chapter ii - inflibnetshodhganga.inflibnet.ac.in/bitstream/10603/40412/6/06_chapter2.pdf · chapter...
TRANSCRIPT

Chapter II
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

47
4. Chapter II: The effect of chromium on arbuscular mycorrhizal and
dark septate endophyte fungal associations
4.1. Introduction
The past decades have witnessed a rapid growth in industries with an accelerated
increase in HM pollution in the biosphere (Gisbert et al., 2000). Even though HMs are
present in all environmental compartments in trace levels, its excess presence can be a
serious cause of concern as it may lead to severe eco-toxicity resulting in loss of
biodiversity and can cause hazardous effect on human health (Shanker et al., 2005).
The long term persistence of these metals is another serious threat to the environment
(Yadav and Chandra, 2011), as they turn out to be conservative pollutants (Kim et al., 2010).
The extent of concern over the adverse effect of HMs may differ, as some are toxic and
others are essential nutrients at various magnitudes.
Large amount of Cr is used as tanning agent in the leather processing and
finishing industry, besides in the production of refractory steel, drilling muds,
electroplating cleaning agents, catalytic manufacture, wood preservation and in the
production of chromic acid (Shanker et al., 2005). As the uptake of Cr into the leather is
not complete and forms insoluble complexes with hydroxyl bridge (Khan et al., 2001),
tannery industry forms the major source of Cr contamination (Chandra et al., 2004a).
But Cr in trace quantity is an essential element for human metabolism as it is required in
glucose metabolism and its deficiency may result in diabetes, arteriosclerosis, growth
problems and cataract in humans (Khan et al., 2001). Another aspect of Cr contamination
is its oxidation state that is, hexavalent Cr is more toxic than trivalent which precipitates
at high pH (Khan et al., 2001) and is mutagenic and carcinogenic (Dixit et al., 2002).
Besides Cr, other HMs like lead (Pb) and cadmium (Cd) are also released in to the
environment in smaller or larger quantities with the tannery effluents and sludge.
Soil micro-organisms especially fungi have the ability to protect the plants against
heavy metal stress by improving plant nutrition by the absorption of various useful
elements (Medina and Azcon, 2010). Arbuscular mycorrhizal fungi, one of the most
beneficial soil fungi which provide a direct link between soil and plant, occur in 80-90%
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

48
of the plant species (Harrison, 1997). Even though the symbiotic association of AM fungi
with plants has been known from centuries, the recent years have witnessed its
improved application in horticulture, agriculture and various decontamination processes
(Ruscitti et al., 2011). Generally AM fungi are seen in the most stressed and nutrient
depleted environments (Biro and Takacs, 2007; Khade and Adholeya, 2009; Rao and
Tak, 2001). The initial colonizers of heavily polluted soils are reported to be metal
tolerant plant species which later tend to develop low mycorrhizal colonization which
improves the soil organic content. This improved soil microclimate may improve the extent
of mycorrhizal colonization which helps in the improved survival rates of the next generation
plants (Regvar and Vogel-Mikus, 2008; Leyval and Joner, 2001; Pawlowska et al., 1996).
Dark septate endophytic fungi are also beneficial plant associates, which extensively
colonize root cortical cells of plants forming melanized or hyaline hyphae which form
microsclerotia or moniliform cells (Muthukumar and Prabha, 2012b, Newsham, 1999).
Dark septate endophytes are seen more prevalent in some heavy metal contaminated sites
than AM fungi (Deram et al., 2008). Even though the DSE fungi have been associated to
stressful habitats (Knaff et al., 2012; Frenot et al., 2005; Korhonen et al., 2004), the effect of
heavy metals like Cr has not been investigated in detail. In addition, co-existence of AM
and DSE fungi are seen in many stressful ecosystems (Wagg et al., 2008). Therefore, it is
important to assess the impact of Cr contamination on AM and DSE fungi, so that further
studies could be focussed on the chromate detoxification using these fungal endophytes.
With this objective in mind, the purpose of this study was to screen the extent of AM and
DSE fungal colonization in a heavily contaminated soil with tannery effluents and sludge.
4.2. Materials and methods
4.2.1. Study sites
Study sites as mentioned in Chapter I.
4.2.2. Sampling
The roots of the seventy four plants species collected from Site-C and Site-NC
(mentioned in Chapter I) were thoroughly rinsed with water to remove adhering soil and
other adherants. The roots were fixed in FAA (formalin-acetic acid-70% ethanol: 5:5:90; v/v)
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

49
and later transported to laboratory for further processing. Rhizospheric soil (soil shaken
from roots and adjacent to roots) was collected and divided into two halves. One half of
the soil sample collected from each plant was packed individually in polythene bags and
stored at 4 ºC for spore enumeration. The second half of the soil samples of all the plants
of a site were bulked and mixed well to form a composite soil sample. This composite
was divided into three sub samples, which was used for assessing soil characteristics.
4.2.3. Determination of soil characters
Soil characters were determined as mentioned in Chapter I.
4.2.4 Preparation of roots for arbuscular mycorrhizal and dark septate endophye
fungal assessment
Fixed roots were washed free of FAA, cut into 1-cm long fragments and cleared
for 45 min. in 2.5% KOH at 100 °C, rinsed in water, acidified for 15 mins in 5 N HCl and
stained in 0.05% trypan blue overnight (Koske and Gemma, 1989). Squashes of the
stained roots were examined for the presence of fungal structures using an Olympus
BX51 microscope (×400). The percentage of root length colonization was quantified
according to the magnified intersection method of McGonigle et al. (1990). The procedure
adopted for assessing AM fungal colonization was also used for examining the extent of
for DSE fungal colonization. We also determined the AM morphological types of the
plant species studied. As only squashed roots were examined, we could not distinguish
the intermediate subtypes described by Dickson (2004).
4.2.5. Enumeration and isolation of arbuscular mycorrhiizal fungal spores
Isolation of spores was done from 25g air dried soil as per the procedure
described by Muthukumar and Udaiyan (2000). Uncollapsed intact AM fungal spores
were transferred to a glass slide containing polyvinyl alcohol lacto glycerol with or
without Melzer’s reagent (1:1, v/v) using a wet needle (Schenck and Perez, 1990). Spores
were identified based on spore morphology and sub-cellular characters. Spore characters
were compared with the original description database established by the Schüβler
laboratory (http://www.Irz-muenchen.de/~schuessler/ amphylo/amphylo_species.html)
and cultural database of INVAM (http://www.invam.caf.wvu.edu).
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

50
4.2.6. Plant and arbuscular mycorrhizal fungal nomenclature
Plant nomenclature and authorities are used by Henry et al. (1989) and for AM
fungi are those of Oehl et al. (2008) and http://www.lrzmuenchen.de/~schuessler/amphylo/
amphylo_species.html.
