to control group (39.57±5.04 U/L serum) in comparison to the toxic group. Level of
SGPT in group which received silymarin was (49.14±4.32 U/L serum) which was equal
to 5th day germination extract. 7
th day extract was on second position (59.1±5.9 U/L
serum) followed by 11th
day extract (81.33±4.2 U/L serum) and seed extract
(100.22±7.1 U/L serum).
Figure 5.5: Effect of N. sativa extracts of different germination phases on SGPT
level against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
0
100
200
300
400
500
600
Group I Group II Group III Group IV Group V Group VI Group VII
SG
PT
(U
/L s
eru
m)
SGPT/ALT

CHAPTER-5
Integral University, Lucknow 191
5.4.1.4 Total bilirubin
Mean value of total bilirubin in control group was 0.19±0.01 (mg/dl serum).
There was a significant (P<0.001) increase in value of total bilirubin level in rats
administered with CCl4 (3ml/kg) for 24 hrs (0.668±0.08 mg/dl serum).
Administration of N. sativa seed extract (non-germinated) and extracts from different
germination phases (5th day, 7
th day and 11
th day) ameliorated the increase in total
bilirubin level due to CCl4 toxicity. In all the four groups receiving N. sativa extracts
rise in total bilirubin level was prevented (Table 5.1 and Figure 5.6). 7th day extract
was on second position followed by 11th
day extract and seed extract to ameliorate
the level of bilitubin.
Figure 5.6: Effect of N. sativa extracts of different germination phases on bilirubin
level against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Group I Group II Group III Group IV Group V Group VI Group VII
Bil
iru
bin
(m
g/d
l se
rum
)
BILIRUBIN(TOTAL)
CHAPTER-5
Integral University, Lucknow 192
5.4.2 Effect of N. sativa extracts of different germination phases on antioxidant
enzymes level against CCL4-induced hepatotoxicity.
Carbon tetrachloride (CCl4) administration caused a significant oxidative stress
as evident by increase in MDA level and decrease in SOD, CAT and GSH levels.
This altered redox status was attenuated by administration of N. sativa extracts of
different phases of seed germination.
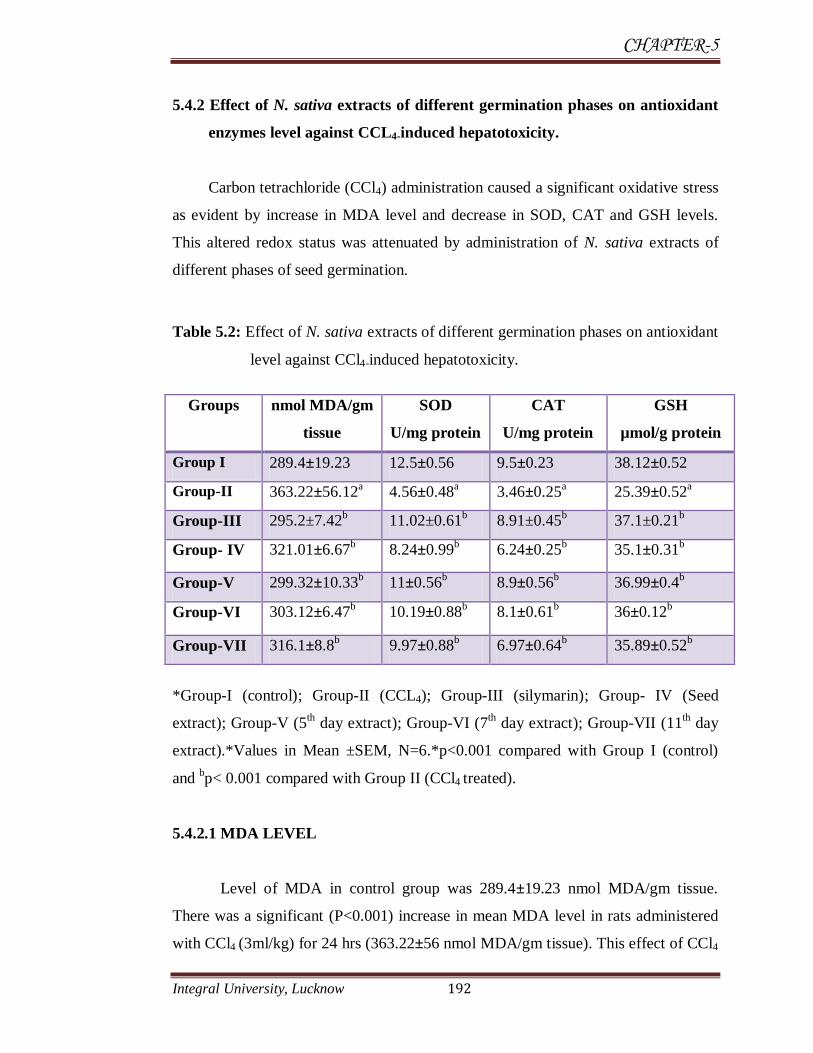
Table 5.2: Effect of N. sativa extracts of different germination phases on antioxidant
level against CCl4-induced hepatotoxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).*Values in Mean ±SEM, N=6.*p<0.001 compared with Group I (control)
and bp< 0.001 compared with Group II (CCl4 treated).
5.4.2.1 MDA LEVEL
Level of MDA in control group was 289.4±19.23 nmol MDA/gm tissue.
There was a significant (P<0.001) increase in mean MDA level in rats administered
with CCl4 (3ml/kg) for 24 hrs (363.22±56 nmol MDA/gm tissue). This effect of CCl4
Groups nmol MDA/gm
tissue
SOD
U/mg protein
CAT
U/mg protein
GSH
µmol/g protein
Group I 289.4±19.23 12.5±0.56 9.5±0.23 38.12±0.52
Group-II 363.22±56.12a 4.56±0.48
a 3.46±0.25
a 25.39±0.52
a
Group-III 295.2±7.42b 11.02±0.61
b 8.91±0.45
b 37.1±0.21
b
Group- IV 321.01±6.67b 8.24±0.99
b 6.24±0.25
b 35.1±0.31
b
Group-V 299.32±10.33b 11±0.56
b 8.9±0.56
b 36.99±0.4
b
Group-VI 303.12±6.47b 10.19±0.88
b 8.1±0.61
b 36±0.12
b
Group-VII 316.1±8.8b 9.97±0.88
b 6.97±0.64
b 35.89±0.52
b

