characterisation of a population of pratylenchus ... of a population of pratylenchus hippeastri from...
TRANSCRIPT

Nematology 2010 Vol 12(6) 847-868
Characterisation of a population of Pratylenchus hippeastri frombromeliads and description of two related new species
P floridensis n sp and P parafloridensis n spfrom grasses in Florida
Francesca DE LUCA 1 Alberto TROCCOLI 1lowast Larry W DUNCAN 2 Sergei A SUBBOTIN 34Lieven WAEYENBERGE 5 Maurice MOENS 56 and Renato N INSERRA 7
1 CNR Istituto per la Protezione delle Piante Via G Amendola 122D Bari 70126 Italy2 University of Florida IFAS Citrus Research and Education Center Department of Entomology Lake Alfred
FL 33850-2299 USA3 Plant Pest Diagnostic Center California Department of Food and Agriculture 3294 Meadowview Road Sacramento
CA 95832 USA4 Center of Parasitology of AN Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences
Leninskii Prospect 33 Moscow 117071 Russia5 Institute for Agricultural and Fisheries Research (ILVO) Burg Van Gansberghelaan 96 9820 Merelbeke Belgium
6 Gent University Laboratory for Agrozoology Coupure links 653 9000 Gent Belgium7 Florida Department of Agriculture and Consumer Services DPI Nematology Section PO Box 147100 Gainesville
FL 32614-7100 USA
Received 8 January 2010 revised 13 February 2010Accepted for publication 13 February 2010
Summary ndash Morphological and molecular analyses confirmed the presence of Pratylenchus hippeastri in regulatory samples collectedin commercial bromeliad operations from genera Guzmania Neoregelia and Vriesea in central and south Florida USA Specimens ofP hippeastri from bromeliads contained males which were not detected in the type population from amaryllis and are described hereinfor the first time The rDNA sequences of these males matched those of P hippeastri female type material Pratylenchus hippeastri androot-lesion nematodes from several hosts in Florida were characterised at the morphological and molecular level whereas other samplesfrom Russia and South Africa were characterised at the molecular level only Sequence and phylogenetic analysis using the ITS rRNAgene of these root-lesion nematodes revealed the presence of eight putative new species (spH1-H8) closely related to P hippeastriHere we describe two Florida representatives of the amphimictic root-lesion nematodes from Bahia grass (N1) and maidencane (N2)previously characterised by Inserra et al in 1996 and Duncan et al in 1999 as two new species phylogenetically related to P hippeastriand named P floridensis n sp and P parafloridensis n sp respectively The small round or oval (rarely rectangular and occasionallyoblong) and enlarged spermatheca and the bluntly pointed or subacute tail with smooth and occasionally indented terminus separateP floridensis n sp from P parafloridensis n sp which has a quadrangular spermatheca and a subhemispherical or bluntly pointed tailwith generally smooth and rarely indented terminus However these characters may overlap in some specimens making a morphologicalseparation problematic without the use of molecular analysis The close phylogenetic relationships shared by the species characterisedin this study indicate that they are representatives of a P hippeastri species complex
Keywords ndash Bahia grass bottlebrush Callistemon rigidus Fraxinus caroliniana maidencane molecular morphology morphometricsPanicum hemitomon Paspalum notatum phylogeny pop ash species complex St Augustine grass taxonomy
In the early 1990s two amphimictic populations ofroot-lesion nematodes were found on Bahia grass (Pas-palum notatum Flueggeacute) (N1) and maidencane (Panicum
lowast Corresponding author e-mail atroccolibaippcnrit
hemitomon Schultes) (N2) in Florida USA The popula-tions have regulatory significance because they are mor-phologically similar to Pratylenchus coffeae (Zimmer-
copy Koninklijke Brill NV Leiden 2010 DOI101163138855410X495809Also available online - wwwbrillnlnemy 847
F De Luca et al
mann 1898) Filipjev amp Schuurmans Stekhoven 1941which is a regulated nematode in Florida Morpho-biological studies of these two populations provided evi-dence that they share similarities (undivided smooth facewith two lip annuli and prominent spermatheca filled withsperm) with P loosi Loof 1960 a species morphologi-cally closely related to P coffeae In spite of the fact thatthese populations have a more anterior vulva (75-80)than P loosi (79-85) they were tentatively identified asFlorida populations of P loosi (Inserra et al 1996) Sub-sequent molecular analyses of these populations showedthat they were phylogenetically unrelated to both P cof-feae and P loosi (Duncan et al 1999) and consisted oftwo morphologically similar species of root-lesion ne-matodes designated as Pratylenchus N1 and N2 (N1 =P loosi Zolfo Springs and N2 = P loosi Lithia in Table 2)Detailed morphological characteristics were given by In-serra et al (1996) and Duncan et al (1999)
The recently described P hippeastri Inserra TroccoliGozel Bernard Dunn amp Duncan 2007 from amaryllisin Florida USA was characterised by absence of malesa flat and smooth face with two lip annuli a rectangularempty spermatheca a conoid tail with a bluntly pointedterminus and anterior vulva position (75-78) (Inserra etal 2007) Pratylenchus hippeastri clustered in a cladewith the amphimictic Pratylenchus N1 and N2 popula-tions from pasture grasses in Florida when compared byD2-D3 expansion segments of the 28S rDNA sequenceswith those of 32 Pratylenchus populations studied by In-serra et al (2007) The close phylogenetic relationshipsbased on D2-D3 between P hippeastri and PratylenchusN1 and N2 were confirmed by Subbotin et al (2008)who considered Pratylenchus N1 and N2 conspecific withP hippeastri in spite of the difference in their reproduc-tive behaviour and morphological differences
Recently root-lesion nematodes with few males andnumerous females morphologically similar to P hippeas-tri were detected in regulatory samples collected in com-mercial bromeliad operations in central and south FloridaBromeliads (Bromeliaceae) are ornamental epiphytes thatproduce roots that anchor the plant to branches and twigsof trees but also take up nutrients when in contact with orgrown in soil and other media In spite of the presence ofmales unknown from the type population of P hippeastrithese root-lesion nematodes from bromeliads share ma-jor diagnostic morphological features with P hippeastriAdditionally male-less and amphimictic root-lesion ne-matodes with two or occasionally three lip annuli andmorphologically similar to P hippeastri and N1 and N2
occur in Florida on turf grasses ornamentals and na-tive trees However the taxonomic status of these cryp-tic species (morphologically similar but genetically dif-ferent) as well that of N1 and N2 and those infectingbromeliads is uncertain and requires clarification
The main objectives of this study were to i) charac-terise molecularly and morphologically populations re-sembling P hippeastri from bromeliads and confirm theirspecies identity ii) provide updated molecular and mor-phological data on the root-lesion nematodes N1 and N2and describe them as two new species and iii) recon-struct phylogenetic relationships between P hippeastriand N1 and N2 along with other closely related speciesfrom Florida Russia and South Africa using the ITSand D2-D3 expansion segments of 28S rRNA gene se-quences
Materials and methods
ROOT-LESION NEMATODE POPULATIONS FROM
BROMELIADS GRASSES AND OTHER PLANTS
Three bromeliad production operations located nearApopka and Miami Florida were surveyed during all sea-sons in 2007-2008 Sixty composite root and soil sampleswere collected from containerised bromeliads (Table 1)in all of the production operations Ten root-lesion nema-tode females (originally from Goulds south Florida) werehand picked from infested bromeliad samples and trans-ferred to carrot disks at 23C (Huettel 1985) Culturednematodes were used for molecular and morphologicalanalyses and sex ratio determination
Pratylenchus hippeastri from amaryllis and relatedmale-less root-lesion nematodes from bromeliads andother hosts in Florida (spH5 spH7 and spH8) tentativelyidentified morphologically as representative of P hippeas-tri P zeae Graham 1951 and P jordanensis Hashim1983 (= P zeae) (Table 2) were selected for this study
Table 1 Bromeliad species and cultivars sampled in Floridalowast
Genus Cultivar
Guzmania Confetti Eloy Intro Indian night Irene MarjanOptimalowast Orange Ostara
Neoregelia Passionlowast Ardielowast Franklowast Infernolowast MartinlowastTricolorlowast
Vriesea splowast ndash
Bromeliads marked by an asterisk were infected by root lesionnematodes
848 Nematology
Pratylenchus hippeastri group from Florida
Table 2 Pratylenchus species and populations used in this study
Identification based Preliminary Host plant Locality Collection codes GenBank GenBank Source ofon ITS and D2-D3 identification for DNA or accession accession materials orrDNA sequences based on nematode number number for reference
morphology cultures for ITS D2-D3 of 28SrDNA
P hippeastri P hippeastri Amaryllis Tampa Hillsborough PhippTampa FJ712932- GU214112 L Duncan(Hippeastrum sp) County FL USA FJ712936 GU214113
P hippeastri P hippeastri Bromeliads Goulds Dade County FloridaPh N554883- N554879- L Duncan(Neoregelia spp) FL USA N554887 N554882
P hippeastri P hippeastri Amaryllis Gainesville Alachua FloridaPh FN554888 DQ498829 Inserra et al(Hippeastrum sp) County FL USA FN554889 DQ498831 (2007)
P parafloridensis P loosi Maidencane Lithia Hillsborough Ploosi Lithia GQ988377 AF170438 L Duncann sp Pratylenchus N2 (Panicum County FL USA GQ988378 GU214114
hemitomon) GU214115P floridensis n sp P loosi Bahia grass Zolfo Springs Hardee PloosiZolfoN1 GQ988375 AF170437 L Duncan
Pratylenchus N1 (Paspalum notatum) County FL USA GQ988376 GU214116GU214117
Pratylenchus spH1 Pratylenchus sp Pop ash (Fraxinus Perry Taylor county CD580 GU131132- GU131127- R Inserracaroliniana) FL USA GU131135 GU131129
Pratylenchus spH2 P zeae Unknown Upington South-Africa PzUping FJ713012- GU214121 E Van den BergFJ713016 GU214122
Pratylenchus spH3 P subranjani Grassland Russia PsubMi8 GQ988369 ndash A RyssGQ988370
Pratylenchus spH4 P scribneri Corn (Zea mays) Florida USA PscribFloridaUSA FJ712997- ndash J PinochetFJ713001
Pratylenchus spH5 P hippeastri Bottlebrush Hastings St John CD544 GU131136 GU131130 R Inserra(Callistemon rigidus) County FL USA GU131137 GU131131
Pratylenchus spH6 Pratylenchus sp St Augustine grass Arcadia De Soto County CD547 CD548 GU131138- GU131123- R Inserra(Stenotaphrum FL USA GU131141 GU131126secundatum)
Pratylenchus spH7 P zeae Turf Florida USA PzInserra GQ988371 GU214123 L DuncanGQ988372 GU214124
Pratylenchus spH8 P jordanensis Grassland La Belle Hendry County PjordInserra GQ988373 GU214118- L DuncanP zeae FL USA GQ988374 GU214120
P jaehni P jaehni Citrus (Citrus Sao Paolo Brasil Pjaehni FJ712937- AF170426 L Duncanaurantium) FJ712941 AF170427
Two additional Florida amphimictic root-lesion nema-todes (spH1 and spH6) similar to N1 and N2 identified asPratylenchus sp (Table 2) were also included for molec-ular analysis only along with two other samples identifiedas P subranjani Mizukubo Toida Keereewan amp Yoshida1990 and P zeae (spH3 spH2) from Russia and SouthAfrica respectively and a Florida population with dividedface identified as P scribneri Steiner in Sherbakoff ampStanley 1943 (spH4) (Hernaacutendez et al 2000) (Table 2)Nematodes were extracted from soil by the sieving de-canting and centrifugal flotation method (Jenkins 1964)and from bromeliad bottlebrush pop ash St Augustinegrass and mixed species of turf grass roots by incubationin jars
MOLECULAR ANALYSIS
DNA was extracted from individuals of both female andmale root-lesion nematode specimens DNA extractionfrom individual root-lesion nematode specimens PCRcloning and sequencing took place in three laborato-ries IPP Italy ILVO Belgium and PPDC CDFA USAThe protocols were described in detail by De Luca etal (2004) Waeyenberge et al (2009) and Subbotin etal (2008) respectively The following sets of primerswere used for amplification of two gene fragments in thepresent study i) D2-D3 expansion segments of 28S rRNAusing forward D2A (5prime-ACAAGTACCGTGGGGAAAGTTG-3prime) and reverse D3B (5prime-TCGGAAGGAACCAGCTACTA-3prime) and ii) ITS1-58-ITS2-rRNA using forward
Vol 12(6) 2010 849
F De Luca et al
18S-Int (5prime-CGTAACAAGGTAGCTGTAGG-3prime) and re-verse 26S-Int (5prime-TCCTCCGCTAAATGATAT-3prime) for-ward TW81 (5prime-GTTTCCGTAGGTGAACCTGC-3prime) andreverse AB28 (5prime-ATATGCTTAAGTTCAGCGGGT-3prime) orforward PRATTW81 (5prime-GTAGGTGAACCTGCTGCTG-3prime) and reverse AB28 PCR products were purified usingthe protocol listed by the manufacturers of NucleospinExtract II (Macherey-Nagel Germany) or QIAquick (Qi-agen USA) gel extraction kits and used for cloning or di-rect sequencing in both directions with the primers givenabove or M13 forward and M13 reverse primers TOPO-TA cloning kit (Invitrogen) or pGEM-T Vector System IIkit (Promega USA) were used for cloning of PCR prod-ucts Newly obtained sequences were deposited in Gen-Bank under accession numbers given in Table 2
The newly obtained sequences for both ribosomal re-gions of P hippeastri from bromeliad and of root-lesionnematodes from other hosts including those from amaryl-lis Bahia grass (N1) maidencane (N2) (Table 2) and ofP jaehni Inserra Duncan Troccoli Dunn dos SantosKaplan amp Vovlas 2001 (Duncan et al 1999 Waeyen-berge unpubl) were aligned using ClustalW (Thompsonet al 1997) with default parameters Pratylenchus jaehniwas used as an outgroup taxon (Subbotin et al 2008)Phylogenetic analysis of the sequence data sets were per-formed with maximum parsimony (MP) using PAUPlowast4b10 (Swofford 2002) and Bayesian inference (BI) usingMrBayes 312 (Huelsenbeck amp Ronquist 2001) For MPwe used heuristic search setting with ten replicates of ran-dom taxon addition tree bisection-reconnection branchswapping to seek the most parsimonious trees Gaps weretreated as missing data To obtain an estimation of the sup-port for each node a bootstrap analysis (BS) with 1000replicates was done BI analysis under the GTR + I + Gmodel for each gene was initiated with a random startingtree and was run with four chains for 10 times 106 genera-tions The Markov chains were sampled at intervals of 100generations Two runs were performed for each analysisThe log-likelihood values of the sample points stabilisedafter approximately 103 generations The topologies wereused to generate a 50 majority rule consensus tree Pos-terior probabilities (PP) are given on appropriate clades