4.2.7. Statistical analysis
One-way analysis of variance (ANOVA) was used to test the significance of
variation within AM and DSE fungal variables. Variations among the plant species
occurring in both sites were assessed using t-test. Pearson’s correlation was used to assess
the relationship between AM and DSE fungal variables (SPSS, Windows Version 9)
Spore numbers were log transformed and percentage data on root colonization was
arcsine transformed prior to analysis.
4.3. Results
4.3.1. Physico-chemical characteristics of soil samples
The physico-chemical characteristics of soil samples are mentioned in Chapter I
4.3.2. Occurrence of arbuscular mycorrhizal fungi
In the present study, 74 plant species were examined for their association with
AM fungi. Out of the 41 plants species collected from Site-C (Table 4.1), only 19 (46%)
had AM fungal association. Sixty seven percentage (Acrachne racemosa, Alternanthera
tenella, Alternanthera triandra, Blumea oxyodonta, Brachiaria ramosa, Calotropis
gigantea, Canna indica, Chromolaena odorata, Coccinia grandis, Eleusine indica,
Eupatorium odoratum, Lantana camara, Lindernia anagallis, Mikania scandens,
Pennisetum purpureum, Peperomia pellucida, Phyllanthus amarus, Phyllanthus
reticulates, Ricinus communis, Scoparia dulcis, Synedrella nodiflora, Vernonia cinerea)
of plant roots examined from Site-NC (Table 4.2) had AM fungal structures. The AM
fungal colonization was characterized by the presence of hyphae, hyphal coils,
arbuscules, arbusculate coils and vesicles. Plant species like Carex pellita, Commelina
attenuata, Cyperus rotundus, Eragrostis japonica, Lantana camara, Mikania scandens,
Peperomia pellucida and Scoparia dulcis collected from Site-C (Table 4.2) had
significantly higher AM colonization when compared with their counterparts in Site-NC
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Table 4.1. Arbuscular mycorrhizal (AM) and dark septate endophyte (DSE) fungal association, AM morphology and spore
numbers in plant species from heavily chromium (Cr) contaminated site (C) and non contaminated site (NC) sites
Families Plant species Sites Fungal
association AM type
Spore
numbersa
Aizolaceae Trianthema portulacastrum L. C AM/DSE Paris 2.67±0.88
Amaranthaceae Alternanthera tenella Moq. C DSE … 1.67 ± 1.20
NC AM/DSE Paris 5.00 ± 0.58
Alternanthera triandra Steud. C DSE … 2.00 ± 1.00
NC AM/DSE Paris 2.67 ± 0.67
Amaranthus spinosus L. C … … 2.00 ±1.00
NC DSE … 1.67 ±0.33
Amaranthus viridis Pollich ex Moq. C DSE … 1.33 ± 0.88
NC DSE … 5.00 ± 0.58
Araceae Caladium bicolor Vent. C DSE … 1.67 ± 0.67
Colocasia esculenta C DSE 1.33±0.33
NC DSE … 2.67±0.88
Asclepiadaceae Calotropis gigantea (L.) W.T.Aiton NC AM/DSE Paris 1.00 ± 0.00
Asteraceae Artemisia japonica Thunb. C DSE … 4.33 ± 0.33
Blumea oxyodonta DC. NC AM/DSE Paris 6.33 ± 0.67
Chromolaena odorata (L.) R.M.King & H.Rob. NC AM/DSE Arum 3.67 ± 0.33
Eclipta prostrata (L.) L. C …. … 1.67±0.67
Eupatorium odoratum L. NC AM/DSE Paris 1.00±0.00
Mikania scandens (L.) Willd. C AM … 1.33±0.33
NC AM/DSE Arum 2.00±0.58
Synedrella nodiflora Gaertn. NC AM Paris 1.67±0.67
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Families Plant species Sites Fungal
association AM type
Spore
numbersa
Cannaceae Canna indica Redoute C AM Paris 1.00 ± 0.58
NC AM Paris 2.00 ± 0.58
Capparaceae Cleome viscosa L. C …. … 1.67 ±1.20
NC DSE … 5.00 ± 1.00
Commelinaceae Commelina attenuata B. Heyne ex Steud. C AM/DSE Paris 3.33±1.20
NC DSE … 2.67±1.67
Compositae Vernonia cinerea (L.) Less C AM/DSE Arum 2.67±0.88
NC AM/DSE Arum 1.33±0.33
Convolvulaceae Ipomea obscura (L.) Less C DSE … 1.67±0.67
Ipomea quamoclit L. C DSE … 1.33±0.33
Cucurbitaceae Coccinia grandis (L.) Voigt C AM/DSE Arum 1.00±0.00
NC AM/DSE Arum 2.33±0.67
Cyperaceae Carex pellita Muhl. ex Schkuhr C AM/DSE Arum 1.67 ± 0.67
NC DSE … 3.33 ± 1.20
Cyperus rotundus Benth. C AM Paris 2.33±0.67
NC DSE … 2.33±0.67
Cyperus stoloniferus Retz. C DSE … 1.67±0.33
NC DSE … 1.00±0.00
Elaeocarpaceae Muntingia calabura L. C …. … 3.33±1.20
Euphorbiaceae Phyllanthus amaras Schumach. C DSE … 1.00±0.00
NC AM/DSE Paris 2.00±1.00
Phyllanthus reticulatus Peir. C AM Paris 2.33±0.67
NC AM/DSE Paris 3.67±0.33
Ricinus communis L. C AM Arum 1.00±0.00
NC AM Arum 1.33±0.88
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Families Plant species Sites Fungal
association AM type
Spore
numbersa
Nephrolepidaceae Nephrolepis sp. C DSE … 3.00±0.58
Onagraceae Ludwigia octovalvis (Jacq.) P.H.Raven C DSE … 2.33±0.88
Passifloraceae Passiflora foetida Vell. C AM Arum 3.00±1.00
Piperaceae Peperomia pellucida (L.) Kunth C AM/DSE Arum 1.00±0.00
NC AM/DSE Arum 3.67±0.33
Poaceae Acrachne racemosa Ohwi C DSE … 1.67 ± 0.67