CHAPTER-5
Integral University, Lucknow 193
toxicity was ameliorated in rats which received N. sativa extracts from different
germination phases of seed. In all the four groups receiving N. sativa extracts rise in
MDA level was prevented (Table 5.2 and Figure 5.7). Among all extracts 5th day
germination extract showed significant protective effect against CCl4-induced
toxicity and reduced MDA level as control (299.32±10.33 nmol MDA/gm tissue).
Silymarin also reduced MDA level (295.2±7.42 nmol MDA/gm tissue) significantly.
7th day, 11
th day and seed extract (non-germinated) also reduced the MDA level.
Figure 5.7: Effect of N. sativa extracts of different germination phases on MDA
level against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
5.4.2.2 Superoxide dismutase
There was a significant (P<0.001) decrease in SOD level observed in rats
administered with CCl4 (4.56±0.48 U/mg) in comparison to control group (12.5±0.56
U/mg). Extracts of N. sativa raise the level of superoxide dismutase enzyme and
maintain the level of catalase as in control group. The mean value of 5th day, 7
th day,
11th day and seed extract was 11±0.568 U/mg, 10.19±0.88 U/mg, 9.97±0.88 U/mg
0
50
100
150
200
250
300
350
400
450
Group I Group II Group III Group IV Group V Group VI Group VII
MD
A (
nm
ol/
gm
tis
sue)

CHAPTER-5
Integral University, Lucknow 194
and 8.24±0.99 U/mg respectively (Table 5.2 and Figure 5.8). Results showed that 5th
day germinated extract significantly increased the level of SOD. Extract of 7th
day
germination phase was on second position followed by 11th
and seed extract (non-
germinated).
Figure 5.8: Effect of N. sativa extracts of different germination phases on SOD level
against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
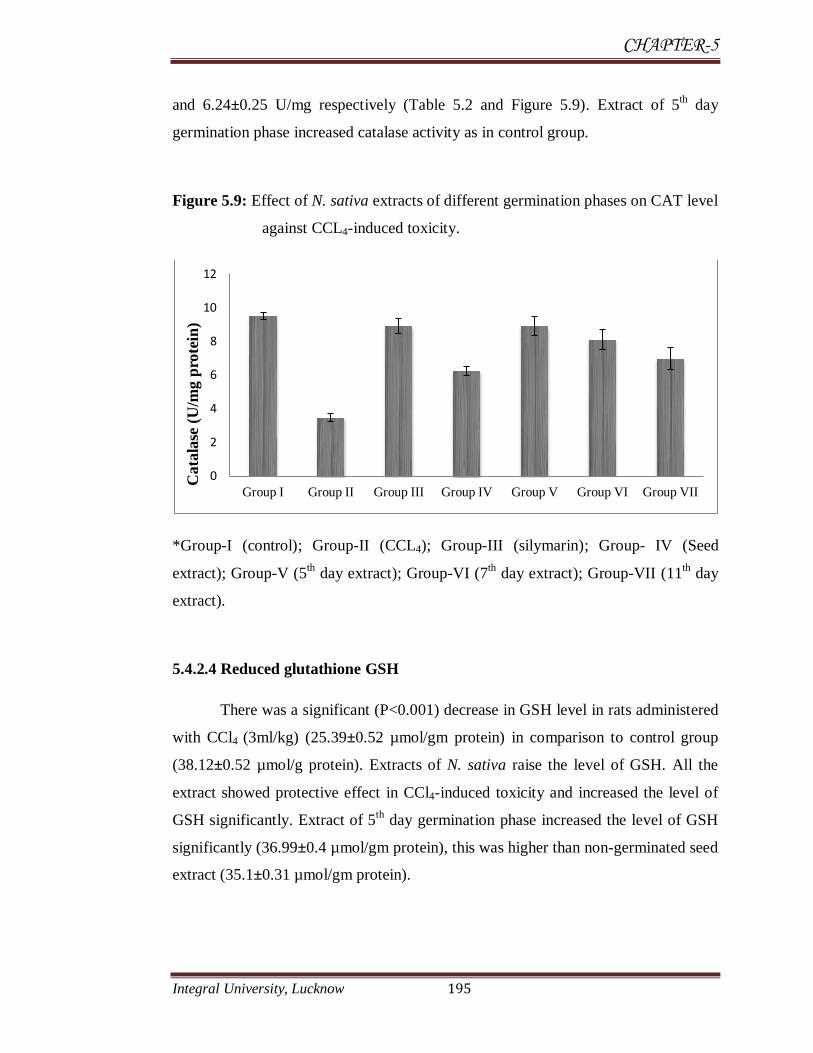
5.4.2.3 Catalase
There was a significant (P<0.001) decrease in CAT enzyme level in rats
administered with CCl4 (3ml/kg) for 24 hrs (3.46±0.25 U/mg) in comparison to
control group (9.5±0.23 U/mg). Extracts of N. sativa from germination phases of
seed raise the level of catalase and maintain the level of catalase as was in control
group. The mean value of silymarin, 5th
day, 7th
day, 11th day and seed extract (non-
germinated) was 8.91±0.45 U/mg, 8.9±0.56 U/mg, 8.1±0.61 U/mg, 6.97±0.64 U/mg
0
2
4
6
8
10
12
14
Group I Group II Group III Group IV Group V Group VI Group VII
SO
D (
U/m
g p
rote
in)
CHAPTER-5
Integral University, Lucknow 195
and 6.24±0.25 U/mg respectively (Table 5.2 and Figure 5.9). Extract of 5th day
germination phase increased catalase activity as in control group.
Figure 5.9: Effect of N. sativa extracts of different germination phases on CAT level
against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
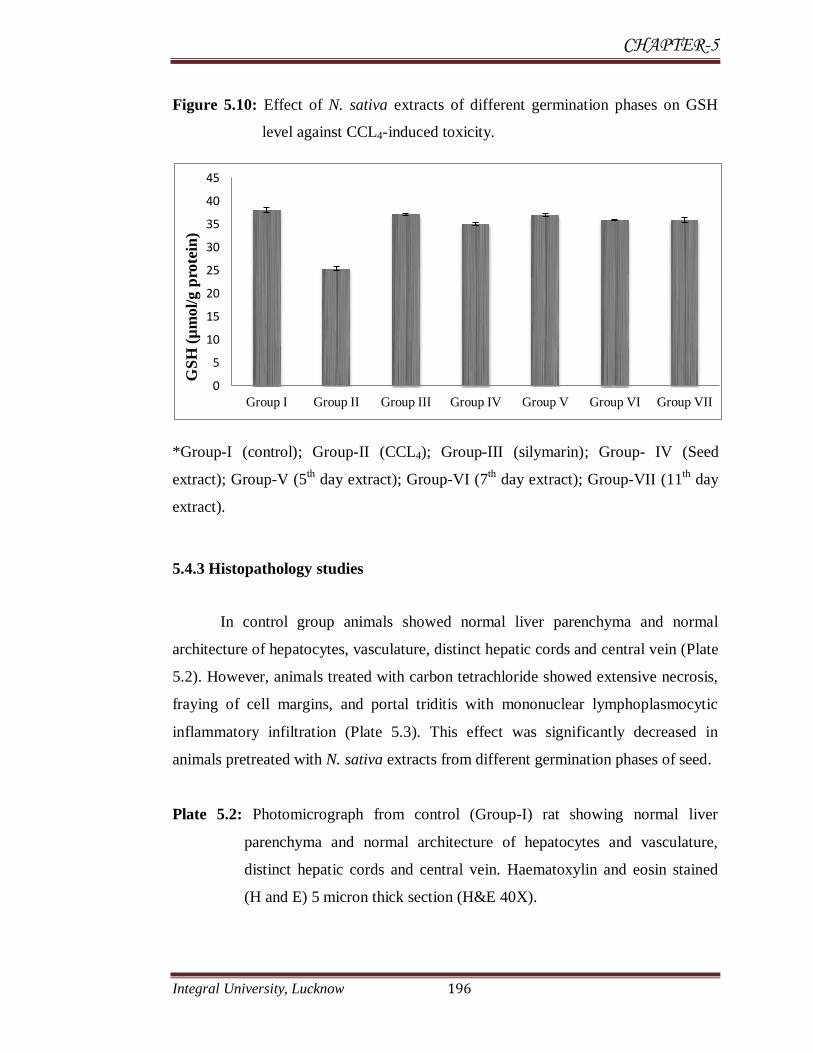
5.4.2.4 Reduced glutathione GSH
There was a significant (P<0.001) decrease in GSH level in rats administered
with CCl4 (3ml/kg) (25.39±0.52 µmol/gm protein) in comparison to control group
(38.12±0.52 µmol/g protein). Extracts of N. sativa raise the level of GSH. All the
extract showed protective effect in CCl4-induced toxicity and increased the level of
GSH significantly. Extract of 5th
day germination phase increased the level of GSH
significantly (36.99±0.4 µmol/gm protein), this was higher than non-germinated seed
extract (35.1±0.31 µmol/gm protein).
0
2
4
6
8
10
12
Group I Group II Group III Group IV Group V Group VI Group VII
Cata
lase
(U
/mg p
rote
in)
CHAPTER-5
Integral University, Lucknow 196
Figure 5.10: Effect of N. sativa extracts of different germination phases on GSH
level against CCL4-induced toxicity.
*Group-I (control); Group-II (CCL4); Group-III (silymarin); Group- IV (Seed
extract); Group-V (5th day extract); Group-VI (7
th day extract); Group-VII (11
th day
extract).
5.4.3 Histopathology studies
In control group animals showed normal liver parenchyma and normal
architecture of hepatocytes, vasculature, distinct hepatic cords and central vein (Plate
5.2). However, animals treated with carbon tetrachloride showed extensive necrosis,
fraying of cell margins, and portal triditis with mononuclear lymphoplasmocytic
inflammatory infiltration (Plate 5.3). This effect was significantly decreased in
animals pretreated with N. sativa extracts from different germination phases of seed.
Plate 5.2: Photomicrograph from control (Group-I) rat showing normal liver
parenchyma and normal architecture of hepatocytes and vasculature,
distinct hepatic cords and central vein. Haematoxylin and eosin stained
(H and E) 5 micron thick section (H&E 40X).
0
5
10
15
20
25
30
35
40
45
Group I Group II Group III Group IV Group V Group VI Group VII
GS
H (
µm
ol/
g p
rote
in)