MORPHOMETRIC AND MORPHOLOGICAL ANALYSIS
Adult root-lesion nematodes from bromeliads wereused for this study Live specimens were immobilised bygently heating and then mounted in water agar on a slide(Esser 1986) for measurements and photographs Addi-tional measurements and drawings were made from spec-
imens killed and fixed in hot aqueous 2 formaldehyde +1 propionic acid dehydrated in ethanol vapour andmounted in dehydrated glycerin (Hooper 1970) Mea-surements of specimens were made with an ocular mi-crometer and drawings with a camera lucida Abbrevi-ations used are defined in Siddiqi (2000) Photographswere taken with two Leica (Wild MPS 4652 and LeicaDFC 320) cameras mounted on Nikon (Optiphot) and Le-ica DM 2500 compound microscopes
The morphological information on the root-lesion ne-matodes from Bahia grass (N1) and maidencane (N2) pro-vided by Inserra et al (1996) and Duncan et al (1999)was complemented by further microscopic observationsof additional preserved specimens kept in the nematodecollection (CNR-IPP Bari) by the second author of thispaper Morphometrics of mature females of root-lesionnematode species studied by Duncan et al (1999) andP hippeastri from bromeliad and the original descrip-tion were subjected to principal component analysis us-ing Minitab 13 (Minitab USA) The populations werecharacterised based on the lip morphology (smooth or di-vided) and the weakly-allometric charactersratios V aand stylet length These characters were reported to dis-criminate relationships among P coffeae-group speciesthat conformed closely to the phylogenetic relationshipsinferred by analyses of the D2-D3 region of the 28S rDNAsequence (Duncan et al 1999)
Specimens for scanning electron microscope (SEM)observations were cold-fixed in glutaraldehyde bufferedwith 01 M phosphate buffer (pH 72) post fixed 1 h in2 osmium tetroxide dehydrated in a graded series ofethanol critical point dried with CO2 and sputter coatedwith gold palladium (Eisenback 1985) Nematodes wereobserved with a Hitachi S530 microscope at 15-20 kVaccelerating voltage
SPECIES DELIMITATION IN STUDIED PRATYLENCHUS
Species delimitation of the studied populations wasdone by applying an integrated or polyphasic approachwhich was based on consideration of results of morpho-logical and morphometrical studies phylogenetic and se-quencing analysis and analysis of host plants and geo-graphic distribution of studied samples This approach in-tegrates any significant information on the organisms andresults in a consensus and transition type of classification(Subbotin amp Moens 2006) Two new species named hereas P floridensis n sp and P parafloridensis n sp andseveral unidentified putative new species defined here asPratylenchus spH1-H8 were delimited in this study using
850 Nematology
Pratylenchus hippeastri group from Florida
this approach More detailed morphological and molecu-lar analysis is still required to confirm the unique speciesstatus of Pratylenchus spH1-H8
Results
ROOT-LESION NEMATODE POPULATIONS COLLECTED
FROM BROMELIADS AND OTHER PLANTS
Bromeliads belonging to the genera Guzmania Ruizamp Pav Neoregelia L B Sm and Vriesea Lindl werefound to be infected by root-lesion nematodes similar toP hippeastri (Table 1) Population levels were usuallylt10 specimensg fresh roots In some cases bromeliadroots were found infected concomitantly with a few spec-imens of P brachyurus (Godfrey 1929) Filipjev amp Schu-urmans Stekhoven 1941 endoparasitic Helicotylenchusdihystera (Cobb 1893) Sher 1961 and Meloidogyne are-naria (Neal 1889) Chitwood 1949 Carrot cultures inoc-ulated with female root-lesion nematodes from bromeli-ads produced a large number of nematodes at differentlife stages and only a few males (usually three malesper 100 females) The population levels of other Floridaroot-lesion nematodes varied from 30 160 and 100 speci-mensg fresh roots for spH1 spH5 and spH6 respectivelyThe number of males in the amphimictic spH1 and spH6was ca 40100 females
MOLECULAR CHARACTERISATION OF P HIPPEASTRI
SPECIES COMPLEX
The amplification of the ITS containing region pro-duced a single fragment of ca 970-1000 bp for the studiedsamples The sequence alignment for P hippeastri and re-lated species with consensus sequence for each putativespecies is given in Figure 1 The ITS alignment included44 sequences and was 1050 bp in length Sequence di-versity within all studied root-lesion nematodes includingP jaehni reached 19 (174 nucleotides) for P hippeas-tri from bromeliad and from amaryllis it varied from 0 to06 (0-6 nucleotides) whereas sequence diversity withinthe other root-lesion nematodes related to P hippeastrireached 62 (57 nucleotides) Phylogenetic relationshipswithin Pratylenchus species as inferred from Bayesianinference are given in Figure 2 Four main moderate orhighly supported clades (PP = 90-100) were distinguishedwithin the tree Clade 1 grouped P hippeastri populationsalong with root-lesion nematodes spH1-H5 Populationsof P hippeastri from amaryllis and bromeliads clustered
together forming one highly supported (PP = 100) sub-clade within clade 1 The ITS sequences for PratylenchusspH2 and spH3 did not form distinct subclades and re-lationships between them were not resolved The root-lesion nematode N2 (= P parafloridensis n sp in Table 2and Figs 1-3) formed a moderately supported (PP = 90)clade 2 together with Pratylenchus spH6 Root-lesion ne-matodes spH7 and spH8 clustered together and were notwell separated The two sequences of N1 (= P floridensisn sp in Table 2 and Figs 1-3) formed highly supportedclade 4 at the basal position of the tree
Sequence alignments of the D2-D3 of 28S rDNA in-cluded 32 sequences of 713 bp length Sequence diver-sity reached 74 (47 nucleotides) for all root-lesion ne-matodes studied 21 (14 nucleotides) for the speciesclosely related to P hippeastri and varied from 0-03 (0-2 nucleotides) within P hippeastri populations Phyloge-netic relationships within Pratylenchus species based onD2-D3 of 28S rDNA sequences is given in Figure 3 Fivemain weakly to highly supported clades (PP = 72-100)were distinguished within the tree which corresponded tothe clades on the ITS-rRNA tree Populations of P hip-peastri from amaryllis and bromeliads formed a moder-ately supported (PP = 93) clade 1 together with Praty-lenchus spH1 (Florida USA) spH2 (South Africa) andspH5 (Florida USA) Root-lesion nematodes spH7 andspH8 clustered in a moderately supported clade 3 Theamphimictic N1 formed a highly supported clade 4 at thebasal position of the tree MP and BI analyses generatedcongruent trees with similar branch supports for the ITSand D2-D3 gene alignments respectively
The close phylogenetic relationships shared by thespecies (spH1-H8) characterised in this study indicate thatthey are representatives of a P hippeastri species com-plex These results confirmed the identity of males and fe-males of bromeliad populations as P hippeastri and pro-vided evidence that the rDNA sequences of P hippeas-tri males from bromeliads matched those of P hippeastrifemale type material Furthermore the phylogenetic find-ings also provided support for the description of N1 andN2 as two new species named P floridensis n sp andP parafloridensis n sp respectively
MORPHOLOGICAL CHARACTERISATION OF
PRATYLENCHUS HIPPEASTRI
Comparative measurements of females and males ofP hippeastri from bromeliads and those reported fromamaryllis in the original description are reported inTable 3
Vol 12(6) 2010 851
F De Luca et al
852 Nematology
Pratylenchus hippeastri group from Florida
Fig 2 The 50 majority rule consensus tree from Bayesian analysis generated from the ITS sequence dataset for the Pratylenchushippeastri species complex using the GTR + I + G model Posterior probability more than 70 is given for appropriate clades
Female
Morphometric values of the females from bromeliadsdid not differ from those of P hippeastri from amaryllisexcept for the tail length in fixed specimens which wasslightly shorter than that of females from amaryllis Smalldifferences were observed also in the mean values of some
characters of living specimens from bromeliad such asbody tail and post-uterine sac length which were shorterthan those of specimens from amaryllis Bromeliad fe-males also showed smaller maximum and vulval body dia-meter and a shorter vulva-anus distance However theirrange values overlapped Their lip pattern was also simi-lar to that of P hippeastri and consisted of a flat and un-
Fig 1 Sequence alignment of partial 18S complete ITS1 58S ITS2 and partial 28S rRNA for the Pratylenchus hippeastri speciescomplex The 18S 58S and 28S rRNA gene sequences are marked in bold the primers sequences are underlined Consensus sequenceis given for each species only Single letter code recommended by NC-IUB was used to specify nucleotide if two or more bases werepermitted at a particular position in a species subalignment Lower case symbols indicate presence of one or several gaps in a particularposition in sequences for a species subalignment
Vol 12(6) 2010 853
F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
mann 1898) Filipjev amp Schuurmans Stekhoven 1941which is a regulated nematode in Florida Morpho-biological studies of these two populations provided evi-dence that they share similarities (undivided smooth facewith two lip annuli and prominent spermatheca filled withsperm) with P loosi Loof 1960 a species morphologi-cally closely related to P coffeae In spite of the fact thatthese populations have a more anterior vulva (75-80)than P loosi (79-85) they were tentatively identified asFlorida populations of P loosi (Inserra et al 1996) Sub-sequent molecular analyses of these populations showedthat they were phylogenetically unrelated to both P cof-feae and P loosi (Duncan et al 1999) and consisted oftwo morphologically similar species of root-lesion ne-matodes designated as Pratylenchus N1 and N2 (N1 =P loosi Zolfo Springs and N2 = P loosi Lithia in Table 2)Detailed morphological characteristics were given by In-serra et al (1996) and Duncan et al (1999)
The recently described P hippeastri Inserra TroccoliGozel Bernard Dunn amp Duncan 2007 from amaryllisin Florida USA was characterised by absence of malesa flat and smooth face with two lip annuli a rectangularempty spermatheca a conoid tail with a bluntly pointedterminus and anterior vulva position (75-78) (Inserra etal 2007) Pratylenchus hippeastri clustered in a cladewith the amphimictic Pratylenchus N1 and N2 popula-tions from pasture grasses in Florida when compared byD2-D3 expansion segments of the 28S rDNA sequenceswith those of 32 Pratylenchus populations studied by In-serra et al (2007) The close phylogenetic relationshipsbased on D2-D3 between P hippeastri and PratylenchusN1 and N2 were confirmed by Subbotin et al (2008)who considered Pratylenchus N1 and N2 conspecific withP hippeastri in spite of the difference in their reproduc-tive behaviour and morphological differences
Recently root-lesion nematodes with few males andnumerous females morphologically similar to P hippeas-tri were detected in regulatory samples collected in com-mercial bromeliad operations in central and south FloridaBromeliads (Bromeliaceae) are ornamental epiphytes thatproduce roots that anchor the plant to branches and twigsof trees but also take up nutrients when in contact with orgrown in soil and other media In spite of the presence ofmales unknown from the type population of P hippeastrithese root-lesion nematodes from bromeliads share ma-jor diagnostic morphological features with P hippeastriAdditionally male-less and amphimictic root-lesion ne-matodes with two or occasionally three lip annuli andmorphologically similar to P hippeastri and N1 and N2
occur in Florida on turf grasses ornamentals and na-tive trees However the taxonomic status of these cryp-tic species (morphologically similar but genetically dif-ferent) as well that of N1 and N2 and those infectingbromeliads is uncertain and requires clarification
The main objectives of this study were to i) charac-terise molecularly and morphologically populations re-sembling P hippeastri from bromeliads and confirm theirspecies identity ii) provide updated molecular and mor-phological data on the root-lesion nematodes N1 and N2and describe them as two new species and iii) recon-struct phylogenetic relationships between P hippeastriand N1 and N2 along with other closely related speciesfrom Florida Russia and South Africa using the ITSand D2-D3 expansion segments of 28S rRNA gene se-quences
Materials and methods
ROOT-LESION NEMATODE POPULATIONS FROM
BROMELIADS GRASSES AND OTHER PLANTS
Three bromeliad production operations located nearApopka and Miami Florida were surveyed during all sea-sons in 2007-2008 Sixty composite root and soil sampleswere collected from containerised bromeliads (Table 1)in all of the production operations Ten root-lesion nema-tode females (originally from Goulds south Florida) werehand picked from infested bromeliad samples and trans-ferred to carrot disks at 23C (Huettel 1985) Culturednematodes were used for molecular and morphologicalanalyses and sex ratio determination
Pratylenchus hippeastri from amaryllis and relatedmale-less root-lesion nematodes from bromeliads andother hosts in Florida (spH5 spH7 and spH8) tentativelyidentified morphologically as representative of P hippeas-tri P zeae Graham 1951 and P jordanensis Hashim1983 (= P zeae) (Table 2) were selected for this study
Table 1 Bromeliad species and cultivars sampled in Floridalowast
Genus Cultivar
Guzmania Confetti Eloy Intro Indian night Irene MarjanOptimalowast Orange Ostara
Neoregelia Passionlowast Ardielowast Franklowast Infernolowast MartinlowastTricolorlowast
Vriesea splowast ndash
Bromeliads marked by an asterisk were infected by root lesionnematodes
848 Nematology
Pratylenchus hippeastri group from Florida
Table 2 Pratylenchus species and populations used in this study
Identification based Preliminary Host plant Locality Collection codes GenBank GenBank Source ofon ITS and D2-D3 identification for DNA or accession accession materials orrDNA sequences based on nematode number number for reference
morphology cultures for ITS D2-D3 of 28SrDNA
P hippeastri P hippeastri Amaryllis Tampa Hillsborough PhippTampa FJ712932- GU214112 L Duncan(Hippeastrum sp) County FL USA FJ712936 GU214113
P hippeastri P hippeastri Bromeliads Goulds Dade County FloridaPh N554883- N554879- L Duncan(Neoregelia spp) FL USA N554887 N554882
P hippeastri P hippeastri Amaryllis Gainesville Alachua FloridaPh FN554888 DQ498829 Inserra et al(Hippeastrum sp) County FL USA FN554889 DQ498831 (2007)