NC AM/DSE Arum 1.33 ± 0.88
Brachiaria ramosa (L.) Stapf NC AM/DSE Paris 4.67 ± 0.67
Cynodon dactylon (L.) Pers. C AM/DSE Paris 6.33±0.33
Digitaria radicosa (J.Presl) Miq. C DSE … 3.33±1.20
Eleusine indica Steud. NC AM/DSE Arum 2.33±0.67
Eragrostis japonica Trin. C AM Paris 1.00±0.00
Pennisetum purpureum Schumach. NC AM/DSE Paris 2.33±0.67
Polygonaceae Persicaria chinensis (L.) Nakai NC DSE … 1.67±0.33
Pteridaceae Acrostichum speciosumWilld. C AM/DSE Intermediate 3.00 ±1.15
Scrophulariaceae Lindernia anagallis (Burm.f.) Pennell C DSE … 3.67±0.88
NC AM/DSE Paris 1.67±0.33
Lindernia crustacea (L.) F. Muell. C AM/DSE Arum 2.00±1.00
Scoparia dulcis L. C AM Paris 2.67±0.88
NC AM/DSE Paris 1.67±0.67
Solanaceae Datura stramonium L. C AM Arum 1.67±0.67
Physalis minima L. NC DSE … 3.33±0.88
Solanum nigrum L. C DSE … 2.67±0.88
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Families Plant species Sites Fungal
association AM type
Spore
numbersa
Thelypteridaceae Macrothelypteris torresiana (Gaudich.) Ching C DSE … 1.33 ± 0.33
Urticaceae Pouzolzia auriculata Wight NC DSE … 2.00 ± 1.00
Verbenaceae Lantana camara L. C AM/DSE Paris 3.00±1.00
NC AM/DSE Paris 2.67±0.67
Woodsiaceae Diplazium esculentum (Retz.) Sw. C DSE … 1.67±0.33 aSpore numbers/100g of soil
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Table 4.2. Extent of arbuscular mycorrhizal (AM) and dark septate endophyte (DSE) fungal in plants species collected from chromium
contaminated site (Site-C)
Plant name AM colonization (%)
a
DSE colonization (%)
b
RLH RLHC RLA RLAC RLV RLT DSEH DSEMI DSEMO DSET
Acrachne racemosa -- -- -- -- -- -- 43.33 ± 1.76 -- 2.67 ± 0.67 46.00 ± 1.15
Acrostichum speciosum 25.55± 2.72c 12.59 ± 1.45 -- 3.08 ± 1.34 12.44 ± 2.48 53.62 ± 2.58 14.30 ± 1.35 -- -- 14.30 ± 1.35
Alternanthera tenella -- -- -- -- -- -- 48.17 ± 4.72 -- 14.60 ± 2.72 62.77± 4.98
Alternanthera triandra -- -- -- -- -- -- 26.67 ± 3.71 -- -- 26.67 ± 3.71
Artemisia japonica -- -- -- -- -- -- 18.67 ± 0.67 -- 3.33±0.67 22.00 ± 1.15
Amaranthus spinosus -- -- -- -- -- -- -- -- -- --
Amaranthus viridis -- -- -- -- -- -- 22.00 ± 3.06 2.67 ± 0.67 -- 24.67 ± 3.71
Caladium bicolor -- -- -- -- -- -- 33.33 ± 1.76 -- -- 33.33 ± 1.76
Canna indica _ 16.57 ± 1.33 _ 5.72 ± 1.07 6.36 ± 1.26 28.65 ± 2.88 -- -- -- --
Carex pellita 12.30 ± 1.44 -- 3.10 ± 0.64 -- 4.31 ± 0.57 19.71 ± 1.91 34.53 ± 1.82 3.71 ± 0.07 -- 38.24 ± 1.79
Cleome viscosa -- -- -- -- -- -- -- -- -- --
Coccinia grandis 11.73±0.94 -- 6.51±0.41 -- -- 18.24±1.03 21.32±0.93 -- -- 21.32±0.93
Colocasia esculenta -- -- -- -- -- -- 30.00 ± 1.15 -- 3.33 ± 0.67 33.33 ± 0.67
Commelina attenuata -- 6.24 ± 1.62 -- 3.77 ± 1.13 -- 10.02 ± 2.27 29.32 ± 2.41 11.89±2.28 -- 41.21 ± 3.49
Cynodon dactylon -- 3.79 ±1.12 -- 5.08 ± 2.34 -- 8.87 ± 3.26 38.33 ± 2.14 -- 25.72 ± 3.27 64.05 ± 4.75
Cyperus rotundus 24.64 ± 0.85 -- 35.19 ± 0.06 -- -- 59.83 ± 0.80 -- -- -- --
Cyperus stoloniferus -- -- -- -- -- -- 31.33 ± 1.76 -- -- 31.33 ± 1.76
Datura stramonium 21.29 ±0.20 -- 33.14 ± 0.59 -- 6.39 ± 1.74 60.82 ± 1.95 -- -- -- --
Digitaria radicosa -- -- -- -- -- -- 66.85 ± 4.14 -- 5.16 ± 2.29 72.00 ± 6.31
Diplazium esculentum -- -- -- -- -- -- 25.33 ± 1.33 -- -- 25.33 ± 1.33
Eclipta prostrata -- -- -- -- -- -- -- -- -- --
Eragriostis japonica -- 18.32 ± 1.00 -- 26.04 ± 0.66 7.06 ± 1.71 51.92 ± 2.08 -- -- -- --
Ipomea obscura -- -- -- -- -- -- 33.46 ± 2.03 -- 3.92 ± 1.20 37.38 ± 0.84
Ipomea quamoclit -- -- -- -- -- -- 27.24±0.99 -- -- 27.24 ± 0.99
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Plant name AM colonization (%)
a
DSE colonization (%)
b
RLH RLHC RLA RLAC RLV RLT DSEH DSEMI DSEMO DSET
Lantana camara 11.50 ± 2.50 -- 5.75 ± 1.52 -- -- 17.24 ± 3.98 45.40 ± 3.20 -- 18.97 ± 1.99 64.37 ± 3.50
Lindernia anagallis -- -- -- -- -- -- 42.00 ± 2.31 -- -- 42.00 ± 2.31
Lindernia crustacea 18.26 ± 0.76 -- 3.11 ± 0.60 -- 3.17 ± 0.68 24.53 ± 0.52 15.69 ± 1.43 -- -- 15.69 ± 1.43
Ludwigia octovalvis -- -- -- -- -- -- 35.10 ± 2.21 -- 9.72 ± 1.87 44.82 ± 0.59
Macrothelypteris torresiana -- -- -- -- -- -- 40.13 ± 0.84 9.50 ± 1.73 -- 49.62 ± 2.20
Mikania scandens 26.43 ± 1.64 -- 31.35 ± 0.16 -- 7.57 ± 0.53 65.35 ± 1.87 -- -- -- --
Muntingia calabura -- -- -- -- -- -- -- -- -- --
Nephrolepis sp -- -- -- -- -- -- 10.67 ± 1.76 -- -- 10.67 ± 1.76
Passiflora foetida 31.61 ± 2.60 -- 10.05 ± 2.37 -- 3.65 ± 1.14 45.31 ± 3.82 -- -- -- --
Peperomia pellucida 26.35 ± 0.53 -- 30.54 ± 1.11 -- -- 56.89 ± 0.64 6.56 ± 1.11 -- -- 6.56 ± 1.11
Phyllanthus amarus -- -- -- -- -- 36.67 ± 1.67 -- 36.67 ± 1.67
Phyllanthus reticulatus -- 9.30±0.86 -- 3.74±0.88 -- 13.04±0.86 -- -- -- --
Ricinus communis 35.58 ± 1.14 -- 20.86 ± 1.66 -- -- 56.44 ± 0.53 -- -- -- --