CHAPTER-5
Integral University, Lucknow 197
Hepatic cord
Central vein
Plate 5.3: Photomicrograph of CCl4 treated rats (Group-II) showing extensive
necrosis, fraying of cell margins, portal triditis with mononuclear
lymphoplasmocytic inflammatory infiltration, strip focal necrosis, feathery
degeneration (in the mid zone) involving several hepatocytes, necrosis of several
hepatocyte extended in band like fashion in the mid zonal area and occasional
apoptotic bodies.

CHAPTER-5
Integral University, Lucknow 198
Plate 5.4: Liver section from rat orally treated with seed extract (non-germinated
1g/kg) showing a slight protective effect but shows some
lymphoplasmocytic inflammatory infiltration.
Plate 5.5: Liver section from rat orally treated with 5th
day germination extract (1
g/kg), showing highly protective effect without lymphoplasmocytic
inflammatory infiltration
CHAPTER-5
Integral University, Lucknow 199
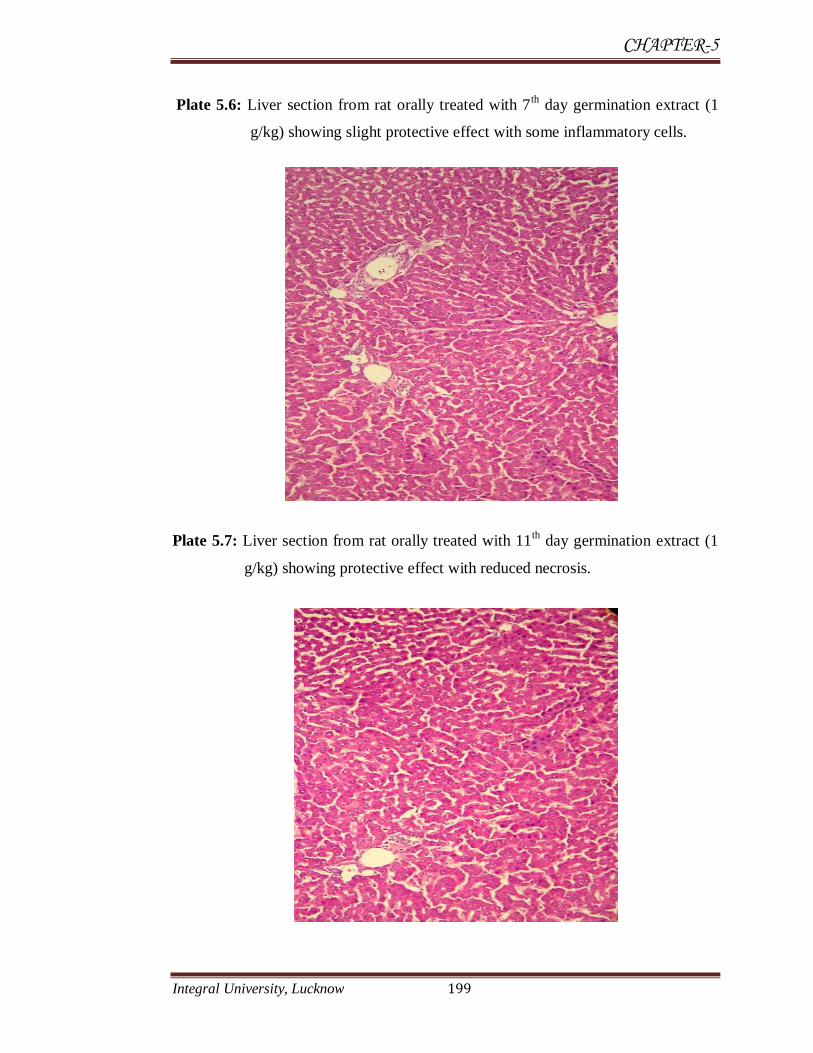
Plate 5.6: Liver section from rat orally treated with 7th
day germination extract (1
g/kg) showing slight protective effect with some inflammatory cells.
Plate 5.7: Liver section from rat orally treated with 11th
day germination extract (1
g/kg) showing protective effect with reduced necrosis.

