characterization and enzyme engineering of laccases
TRANSCRIPT

University of Kentucky University of Kentucky
UKnowledge UKnowledge
Theses and Dissertations--Biosystems and Agricultural Engineering Biosystems and Agricultural Engineering
2020
CHARACTERIZATION AND ENZYME ENGINEERING OF LACCASES CHARACTERIZATION AND ENZYME ENGINEERING OF LACCASES
TOWARDS LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS TOWARDS LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS
Joseph Stevens University of Kentucky, [email protected] Author ORCID Identifier:
https://orcid.org/0000-0002-2595-9256 Digital Object Identifier: https://doi.org/10.13023/etd.2020.346
Right click to open a feedback form in a new tab to let us know how this document benefits you. Right click to open a feedback form in a new tab to let us know how this document benefits you.
Recommended Citation Recommended Citation Stevens, Joseph, "CHARACTERIZATION AND ENZYME ENGINEERING OF LACCASES TOWARDS LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS" (2020). Theses and Dissertations--Biosystems and Agricultural Engineering. 72. https://uknowledge.uky.edu/bae_etds/72
This Doctoral Dissertation is brought to you for free and open access by the Biosystems and Agricultural Engineering at UKnowledge. It has been accepted for inclusion in Theses and Dissertations--Biosystems and Agricultural Engineering by an authorized administrator of UKnowledge. For more information, please contact [email protected].

STUDENT AGREEMENT: STUDENT AGREEMENT:
I represent that my thesis or dissertation and abstract are my original work. Proper attribution
has been given to all outside sources. I understand that I am solely responsible for obtaining
any needed copyright permissions. I have obtained needed written permission statement(s)
from the owner(s) of each third-party copyrighted matter to be included in my work, allowing
electronic distribution (if such use is not permitted by the fair use doctrine) which will be
submitted to UKnowledge as Additional File.
I hereby grant to The University of Kentucky and its agents the irrevocable, non-exclusive, and
royalty-free license to archive and make accessible my work in whole or in part in all forms of
media, now or hereafter known. I agree that the document mentioned above may be made
available immediately for worldwide access unless an embargo applies.
I retain all other ownership rights to the copyright of my work. I also retain the right to use in
future works (such as articles or books) all or part of my work. I understand that I am free to
register the copyright to my work.
REVIEW, APPROVAL AND ACCEPTANCE REVIEW, APPROVAL AND ACCEPTANCE
The document mentioned above has been reviewed and accepted by the student’s advisor, on
behalf of the advisory committee, and by the Director of Graduate Studies (DGS), on behalf of
the program; we verify that this is the final, approved version of the student’s thesis including all
changes required by the advisory committee. The undersigned agree to abide by the statements
above.
Joseph Stevens, Student
Dr. Jian Shi, Major Professor
Dr. Donald Colliver, Director of Graduate Studies

CHARACTERIZATION AND ENZYME ENGINEERING OF LACCASES TOWARDS
LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS
________________________________________
DISSERTATION
________________________________________
A dissertation submitted in partial fulfillment of the
requirements for the degree of Doctor of Philosophy in the
Colleges of Agriculture and Engineering
at the University of Kentucky
By
Joseph Stevens
Lexington, Kentucky
Co- Directors: Dr. Jian Shi, Assistant Professor of Biosystems and Agricultural
Engineering
and Dr. Sue Nokes, Professor of Biosystems and Agricultural Engineering
Lexington, Kentucky
2020
Copyright © Joseph Stevens 2020
https://orcid.org/0000-0002-2595-9256

ABSTRACT OF DISSERTATION
CHARACTERIZATION AND ENZYME ENGINEERING OF LACCASES TOWARDS
LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS
Lignin is one of the most abundant polymers found in nature, making up 15 – 40%
of the weight of terrestrial biomass. Due to the structural and monomeric heterogeneity of
lignin, it is recalcitrant thermochemical and biological valorization methods. Converting
lignin to value-added products via sustainable and cost-effective pathways will reduce
waste and add value to future cellulosic biorefineries. Biological methods for lignin
valorization (e.g. lignin degrading enzymes or microbes) is limited by low lignin solubility
in biocompatible solvents, resulting in low product yield. Recent reports on biocatalysts
for lignin valorization have focused on the lignolytic multicopper oxidase laccase, which
operates in a wide range of conditions, does not require additional solvents such as
hydrogen peroxide, and can oxidize nonphenolic linkages when used alongside a small
molecule mediator.
To improve the yield of products from biological lignin valorization, it is necessary
to use a biocompatible solvent that is capable of solubilizing lignin in mild conditions
alongside a biocatalyst capable of operating in high concentrations of the solvent. Ionic
liquids (ILs) are molten salts that are liquid at temperatures < 100oC. The properties of ILs
can be tuned by selecting the appropriate cation and anion. Many ILs have been developed
for biomass pretreatment, for example 1-ethyl-3-methylimidazolium acetate
([C2C1Im][OAc]), cholinium lysinate ([Ch][Lys]), and diethylamine hydrogen sulfate
([DEA][HSO4]). We screened the biocompatibility of these 3 ILs with the activity of a
mesophilic fungal laccase from Trametes versicolor (TvL). [C2C1Im][OAc] and [Ch][Lys]
severely inhibited TvL activity in low concentrations (<5%), whereas minimal loss of TvL
activity was observed in 10% [DEA][HSO4]. Treating Kraft lignin with the laccase-
mediator-IL system resulted in low product yields and minimal changes to the lignin

structure, but nonetheless served as a promising proof of concept upon which future studies
can improve.
The extreme conditions (pH, temperature, salinity) in which thermophilic enzymes
operate makes them desirable biocatalysts for use in many biotechnological applications,
including lignin valorization. Thermophilic cellulases show minimal loss of activity in
20% [C2C1Im][OAc]. The hyperthermophilic bacterium Thermus thermophilus is reported
to produce the most thermophilic laccase (TtL) to date, demonstrating optimal substrate
oxidation at 92oC. Despite the thermophilicity of TtL, severe activity loss was observed in
<3% [C2C1Im][OAc] via a mixed competitive and noncompetitive inhibition mechanism.
A rational design strategy, guided by previous enzyme engineering studies for improving
laccase activity in ILs, produced 8 single amino acid mutants, none of which improved the
activity of TtL in [C2C1Im][OAc] compared to the wild type. These results highlight the
challenges faced when designing a biological lignin valorization system with a laccase and
an aqueous ILs.
Recent reports show the IL 1-ethyl-3-methylimidazolium ethylsulfate
([C2C1Im][EtSO4]) is biocompatible with the activity of a laccase from the thermophilic
fungus Myceliophthora thermophila (MtL) up to 25% IL concentration. When we screened
the biocompatibility of [C2C1Im][OAc] with MtL, severe inhibition of MtL activity was
observed in >15% [C2C1Im][OAc]. The surface charges of MtL were modified to increase
activity in [C2C1Im][OAc]. Increasing the acid or amine residues on the enzyme surface
did not affect the activity or stability of MtL in [C2C1Im][OAc]. Docking simulations
showed that the modifications do not significantly alter the interactions between the IL and
the enzyme surface. These results suggest that IL inhibition of laccases is not due to IL
docking on the enzyme surface, but rather destabilization of the coppers.
We applied a mixed technique approach using in vitro and in silico methods to
understand the interactions between Toxicodendron vernicifluum laccase (RvL) and
[Ch][Lys]. Biocompatibility, kinetics, and docking simulations support that [Ch][Lys]
inhibits RvL through a mixed competitive and noncompetitive inhibition mechanism.
Molecular dynamics (MD) simulations showed that the IL affects the stability of the
substrate in the active site.
Collectively, these studies characterize the interactions between several mesophilic
and thermophilic laccases and ILs used for biomass pretreatment with a variety of in vitro
and in silico techniques. The information provided by these studies can be used to inform
the design of future biological lignin valorization systems using laccase and a
biocompatible, aqueous IL.
KEYWORDS: Lignin, biocatalysis, biorefinery, ionic liquids, enzyme engineering
Joseph Craig Stevens
(Name of Student)
July 27, 2020
Date

CHARACTERIZATION AND ENZYME ENGINEERING OF LACCASES
TOWARDS LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS
By
Joseph Craig Stevens
Dr. Jian Shi
Co- Director of Dissertation
Dr. Sue Nokes
Co- Director of Dissertation
Dr. Donald Colliver
Director of Graduate Studies
July 27, 2020
Date

iii
ACKNOWLEDGMENTS
I would like to first and foremost thank my adviser of the last 4 years, Dr. Jian Shi.
Without his guidance I would not be the researcher I am today. Without his patience I
would not understand the virtue of patience in research and understanding that good
science takes time. Without his faith in me I would not have the confidence to pursue my
future in research. I owe him a great debt of gratitude for helping shape my personal and
professional future.
I would also like to thank Dr. David Rodgers for his instruction in my first
biochemistry courses that influenced my research interests during my Ph.D. studies. You
have shown me a great deal of patience, kindness, and insight while helping me with my
research and serving on my Ph.D. committee. I would also like to thank my other
committee members, Dr. Sue Nokes and Dr. Michael Montross, who provided me with
valuable insight and helped me build confidence in myself during my Ph.D. I would also
like to thank my committee for having faith in me to pursue a direct-to-Ph.D. pathway and
to spend my 9 months in France wisely.
For my time in Toulouse I owe a great deal to my collaborators and friends at the
Toulouse Biotechnology Institute (TBI) in France and the Chateaubriand Fellowship from
the Office for Science & Technology of the Embassy of France in the United States. By
agreeing to be my research host for the fellowship, Dr. Claire Dumon has played a crucial
role in shaping my confidence and abilities as a person and a researcher. The patience of
Dr. Isabelle André and Dr. Jérémy Esque knows no bounds and I thank them for spending
many hours working with me on the bioinformatics portions of my later chapters. I also
owe a deep debt of gratitude to the kindness and English-speaking abilities of my co-

iv
workers at TBI, Dr. Cédric Montanier, Dr. Régis Fauré, Dr. Jiao Zhao, Dr. Tarun Narwani,
and Thomas Enjalbert. I would also like to thank my host family in Toulouse, Dominique,
Nathalie, Lucy, and Jenny Bergonier for helping me navigate Toulouse upon my arrival
and for sharing many Sunday lunches with me. To all of my friends in Toulouse: you
made my 9 months there a once-in-a-lifetime experience and I promise that I will learn
more French before I return.
I would like to extend a special thank you to Dr. Lalitendu Das, a former post-doc
in Dr. Shi’s lab and a future co-worker (again), for lecturing and challenging me that first
morning at the conference in San Francisco. I owe you a great deal for the experiences
I’ve had the last 4 years, not least of which are going to Spalding’s for an early morning
doughnut, and you have played a critical role in shaping my future. I look forward to
working with you again.
I would like to thank the friends I have made in Lexington and Toulouse these last
4 years: Ryan, Woomer, Wenqi shishou, Lalit, Jameson, Jérémy, and Tarun. You all mean
more to me than I can express in words because I only know how to say how amazing
everything is, how much I love the simplicity of Ramirez tacos and a burger from Five
Guys, and how that really was the best pizza we’ve ever made.
I would like to thank the rest of the Shi lab group, Enshi Liu, Luke Dodge, Truc
Nguyen, Siquan Xu, Makua Vin-Nnajiofor, Can Liu, Dr. Binling Ai and Yuxuan Zhang
along with Dr. Udeep Chawla from Dr. Rodgers’ lab for providing me with a great deal
of support and great memories the past several years. I look forward to hearing about all
of your future research endeavors.

v
To all of the faculty, staff, and fellow graduate students in the BAE department,
thank you for your kindness and for allowing a diehard Louisville Cardinal fan to work
alongside you all. Maybe Cardinals and Wildcats aren’t so different after all.
I would like to thank my middle school science teacher, Mr. Fred Whittaker, for
instilling in me a passion for science and, above all else, compassion for my fellow
humans.
I would like to thank my parents, Barry and Mary Beth, and my brother, Porter,
for pretending to know what a “lignin” is and for showing me unconditional love and
support for the last 26 years.
A special, albeit strange, thank you to Malcolm James McCormick for helping me
to appreciate the things I have yet to experience. One day I’ll be brave enough to listen to
Circles, but Swimming is still teaching me what it’s all about.
And lastly, I would like to thank Robert “Papa” Rowan for keeping me busy on
Sundays in Louisville. I sure hope San Francisco is as amazing as you made it out to be.

vi
TABLE OF CONTENTS
ACKNOWLEDGMENTS ........................................................................................................................... III
LIST OF TABLES ....................................................................................................................................... XI
LIST OF FIGURES ................................................................................................................................... XII
LIST OF ADDITIONAL FIGURES .........................................................................................................XV
CHAPTER 1. BIOCATALYSIS IN AQUEOUS IONIC LIQUIDS FOR LIGNIN
VALORIZATION: OPPORTUNITIES AND RECENT DEVELOPMENTS1 ....................................... 1
1.1 INTRODUCTION ................................................................................................................................ 1
1.2 LIGNIN............................................................................................................................................. 3
1.2.1 Lignin structure and characteristics ................................................................................... 3
1.2.2 Lignin depolymerization methods ....................................................................................... 7
1.3 LIGNIN DEGRADING ENZYMES ......................................................................................................... 9
1.3.1 Peroxidases ......................................................................................................................... 9
1.3.2 Laccases ............................................................................................................................ 11
1.3.3 Other LDEs ....................................................................................................................... 14
1.4 IONIC LIQUIDS ............................................................................................................................... 18
1.4.1 IL properties...................................................................................................................... 18
1.4.2 Biomass pretreatment with ILs ......................................................................................... 21
1.4.3 Effect of IL treatment on lignin characteristics ................................................................ 25
1.4.4 Rationale for pairing LDE and IL for lignin depolymerization ........................................ 26
1.5 ENZYME-IL INTERACTIONS ........................................................................................................... 27
1.5.1 Enzyme activity and stability in ILs .................................................................................. 27
1.5.2 Effect of IL on enzyme ....................................................................................................... 30
1.5.3 Methods for improving enzyme stability in ILs ................................................................. 32
1.6 PROTEIN ENGINEERING TO IMPROVE IL COMPATIBILITY ............................................................... 35

vii
1.6.1 Structure-function relationship of proteins ....................................................................... 35
1.6.2 Protein engineering for improved function....................................................................... 38
1.6.3 Protein crystallization in ILs ............................................................................................ 41
1.7 PRODUCT SELECTIVITY ................................................................................................................. 41
1.7.1 Controlling product formation via biocatalyst selectivity ................................................ 42
1.7.2 Controlling product formation via IL selectivity .............................................................. 42
1.7.3 Preventing lignin repolymerization and improving yield ................................................. 43
1.8 CONCLUSIONS AND PERSPECTIVES ................................................................................................ 45
1.9 ORGANIZATION OF CHAPTERS....................................................................................................... 46
CHAPTER 2. UNDERSTANDING LACCASE-IONIC LIQUID INTERACTIONS TOWARDS
BIOCATALYTIC LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS1 ............................. 49
2.1 ABSTRACT ..................................................................................................................................... 49
2.2 INTRODUCTION .............................................................................................................................. 50
2.3 METHODS AND MATERIALS .......................................................................................................... 53
2.3.1 Materials ........................................................................................................................... 53
2.3.2 Biocompatibility screening ............................................................................................... 54
2.3.3 Inhibition kinetics.............................................................................................................. 54
2.3.4 Molecular docking simulations ......................................................................................... 55
2.3.5 Lignin depolymerization in aqueous IL ............................................................................ 55
2.3.6 Lignin characterization ..................................................................................................... 57
2.4 RESULTS AND DISCUSSION ............................................................................................................ 60
2.4.1 Biocompatibility screening ............................................................................................... 60
2.4.2 Inhibition mechanisms ...................................................................................................... 62
2.4.3 Docking simulations.......................................................................................................... 63
2.4.4 GGE dimer transformation ............................................................................................... 64
2.4.5 Depolymerization of alkaline lignin.................................................................................. 68
2.5 CONCLUSIONS ............................................................................................................................... 76
2.6 AUTHOR CONTRIBUTIONS ............................................................................................................. 76

viii
2.7 ACKNOWLEDGEMENTS .................................................................................................................. 77
2.8 SUPPLEMENTAL INFORMATION ..................................................................................................... 78
CHAPTER 3. CHARACTERIZATION AND ENZYME ENGINEERING OF A
HYPERTHERMOPHILIC LACCASE TOWARD IMPROVING ITS ACTIVITY IN IONIC
LIQUID1 83
3.1 ABSTRACT ..................................................................................................................................... 83
3.2 INTRODUCTION .............................................................................................................................. 84
3.3 MATERIALS AND METHODS .......................................................................................................... 88
3.3.1 Materials ........................................................................................................................... 88
3.3.2 Expression of T. thermophilus Laccase ............................................................................ 88
3.3.3 Purification and copper-induced laccase folding ............................................................. 89
3.3.4 Biocompatibility screening ............................................................................................... 89
3.3.5 Inhibition kinetics.............................................................................................................. 90
3.3.6 Docking Simulations ......................................................................................................... 90
3.3.7 Site-directed mutagenesis of TtL ....................................................................................... 91
3.4 RESULTS ........................................................................................................................................ 92
3.4.1 WT TtL Biocompatibility Screening .................................................................................. 92
3.4.2 Enzyme Kinetics ................................................................................................................ 93
3.4.3 Docking simulations.......................................................................................................... 94
3.4.4 Rational Design of TtL ...................................................................................................... 96
3.5 DISCUSSION ................................................................................................................................... 98
3.6 AUTHOR CONTRIBUTIONS ............................................................................................................. 99
3.7 ACKNOWLEDGEMENTS .................................................................................................................. 99
3.8 SUPPLEMENTAL INFORMATION ................................................................................................... 100
CHAPTER 4. MODIFYING SURFACE CHARGES OF A THERMOPHILIC LACCASE
TOWARDS IMPROVING ACTIVITY AND STABILITY IN AQUEOUS IONIC LIQUID ........... 104
4.1 ABSTRACT ................................................................................................................................... 104

ix
4.2 INTRODUCTION ............................................................................................................................ 105
4.3 METHODS AND MATERIALS ........................................................................................................ 110
4.3.1 Materials ......................................................................................................................... 110
4.3.2 Surface Charge Modifications ........................................................................................ 110
4.3.3 Biocompatibility Screening ............................................................................................. 111
4.3.4 MtL Thermostability........................................................................................................ 112
4.3.5 Docking Simulations ....................................................................................................... 112
4.4 RESULTS AND DISCUSSION .......................................................................................................... 113
4.4.1 Biocompatibility Screening ............................................................................................. 113
4.4.2 Stability [C2C1Im][OAc] ................................................................................................. 115
4.4.3 Docking Simulations ....................................................................................................... 117
4.5 CONCLUSIONS AND FUTURE WORK ............................................................................................ 119
4.6 AUTHOR CONTRIBUTIONS ........................................................................................................... 123
4.7 ACKNOWLEDGEMENTS ................................................................................................................ 123
CHAPTER 5. A MIXED TECHNIQUE APPROACH TOWARDS UNDERSTANDING IONIC
LIQUID INHIBITION OF A PLANT LACCASE ................................................................................. 125
5.1 ABSTRACT ................................................................................................................................... 125
5.2 INTRODUCTION ............................................................................................................................ 126
5.3 METHODS AND MATERIALS ........................................................................................................ 129
5.3.1 Materials ......................................................................................................................... 129
5.3.2 Plant laccase purification ............................................................................................... 129
5.3.3 Biocompatibility screening ............................................................................................. 130
5.3.4 Enzyme Kinetics .............................................................................................................. 130
5.3.5 Plant laccase modelling and copper parameterization .................................................. 131
5.3.6 Docking Simulations ....................................................................................................... 132
5.4 RESULTS AND DISCUSSION .......................................................................................................... 132
5.4.1 Biocompatibility of AILs with laccase activity................................................................ 132
5.4.2 Inhibition Mechanisms .................................................................................................... 134