Scoparia dulcis -- 31.22 ± 1.71 -- 10.92 ± 1.91 6.95 ± 1.07 49.09 ± 1.81 -- -- -- --
Solanum nigrum -- -- -- -- -- -- 28.31 ±1.87 4.59 ± 0.59 -- 32.90 ± 1.74
Trianthema portulacastrum -- 13.75 ± 1.79 -- 4.53 ± 1.02 5.47 ± 1.03 23.75 ± 3.29 34.20 ± 2.32 -- -- 34.20 ± 2.32
Vernonia cinerea 12.57 ± 2.14 -- 6.00 ± 1.34 -- -- 18.58 ± 3.48 20.01 ± 2.71 -- -- 20.01 ± 2.71 aRLH - root length with hyphae, RLHC -root length with hyphal coils, RLA - root length with arbuscules, RLAC -root length arbusculate coils, RLV -root length with vesicles,
RLT- root length with total colonization bDSEH- root length with dark septate fungal hyphae, DSEMI- root length with moniliform hyphae, DSEMO-root length with microsclerotia, DTC- root length with total
colonization cMean ± S.E
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

51
irrespective of the high Cr content in the soil. Fungal structures were absent in
Carex pellita, Commelina attenuata and Cyperus rotundus collected from Site-NC,
whereas, the same plant species were mycorrhizal in Site-C. Arbuscular mycorrhizal
fungal structures were absent in the roots of Acrachne racemosa, Alternanthera tenella,
Alternanthera triandra, Artemisia japonica, Amaranthus spinosus, Amaranthus viridis,
Caladium bicolor, Cleome viscosa, Colocasia esculenta, Cyperus stoloniferus, Digitaria
radicosa, Diplazium esculentum, Eclipta prostrata, Ipomea obscura, Ipomea quamoclit,
Lindernia anagallis, Ludwigia octovalvis, Macrothelypteris torresiana, Muntingia
calabura, Nephrolepis sp., Phyllanthus amarus and Solanum nigrum from Site-C.
Roots of A. spinosus, A. viridis, Cleome viscosa, Colocasia esculenta, Cyperus
stoloniferus lacked AM fungal structures in both the sites.
Out of the four ferns (Acrostichum speciosum, Diplazium esculentum,
M. torresiana, Nephrolepis sp.) examined from Site-C, only roots of Acrostichum
speciosum contained AM fungal structures. All the members of the Poaceae examined
from both the sites, except for Acrachne racemosa and Digitaria radicosa from Site-C
were mycorrhizal. Eragrostis japonica showed a colonization rate of 52% even in Site-C.
Although 50% of the taxa from the predominant family Amaranthaceae were mycorrhizal
at Site-C they were non-mycorrhizal at Site-NC (Table 4.3). Eclipta prostrata and
Artemisia japonica (Site-C) lacked AM fungal association among Asteraceae.
4.3.3 Extent of arbuscular mycorrhizal fungal colonization
The different AM fungal variables exhibited significant variations among plant
species. The percentage of total root length colonization (RLT) ranged from 8.87
(Cynodon dactylon) to 65.35 (M. scandens) in Site-C (Table 4.2) and 7.70 (Blumea
oxyodonta) to 61.30 (Ricinus communis) in Site-NC (Table 4.3). The extent of AM fungal
structures including percentage root length with hyphae (RLH, F40,82 =144.69), hyphal
coils (RLHC, F40,82 =115.93), arbuscules (RLA, F40,82 =246.98), arbusculate coils (RLAC
F40,82 =58.81) and vesicles (RLV, F40,82 =20.95) significantly (P<0.01) varied among plant
species at Site-C. In Site-C, RLH, RLHC, RLA, RLAC and RLV ranged between 11.50
(L. camara) and 35.58 (R. communis), 3.79 (Cynodon dactylon) and 31.22 (S. dulcis),
3.10 (Carex pellita) and 35.19 (Cyperus rotundus), 3.08 (Acrostichum speciosum) and
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Table 4.3. Extent of arbuscular mycorrhizal (AM) and dark septate endophyte (DSE) fungal colonization in plant species collected from
non contaminated site (Site NC)
Plant species AM colonization (%)
a
DSE colonization (%)
b
RLH RLHC RLA RLAC RLV RLT DSEH DSEMI DSEMO DSET
Acrachne racemosa 15.46 ± 0.87c -- 7.40 ± 1.36 -- 5.93 ± 1.39 28.80 ± 1.59 32.82 ± 2.74 10.48 ± 1.74 -- 43.29 ± 4.44
Alternanthera tenella -- 14.89±2.10 -- 12.49 ± 1.89 2.53 ± 0.42 29.91 ± 4.10 39.45 ± 2.28 5.96 ± 2.35 -- 45.41 ± 3.86
Alternanthera triandra -- 25.02±2.37 -- 34.9 ± 0.82 -- 59.92 ± 2.84 21.83 ± 1.75 2.75 ± 0.52 -- 24.55 ± 1.48
Amaranthus spinosus -- -- -- -- -- -- 6.89 ± 0.35 -- -- 6.89 ± 0.35
Amaranthus viridis -- -- -- -- -- -- 13.19 ±1.51 3.02±0.29 -- 16.21±1.45
Blumea oxyodonta -- 5.76 ± 0.95 -- 1.94 ± 0.06 -- 7.70 ± 0.90 30.67 ± 1.76 -- -- 30.67 ± 1.76
Brachiaria ramosa -- 15.91 ± 4.08 -- 4.31 ± 0.58 4.91 ± 0.50 25.13 ± 4.97 12.50 ± 2.64 3.11 ± 1.13 -- 15.61 ± 2.53
Calotropis gigantea -- 9.09 ± 1.48 -- 5.68 ± 1.49 -- 14.76 ± 2.97 44.31 ± 1.59 -- 3.97 ± 0.55 48.28 ± 2.14
Canna indica -- 15.00 ± 0.96 -- 41.66 ± 1.67 56.66 ± 0.96 -- -- -- --
Carex pellita -- -- -- -- -- -- 17.33 ± 1.76 -- -- 17.33 ± 1.76
Chromolaena odorata 24.03 ± 0.55 -- 5.65 ± 1.99 -- 2.53 ± 0.74 32.21 ± 1.34 24.80 ± 1.62 -- -- 24.80 ± 1.62
Cleome viscosa -- -- -- -- -- -- 50.98 ± 2.99 2.61 ± 0.65 -- 53.59 ± 3.27