CHAPTER-5
Integral University, Lucknow 200
A number of chemicals including various environmental toxicants and
clinically useful drugs can cause severe cellular damages in different organs of our
body through the metabolic activation to highly reactive substances such as free
radicals. CCl4 is one of such widely studied environmental toxicant (Sil et al., 2006).
Up to the present time the etiology and treatment of most liver diseases are not
known. The liver is the commonest site affected during the toxic manifestation of
many drugs. Toxicity in liver due to CCl4 and other chemicals is attributed to the
toxic metabolites formed which are responsible for the initiation of CCl4 dependent
lipid peroxidation (Arulkumaran et al., 2007). In the liver CCl4 is metabolized by the
cytochrome P450-dependent monooxygenase systems followed by its conversion to
more chemically active form namely trichloromethyl radical (CCl3•-
) that bind
covalently to the macromolecules and induce peroxidative degradation of membrane
lipids of endoplasmic reticulum rich in polyunsaturated fatty acids which leads to the
formation of lipid peroxides. This lipid peroxidative degradation of biomembranes is
one of the principle causes of hepatotoxicity of CCl4 (Kaplowitz et al., 1986). CCL4
also causes depletion of glutathione content and reduced the level of SOD and CAT
enzymes (Kamiyama et al., 1993).
In the assessment of liver damage by CCl4 the determination of enzyme
levels such as SGOT (AST), SGPT (ALT) and bilirubin is largely used. Necrosis or
membrane smash up releases the enzyme into circulation and hence they can be
measured in the serum. Increase levels of SGOT indicate liver damage in the same
manner caused by viral hepatitis as well as cardiac infarction and muscle injury. This
enzyme catalyses the conversion of alanine to pyruvate and glutamate and is released
in a similar manner. Therefore SGPT is more specific to the liver and is a better
parameter for detecting liver injury. Elevated levels of serum enzymes are indicative
of cellular leakage and loss of functional integrity of cell membrane in liver
(Drotman et al., 1978). On the other hand serum ALP, bilirubin and total protein
levels are related to the function of hepatic cell. Increase level of ALP in serum is
due to increased synthesis in presence of increasing biliary pressure (Muriel et al.,
1992).

CHAPTER-5
Integral University, Lucknow 201
As the results showed that oral administration of CCl4 caused a significant
(P<0.001) elevation of enzyme levels such as ALP, SGPT, SGOT, total bilirubin and
decrease in antioxidant system increased in MDA level and depletion of glutathione
content, SOD and CAT activity when compared to control group. There was a
significant (P<0.001) restoration of these enzyme levels on oral administration of the
N. sativa extracts from different germination phases in day-dependant manner and
also by silymarin at a dose of 50 mg/kg. Among all tested extracts 5th
day
germination extract showed significant protective effect like standard drug silymarin.
Extract of 7th
day germination was on second position followed by 11th
day and seed
extract. The reversal of increased serum enzymes in CCl4-induced liver damage by
the extract may be due to the prevention of the leakage of intracellular enzymes by
its membrane stabilizing activity. This is in agreement with the commonly accepted
view that serum levels of transaminases returns to normal with the healing of hepatic
parenchyma and the regeneration of hepatocytes (Thabrew et al., 1987). Effective
control of ALP and bilirubin levels point towards an early improvement in the
secretary mechanism of the hepatic cells.
The worth of any hepatoprotective drug is reliant on its capability of either
reducing the harmful effect or restoring the normal hepatic physiology that has been
disturbed by a hepatotoxin. Silymarin, the standard hepatoprotective drug used in the
present study exerted marked protective effects against CCl4-induced liver injury. It
has been reported in several in vitro and in vivo studies (Letteron et al., 1990). The
crucial protective mechanism of silymarin is an inhibition of lipid peroxidation by its
free radical scavenging properties (Farghali et al., 2000). Silymarin prevented CCl4
induced lipid peroxidation and hepatotoxicity in mice by decreasing the metabolic
activation of CCl4 and by acting as a chain-breaking antioxidant (Letteron et al.,
1990). Both silymarin and the N. sativa extracts of germination phases (specially of
5th day germination extract) decreased CCl4 induced elevated enzyme levels in tested
groups indicating the protection of structural integrity of hepatocyte cell membrane
or regeneration of damaged liver cells as they showed in histopathology results. The
increase in MDA (LPO) level in liver induced by CCl4 suggests enhanced lipid

CHAPTER-5
Integral University, Lucknow 202
peroxidation leading to tissue damage and failure of antioxidant defense mechanism
to prevent formation of excessive free radicals. Treatment with N. sativa
significantly reverses these changes. Hence it is likely that the mechanism of
hepatoprotection of N. sativa is due to its antioxidant effect. N. sativa reduced lipid
peroxidation significantly during germination phases by scavenging free hydroxyl
radical was reported in our previous research (Kamal et al., 2010).
Superoxide dismutase (SOD) has been reported as one of the most important
enzymes in the enzymatic antioxidant defense system which scavenges the
superoxide anion to form hydrogen peroxide and thus diminishing the toxic effect
caused by this radical. Decrease in enzyme activity of SOD is a sensitive index in
hepatocellular damage and is the most sensitive enzymatic index in live injury
(Curtis et al., 1972). Catalase (CAT) is an enzymatic antioxidant widely distributed
in all animal tissues and the highest activity is found in the red blood cells and liver.
CAT decomposes hydrogen peroxide and protects the tissues from highly reactive
hydroxyl radicals (Chance et al., 1992). N. sativa extracts from germination phases
cause a significant (P<0.001) increase in hepatic SOD and CAT activity and thus
reduces reactive free radical induced oxidative damage to liver. Previous research
has been proved that N. sativa shows significant high antioxidant activity during
germination phases as compared non-germinated seed (Kamal et al., 2010).
Glutathione is one of the most abundant tripeptide, non-enzymatic biological
antioxidant present in the liver. It removes free radical species such as hydrogen
peroxide, superoxide radicals and maintains membrane protein thiols. Also it is
substrate for glutathione peroxidase (Prakash et al., 2001). Decreased level of GSH
is associated with an enhanced lipid peroxidation in CCl4 treated rats. Administration
of N. sativa significantly (P<0.001) increased the level of GSH especially in 5th
day
germination extract treated group.
Many studies have demonstrated hepatoprotective effect of N. sativa seeds
and reported significant improvement in liver functional test. After giving N. sativa
oil significant improvement of deranged liver functional test parameters against