x
5.4.3 Copper parameterization and stabilization during simulations ..................................... 135
5.4.4 IL Docking ...................................................................................................................... 136
5.4.5 Steered Molecular Dynamics of DMP in [Ch][Lys]....................................................... 137
5.5 AUTHOR CONTRIBUTIONS ........................................................................................................... 138
5.6 ACKNOWLEDGEMENTS ................................................................................................................ 139
5.7 SUPPLEMENTAL INFORMATION ................................................................................................... 140
CHAPTER 6. CONCLUSIONS AND FUTURE WORK .................................................................. 142
6.1 CONCLUSIONS ............................................................................................................................. 142
6.2 FUTURE WORK ............................................................................................................................ 144
BIBLIOGRAPHY ...................................................................................................................................... 145
VITA ........................................................................................................................................................... 167

xi
LIST OF TABLES
Table 1-1. Applications of lignolytic enzymes ................................................................. 17
Table 1-2. Studies of enzyme activity and stability in ILs ............................................... 28
Table 2-1. Kinetic constants of ABTS oxidtion by T. versicolor laccase in AILs ........... 62
Table 2-2. Molecular weight distribution of GGE before (0 min), during (30 min), and after
(120 min) treatment with TvL, ABTS, and ILs1................................................................ 66
Table 2-3. Converstion and yield of alkaline lignin degradation products ....................... 69
Table 2-4. Processed lignin molecular weight distribution and polydispersity of lignin
streams1 ............................................................................................................................. 71
Table 3-1. Kinetic constants of the WT TtL in different concentrations of [C2C1Im][OAc].
Values shown are mean ± standard error (SE) of 3 experiments. ..................................... 94
Table 3-2. Activity of WT and mutants relative to activity in buffer in different
concentrations of [C2C1Im][OAc]. Values shown are mean ± standard deviation (SD) of 3
experiments1. ..................................................................................................................... 97
Table 4-1. Summary of mutations to used mimic surface charge modifications in PyMol
......................................................................................................................................... 113
Table 4-2. Previous studies using various methods to improve enzyme activity in aqueous
ILs ................................................................................................................................... 123
Table 5-1. Kinetic constants of RvL in different concentrations of [Ch][Lys]. Values shown
are mean ± standard error of 3 experiments .................................................................... 134

xii
LIST OF FIGURES
Figure 1-1. Representative lignin structure consisting syringyl and guaiacyl units (adapted
from 42). ............................................................................................................................... 4
Figure 1-2. Formation of inter-unit linkages during polymerization of coniferyl alcohol
monomers (adapted from 41). .............................................................................................. 5
Figure 1-3. a) Lignin peroxidase from Phanerochaete chrysosporium (PDBID: 1B82) 72,
b) Manganese peroxidase from Phanerochaete chrysosporium (PDBID: 1MNP) 73 and c)
Versatile peroxidase from Pleurotus eryngii (PDBID: 2BOQ) 74. Figures downloaded from
the Research Collaboratory for Structural Bioinformatics’ Protein Data Bank (RCSB
PDB)75 ............................................................................................................................... 10
Figure 1-4. Laccase from Trametes versicolor (PDBID: 1GYC) 86. Figure downloaded
from the Research Collaboratory for Structural Bioinformatics’ Protein Data Bank (RCSB
PDB) 75. ............................................................................................................................. 12
Figure 1-5. Lignolytic pathways of Sphingobium sp. strain SYK-6 121. .......................... 15
Figure 1-6. a) [C2C1Im][OAc] has served as the benchmark IL for biomass pretreatment
for several years. Recently, bioderived ILs like b) choline lysinate ([Ch][Lys]) and c)
diethylamine hydrogen sulfate ([DEA][HSO4]) have attracted interest due to their low cost.
........................................................................................................................................... 21
Figure 1-7. Butylimidazolium ILs. These ILs have all been effectively utilized in biomass
pretreatment strategies with different effects on downstream processing of biomass
(adapted from 22). .............................................................................................................. 23
Figure 1-8. Effect of surface charge modification on IL interaction with enzymes
(Nordwald, 2015). ............................................................................................................. 35
Figure 1-9. Project summary............................................................................................. 46
Figure 2-1. Activity of T. versicolor laccase in AIL solutions. H = high laccase
concentration (3.0 μg mL-1), M = medium laccase concentration (1.2 μg mL-1), L = low
laccase concentration (0.6 μg mL-1). Values indicate the ratio of TvL activity in ILs relative
to TvL activity in buffer (0% IL)....................................................................................... 61
Figure 2-2. Results of simulations showing the docking of b) [C2C1Im][OAc], c) [Ch][Lys],
and d) [DEA][HSO4] near the Type I copper of T. versicolor laccase. The red arrow in a)
points to the Type I copper. The ILs are represented by the multi-colored stick models in
panels b-d and the His residues that coordinate the coppers are colored blue. ................ 63
Figure 2-3. HPLC chromatograms of GGE incubated with laccase and different AILs a)
without ABTS present during the reaction. b) ABTS present during the reaction. GGE
elutes at ~23 min and reaction products elute in the 2-5 min range. ................................ 65

xiii
Figure 2-4. Molecular weight distributions of products after GGE reaction with TvL,
ABTS, and a) buffer without IL, b) 2% [C2C1Im][OAc], c) 1% [Ch][Lys], d) 10%
[DEA][HSO4] .................................................................................................................... 67
Figure 2-5. GC-MS chromatograms of alkaline lignin breakdown products after treatment
with laccase, AILs, and ABTS. Major products identified were 1) guaiacol, 2) vanillin, 3)
acetovanillone, 4) homovanillic acid, 5) syringaldehyde, and 6) acetosyringone. ........... 70
Figure 2-6. 13C-1H (HSQC) spectra of aliphatic (top) and aromatic (bottom) regions of a)
raw alkaline lignin (KL), b) alkaline lignin treated with laccase and ABTS in buffer (KLA),
and c) alkaline lignin treated with laccase and ABTS in 2% [C2C1Im][OAc] (EKLA).
Figure S6 in the supplementary material contains all of the spectra from the study. The
structures of side-chain linkages and lignin compositional units, shown in the middle of the
figure, were coded with colors corresponding to the cross peaks in the spectra aliphatic and
aromatic regions, respectively. Linkage percentages were calculated via volume
integration of the Cα-H of the β-O-4', β-5', and β-β' linkages and treated as a relative ratio
of the three. Volume integrations are intended to be only semi-quantitative due to problems
inherent to HSQC experiments including a wide array of potential 1JC-H values and rates of
T2 relaxation 260. ................................................................................................................ 73
Figure 3-1. Activity of the WT TtL in different concentrations of [C2C1Im][OAc] (A).
Michaelis-Menten curves of TtL in different concentrations of [C2C1Im][OAc] (B). Values
shown are mean with error bars representing the SD of 3 experiments. .......................... 92
Figure 3-2. Top docking locations from AutoDock of [C2C1Im][OAc] to the surface of [E]
(A) and [E][S] (B). The IL is shown as magenta spheres, the coppers are shown as brown
spheres, and ABTS is shown as red sticks. ....................................................................... 95
Figure 4-1. Clustal Omega sequence alignment of laccases from a bacterium (T.
thermophilus – 2XU9), a plant (T. vernicifluum - Plant), and fungi (T. versicolor – 1GYC,
M. thermophila – 6F5K). Conserved HXH motifs and T1-coordinating
methionine/phenylalanine/leucine are highlighted with thick blue bars......................... 108
Figure 4-2. Heatmap showing the activity of MtL and the charge variants in
[C2C1Im][OAc] relative to MtL activity in buffer. ......................................................... 114
Figure 4-3. Heatmaps showing the residual activity of MtL charge variants after incubation
in 50 mM citrate/100 mM phosphate buffer and 2.5 or 5% [C2C1Im][OAc] at pH 4.5 and
40oC................................................................................................................................. 116
Figure 4-4. Major docking locations of [C2C1Im][OAc] to the surface of unmodified (a),
succinylated/acetylated (b), and cationized/neutralized MtL (c). [C2C1Im]+ is shown as
magenta spheres, [OAc]- is shown as red spheres, and coppers are shown as brown spheres
......................................................................................................................................... 118
Figure 5-1. Effect of aqueous [Ch][Lys] on the activity of RvL. Values shown are the mean
± SD of 3 experiments .................................................................................................... 133

xiv
Figure 5-2. Distances between (top) T1 copper, (middle) T2 copper, and (bottom) T3
coppers and coordinating residues during 100 ns simulations........................................ 136
Figure 5-3. Major docking locations of [Ch][Lys] to the surface of [E] (panel a, green) and
[E][S] (panel b, cyan). [Ch]+ is shown as red spheres, [Lys]+ is shown as magenta spheres,
DMP is shown as yellow spheres, and coppers are shown as brown spheres. ............... 137
Figure 5-4. Free energy profile of DMP in 0 (blue line), 2 (green line), and 4% (red line)
[Ch][Lys]. The measurement of the distance between the T1 copper of DMP hydroxyl
group is also shown for reference. .................................................................................. 138

xv
LIST OF ADDITIONAL FIGURES
Table S 3-1. List of primer names and sequences for making the mutants in this study.
Mutated nucleotides are underlined. ............................................................................... 100
Figure S 2-1. Hanes-Woolf curves of T. versicolor laccase in a) [C2C1Im][OAc], b)
[Ch][Lys], and c) [DEA][HSO4] ....................................................................................... 78
Figure S 2-2. Structure of proposed a) coupled GGE dimers, b) three coupled GGE dimers,
and c) addition of a G monomer unit to the S2b structure formed during GGE
polymerization reaction with T. versicolor laccase, ABTS, and AILs. ............................ 79
Figure S 2-3. a) FTIR spectra of alkaline lignin before and after treatment with laccase,
AILs, and ABTS (The red arrow is indicating the corresponding S-lignin peak); b) Thermal
profiles of alkaline lignin before and after treatment with laccase, AILs, and ABTS. ..... 80
Figure S 2-4. Molecular weight distribution of a) solid-phase and b) liqui-phase alkaline
lignin after treatment with laccase, AILs, and ABTS ....................................................... 81
Figure S 2-5. 13C-1H (HSQC) spectra of alkyl (top) and aromatic (bottom) regions of a)
raw alkaline lignin (KL), b) alkaline lignin treated with laccase and ABTS, and alkaline
lignin treated with laccase and ABTS in aqueous ionic liquids: c) [Ch][Lys], d)
[DEA][HSO4], and e) [C2C1Im][OAc]. *The structures of side-chain linkages and lignin
compositional units were coded with colors corresponding to the cross peaks in the spectra
alkyl and aromatic regions, respectively. .......................................................................... 82
Figure S 3-1. Crystal structures of TtL (PDBID: 2XU9, left) and TvL (PDBID: 1GYC,
right) showing domain 1 (green), domain 2 (orange), domain 3 (magenta), and the L1 loop
(cyan) connecting domains 2 and 3 (A). Sequence alignment of TtL and B. subtilis CotA,
homologous Glutamate residues are highlighted (B)...................................................... 101
Figure S 3-2. Aromatic and anionic residues around E170 in TtL (A) and E188 in B. subtilis
CotA (B). Glutamates targeted for mutations are shown in cyan while all other residues
are shown in green. All residues shown are within 5Å of the respective Glutamate. ... 102
Figure S 3-3. Codon optimized sequence of TtL (NCBI Accession Number: I7AL37) . 103
Figure S 5-1. Michaelis-Menten curves of RvL in aqueous [Ch][Lys]. Data were plotted
and models were fit in Sigma-Plot. Values plotted are mean ± SD of 3 experiments. .. 140
Figure S 5-2. Final charges and geometry of coppers and coordinating residues .......... 141

1
CHAPTER 1. BIOCATALYSIS IN AQUEOUS IONIC LIQUIDS FOR LIGNIN VALORIZATION:
OPPORTUNITIES AND RECENT DEVELOPMENTS1
1.1 Introduction
Lignin is synthesized biologically by plants and is one of the most abundant
biopolymers on the planet. Besides serving as one of the primary building blocks for plant
cell walls, lignin plays important roles such as facilitating water transport and protecting
the plant from pathogenic attack. As a result of human’s practice in utilizing plant materials
(i.e., paper and pulping industry), lignin is extracted from the cellulose portion and
commonly treated as a low-value/waste stream. Due to the recent surge in cellulosic
biofuel production, the volume of lignin from cellulosic biorefineries is predicted to reach
60 million tons annually by 2022, which will add on top of the lignin produced by the paper
industry 1. Currently, lignin is primarily burned in boilers to generate heat and steam that
power upstream processes 2, 3. Converting lignin to value-added chemicals will add
revenue to the biorefineries and reduce emitted CO2 when burned 3, 4. Despite its great
potential, lignin valorization is still challenging.
Lignin depolymerization is the first step toward chemical production since lignin
in its polymeric form has limited uses. The breakdown of lignin to low molecular weight
compounds helps to channel desirable functions and make suitable intermediates for
upgrading to fuels and chemicals. Lignin depolymerization can be categorized into
thermochemical routes such as high temperature pyrolysis, hydrogenolysis, and catalytic
oxidation and biological routes using lignin degrading enzymes (LDEs) and/or
microorganisms5-8. Tremendous efforts have been focused on improving the catalyst
1This chapter in its whole has been published in Biotechnology Advances. 2019, 37 (8), 107418.
DOI: 10.1016/j.biotechadv.2019.107418.

2
efficiency and selectivity in a thermochemical conversion process 9, 10. However, the
effectiveness and selectivity of the catalyst as developed on lignin model compounds often
get compromised when applied to plant derived lignin due to its intrinsic structural and
compositional complexity 11.
Biological lignin deconstruction can be performed at lower temperature and
pressure than thermochemical methods while product selectivity can be potentially
improved via the specificity of biocatalysts 12. The dominant lignolytic enzymes produced
by white and brown rot fungi or some Actinobacteria are lignin peroxidases, versatile
peroxidases, manganese peroxidases, and laccases. In addition, several unique pathways
for the cleavage of lignin linkages by nicotinamide adenine dinucleotide (NAD) or
glutathione dependent enzymes have been discovered in certain soil bacteria such as
Sphingobium sp. SYK-6 13-15. Application of these lignolytic enzymes in an industrially
relevant lignin valorization process is still hindered due to the high cost of enzyme
production and by the very low lignin solubility (for many industrial relevant lignin
streams) in the aqueous environment compatible with enzyme activity 16, 17. However,
most of the known effective lignin solvents such as hexane, dimethylsulfoxide (DMSO),
acetone, and alkalis are detrimental to enzyme activity in their pristine form or aqueous
solutions 18, 19 and often require high temperature for complete lignin dissolution especially
when dealing with plant derived native lignins 20. Thus, a better solvent system with both
high lignin solubility and enzyme compatibility is essential for developing a better
biological lignin depolymerization process.
As alternatives to common organic solvents, certain ionic liquids (ILs) have
demonstrated a remarkable ability to solubilize and depolymerize lignin during biomass

3
pretreatment 21-23. The cellulose portion recovered from IL pretreatment is highly
digestible by using cellulolytic enzymes while lignin in the liquid stream can undergo
additional processing and be converted to value-added chemicals 21. Taking advantage of
IL solvent systems, a number of base, acid and metal catalysts have been tested previously
for catalytically depolymerizing lignin in IL media 10, 24-28. Meanwhile the use of
biocatalysts in IL, i.e. LDEs has attracted increasing interest in recent years due to potential
advantages such as better selectivity and affinity, in milder reaction conditions. The
toxicity of certain ILs to enzymes and microbes however hinders direct product formation
via biological lignin depolymerization in pure IL or its aqueous solution.
There have been several articles reviewing the effects of ILs on enzymes such as
lipase, xylanase, cellulase, and laccase, etc. 29-37. To the best of our knowledge, however,
there has not been a review of the effects of ILs on LDEs in the context of lignin
valorization. This paper emphasizes the effects of IL on lignin characteristics, the
relationships between LDE protein structure and function and ILs (enzyme-IL
interactions), along with strategies for improving the efficiency and product selectivity of
LDEs in ILs for lignin depolymerization. Finally, possible pathways toward a lignin
valorization system using a biocompatible IL and LDE are proposed; challenges and
perspectives associated with designing such a system are also discussed.
1.2 Lignin
1.2.1 Lignin structure and characteristics
Lignin is a heterogeneous biopolymer, responsible for providing plants with
structural support, microbial resistance and channels for water and nutrient transport 38.