P parafloridensis P loosi Maidencane Lithia Hillsborough Ploosi Lithia GQ988377 AF170438 L Duncann sp Pratylenchus N2 (Panicum County FL USA GQ988378 GU214114
hemitomon) GU214115P floridensis n sp P loosi Bahia grass Zolfo Springs Hardee PloosiZolfoN1 GQ988375 AF170437 L Duncan
Pratylenchus N1 (Paspalum notatum) County FL USA GQ988376 GU214116GU214117
Pratylenchus spH1 Pratylenchus sp Pop ash (Fraxinus Perry Taylor county CD580 GU131132- GU131127- R Inserracaroliniana) FL USA GU131135 GU131129
Pratylenchus spH2 P zeae Unknown Upington South-Africa PzUping FJ713012- GU214121 E Van den BergFJ713016 GU214122
Pratylenchus spH3 P subranjani Grassland Russia PsubMi8 GQ988369 ndash A RyssGQ988370
Pratylenchus spH4 P scribneri Corn (Zea mays) Florida USA PscribFloridaUSA FJ712997- ndash J PinochetFJ713001
Pratylenchus spH5 P hippeastri Bottlebrush Hastings St John CD544 GU131136 GU131130 R Inserra(Callistemon rigidus) County FL USA GU131137 GU131131
Pratylenchus spH6 Pratylenchus sp St Augustine grass Arcadia De Soto County CD547 CD548 GU131138- GU131123- R Inserra(Stenotaphrum FL USA GU131141 GU131126secundatum)
Pratylenchus spH7 P zeae Turf Florida USA PzInserra GQ988371 GU214123 L DuncanGQ988372 GU214124
Pratylenchus spH8 P jordanensis Grassland La Belle Hendry County PjordInserra GQ988373 GU214118- L DuncanP zeae FL USA GQ988374 GU214120
P jaehni P jaehni Citrus (Citrus Sao Paolo Brasil Pjaehni FJ712937- AF170426 L Duncanaurantium) FJ712941 AF170427
Two additional Florida amphimictic root-lesion nema-todes (spH1 and spH6) similar to N1 and N2 identified asPratylenchus sp (Table 2) were also included for molec-ular analysis only along with two other samples identifiedas P subranjani Mizukubo Toida Keereewan amp Yoshida1990 and P zeae (spH3 spH2) from Russia and SouthAfrica respectively and a Florida population with dividedface identified as P scribneri Steiner in Sherbakoff ampStanley 1943 (spH4) (Hernaacutendez et al 2000) (Table 2)Nematodes were extracted from soil by the sieving de-canting and centrifugal flotation method (Jenkins 1964)and from bromeliad bottlebrush pop ash St Augustinegrass and mixed species of turf grass roots by incubationin jars
MOLECULAR ANALYSIS
DNA was extracted from individuals of both female andmale root-lesion nematode specimens DNA extractionfrom individual root-lesion nematode specimens PCRcloning and sequencing took place in three laborato-ries IPP Italy ILVO Belgium and PPDC CDFA USAThe protocols were described in detail by De Luca etal (2004) Waeyenberge et al (2009) and Subbotin etal (2008) respectively The following sets of primerswere used for amplification of two gene fragments in thepresent study i) D2-D3 expansion segments of 28S rRNAusing forward D2A (5prime-ACAAGTACCGTGGGGAAAGTTG-3prime) and reverse D3B (5prime-TCGGAAGGAACCAGCTACTA-3prime) and ii) ITS1-58-ITS2-rRNA using forward
Vol 12(6) 2010 849
F De Luca et al
18S-Int (5prime-CGTAACAAGGTAGCTGTAGG-3prime) and re-verse 26S-Int (5prime-TCCTCCGCTAAATGATAT-3prime) for-ward TW81 (5prime-GTTTCCGTAGGTGAACCTGC-3prime) andreverse AB28 (5prime-ATATGCTTAAGTTCAGCGGGT-3prime) orforward PRATTW81 (5prime-GTAGGTGAACCTGCTGCTG-3prime) and reverse AB28 PCR products were purified usingthe protocol listed by the manufacturers of NucleospinExtract II (Macherey-Nagel Germany) or QIAquick (Qi-agen USA) gel extraction kits and used for cloning or di-rect sequencing in both directions with the primers givenabove or M13 forward and M13 reverse primers TOPO-TA cloning kit (Invitrogen) or pGEM-T Vector System IIkit (Promega USA) were used for cloning of PCR prod-ucts Newly obtained sequences were deposited in Gen-Bank under accession numbers given in Table 2
The newly obtained sequences for both ribosomal re-gions of P hippeastri from bromeliad and of root-lesionnematodes from other hosts including those from amaryl-lis Bahia grass (N1) maidencane (N2) (Table 2) and ofP jaehni Inserra Duncan Troccoli Dunn dos SantosKaplan amp Vovlas 2001 (Duncan et al 1999 Waeyen-berge unpubl) were aligned using ClustalW (Thompsonet al 1997) with default parameters Pratylenchus jaehniwas used as an outgroup taxon (Subbotin et al 2008)Phylogenetic analysis of the sequence data sets were per-formed with maximum parsimony (MP) using PAUPlowast4b10 (Swofford 2002) and Bayesian inference (BI) usingMrBayes 312 (Huelsenbeck amp Ronquist 2001) For MPwe used heuristic search setting with ten replicates of ran-dom taxon addition tree bisection-reconnection branchswapping to seek the most parsimonious trees Gaps weretreated as missing data To obtain an estimation of the sup-port for each node a bootstrap analysis (BS) with 1000replicates was done BI analysis under the GTR + I + Gmodel for each gene was initiated with a random startingtree and was run with four chains for 10 times 106 genera-tions The Markov chains were sampled at intervals of 100generations Two runs were performed for each analysisThe log-likelihood values of the sample points stabilisedafter approximately 103 generations The topologies wereused to generate a 50 majority rule consensus tree Pos-terior probabilities (PP) are given on appropriate clades
MORPHOMETRIC AND MORPHOLOGICAL ANALYSIS
Adult root-lesion nematodes from bromeliads wereused for this study Live specimens were immobilised bygently heating and then mounted in water agar on a slide(Esser 1986) for measurements and photographs Addi-tional measurements and drawings were made from spec-
imens killed and fixed in hot aqueous 2 formaldehyde +1 propionic acid dehydrated in ethanol vapour andmounted in dehydrated glycerin (Hooper 1970) Mea-surements of specimens were made with an ocular mi-crometer and drawings with a camera lucida Abbrevi-ations used are defined in Siddiqi (2000) Photographswere taken with two Leica (Wild MPS 4652 and LeicaDFC 320) cameras mounted on Nikon (Optiphot) and Le-ica DM 2500 compound microscopes
The morphological information on the root-lesion ne-matodes from Bahia grass (N1) and maidencane (N2) pro-vided by Inserra et al (1996) and Duncan et al (1999)was complemented by further microscopic observationsof additional preserved specimens kept in the nematodecollection (CNR-IPP Bari) by the second author of thispaper Morphometrics of mature females of root-lesionnematode species studied by Duncan et al (1999) andP hippeastri from bromeliad and the original descrip-tion were subjected to principal component analysis us-ing Minitab 13 (Minitab USA) The populations werecharacterised based on the lip morphology (smooth or di-vided) and the weakly-allometric charactersratios V aand stylet length These characters were reported to dis-criminate relationships among P coffeae-group speciesthat conformed closely to the phylogenetic relationshipsinferred by analyses of the D2-D3 region of the 28S rDNAsequence (Duncan et al 1999)
Specimens for scanning electron microscope (SEM)observations were cold-fixed in glutaraldehyde bufferedwith 01 M phosphate buffer (pH 72) post fixed 1 h in2 osmium tetroxide dehydrated in a graded series ofethanol critical point dried with CO2 and sputter coatedwith gold palladium (Eisenback 1985) Nematodes wereobserved with a Hitachi S530 microscope at 15-20 kVaccelerating voltage
SPECIES DELIMITATION IN STUDIED PRATYLENCHUS
Species delimitation of the studied populations wasdone by applying an integrated or polyphasic approachwhich was based on consideration of results of morpho-logical and morphometrical studies phylogenetic and se-quencing analysis and analysis of host plants and geo-graphic distribution of studied samples This approach in-tegrates any significant information on the organisms andresults in a consensus and transition type of classification(Subbotin amp Moens 2006) Two new species named hereas P floridensis n sp and P parafloridensis n sp andseveral unidentified putative new species defined here asPratylenchus spH1-H8 were delimited in this study using
850 Nematology
Pratylenchus hippeastri group from Florida
this approach More detailed morphological and molecu-lar analysis is still required to confirm the unique speciesstatus of Pratylenchus spH1-H8
Results
ROOT-LESION NEMATODE POPULATIONS COLLECTED
FROM BROMELIADS AND OTHER PLANTS
Bromeliads belonging to the genera Guzmania Ruizamp Pav Neoregelia L B Sm and Vriesea Lindl werefound to be infected by root-lesion nematodes similar toP hippeastri (Table 1) Population levels were usuallylt10 specimensg fresh roots In some cases bromeliadroots were found infected concomitantly with a few spec-imens of P brachyurus (Godfrey 1929) Filipjev amp Schu-urmans Stekhoven 1941 endoparasitic Helicotylenchusdihystera (Cobb 1893) Sher 1961 and Meloidogyne are-naria (Neal 1889) Chitwood 1949 Carrot cultures inoc-ulated with female root-lesion nematodes from bromeli-ads produced a large number of nematodes at differentlife stages and only a few males (usually three malesper 100 females) The population levels of other Floridaroot-lesion nematodes varied from 30 160 and 100 speci-mensg fresh roots for spH1 spH5 and spH6 respectivelyThe number of males in the amphimictic spH1 and spH6was ca 40100 females
MOLECULAR CHARACTERISATION OF P HIPPEASTRI
SPECIES COMPLEX
The amplification of the ITS containing region pro-duced a single fragment of ca 970-1000 bp for the studiedsamples The sequence alignment for P hippeastri and re-lated species with consensus sequence for each putativespecies is given in Figure 1 The ITS alignment included44 sequences and was 1050 bp in length Sequence di-versity within all studied root-lesion nematodes includingP jaehni reached 19 (174 nucleotides) for P hippeas-tri from bromeliad and from amaryllis it varied from 0 to06 (0-6 nucleotides) whereas sequence diversity withinthe other root-lesion nematodes related to P hippeastrireached 62 (57 nucleotides) Phylogenetic relationshipswithin Pratylenchus species as inferred from Bayesianinference are given in Figure 2 Four main moderate orhighly supported clades (PP = 90-100) were distinguishedwithin the tree Clade 1 grouped P hippeastri populationsalong with root-lesion nematodes spH1-H5 Populationsof P hippeastri from amaryllis and bromeliads clustered
together forming one highly supported (PP = 100) sub-clade within clade 1 The ITS sequences for PratylenchusspH2 and spH3 did not form distinct subclades and re-lationships between them were not resolved The root-lesion nematode N2 (= P parafloridensis n sp in Table 2and Figs 1-3) formed a moderately supported (PP = 90)clade 2 together with Pratylenchus spH6 Root-lesion ne-matodes spH7 and spH8 clustered together and were notwell separated The two sequences of N1 (= P floridensisn sp in Table 2 and Figs 1-3) formed highly supportedclade 4 at the basal position of the tree
Sequence alignments of the D2-D3 of 28S rDNA in-cluded 32 sequences of 713 bp length Sequence diver-sity reached 74 (47 nucleotides) for all root-lesion ne-matodes studied 21 (14 nucleotides) for the speciesclosely related to P hippeastri and varied from 0-03 (0-2 nucleotides) within P hippeastri populations Phyloge-netic relationships within Pratylenchus species based onD2-D3 of 28S rDNA sequences is given in Figure 3 Fivemain weakly to highly supported clades (PP = 72-100)were distinguished within the tree which corresponded tothe clades on the ITS-rRNA tree Populations of P hip-peastri from amaryllis and bromeliads formed a moder-ately supported (PP = 93) clade 1 together with Praty-lenchus spH1 (Florida USA) spH2 (South Africa) andspH5 (Florida USA) Root-lesion nematodes spH7 andspH8 clustered in a moderately supported clade 3 Theamphimictic N1 formed a highly supported clade 4 at thebasal position of the tree MP and BI analyses generatedcongruent trees with similar branch supports for the ITSand D2-D3 gene alignments respectively
The close phylogenetic relationships shared by thespecies (spH1-H8) characterised in this study indicate thatthey are representatives of a P hippeastri species com-plex These results confirmed the identity of males and fe-males of bromeliad populations as P hippeastri and pro-vided evidence that the rDNA sequences of P hippeas-tri males from bromeliads matched those of P hippeastrifemale type material Furthermore the phylogenetic find-ings also provided support for the description of N1 andN2 as two new species named P floridensis n sp andP parafloridensis n sp respectively
MORPHOLOGICAL CHARACTERISATION OF
PRATYLENCHUS HIPPEASTRI
Comparative measurements of females and males ofP hippeastri from bromeliads and those reported fromamaryllis in the original description are reported inTable 3
Vol 12(6) 2010 851
F De Luca et al
852 Nematology
Pratylenchus hippeastri group from Florida
Fig 2 The 50 majority rule consensus tree from Bayesian analysis generated from the ITS sequence dataset for the Pratylenchushippeastri species complex using the GTR + I + G model Posterior probability more than 70 is given for appropriate clades
Female
Morphometric values of the females from bromeliadsdid not differ from those of P hippeastri from amaryllisexcept for the tail length in fixed specimens which wasslightly shorter than that of females from amaryllis Smalldifferences were observed also in the mean values of some
characters of living specimens from bromeliad such asbody tail and post-uterine sac length which were shorterthan those of specimens from amaryllis Bromeliad fe-males also showed smaller maximum and vulval body dia-meter and a shorter vulva-anus distance However theirrange values overlapped Their lip pattern was also simi-lar to that of P hippeastri and consisted of a flat and un-
Fig 1 Sequence alignment of partial 18S complete ITS1 58S ITS2 and partial 28S rRNA for the Pratylenchus hippeastri speciescomplex The 18S 58S and 28S rRNA gene sequences are marked in bold the primers sequences are underlined Consensus sequenceis given for each species only Single letter code recommended by NC-IUB was used to specify nucleotide if two or more bases werepermitted at a particular position in a species subalignment Lower case symbols indicate presence of one or several gaps in a particularposition in sequences for a species subalignment
Vol 12(6) 2010 853
F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
Table 2 Pratylenchus species and populations used in this study
Identification based Preliminary Host plant Locality Collection codes GenBank GenBank Source ofon ITS and D2-D3 identification for DNA or accession accession materials orrDNA sequences based on nematode number number for reference
morphology cultures for ITS D2-D3 of 28SrDNA