CHAPTER-5
Integral University, Lucknow 203
various hepatotoxic compounds (Mohideen et al., 2003; Al-Ghamdi et al., 2003;
Kanter et al., 2005; El-Gharieb et al., 2010; Janakat et al., 2010). N. sativa
administration in several model of induced hepatotoxicity has been shown to exert
hepatoprotective effect along with restoration of reduced glutathione, superoxide
dismutase, lactate dehydrogenase, catalase and decrease in lipid per oxidation
reaction (Al-Ghamdi et al., 2003; Mahmoud et al., 2002; Kanter et al., 2005; El-
Gharieb et al., 2010). Present study is the first study to investigate hepatoprotective
effect of N. sativa during germination phases of seed. As results showed that N.
sativa possesses better hepatoprotective effect during germination phases (especially
during 5th day) than non-germinated seed.
Protective effect of N. sativa may be due to the presence of high amount of
bioactive compounds such as thymoquinone (TQ), thymol, carvacrol and other
phytochemicals like flavonoids, phenols and tannins. Both biochemical and
histopathological data showed that there was no significant difference in extract
treatment when compared to control group. Extract of 5th day germination phase was
highly protective among all the extracts due to presence of higher quantity of
secondary metabolites, TQ, thymol and other active constituents were higher in 5th
day germination extract (chapter 2).
TQ has been reported to have potent superoxide anion (O2−) scavenging
abilities and to inhibit iron-dependent microsomal lipid peroxidation (Badary et al.,
2003). TQ may protect against CCl4-induced hepatotoxicity by a combination of two
mechanisms, i.e. TQ antioxidant potential (Houghton et al., 1995) and a previous
report of the quinone to have in vitro and in vivo superoxide anion radical
scavenging ability (Nagi and Mansour, 2000). TQ provided good protection against
lipid peroxidation and the oxidative damage caused by several toxic agents, as in
cisplatin nephrotoxicity (Badary et al., 1997), doxorubicin cardiotoxicity (Al-
Shabanah et al., 1998) and benzo(a)pyrene-induced forestomach carcinogenesis
(Badary et al., 1999). Burits and Bucar (2000) found that TQ had a scavenger effect
against the OH˙ radical in vitro. Moreover, Houghton et al. (1995) reported that TQ

CHAPTER-5
Integral University, Lucknow 204
had potent anti-inflammatory and inhibitory effects on non-enzymatic peroxidation
of brain phospholipid liposomes. This was also reported by Abdulrahman L. Al-
Malki (2010) that pretreatment with thymol ameliorates the deleterious effect of
CCl4. It was speculated that thymol exert their effects by decreasing lipid
peroxidation and enhancing the activities of antioxidant enzymes. For this reason,
thymol could be used as hepatoprotective agent with free medication side effects
(Abdulrahman L. Al-Malki, 2010). Carvacrol, another constituent of N. sativa
affords significant hepatoprotective and antioxidant effect against D-GalN-induced
hepatotoxicity rats (Baser et al., 2008; Aristatile et al., 2009). The protective effects
of alpha-Hederin (constituent of N. sativa) on carbon tetrachloride-induced
hepatotoxicity were also observed by Jeong et al., (1998). N. sativa seed have been
shown to significantly decrease TNF-α (tumor necrosis factor α), IFN-γ (interferon
γ), IL-β (interleukin β) in CCl4-induced hepatotoxicity (Al-Ghamdi, 2003). These
hepatoprotective effects were probably due to antioxidant activities of N. sativa and
their active constituent TQ.
The above observation shows that N. sativa produces hepatoprotective effect
by restoration of anti oxidation enzyme system of the liver, reduction of lipid
peroxidation and inhibition of neutrophil infiltration and release of cytokines from
the inflammatory cells. Our study is also in conformity with Mahmoud et al., (2002)
who reported that active components of N. sativa increased hepatic antioxidant
enzymes activities (superoxide dismutase, catalase, glutathione) and reduces the
hepatic lipid peroxidation.

CHAPTER-5
Integral University, Lucknow 205
5.5 Conclusion
It may be concluded the N. sativa methanolic extracts from different
germination phases showed high protective activity in CCl4-induced hepatotoxicity.
The extracts of germination stages were more protective than non-germinated seed
extract. This may be due to the fact that during germination many phytoconstituents
are produced in high quantities like flavonoids, phenols, tannins, which increase the
level of antioxidant enzymes and reduce the toxicity of CCl4. In the present study 5th
day germination phase followed by 7th
day extracts possesses significant
hepatoprotective activity and reduces the CCl4-induced hepatotoxicity compared
than non-germinated seed extract. The activity seems to be due to active component
thymoquinone which increase the antioxidant enzyme level. Further study is required
in this area as preliminary results are very encouraging to investigate the mechanism
of action of the compound to minimize toxicity.

CHAPTER-5
Integral University, Lucknow 206
5.6 REFERENCES
Abdulrahman, L., Al-Malki. (2010). Antioxidant Properties of Thymol and
Butylated Hydroxytoluene in Carbon Tetrachloride Induced Mice Liver
Injury. JKAU.AU. Sci., 22(1):239-248.
Aebi, H. (1974).Catalase In: Bergmeyer HV, editor.Methods in enzymatic an
alysis. New York: Acadamic press. 2:674‐84.
Ahmad, I.Z., Kamal, A., Islam, M.H. (2010). Alteration in the Activity of
Antioxidant Enzymes in Nigella sativa Seed during Different Phases of
Germination. M. Kalogiannakis, D. Stavrou & P. Michaelidis (Eds.)
Proceedings of the 7th International Conference on Hands-on Science.
25-31 July, pp. 423-426 http://www.clab.edc.uoc.gr/HSci2010
Al-Ghamdi, M.S. (2003). Protective Effect of Nigella sativa Seeds Against
Carbon Tetrachloride-induced Liver Damage. American J. Chinese
Med., 31 (5):721-728.
Asif Mir, Farida Anjum, Naveeda Riaz, Hina Iqbal et al., (2010). Carbon
Tetrachloride (CCl4) induced hepatotoxicity in rats: Curative role of
Solanum nigrum. J. Med. Plants Res., 4:2525-32
Arulkumaran, S., Ramprasath, V.R., Shanthi, P, Sachdanandam, P. (2007).
Alteration of DMBA-induced oxidative stress by additive action of a
modified indigenous preparation Kalpaamruthaa. Chem. Biol. Interact.,
167:99–106.
Ali, B.H., Blunden, G. (2003). Pharmacological and toxicological properties
of Nigella sativa. Phytother. Res., 17: 299–305.
Al-Shabanah, O.A., Badary, O.A., Nagi, M.N., Al-Gharably, N.M., Al-
Rikabi, A.C., Al-Bekairi, A.M. (1998), Thymoquinone protects against
doxorubicin- induced cardiotoxicity without compromising its antitumor
activity. J. Exp. Clin. Cancer Res., 17: 193-198.
Aristatile, B., Al-Numair, K.S., Veeramani, C., Pugalendi, K.V. (2009).
Effect of carvacrol on hepatic marker enzymes and antioxidant status in D-
galactosamine-induced hepatotoxicity in rats. Fundam. Clin. Pharmacol.,
23(6):757-765.
Aruoma, O.I. (1998). Free radicals, oxidative stress, and antioxidants in
human health and disease. J. Am. Oil Chem. Soc., 75:199-212.