4
Lignin is constructed of three major subunits, p-coumaryl alcohol, coniferyl alcohol, and
sinapyl alcohol 39. These monolignols are differentiated based on the degree of
methoxylation of their phenyl rings; p-coumaryl alcohol has no methoxy group, coniferyl
alcohol has one methoxy group (2-methoxyphenol), and sinapyl alcohol has two methoxy
groups (2,6-dimethoxyphenol). These heterogeneous building blocks are the precursors to
what will ultimately become lignin. During lignin biosynthesis, the monolignols undergo
slight structural changes as they are converted to phenylpropanoids 39. The p-coumaryl
alcohol is converted to hydroxyphenol (H-lignin), coniferyl alcohol is converted to
guaiacyl (G-lignin), and sinapyl alcohol is converted to syringyl (S-lignin) 40, 41. Figure 1-
1 highlights the structural differences between the monolignols before and after lignin
biosynthesis; most notable is the oxidation of the double bond between the and tail
carbons.
Figure 1-1. Representative lignin structure consisting syringyl and guaiacyl units (adapted
from 42).
Monolignols are linked via a heterogeneous network of inter-unit linkages. The
inter-unit linkages seen in lignin are largely dictated by the monolignols present. The most
common linkage is the β-O-4’ linkage between H-, G-, or S-lignin 39, 41, 43. G- and H-lignin

5
can be linked together via β-5’, 5-5’-O-4, or 4-O-5’ linkages. H-, G-, and S-lignin can
further be linked together via β-β’ or β-1’ linkages 43. Figure 1-2 highlights the complexity
of the most common inter-unit linkages. The combination of different monolignols
alongside the network of inter-unit linkages, contributes to lignin’s inherent heterogeneity.
Figure 1-2. Formation of inter-unit linkages during polymerization of coniferyl alcohol
monomers (adapted from 41).
The source of the biomass determines important properties of lignin. The fraction
of lignin present in the biomass is dependent upon the source; softwoods contain 25-31%
lignin, while hardwoods contain 16-24% lignin 44. Additionally, the ratio of monolignols
present depends upon the source of biomass: softwoods primarily contain G-lignin,
hardwoods primarily contain G- and S-lignin, and grasses contain different ratios of H-, G-
, and S-lignin 44. It is this heterogeneity present at all levels of lignin that is responsible
for the challenges in selectively depolymerizing these molecules 45.
Plant cell wall engineering has attracted increasing interest in recent years. Genetic
modification/engineering the monolignol composition and/or inter-unit linkages present in
the native lignin, such as β-O-4’ linkages, could make lignin decomposition processes more

6
efficient. For example, Wilkerson et al. genetically modified poplar trees to synthesize
lignin from monolignol ferulate conjugates and demonstrated improved cell wall
digestibility after mild alkaline pretreatment of the engineered plants 46. In a follow-up
study, Parsell et al. applied a bimetallic Zn/Pd/C catalyst to breakdown engineered poplar
trees that contained higher levels of S-lignin; results suggest that the ester linkages present
in the genetically modified poplar resulted in higher levels of lignin digestion to a single
product (dihydroeugenol) when compared to native poplar 47.
Since lignin is such a vital component for plant growth, altering lignin biosynthesis
may have deleterious effects. Repression of a gene responsible for lignin biosynthesis
(hydroxycinnamoyl CoA:shikimate hydroxycinnamoyl transferase, HCT or p-coumaroyl
shikimate 3'-hydroxylase, C3'H) in Arabidopsis thaliana led to stunted growth of the plant,
a result of reduced auxin transport throughout the plant 48. Bonawitz et al. were able to
engineer an Arabidopsis mutant and simultaneously retain its growth and engineered lignin
properties 49. It was shown that disruption of a transcriptional regulatory complex, complex
subunits MED5a and MED5b restored lignin levels to those found in wild type
Arabidopsis. Interestingly, the resulting lignin was primarily made of H-lignin (~95%),
with trace amounts of G- and S- lignin, a trait greatly improving enzymatic digestibility of
this mutant after pretreatment 50. Engineering plants with lower lignin content, different
lignin composition and inter-unit linkages, and lignin deposition creates opportunities to
design better energy crops with traits suitable for bioconversion to biofuels and chemicals
40, 51.

7
1.2.2 Lignin depolymerization methods
Lignin is often underutilized in its polymeric form. Most of the lignin is burned to
produce heat and power; however, a small portion finds its applications as dispersants and
binders 52. The breakdown of lignin opens access to desirable functions that are not
achievable when lignin is a polymer; while the resulting low molecular weight compounds
become suitable for upgrading to fuel and chemicals. Several lignin depolymerization
pathways are being developed, including catalytic oxidation, pyrolysis, gasification, and
hydrogenolysis, etc. These methods utilize a suite of tools like high temperature, chemical
catalysts, and solvents to break down lignin. For example, Alkali processes typically use
sodium hydroxide (NaOH) in combination with an oxidative agent for lignin
decomposition, which are the most common methods in the paper industry. The two most
common alkali pretreatment methods are the Soda and Kraft processes, both of which use
NaOH along with other chemicals such as sodium sulfide and anthraquinone 53.
Ammonium hydroxide has also been shown to be effective for lignin decomposition 21.
While use of high pH methods is effective, chemistry was shown to be effective in
depolymerizing lignin in acid, alkaline, or organic solvents. Acid-catalyzed lignin
decomposition uses a variety of catalysts including mineral acids, Lewis acids, and ILs 54,
most notably through the use of dilute sulfuric acid (H2SO4). Lignin extracted from dilute
H2SO4 pretreated switchgrass was found to have a reduced molecular weight compared to
untreated lignin 21. One marker of lignin degradation is the formation of aromatic products
due to the aromatic nature of monolignols. Rahimi et al 55 used formic acid for lignin
decomposition, resulting in a yield of over 60 wt% aromatics under mild reaction
conditions. While oxidative chemistry has been shown to be effective to depolymerize

8
lignin in acid, alkaline or solvents, reductive chemistry using hydrogen molecules or
hydrogen donor provides another pathway.
Hydrogenolysis breaks down lignin using hydrogen with help from metal catalysts.
During hydrogenolysis, reductive bond cleavage takes place within lignin and/or lignin
model compounds in the presence of hydrogen as a reducing agent 56. A range of catalysts,
including noble metals (such as platinum), transition metals, and bimetallic catalysts, such
as Zn/Pd/C and Pd/CeO2, were reported to facilitate the cleavage of lignin into aromatic
products 47, 57. Alternatively, catalytic transfer hydrogenolysis (CTH) has shown great
promise 58. In the CTH reaction scheme, an equivalent of H2 is transferred from a donor
molecule to the acceptor molecule. Hydrogen donor molecules are often inexpensive
organic alcohols capable of readily generating hydrogen molecules and the same time
serving as solvents for lignin 59. A variety of hydrogen donating agents have been tested
including formic acid, methanol, ethanol, tetralin etc., among which isopropyl alcohol
remains a popular choice due to its relative low cost and easy subsequent separation from
the reaction mixture 60.
Pyrolysis is the breakdown of large molecules into smaller ones by the application
of heat in the absence of oxygen. During pyrolysis, lignin is heated to temperatures
between 160-900 °C where cleavage of the ether (C-O) and C-C linkages takes place 61.
Lignin pyrolysis produces a range of pyrolytic aromatic compounds in oil form in addition
to gas products and residual char 62, 63. The yield and composition of pyrolytic oil are
influenced by many factors, including lignin type and operation conditions 64.
As opposed to thermochemical lignin depolymerization methods, which require
intense energy input and/or harsh chemicals and costly catalysts, biological lignin

9
depolymerization takes advantage of microorganisms and their enzymatic excreta to
cleavage the inter-unit lignin linkages and lignin-carbohydrate linkages. The LDEs play
key roles in biological lignin depolymerization in nature. The next section summarizes the
key LDEs with emphasis on their temperature and pH optima (parameters relevant to IL
tolerance) and potential applications in lignin depolymerization.
1.3 Lignin degrading enzymes
1.3.1 Peroxidases
Lignin peroxidase (LiP), manganese peroxidase (MnP), and versatile peroxidase
(VP) are a series of iron-based LDEs present in nature 65-67. Numerous forms of these
LDEs have been isolated from a variety of lignolytic fungi and bacteria 65, 68-71.
Additionally, isozymes have been identified within individual species. For instance, 16
forms of LiP and 5 forms of MnP were isolated from liquid culture containing Trametes
versicolor 69. While these enzymes serve a similar purpose in nature, many have been found
to have unique properties.

10
Figure 1-3. a) Lignin peroxidase from Phanerochaete chrysosporium (PDBID: 1B82) 72,
b) Manganese peroxidase from Phanerochaete chrysosporium (PDBID: 1MNP) 73 and c)
Versatile peroxidase from Pleurotus eryngii (PDBID: 2BOQ) 74. Figures downloaded from
the Research Collaboratory for Structural Bioinformatics’ Protein Data Bank (RCSB
PDB)75
Peroxidases can retain activity in a wide range of pH and temperature. The optima
are dependent upon the enzyme’s source and the oxidation substrate. It was found that LiP
and MnP from a soil fungus Deuteromycete retain activity over a pH range of 4-11 and a
temperature range of 30-55 ºC 68. Fourteen forms of MnP and LiP from Pleurotus ostreatus
and Ceriporiopsis subvermispora were expressed in Escherichia coli; remarkably, an
isozyme of MnP from C. subvermispora retained over 80% activity at pH 2.0 76.
Recombinant VP from P. eryngii expressed in P. chrysosporium was found to have optimal
activity at pH 3.0, while retaining activity at temperatures up to 40 ºC 77. Fujii et al.
observed different pH optima for the LDEs collected at different depths from the forest

11
floor of regions in Japan, Thailand, and Indonesia. It was shown that MnP activity was
detected at pH greater than 5.0, while LiP activity was detected at pH less than 5.0 78. The
different pH optima suggest that LiP and MnP fulfill important roles in biodegradation of
plant matter at different depths of the forest floor. The optimal temperature for LDE
activity has also been observed to vary with respect to the substrate present. A peroxidase
from Bacillus subtilis had an optimal pH of 3.0, while maximum substrate oxidation was
observed at different temperatures from 30-50 ºC 79. LiP displayed optimal removal of a
nonylphenol at pH range 4.0-7.0 and temperature range of 25-50 ºC 66. Evidence has
suggested that the difference in pH and temperature optima between LDEs serves
important functions for the microbes to adapt to various environments in nature.
LiP, MnP, and VP have a wide range of applications beside biodegradation of
lignin. This is due to the capability of LDEs to oxidize a wide variety of phenolic substrates
66, 71, 80, 81. For example, LiP was shown to remove a nonylphenol from wastewater 66, while
a peroxidase expressed in B. subtilis can be used in dye-decolorizing operations 79. MnP
from Bjerkandera adusta was used to depolymerize chemically-pretreated coal, forming
aromatic compounds in the process 82. Also reported is a MnP from Ganoderma lucidum
for use in reducing both color and turbidity of apple and orange juices 83. The oxidation
potential of peroxidases allows for applications besides oxidizing phenolic substrates
(Table 1-1).
1.3.2 Laccases
First discovered in 1883 in an extract from the tree Rhus vernicifera, laccases are a
member of the superfamily of multicopper oxidases 84, 85. Laccases utilize four coppers to
reduce O2 to H2O and, in the process, oxidize the substrate 86, 87. This oxidative mechanism

12
stands in stark contrast to the mechanism of the peroxidase LDEs; the use of O2 in place of
H2O2 results in a milder chemical environment. This unique trait has led laccases to attract
interest in the lignin valorization community. The most commonly investigated laccases
are produced commercially from a polypore basidiomycete, T. versicolor (Figure 1-4). The
laccases secreted by T. versicolor have moderate degrees of stability at pH from 5 to 8 and
at temperatures from 10 to 45 °C 86, 88, 89.
Figure 1-4. Laccase from Trametes versicolor (PDBID: 1GYC) 86. Figure downloaded
from the Research Collaboratory for Structural Bioinformatics’ Protein Data Bank (RCSB
PDB) 75.
1.3.2.1 Thermophilic laccases
Laccases from thermophilic microorganisms have received particular interest for
use in lignin valorization and other applications because of the improved thermostability.
These laccases have been shown to oxidize a wide variety of phenolic substrates. Kiiskinen
et al. expressed a thermophilic laccase from Melanocarpus albomyces in Trichoderma
reesei. This laccase is capable of oxidizing 2,6-dimethoxyphenol (DMP), syringaldazine,
and guaiacol at temperatures up to 50 oC 90. Recently, An et al. expressed a thermostable
fungal laccase (PPLCC2) from Postia placenta in Pichia pastoris 85. PPLCC2 displayed

13
optimum oxidation of 2,2’-azino-bis(3-ethylbenzthioazoline-6-sulphonic acid) (ABTS)
and DMP at 60 ºC. While continuing to search for thermophilic laccases in fungi,
researchers have expanded their search to bacterial and archaeal laccases.
There have been several thermophilic laccases and laccase-like enzymes
discovered in bacteria in the past decades. B. subtilis was found to produce a thermostable
form of a laccase-like enzyme, CotA 91, which has maximum enzyme activity at 75 °C with
ABTS as substrate 92. Laccase from the bacterium Thermus thermophilus (TtL) is the most
thermostable laccase to date 93; optimal substrate oxidation was observed at 92 °C. A
recent work reported a thermostable laccase (LccA) isolated from the Dead Sea archaeon
Haloferax volcanii 94. It was found that LccA oxidized the phenolic substrates DMP and
syringaldazine along with the nonphenolic substrate ABTS. Remarkably, LccA retained
significant activity at 50 °C and in salt concentrations up to 1.4 M.
These laccases have been observed to be reliably thermophilic; however, they have
been observed to vary in pH optima. The pH optima of TtL and PPLCC2 change depending
upon which oxidation substrate is present. PPLCC2 exhibits optimal oxidation of ABTS
and DMP at pH 3.5 and 5.0, respectively 85. TtL exhibits optimal oxidation of ABTS and
syringaldazine at pH 4.5 and 5.5, respectively 93. The diverse temperature, pH, and
substrate affinity exhibited by laccases enable their use in a number of occasions.
1.3.2.2 Laccase applications
Laccases have a wide variety of applications with a few examples listed in Table 1-
1 95, 96. In the pulp and paper industry, laccases have been used to reduce the lignin content
of wood pulp 97. In the textile industry, laccases have been used to decolorize dyes and

14
treat a number of fabrics 98-101. Laccases can also be used to remove chemicals from
wastewater and contaminated soil 102-106. In food industry, laccase can be used in
processing fruit juice and treating wastewater from various food processes 95, 107-109. In
addition, laccases have found use in the pharmaceutical industry, producing compounds
that have various medical applications 110, 111. The diverse list of laccase applications
continues to expand with research into bioenergy.
The primary application of laccases for bioenergy is for biomass deconstruction.
Laccase is capable of removing lignin from the biomass, resulting in increased digestibility
of the cellulose and hemicellulose fractions 112. Laccase-mediator systems have been
reported to fractionate and modify lignin to produce quality carbon fibers 113; such a system
converts lignin to a high molecular weight fraction suitable for carbon fiber and a soluble
low molecular weight fraction suitable for biological conversion to lipids 114. Laccases
have also been used in microbial fuel cells (MFCs), in such cases laccases function as a
biocathode, providing a cheaper alternative to platinum 115. Immobilization of laccase on
carbon nanotubes further improves laccase activity in MFCs over time 116.
1.3.3 Other LDEs
In addition to the peroxidases and laccases, the search for LDEs led to the discovery
of novel enzyme consortia in bacteria. Several classes of the bacterial dye-decolorizing
peroxidases, multi-copper oxidases and β-etherases have been reviewed previously 16, 117,
118. The soil bacterium Sphingobium sp. SYK-6 is capable of growing on lignin-derived
monoaryls and biaryls substrates via expression of several LDEs as shown in Figure 1-5
119-121. SYK-6 has been shown to be extremely tolerant of high pH conditions, a desirable
characteristic for lignin valorization 14. Additionally, SYK-6 was found to prefer vanillin

15
when grown in the presence of several carbon sources; through further observations, it was
determined that ligninolysis is a mechanism required by SYK-6 for survival 14. The
vanillin catabolic pathway is responsible for providing SYK-6 with sufficient
concentrations of nicotinamide adenine dinucleotide (NADH) and nicotinamide adenine
dinucleotide phosphate (NADPH), important reducing species vital for microbial growth.
These unique lignolytic characteristics of SYK-6 have made it an attractive target for
designing lignin valorization strategies. The complete genome of SYK-6 has been
sequenced, which consists of a 4,199,332-bp-long chromosome and a 148,801-bp-long
plasmid 122.
Figure 1-5. Lignolytic pathways of Sphingobium sp. strain SYK-6 121.
The lignolytic capabilities of Sphingobium have been leveraged in plant
engineering for lignin valorization. A family of LDEs from Sphingobium, LigDFG, were

16
expressed in A. thaliana, resulting in high levels of oxidized G- and S-lignin subunits in
plants 123. Furthermore, E. coli was engineered by combining vanillin degradation
pathways (LigV, LigM) from SYK-6 and a protocheuate decarboxylase (aroY) and a
catechol dioxygenase (CatA) from tobacco plant 124. In a follow-up study, an
autoregulatory system was constructed into the engineered E. coli chassis and the new
strategy demonstrated bioconversion of the central intermediates such as vanillin into cis,
cis-muconic acid at high yield in E. coli 125. Such approaches highlight the opportunities
of developing novel applications of bacterial LDEs for lignin depolymerization.

17
Table 1-1. Applications of lignolytic enzymes
Enzyme Source Application References
Peroxidase B. subtilis Dye decolorization 79
MnP Bjerkandera adusta Coal treatment 82
MnP
Laccase
G. lucidum
Trametes hirsuta Food industry
83
107
95
108
Laccase T. versicolor Lignin removal from wood
pulp 97
Laccase
T. hirsute
Trametes villosa
Sclerotium rolfsii
T. versicolor
Dye decolorizing in textiles
98
99
100
101
Laccase
LiP
Trametes hirsute
Various lignolytic
microbes
T. versicolor
T. villosa
Lentimula edodes
Botrytis cinerea
Wastewater treatment
66
102, 103
104
105
106
109
Laccase
Myceliophtora
thermophyla
Trametes pubescens
Saccharomyces
cerevisiae
Production of pharmaceuticals 110
111
Laccase T. versicolor
T. hirusta Biocathodes
115
116

18
1.4 Ionic liquids
ILs are pairs of ions that exist in liquid form, often below 100 ℃ (also called room
temperature ILs). Given the enormous selections of cations and anions, a large number
(1018 theoretically) of ILs can be synthesized. For the same reason, ILs can be designed
with fine-tuned properties to suit the needs of a process in a highly specific manner. For
instance, by selecting environmentally friendly ion pairs, ILs can be synthesized to replace
environmentally harmful volatile organic solvents commonly used in industry 126, 127.
Many ILs have little to no vapor pressure, minimizing harmful emissions that are
associated with more common organic solvents 128. There have been, however, some
circumstances that challenge this notion 129. Many alkylimidazolium ILs were shown to
be flammable when exposed to a heat source 130. ILs have a wide range of applications,
including biomass processing, CO2 capture, compound extraction, thermostable lubricant,
reaction medium/catalyst for catalysis, pharmaceuticals, etc. which can be seen from
several review articles 126, 131, 132. The foci of this section are to review the properties of
ILs that are relevant to lignin fractionation and enzyme activity/stability and the effect of
IL treatment on lignin characteristics.
1.4.1 IL properties
Solvent properties, hydrophobicity, and biocompatibility/toxicity are important
properties of ILs that determine their potential applications. Those properties are directly
influenced by the cation and anion species. In other words, it is possible to tune the IL
properties by changing the anion/cation combinations 133. The Kamlet-Taft system is
commonly used to describe IL’s solvent properties in three separate terms/parameters:
polarizability (π*), hydrogen bond donator capacity (α) and hydrogen bond acceptor

19
capacity (β) 134. Kamlet-Taft parameters were not only used for measuring solvent
properties of a single IL but also applied to describing the average or bulk solvent
properties of binary and ternary IL/H2O mixtures 135, 136. Primarily determined by the
anion, the β value has been used to predict ILs’ capacity to disrupt cellulose 137, 138. As
proposed by Doherty et al., higher β values indicate that ILs tend to form strong attractions
with the hydroxyl protons of cellulose; this interaction leads to disruption of the crystalline
lattice 139. More computational-based evidence was reported by Sun and co-workers
linking the predicted interaction energies with experimentally determined Kamlet-Taft
parameters140. A positive correlation was observed between cellulose conversion and β
values. ILs with larger differences between β and α, net basicity (β-α), tend to dissolve
cellulose more efficiently141, 142.
The hydrophobicity of IL is relevant to enzyme stability/activity in IL; some ILs
have been observed to deactivate enzymes with decreasing hydrophobicity. When a series
of alkylimidazolium ILs were screened for hydrophobicity, the alkyl chain length of the
cation was well correlated with the hydrophobicity of the ILs 143. In addition,
hydrophobicity of the anion also contributes largely to the IL’s hydrophobicity. For
example, ILs with a bis(trifluoromethylsulfonyl)amide ([NTf2]-) anion have been used to
stabilize lithium electrochemistry due to the anion’s hydrophobic nature 144-147. It has been
reported that alkylimidazolium ILs with alkyl sulfate anions, with decreasing anion chain
length, resulted in higher T. versicolor laccase deactivation 148, 149. Another predictor of
the interaction between an IL and an enzyme is the Hofmeister series (aka. lyotropic series),
a classification of ions in order of their ability to salt out or salt in proteins. It has been
shown that kosmotropic anions and chaotropic cations are preferable for enzyme stability,