P hippeastri P hippeastri Amaryllis Tampa Hillsborough PhippTampa FJ712932- GU214112 L Duncan(Hippeastrum sp) County FL USA FJ712936 GU214113
P hippeastri P hippeastri Bromeliads Goulds Dade County FloridaPh N554883- N554879- L Duncan(Neoregelia spp) FL USA N554887 N554882
P hippeastri P hippeastri Amaryllis Gainesville Alachua FloridaPh FN554888 DQ498829 Inserra et al(Hippeastrum sp) County FL USA FN554889 DQ498831 (2007)
P parafloridensis P loosi Maidencane Lithia Hillsborough Ploosi Lithia GQ988377 AF170438 L Duncann sp Pratylenchus N2 (Panicum County FL USA GQ988378 GU214114
hemitomon) GU214115P floridensis n sp P loosi Bahia grass Zolfo Springs Hardee PloosiZolfoN1 GQ988375 AF170437 L Duncan
Pratylenchus N1 (Paspalum notatum) County FL USA GQ988376 GU214116GU214117
Pratylenchus spH1 Pratylenchus sp Pop ash (Fraxinus Perry Taylor county CD580 GU131132- GU131127- R Inserracaroliniana) FL USA GU131135 GU131129
Pratylenchus spH2 P zeae Unknown Upington South-Africa PzUping FJ713012- GU214121 E Van den BergFJ713016 GU214122
Pratylenchus spH3 P subranjani Grassland Russia PsubMi8 GQ988369 ndash A RyssGQ988370
Pratylenchus spH4 P scribneri Corn (Zea mays) Florida USA PscribFloridaUSA FJ712997- ndash J PinochetFJ713001
Pratylenchus spH5 P hippeastri Bottlebrush Hastings St John CD544 GU131136 GU131130 R Inserra(Callistemon rigidus) County FL USA GU131137 GU131131
Pratylenchus spH6 Pratylenchus sp St Augustine grass Arcadia De Soto County CD547 CD548 GU131138- GU131123- R Inserra(Stenotaphrum FL USA GU131141 GU131126secundatum)
Pratylenchus spH7 P zeae Turf Florida USA PzInserra GQ988371 GU214123 L DuncanGQ988372 GU214124
Pratylenchus spH8 P jordanensis Grassland La Belle Hendry County PjordInserra GQ988373 GU214118- L DuncanP zeae FL USA GQ988374 GU214120
P jaehni P jaehni Citrus (Citrus Sao Paolo Brasil Pjaehni FJ712937- AF170426 L Duncanaurantium) FJ712941 AF170427
Two additional Florida amphimictic root-lesion nema-todes (spH1 and spH6) similar to N1 and N2 identified asPratylenchus sp (Table 2) were also included for molec-ular analysis only along with two other samples identifiedas P subranjani Mizukubo Toida Keereewan amp Yoshida1990 and P zeae (spH3 spH2) from Russia and SouthAfrica respectively and a Florida population with dividedface identified as P scribneri Steiner in Sherbakoff ampStanley 1943 (spH4) (Hernaacutendez et al 2000) (Table 2)Nematodes were extracted from soil by the sieving de-canting and centrifugal flotation method (Jenkins 1964)and from bromeliad bottlebrush pop ash St Augustinegrass and mixed species of turf grass roots by incubationin jars
MOLECULAR ANALYSIS
DNA was extracted from individuals of both female andmale root-lesion nematode specimens DNA extractionfrom individual root-lesion nematode specimens PCRcloning and sequencing took place in three laborato-ries IPP Italy ILVO Belgium and PPDC CDFA USAThe protocols were described in detail by De Luca etal (2004) Waeyenberge et al (2009) and Subbotin etal (2008) respectively The following sets of primerswere used for amplification of two gene fragments in thepresent study i) D2-D3 expansion segments of 28S rRNAusing forward D2A (5prime-ACAAGTACCGTGGGGAAAGTTG-3prime) and reverse D3B (5prime-TCGGAAGGAACCAGCTACTA-3prime) and ii) ITS1-58-ITS2-rRNA using forward
Vol 12(6) 2010 849
F De Luca et al
18S-Int (5prime-CGTAACAAGGTAGCTGTAGG-3prime) and re-verse 26S-Int (5prime-TCCTCCGCTAAATGATAT-3prime) for-ward TW81 (5prime-GTTTCCGTAGGTGAACCTGC-3prime) andreverse AB28 (5prime-ATATGCTTAAGTTCAGCGGGT-3prime) orforward PRATTW81 (5prime-GTAGGTGAACCTGCTGCTG-3prime) and reverse AB28 PCR products were purified usingthe protocol listed by the manufacturers of NucleospinExtract II (Macherey-Nagel Germany) or QIAquick (Qi-agen USA) gel extraction kits and used for cloning or di-rect sequencing in both directions with the primers givenabove or M13 forward and M13 reverse primers TOPO-TA cloning kit (Invitrogen) or pGEM-T Vector System IIkit (Promega USA) were used for cloning of PCR prod-ucts Newly obtained sequences were deposited in Gen-Bank under accession numbers given in Table 2
The newly obtained sequences for both ribosomal re-gions of P hippeastri from bromeliad and of root-lesionnematodes from other hosts including those from amaryl-lis Bahia grass (N1) maidencane (N2) (Table 2) and ofP jaehni Inserra Duncan Troccoli Dunn dos SantosKaplan amp Vovlas 2001 (Duncan et al 1999 Waeyen-berge unpubl) were aligned using ClustalW (Thompsonet al 1997) with default parameters Pratylenchus jaehniwas used as an outgroup taxon (Subbotin et al 2008)Phylogenetic analysis of the sequence data sets were per-formed with maximum parsimony (MP) using PAUPlowast4b10 (Swofford 2002) and Bayesian inference (BI) usingMrBayes 312 (Huelsenbeck amp Ronquist 2001) For MPwe used heuristic search setting with ten replicates of ran-dom taxon addition tree bisection-reconnection branchswapping to seek the most parsimonious trees Gaps weretreated as missing data To obtain an estimation of the sup-port for each node a bootstrap analysis (BS) with 1000replicates was done BI analysis under the GTR + I + Gmodel for each gene was initiated with a random startingtree and was run with four chains for 10 times 106 genera-tions The Markov chains were sampled at intervals of 100generations Two runs were performed for each analysisThe log-likelihood values of the sample points stabilisedafter approximately 103 generations The topologies wereused to generate a 50 majority rule consensus tree Pos-terior probabilities (PP) are given on appropriate clades
MORPHOMETRIC AND MORPHOLOGICAL ANALYSIS
Adult root-lesion nematodes from bromeliads wereused for this study Live specimens were immobilised bygently heating and then mounted in water agar on a slide(Esser 1986) for measurements and photographs Addi-tional measurements and drawings were made from spec-
imens killed and fixed in hot aqueous 2 formaldehyde +1 propionic acid dehydrated in ethanol vapour andmounted in dehydrated glycerin (Hooper 1970) Mea-surements of specimens were made with an ocular mi-crometer and drawings with a camera lucida Abbrevi-ations used are defined in Siddiqi (2000) Photographswere taken with two Leica (Wild MPS 4652 and LeicaDFC 320) cameras mounted on Nikon (Optiphot) and Le-ica DM 2500 compound microscopes
The morphological information on the root-lesion ne-matodes from Bahia grass (N1) and maidencane (N2) pro-vided by Inserra et al (1996) and Duncan et al (1999)was complemented by further microscopic observationsof additional preserved specimens kept in the nematodecollection (CNR-IPP Bari) by the second author of thispaper Morphometrics of mature females of root-lesionnematode species studied by Duncan et al (1999) andP hippeastri from bromeliad and the original descrip-tion were subjected to principal component analysis us-ing Minitab 13 (Minitab USA) The populations werecharacterised based on the lip morphology (smooth or di-vided) and the weakly-allometric charactersratios V aand stylet length These characters were reported to dis-criminate relationships among P coffeae-group speciesthat conformed closely to the phylogenetic relationshipsinferred by analyses of the D2-D3 region of the 28S rDNAsequence (Duncan et al 1999)
Specimens for scanning electron microscope (SEM)observations were cold-fixed in glutaraldehyde bufferedwith 01 M phosphate buffer (pH 72) post fixed 1 h in2 osmium tetroxide dehydrated in a graded series ofethanol critical point dried with CO2 and sputter coatedwith gold palladium (Eisenback 1985) Nematodes wereobserved with a Hitachi S530 microscope at 15-20 kVaccelerating voltage
SPECIES DELIMITATION IN STUDIED PRATYLENCHUS
Species delimitation of the studied populations wasdone by applying an integrated or polyphasic approachwhich was based on consideration of results of morpho-logical and morphometrical studies phylogenetic and se-quencing analysis and analysis of host plants and geo-graphic distribution of studied samples This approach in-tegrates any significant information on the organisms andresults in a consensus and transition type of classification(Subbotin amp Moens 2006) Two new species named hereas P floridensis n sp and P parafloridensis n sp andseveral unidentified putative new species defined here asPratylenchus spH1-H8 were delimited in this study using
850 Nematology
Pratylenchus hippeastri group from Florida
this approach More detailed morphological and molecu-lar analysis is still required to confirm the unique speciesstatus of Pratylenchus spH1-H8
Results
ROOT-LESION NEMATODE POPULATIONS COLLECTED
FROM BROMELIADS AND OTHER PLANTS
Bromeliads belonging to the genera Guzmania Ruizamp Pav Neoregelia L B Sm and Vriesea Lindl werefound to be infected by root-lesion nematodes similar toP hippeastri (Table 1) Population levels were usuallylt10 specimensg fresh roots In some cases bromeliadroots were found infected concomitantly with a few spec-imens of P brachyurus (Godfrey 1929) Filipjev amp Schu-urmans Stekhoven 1941 endoparasitic Helicotylenchusdihystera (Cobb 1893) Sher 1961 and Meloidogyne are-naria (Neal 1889) Chitwood 1949 Carrot cultures inoc-ulated with female root-lesion nematodes from bromeli-ads produced a large number of nematodes at differentlife stages and only a few males (usually three malesper 100 females) The population levels of other Floridaroot-lesion nematodes varied from 30 160 and 100 speci-mensg fresh roots for spH1 spH5 and spH6 respectivelyThe number of males in the amphimictic spH1 and spH6was ca 40100 females
MOLECULAR CHARACTERISATION OF P HIPPEASTRI
SPECIES COMPLEX
The amplification of the ITS containing region pro-duced a single fragment of ca 970-1000 bp for the studiedsamples The sequence alignment for P hippeastri and re-lated species with consensus sequence for each putativespecies is given in Figure 1 The ITS alignment included44 sequences and was 1050 bp in length Sequence di-versity within all studied root-lesion nematodes includingP jaehni reached 19 (174 nucleotides) for P hippeas-tri from bromeliad and from amaryllis it varied from 0 to06 (0-6 nucleotides) whereas sequence diversity withinthe other root-lesion nematodes related to P hippeastrireached 62 (57 nucleotides) Phylogenetic relationshipswithin Pratylenchus species as inferred from Bayesianinference are given in Figure 2 Four main moderate orhighly supported clades (PP = 90-100) were distinguishedwithin the tree Clade 1 grouped P hippeastri populationsalong with root-lesion nematodes spH1-H5 Populationsof P hippeastri from amaryllis and bromeliads clustered
together forming one highly supported (PP = 100) sub-clade within clade 1 The ITS sequences for PratylenchusspH2 and spH3 did not form distinct subclades and re-lationships between them were not resolved The root-lesion nematode N2 (= P parafloridensis n sp in Table 2and Figs 1-3) formed a moderately supported (PP = 90)clade 2 together with Pratylenchus spH6 Root-lesion ne-matodes spH7 and spH8 clustered together and were notwell separated The two sequences of N1 (= P floridensisn sp in Table 2 and Figs 1-3) formed highly supportedclade 4 at the basal position of the tree
Sequence alignments of the D2-D3 of 28S rDNA in-cluded 32 sequences of 713 bp length Sequence diver-sity reached 74 (47 nucleotides) for all root-lesion ne-matodes studied 21 (14 nucleotides) for the speciesclosely related to P hippeastri and varied from 0-03 (0-2 nucleotides) within P hippeastri populations Phyloge-netic relationships within Pratylenchus species based onD2-D3 of 28S rDNA sequences is given in Figure 3 Fivemain weakly to highly supported clades (PP = 72-100)were distinguished within the tree which corresponded tothe clades on the ITS-rRNA tree Populations of P hip-peastri from amaryllis and bromeliads formed a moder-ately supported (PP = 93) clade 1 together with Praty-lenchus spH1 (Florida USA) spH2 (South Africa) andspH5 (Florida USA) Root-lesion nematodes spH7 andspH8 clustered in a moderately supported clade 3 Theamphimictic N1 formed a highly supported clade 4 at thebasal position of the tree MP and BI analyses generatedcongruent trees with similar branch supports for the ITSand D2-D3 gene alignments respectively
The close phylogenetic relationships shared by thespecies (spH1-H8) characterised in this study indicate thatthey are representatives of a P hippeastri species com-plex These results confirmed the identity of males and fe-males of bromeliad populations as P hippeastri and pro-vided evidence that the rDNA sequences of P hippeas-tri males from bromeliads matched those of P hippeastrifemale type material Furthermore the phylogenetic find-ings also provided support for the description of N1 andN2 as two new species named P floridensis n sp andP parafloridensis n sp respectively
MORPHOLOGICAL CHARACTERISATION OF
PRATYLENCHUS HIPPEASTRI
Comparative measurements of females and males ofP hippeastri from bromeliads and those reported fromamaryllis in the original description are reported inTable 3
Vol 12(6) 2010 851
F De Luca et al
852 Nematology
Pratylenchus hippeastri group from Florida
Fig 2 The 50 majority rule consensus tree from Bayesian analysis generated from the ITS sequence dataset for the Pratylenchushippeastri species complex using the GTR + I + G model Posterior probability more than 70 is given for appropriate clades
Female
Morphometric values of the females from bromeliadsdid not differ from those of P hippeastri from amaryllisexcept for the tail length in fixed specimens which wasslightly shorter than that of females from amaryllis Smalldifferences were observed also in the mean values of some
characters of living specimens from bromeliad such asbody tail and post-uterine sac length which were shorterthan those of specimens from amaryllis Bromeliad fe-males also showed smaller maximum and vulval body dia-meter and a shorter vulva-anus distance However theirrange values overlapped Their lip pattern was also simi-lar to that of P hippeastri and consisted of a flat and un-
Fig 1 Sequence alignment of partial 18S complete ITS1 58S ITS2 and partial 28S rRNA for the Pratylenchus hippeastri speciescomplex The 18S 58S and 28S rRNA gene sequences are marked in bold the primers sequences are underlined Consensus sequenceis given for each species only Single letter code recommended by NC-IUB was used to specify nucleotide if two or more bases werepermitted at a particular position in a species subalignment Lower case symbols indicate presence of one or several gaps in a particularposition in sequences for a species subalignment
Vol 12(6) 2010 853