CHAPTER-5
Integral University, Lucknow 207
Badary, O.A., Al-Shabanah, O.A., Nagi, M.N. et al., (1997). Thymoquinone
ameliorates the nephrotoxicity induced by cisplatin in rodents and potentiates
its antitumor activity. Can. J. Physiol. Pharmacol., 75:1356-1361.
Badary, O.A., Al-Shabanah, O.A., Nagi, M.N., Al- Rikabi, A.C., Elmazar,
M.M. (1999). Inhibition of benzo (a) pyrene-induced forestomach
carcinogenesis in mice by thymoquinone. Eur. J. Cancer Prev., 8:435-440.
Badary, O.A., Taha, R.A., Gamal, el-Din, A.M., Abdel-Wahab, M.H. (2003).
Thymoquinone is a potent superoxide anion scavenger. Drug Chem. Toxicol.,
26:87–98.
Baser, K.H. (2008). Biological and pharmacological activities of carvacrol
and carvacrol bearing essential oils. Curr. Pharm. Des., 14(29):3106-3119.
Beckman, K.B., Ames, B.N. (1990). The free radical theory of aging matures.
Physiol. Rev., 78: 547–581.
Benedetti, A.M. (1987). Bioelectro.chem.Bioenrg,18.Biochemistry, Edn.4th
309-326.
Burits, M., Bucar, F. (2000). Antioxidant activity of Nigella sativa essential
oil. Phytother. Res., 14:323-328.
Burns, J., Gardne, P.T., Matthews, D., Duthie, G.G., Lean, M.E., Crozier, A.
(2001). Extraction of phenolics and changes in antioxidant activity of red
wines during vinification. J. Agric. Food Chem., 49:5797-5808.
Chance, B., Greenstein, D.S. (1992). The mechanism of catalase actions-
steady state analysis. Arch. Biochem. Biophys., 37:301–339.
Chattopadhyay, R.R., Bhattacharyya, S.K. (2007). Terminalia chebula: An
update. Pharmacog., 1(1):439–45.
Cook, N.C., Samman, S. (1996). Flavonoids chemistry, metabolism,
cardioprotective effects, and dietary sources. Nutri. Biochem., 7:66-76.
Cullen, J.M., Miller, R.T. (2006). The role of pathology in the identification
of druginduced hepatic toxicity. Expe. Opin. Drug Meta. Toxicol., 2: 241-
247.
Curtis, J.J. Mortiz, M. (1972). Serum enzymes derived from liver cell fraction
and response to carbon tetrachloride intoxication in rats. Gastroenterol, 62:
84–92.
Daba, M.H., Abdel-Rahman, M.S. (1998). Hepatoprotective activity of
thymoquinone in isolated rat hepatocytes. Toxicol Lett., 16:95(1):23-9.

CHAPTER-5
Integral University, Lucknow 208
Deleve, L.D., Kaplowitz, N. (1995). Mechanism of drug induced disease.
Gasteroenterol. clin. NAM., 24:787-810.
Diaz, M.N., Frei, B., Vita, J.A., Keaney, J.F. (1997). Antioxidants and
atherosclerotic heart disease. N. Engl. J. Med., 337:408-416.
Drotman, R., Lawhan, G. (1978). Serum enzymes are indications of chemical
induced liver damage. Drug Chem. Toxicol. 1:163–171.
Nayak, D.P., Dinda, S.C., Swain, P.K., Kar, B., Patro, V. J. (2012).
Hepatoprotective activity against CCL4-induced hepatotoxicity in rats of
Chenopodium album aerial parts .J. Phytothe. Pharmacol., 41(2):33-41.
El-Gharieb, M.A., El-Masry, T.A., Emara, A.M., Hashem, M.A. (2010).
Potential hepatoprotective effects of vitamin E and Nigella sativa oil on
hepatotoxicity induced by chronic exposure to malathion in human and male
albino rats Toxicological and Environmental Chemistry, 92 (2):395-412.
Eliman, G. L. (1959). Tissue sulfhydryl group. Arch. Biochem. Biophys.,
82:70-77.
Farghali, H., Kamenikova, L., Hynie, S., Kmonickova, E. (2000). Silymarin
effects on intracellular calcium and cytotoxicity: a study in perfused rat
hepatocytes after oxidative stress injury. Pharmacol. Res., 41: 231-237.
Gülçin, I., Büyükokuroglu, M.F., Oktay, M., Küfrevio, glu, O.I. (2002). On
the in vitro antioxidant properties of melatonin. J. Pineal Res., 33:167–171.
Gupta, A.K., Misra, N. (2006). Hepatoprotective activity of aqueous
ethanolic extract of Chamomile capitula in paracetamol intoxicated albino
rats. American J. Pharmacol. Toxicol., 1:17-20.
Halliwell, B., and Gutteridge J. (1984). Oxygen toxicity oxygen radicals,
transition metals and disease. Biochem. J., 219:1–14.
Halliwell, B., and Gutterridge, J.M.C. (1989). Free Radicals in Biology and
Medicine, seconded Clarendon Press, Oxford.
Handa, S.S. (1991). Management of hepatic aliments. Pharmatimes,
23(4):13.
Haq, A., Abdullatif, M., Lobo, P.I., Khabar, K.S., Sheth, K.V., al-Sedairy,
S.T. (1995). Nigella sativa: effect on human lymphocytes and
polymorphonuclear leukocyte phagocytic activity. Immunopharmacology,
30(2):147-55