Coccinia grandis 10.19 ± 3.22 -- 5.06 ± 2.17 -- -- 15.25 ± 4.38 23.95 ± 1.80 -- 5.84 ± 2.33 29.79 ± 3.13
Colocasia esculenta -- -- -- -- -- -- 38.00 ± 1.15 -- -- 38.00 ± 1.15
Commelina attenuata -- -- -- -- -- -- 43.65 ± 3.83 3.30 ± 1.35 -- 46.94 ± 2.94
Cyperus rotundus -- -- -- -- -- -- 36.00 ± 2.00 2.67± 0.67 -- 38.67 ± 1.76
Cyperus stoloniferus -- -- -- -- -- -- 23.51±1.15 -- -- 23.51±1.15
Eleusine indica 20.53 ± 2.09 -- 10.58 ± 1.73 -- 7.66 ± 1.56 38.77 ± 4.42 8.21 ± 1.07 -- -- 8.21 ± 1.07
Eupatorium odoratum -- 14.24 ± 2.73 -- 6.28 ± 1.92 -- 20.52 ± 4.48 38.62 ± 0.88 -- 3.10 ± 0.67 41.72 ± 1.33
Lantana camara 23.14 ± 2.11 -- 13.86 ± 0.94 -- 5.20 ± 1.00 42.20 ± 3.48 12.14 ± 1.99 4.06 ± 1.56 -- 16.20 ± 2.93
Lindernia anagallis -- 15.32 ± 1.02 -- 17.11 ± 1.04 3.11 ± 0.70 35.54 ± 1.49 26.35 ± 0.93 -- -- 26.35 ± 0.93
Mikania scandens 13.38 ± 1.93 -- 5.09 ± 1.16 -- 6.42 ± 1.30 24.88 ± 3.31 -- -- -- --
Pennisetum purpureum -- 5.27 ± 0.67 -- 2.64 ± 0.68 -- 7.91 ± 1.19 11.18 ± 1.28 2.63 ± 0.65 -- 13.80 ± 1.92
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Plant species AM colonization (%)
a
DSE colonization (%)
b
RLH RLHC RLA RLAC RLV RLT DSEH DSEMI DSEMO DSET
Peperomia pellucida 28.99 ± 0.81 -- 22.91 ± 3.52 -- -- 51.90 ± 4.24 9.95 ± 1.61 -- -- 9.95 ± 1.61
Persicaria chinensis -- -- -- -- -- -- 28.67 ± 1.76 -- -- 28.67 ± 1.76
Phyllanthus amarus -- 11.60 ± 1.89 -- 6.43 ± 1.26 2.58 ± 0.64 20.61 ± 2.70 27.70 ± 3.26 -- -- 27.70 ± 3.26
Phyllanthus reticulatus -- 21.12 ± 2.80 -- 23.53 ± 2.71 -- 44.64 ± 1.42 20.89 ± 1.13 -- 3.36±0.95 24.25 ± 0.57
Physalis minima -- -- -- -- -- -- 22.00 ± 1.15 -- -- 22.00 ± 1.15
Pouzolzia auriculata -- -- -- -- -- -- 42.66 ± 1.96 3.50 ± 1.00 1.75 ± 0.02 47.91 ± 2.06
Ricinus communis 28.14 ± 1.66 -- 33.15 ± 1.02 -- -- 61.30 ± 2.14 -- -- -- --
Scoparia dulcis -- 13.35 ± 2.13 -- 5.04 ± 1.12 2.52 ± 0.56 20.91± 2.46 5.74 ± 0.16 -- -- 5.74 ± 0.16
Synedrella nodiflora -- 22.89 ± 1.97 -- 4.50 ± 1.76 3.17 ± 1.25 30.56 ± 1.73 -- -- -- --
Vernonia cinerea 22.79 ± 3.26 -- 13.03 ± 2.45 -- -- 35.82 ± 5.47 22.53 ± 2.00 -- -- 22.53 ± 2.00 aRLH - root length with hyphae, RLHC -root length with hyphal coils, RLA - root length with arbuscules, RLAC -root length arbusculate coils, RLV - root length with vesicles,
RLT- root length with total colonization bDSEH - root length with dark septate fungal hyphae, DSEMI - root length with moniliform hyphae, DSEMO - root length with microsclerotia, DTC - root length with total colonization cMean ± S.E
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

52
26.04 (E. japonica) and 3.17 (Lindernia crustacea) and 12.44 (Acrostichum speciosum)
respectively (Figure 4.1). Likewise, at Site-NC also, the RLH (F32,66 =85.54), RLHC
(F32,66 =36.39), RLA ((F32,66 =55.37), RLAC (F32,66 =121.41) and RLV (F32,66 =15.90)
showed significant variations (P<0.01) among the plant species and ranged between
10.19 (Coccinia grandis) and 28.99 (P. pellucida), 5.27 (Pennisetum purpureum) and
25.02 (Alternanthera triandra), 5.06 (Coccinia grandis) and 33.15 (R. communis), 1.94
(B. oxyodonta) and 41.66 (Canna indica), 2.52 (Scoparia dulcis) and 7.66 (Eleusine
indica) respectively. Significant variation was seen in all the AM fungal variables for
plant species occurring at both the sites except for the RLA (t2,4=0.66, P>0.05),
RLH (t2,4=0.46, P>0.05) and RLT (t2,4=0.94, P>0.05) of Coccinia grandis, RLHC
(t2,4=0.95, P>0.05) of Canna indica and RLV (t2,4=0.82, P>0.05) of M. scandens (Table 4.4).
4.3.4. Arbuscular mycorrhizal morphological types
Almost all the mycorrhizal plants (99%) collected from both the sites had Arum or
Paris type AM morphology except Acrostichum speciosum collected from Site-C
(Table 4.1). This member of Pteridaceae family had intermediate type morphology.
Typical Paris type morphology was more prevalent in Site-C.
4.3.5. Arbuscular mycorrhizal fungal spores
Low density of AM fungal spores was observed in the rhizospheric soil of all the
74 plant species collected from both the sites (Site-C and Site-NC). Both the sites
harboured only a single spore morphotype belonging to Scutellospora calospora. There was
not much difference in the number of spores of each plant species from both the sites as
AM fungal spores ranged between 1 (Canna indica, Coccinia grandis, E. japonica,
Peperomia pellucida, Phyllanthus amarus, R. communis) to 6 (Cynodon dactylon) per
100 g of soil at Site-C and 1 (Calotropis gigantea, Cyperus stoloniferus, Eupatorium
odoratum) to 6 (B. oxyodonta) per 100 g of soil at Site-NC (Table 4.1). Significant
(P<0.01) variation was seen in spore numbers among plant species at both Site-C
(F40, 82 =2.08) and Site-NC (F32, 66 =3.51).
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.