20
with the anion’s properties largely determining enzyme-IL interactions 33, 150. Molecular
dynamics simulations of a lipase in the ILs 1-butyl-3-methylimidazolium
hexafluorophosphate ([C4C1Im][PF6]) and 1-butyl-3-methylimidazolium nitrate
([C4C1Im][NO3]) showed that IL-enzyme interactions are dominated by the anion 151.
Collection of these properties leads to a more generic definition of IL’s
biocompatibility/toxicity.
Quantitative structure–property relationship (QSPR) modeling is a powerful tool
predicting IL toxicity. Results from Rank et al. show strong correlation between
lipophilicity of the cations and IL cytotoxicity by comparing experimental with
computational data over a set of 74 ILs with imidazolium, pyrrolidinium, pyridinium,
quinolinium, quaternary phosphonium, and quaternary ammonium cations and the small
anions such as Cl-, Br-, BF4- or PF6- 152. Results from the same group suggest that anion’s
lipophilicity and/or vulnerability to hydrolytic cleavage also appeared to be a crucial
structural feature leading to cytotoxicity 153. Using a set of electronic, spatial, structural,
thermodynamic, and topological descriptors, Couling et al. constructed QSPR models to
predict IL toxicity; results show an increasing trend of toxicity based on cation type of
ammonium, pyridinium, imidazolium, triazolium, tetrazolium and decreasing trend of
toxicity with increasing number of negatively charged atoms and ring methylation on the
cation 154. A more recent work from Yoo et al. 155, adopted an integrated experimental and
computational approach to explore the molecular-level details of IL’s cytotoxicity.
Molecular dynamics (MD) simulations based on 1-palmiltoyl-2-oleoyl-
phosphatidylcholine lipid bilayer suggest that cytotoxicity increases with increasing alkyl
chain length of the cation, due to the ability of the longer alkyl chain to insert in and disrupt

21
the cell membrane 155. Collectively, it is concluded that both the cation and anion play
important roles in determining IL’s toxicity/biocompatibility through their interactions
with biomolecules.
1.4.2 Biomass pretreatment with ILs
The ability of certain ILs to solubilize one or all three major components of
lignocellulose has made them attractive solvents for biomass pretreatment 156. This
capability is largely attributed to IL’s ability to form strong hydrogen bonds with the
cellulose, hemicellulose, and lignin polymers present in biomass 157, 158. A computational
approach was used to predict the ability of 750 ILs to solubilize cellulose and lignin using
p-coumaryl alcohol as a lignin model compound; it was predicted that a very wide range
of anion/cation combinations can form ILs capable of solubilizing lignin 159. The factors
affecting IL pretreatment performance and challenges facing this pretreatment technology
are discussed below.
Figure 1-6. a) [C2C1Im][OAc] has served as the benchmark IL for biomass pretreatment
for several years. Recently, bioderived ILs like b) choline lysinate ([Ch][Lys]) and c)
diethylamine hydrogen sulfate ([DEA][HSO4]) have attracted interest due to their low cost.
Effect of cation/anion on pretreatment
The properties of ILs are sensitive to the cation and anion species present in the IL.
As a result, these species can significantly alter the products obtained from biomass after

22
IL pretreatment. The ability of ILs to dissolve cellulose is dependent upon the anion and
cation species. ILs with the 1-butyl-3-methylimidazolium ([C4C1Im]) cation are capable
of dissolving cellulose when paired with a chloride anion ([Cl]) 160. When the same cation
is paired with [NTf2] anion, the IL can no longer dissolve cellulose. Furthermore, when
[Cl] is paired with a pyrrolidinium or piperidinium cation, the IL can no longer dissolve
cellulose 161. The ability of choline amino acid ([Ch][AA]) ILs to solubilize the
components of biomass have been shown to be sensitive to the choice of amino acid. Liu
et al. screened a series of [Ch][AA] ILs for lignin, cellulose, and hemicellulose solubility
and found hemicellulose solubility was heavily dependent upon the amino acid anions 162.
When pretreating wheat straw with dilute [Ch][AA] ILs, it was found that glutamic acid
significantly reduced lignin removal during pretreatment 163. In a follow up study, Hou et
al. 164 established a positive correlation between the pKa of the anion and the
delignification observed during pretreatment.
Alkylimidazolium ILs have served as benchmark solvents for pretreatment of
lignocellulosic biomass (Figure 1-6). These ILs have been shown to improve cellulose and
hemicellulose digestibility after pretreatment, via lignin removal and reduction or change
in the crystallinity of cellulose. Enzymatic hydrolysis sugar yields can be greatly improved
after pretreating a variety of biomass feedstocks with alkylimidazolium ILs 22, 165. In one
example, it is shown that [C2C1Im][OAc] reduces the crystallinity of the cellulose in wood
flour and removed 40% of lignin, leading to over 90% glucose yield from enzymatic
hydrolysis of the pretreated biomass 166. Despite their effectiveness in pretreatment
strategies, the high cost of alkyl imidazolium ILs has hindered their widespread adoption
in biorefineries.

23
Figure 1-7. Butylimidazolium ILs. These ILs have all been effectively utilized in biomass
pretreatment strategies with different effects on downstream processing of biomass
(adapted from 22).
Cost reduction strategies to enable IL pretreatment
Three primary avenues for IL cost reduction are being explored: the use of
inexpensive feedstocks for IL synthesis, use of aqueous ILs, and reusing or recovering ILs
for repeating pretreatment cycles 23.
a. Inexpensive feedstock for IL synthesis
There are a number of low-cost precursors that can be used for IL synthesis. To
serve as viable replacements, however, these new ILs must match the pretreatment efficacy
of alkylimidazolium ILs. Cations and anions derived from renewable resources such as
protein, sugars, and biomass-derived carboxylic acids are of particular interest. George et
al. synthesized low-cost ILs from ammonium cations and the [HSO4]- anion and tested
their effectiveness in pretreating switchgrass; results show that sugar yields and lignin
removal using the low-cost ILs were comparable to 80% [C2C1Im][OAc] 167. ILs
synthesized from choline chloride and bio-derived amino acids have been shown to be
highly effective in solubilizing hemicellulose and lignin without solubilizing cellulose 168.
Eight choline amino acid ([Ch][AA]) ILs were screened for their ability to remove lignin
during pretreatment of rice straw. The majority of the ILs were shown to remove over 50%

24
of lignin during pretreatment, with [Ch][Lys] removing 60.4% of lignin during
pretreatment 169. These new ILs are potentially more biocompatible and cheaper compared
to petroleum based alkylimidazolium ILs 170. In addition to lowering the cost of IL itself,
reducing the dosage of IL by using IL-water mixtures (aqueous IL) will further lower the
chemical cost of pretreatment.
b. Aqueous ILs
Instead of using pure, or “neat” ILs, using IL-water mixtures reduces the amount
of IL needed to achieve a desirable pretreatment outcome, thus reducing the overall cost.
Elucidating the role of water on the IL pretreatment efficiency helps to design a better
pretreatment process using aqueous IL. In a previous study, the effect of water on IL
pretreatment was investigated by pretreating switchgrass with a series of [C2C1Im][OAc]-
water mixtures. It was found that pretreatment with dilute [C2C1Im][OAc] at 50-80% IL
concentration led to comparable glucose yields and lignin removal to those obtained with
100% [C2C1Im][OAc] 171. Pretreatment with dilute [Ch][AA] ILs was also shown to
improve sugar yields and lignin removal when compared to pretreatment using pure ILs
163. These findings provide insights into the interplay of water as a co- and anti-solvent in
aqueous IL system. Using aqueous IL as pretreatment agents offers additional benefits
including reduced viscosity of the IL medium, less gel formation, and lower energy
requirements and costs associated with IL recycling 171, 172.
c. IL reuse and lignin recovery
Recovery and reuse of ILs following pretreatment can reduce the demand for
purchasing or synthesizing nascent ILs. Oftentimes these ILs can be reused several times

25
without a significant reduction in pretreatment efficacy. For instance, reuse of
[C2C1Im][OAc] for 5 times was possible without observing a significant change in the
pretreatment maple wood flour 166. [Ch][Lys] can be recovered and reused for pretreatment
of rice straw 5 times until a significant reduction in pretreatment efficacy was observed 169.
Separation technologies such as evaporation/distillation, electrodialysis, reverse osmosis,
and pervaporation have been investigated for concentrating ILs for reuse 173-175. However,
sugars and lignin need to be separated from the IL before sending the IL back for the next
batch; the cost associated with this step can be prohibitive.
The use of IL-based solvents for biomass dissolution and lignin depolymerization
is relatively new and much needs to be learned before an industrial scale process can be
implemented. System complexities associated with IL recycling, biomass-solute
separation and downstream processing remain the key challenges in the commercialization
176. Recent studies have identified some key areas that influence the overall process
economics of IL-based biorefinery processes 175, 177, 178. These areas include the use of low-
cost IL at high solid loading in the reaction, development of biocompatible ILs or low price
IL-tolerant enzyme mixtures, development of efficient IL recycle and product recovery
technologies, and system intensification and optimization. In-situ lignin valorization is one
of the possible ways to lower the product recovery cost and in turn improve the overall
economics.
1.4.3 Effect of IL treatment on lignin characteristics
The structure of lignin undergoes numerous changes during IL pretreatment.
Molecular weight distribution can be quantified with gel permeation chromatography
(GPC), while linkage frequency can be quantified with Heteronuclear Single Quantum

26
Coherence Nuclear Magnetic Resonance (HSQC NMR). Changes in lignin structure as a
result of pretreatment with [C2C1Im][OAc] were investigated at different treatment
temperatures and times. Changes in β-O-4’, β-5’, and β-β’ linkage frequencies were
observed, along with a reduction in the molecular weight distribution 179. GPC analysis of
lignin extracted from switchgrass with dilute [Ch][Lys] found lignin in the liquid stream
had a lower molecular weight than lignin found in the liquid stream after dilute acid and
ammonium hydrolysis pretreatment 21. GPC and HSQC NMR provide helpful information
regarding changes to the lignin structure. Due to the heterogeneous nature of lignin, it is
challenging to elucidate lignin depolymerization mechanisms in ILs. β-O-4’ linked lignin
model compounds have been used to elucidate probable depolymerization mechanisms of
methylimidazolium ILs 180. Model lignin compounds enable researchers to better
investigate the effects of IL treatment on common aryl ether or carbon-carbon linkages
found in real lignin 25.
1.4.4 Rationale for pairing LDE and IL for lignin depolymerization
Taken together, given the unique properties of certain ILs that provide for lignin
solubility and biocompatibility to enzymes and microbes, there is a great opportunity to
develop a new strategy for lignin extraction, depolymerization, and upgrading via
biocatalysis in ILs. Instead of using harsh thermochemical processes, the relatively mild
biochemical process facilitates the selective deconstruction of lignin and retains the
functionality of the oligomeric lignols, thus helping to channel desirable aromatic products.
Despite previous exploration of catalysis and enzyme activity in ILs, there is a gap in our
fundamental understanding of the mechanisms determining the interactions of lignolytic
enzyme and these solvents. Further exploration is needed into how the solvent properties

27
(acidity, basicity and polarity) as determined by the cation and anion pairs impact the lignin
solubility and enzyme activity in aqueous IL systems. Furthermore, in order to enable
enzymatic reactions in IL, two aspects need further investigation: the development of
biocompatible ILs with improved lignin solubility, and the selection of IL tolerant
enzymes. These developments rely on a better understanding of the enzyme-IL
interactions, such as the structural changes, surface charges, inhibition kinetics, enzyme
stability and the structure-function relationship of enzyme in IL media. Based on that, the
LDE-IL system can be tuned to improve the product yield and selectivity.
1.5 Enzyme-IL interactions
1.5.1 Enzyme activity and stability in ILs
Certain ILs, as part of their highly tunable nature, can provide an environment
suitable for enzymatic activity. Cellulase 181, laccase 150, 182, 183, lipase 184, 185, α-
chymotrypsin 185-187, tyrosinase 185, and lysozyme 188 activity have all been tested in the
presence of IL (Table 1-2). Numerous studies have attempted to elucidate the IL
characteristics affecting enzyme’s activity and stability in IL. Efforts have also been made
to identify enzymes with improved resistance to enzymatic inhibition caused by IL.

28
Table 1-2. Studies of enzyme activity and stability in ILs
Enzyme Source Ionic Liquid Reference
Laccase T. versicolor Alkyl imidazolium
182
183
Laccase T. versicolor 56 ILs 189
Cellulase T. reesei [C2C1Im][OAc] 190
β-Glucosidase A. niger [C2C1Im][OAc] 190
Laccase T. versicolor [Ch][H2PO4] 189
Laccase Aspergillus (DeniLite
Base) [C2C1Im][MDEGSO4] 191
Lysozyme Chicken egg white
Ethanolammonium formate
Ethylammonium nitrate
Bis(2-methoxyethyl)ammonium acetate
N,N-dimethylethanolammonium glycolate
[Ch][H2PO4]
188
Laccase T. versicolor
[C2C1Im][EtSO4]
[C2C1Im][OAc]
183
Laccase T. versicolor [Ch][H2PO4] 189
α-Chymotrypsin Bovine pancreas type
II
Triethylammonium acetate
Triethylammonium phosphate
1-benzyl-3-methylimidazolium chloride [C6C1Im][Cl]
1-benzyl-3-methylimidazolium tetrafluoroborate
[C6C1Im][BF4]
Tetra-butylphosphonium bromide
186
RNase A N/A
[Ch][H2PO4]
[C2C1Im][DCA]
192
1.5.1.1 Enzyme activity in ILs
A wide variety of factors are thought to play a role in enzyme activity in ILs. There
is a clear trend that persists among studies performed: the higher the IL concentration, the
more severe the inhibition observed. Laccase from T. versicolor was screened for activity

29
and stability in a series of methylimidazolium ILs 193. Increasing concentrations of ILs
resulted in increased levels of enzymatic inhibition; while 1-ethyl-3-methylimidazolum
ethylsulfate ([C2C1Im][EtSO4]) caused less inhibition to laccase activity as compared to
[C4C1Im][Cl]. Similarly, as shown in another study, T. versicolor laccase activity
decreased with increasing IL concentration of the two tested alkylimidazolium ILs,
[C2C1Im][EtSO4] and [C2C1Im][OAc] 183. This trend was not always true as improved
laccase activity has been observed in some ILs. Galai et al. screened T. versicolor laccase
activity in 56 ILs and found that 13 of the ILs improved laccase activity; the most notable
improvement was seen in choline dihydrogenphosphate ([Ch][H2PO4]) which increased
laccase activity 451% 150. In addition to the enzyme activity, which is determined by short-
term inhibitory effect, the long-term effect of IL on enzyme, in other words, the enzyme
stability in IL, is equally important.
1.5.1.2 Enzyme stability in ILs
ILs have been shown to stabilize enzymatic activity over varying periods of time.
The long-term effects vary with respect to the IL-enzyme pair and IL concentration. Some
ILs, like [Ch][H2PO4], continue to positively impact enzymatic activity over time 150, 188.
However, the long-term effects of several alkylimidazolium ILs stands in contrast to their
short-term effects. T. versicolor laccase in [C4C1Im][Cl], [C2C1Im][OAc], 1-ethyl-3-
methylimidazolium MDEGSO4 ([C2C1Im][MDEGSO4]), and [C2C1Im][EtSO4] exhibited
improved stability over varying periods of time, however, all ILs caused inhibition to
enzyme activity in short term 193.
Certain ILs have also been shown to improve enzymatic resistance to inhibition
caused by pH and thermal deactivation. For example, T. versicolor laccase in 10 mM

30
[Ch][H2PO4] retained activity up to pH 12, compared to pH 9 without IL 150. Short alkyl
chain ammonium ILs, triethyl ammonium acetate ([TEA][OAc]) and triethyl ammonium
phosphate ([TEA][PO4]) were shown to improve the thermal stability of α-chymotrypsin
when compared to enzyme activity in 0.05 M pH 8.2 Tris-HCl buffer 186. Not surprisingly,
the ability of ILs to stabilize enzymes is sensitive to the cation and anion present; the
thermostability of RNase A was improved in [Ch][H2PO4] whereas choline chloride
([Ch][Cl]) did little to enhance thermostability 192. This anion-specific effect can be
partially explained by the Hofmeister series: H2PO4- has been observed to be less
chaotropic than Cl- when paired with the same cation 37, 148, 194. In spite of the numerous
studies of IL-enzyme interactions, the exact mechanisms by which ILs affect enzyme
stability are still largely uncertain.
1.5.2 Effect of IL on enzyme
Investigating the changes of enzyme structure provides important information
about enzyme stability in different ILs with varying properties. The structure of an enzyme
is critical for its function, whereas enzyme-IL interactions may cause structural changes of
the enzyme that further impact its activity and stability. The enzyme-IL interactions are
dependent on IL properties and the enzyme sources. Thus, an in-depth understanding of
enzyme structure and enzyme-IL interactions is critical for designing future biocatalytic
systems in ILs.
1.5.2.1 Enzyme structure
Circular Dichroism (CD) spectroscopy can help understand changes made to
enzyme structure in ILs by highlighting secondary structures of the enzyme. CD

31
spectroscopy of T. versicolor in [Ch][H2PO4] revealed that addition of IL resulted in a
conformation change around the active site, along with a shift in the ratio of α/β structures
150. Enzyme crystallization in IL and resolving the structure could provide a more
comprehensive view of structural changes; this approach will be discussed later in the
review.
1.5.2.2 IL properties affecting enzymatic activity
There have been a myriad of in-depth reviews discussing the different IL properties
responsible for enzymatic inhibition and stability 29-37. Each of the reviews contains tables
summarizing studies examining different IL properties and their effect on enzymatic
activity and stability. Polarity, alkyl chain length of the cation, hydrophobicity, anion, and
viscosity are common factors discussed in each of the reviews. Increased cation chain
length, increased anion size, and decreased polarity (a consequence of chain length and
anion size) were all factors associated with reduced α-chymotrypsin activity 184.
Thermostability of α-chymotrypsin in ILs revealed that ILs with imidazolium and
phosphonium-based cations were weak stabilizers, while ILs with ammonium-based
cations were strong stabilizers 186.
Increasing cation hydrophobicity and decreasing anion nucleophilicity has been
shown to increase lipase activity 195. Two enzymes, a lipase and a tyrosinase, were shown
to be biocompatible with aqueous IL made of chaotropic cations and kosmotropic anions
196. Results indicate that in general ILs with choline or ammonium cations activated lipase
while imidazolium ILs deactivated the enzyme. More specifically, ammonium ILs
composed of chaotropic cations (favorably with H-bonding capability) and kosmotropic
anions are favored for enzyme catalysis 196. The interactions between a lipase and several