CHAPTER-5
Integral University, Lucknow 209
Hesham, R.E., Shgeru, N. (2002). Chemistry of Bioflavonoids. Indian. J.
Pharm. Educ., 36:191-194.
Houghton, P.J., Zarka, R., De las Heras, B., Hoult, J.R.S. (1995). Fixed oil of
Nigella sativa and derived thymoquinone inhibit eicosanoid generation in
leukocytes and membrane lipid peroxidation. Planta Medica., 61: 33-36.
Jaeschke, H., Gores, G.J., Cederbaum, A.I., Hinson, J.A., Pessayre, D.,
Lemasters, J.J. (2002). Mechanisms of hepatotoxicity. Toxicol. Sci., 65
(2):166–76
Janakat, S., Nassar, M. (2010). Hepatoprotective activity of desert truffle
(Terfezia claveryi) in comparison with the effect of Nigella sativa in the rat.
Pakistan J. Nutr., 9 (1): 52-56.
Jeong, H.G., Park, H.Y. (1998). The prevention of carbon tetrachloride-
induced hepatotoxicity in mice by alpha-hederin: inhibiton of cytochrome
P450 2E1 expression. Biochem. Mol. Biol. Int., 45(1):163-170.
Johnston, D.E., Knoerning, C. (1998). Mechanism of early CCl4 toxicity in
cultured rate hepatocytes. pharmacol. Toxicol., 83:231-39.
Kamal, A., Islam, H., Ahmad, I.Z. (2010). Hydroxyl free radical
scavenging activity of Nigella Sativa L. seed extracts in various
Germinating Stages under Cadmium Stress, Int. J. Bio. Sci. Eng., 1(4):203-
220.
Kadiiska, M.B., Gladen, B.C., Baird, D.D., Dikalova, A.E., Sohal,
R.S., Hatch, G.E., Jones, D.P., Mason, R.P., Barrett, J.C. (2000). Biomarkers
of oxidative stress study: are plasma antioxidants markers of CCl4 poisoning?
Free Radic. Biol. Med., 28(6):838-45.
Kamiyama, T., Sato, C., Liu, J. (1993). Role of lipid peroxidation in
acetaminophen induced hepatotoxicity; comparision with
carbontetrachloride. Toxicol Lett., 66:7-12.
Kamal, A., Ahmad, I. Z., Islam, H. (2010). Alteration in the activity of
antioxidant enzymes in Nigella sativa seed during different phases of
germination. Proceedings of the 7th International Conference on Hands-
on Science., Rethymno-Crete, 423-426 http://www.clab.edc.uoc.gr/HSci2010
Kanter, M., Coskun, O., Budancamanak, M. (2005). Hepatoprotective effects
of Nigella sativa L and Urtica dioica L on lipid peroxidation, antioxidant
enzyme systems and liver enzymes in carbon tetrachloride-treated rats. World
J. Gastroenterolo., 11 (42): 6684-6688.

CHAPTER-5
Integral University, Lucknow 210
Kaplowitz, N., Aw, T.Y., Simon, F.R., Stolz, A. (1986).Drug induced
hepatotoxicity. Ann. Intern. Med. 104: 826–839.
Kauppinen, T., Toikkanen, J., Pedersen, D., et al. (1998). Occupational
Exposure to Carcinogens in the European Union in 1990–93, Carex
(International Information System on Occupational Exposure to
Carcinogens), Helsinki, Finnish Institute of Occupational Health.
Keeffe, Emmet, B., Friedman, Lawrence, M. (2004). Handbook of liver
diseases. Edinburgh: Churchill Livingstone. 104–123.
Keli-chen, Geoff, W., Plumb, Richard, N., Bennett, Youngping, Bao. (2005).
Antioxidant activities of extracts from five anti-viral medicinal plants. J.
Ethnopharmacol., 96:201-205.
Kitase, A., Hino, K., Furutani, T., Okuda, M., Gondo, T., Hidaka, I., et al.,
(2005). In situ detection of oxidized n-3 polyunsaturated fatty acids in
chronic hepatitis C: correlation with hepatic steatosis. J. Gastroenterolo., 40:
617-624.
Kohle, C., Schwarz, M., Bock, K.W. (2008). Promotion of
hepatocarcinogenesis in humans and animal models. Arch. Toxicol., 82: 623-
631.
Lee, K.W., Hur, H.J., Lee, H.J., Lee, C.Y. (2005). Antiproliferative effects of
dietary phenolic substances and hydrogen peroxide. J. Agric. Food Chem.,
53:1990–1995.
Lee, W.M. (2003). Acute liver failure in the United States. Seminars in Liver
Disease, 23: 217-226.
Letteron, P., Labbe, G., Degott, C., Berson, A., et al., (1990): Mechanism of
the protective effects of silymarin against carbon tetrachloride-induced lipid
peroxidation and hepatotoxicity in mice: evidence that silymarin acts both as
an inhibitor of metabolic activation and as a chainbreaking antioxidant.
Biochem. Pharmacol., 39:2027-2034.
Lewis, H.W., Elvin-Lewis, M.P.H. (1977). Medical Botany: Plants
Affecting Man’s Health. John Wiley and Sons, New York, PP. 217-18.
Lewis, R.J. (1993). Hawley’s Condensed Chemical Dictionary, Edn. 12th
New York, Van Nostrand Reinhold, pp. 221–222.
Liu, S.L., Degli Esposti, S., Yao, T., Diehl, A.M., Zern, M.A. (1995).
Vitamin E therapy of acute CCl4-induced hepatic injury in mice is associated