F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
18S-Int (5prime-CGTAACAAGGTAGCTGTAGG-3prime) and re-verse 26S-Int (5prime-TCCTCCGCTAAATGATAT-3prime) for-ward TW81 (5prime-GTTTCCGTAGGTGAACCTGC-3prime) andreverse AB28 (5prime-ATATGCTTAAGTTCAGCGGGT-3prime) orforward PRATTW81 (5prime-GTAGGTGAACCTGCTGCTG-3prime) and reverse AB28 PCR products were purified usingthe protocol listed by the manufacturers of NucleospinExtract II (Macherey-Nagel Germany) or QIAquick (Qi-agen USA) gel extraction kits and used for cloning or di-rect sequencing in both directions with the primers givenabove or M13 forward and M13 reverse primers TOPO-TA cloning kit (Invitrogen) or pGEM-T Vector System IIkit (Promega USA) were used for cloning of PCR prod-ucts Newly obtained sequences were deposited in Gen-Bank under accession numbers given in Table 2
The newly obtained sequences for both ribosomal re-gions of P hippeastri from bromeliad and of root-lesionnematodes from other hosts including those from amaryl-lis Bahia grass (N1) maidencane (N2) (Table 2) and ofP jaehni Inserra Duncan Troccoli Dunn dos SantosKaplan amp Vovlas 2001 (Duncan et al 1999 Waeyen-berge unpubl) were aligned using ClustalW (Thompsonet al 1997) with default parameters Pratylenchus jaehniwas used as an outgroup taxon (Subbotin et al 2008)Phylogenetic analysis of the sequence data sets were per-formed with maximum parsimony (MP) using PAUPlowast4b10 (Swofford 2002) and Bayesian inference (BI) usingMrBayes 312 (Huelsenbeck amp Ronquist 2001) For MPwe used heuristic search setting with ten replicates of ran-dom taxon addition tree bisection-reconnection branchswapping to seek the most parsimonious trees Gaps weretreated as missing data To obtain an estimation of the sup-port for each node a bootstrap analysis (BS) with 1000replicates was done BI analysis under the GTR + I + Gmodel for each gene was initiated with a random startingtree and was run with four chains for 10 times 106 genera-tions The Markov chains were sampled at intervals of 100generations Two runs were performed for each analysisThe log-likelihood values of the sample points stabilisedafter approximately 103 generations The topologies wereused to generate a 50 majority rule consensus tree Pos-terior probabilities (PP) are given on appropriate clades
MORPHOMETRIC AND MORPHOLOGICAL ANALYSIS
Adult root-lesion nematodes from bromeliads wereused for this study Live specimens were immobilised bygently heating and then mounted in water agar on a slide(Esser 1986) for measurements and photographs Addi-tional measurements and drawings were made from spec-
imens killed and fixed in hot aqueous 2 formaldehyde +1 propionic acid dehydrated in ethanol vapour andmounted in dehydrated glycerin (Hooper 1970) Mea-surements of specimens were made with an ocular mi-crometer and drawings with a camera lucida Abbrevi-ations used are defined in Siddiqi (2000) Photographswere taken with two Leica (Wild MPS 4652 and LeicaDFC 320) cameras mounted on Nikon (Optiphot) and Le-ica DM 2500 compound microscopes
The morphological information on the root-lesion ne-matodes from Bahia grass (N1) and maidencane (N2) pro-vided by Inserra et al (1996) and Duncan et al (1999)was complemented by further microscopic observationsof additional preserved specimens kept in the nematodecollection (CNR-IPP Bari) by the second author of thispaper Morphometrics of mature females of root-lesionnematode species studied by Duncan et al (1999) andP hippeastri from bromeliad and the original descrip-tion were subjected to principal component analysis us-ing Minitab 13 (Minitab USA) The populations werecharacterised based on the lip morphology (smooth or di-vided) and the weakly-allometric charactersratios V aand stylet length These characters were reported to dis-criminate relationships among P coffeae-group speciesthat conformed closely to the phylogenetic relationshipsinferred by analyses of the D2-D3 region of the 28S rDNAsequence (Duncan et al 1999)
Specimens for scanning electron microscope (SEM)observations were cold-fixed in glutaraldehyde bufferedwith 01 M phosphate buffer (pH 72) post fixed 1 h in2 osmium tetroxide dehydrated in a graded series ofethanol critical point dried with CO2 and sputter coatedwith gold palladium (Eisenback 1985) Nematodes wereobserved with a Hitachi S530 microscope at 15-20 kVaccelerating voltage
SPECIES DELIMITATION IN STUDIED PRATYLENCHUS
Species delimitation of the studied populations wasdone by applying an integrated or polyphasic approachwhich was based on consideration of results of morpho-logical and morphometrical studies phylogenetic and se-quencing analysis and analysis of host plants and geo-graphic distribution of studied samples This approach in-tegrates any significant information on the organisms andresults in a consensus and transition type of classification(Subbotin amp Moens 2006) Two new species named hereas P floridensis n sp and P parafloridensis n sp andseveral unidentified putative new species defined here asPratylenchus spH1-H8 were delimited in this study using
850 Nematology
Pratylenchus hippeastri group from Florida
this approach More detailed morphological and molecu-lar analysis is still required to confirm the unique speciesstatus of Pratylenchus spH1-H8
Results
ROOT-LESION NEMATODE POPULATIONS COLLECTED
FROM BROMELIADS AND OTHER PLANTS
Bromeliads belonging to the genera Guzmania Ruizamp Pav Neoregelia L B Sm and Vriesea Lindl werefound to be infected by root-lesion nematodes similar toP hippeastri (Table 1) Population levels were usuallylt10 specimensg fresh roots In some cases bromeliadroots were found infected concomitantly with a few spec-imens of P brachyurus (Godfrey 1929) Filipjev amp Schu-urmans Stekhoven 1941 endoparasitic Helicotylenchusdihystera (Cobb 1893) Sher 1961 and Meloidogyne are-naria (Neal 1889) Chitwood 1949 Carrot cultures inoc-ulated with female root-lesion nematodes from bromeli-ads produced a large number of nematodes at differentlife stages and only a few males (usually three malesper 100 females) The population levels of other Floridaroot-lesion nematodes varied from 30 160 and 100 speci-mensg fresh roots for spH1 spH5 and spH6 respectivelyThe number of males in the amphimictic spH1 and spH6was ca 40100 females
MOLECULAR CHARACTERISATION OF P HIPPEASTRI
SPECIES COMPLEX
The amplification of the ITS containing region pro-duced a single fragment of ca 970-1000 bp for the studiedsamples The sequence alignment for P hippeastri and re-lated species with consensus sequence for each putativespecies is given in Figure 1 The ITS alignment included44 sequences and was 1050 bp in length Sequence di-versity within all studied root-lesion nematodes includingP jaehni reached 19 (174 nucleotides) for P hippeas-tri from bromeliad and from amaryllis it varied from 0 to06 (0-6 nucleotides) whereas sequence diversity withinthe other root-lesion nematodes related to P hippeastrireached 62 (57 nucleotides) Phylogenetic relationshipswithin Pratylenchus species as inferred from Bayesianinference are given in Figure 2 Four main moderate orhighly supported clades (PP = 90-100) were distinguishedwithin the tree Clade 1 grouped P hippeastri populationsalong with root-lesion nematodes spH1-H5 Populationsof P hippeastri from amaryllis and bromeliads clustered
together forming one highly supported (PP = 100) sub-clade within clade 1 The ITS sequences for PratylenchusspH2 and spH3 did not form distinct subclades and re-lationships between them were not resolved The root-lesion nematode N2 (= P parafloridensis n sp in Table 2and Figs 1-3) formed a moderately supported (PP = 90)clade 2 together with Pratylenchus spH6 Root-lesion ne-matodes spH7 and spH8 clustered together and were notwell separated The two sequences of N1 (= P floridensisn sp in Table 2 and Figs 1-3) formed highly supportedclade 4 at the basal position of the tree
Sequence alignments of the D2-D3 of 28S rDNA in-cluded 32 sequences of 713 bp length Sequence diver-sity reached 74 (47 nucleotides) for all root-lesion ne-matodes studied 21 (14 nucleotides) for the speciesclosely related to P hippeastri and varied from 0-03 (0-2 nucleotides) within P hippeastri populations Phyloge-netic relationships within Pratylenchus species based onD2-D3 of 28S rDNA sequences is given in Figure 3 Fivemain weakly to highly supported clades (PP = 72-100)were distinguished within the tree which corresponded tothe clades on the ITS-rRNA tree Populations of P hip-peastri from amaryllis and bromeliads formed a moder-ately supported (PP = 93) clade 1 together with Praty-lenchus spH1 (Florida USA) spH2 (South Africa) andspH5 (Florida USA) Root-lesion nematodes spH7 andspH8 clustered in a moderately supported clade 3 Theamphimictic N1 formed a highly supported clade 4 at thebasal position of the tree MP and BI analyses generatedcongruent trees with similar branch supports for the ITSand D2-D3 gene alignments respectively
The close phylogenetic relationships shared by thespecies (spH1-H8) characterised in this study indicate thatthey are representatives of a P hippeastri species com-plex These results confirmed the identity of males and fe-males of bromeliad populations as P hippeastri and pro-vided evidence that the rDNA sequences of P hippeas-tri males from bromeliads matched those of P hippeastrifemale type material Furthermore the phylogenetic find-ings also provided support for the description of N1 andN2 as two new species named P floridensis n sp andP parafloridensis n sp respectively
MORPHOLOGICAL CHARACTERISATION OF
PRATYLENCHUS HIPPEASTRI
Comparative measurements of females and males ofP hippeastri from bromeliads and those reported fromamaryllis in the original description are reported inTable 3
Vol 12(6) 2010 851
F De Luca et al
852 Nematology
Pratylenchus hippeastri group from Florida
Fig 2 The 50 majority rule consensus tree from Bayesian analysis generated from the ITS sequence dataset for the Pratylenchushippeastri species complex using the GTR + I + G model Posterior probability more than 70 is given for appropriate clades
Female
Morphometric values of the females from bromeliadsdid not differ from those of P hippeastri from amaryllisexcept for the tail length in fixed specimens which wasslightly shorter than that of females from amaryllis Smalldifferences were observed also in the mean values of some
characters of living specimens from bromeliad such asbody tail and post-uterine sac length which were shorterthan those of specimens from amaryllis Bromeliad fe-males also showed smaller maximum and vulval body dia-meter and a shorter vulva-anus distance However theirrange values overlapped Their lip pattern was also simi-lar to that of P hippeastri and consisted of a flat and un-
Fig 1 Sequence alignment of partial 18S complete ITS1 58S ITS2 and partial 28S rRNA for the Pratylenchus hippeastri speciescomplex The 18S 58S and 28S rRNA gene sequences are marked in bold the primers sequences are underlined Consensus sequenceis given for each species only Single letter code recommended by NC-IUB was used to specify nucleotide if two or more bases werepermitted at a particular position in a species subalignment Lower case symbols indicate presence of one or several gaps in a particularposition in sequences for a species subalignment
Vol 12(6) 2010 853
F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
this approach More detailed morphological and molecu-lar analysis is still required to confirm the unique speciesstatus of Pratylenchus spH1-H8
Results
ROOT-LESION NEMATODE POPULATIONS COLLECTED
FROM BROMELIADS AND OTHER PLANTS
Bromeliads belonging to the genera Guzmania Ruizamp Pav Neoregelia L B Sm and Vriesea Lindl werefound to be infected by root-lesion nematodes similar toP hippeastri (Table 1) Population levels were usuallylt10 specimensg fresh roots In some cases bromeliadroots were found infected concomitantly with a few spec-imens of P brachyurus (Godfrey 1929) Filipjev amp Schu-urmans Stekhoven 1941 endoparasitic Helicotylenchusdihystera (Cobb 1893) Sher 1961 and Meloidogyne are-naria (Neal 1889) Chitwood 1949 Carrot cultures inoc-ulated with female root-lesion nematodes from bromeli-ads produced a large number of nematodes at differentlife stages and only a few males (usually three malesper 100 females) The population levels of other Floridaroot-lesion nematodes varied from 30 160 and 100 speci-mensg fresh roots for spH1 spH5 and spH6 respectivelyThe number of males in the amphimictic spH1 and spH6was ca 40100 females
MOLECULAR CHARACTERISATION OF P HIPPEASTRI
SPECIES COMPLEX
The amplification of the ITS containing region pro-duced a single fragment of ca 970-1000 bp for the studiedsamples The sequence alignment for P hippeastri and re-lated species with consensus sequence for each putativespecies is given in Figure 1 The ITS alignment included44 sequences and was 1050 bp in length Sequence di-versity within all studied root-lesion nematodes includingP jaehni reached 19 (174 nucleotides) for P hippeas-tri from bromeliad and from amaryllis it varied from 0 to06 (0-6 nucleotides) whereas sequence diversity withinthe other root-lesion nematodes related to P hippeastrireached 62 (57 nucleotides) Phylogenetic relationshipswithin Pratylenchus species as inferred from Bayesianinference are given in Figure 2 Four main moderate orhighly supported clades (PP = 90-100) were distinguishedwithin the tree Clade 1 grouped P hippeastri populationsalong with root-lesion nematodes spH1-H5 Populationsof P hippeastri from amaryllis and bromeliads clustered
together forming one highly supported (PP = 100) sub-clade within clade 1 The ITS sequences for PratylenchusspH2 and spH3 did not form distinct subclades and re-lationships between them were not resolved The root-lesion nematode N2 (= P parafloridensis n sp in Table 2and Figs 1-3) formed a moderately supported (PP = 90)clade 2 together with Pratylenchus spH6 Root-lesion ne-matodes spH7 and spH8 clustered together and were notwell separated The two sequences of N1 (= P floridensisn sp in Table 2 and Figs 1-3) formed highly supportedclade 4 at the basal position of the tree