32
ILs were simulated with molecular dynamics (MD) simulations, revealing dominant
Coulombic and van der Waals interactions 29. The same simulations also showed some
[C4C1Im] cations diffused into the active site, implicating competitive inhibition. There is
no consensus as to which, if any, how IL property dominates the impact on enzyme activity
and stability. Regardless, there have been numerous efforts focused on improving enzyme
activity in ILs.
1.5.3 Methods for improving enzyme stability in ILs
In an effort to combat the inhibitory effects of ILs on enzymatic activity, a number
of methods have been explored. Most predominant of these approaches are immobilization
of the enzyme on a substrate, screening enzymes from thermophilic and halotolerant
organisms, and engineering the enzyme surface residues. Each of these methods has been
successfully demonstrated with different enzymes.
1.5.3.1 Immobilization
Enzymes can be attached to an inert, insoluble material via different mechanisms
such as adsorption, entrapment, cross-linking, etc. Enzyme immobilization has proven to
be an effective method of improving enzyme activity and/or stability in less than optimal
conditions 197. Furthermore, immobilized enzymes have better recyclability than free
enzymes. Immobilization strategies have been successfully employed to improve enzyme
activity in ILs. In one study, immobilization of lipase on acrylic resins improved yields of
biodiesel in 1-butyl-3-methylimidazolium hexafluorophosphate ([C4C1Im][PF6]) 185.
Laccase from Myceliophthora thermophila was immobilized on glyoxyl-agarose beads,
improving thermostability in [C2C1Im][EtSO4] at 70 °C 198. Several other enzyme

33
immobilization strategies (on solid support, sol–gel, and cross-linked enzyme aggregate,
CLEA), have been reviewed previously and will not be discussed here 30.
1.5.3.2 Thermophilic and halotolerant enzymes
Thermotolerant and halophilic organisms have evolved to produce enzymes with
resistance to harsh environments. This selective pressure from the environment has
enabled these enzymes to potentially resist the inhibitory effects of ILs. Cellulases from a
hyperthermophilic bacterium and archaeon retained activity in two-fold higher
[C2C1Im][OAc] concentration than their mesophilic counterparts 199. In a follow-up study,
Zhang et al. identified a halophilic cellulase from a halophilic archaeon and found it can
tolerate relatively high concentrations (20% w/w) of several alkylimdazolium ILs 200. The
halophilic and IL-tolerant cellulase identified by Zhang et al. was found to have a large
number of negatively-charged surface residues 200. Similarly, cellulases from the
halophilic Aspergillus terreus strains displayed improved activity and stability in high salt
conditions and a series of alkylimidazolium ILs, most notably [C2C1Im][OAc] 201.
Although, limited number of structural evidences exist to explain what causes the improved
resistance of those enzymes to IL, it’s possible that the thermophilic cellulases evolved to
have more negatively-charged surface residues than the mesophilic cellulase. Several
studies have suggested that reducing the number of positively-charged surface residues and
increasing the number of negatively-charged surface residues improve enzyme resistance
to ILs 202, 203. The surface characteristics enabling enzyme tolerance to IL have inspired
artificial ways to modify enzyme surfaces.
1.5.3.3 Charge engineering

34
Altering the charged residues on the surface of the enzyme could affect the ability
of the ILs to bind to the enzyme (Figure 1-7). This reduction in binding affinity would
reduce IL inhibition. Reducing the amine:acid ratio of surface residues has been shown to
improve activity of several enzymes in [C4C1Im][Cl] 202. Later molecular dynamics
simulations of modified lipase and α-chymotrypsin showed reduced anion binding and
increased cation binding on the enzyme surface 203. Surface modification of enzyme by
introducing protein–polymer surfactant nanoconstructs was shown to greatly improve
enzyme solubility in hydrophilic and hydrophobic ILs 204. Applying this method, Brogan
et al. 205 recently demonstrated that chemically modified glucosidase was thermally stable
at up to 137 °C and 30 times more active towards cellulose in [C2C1Im][EtSO4] as
compared to reactions in aqueous media due to solvent-induced substrate promiscuity of
glucosidase.

35
Figure 1-8. Effect of surface charge modification on IL interaction with enzymes
(Nordwald, 2015).
1.6 Protein engineering to improve IL compatibility
1.6.1 Structure-function relationship of proteins
The structural hierarchy of proteins can be broken down into 4 levels: primary,
secondary, tertiary, and quaternary. The primary structure of a protein is the sequence of
amino acids. The secondary structure captures how those amino acids are folded into α-
helices, β-sheets, or loops. The tertiary structure captures how the secondary structures
fold together to form the final protein structure; oftentimes this is accomplished by an
intricate network of hydrogen bonds, salt bridges, and disulfide bonds 206, 207. The
quaternary structure is defined by tertiary structures forming higher-order structures such
as dimers or trimers. Each of these structural levels plays a key role in the function of
proteins.

36
1.6.1.1 Primary structure
Directed mutagenesis of key residues in a variety of enzymes has implicated the
ubiquitous role primary structure plays in protein function. These residues have been
shown to be responsible for a variety of key functions of the protein.
Substrate recognition and binding
In many proteins, key amino acids have been implicated in the binding of the
substrate. Mutations of 2 residues near the heme access channel, H82A and Q222A, and
2 residues near the surface of the protein, W171A and F267L, revealed the importance of
each residue for LiP substrate binding and function 208. Mutation of Asp376 to Asn, Glu,
or Thr at the catalytic phosphorylation site of the α subunit leads to loss of ATPase activity
209. It was found that leptin binding to the leptin receptor of a variety of species is highly
specific to the residues present on both the receptor and leptin 210. Mutations of residues
near the surface of Type III antifreeze protein (AFP) resulted in loss of activity 211.
Oftentimes, the residues responsible for enzymatic activity have a net charge at
physiological pH.
Altering the primary structure of an inactive enzyme can alter the enzyme activity
and substrate affinity. A heme peroxidase from Coprinus cinereus was unable to catalyze
the oxidation of veratryl oxidase, a common substrate used for screening lignolytic
enzymes. The inactive heme peroxidase has a structure similar to LiP, so a number of
mutations were made using the active LiP as a template 212. A Trp residue (Trp179) was
introduced to interact with veratryl alcohol, while D179W, R258E, and R272D mutations
created an environment mimicking that found around the catalytic Trp residue in the active

37
LiP 212. All of these mutations enabled the previously inactive heme peroxidase to oxidize
veratryl alcohol. This approach allows identification of primary structure that are crucial
for proper enzyme function.
Charged residues
Charged residues on the surface of proteins can be implicated when binding to a
charged substrate. For instance, alanine mutations of a series of positively-charged Lys
and Arg residues resulted in decreased activity in proliferating cell nuclear antigen (PCNA)
213. Key mutations of charged residues of TRPV1 and TRPV4 dramatically reduce the
permeability of calcium and magnesium ions 214. TRPC1 and TRPC5 channel permeation
properties have been shown to be highly dependent upon negatively charged residues close
to the pore region 214. As discussed in section 5.3, modification to the charged residues on
surface of the enzyme has proven effective to reduce IL inhibition by altering IL binding
affinity to enzymes.
1.6.1.2 Secondary structure
As amino acids begin interacting with each other, they start forming higher-order
structures. The presence of these secondary structures can impact the function and stability
of a protein. T. versicolor laccase experienced a shift in the ratio of α-helices and β-sheets
in the presence of [Ch][H2PO4], likely due to anion-peptide interactions, though the exact
reason why this shift occurred was not confirmed 150. The β-sheet near the C-terminus of
a Type III antifreeze protein is thought to be responsible for antifreeze activity 211. The
secondary structure of a laccase is related to its thermostability. For instance, bacterial
laccase has a coiled section connecting 2 domains, whereas plant and fungal laccases do

38
not; for that reason, among many others, bacterial laccases are typically more thermostable
than fungal and plant laccases 206, 207.
1.6.1.3 Tertiary and quaternary structure
Tertiary and quaternary structure have been shown to affect a wide variety of
protein functions. Changes to the tertiary and quaternary structure of the Tobacco Mosaic
Virus Coat Protein have been shown to significantly affect the immune system response of
the host 215. An N-terminus domain of a protein from Bacillus stearothermophilus is
responsible for anchoring the protein to the cell wall of the organism 216. The structure of
G-protein coupled receptors (GPCRs) is critical in determining their conformation and, as
a result, their function 217. When a GPCR receptor is unbound, it adopts an inactive
conformation; once it is bound by an agonist, the structure of the GPCR receptor changes
to an active form 217. Although few studies have examined IL effects on tertiary structure,
it is reported that the tertiary structure of the immunoglobulin binding domain of
streptococcal protein B disrupted in high concentrations (60% v/v) of [C4C1Im][Br] using
high-resolution magic-angle-spinning NMR spectroscopy 218. Knowledge of IL’s impact
on protein structure provides vital information for protein engineering strategies to improve
enzymes’ IL tolerance.
1.6.2 Protein engineering for improved function
To improve function of proteins under environmental stresses (high pH, high
temperature, etc.), key residues can be identified and mutated. The library obtained from
screening mutations of residues grows at an enormous rate as more sites are mutated, a
consequence of the number of possible amino acids at each position (20n, n = number of

39
sites mutated). When screening mutations at 4 sites, the library will contain 204 = 160,000
entries. Addition of just 1 more site mutation increases the size of the library to over 3
million entries. It’s for this reason that most mutation studies are limited to just a few sites.
Despite this limitation, impressive results have been obtained by mutations of just a few
sites on many proteins. Two protein engineering approaches are commonly used for
improving protein function: directed evolution and rational design 219. The ultimate goal
of both approaches is the same, which is a protein with improved stability in a harsh
environment. What sets the approaches apart is the size of the generated library of mutants.
Directed evolution usually generates a much larger library of mutants than rational design.
1.6.2.1 Directed evolution
Directed evolution mimics the environmental stresses to an organism which is
performed in lab using methods like error-prone PCR for diversity generation (random
mutagenesis) and screenings in iterative cycles 220. The organisms producing the best
performing enzymes are subjected to another round of, and in most cases more severe,
environmental stress. The screening procedure is repeated until a desired optimal protein
is obtained. Throughout the screenings the proteins are sequenced and key residues that
improve performance are identified. In one example, Martin et al. generated a
thermostable mutant of a mesophilic cold shock protein through the use of directed
evolution 221. Through application of high heat and high salt stresses, a mutant was
produced that outperformed a thermophilic cold shock protein. By sequencing the mutants,
key residues responsible for improving the thermostability of the protein can be identified.
A thermophilic fungal laccase from M. thermophila was subjected to repeated rounds of
directed evolution in organic solvents, in the process developing an affinity for higher pH

40
environments 106. Other directed evolution strategies involving laccases are primarily
focused on improving expression in non-native organisms, i.e. fungal laccase expression
in yeast 222.
1.6.2.2 Rational design
Rational design requires the knowledge of the residues thought to be responsible
for improving a protein’s tolerance to environmental stresses. These residues are often
identified through the use of a homolog. The crystal structure of MnP from C.
subvermispora was used as a template for directed mutagenesis of a P. ostreatus versatile
peroxidase (VP), producing 2 variants with improved resistance to a low pH environment
223. Identifying residues in ancestral forms of the enzyme, a triple mutant
(H239F/T240L/I241L) LiP from P. chrysosporium was engineered for improved activity
at high temperatures and stability in the presence of H2O2 80. Threonine residues
surrounding the type I copper in B. subtilis CotA, T415 and T418, were the focus of a
rational design strategy by Khodakarami et al. 224 with the goal of improving activity and
substrate selectivity. The T415I mutation improved activity with ABTS 4-fold, whereas
selectivity towards ABTS was 11 times higher than syringaldazine in the same mutant 224.
A fungal laccase from T. versicolor exhibited improved resistance to the IL
[C2C1Im][EtSO4] when mutations were made to loop residues connecting domains 2 and 3
225. While rational design can be a powerful tool for improving enzyme function in stressful
environments, it can be limited by lack of knowledge about the protein’s sequence and
structure.

41
1.6.3 Protein crystallization in ILs
Crystallizing proteins in ILs could provide more detailed insight into the binding
locations of ILs on the surface of proteins. Crystallization of proteins in ILs has been
shown to, in some cases, improve crystal quality, size, and number. When trypsin and
lysozyme were crystallized in ILs, crystal number and quality were improved; resolution
in [C4C1Im][BF4] was improved from 2.8 Å to 2.1 Å 226. Addition of ILs resulted in
formation of larger lysozyme crystals 188. When trypsin was crystallized in ILs, some
fragments of the ILs were found to have been incorporated as part of the protein structure
227. Numerous studies have highlighted the benefits of using ILs for protein crystallization,
though none have identified IL binding sites on the protein’s surface. Docking simulations
can be used to identify IL binding sites, as seen in the 2016 study by Anbarasan et al. with
a thermophilic xylanase in biomass-dissolving ILs 228. To get a clearer picture, however,
crystallization of proteins in ILs is critical for elucidating the effects of enzyme-IL
interactions.
1.7 Product selectivity
Due to the complex nature of lignin, controlling product formation from lignin
depolymerization proves difficult. A biocatalysis system coupling the LDEs and ILs opens
opportunities in directing product formation from lignin degradation. The selectivity is
rooted in both the tunable properties of ILs and the specificity of LDEs. Additionally, use
of a mediator alongside LDEs (e.g. laccase) can increase the oxidation window thus
extending the suite of products obtained from lignin. Use of capping agents has also been
shown to prevent condensation of lignin depolymerization products.

42
1.7.1 Controlling product formation via biocatalyst selectivity
The subject of directing formation of products from lignin has already been
explored through the use of a number of base, acid, and metal catalysts for catalytically
depolymerizing lignin in IL media 10, 24-28. The use of a biocatalyst, however, could
potentially offer improved selectivity at less severe reaction conditions. Laccase uses O2
as the final electron acceptor in its oxidative mechanism, and there is no need for harsh
temperature/pressure and costly catalysts. In addition, a number of ways can be used to
tune the biocatalyst for different products. One way is via the use of a mediator such as 1-
hydroxybenzotriazole (HBT) and ABTS. Introduction of a mediator helps increase the
oxidative potential of laccase, thus increasing the number of products obtained by enabling
laccase to oxidize non-phenolic compounds 229, 230. Furthermore, the source of enzymes
could potentially affect the products obtained from lignin as a consequence of the oxidative
mechanisms. For instance, white rot fungi typically utilize several oxidative enzymes, such
as LiP, MnP, and laccases 17 while brown rot fungi utilize a Fenton redox reaction when
degrading lignin 231, 232. Recent discoveries of new classes of LDEs that selectively cleave
C-C bonds, hold great promise since this capacity is unrivalled by common metal catalysts
233, 234.
1.7.2 Controlling product formation via IL selectivity
ILs are capable of directly influencing the products obtained from lignin
degradation. It has been demonstrated that acidic ILs are capable of cleaving the β-O-4’
linkage 180. Additionally, Dutta et al. 235 demonstrated that product formation from lignin
degradation was a function of the IL used and the treatment temperature. At high

43
temperatures [Ch][Lys], [TEA][HSO4], and [C2C1Im][OAc] were shown to favor
formation of guaiacol, guaiacylacetone, and both guaiacol and guaiacylacetone,
respectively. This product selectivity was only observed at high temperatures and high IL
concentrations.
Several ILs have been shown to be capable of shifting product formation of
biocatalysts. Increasing concentration of ILs was correlated with an increase in the
monophenolase:diphenolase activity ratio of a tyrosinase 236. A protic IL,
triethylammonium mesylate increased the selectivity of lipase towards omega-3 fatty acids
237. Product selectivity of a sialidase was shown to be highly sensitive to the amount and
the type of IL present in the system 238. Pairing the product selectivity offered by both the
LDEs and ILs could potentially lead to more effective lignin depolymerization pathways.
1.7.3 Preventing lignin repolymerization and improving yield
One of the primary issues encountered when dealing with lignin decomposition,
along with directing product formation, is that of lignin repolymerization. The
intermediates formed during lignin decomposition are susceptible to repolymerizing rather
than forming monomeric, dimeric, or oligomeric products. Under acid and/or high
temperatures conditions commonly used for lignin extraction, stable carbon-carbon (C–C)
bonds are formed once the lignin ether bonds are cleaved. This leads to severe and
irreversible condensation that dramatically affects further upgrading of lignin. When
examining depolymerization of organosolv lignin with base catalysts, it also was found
that lignin repolymerization occurred; S-lignin was the primary subunit involved in the
repolymerization reactions 239. Condensation has also been observed in enzymatic
depolymerization strategies. Lignin model compounds and dimers underwent

44
condensation when treated with fungal laccases from Melanocarpus albomyces and
Trametes hirsuta 240. Interestingly, formation of some condensed lignin structures
appeared at different times dependent upon the redox potential of the laccase. Preventing
repolymerization could improve product yields from lignin depolymerization.
Perhaps the most promising and widely researched method for preventing lignin
repolymerization is the use of capping agents. Several capping agents have been
demonstrated to improve lignin product yield. Boric acid and phenol were used as capping
agents, with phenol improving product formation from organosolv olive tree pruning lignin
241. A phenol:lignin ratio of 2:1 resulted in the highest oil yield and lowest
repolymerization. Phenol as a capping agent is not without its limitations, however;
increasing the phenol:lignin ratio beyond 2:1 resulted in the formation of higher molecular
weight lignin, which is likely a result of the excess phenol reacting with products and
residual lignin. Formaldehyde has also been used as a capping agent for stabilizing lignin
during biomass pretreatment, greatly improving yields of guaiacyl and syringyl monomers
close to the their theoretical limits 242. Using a capping agent alongside an enzyme-IL pair
could improve product yields while enabling selective product formation.
The time and enzyme quantity required for significant biocatalytic lignin
valorization will also need to be investigated as a potential roadblock towards improving
yield. Most studies reported hours to days of reaction time used for enzymatic lignin
depolymerization in ILs. These reaction times are generally comparable to enzymatic
depolymerization of other biopolymers in IL, such as cellulose and hemicellulose. Studies
investigating the sugar (xylose and glucose) yields from enzymatic saccharification after
pretreatment of biomass with dilute [TEA][HSO4] or [C2C1Im][OAc] required 24-72h and

45
several mg protein/g biomass to achieve the maximum sugar yields 171, 243. Given the
heterogeneous nature of lignin it’s likely that the reaction times and enzyme quantities
required for enzymatic lignin depolymerization, prior to enzyme engineering, will be
similar or even greater. However, it is projected that the efficacy can be improved by
enzyme engineering for better enzymes and/or by process intensification and optimization
(i.e. immobilized enzymes) for better enzyme stability and recyclability.
1.8 Conclusions and perspectives
The great potential for lignin to be a renewable feedstock for commercial chemicals
is hindered by its structural heterogeneity and recalcitrance. With such an intense research
focus in recent years, great strides have been made in developing lignin valorization
strategies. ILs have been proven to be a capable class of lignin solvents and guiding
principles of designing biocompatible ILs have been proposed based on both experimental
and computational studies. Thermophilic and IL-tolerant LDEs have been explored for
increased resistance to harsh environments, highlighting their potential to be used
alongside biocompatible ILs. However, further research is needed to elucidate the LDE-
IL interactions for the purpose of lignin valorization. Toward this end, several research
areas appear important: 1) utilizing the suite of structural biology tools to explore the LDE-
IL interactions and how they affect the lignolytic capabilities of the biocatalysts, 2) protein
engineering strategies can be applied to create novel LDEs that can withstand high
concentrations of ILs while maintaining their lignolytic properties, 3) understanding the
interaction between IL and LDEs for better product selectivity and developing strategies
to prevent lignin condensation in IL; and 4) testing the effectiveness of lignin-IL-enzyme
systems on industrial relevant polymeric lignin streams on top of model compounds.