CHAPTER-5
Integral University, Lucknow 211
with inhibition of nuclear factor kappa B binding. Hepatology, 22(5):1474-
81.
Mahmoud, M.R., El-Abhar, H.S., Saleh, S. (2002). The effect of Nigella
sativa oil against the liver damage induced by Schistosoma mansoni
infection in mice J. Ethnopharmacol., 79 (1):1-11.
Mansour, M.A., Ginwai, O.T., El-Hadiya, T., El-Khatib, A.S., Al-Shabanah,
O.A., Al-Sawaf, H.A. (2001). Effects of volatile oil constituents of Nigella
sativa on carbon tetrachloride-induced Hepatotoxicity in mice: evidence for
antioxidant effects of thymoquinone. Res. Commun. Mol. Pathol.
Pharmacol., 110(3–4):239–51.
Marx, J. (2004). Inflammation and cancer: the link grows stronger. Science,
36:966–1008.
Mastour, S., Al-Ghamdi, (2003). Protective Effect of Nigella sativa Seeds
Against Carbon Tetrachloride-induced Liver Damage. American J. Chinese
Med., 31(5):721–728.
Mates, J.M. (2000). Effect of antioxidant enzymes in the molecular control of
reactive oxygen species toxicology. Toxicol., 153:83-104.
Mau, J.L., Chao, G.R., Wu, K.T. (2001). Antioxidant properties of
methanolic extracts from several mushrooms. J. Agri. Food Chem., 49:5461–
5467.
McCord, J. M., Fridovich, I. (1969). Superoxide dismutase: an enzymic
function for erythrocuprein (hemocuprein). J. Biol. chem. 244:6049-55.
Mehta, B.K., Pandit, V., Gupta, M. (2009). New principles from seed of
Nigella sativa. Nat. Prod. Res., 23(2):138-48.
Miyoshi, H., Rust, C., Roberts, P.J., Burgart, L.J., Gores, G.J. (1999).
Hepatocyte apoptosis after bile duct ligation in the mouse involves Fas.
Gastroenterology 117: 669-677.
Mohideen, S., Ilavarasan, Sasikala, R.E., Thirumalai, Kumaran, R. (2003).
Hepatoprotective activity of Nigella sativa Linn. Indian J. Pharma.
Sci., 65 (5): 550-551.
Momin, A. (1987). Role of indigenous medicine in primary health care.
1st International Seminar on Unani Medicine, New Delhi, India, pp. 54.
Mumoli, N., Cei, M., Cosimi, A. (2006). Drug-related hepatotoxicity. N.
Engl. J. Med. 354 (20):2191–2193

CHAPTER-5
Integral University, Lucknow 212
Muriel, P., Garcipiana, T. (1992). Silymarin protects against paracetamol-
induced lipid peroxidation and liver damage. J. Appl. Toxicol. 12: 439–442.
Nagi, M.N., Mansour, M.A. (2000). Protective effect of thymoquinone
against doxorubicin-induced cardiotoxicity in rats: a possible mechanism of
protection. Pharmacol. Res., 41:283–289.
Negi, A.S., Kumar, J.K., Luqman, S., Shanker, K., Gupta, M.M., Khanuja,
S.P.S. (2008). Recent advances in plant hepatoprotectives: A chemical and
biological profile of some important leads. Med. Res. Rev., 28:746-772.
Ohkawa, H., Ohishi, N., Yagi, K. (1979). Assay for lipid peroxides in animal
tissue by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
Ostapowicz, G., Fontana, R.J., Schiodt, F.V., et al. (2002). Results of a
prospective study of acute liver failure at 17 tertiary care centers in the
United States. Ann. Intern. Med., 137 (12): 947–54.
Pablo Muriel (2009). Role of free radicals in liver diseases. Hepatol.
Int., 3(4): 526–536.
Patel, T., Roberts, L.R., Jones, B.A., Gores, G.J. (1998). Dysregulation of
apoptosis as a mechanism of liver disease: an overview. Semin. Liver Dis.,
18 (2):105–14.
Palanivel, M.G., Rajkapoor, B., Ra Kumar, R.S. (2008). Hepatoprotective
and Antioxidant Effect of Pisonia aculeata L. against CCl4- Induced Hepatic
Damage in Rats. Sci. Pharm.; 76: 203–215.
Parkin, D.M., Bray, F., Ferlay, J., Pisani, P. (2005). Global cancer statistics,
2002. CA Cancer J Clin., 55:74–108.
Pisani, P., Parkin, D.M., Bray, F., Ferlay, J. (1999). Estimates of the
worldwide mortality from 25 cancers in 1990. Int. J. Cancer., 83:18–29.
Prakash, J., Gupta, S.K., Singh, N. (2001). Chemopreventive activity of
Withania somnifera in experimentally induced fibro sarcoma tumors in Swiss
albino rats. Phytother. Res., 15: 200–204.
Randhawa, M.A., Al- Ghamdi, M.J. (2002). A review of
pharmacotherapeutic effects of Nigella sativa. Pak. Jr. of Med. Res.,
41(2):77-83.
Rice-Evans, C.A., Miller, J., Paganga, G. (1997). Antioxidant properties of
phenolic compounds. Trends Plant Sci., 2:152–159.

CHAPTER-5
Integral University, Lucknow 213
Ronsein, G.E., Guidi, D.B., Benassi, J.C., Wilhelm Filho, D., Pedrosa, R.C.
(2005). Cytoprotective effects of carvedilol against oxygen free radical
generation in rat liver. Redox. Rep., 10: 131-137.
Seitz, H.K., Stickel, F. (2006). Risk factors and mechanisms of
hepatocarcinogenesis with special emphasis on alcohol and oxidative
stress. Biol. Chem., 387:349–360.
Seki, S., Kitada, T., Sakaguchi, H. (2005). Clinicopathological significance
of oxidative cellular damage in non-alcoholic fatty liver diseases. Hepato.
Res. 33:132-134.
Sil, P.C., Manna, P., Sinha, M. (2006). Aqueous extract of Terminalia arjuna
prevents carbon tetrachloride induced hepatic and renal disorders. B.M.C.
Comp. Alte. Med.., 6: 33.
Smith, M.A., Perry, G., Richey, P.L., Sayre, L.M., Anderson, V.E., Beal,
M.F. (1996). Oxidative damage in Alzheimer’s. Nature., 382:120-121.
Subodh, K. Chauhan, Thapliyal, R.P., Sanjay, K. Ojha, Himanshu Rai,
Puneet Singh, Manendra Singh, Amir Khan. (2011). Therapeutic role of
Nigella sativa on turpentine oil induced acute inflammation in rat liver. Inte.
J. Chem. Anal. Sci., 2(8):123-127
Tessitore, L., Bollito, E. (2006). Early induction of TGF-beta1 through a
fasting refeeding regimen promotes liver carcinogenesis by a sub-initiating
dose of diethylnitrosamine. Cell Proliferation., 39:105-116.
Thabrew, M., Joice, P. (1987). A comparative study of the efficacy of Pavetta
indica and Osbeckia octanda in the treatment of liver dysfunction. Planta.
Med. 53: 239–241.
Tiwari, A.K. (2001). Imbalance in antioxidant defense and human disease:
Multiple approach of natural antioxidants therapy. Curr. Sci., 81:1179–1187.
Turkdogan, M.K., Ozbek, H., Yener, Z., Tuncer, I., Uygan, I., Ceylan, E.,
(2003). The role of Urtica dioica and Nigella sativa in the prevention of
carbon tetrachloride-induced hepatotoxicity in rats. Phytother. Res., 17:942-
946.
Weber, L.W., Boll, M., et al. (2003). Hepatotoxicity and mechanism of
action of haloalkanes: carbon tetrachloride as a toxicological model. Cri.t
Rev. Toxicol., 33(2): 105-36.
Zimmerman, H.J. (1978). Drug-induced liver disease. Drugs, 16 (1): 25–45.