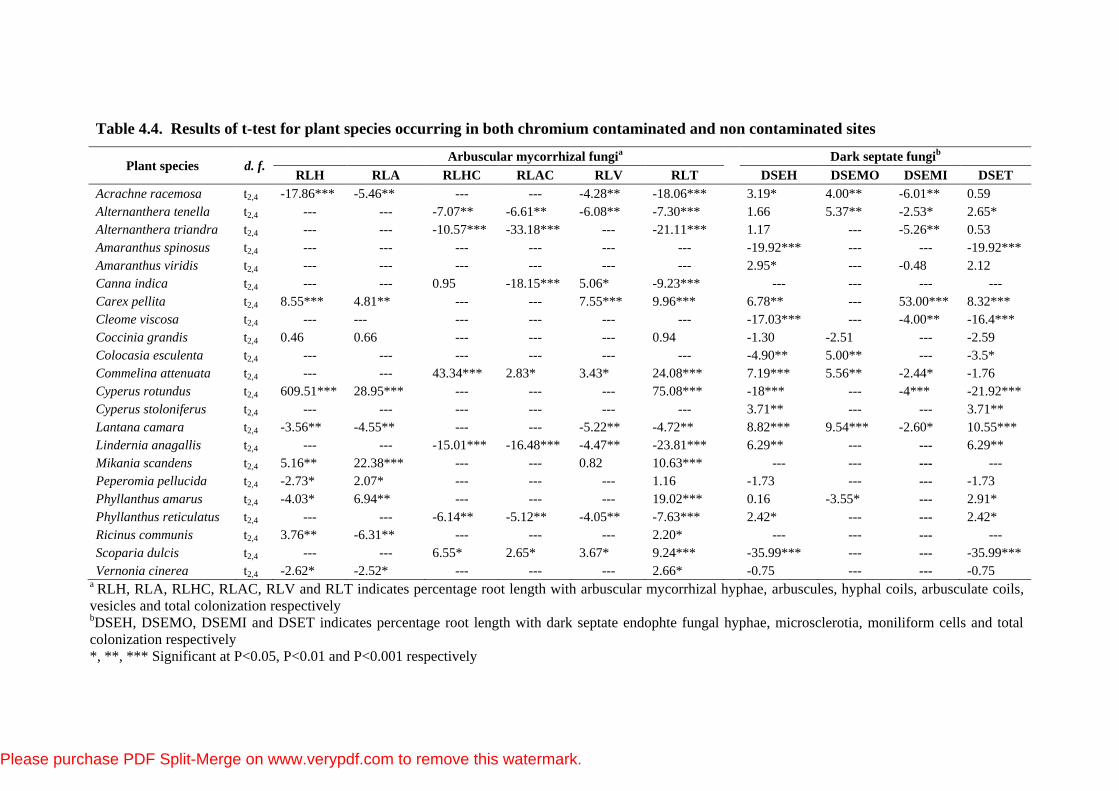
Table 4.4. Results of t-test for plant species occurring in both chromium contaminated and non contaminated sites
Plant species d. f. Arbuscular mycorrhizal fungi
a Dark septate fungi
b
RLH RLA RLHC RLAC RLV RLT DSEH DSEMO DSEMI DSET
Acrachne racemosa t2,4 -17.86*** -5.46** --- --- -4.28** -18.06*** 3.19* 4.00** -6.01** 0.59
Alternanthera tenella t2,4 --- --- -7.07** -6.61** -6.08** -7.30*** 1.66 5.37** -2.53* 2.65*
Alternanthera triandra t2,4 --- --- -10.57*** -33.18*** --- -21.11*** 1.17 --- -5.26** 0.53
Amaranthus spinosus t2,4 --- --- --- --- --- --- -19.92*** --- --- -19.92***
Amaranthus viridis t2,4 --- --- --- --- --- --- 2.95* --- -0.48 2.12
Canna indica t2,4 --- --- 0.95 -18.15*** 5.06* -9.23*** --- --- --- ---
Carex pellita t2,4 8.55*** 4.81** --- --- 7.55*** 9.96*** 6.78** --- 53.00*** 8.32***
Cleome viscosa t2,4 --- --- --- --- --- --- -17.03*** --- -4.00** -16.4***
Coccinia grandis t2,4 0.46 0.66 --- --- --- 0.94 -1.30 -2.51 --- -2.59
Colocasia esculenta t2,4 --- --- --- --- --- --- -4.90** 5.00** --- -3.5*
Commelina attenuata t2,4 --- --- 43.34*** 2.83* 3.43* 24.08*** 7.19*** 5.56** -2.44* -1.76
Cyperus rotundus t2,4 609.51*** 28.95*** --- --- --- 75.08*** -18*** --- -4*** -21.92***
Cyperus stoloniferus t2,4 --- --- --- --- --- --- 3.71** --- --- 3.71**
Lantana camara t2,4 -3.56** -4.55** --- --- -5.22** -4.72** 8.82*** 9.54*** -2.60* 10.55***
Lindernia anagallis t2,4 --- --- -15.01*** -16.48*** -4.47** -23.81*** 6.29** --- --- 6.29**
Mikania scandens t2,4 5.16** 22.38*** --- --- 0.82 10.63*** --- --- --- ---
Peperomia pellucida t2,4 -2.73* 2.07* --- --- --- 1.16 -1.73 --- --- -1.73
Phyllanthus amarus t2,4 -4.03* 6.94** --- --- --- 19.02*** 0.16 -3.55* --- 2.91*
Phyllanthus reticulatus t2,4 --- --- -6.14** -5.12** -4.05** -7.63*** 2.42* --- --- 2.42*
Ricinus communis t2,4 3.76** -6.31** --- --- --- 2.20* --- --- --- ---
Scoparia dulcis t2,4 --- --- 6.55* 2.65* 3.67* 9.24*** -35.99*** --- --- -35.99***
Vernonia cinerea t2,4 -2.62* -2.52* --- --- --- 2.66* -0.75 --- --- -0.75 a RLH, RLA, RLHC, RLAC, RLV and RLT indicates percentage root length with arbuscular mycorrhizal hyphae, arbuscules, hyphal coils, arbusculate coils,
vesicles and total colonization respectively bDSEH, DSEMO, DSEMI and DSET indicates percentage root length with dark septate endophte fungal hyphae, microsclerotia, moniliform cells and total
colonization respectively
*, **, *** Significant at P<0.05, P<0.01 and P<0.001 respectively
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Figure 4.1. Arbuscular mycorrhizal colonization in plant species growing on chromium
contaminated soil:
a. Appresorium (arrow head) in Vernonia cinerea
b. Arbusculate coils (ac) in Commelina attenuata
c. Arbuscules (ar) in Lantana camara
d. vesicles (v) in Alternanthera tenella
e. hyphal coils (hc) and vesicles (v) in Eragriostis japonica.
Scale bars: a=100 μm; b-e=50 μm.