Sequence alignments of the D2-D3 of 28S rDNA in-cluded 32 sequences of 713 bp length Sequence diver-sity reached 74 (47 nucleotides) for all root-lesion ne-matodes studied 21 (14 nucleotides) for the speciesclosely related to P hippeastri and varied from 0-03 (0-2 nucleotides) within P hippeastri populations Phyloge-netic relationships within Pratylenchus species based onD2-D3 of 28S rDNA sequences is given in Figure 3 Fivemain weakly to highly supported clades (PP = 72-100)were distinguished within the tree which corresponded tothe clades on the ITS-rRNA tree Populations of P hip-peastri from amaryllis and bromeliads formed a moder-ately supported (PP = 93) clade 1 together with Praty-lenchus spH1 (Florida USA) spH2 (South Africa) andspH5 (Florida USA) Root-lesion nematodes spH7 andspH8 clustered in a moderately supported clade 3 Theamphimictic N1 formed a highly supported clade 4 at thebasal position of the tree MP and BI analyses generatedcongruent trees with similar branch supports for the ITSand D2-D3 gene alignments respectively
The close phylogenetic relationships shared by thespecies (spH1-H8) characterised in this study indicate thatthey are representatives of a P hippeastri species com-plex These results confirmed the identity of males and fe-males of bromeliad populations as P hippeastri and pro-vided evidence that the rDNA sequences of P hippeas-tri males from bromeliads matched those of P hippeastrifemale type material Furthermore the phylogenetic find-ings also provided support for the description of N1 andN2 as two new species named P floridensis n sp andP parafloridensis n sp respectively
MORPHOLOGICAL CHARACTERISATION OF
PRATYLENCHUS HIPPEASTRI
Comparative measurements of females and males ofP hippeastri from bromeliads and those reported fromamaryllis in the original description are reported inTable 3
Vol 12(6) 2010 851
F De Luca et al
852 Nematology
Pratylenchus hippeastri group from Florida
Fig 2 The 50 majority rule consensus tree from Bayesian analysis generated from the ITS sequence dataset for the Pratylenchushippeastri species complex using the GTR + I + G model Posterior probability more than 70 is given for appropriate clades
Female
Morphometric values of the females from bromeliadsdid not differ from those of P hippeastri from amaryllisexcept for the tail length in fixed specimens which wasslightly shorter than that of females from amaryllis Smalldifferences were observed also in the mean values of some
characters of living specimens from bromeliad such asbody tail and post-uterine sac length which were shorterthan those of specimens from amaryllis Bromeliad fe-males also showed smaller maximum and vulval body dia-meter and a shorter vulva-anus distance However theirrange values overlapped Their lip pattern was also simi-lar to that of P hippeastri and consisted of a flat and un-
Fig 1 Sequence alignment of partial 18S complete ITS1 58S ITS2 and partial 28S rRNA for the Pratylenchus hippeastri speciescomplex The 18S 58S and 28S rRNA gene sequences are marked in bold the primers sequences are underlined Consensus sequenceis given for each species only Single letter code recommended by NC-IUB was used to specify nucleotide if two or more bases werepermitted at a particular position in a species subalignment Lower case symbols indicate presence of one or several gaps in a particularposition in sequences for a species subalignment
Vol 12(6) 2010 853
F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
852 Nematology
Pratylenchus hippeastri group from Florida
Fig 2 The 50 majority rule consensus tree from Bayesian analysis generated from the ITS sequence dataset for the Pratylenchushippeastri species complex using the GTR + I + G model Posterior probability more than 70 is given for appropriate clades
Female
Morphometric values of the females from bromeliadsdid not differ from those of P hippeastri from amaryllisexcept for the tail length in fixed specimens which wasslightly shorter than that of females from amaryllis Smalldifferences were observed also in the mean values of some
characters of living specimens from bromeliad such asbody tail and post-uterine sac length which were shorterthan those of specimens from amaryllis Bromeliad fe-males also showed smaller maximum and vulval body dia-meter and a shorter vulva-anus distance However theirrange values overlapped Their lip pattern was also simi-lar to that of P hippeastri and consisted of a flat and un-
Fig 1 Sequence alignment of partial 18S complete ITS1 58S ITS2 and partial 28S rRNA for the Pratylenchus hippeastri speciescomplex The 18S 58S and 28S rRNA gene sequences are marked in bold the primers sequences are underlined Consensus sequenceis given for each species only Single letter code recommended by NC-IUB was used to specify nucleotide if two or more bases werepermitted at a particular position in a species subalignment Lower case symbols indicate presence of one or several gaps in a particularposition in sequences for a species subalignment
Vol 12(6) 2010 853
F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
Fig 2 The 50 majority rule consensus tree from Bayesian analysis generated from the ITS sequence dataset for the Pratylenchushippeastri species complex using the GTR + I + G model Posterior probability more than 70 is given for appropriate clades
Female
Morphometric values of the females from bromeliadsdid not differ from those of P hippeastri from amaryllisexcept for the tail length in fixed specimens which wasslightly shorter than that of females from amaryllis Smalldifferences were observed also in the mean values of some
characters of living specimens from bromeliad such asbody tail and post-uterine sac length which were shorterthan those of specimens from amaryllis Bromeliad fe-males also showed smaller maximum and vulval body dia-meter and a shorter vulva-anus distance However theirrange values overlapped Their lip pattern was also simi-lar to that of P hippeastri and consisted of a flat and un-
Fig 1 Sequence alignment of partial 18S complete ITS1 58S ITS2 and partial 28S rRNA for the Pratylenchus hippeastri speciescomplex The 18S 58S and 28S rRNA gene sequences are marked in bold the primers sequences are underlined Consensus sequenceis given for each species only Single letter code recommended by NC-IUB was used to specify nucleotide if two or more bases werepermitted at a particular position in a species subalignment Lower case symbols indicate presence of one or several gaps in a particularposition in sequences for a species subalignment
Vol 12(6) 2010 853
F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
Fig 3 The 50 majority rule consensus tree from Bayesian analysis generated from the D2-D3 of 28S rRNA gene sequence datasetfor the Pratylenchus hippeastri species complex using the GTR + I + G model Posterior probability is given for appropriate clades
divided face with two lip annuli (in some specimens withan incomplete third annulus) and with the second annuluslarger and thicker than the first These females also hada small and empty spermatheca indicating that fertilisa-tion by the males in the cultures may not occur The smalland empty spermatheca was also observed in females col-lected directly from bromeliad roots from the nurseriesThese morphometric and morphological characters indi-
cate that the lesion nematodes from bromeliads are P hip-peastri
The principal components analysis for the morphomet-rics listed in Table 3 and those reported by Duncan et al(1999) positioned bromeliad P hippeastri in a group (VI)with N1 and N2 and separate from the other Pratylenchusspecies studied by Duncan et al (1999) (Fig 4) Consider-ing the four characters used the two principal components
854 Nematology
Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
Table 3 Morphometrics of Pratylenchus hippeastri from Florida All measurements are in μm and in the form mean plusmn standarddeviation (range)
Character Population from bromeliads (present study) Population from amaryllis(Inserra et al 2007)
Female MaleFemale
n 12 (live) 10 (fixed) 5 (fixed) 21 (fixed)L 527 plusmn 485 (447-616) 614 plusmn 224 (585-651) 421 plusmn 323 (370-452) 590 plusmn 218 (545-627)a 283 plusmn 16 (246-307) 252 plusmn 20 (237-265) 306 plusmn 33 (258-339) 255 plusmn 12 (232-279)b 58 plusmn 04 (52-66) 66 plusmn 04 (59-72) 55 plusmn 02 (53-58) 65 plusmn 04 (57-71)bprime 37 plusmn 03 (33-42) 45 plusmn 04 (40-53) 35 plusmn 03 (32-38) 44 plusmn 03 (39-51)c 179 plusmn 12 (157-199) 186 plusmn 20 (164-233) 184 plusmn 14 (161-193) 161 plusmn 10 (146-187)cprime 24 plusmn 02 (20-26) 22 plusmn 02 (18-25) 24 plusmn 01 (23-25) 26 plusmn 02 (22-29)V or T 776 plusmn 13 (756-796) 777 plusmn 12 (757-794) 46 plusmn 28 (425-490) 77 plusmn 08 (75-78)Stylet length 154 plusmn 03 (151-158) 158 plusmn 04 (153-167) 143 plusmn 03 (140-147) 155 plusmn 04 (15-16)DGO from stylet base 34 plusmn 03 (30-38) 25 plusmn 03 (20-27) 22 plusmn 04 (17-27) 29 plusmn 02 (25-30)o 22 plusmn 16 (192-252) 16 plusmn 17 (128-174) 154 plusmn 31 (116-190) 19 plusmn 12 (16-20)Anterior end to
centre of metacorpus 58 plusmn 26 (55-64) 62 plusmn 19 (58-65) 506 plusmn 07 (50-51) 63 plusmn 19 (59-66)cardia 90 plusmn 30 (85-95) 93 plusmn 53 (87-106) 79 plusmn 20 (767-813) 92 plusmn 33 (83-98)end of pharyngeal 139 plusmn 97 (128-153) 137 plusmn 88 (123-147) 126 plusmn 70 (117-132) 134 plusmn 66 (116-145)
gland lobesecretoryexcretory pore 88 plusmn 55 (774-95) 94 plusmn 29 (89-99) 73 plusmn 42 (66-76) 91 plusmn 25 (85-95)
Pharyngeal overlap 50 plusmn 80 (385-615) 45 plusmn 76 (33-58) 48 plusmn 89 (35-55) 43 plusmn 54 (32-51)Max body diam 19 plusmn 15 (156-215) 24 plusmn 07 (233-257) 138 plusmn 08 (127-145) 23 plusmn 14 (21-27)Vulval body diam 17 plusmn 15 (142-205) 22 plusmn 16 (193-24) ndash 21 plusmn 11 (180-230)Anal body diam 12 plusmn 12 (107-147) 15 plusmn 04 (147-16) 95 plusmn 03 (93-10) 144 plusmn 08 (13-16)Vulva to anus distance 86 plusmn 109 (705-103) 103 plusmn 54 (92-109) ndash 98 plusmn 61 (88-112)Anterior genital 133 plusmn 603 (108-170) 268 plusmn 603 (200-387) 194 plusmn 204 (165-220) 254 plusmn 472 (181-360)
tract lengthPUS 23 plusmn 32 (186-294) 35 plusmn 30 (30-393) ndash 30 plusmn 49 (21-45)Tail length 30 plusmn 25 (272-357) 33 plusmn 30 (28-373) 23 plusmn 06 (220-237) 37 plusmn 22 (320-420)Spicule length ndash ndash 19 plusmn 06 (180-193) ndashGubernaculum length ndash ndash 53 plusmn 06 (47-60) ndashNo of tail annuli 24 plusmn 19 (21-26) 20 plusmn 26 (17-25) ndash 22 plusmn 21 (19-26)
explained 69 of the morphological variation betweenthe populations (n = 797 specimens from 45 populations)The correlations between the characters and the scores forprincipal components 1 and 2 respectively were V =075 and minus035 a = minus038 and minus075 stylet = 068 andminus050 lip morphology = 059 and minus054 Multiple re-gression of each character against the two principal com-ponent scores explained 68 71 72 and 64 of the varia-tion in V a stylet and lip morphology respectively
Male
The few males present in carrot disk cultures andbromeliad roots exhibited a slight sexual dimorphismMales had smaller lip region diameter (60-67 vs 7-
78 μm) stylet knobs (23-30 across and 17-20 high vs30-47 and 20-27 μm) and metacorpus (73-87 diam times100-113 high vs 100-113 times 133-173 μm) comparedto those of the females This sexual dimorphism has beenreported for many amphimictic root-lesion nematodesThe male lip pattern in spite of a slightly more collapsedappearance of the cuticle did not differ from that ofthe female and showed a flat and undivided face withtwo lip annuli of different size with the second annuluslarger and thicker than the first (Figs 5B 6C 7C D)The oral disc was slightly raised and the amphidialapertures were broader than in the female The styletwas more slender than that of the female and the knobsellipsoidal to triangular in profile with rounded margins
Vol 12(6) 2010 855
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology
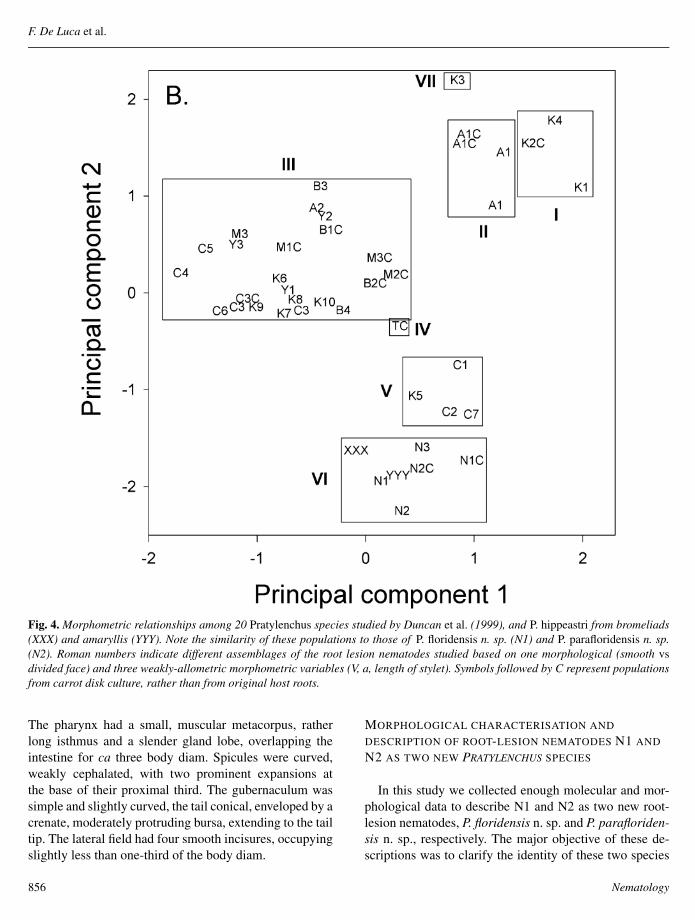
F De Luca et al
Fig 4 Morphometric relationships among 20 Pratylenchus species studied by Duncan et al (1999) and P hippeastri from bromeliads(XXX) and amaryllis (YYY) Note the similarity of these populations to those of P floridensis n sp (N1) and P parafloridensis n sp(N2) Roman numbers indicate different assemblages of the root lesion nematodes studied based on one morphological (smooth vsdivided face) and three weakly-allometric morphometric variables (V a length of stylet) Symbols followed by C represent populationsfrom carrot disk culture rather than from original host roots
The pharynx had a small muscular metacorpus ratherlong isthmus and a slender gland lobe overlapping theintestine for ca three body diam Spicules were curvedweakly cephalated with two prominent expansions atthe base of their proximal third The gubernaculum wassimple and slightly curved the tail conical enveloped by acrenate moderately protruding bursa extending to the tailtip The lateral field had four smooth incisures occupyingslightly less than one-third of the body diam
MORPHOLOGICAL CHARACTERISATION AND
DESCRIPTION OF ROOT-LESION NEMATODES N1 AND
N2 AS TWO NEW PRATYLENCHUS SPECIES
In this study we collected enough molecular and mor-phological data to describe N1 and N2 as two new root-lesion nematodes P floridensis n sp and P parafloriden-sis n sp respectively The major objective of these de-scriptions was to clarify the identity of these two species
856 Nematology
Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
Fig 5 Camera lucida line drawings of male of Pratylenchushippeastri A Entire body B En face view showing oral discfused with median and lateral lip sectors C Pharyngeal regionD Anterior end E F Tail region
that have been reported in the literature and GenBank withacronyms The morphological description of the other pu-tative species in the P hippeastri species complex fromFlorida and other countries was not attempted becausethey are cryptic species not separable by morphologicalanalysis
Pratylenchus floridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N1 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
Measurements of this species originally identified asP loosi from Bahia grass and later as N1 root-lesion ne-matode were reported in Inserra et al (1996) and Dun-can et al (1999) respectively Additional measurements(present study) of preserved specimens kept in the CNR-IPP Barirsquos nematode collection are reported in Table 4
DESCRIPTION
Female