46
Collectively, this review highlights the tremendous potential of designing LDE-IL pairs for
lignin valorization in the IL medium.
1.9 Organization of Chapters
Figure 1-9. Project summary
The project summary in Figure 1-9 provides a general overview of the goal of this
dissertation. This dissertation seeks to design a system for biological lignin valorization
through the use of an aqueous IL and a laccase that can retain high levels of activity in the
aqueous IL. The aqueous IL will be used to pretreat biomass and extract lignin into the
aqueous phase during pretreatment. The laccase will then be introduced into the lignin-
containing aqueous phase to convert the extracted lignin into value-added products.
Chapter 1 of this dissertation provides a general overview of several topics as they relate
to the goal of this dissertation.
Chapter 2 of this dissertation seeks to establish a framework that subsequent
chapters can follow. This chapter also provides a proof-of-concept biocatalytic lignin
Lignin Ionic Liquids Biocatalyst
Chemicals

47
valorization system. The interactions between 3 aqueous ILs and a fungal laccase are
characterized using several techniques (biocompatibility screening, enzyme kinetics, and
molecular docking simulations) that will be used in subsequent chapters. The effect of the
ILs on lignin oxidation are also assessed to provide a foundation upon which the
subsequent chapters can improve.
Chapters 3 and 4 of this dissertation seek to produce a laccase that can retain high
levels of activity in concentrations of aqueous IL such that the aqueous IL is effective for
biomass pretreatment. Chapter 3 characterizes the interactions between aqueous ILs and a
hyperthermophilic bacterial laccase. Several single amino acid variants of this laccase are
produced in an effort to improve activity in the aqueous IL. Chapter 4 characterizes the
interactions between an aqueous IL and a thermophilic fungal laccase. Once again, several
variants of this laccase are produced, this time by engineering the enzyme surface charge,
in an effort to improve its activity in the aqueous IL.
Chapter 5 of this dissertation focuses on characterizing the interactions between a
mesophilic plant laccase and an ionic liquid. The interactions are characterized using the
same framework established in the Chapter 1 (biocompatibility screening, kinetics, and
molecular docking simulations) along with molecular dynamics (MD) simulations. The
goal of the MD simulations is to improve the understanding of the laccase-IL interactions
using the atomistic resolution afforded by MD simulations. This final chapter further
expands upon the framework used in previous chapters to understand laccase-IL
interactions.
The final chapter summarizes all of the work contained within the dissertation and
discusses many of the challenges faced during the experiments. This chapter also discusses

48
many of the challenges that could be faced in future studies that seek to achieve a similar
goal as this dissertation: characterizing the complex laccase-IL interactions and improving
laccase activity in aqueous ILs. Future work, both based on the results of this dissertations
and recent developments in the field of biomass pretreatment and lignin valorization, is
proposed and discussed as well.

49
CHAPTER 2. UNDERSTANDING LACCASE-IONIC LIQUID INTERACTIONS TOWARDS
BIOCATALYTIC LIGNIN VALORIZATION IN AQUEOUS IONIC LIQUIDS1
2.1 Abstract
Lignin is a crucial component of plant matter; however, it is also largely responsible
for the recalcitrance of lignocellulosic biomass when subjected to pretreatment processes.
Lignin is generated in large volumes as a waste product from paper and pulping industry
as well as cellulosic biorefineries. As a result, lignin valorization is critical to the
successful implementation of cellulosic biofuels. To this end, we investigated interactions
between three ionic liquids (AILs) and the lignolytic enzyme laccase toward biocatalytic
lignin conversion to aromatic monomers. Laccase exhibited minimal loss of activity in
10% [DEA][HSO4]. Changes in Vmax and Km of laccase with respect to IL concentration
indicate [DEA][HSO4] is a non-competitive inhibitor, whereas [Ch][Lys] and
[C2C1Im][OAc] are mixed inhibitors. Docking simulations suggested that [Ch][Lys] and
[C2C1Im][OAc] disrupt residues leading to the active site. Experiments with a β-O-4'
linked model dimer revealed laccase in [C2C1Im][OAc] and [Ch][Lys] requires the
presence of ABTS to oxidize the β-O-4' linkage, leading to polymerization of the model
dimer. Alkaline lignin treated with laccase, ABTS, and the AILs showed few structural
changes, although the lignin was partially solubilized and converted to degradation
products. The major products obtained from alkaline lignin were vanillin, acetosyringone,
syringaldehyde, and acetovanillone. The results of this study provide, for the first time, an
in-depth explanation of the interactions between laccase and AILs for the purpose of lignin
valorization.
1This chapter in its whole has been published in ACS Sustainable Chemistry & Engineering,
2019, 7 (19), 15928-15938. DOI: 10.1021/acssuschemeng.9b02151.

50
2.2 Introduction
Lignin is among the most abundant biopolymers found in nature and makes up
around one quarter of lignocellulosic biomass 244-246. Lignin is primarily made of three
phenylpropanoid subunits synthesized from corresponding alcohol precursors: p-coumaryl
alcohol (H), coniferyl alcohol (G), sinapyl alcohol (S) 40, 41. These monolignols are joined
via a complex network of linkages including β-aryl ether (β-O-4'), phenylcoumaran (β-5'),
and resinol (β-β') as the dominant linkages 43, 45. The nature of the linkages is largely
determined by the monolignol content of the biomass which, in turn, is dependent upon the
source of the biomass 44. The compositional and structural heterogeneity contributes
largely to lignin’s recalcitrance to biological or chemical decomposition. Currently, lignin
is generated in large volume as a waste product from the paper and pulping industry as well
as a byproduct of the cellulosic biorefinery 1, 247. Developing technologies to convert lignin
to value-added products will not only reduce waste generation but also increase revenue
for the paper industry as well as future cellulosic biorefineries 3, 4.
Lignin has limited use in its polymeric form. In contrast, the phenolic compounds
derived from lignin depolymerization can be used as fuels or building blocks to synthesize
chemicals for the food, plastic, and pharmaceutical industries, which are, at present,
primarily derived from petroleum 248-250. Lignin deploymerization methods can be
categorized into thermochemical, such as high temperature pyrolysis, hydrogenolysis,
catalytic oxidation, and biological, using lignolytic enzymes or microbes 5, 6, 8, 78. In
general, biological lignin depolymerization can be performed at milder temperatures and
pressures compared to thermochemical methods, potentially improving product selectivity
via the specificity of the biocatalysts 6, 12. A range of lignin degrading enzymes (LDEs)

51
have been investigated including lignin peroxidases, versatile peroxidases, manganese
peroxidases, laccases from white and brown rot fungi, and the newly discovered NAD or
glutathione dependent enzymes from certain soil bacteria, such as Sphingobium SYK-6 13-
15. However, application of these lignolytic enzymes in large-scale operations valorizing
lignin to aromatic monomers is still hindered due to the high cost of enzyme production
and poor yield of aromatic monomers due to the very low lignin solubility (for many of the
industrially relevant lignin streams) in the aqueous environment compatible with enzyme
activity as well as the poor selectivity of the lignolytic enzymes to produce defined
monomers.16, 17.
Most of the known effective lignin solvents (e.g., DMSO, acetone, or alkaline
solutions) are detrimental to enzyme activity when added to aqueous solutions 18, 19. In
addition, they often require high temperature for complete lignin dissolution, especially
when dealing with plant derived native lignins 20, which again can be incompatible with
the use of enzymes. Thus, a solvent system with both high lignin solubility and enzyme
compatibility is preferred for developing a better biological lignin depolymerization
process. Ionic liquids (ILs) are molten salts made of organic cations and anions that are
liquid at temperatures <100 oC 127. By selecting the cation and anion, the properties of an
IL can be tuned for a wide range of applications128, 131, 133. Alkylimidazolium based ILs
have been extensively studied for biomass pretreatment and lignin fractionation251-253.
Recently, ILs from bio-derived cations (such as choline and ammonium) and anions (such
as amino acids and carboxylic acids) have been investigated as lower-cost alternatives to
imidazolium-based ILs 167. For example, pretreating switchgrass with 10% cholinium
lysinate ([Ch][Lys]) in water resulted in >80% of the lignin being solubilized into the liquid

52
stream 254. Results also suggest that tuning aqueous IL (AIL) mixtures can favor specific
products during a catalytic oxidation process 255.
Laccases are a member of the superfamily of multi-copper oxidases (E.C. 1.10.3.2).
First discovered in extracts from the Chinese lacquer tree (Toxicodendron vernicifluum,
previously named Rhus vernicifera), laccases have since been found in a variety of
organisms including plants, fungi, bacteria, and archaea 84, 85, 90, 91. Laccases utilize a multi-
copper cofactor for substrate oxidation 86. Rather than requiring H2O2 as in other lignolytic
enzymes, laccases use O2 as the final electron acceptor 66, 71, 80. Furthermore, oxidation of
small-molecule mediators by laccase, such as 1-hydroxybenzotriazole (HBT) or 2,2'-azino-
bis(3-ethylbenzothiazoline-6-sulphonic acid) (ABTS), enables the oxidation of non-
phenolic lignin compounds 229, 230. This feature, alongside their inherent lignolytic
capabilities and their activity under milder reaction conditions, makes laccases a desirable
biocatalyst for lignin upgrading strategies 97, 256. ILs have been found to be capable of
supporting and, in a few cases, improving activity and stability of enzymes including
laccases 31, 32, 186, 257. T. versicolor laccase activity decreased with increasing IL
concentration of the two alkylimidazolium ILs tested, [C2C1Im][EtSO4] and
[C2C1Im][OAc] 183. However, Galai et al. screened T. versicolor laccase activity in 56
AILs and found that 13 of the AILs improved laccase activity; the most notable
improvement was seen in choline dihydrogenphosphate ([Ch][H2PO4]) which increased
laccase activity 451% 150.
Given the unique properties that certain AILs provide regarding lignin solubility
and biocompatibility, the utilization of AIL and LDEs confluently holds great potential for
developing a new strategy for lignin extraction and depolymerization. Despite previous

53
exploration of catalysis and enzyme activity in AILs, there is a gap in our understanding
of the mechanisms underlying the interactions between lignolytic enzymes and these
solvents. This study aims to elucidate the interactions between laccase and ILs, inhibition
mechanism and product profiling on such a biocatalytic system with laccase and three
AILs. To achieve this goal, we screened the ILs 1-ethyl-3-methylimidazolium acetate
([C2C1Im][OAc]), [Ch][Lys], and diethylamine hydrogensulfate ([DEA][HSO4]) for their
biocompatibility with a fungal laccase. We then investigated enzyme kinetics and the
inhibition mechanism using docking simulations with the ILs and the active site. Finally,
the IL-laccase system was tested on a β-O-4' linked model dimer (GGE) and commercial
kraft lignin in the presence of the mediator ABTS; the resulting residual lignin and lignin-
derived products were then characterized. This work provides a better understanding of
the inhibition mechanisms of ILs on laccase for developing a biocatalytic lignin
valorization process using AILs and LDEs.
2.3 Methods and Materials
2.3.1 Materials
Laccase from T. versicolor (0.66 U/mg), 2,2'-azino-bis(3-ethylbenzothiazoline-6-
sulphonic acid) (ABTS), alkaline lignin (lot# MKBV5831V), choline hydroxide, L-lysine,
and the IL [C2C1Im][OAc] were purchased from MilliporeSigma (St. Louis, MO, USA).
Diethylamine (DEA), sulfuric acid (H2SO4), guaiacylglycerol-β-guaiacyl ether (GGE),
tetrahydrofuran (THF), 2-methyltetrahydrofuran (MeTHF), chromium (III)
acetylacetonate (Cr(acac)3), and ethyl acetate were purchased from Tokyo Chemical
Industry Co. (Portland, OR, USA). The ILs, cholinium lysinate ([Ch][Lys]), and

54
diethylamine hydrogensulfate ([DEA][HSO4]) were synthesized using the anion and cation
precursors according to previously developed protocols 167.
2.3.2 Biocompatibility screening
The biocompatibility of three ILs, [C2C1Im][OAc], [Ch][Lys], and [DEA][HSO4]
in their aqueous solution (approximate 1-10% w/v), was screened with T. versicolor
laccase. To delineate the effect of pH, the IL solutions were adjusted to pH 5.0 using 1 M
sodium hydroxide or 1 M hydrochloric acid prior to testing. Activity was screened with
20 mM sodium citrate buffer, AIL (0, 1, 2, 3, 4, or 10% w/v) and 2 mM ABTS in clear, flat
bottom 96-well Costar assay plates (Corning Inc., Kennebunk, ME). Laccase was loaded
at 3 concentrations: low (0.6 μg mL-1), medium (1.2 μg mL-1), and high (3.0 μg mL-1). The
plate was incubated at 40oC in a SpectraMax M2 plate reader (Molecular Devices,
Sunnyvale, CA). Absorbance readings were taken every 30 seconds for 5 min at a
wavelength of 420 nm. Plates were shaken for 5 seconds prior to initial reading and 3
seconds before each reading to ensure well mixing. Oxidation of ABTS in buffer and AILs
without laccase were measured as blanks.
2.3.3 Inhibition kinetics
Michaelis-Menten curves were generated for different IL concentrations by
measuring initial velocities for 11 different ABTS concentrations at a medium loading (1.2
μg mL-1) of laccase. ABTS oxidation was measured using the same method as described
in the biocompatibility screening in 96-well plates. The oxidized ABTS was quantified
using an absorbance coefficient ε = 36,000 M-1 cm-1 258. A unit of activity was defined as
the amount of laccase that oxidizes 1 μmol of ABTS per minute. Laccase initial activity

55
as a function of ABTS concentration ([S]) was fit to the Michaelis-Menten curve, shown
in equation 2-1.
V = Vmax×[S]
Km+[S] (2-1)
These curves were linearized using the Hanes-Woolf method, resulting in equation 2-2.
[S]
V=
1
Vmax× [S] +
Km
Vmax (2-2)
Equation 2 was used to calculate the kinetic coefficients, Km and Vmax of laccase in different
AILs.
2.3.4 Molecular docking simulations
In order to determine likely interactions between IL molecules and laccase, docking
simulations were performed using the YASARA Structure software package (YASARA
Biosciences GmbH, Vienna, Austria) with the AutoDock Vina plugin 259. Ion structures
and parameters were generated in YASARA from SMILES representations and energy
minimized. The crystal structure of T. versicolor laccase (PDB ID: 1GYC) was used as a
target in the docking simulations 86. Cations and anions were treated as separate ligands,
with 25 simulations run for each, and the target laccase was held rigid throughout all
simulations. Results of docking were visualized with PyMol (Schrodinger LLC, New
York, NY).
2.3.5 Lignin depolymerization in aqueous IL
GGE dimer: Prior to experiments with the model compound, all solutions were
adjusted to pH 5.0 with 1 M sodium hydroxide or 1 M hydrochloric acid. Laccase was
loaded to a final concentration of 1.2 μg mL-1 and ABTS was added for a final
concentration of 2 mM. The GGE model compound was added for a final concentration

56
of 0.5 mg mL-1. AILs were added for final concentrations of 2% (w/v) [C2C1Im][OAc],
1% [Ch][Lys], and 10% [DEA][HSO4]. Reactions were carried out in 1 mL glass tubes
under 30 psi O2 pressure at 40oC inside a 100 mL Parr reactor (4593 bench top reactor,
Alloy C276, Parr Instruments, IL). The reactor was purged with pure O2 for 1 minute and
then the release valve was closed to maintain O2 pressure. The reaction was allowed to
proceed for an additional 2 h.
After reaction, product profiles were analyzed by isocratic HPLC using an 80%
water – 20% acetonitrile mobile phase at a flow of 0.5 mL min-1 for 30 minutes through an
Ultimate 3000 HPLC system (Dionex Corporation, Sunnyvale, CA) equipped with
ultraviolet (UV) detector. Separation was achieved through use of a Zorbax Eclipse XDB-
C18 column (5 μm particle size, 150 x 4.6 mm i.d., Agilent Technologies, Santa Clara,
CA) at 25oC. Elution profiles were measured by UV absorbance at 280 nm. The molecule
weight distribution of the products was characterized using gel permeation
chromatography (GPC) while liquid chromatography–mass spectrometry (LC-MS) was
used to identify the major products in GGE experiment as described in the “Lignin
Characterization” section.
Alkaline lignin: The same reaction protocol was used for alkaline lignin samples as
for the GGE dimer except that 500 mg of kraft lignin was loaded with the reactants (1.2 μg
mL-1 laccase, 2 mM ABTS, AILS) in the Parr reactor with a final reaction volume of 10
mL. After the reaction, residual lignin was precipitated by addition of 20 mL water and
centrifugation at 7,000 rpm for 15 minutes. The aqueous fraction was subjected to liquid-
liquid extraction using a 1:1 volume of ethyl acetate: liquid fraction, with the extraction

57
repeated 3 times. The ethyl acetate was evaporated in a vacuum oven set at room
temperature for 24 h to leave a thick oil. Oil yield was calculated using equation 2-3.
Oil Yield (%yield) = weight of oil
weight of initial lignin× 100 (2-3)
The precipitated solids (residual lignin) were washed with 50 mL water three times
to remove any excess IL and dried in a convection oven at 45oC for 48 h. Lignin conversion
was calculated using equation 2-4.
Conversion (%) = weight of initial lignin−weight of remaining lignin
weight of initial lignin× 100 (2-4)
The oil was dissolved in 750 μL MeTHF and analyzed by GC-MS for lignin
decomposition products. Identification and quantification of the monomeric products from
the depolymerization reaction were performed on an Agilent 7890B GC coupled to a
5977B MS with an HP-5ms (60 m × 0.32 mm) capillary column 260. The temperature
program started at 50°C and increased to 120°C at 10°C min−1 with a holding time of 5
min; then it was raised to 280°C at 10°C min−1 with a holding time of 8 min and to 300°C
at 10°C min−1 with holding time of 2 min 261. The inlet was held in splitless mode at 250oC
using helium as a carrier gas at a flow rate of 1.2 mL min−1. Calibration curves were created
using commercially available pure compounds: guaiacol, syringol, vanillin, acetovanillone,
and homovanillic acid (Sigma Aldrich, St. Louis, MO). Monomer yields were calculated
on basis of mg/g starting lignin.
2.3.6 Lignin characterization
Gel Permeation Chromatography (GPC): The weight-average molecular weight
(Mw) and number-average molecular weight (Mn) of both precipitated lignin and lignin
products in the liquid phase following reactions were determined by GPC. The precipitated