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

53
4.3.6. Occurrence and extent of dark septate endophyte fungi
Of the 74 plant species examined for DSE association 88% from Site-NC and
68 % from Site-C had DSE fungal association (Table 4.1). Only 28 of the 41 plant
species examined from Site-C had DSE fungal structures (Table 4.1). Except Canna
indica (Cannaceae), M. scandens (Asteraceae), R. communis (Euphorbiaceae) and
Synedrella nodiflora (Solanaceae), all the other plant species collected from Site-NC had
DSE fungal association. Canna indica, M. scandens and R. communis occuring in both
the sites lacked DSE fungal association. Moderate rate of DSE colonization was seen in
Amaranthus viridis, Alternanthera tenella, Carex pellita, Cyperus stoloniferus, Lantana
camara, Lindernia anagallis, and Phyllanthus amarus collected from Site-C while the
same plant species had low or absence of DSE fungal structures at Site-NC.
Dark septate endophytic fungal microsclerotia were found only in 16 plant
species, of which 5 plants (Amaranthus viridis, Carex pellita, Commelina attenuata,
M. torrensiana, S. nigrum) belonged to Site-C. The roots of A. racemosa, Alternanthera
tenella, Alternanthera triandra, Cleome viscosa, Cyperus rotundus, Commelina
attenuata, and L. camara collected from Site-NC contained microsclerotia, whereas
microsclerotia were absent in many of the DSE fungal associated plants in heavily
polluted Site-C. Moniliform cells were seen in roots of 22% of plants examined from
Site-C and 15% plants examined from Site-NC.
The percentage of total root length colonized by DSE fungi (DSET) ranged from
6.56 (Peperomia pellucida) to 72 (D. radicosa) and 5.74 (S. dulcis) to 53.59 (Cleome viscosa)
procured from Site-C and Site-NC respectively (Table 4.2 and 4.3). Similarly, the
percentage root length with DSE fungal hyphae (DSEH) ranged from 5.74 (Peperomia
pellucida) to 66.85 (D. radicosa) in Site-C and 5.74 (S. dulcis) to 50.98 (Cleome viscosa)
in Site-NC (Figure 4.2). In both the sites, the minimum percentage root length with
microsclerotia (DSEMI) and moniliform cells (DSEMO) were seen in Cleome viscosa
(2.61) and Pouzolzia auriculata (1.75) collected from Site-NC (Figure 4.2). Likewise, the
maximum %DSEMI (11.89) and %DSEMO (25.72) were observed in Commelina
attenuata and Cynodon dactylon examined from Site-C. Significant (P<0.01) variation
existed for DSEH, DSEMI, DSEMO and DSET among plant species in Site-C
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Figure 4.2. Dark septate endophyte fungal colonization in plant species growing on
chromium contaminated soil:
a. Dark septate endophytic fungal hyphae (dh) in Artemisia japonica
b. DSE fungal hyphae (dh) and microsclerotia (ms) in Chromolaena odorata
c. microsclerotia (ms) in Commelina attenuata
d. microsclerotia (ms) in Cleome viscosa.
Scale bars: a-d= 50 μm.
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

54
(F40,82 =86.62, F40,82 =28.26, F40,82 =37.00, F40,82 =93.8) and Site-NC (F32,66 =48.19,
F32,66 =10.61, F32,66 =10.04, F32,66 =47.07) respectively. Significant negative correlation
existed between RLT and DSET at Site-C (r= -0.534; P<0.01; n=41). Spore numbers and
DSEMO also showed significant positive correlation at Site-C (r=0.517; P<0.01; n=41).
4.4 Discussion
The tannery effluent contaminated soil had high concentrations of Cr, which is a
clear indication of the extreme selection pressure on the AM and DSE fungal endophytes.
The incomplete uptake of Cr during tanning explains for the presence of high Cr content
in the highly contaminated soil. On a physico-chemical examination of tannery effluent
laden soil, Verma and Biswas (2013) reported the alkaline nature of contaminated soil
(7.5-7.2) which falls in line with the present study. The high levels of N, P and K in Site-C
could be attributed to the organic content present in the tannery sludge (Munn et al., 2000).
There are also reports on the presence of N, Ca, Mg, P and small amounts of sodium in
tannery effluents (Ogbonna et al., 1998; Frank, 1998).
Less than half the percentage of total plants examined from Site-C had AM fungal
association in spite of the fact that high concentration of HMs are known to adversely
affect the AM fungal formation and function (Del Vel et al., 1999; Pichardo et al., 2012).
The adverse effects of arsenic on AM fungal colonization were reported in Nicotiana
tabacum by Jianfeng et al. (2009). Likewise, low levels of AM colonization was seen in
multi metal (Cu, Zn and Mn) contaminated soil inoculated with indigenous (31%) or
introduced (19%) AM fungi (Wang et al., 2005). Maximum AM root colonization of 35%
was reported for clover growing on mine spoils heavily contaminated with Zn and Cd
(Gildon and Tinker, 1981). In a calamine spoil mine rich in Pb and Zn in southern
Poland, only 17% of the absolute cover was harboured with mycorrhizal plant species
(Pawlowska et al., 1996). In the present study, the high nutrient levels in contaminated
soil could also be a reason of lack of AM colonization in plant species, as soils with high
fertility tend to decrease the dependence of plants on AM fungi (Siqueira and Saggin -
Junior, 1995).
Nevertheless, irrespective of the high Cr concentration in the soil, AM association
was present in many plant species suggesting the avoidance or tolerance of AM fungi to
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

55
heavy metal stress (Leyval et al., 1997). High rate of AM colonization was also seen in
wheatgrass examined from a subalpine coalmine soil and in oil sand tailings (Zak and
Parkinson, 1982). The maximum RTC in Site-C occurred in M. scandens (65.35%) which
exceeded the maximum colonization levels recorded in R. communis (61.30%) at
Site-NC. This is in line with the findings of Khade and Adholeya (2009) where 100% of
the root length was colonized by AM fungi in Parthenium sp growing on a tannery
effluent contaminated soil. The highly mycorrhizal plants would have managed to escape
the high metal enriched soil micropockets in Site-C as high rate of AM colonization was
seen in Agrostis capillaris on a Zn and Cd contaminated site close to a zinc refinery
(Griffieon et al., 1994). Oxalis acetosella examined from a multiple HM in (Cd-Zn, Pb)
contaminated soil had higher colonization levels than those examined from
uncontaminated soil (Turnau et al., 1996). Weissenhorn et al. (1995a, b) also reported
high rate of AM colonization in plants collected from soil that received multiple metal
contaminations. The slightly alkaline nature of the contaminated soil may also
immobilize Cr to certain extent, making it less bioavailable to the plants (Aydinalp and
Marinova, 2003) thus reducing its toxicity.
The lack of AM colonization in the roots of certain plant species like A. spinosus,
A. viridis, Cleome viscosa, Colocasia esculenta, Cyperus stoloniferus in both the sites
suggests the influence of host specificity, soil pH, presence, type, availability of AM
inoculum and nutrient levels (Friese and Allen 1991; Sanders and Fitter 1992a) other than
the role of HM pollution on the formation of AM fungal association.