Body of dead females almost straight Labial regionwith two annuli 2 μm high times 7 μm broad on averageoffset from body by a slight constriction second lipannulus wider and higher than first SEM en face viewcharacterised by undivided pattern all labial sectors fusedtogether and partially with an oval oral disc amphidialopenings rather wide obliquely orientated at sides oforal disc Stylet with ellipsoidal knobs or rounded withslightly flattened anterior surface 4 μm across 2 μmhigh (mean values) Dorsal pharyngeal gland opening 2-25 μm posterior to stylet base (o range = 129-167)Pharyngeal metacorpus oval 11-13 μm high times 85-11 μmin diam Isthmus slender encircled by nerve ring in upperpart located just posterior to metacorpus Pharyngealgland lobe overlapping intestine ventrally 35 plusmn 78 (27-52) μm in length Lateral fields with four lines notareolated Anterior genital tract 144-240 μm or 32-47of body length long Spermatheca small rounded or ovalrarely rectangular in shape occasionally oblong and largefilled with sperm Post-uterine sac ca 15 body diam longundifferentiated Tail bluntly pointed (sensu Frederick ampTarjan 1989) or subacute with smooth terminus In fewspecimens slight indentation was observed at tail tip
Specific epithet derived from Florida the only geographicalarea where this species has been detected
Vol 12(6) 2010 857
F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
Fig 6 Light micrographs of male of Pratylenchus hippeastri A Entire body B Pharyngeal region (live specimen) C Anterior endD Lateral field at mid-body E-G Tail region at different foci (Scale bars A = 50 μm B-G = 20 μm)
858 Nematology
Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
Fig 7 SEM morphology of Pratylenchus hippeastri from bromeliads in Florida A B Female C D Male A B Undivided face patternwith all labial sectors fused together and with oral disc Note second lip annulus thicker than first C D Undivided face pattern similarto that of female but with broader amphidial apertures
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand slightly smaller body size Lip region slightly off-set 2 μm high and 57 μm broad with two annuli(second annulus higher than first) Stylet more slen-der and shorter than in female with minute slightlycupped knobs 27 μm across 15 μm high Lip pat-tern in SEM en face view showing a plane undividedface two lip annuli (second annulus larger and thicker
than first) and an oral disc more rounded than in fe-males Pharynx with oval metacorpus (105 times 73 μmin longitudinal and cross diam respectively) and glandlobe overlapping intestine for 37 μm Hemizonid justanterior to secretory-excretory pore hemizonion eightannuli posterior to it Lateral field with four smoothlines Testis outstretched 238 μm long Tail conoidwith narrowed hyaline tip 4 μm long Spicules arcuateweakly cephalated gubernaculum simple slightly arcu-ate
Vol 12(6) 2010 859
F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
Fig 8 Camera lucida line drawings of Pratylenchus floridensis n sp (A-L) and P parafloridensis n sp (M-X) A N Female pharyngealregion B O Male pharyngeal region C P Female posterior region D Q Female anterior end E M Female entire body F RFemale vulval region with spermatheca G S Male tail H-L T-X Female tail
860 Nematology
Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
Fig 9 Light micrographs of Pratylenchus floridensis n sp (A-D) and P parafloridensis n sp (E-H) A E Female pharyngeal regionB F Female anterior end C G Female vulval region (brackets indicate position and the extent of spermatheca) D H Femaleposterior body portion (spermatheca in brackets) (Scale bars A-C E-G = 20 μm D H = 50 μm)
TYPE HOST AND LOCALITY
Bahia grass (Paspalum notatum Flueggeacute) roots col-lected from a sod farm in Zolfo Springs Hardee Coun-ty FL USA (latitude 2724prime67primeprimeN longitude 8138prime41primeprimeW) The soil type is sandy and the climate is subtrop-ical
TYPE MATERIAL
Holotype female 25 female and one male paratypedeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0939 to J-0944) Additional paratypes were distributed to the USDA
Vol 12(6) 2010 861
F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
Fig 10 Comparative light micrographs of tail region of Pratylenchus floridensis n sp and P parafloridensis n sp (broken line showsanus level)
Nematode Collection Beltsville MD USA (collectionnumber IPP-J-0945) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0938)
DIAGNOSIS AND RELATIONSHIPS
The P floridensis n sp female is characterised bythe following morphological characters slender bodyundivided plain and smooth face with all labial sectorsfused together and partially with an oval oral disclip region with two lip annuli and with the secondannulus larger and thicker than the first ellipsoidal styletknobs or rounded with slightly flattened anterior surfacerounded oval or rarely rectangular spermatheca filledwith sperm tail bluntly pointed with smooth terminus (inrare specimens slightly indented) The matrix code (sensuCastillo amp Vovlas 2007) for this species is A1 B2 C2D2 E2 F4 G3 H1 I2 J1 K1
Few morphological and morphometrical characters sep-arate P floridensis n sp from P parafloridensis n sphereinafter described The present study revealed thatP floridensis n sp differs from P parafloridensis nsp in having a shorter female body (average 450 vs532 μm) an oval vs round oral disc a small round tooval and sometimes rectangular spermatheca vs quadran-gular or large rectangular in P parafloridensis n sp anda bluntly to finely pointed (rarely indented) tail tip vs sub-
hemispherical or bluntly pointed tail with smooth or lessfrequently indented tail terminus However these charac-ters may overlap in some specimens making the morpho-logical separation of these two species unreliable withoutthe corroboration of the molecular analysis and thus theyare considered as cryptic species
The amphimictic reproductive habits presence of malesand a large spermatheca filled with sperm morphologi-cally separate P floridensis n sp from other male-lessPratylenchus species with undivided and smooth facetwo lip annuli and having a non-functional spermathecasuch as P acuticaudatus Braasch amp Decker 1989 P an-gulatus Siddiqi 1994 P brachyurus P estoniensis Ryss1982 P hippeastri and P tenuis Thorne amp Malek 1968This new species differs from the amphimictic Praty-lenchus with the same lip region features by the followingcharacters from P alleni Ferris 1961 it differs in the tailshape (bluntly pointed vs rounded) and a more anteriorvulva position (77 vs 80) from P araucensis MuacuteneraBert amp Decraemer 2009 by the long vs short pharyngealoverlap lateral field smooth vs areolated in outer lateralridges and shape of tail terminus (smooth vs variable)from P artemisiae Zheng amp Chen 1994 by the longerstylet length (14-155 vs 115-145 μm) and more ante-rior vulva position (75-80 vs 76-81) from P brzeskiiKarssen Waeyenberge amp Moens 2000 by the shorterstylet (14-155 vs 18-19 μm) from P coffeae by the tailshape (bluntly pointed vs rounded truncate or indented)
862 Nematology
Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
Fig 11 Comparison of SEM lip region morphology of Pratylenchus floridensis n sp (A-C) and P parafloridensis n sp (D-F) A BFemale face view showing undivided patterns with all labial sectors fused together and partially with oval oral disc C Male facepattern D-F Female undivided face patterns with all labial sectors fused together and partially with round oral disc Note second lipannulus larger and thicker than first in both species
and a more anterior vulva position (77 vs 81) fromP flakkensis Seinhorst 1968 by the tail (bluntly pointedwith smooth terminus vs conical with faintly annulatedterminus) and stylet knob shape (ellipsoid to rounded vsanteriorly pointed) from P gutierrezi Golden Lopez ampVilchez 1992 by the undivided face vs divided and fromP kumamotoensis Mizukubo Sugimura amp Uesugi 2007by the pharyngeal gland lobe (ventral vs frequently dor-sal) shorter PUS (23-32 vs 37-45 μm) and lateral field(smooth vs areolated in vulval region) Pratylenchus flori-densis n sp also differs from P jaehni P loosi P neo-brachyurus Siddiqi 1994 P panamenis Siddiqi Lopezamp Vilchez 1991 P roseus Zarina amp Maqbool 1998 and
Considered as junior synonym of P panamaensis by Siddiqi(2000) Castillo and Vovlas (2007) and Handoo et al (2008)
P silvaticus Brzeski 1998 by the more anterior vulva po-sition (75-80 vs 77-80 79-85 80-84 77-83 81-83 and80-83 respectively) In addition P floridensis n sp hasa longer tail than P jaehni (25-29 vs 21-31 μm) a longerbody than P neobrachyurus (387-507 vs 310-410 μm)a different tail shape to P panamensis (bluntly pointedwith mostly smooth terminus vs subclavate with annu-lated terminus) different vulval margins number of lat-eral lines and tail terminus than P roseus (no vulval flapsfour lateral lines and smooth tail terminus vs presence ofvulval flaps six lateral lines and coarsely annulated tailterminus) and different tail shape to P silvaticus (slightlyclavate with irregularly striated tail terminus)
We would like to point out that lip patterns of P acuti-caudatus P alleni P artemisiae P angulatus P brzeskii
Vol 12(6) 2010 863
F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
Table 4 Morphometrics of Pratylenchus floridensis n sp (Zolfo Springs population) and P parafloridensis n sp (Lithia population)All measurements are in μm and in the form mean plusmn sd (range)
Character P floridensis n sp P parafloridensis n sp
Female Male Female Male
Holotype Paratypes Paratype Holotype Paratypes Paratypes
n ndash 9 1 ndash 10 5L 457 450 plusmn 315 (387-507)lowast 457 563 532 plusmn 416 (475-603)lowastlowast 448 plusmn 379 (414-494)a 295 264 plusmn 18 (207-301) 254 256 290 plusmn 33 (252-37) 297 plusmn 38 (25-353)b 59 56 plusmn 03 (53-59) 61 66 59 plusmn 03 (53-66) 56 plusmn 04 (53-60)bprime 39 39 plusmn 03 (34-43) 41 44 40 plusmn 04 (35-46) 37 plusmn 04 (34-40)c 157 168 plusmn 08 (153-178) 146 179 168 plusmn 14 (149-185) 179 plusmn 20 (15-191)cprime 29 26 plusmn 03 (22-31) 29 24 29 plusmn 03 (24-33) 28 plusmn 04 (24-33)V or T 77 77 plusmn 16 (75-80)lowast 52 78 77 plusmn 16 (75-80)lowastlowast 46 plusmn 79 (35-52)Stylet length 15 15 plusmn 06 (140-158) 22 158 154 plusmn 06 (145-160) 14 plusmn 03 (135-140)Anterior end to centre 56 54 plusmn 41 (48-59) 53 57 59 plusmn 36 (52-64) 54 plusmn 43 (51-61)
of metacorpusend of pharyngeal 117 118 plusmn 105 (104-140) 113 127 134 plusmn 68 (127-145) 128 plusmn 15 (118-145)
gland lobesecretoryexcretory 79 78 plusmn 66 (66-86) 75 83 88 plusmn 53 (81-96) 76 plusmn 36 (72-81)
poreSpermatheca length 16 17 plusmn 39 (13-245) ndash 21 24 plusmn 80 (15-38) ndashSpermatheca diam 10 12 plusmn 14 (95-13) ndash 17 13 plusmn 22 (95-16) ndashPUS 28 27 plusmn 35 (23-32) ndash 31 28 plusmn 52 (21-37) ndashTail length 29 28 plusmn 14 (25-29) 313 315 32 plusmn 27 (28-35) 25 plusmn 54 (22-33)Anal body diam 10 11 plusmn 12 (90-120) 107 13 11 plusmn 07 (105-13) 90 plusmn 17 (67-107)Spicules ndash ndash 19 ndash ndash 185 plusmn 06 (178-19)Gubernaculum ndash ndash 6 ndash ndash 53 plusmn 0 (53)
Measurements taken on 18 specimens Measurements taken on 19 specimens
P estoniensis P flakkensis P gibbicaudatus P kumamo-toensis P neobrachyurus P panamensis P roseus P sil-vaticus and P tenuis are not known (Castillo amp Vovlas2007)
Pratylenchus parafloridensis n sp= P loosi apud Inserra et al (1996) nec Loof (1960)
= Pratylenchus N2 of Duncan et al (1999)(Figs 8-11)
MEASUREMENTS
See Table 4 and Inserra et al (1996) and Table 2 andResults in Duncan et al (1999)
Specific epithet consisting of para = close + floridensis andindicating the close similarity of this species with P floridensisn sp
DESCRIPTION
Female
Body of dead females almost straight or in open CLabial region with two annuli 23 μm high 75 μm broadon average offset from body by a slight constrictionsecond lip annulus distinctly wider and higher than firstSEM en face view characterised by undivided patternwith all labial sectors fused together and with a roundedoral disc amphidial openings obliquely orientated atsides of oral disc Stylet with rounded or ellipsoidalknobs 4 μm across 21 μm high (mean values) Dorsalpharyngeal gland opening 2-25 μm posterior to styletbase (o range = 125-172) Pharyngeal metacorpusoval 10-15 μm high times 75-13 μm diam Isthmus slenderencircled by nerve ring in anterior half Pharyngeal glandlobe rather long overlapping intestine ventrally 43 plusmn93 (30-56) μm in length Lateral fields with four lines
864 Nematology
Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
not areolated Anterior genital tract 171-211 μm or 31-39 of body length long Spermatheca filled with spermquadrangular or large rectangular in shape sometimeswith constriction in equatorial diam giving appearanceof a bilobed structure Post-uterine sac ca 15 body diamlong often with rudimentary cellular elements at tip Tailconoid subhemispherical or bluntly pointed with smoothor slightly indented (ca 30 of specimens observed)terminus
Male
Common in field populations Body straight when heat-relaxed similar to female except for sexual dimorphismand body size which is slightly smaller Lip region 20 plusmn01 (2-21) μm high and 64 plusmn 03 (6-67) μm broadStylet more slender and shorter than in female withrounded knobs 2-27 μm across 2 μm high Pharyngealmetacorpus rounded to oval 105 times 77 μm (longitudi-nal and cross diam respectively) Pharyngeal gland lobeoverlapping intestine for 43 plusmn 74 (37-51) μm Hemi-zonid just anterior to excretory pore Lateral field withfour smooth lines Tail conical rather short Testis out-stretched 211 plusmn 346 (174-254) μm long Spicules ar-cuate slender weakly cephalated gubernaculum simpleslightly arcuate
TYPE HOST AND LOCALITY
Maidencane (Panicum hemitomon Schultes) roots col-lected from a pasture land in Lithia Hillsborough FLUSA (latitude 2779prime63primeprimeN longitude 8221prime13primeprimeW) Thesoil type is sandy and the climate is subtropical
TYPE MATERIAL
Holotype female 29 female and four male paratypesdeposited at the Istituto per la Protezione delle Piante(IPP) of Consiglio Nazionale delle Ricerche (CNR) Sededi Bari Bari Italy (collection numbers IPP-J-0946 to J-0952) Additional paratypes were distributed to the USDANematode Collection Beltsville MD USA (collectionnumber IPP-J-0953) and University of California River-side Nematode Collection Riverside CA USA (collec-tion number IPP-J-0954)
DIAGNOSIS AND RELATIONSHIPS
The P parafloridensis n sp female is characterisedby the following morphological characters slender bodyundivided plain and smooth face with all labial sectors
fused together and with a round oral disc lip regionwith two lip annuli and with the second annulus largerand thicker than the first generally rounded stylet knobsquadrangular or large rectangular sometimes bilobedspermatheca filled with sperm tail subhemispherical orbluntly pointed with smooth or less frequently slightlyindented terminus The matrix code (sensu Castillo ampVovlas 2007) for this species is A1 B2 C2 D2 E2 F5G3 H1 I3 J1 K1