58
lignin was acetylated via acetobromination 262. Briefly, 10 mg of the lignin was dissolved
in 2.5 mL of 92:8 (v/v) acetic acid and acetyl bromide and mixed at 50oC for 2 h. Excess
acetic acid and acetyl bromide was evaporated by purging with N2. The acetylated lignin
was dissolved in 750 uL THF and analyzed by direct injection into an HPLC system
equipped with a Mixed-D PLgel column (5 μm particle size, 300 x 7.5 mm i.d., molecular
weight range of 200 – 400,000 μm, Polymer Laboratories, Amherst, MA, USA) at 50°C.
The THF flow rate was 0.5 mL min-1. Eluted molecular species were detected by
absorbance at 300 nm, which was calibrated using a polystyrene standards kit (Sigma-
Aldrich, St. Louis, MO). The molecular weight distribution of model compound products
was analyzed by direct injection into the HPLC system using the aforementioned method.
Fourier Transform Infrared Spectroscopy (FTIR): Chemical structures of alkaline
lignin and treated lignin were determined using a Thermo Nicolet 870 FTIR-ATR
spectrometer. Lignin sample spectra were obtained with 64 scans at wavelengths from 700
to 4000 cm-1 with a spectral resolution of 2 cm-1. The raw FTIR spectra were baseline
corrected and normalized using Omnic 6.1a software and compared in the range 750-4000
cm−1.
Differential Scanning Calorimetry (DSC): DSC measurements of alkaline and
treated lignin samples were performed using a DSC Q20 (TA Instruments, New Castle,
DE) equipped with an autosampler. Analysis was carried out in a temperature range of 40-
500°C at a rate of 10°C min-1 and N2 flow of 50 mL min-1.
1H-13C HSQC NMR: Approximately 100 mg lignin sample along with ca. 1 mg
Cr(acac)3 were dissolved in DMSO-d6/pyridine-d5 (4:1) under mild heat and sonication in
a NMR tube until a homogeneous mixture was obtained. NMR spectra were acquired on

59
a 500 MHz JEOL ECZr (Peabody, MA, USA) spectrometer equipped with a 5-mm Royal
Probe. The central DMSO solvent peak was used as an internal reference (C 39.5, H 2.5
ppm). The 1H–13C correlation experiment was an HSQC experiment (JEOL standard pulse
sequence ‘hsqc_dec_club_pn’). HSQC experiments were carried out using the following
parameters: acquired from 12 to 0 ppm in f2 (1H) with 1024 data points (acquisition time
136 ms), 220 to 0 ppm in f1 (13C) with 256 increments and 150 scans with a 1 s interscan
delay. In all cases, processing used typical sine bell (90o) in f2 and squared sine-bell (90o)
in f1 (first point 0.5). Volume integration of contours in HSQC plots used Maetrelabs
MestReNova 12.0 (Mac version) software. Integrals were from volume-integration of C/H
pairs with similar properties, the α-C/H correlations of A (β-O-4'), B (β-β'), and C (β-5)
units. Spectra are displayed in absolute value mode and color coded (in Adobe Illustrator
CC 2018) using literature reference standards 263.
Model Compound Mass Spectrometry: Mass spectrometry, to identify large
molecular weight products from GGE experiments, was performed on a Q-Exactive
(ThermoScientific, Waltham, MA, USA) orbitrap spectrometer. The instrument
parameters were as follows; a 3.8 kV spray voltage, 225oC inlet temperature, sheath
auxiliary gas flow of 2 (arbitrary units), a mass resolution of 140,000, with nitrogen as the
HCD collision gas. A 10 L aliquot of each sample was reduced to dryness under nitrogen.
The dry residue was dissolved in 100 L of a solution containing 50% acetonitrile and 50%
10 mM aqueous sodium hydroxide. The final solution was infused directly into the mass
spectrometer by syringe pump at a flow rate of 3 mL per minute. For the HCD tandem
mass spectrometry experiments, precursor ions were isolated using a window of 3 m/z, and

60
a normalized collision energy of 25 NCE was applied. Data were processed using Xcalibur
software (ThermoScientific, Waltham, MA, USA).
2.4 Results and Discussion
2.4.1 Biocompatibility screening
In order to test whether laccase is still active in AIL solution, three ILs were
screened for their biocompatibility with T. versicolor laccase (TvL) at concentrations
ranging from 1-10% IL in water. Results indicate that [Ch][Lys] and [C2C1Im][OAc] are
more inhibitory to TvL with 2% [Ch][Lys] or 5% [C2C1Im][OAc] causing significant loss
(>70%) of TvL activity (Fig. 2-1). In contrast, [DEA][HSO4] is more compatible with TvL,
as indicated by the modest loss (<20%) of TvL activity even at 10% concentration. The
tested IL concentrations represented a reported range of AIL biomass pretreatment
conditions. For example, Liu et al. showed that 10% [Ch][Lys] was highly effective in
removing lignin from switchgrass varieties during pretreatment 172, and [C2C1Im][OAc] at
20-50% in water can remove a large portion of the hemicellulose and lignin when
compared to pure IL 171, 264. Gschwend et al. have demonstrated that 80 wt%
triethylammonium hydrogen sulfate ([TEA][HSO4]) is capable of improving Miscanthus
saccharification yields and lignin removal after pretreatment for 15 minutes at 180oC 243.

61
Figure 2-1. Activity of T. versicolor laccase in AIL solutions. H = high laccase
concentration (3.0 μg mL-1), M = medium laccase concentration (1.2 μg mL-1), L = low
laccase concentration (0.6 μg mL-1). Values indicate the ratio of TvL activity in ILs relative
to TvL activity in buffer (0% IL).
Both the cations and anions of the AILs could impact their activity. It has been
shown that the toxicity of imidazolium-based ILs increases with increasing cation size 265.
[C2C1Im][OAc] at 2% concentration strongly inhibited cellulase activities and growth of
common biofuel fermentation microbes such as yeast and E. coli 266 while [Ch][Lys]
exhibited low toxicity to yeast and E. coli 170. A more recent study further suggested that
cellulase enzymes and yeast can tolerate 5% [Ch][Lys] 267. However, it appeared that
[Ch][Lys] was more inhibitory to the TvL than [C2C1Im][OAc] and [DEA][HSO4]. This
case agrees with previous reports showing that ILs with sulfate anions are generally more
biocompatible with laccases 149, 268. However, we did not control for the cation between
each AIL so the biocompatibility cannot be solely attributed to the anion in this case.
1.00-0.83
0.82-0.66
0.65-0.50
0.49-0.33
0.32-0.16
0.15-0.00

62
2.4.2 Inhibition mechanisms
To better understand the different effects of the three ILs on TvL, we determined
their inhibition kinetics using ABTS as a substrate. [C2C1Im][OAc] and [Ch][Lys] are
mixed inhibitors of TvL, decreasing Vmax and increasing Km (Table 2-1). This type of
inhibition suggests that the ILs do not directly compete with the substrate for the active
site, but rather affect the residues surrounding the active site to interfere with substrate
binding. [DEA][HSO4] in contrast is a non-competitive inhibitor, not affecting substrate
binding but decreasing the catalytic rate of the enzyme. To the best of our knowledge,
there have not been any previous studies assessing the inhibitory mechanisms of [Ch][Lys]
and [DEA][HSO4] on laccases. The effects of alkylimidazolium ILs on TvL kinetics have
been reported, though no studies have been performed with [C2C1Im][OAc]. Tavares et
al. found that 3 ILs with a [C2C1Im]+ cation reduced both the Vmax and Km of TvL with
ABTS as a substrate, supporting our finding with [C2C1Im][OAc] 191.
Table 2-1. Kinetic constants of ABTS oxidtion by T. versicolor laccase in AILs
Ionic Liquid Concentration
Ionic Liquid 0% (w/w) 2% (w/w) 5% (w/w)
Vmax (U mg-1 TvL)
[C2C1Im][OAc] 23.1 ± 1.0a 16.2 ± 2.4b 9.0 ± 0.4c
[Ch][Lys] 23.1 ± 1.0a 7.2 ± 0.3b 5.8 ± 0.3b
[DEA][HSO4] 23.1 ± 1.0a 17.9 ± 0.3b 21.6 ± 0.3a
Km (mM ABTS)
[C2C1Im][OAc] 0.065 ± 0.023a 0.531 ± 0.090b 1.469 ± 0.012c
[Ch][Lys] 0.065 ± 0.023a 1.154 ± 0.124b 2.847 ± 0.300c
[DEA][HSO4] 0.065 ± 0.023a 0.104 ± 0.013a 0.111 ± 0.023a

63
2.4.3 Docking simulations
To gain a deeper insight into the inhibition mechanisms, docking simulations were
carried out with the IL cations and anions (Fig. 2-2). A high probability binding site for
[C2C1Im][OAc] and [Ch][Lys] was found near the Type I copper, thus possibly decreasing
the catalytic rate by obstructing removal of electrons during substrate oxidation at the Type
I copper. Binding of these ILs at this site may also interfere with ABTS interaction,
accounting for the observed increase in Km. In contrast, a binding site for [DEA][HSO4]
was not identified, with any significant probability near the active site, consistent with the
observed weak noncompetitive inhibition.
Figure 2-2. Results of simulations showing the docking of b) [C2C1Im][OAc], c) [Ch][Lys],
and d) [DEA][HSO4] near the Type I copper of T. versicolor laccase. The red arrow in a)
points to the Type I copper. The ILs are represented by the multi-colored stick models in
panels b-d and the His residues that coordinate the coppers are colored blue.
Docking simulation of [C2C1Im][OAc] in this study corroborates a previous study
on alkylimidazolium ILs and laccases. MD simulations with alkylimidazolium cations
found that cations of varying chain length interacted with the active site via hydrophobic
interactions 269. The shorter chain length cations (C2-C6) were capable of diffusing into the
active site; longer chain length cations (C8-C10) were limited to binding to Leu residues
around the active site entrance. Several studies have shown that enzyme-IL interactions,
(a) (b) (c) (d)

64
particularly interactions with the anion, lead to disruption of protein stability as predicted
by the Hofmeister series 133, 270, 271. The more chaotropic anions lead to disruption of the
water shell around a protein, causing destabilization of the protein. The specific anion
effect can only be assessed when the cation is consistent, but [HSO4]- has been observed
to be among the most stabilizing of the anions 33.
2.4.4 GGE dimer transformation
The activity screening results support the possibility of using TvL in dilute or more
concentrated AIL solutions, depending on the IL introduced, to process lignin. The
phenolic lignin dimer (GGE) was first tested because the dimer is linked by β-O-4' bond
which is the most common interunit linkage in lignin. In absence of ABTS, TvL in 1%
[Ch][Lys] or 2% [C2C1Im][OAc] was inhibited from processing GGE, whereas TvL in 10%
[DEA][HSO4] experiences no inhibition to processing GGE (Fig. 2-3a). Some studies have
found that acidic ILs are capable of cleaving the β-O-4' linkage on their own at elevated
temperatures 272. The temperature used in this study (40oC) was much lower than previous
studies (150oC), so it is unlikely the [DEA][HSO4] itself was responsible for any major
changes made to GGE during the reaction. Upon addition of ABTS, TvL is able to process
GGE in the presence of all ILs (Fig. 2-3b).

65
Figure 2-3. HPLC chromatograms of GGE incubated with laccase and different AILs a)
without ABTS present during the reaction. b) ABTS present during the reaction. GGE
elutes at ~23 min and reaction products elute in the 2-5 min range.
To better assess changes made to GGE during the reaction, the products were
further analyzed with GPC and LC-MS. The weight-average molecular weight (Mw),
number-average molecular weight (Mn), and the polydispersity index (PDI) of all samples
increased during the reaction with GGE (Table 2-2). The samples with an IL present had
(a)
(b)

66
a lower Mw and higher PDI after 120 minutes compared to the control without AIL. The
Mw of the control after 120 minutes was 1,315 g mol-1, whereas the highest Mw of samples
with IL was 1,087 g mol-1 in 1% [Ch][Lys]. The PDI of the No IL control was 1.18 after
120 minutes, whereas the PDI of samples with ILs were 1.57 (1% [Ch][Lys]), 1.69 (2%
[C2C1Im][OAc]), and 1.90 (10% [DEA][HSO4]). The Mn of IL samples decreased between
30 and 120 minutes, which led to the substantial increase in PDI at the end of the reaction.
Molecular weight distribution plots (Fig. 2-4) suggest this is a result of low-molecular
weight species appearing in the 120 min samples. To identify any higher-order structures,
e.g. dimers, trimers, or oligomers, extracted products were identified with LC-MS.
Table 2-2. Molecular weight distribution of GGE before (0 min), during (30 min), and after
(120 min) treatment with TvL, ABTS, and ILs1
Mw (g mol-1)
IL 0 min 30 min 120 min
No IL 447 941 1315
2% [C2C1Im][OAc] 447 877 975
1% [Ch][Lys] 447 881 1087
10% [DEA][HSO4] 447 993 984
Mn (g mol-1)
IL 0 min 30 min 120 min
No IL 402 817 1118
2% [C2C1Im][OAc] 402 742 576
1% [Ch][Lys] 402 741 694
10% [DEA][HSO4] 402 762 519
PDI
IL 0 min 30 min 120 min
No IL 1.11 1.15 1.18
2% [C2C1Im][OAc] 1.11 1.18 1.69
1% [Ch][Lys] 1.11 1.19 1.57
10% [DEA][HSO4] 1.11 1.30 1.90
1Mw: weight-average molecular weight, Mn: number-average molecular weight, PDI:
polydispersity index

67
Figure 2-4. Molecular weight distributions of products after GGE reaction with TvL,
ABTS, and a) buffer without IL, b) 2% [C2C1Im][OAc], c) 1% [Ch][Lys], d) 10%
[DEA][HSO4]
Direct infusion mass spectrometry experiments using a high-resolution accurate-
mass orbitrap mass spectrometer (Q-Exactive) (ThermoScientific, Waltham, MA, USA) in
the negative ion mode with sodium hydroxide as a pH modifier were performed. The
infusion mass spectra were dominated by three ion peaks with m/z values much greater
than the starting GGE dimer. High-resolution, accurate mass spectra and tandem mass
spectrometry allowed the probable identity of these species to be defined. The ion peak at
m/z 637.1919 was consistent with the coupling of two GGE dimers (GGE dimer MW 320
g mol-1) with the proposed structure shown in Figure S2-2a (calculated m/z 637.1927,
observed m/z 637.1918, error 1.3037 ppm). The ion peak at m/z 953.3242 was consistent
Molecular Weight (g mol-1)
101102103104
No
rma
lize
d I
nte
ns
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0 min
30 min120 min
Molecular Weight (g mol-1)
101102103104
No
rma
lize
d I
nte
ns
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0 min30 min120 min
Molecular Weight (g mol-1)
101102103104
No
rma
lize
d I
nte
ns
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0 min30 min120 min
Molecular Weight (g mol-1)
101102103104
No
rma
lize
d I
nte
ns
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0 min30 min120 min
(d) (c)

68
with the coupling of three GGE dimers with the proposed structure shown in Figure S2-2b
(calculated m/z 953.3237, observed m/z 953.3241, error 0.3765 ppm). Finally, the ion
peak at m/z 1073.3454 was consistent with the addition of a G monomer unit to the three
GGE oligomer (m/z 953.3242) with the proposed structure shown in Figure S2c (calculated
m/z 1073.3449, observed m/z 1073.3448, error -0.0172 ppm).
Previous studies have shown that laccases facilitate both the depolymerization and
polymerization of lignin model compounds. LC-MS analysis of extracted products formed
during guaiacylic and syringilic lignin model compound oxidation by Melanocarpus
albomyces and Trametes hirsuta laccases showed that GGE homodimerized to form a
single product via a 5-5' coupling mechanism, identical to that shown in Figure S2-2a 240.
ABTS was also found to participate in the polymerization of a β-O-4' lignin model dimer
and guaiacol when treated with T. hirsuta laccase, although the structures of the products
formed during the reaction were not identified 273. Neither of these studies investigated the
role of ILs during polymerization of lignin model compounds. We have shown that the
presence of ABTS and, possibly the ILs, facilitates the polymerization of GGE into
structures larger than the single GGE homodimer reported previously 240. Additionally,
the molecular weight distributions of the reaction products suggest that the presence of ILs
promotes the formation of low-molecular weight compounds during model compound
polymerization/depolymerization.
2.4.5 Depolymerization of alkaline lignin
To further investigate the utility of TvL-AIL systems, we tested their ability to
depolymerize commercial alkaline lignin with the same reactors used for other studies. In
comparison to a control lacking IL (2.12% conversion based on the mass of residual solids),

69
higher lignin conversions were achieved for TvL catalysis of alkaline lignin in AIL systems
(Table 2-3). The highest conversion was obtained with TvL in 10% [DEA][HSO4]
(21.66%), followed by TvL in 1% [Ch][Lys] (17.24%) and TvL in 2% [C2C1Im][OAc]
(11.72%). However, 1% [Ch][Lys] resulted in the highest yield of oil (1.28%) as compared
to 2% [C2C1Im][OAc] (0.84%) and 10% [DEA][HSO4] (0.74%). Analysis of extracted
products with GC-MS revealed vanillin to be the primary degradation product (consisting
55-70% of the quantifiable monomers), followed by syringaldehyde, acetovanillone, and
guaiacol (Fig. 2-5). The discrepancy between lignin conversion and oil yield, which was
particularly marked for [DEA][HSO4] (21.66% conversion vs. 0.74% oil yield), could be
attributed to less extractable oligomeric compounds formed by the laccase, similar to the
condensation of the GGE model compound, that cannot be detected by GC-MS.
Table 2-3. Converstion and yield of alkaline lignin degradation products
* Fraction extracted using ethyl acetate
Conversion
(%) Lignin oil yield* (%)
Identifiable monomer yield (mg∙g-1)
Sample Guaiacol Vanillin Syringaldehyde Acetovanillone Total
No IL 2.12 0.74 0.00 1.02 0.59 0.46 2.07
2% [C2C1Im][OAc] 11.72 0.84 0.00 1.15 0.68 0.54 2.37
1% [Ch][Lys] 17.24 1.28 0.54 1.86 1.24 0.98 4.62
10% [DEA][HSO4] 21.66 0.74 0.18 1.45 0.81 0.68 3.12

70
Figure 2-5. GC-MS chromatograms of alkaline lignin breakdown products after treatment
with laccase, AILs, and ABTS. Major products identified were 1) guaiacol, 2) vanillin, 3)
acetovanillone, 4) homovanillic acid, 5) syringaldehyde, and 6) acetosyringone.
Gel permeation chromatography (GPC) results revealed all treatments led to a small
increase in average molecular weight when compared to untreated alkaline lignin (Table
2-4). Additionally, the PDI decreased for all treatments when compared to untreated
alkaline lignin. Treatment of alkaline lignin with laccase and 1-hydroxybenzatriazole
(HBT), another mediator, resulted in a reduced PDI and increased molecular weight 113.
GPC of supernatants revealed the presence of higher molecular weight solids in treatments
with ILs compared to treatment without IL. These results indicate the laccase-AIL system
could facilitate both depolymerization of lignin to a low molecular weight fraction as well
as larger products resulting from subsequent polymerization. This polymerization could
be limited by a capping agent like phenol, which has been used to improve monomer yields
from organosolv olive tree pruning lignin 241, although the amount of phenol required is
Time (min)
6 7 8 9 10 16 18 20 22 24
Inte
ns
ity
0
2e+5
4e+5
6e+5
8e+5
1e+6
No IL
2% [C2C1Im][OAc]
1% [Ch][Lys]
10% [DEA][HSO4]
1
2
3
4
5 6