In the present study, the members of Poaceae either had moderate levels of AM
fungal colonization or lacked it except for Eragrostis japonica from Site-C. An examination
on the mycorrhizal status of plants growing on tannery effluent contaminated soils by
Khade and Adholeya (2009), revealed that Cynodon dactylon had higher AM colonization in
contaminated than in non contaminated soil (29.50% vs 10%). However, in the present
investigation C. dactylon (8.87%) had low rate of AM fungal colonization at Site C.
This is contrary to the results of Soares and Siqueira (2008) where grasses were found to
be tolerant to HM stress as evidenced by AM colonization rates as shown for Brachiaria
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

56
decumbens in multi-contaminated soil (Zn, Cd, and Cu). Some indications also exist on
the significance of AM fungi in the phytoextraction of HMs like Pb from contaminated
soil by vetiver grass (Chrysopogon zizanioids) (Pumamiya et al., 2010)
Only E. prostrata and Artemisia japonica lacked AM association among the six
members examined from Asteraceae. Many hyper accumulating plants are known to
occur in Asteraceae and their HM tolerance could be attributed to the rhizospheric
microorganisms like AM which helps the plants to absorb nutrient or immobilize the
HMs (Turnau and Mesjasz-Przybylowicz, 2003). The predominance of Amaranthaceae
members at Site-C could not be attributed to the AM fungi, but to other factors like high
organic content and nutrients. The high crop yield due to the improved physico-chemical
conditions of soil by the application of tannery effluents was reported by Munn et al. (2000).
Even though many studies have reported the role of AM fungi in the hyperacccumulating
capacity of pteridophytes like Pteris vittata (Cai et al., 2004; Leung et al., 2006), the
absence of AM association in pteridophytes in the present study except Acrostichum
speciosum could be because of the high Cr content in the soil. Studies have even shown
the enhanced uptake of arsenic in mycorrhizal P. vittata (88.1g Kg-1
) in comparison with
non mycorrhizal types (60.4 mg Kg-1
) (Leung et al., 2006).
In the present study Paris-type was more prevalent at Site-C. Likewise, in a multi
metal contaminated site (Zn, Cd, Pb) Paris-type AM morphology was found to be
more widespread while in non contaminated site Arum-type was more common
(Voges-Mikus et al., 2005). In contrast, a study by Khade and Adholeya (2009) showed
the prevalence of Arum-type AM morphology in both tannery effluent contaminated and
non contaminated sites. The slow growth rate specific to Paris –type AM morphology
(Yamato, 2004) is found to be the best strategy for the long term survival of plants
growing in high stress environments (Brundrett and Kendrick, 1990).
In the present investigation, both the sites harboured only spores of
Scutellospora calospora. Reports on single species of AM fungi were also reported from
soils which were highly contaminated with Zn, Cu, Pb, Ni, Cd and Mn (Griffioen, 1994).
The low species richness in both contaminated and non contaminated soils was also
reported by Pawlowska et al. (1996) and Regvar et al. (2001). The spore count decreased
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

57
as the total heavy metal (Zn, Cd, Pb, and Cu) concentration increases in Vertisol and
Leptosol soils (Ortega-Larrocea et al., 2001). Low spore numbers were observed in
contaminated soils when compared to less contaminated sites (Leyval et al., 1995).
In contrary, studies done by Khade and Adholeya (2009) showed higher AM spore
diversity in metal contaminated soil in comparison with non contaminated soil.
Moreover, the high P content of both the soils (Site-C and Site-NC) could also be
attributed for the low species richness (Peng et al., 2010).
Elevated levels of Cr resulted in low number of AM spores in sites filled with
tannery sludge when compared to Site-NC in the present study. Similarly del Val et al. (1999)
also showed that toxicity of heavy metals can reduce the number of spores in the
rhizospheric soil. Despite the excessive heavy metal contamination there are several
reports on high species diversity of AM fungi in metal contaminated soil like arsenic
(Gonzalez-Chavez et al., 2002) and magnesite mine spoils in India (Raman et al., 1993).
Low spore density did not affect the percentage colonization of AM fungi in the
present study. This corroborates with the findings of Khade and Adholeya (2009),
Del Val et al. (1999) and Jacquot-Plumey et al. (2001) where low spore number did not
have any negative effect on the development of AM fungal structures. Regvar et al. (2001)
and Whitfield et al. (2003) also reported that the AM fungal development never suffered
from the less abundant nature of AM fungal spores. This could be due to the favourable
environmental conditions for the germination of the AM spores resulting in rapid
colonization and low spore number (Khade and Adholeya, 2009) as spore germination is
less susceptible than any other facet of AM fungal life cycle (Pawlowska and
Charvat, 2007). The significant positive correlation between the spore numbers and
DSMO suggests that, the influence of environmental factors on these fungal variables are
directly proportional (Muthukumar and Vediyappan, 2010).
The high rate of DSE colonization revealed in the plant roots of Site-C
demonstrates that these fungal endophytes are tolerable to high Cr content in the soil.
This corroborates with the findings of Barrow et al. (1997) where high rate of DSE
colonization was seen in dominant shrubs and grasses in arid grasslands of south western
USA. Ban et al (2012) and Zhang et al. (2008) isolated DSE fungi from the roots of plant
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.

58
species grown in lead-zinc mine tailings and waste smelter site respectively which is a
clear indication of its tolerance towards HM stress. Muthukumar and Vediyappan (2010)
also reported the presence of DSE fungal colonization in >90% of the plant species
examined from pulp and paper mill effluent irrigated soil.
Symbiotic association of plants with the dark septate fungal endophytes provides
the plants with the tolerance to dwell with various stresses in the soil in comparison to
AM fungi (Christie and Kilpatrick, 1992; Muthukumar and Vediyappan, 2010). The significant
negative correlation between AM and DSE colonization in the present study falls in line
with the findings of Muthukumar and Vediyappan (2010) where an inverse relationship
between total root colonization of AM and DSE was reported in the plant species
collected from pulp and paper mill effluent irrigated soil. Scervino et al. (2009) also
recently reported the reduction of AM fungal development by the exudates of DSE fungi.
The exudates of Dreschlera sp. negatively influenced the development of extra matrical
phase of the AM fungus Gigaspora rosea (Scervino et al., 2009).
4.5. Conclusions
Understanding the impact of HMs on AM and DSE would be helpful in
developing more innovative, cost effective and successful strategies in minimizing metal
toxicity on plants and in soil, which could be implemented in various re-vegetation
practices. Further studies are necessary in order to confirm the ability of AM and DSE to
survive in high HM concentrations, as they could be possible candidates for
mycoremediation.
Please purchase PDF Split-Merge on www.verypdf.com to remove this watermark.