The relationship of P parafloridensis with other mem-bers of the genus Pratylenchus is similar to that describedabove for P floridensis n sp
Discussion
PRATYLENCHUS HIPPEASTRI FROM BROMELIADS
This study provides evidence that P hippeastri is a trop-ical root-lesion nematode reported so far only in Floridawhere it parasitises tropical ornamentals such as amaryl-lis and bromeliads The application of sequence and phy-logenetic analysis of the ITS-rRNA gene confirmed co-specificity of the root-lesion nematode population foundparasitising bromeliads with P hippeastri a previouslyknown parasite of amaryllis only Our observations in-dicate that populations of this species from bromeliadspresented males in both carrot discs and bromeliad rootsSo far no males have been found in other populations ofthis nematode The function of the males in the bromeliadpopulations is unclear since they are present in very smallnumber and are consistently in association with unmatedfemales showing an empty and small spermatheca Thereare reports of males occurring in parthenogenetic root-lesion nematodes such as P zeae (Loof 1991) The iden-tity of these males may be questioned since contaminatingmale specimens belonging to different species may be as-sociated with parthenogenetic species However in thiscase their identity was confirmed by sequencing rDNAgenes from male specimens In spite of the occurrence ofa few males our observations do not provide any evidencethat P hippeastri is an amphimictic species
USEFULNESS OF THE ITS-RDNA SEQUENCES FOR
SPECIES DIFFERENTIATION IN PRATYLENCHUS
The ITS-containing region allows better discriminationamong the closely related species studied because it hasevolved faster than the D2-D3 expansion segments of 28SrDNA and has accumulated more substitution changes
Vol 12(6) 2010 865
F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
The present analysis of the ITS-rDNA dataset clearly sep-arated P hippeastri from other amphimictic and male-lessroot-lesion nematodes confirming that they are probablynew Pratylenchus species belonging to the P hippeastrispecies complex The fact that they share morphologicalaffinity show minimal sequence differences in the rRNAgene and that sometimes their positions are not well re-solved in the phylogenetic trees suggests that these speciesare derived by recent speciation events with insufficienttime to attain complete morphological differentiation Thephylogenetic analysis of the ITS-rDNA does not confirmthe conclusion of cospecificity of Pratylenchus N1 and N2populations with P hippeastri previously made by Sub-botin et al (2008) based on analysis of the D2-D3 of 28SrDNA but instead shows that each of these populationsrepresents a distinct species Successful application of theITS for species differentiation in Pratylenchus has beenshown by Orui (1996) Waeyenberge et al (2000) andDe la Pentildea et al (2006) with PCR-RFLP These studiesalso revealed heterogeneity in the ITS sequences whichresulted in additional bands on gels after restriction ofPCR products These additional bands constitute complexRFLP profiles that may complicate diagnostics of Praty-lenchus species Our study also revealed heterogeneity inthe ITS sequences for all studied Pratylenchus speciesHowever in most cases the phylogenetic analysis of theITS sequence dataset allowed clear separation of sam-ple populations because except for Pratylenchus spH2and spH3 all sequences obtained from the same sam-ple clustered together Although P parafloridensis n spand Pratylenchus spH6 formed separate subclades on theITS trees relationships between these species based onthe D2-D3 remain uncertain Thus heterogeneity of ITSrRNA did not preclude species discrimination Combinedwith the PCR-RFLP method sequence and phylogeneticanalysis has become a reliable approach for differentia-tion of Pratylenchus species More detailed analysis ofthe ITS sequence alignment (Fig 1) will allow the designof species-specific primers and the discovery of appropri-ate restriction enzymes for diagnostics of P hippeastriP floridensis n sp P parafloridensis n sp and closelyrelated species
THE PRATYLENCHUS HIPPEASTRI SPECIES COMPLEX
Sequence and phylogenetic analysis revealed that acomplex of cryptic species genetically similar to P hip-peastri occurs in Florida USA South Africa and RussiaIn addition to P floridensis n sp and P parafloridensisn sp we conclude that eight other populations should
be considered as putative undescribed species Howeveradditional molecular morphological and biological stud-ies are required to clarify the taxonomic status of theseeight populations It is noteworthy that based on pre-liminary morphological studies these populations wereidentified not only as P hippeastri but as several otherknown species The diagnostic morphological charactersfor P hippeastri and the newly described P floridensisn sp and P parafloridensis n sp overlap to a signifi-cant degree requiring careful examination of many spec-imens for an accurate diagnosis Thus identification ofthe species of P hippeastri-complex is likely to rely in-creasingly on molecular methods Two (N1 N2) of theeight putative species from Florida were described hereinas new species because of their regulatory significanceThe description of the other six Florida putative P hip-peastri-species complex (spH1 spH4-H8) is currently notof crucial interest for agronomic or regulatory purposeshowever information provided in this paper documentstheir existence The description of the putative P hippeas-tri-species complex from South Africa and Russia (spH2spH3) requires more detailed morphological information
The fact that species in the P hippeastri-complex werefound in Florida Russia and South Africa suggests theirworld-wide distribution and a broad host range amongmonocots Moreover our findings suggest the recent evo-lution in Florida of numerous lesion nematodes includ-ing P hippeastri These species are male-less (spH5spH7 and spH8) or amphimictic (P floridensis n sp Pparafloridensis n sp spH1 and spH6) All have an undi-vided face with two and occasionally three lip annuli withthe exception of the male-less spH4 which has a dividedface with two lip annuli and was identified as P scrib-neri by Hernaacutendez et al (2000) This putative P scribneriin the P hippeastri-complex further complicates the taxo-nomic status of P scribneri Many lesion nematodes fromturf grasses in Florida including spH8 have been identi-fied as P zeae The inclusion in the P hippeastri-complexof another putative P zeae population from South Africacasts doubt about the real identity of P zeae and providesevidence that the reports of P zeae in Florida need to berevaluated
Acknowledgements
The technical field assistance provided by Lance ABrown Carrie S Edenfield Karen W Le Boutillier SolF Looker Ana L Ochoa and Howard L Wallace FloridaDepartment of Agriculture and Consumer Services is
866 Nematology
Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

Pratylenchus hippeastri group from Florida
greatly appreciated The authors thank Drs E Van denBerg A Yu Ryss and J Pinochet for supplying nema-tode materials FdL acknowledges support from the Ital-ian Minister of Research (MIUR) ndash Fondo FAR ndash Legge2971999 Art 12lab ndash Project Grant DM19410 SS ac-knowledges support from the NSF PEET DEB 0731516
References
BRAASCH H amp DECKER H (1989) Pratylenchus acu-ticaudatus sp n (Nematoda Pratylenchidae) aus einemgewaumlchshaus in der DDR Nematologica 34 57-61
BRZESKI MW (1998) Nematodes of Tylenchina in Polandand temperate Europe Warsaw Poland Museum and Insti-tute of Zoology Polish Academy of Sciences 398 pp
CASTILLO P amp VOVLAS N (2007) Pratylenchus (NematodaPratylenchidae) diagnosis biology pathogenicity and man-agement In Hunt DJ amp Perry RN (Eds) NematologyMonographs and Perspectives volume 6 Leiden The Nether-lands Brill Academic Publishers 529 pp
DE LUCA F FANELLI E DI VITO M REYES A ampDE GIORGI C (2004) Comparison of the sequences of theD3 expansion of the 26S ribosomal genes reveals differentdegrees of heterogeneity in different populations and speciesof Pratylenchus from the Mediterranean region EuropeanJournal of Plant Pathology 111 949-957
DUNCAN LW INSERRA RN THOMAS WK DUNN DMUSTIKA I FRISSE LM MENDES ML MORRISK amp KAPLAN DT (1999) Molecular and morphologicalanalyses of isolates of Pratylenchus coffeae and closelyrelated species Nematropica 29 61-80
EISENBACK JD (1985) Technique for preparing nematodesfor scanning electron microscopy In Barker KR CarterCC amp Sasser JN (Eds) An advanced treatise on Meloido-gyne volume 2 Raleigh NC USA Dept of Plant PathologyUniversity of North Carolina Press pp 75-105
ESSER RP (1886) A water agar en face technique Proceed-ings of the Helminthological Society of Washington 53 254-255
FERRIS VR (1961) A new species of Pratylenchus (Nemata-Tylenchida) from roots of soybeans Proceedings of theHelminthological Society of Washington 28 109-111
FREDERICK JJ amp TARJAN AC (1989) A compendium ofthe genus Pratylenchus Filipjev 1936 (Nemata Pratylenchi-dae) Revue de Neacutematologie 12 243-256
GOLDEN AM LOPEZ CR amp VILCHEZ RH (1992) De-scription of Pratylenchus gutierrezi n sp (Nematoda Praty-lenchidae) from coffee in Costa Rica Journal of Nematology24 298-304
HANDOO ZA CARTA LK amp SKANTAR AM (2008)Taxonomy morphology and phylogenetics of coffee-associated root-lesion nematodes Pratylenchus spp In
Souza RM (Ed) Plant-parasitic nematodes of coffee Dor-drecht The Netherlands Springer pp 29-50
HERNAacuteNDEZ MA JORDANA R GOLDARACENA A ampPINOCHET J (2000) SEM observations on nine speciesof the genus Pratylenchus Filipjev 1936 (Nematoda Praty-lenchidae) Journal of Nematode Morphology and Systemat-ics 3 165-174
HOOPER DJ (1970) Handling fixing staining and mountingnematodes In Southey J (Ed) Laboratory methods forwork with plant and soil nematodes Technical Bulletin no 25th edition Ministry of Agriculture Fisheries and FoodLondon UK Her Majestyrsquos Stationery Office pp 39-54
HUELSENBECK JP amp RONQUIST F (2001) MRBAYESBayesian inference of phylogeny Bioinformatics 17 754-755
HUETTEL RN (1985) Carrot disc culture In ZuckermanBM Mai WF amp Harrison MB (Eds) Plant nematologylaboratory manual Amherst MA USA Agricultural Exper-iment Station pp 153-154
INSERRA RN DUNCAN LW VOVLAS N amp LOOFPAA (1996) Pratylenchus loosi from pasture grasses incentral Florida Nematologica 32 159-172
INSERRA RN DUNCAN LW TROCCOLI A DUNND MAYA DOS SANTOS J KAPLAN D amp VOVLASN (2001) Pratylenchus jaehni sp n from citrus in Braziland its relationship with P coffeae and P loosi (NematodaPratylenchidae) Nematology 3 653-665
INSERRA RN TROCCOLI A GOZEL U BERNARD ECDUNN D amp DUNCAN L (2007) Pratylenchus hippeastrin sp (Nematoda Pratylenchidae) from amaryllis in Floridawith notes on P scribneri and P hexincisus Nematology 925-42
JENKINS WR (1964) A rapid centrifugal-flotation method forseparating nematodes from soil Plant Disease Reporter 48692
KARSSEN G WAEYENBERGE L amp MOENS M (2000)Pratylenchus brzeskii sp nov (Nematoda Pratylenchidae)a root-lesion nematode from European coastal dunes AnnalesZoologici 50 255-261
LOOF PAA (1960) Taxonomic studies on the genus Praty-lenchus (Nematoda) Tijdschrift voor Plantenziekten 66 29-90
LOOF PAA (1991) The family Pratylenchidae Thorne 1949In Nickle WR (Ed) Manual of agricultural nematologyNew York NY USA Marcel Dekker pp 363-421
MIZUKUBO T SUGIMURA K amp UESUGI K (2007)A new species of the genus Pratylenchus from chrysanthe-mum in Kyushu western Japan (Nematoda Pratylenchidae)Japanese Journal of Nematology 37 63-74
MUacuteNERA G BERT W amp DECRAEMER W (2009) Morpho-logical and molecular characterisation of Pratylenchus arau-censis n sp (Pratylenchidae) a root-lesion nematode associ-ated with Musa plants in Colombia Nematology 11 799-813
Vol 12(6) 2010 867
F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology

F De Luca et al
ORUI Y (1996) Discrimination of the main Pratylenchusspecies (Nematoda Pratylenchidae) in Japan by PCR-RFLPanalysis Applied Entomology and Zoology 31 505-514
DE LA PENtildeA E MOENS M VAN AELST A amp KARSSENG (2006) Description of Pratylenchus dunensis sp n (Ne-matoda Pratylenchidae) a root-lesion nematode associatedwith the dune grass Ammophila arenaria (L) Link Nemato-logy 8 70-88
RYSS A (1982) [New species of plant nematodes fromthe genus Pratylenchus in Estonia] Eesti NSV TeadusteAkaceemia Toimetised Izvestiya Akademii Nauk EstonskoiSSR Biologia 31 22-29
SEINHORST JW (1968) Three new Pratylenchus species witha discussion of the structure of the cephalic framework and ofthe spermatheca in this genus Nematologica 14 497-515
SIDDIQI MR (1994) Nematodes of tropical rainforests 4Two new species of Pratylenchus Afro-Asian Journal ofNematology 4 190-193
SIDDIQI MR (2000) Tylenchida parasites of plants andinsects 2nd edition Wallingford UK CABI Publishing 848pp
SIDDIQI MR DABUR KR amp BJAJ HK (1991) Descrip-tions of three new species of Pratylenchus Filipjev 1936 (Ne-matoda Pratylenchidae) Nematologia Mediterranea 19 1-7
SUBBOTIN SA amp MOENS M (2006) Molecular taxonomyand phylogeny In Perry RN amp Moens M (Eds) Plantnematology Wallingford UK CABI Publishing pp 33-58
SUBBOTIN SA RAGSDALE EJ MULLENS T ROBERTSPA MUNDO-OCAMPO M amp BALDWIN JG (2008)A phylogenetic framework for root lesion nematodes of the
genus Pratylenchus (Nematoda) Evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters Molecular Phylogenetics andEvolution 48 491-505
SWOFFORD DL (2002) PAUPlowast Phylogenetic Analysis UsingParsimony (lowastand other methods) Sunderland MA USASinauer Associates
THOMPSON JD GIBSON TJ PLEWNIAK F JEANMOU-GIN F amp HIGGINS DG (1997) The Clustal X windowsinterface flexible strategies for multiple sequence alignmentaided by quality analysis tools Nucleic Acids Research 244876-4882
THORNE G amp MALEK RB (1968) Nematodes of the North-ern Great Plains I Tylenchida South Dakota AgriculturalExperimental Station Technical Bulletin 31 111 pp
WAEYENBERGE L RYSS A MOENS M PINOCHET Jamp VRAIN TC (2000) Molecular characterisation of 18Pratylenchus species using rDNA restriction fragment lengthpolymorphism Nematology 2 135-142
WAEYENBERGE L VIAENE N amp MOENS M (2009)Species-specific duplex PCR for the detection of Praty-lenchus penetrans Nematology 11 847-857
ZARINA B amp MAQBOOL MA (1998) Descriptions andobservations on two new and two known species of the genusPratylenchus Filipjev 1936 (Nematoda Pratylenchidae) fromPakistan Pakistan Journal of Nematology 16 13-24
ZHENG JW amp CHEN PS (1994) Primary report on a newspecies in the genus Pratylenchus Journal of Shanxi Agricul-tural University 14 387-390
868 Nematology