71
quite high (2:1 phenol:lignin). The results with the model compound suggest that some
AILs could act as biocompatible capping agents, though this possibility will need to be
investigated further in a separate study.
Table 2-4. Processed lignin molecular weight distribution and polydispersity of lignin
streams1
Solid Phase Liquid Phase
Sample Mw (g mol-1) Mn (g mol-1) PDI Mw (g mol-1) Mn (g mol-1) PDI
Alkaline lignin 5677 1974 2.88 N/A N/A N/A
No IL 5457 2470 2.21 693 450 1.54
2% [C2C1Im][OAc] 5358 2482 2.16 709 542 1.31
1% [Ch][Lys] 5772 2454 2.35 654 535 1.22
10% [DEA][HSO4] 6170 2068 2.98 667 526 1.27
1Mw: weight average molecular weight; Mn: number average molecular weight; PDI:
polydispersity; N/A: not applicable.
The chemical and structural properties of the resulting lignin stream were further
characterized using FTIR, DSC and 2D HSQC NMR. Analysis of FTIR spectra indicated
minimal differences between untreated and treated lignin samples (Fig. S2-3a). The only
difference can be seen is in the peak at 1,110 cm-1. This peak corresponds to the aromatic
C-H plane deformation in S-lignin, and is reduced for treated lignin samples when
compared to untreated alkaline lignin, which is consistent with the identification of
derivatives of S-lignin with GC-MS. The bands at 1,220 cm-1 corresponding to C-C, C-O,
and C=C stretching in G-lignin were unchanged for treated lignin when compared to
untreated alkaline lignin. Preferential breakdown of S-lignin has been observed before via
oxidation of alkaline lignin using transition metal catalysts, so it’s possible that the

72
biocatalyst follows the same trend 254, 274. Thermal profiles as shown by differential
scanning calorimetry are similar for all samples of untreated and treated alkaline lignin
(Fig. S2-3b). Exothermic and endothermic regions are indicated by positive and negative
values of Hf, respectively. An exothermic region between 50-100oC was observed for all
samples. Similarly, an endothermic region between 400-450oC was observed for all
samples. The mild reaction conditions, relatively low temperature and low IL
concentrations did not appear to result in any significant changes to the lignin structure.
2D HSQC NMR spectra show a small amount of S-lignin in the starting material,
however there does not appear to be a substantial reduction in these peaks after processing
suggesting only a small portion of the starting material was solubilized during processing
(Fig. 2-6). There appears to be shifts in the ratios of β-O-4' and β-β' linkages in alkaline
lignin after treatment, but these changes are not above experimental variation. The most
substantial changes were the relative ratios of β-O-4' and β-β' linkages in the alkaline lignin
treated with laccase and ABTS (KLA), while alkaline lignin treated with laccase and ABTS
in [C2C1Im][OAc] (EKLA) saw relatively minor changes in the HSQC. β-O-4' linkages in
alkaline lignin treated without any AILs present were reduced from 54% to 41%. Alkaline
lignin treated with aqueous [C2C1Im][OAc] in addition to laccase and ABTS resulted in
increased β-O-4' (54% → 56%) and reduced β-β' (29% → 26%) linkages. Previously, Li
et al., reported the water-insoluble fraction of alkaline lignin after treatment with laccase
and the mediator HBT had increased β-O-4' (61.5% → 68.5%) and reduced β-β’ (5.8% →
5.7%) content compared to the starting material, suggesting the presence of less branched
lignin structure when compared to the starting material 113.

73
Figure 2-6. 13C-1H (HSQC) spectra of aliphatic (top) and aromatic (bottom) regions of a)
raw alkaline lignin (KL), b) alkaline lignin treated with laccase and ABTS in buffer (KLA),
and c) alkaline lignin treated with laccase and ABTS in 2% [C2C1Im][OAc] (EKLA).
Figure S6 in the supplementary material contains all of the spectra from the study. The
structures of side-chain linkages and lignin compositional units, shown in the middle of the
figure, were coded with colors corresponding to the cross peaks in the spectra aliphatic and
aromatic regions, respectively. Linkage percentages were calculated via volume
integration of the Cα-H of the β-O-4', β-5', and β-β' linkages and treated as a relative ratio
of the three. Volume integrations are intended to be only semi-quantitative due to problems
inherent to HSQC experiments including a wide array of potential 1JC-H values and rates of
T2 relaxation 260.
It is possible that there are IL-specific effects that altered the changes in linkage
ratios seen after treatment with laccase and ABTS. The relative content of ether bond was
based on NMR analysis of the residual solids rather than the liquid products. Results
suggest that the relative ether bond content stayed unchanged or slightly increased in the

74
residuals solids. The relatively small increase in ether bonds from KLA to EKLA is likely
negligible, however, it is possible that dehydrogenation reactions of the β-β linkages
rendered a small amount of these linkages without C-H groups at the Cα-H and Cβ-H
position causing a small increase in the β-O-4 content. A similar phenomenon has been
observed when kraft lignin was treated with T. versicolor laccase and 1-
hydroxybenzotriazole hydrate (HBT), another mediator 275. The insoluble lignin fraction
in their study had a higher ether bond content than the starting material. It is also noted
that these numbers should not be over-interpreted as HSQC NMR is a semi-quantitative
method (due to variations in T2 relaxations and 1JC-H constants) for quantifying the linkages
in their relative ratios. It is surprising that S-lignin was identified in the raw and treated
kraft lignin samples. We encountered the similar problem in our previous work 255 where
a small quantity of S-lignin was observed in the original Sigma-Aldrich alkaline lignin.
While the majority of the lignin is likely softwood, it is possible that the lignin streams
used for the Sigma Aldrich kraft lignin are made of a small amount of other species (e.g.,
hardwood).
The source of the lignin itself could play a role in the depolymerization since the
alkaline (kraft) lignin has been reported to be heterogeneous and condensed 276. To fully
understand these effects, a less recalcitrant lignin source may be tested as the commercial
alkaline/kraft lignin tends to be more condensed (more C-C bonds) than the native lignin
in plants 277. Due the intrinsic complexity of real lignin (alkaline lignin in this study) and
the limitations of current lignin analytical approaches, many of the proposed lignin
transformation mechanisms are based lignin model compounds such as monolignols and
GGE dimmer. In this work, we were able to identify the major products in GGE experiment

75
using LC-MS and propose possible reaction pathways. The lignin transformation
mechanism with GGE dimmer could be partially transferable to the alkaline lignin
substrate; however, to fully reveal the alkaline lignin transformation mechanism is still
very challenging based on current lignin analytical techniques.
Summarizing the results of GGE dimer and alkaline lignin depolymerization, it was
found that TvL in aqueous [C2C1Im][OAc] and [Ch][Lys] require the presence of the
mediator ABTS to process the β-O-4' lignin model compound. Alternatively, TvL in
[DEA][HSO4] does not require ABTS to process with the compound. GPC and LC-MS
analysis of the reaction products indicates that GGE was initially polymerized into dimers,
trimers, and oligomers of the model compound. As the reaction proceeded, the ILs limited
this polymerization and appeared to promote the formation of small-molecular weight
products. TvL in AIL promoted both the depolymerization and polymerization of the
alkaline lignin, and the improved monomer yield could be attributed to the cleavage of the
β-O-4' linkages. Obtaining high lignin monomer yield is important from lignin valorization
standpoint. As the main focus of our study is to understand the interactions among
lignolytic enzymes, lignin and IL solvent, we were not set to optimize lignin
depolymerization; indeed, we picked relatively mild reaction conditions to better probe the
initial stage of the reaction and catch the product profile. Further optimizing the reaction
conditions would likely improve the yield of aromatic monomers from enzymatic lignin
depolymerization. Native plant lignin can be used as a substrate in future studies, in place
of the highly condensed kraft lignin used in this study. Additionally, other laccases or
laccase-engineering strategies can be employed to develop a biocatalyst with resistance to
higher ionic liquid concentrations or operating temperatures.

76
2.5 Conclusions
This study provides insight into the interactions between TvL and AILs. The effects
of three AILs with distinct properties on TvL activity, inhibition mechanism, and oxidative
capability were investigated. Results indicate that [DEA][HSO4] did not impede the ability
of TvL to oxidize ABTS, whereas [C2C1Im][OAc] and [Ch][Lys] were inhibitory at all but
the lowest concentrations. Enzyme kinetics and docking simulations suggest that
[C2C1Im][OAc] and [Ch][Lys] may bind at the entrance to the active site to interfere with
substrate interaction and catalysis, while [DEA][HSO4] likely does not. Both
depolymerized and polymerized products were observed when the TvL enzyme was applied
to a model GGE dimer and alkaline lignin in AILs. The reaction conditions still need to
be optimized by using different lignins and thermophilic laccases with better resistance to
AIL inhibition and higher temperatures. Collectively, this study provides a better
understanding of the interactions between a fungal laccase and AILs and serves as a
foundation for developing lignin valorization via biocatalysis in AILs.
2.6 Author Contributions
Joseph Stevens, Lalitendu Das, Justin Mobley, Shardrack Asare, Bert Lynn, David
Rodgers, and Jian Shi conceptualized the work, designed the experiments, and analyzed
the data. Shardrack Asare and Bert Lynn performed the LCMS experiments, generated the
figures, and wrote the experimental portions. Justin Mobley performed the HSQC NMR
experiments, generated the figures, and wrote the experimental portions. Joseph Stevens
performed all other experiments and wrote all other portions of the manuscript.

77
2.7 Acknowledgements
The authors acknowledge the National Science Foundation under Cooperative
Agreement No. 1355438 and 1632854 (2016 – 2020). This is a publication of the Kentucky
Agricultural Experiment Station and is published with the approval of the Director. This
work is also supported by the National Institute of Food and Agriculture, U.S. Department
of Agriculture, Hatch-Multistate project under accession number 1003563 (2018 – 2023).

78
2.8 Supplemental Information
Figure S 2-1. Hanes-Woolf curves of T. versicolor laccase in a) [C2C1Im][OAc], b)
[Ch][Lys], and c) [DEA][HSO4]
(a)
(b)
(c)

79
Figure S 2-2. Structure of proposed a) coupled GGE dimers, b) three coupled GGE dimers,
and c) addition of a G monomer unit to the S2b structure formed during GGE
polymerization reaction with T. versicolor laccase, ABTS, and AILs.
(a)
(c)
(b)

80
Figure S 2-3. a) FTIR spectra of alkaline lignin before and after treatment with laccase,
AILs, and ABTS (The red arrow is indicating the corresponding S-lignin peak); b) Thermal
profiles of alkaline lignin before and after treatment with laccase, AILs, and ABTS.
(a)
(b)

81
Figure S 2-4. Molecular weight distribution of a) solid-phase and b) liqui-phase alkaline
lignin after treatment with laccase, AILs, and ABTS
(a)
(b)

82
Figure S 2-5. 13C-1H (HSQC) spectra of alkyl (top) and aromatic (bottom) regions of a)
raw alkaline lignin (KL), b) alkaline lignin treated with laccase and ABTS, and alkaline
lignin treated with laccase and ABTS in aqueous ionic liquids: c) [Ch][Lys], d)
[DEA][HSO4], and e) [C2C1Im][OAc]. *The structures of side-chain linkages and lignin
compositional units were coded with colors corresponding to the cross peaks in the spectra
alkyl and aromatic regions, respectively.
(a) (b) (c) (d) (e)
Alk
yl r
egi
on
s A
rom
atic
re
gio
ns

83
CHAPTER 3. CHARACTERIZATION AND ENZYME ENGINEERING OF A HYPERTHERMOPHILIC
LACCASE TOWARD IMPROVING ITS ACTIVITY IN IONIC LIQUID1
3.1 Abstract
Ionic liquids (ILs) are organic salts molten at room temperature that can be used for
a wide variety of applications. Many ILs, such as 1-ethyl-3-methylimidazolium acetate
([C2C1Im][OAc]), have been shown to remove a significant fraction of the complex
biopolymer lignin from biomass during pretreatment. Valorizing lignin via biological
pathways (e.g. enzymes) holds promise but is limited by the low biocompatibility of many
ILs used for pretreatment. The discovery of thermostable enzymes and the application of
enzyme engineering techniques have yielded biocatalysts capable of withstanding high
concentrations of ILs. Converting lignin from a waste product to value-added chemicals
is vital to the success of future cellulosic biorefineries. To that end, we screened the activity
of the lignolytic enzyme laccase from a hyperthermophilic bacterium (Thermus
thermophilus) in aqueous [C2C1Im][OAc]. Despite the thermophilicity (Topt > 90 ºC) of
this laccase, significant activity loss (>50%) was observed in only 2% (w/v)
[C2C1Im][OAc]. Kinetics studies show that the IL can bind to the free enzyme and the
enzyme-substrate complex. Docking simulations suggest that the cation favors binding to
a region close to the active site. We then used a rational design strategy to improve the
activity of the laccase in [C2C1Im][OAc]. A total of 8 single amino acid mutations were
made; however, there were no significant improvements in the activity of the mutants in
[C2C1Im][OAc] compared to the wild type. The results of this study shed light on the
1This chapter in its whole has been accepted for a publication in Frontiers in Energy Research,
2020. DOI: 10.3389/fenrg.2020.00158.

84
complex nature of enzyme-IL interactions and the challenges faced when designing a
biological lignin valorization strategy.
3.2 Introduction
Lignin is a complex biopolymer that makes up ~30% of the terrestrial plant biomass
on earth 40, 244. It is made primarily of 3 phenylpropanoid subunits, syringyl (S), guaiacyl
(G) and p-hydroxyphenyl (H) 41, 43. These subunits are joined via complex networks of
ether and carbon-carbon linkages with the β-aryl ether (β-O-4), phenylcoumaran (β-5) and
pinoresinol (β-β) linkages being the most common 45, 278. Additionally, the ratio of subunits
is not homogeneous across all sources of biomass 44. Due to this monomeric and structural
heterogeneity, lignin is recalcitrant to biological and chemical depolymerization processes
for production of aromatic compounds at high selectivity and yield. Lignin is currently
generated as waste in large volumes in the paper and pulping industry and in cellulosic
biofuel production 1, 247. Converting lignin to value-added products will not only reduce
waste production, but will also add value to the paper and pulping industries and future
cellulosic biorefineries 3, 4.
Although applications of lignin are limited in its polymeric form, the phenolic
products derived from lignin can be used for production of fuels or as building blocks for
chemicals used in the plastics, food, or pharmaceutical industries 248, 249. Broadly speaking
current methods used for converting lignin to value-added products can be divided into
thermochemical, including pyrolysis, hydrogenolysis, and catalytic oxidation, and
biological, including the use of lignolytic enzymes or microbes to break down lignin 6, 279,
280. Biological methods utilize milder reaction conditions, with associated cost and safety

85
advantages, and can potentially improve the yield and selectivity of lignin breakdown
products due to the inherent efficiency of biocatalysts 6, 12. Several lignolytic pathways
have been identified in biomass-degrading fungi and bacteria. These include the heme
peroxidases (versatile, lignin, and manganese), laccases, and the recently discovered NAD
or glutathione-dependent enzymes from the soil bacterium Sphingobium SYK-6 13, 15, 281.
The application of these enzymes in an industrial setting has been limited by several
factors, including the high cost of enzyme production, the poor solubility of lignin in a
biocompatible solvent, and low selectivity of lignin-derived monomers by the biocatalyst
282, 283.
Many of the known lignin solvents, e.g. dimethyl sulfoxide (DMSO) or alkaline
solutions, reduce or eliminate enzyme activity 18, 19. Furthermore, these solvents require the
use of high temperatures for enhanced lignin solubilization 284, which also eliminates
enzyme activity. Therefore, a solvent that is both capable of solubilizing lignin and
supporting enzyme activity is required for developing biological lignin valorization
strategies. Ionic liquids are molten, organic salts (Tm < 100 ºC) that can adopt a variety of
properties by selecting the appropriate cation and anion 127, 128, 131, 133. Many
alkylimidazolium ILs, particularly 1-ethyl-3-methylimidazolium acetate
([C2C1Im][OAc]), have been extensively studied for use in biomass pretreatment at
relatively low temperatures 251-253. Due to the high cost of alkylimidazolium ILs, recent
efforts have focused on driving down the cost of ILs by using inexpensive, bio-derived
cations (e.g. choline and ammonium) and anions (e.g. amino acids and carboxylic acids)
and increasing the fraction of water during pretreatment 167, 285. ILs are favored over other
biomass pretreatment solvents due to their ability to reduce biomass recalcitrance, increase

86
yield of fermentable sugars from enzymatic hydrolysis, and solubilize high fractions (5 –
20%) of cellulose 171, 286, 287.
Laccases are a member of the superfamily of multi-copper oxidases (E.C. 1.10.3.2).
First discovered in the extract from the Japanese lacquer tree Toxicodendron vernicifluum,
laccases have since been identified in fungi, bacteria, and archaea 84, 85, 90, 91, 93. Laccases
differ from the other major lignolytic enzymes found in nature, the heme peroxidases, as
they are copper containing enzymes that utilize molecular oxygen, as opposed to stronger
oxidants, in their catalytic mechanism 71, 80, 86. The mild reaction conditions required for
laccase activity have made them attractive targets for use in a variety of biotechnological
applications, including lignin valorization 288. Additionally, laccases can oxidize
nonphenolic lignin compounds when coupled with small molecule mediator compounds,
like 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) or 1-
hydroxybenzotriazole (HBT) 97, 256, 289. Recently, some ILs, including those investigated
for use in biomass pretreatment have been found to be biocompatible with the activity of
laccases. Several alkylimidazolium ILs were found to be biocompatible with the activity
of a mesophilic fungal laccase from Trametes versicolor (TvL) at different concentrations
in water; over 80% of laccase activity remained in 50% 1-ethyl-3-methylimidazolium
ethylsulfate ([C2C1Im][EtSO4]) 193, for example. However, this IL was shown to be the
most biocompatible of the 4 aklylimidazolium ILs screened in the study. TvL activity was
reduced by 50% in less than 40% 1-butyl-3-methylimidazolium chloride ([C4C1Im][Cl]),
1-hexyl-3-methylimidazolium bromide ([C6C1Im][Cl]), and 1-decyl-3-methylimidazolium
chloride ([C10C1][Cl]) 290. Additionally, we showed in a previous study that the same
laccase loses >50% activity in as little as 10% (w/v) [C2C1Im][OAc] and 1% (w/v)

87
[Ch][Lys], far below the IL concentrations required for effective biomass pretreatment 291.
If laccases are to be used as biocatalysts for lignin valorization, they must be able to
withstand higher concentrations of ILs.
Recent strategies to improve enzyme activity and stability in ILs can be classified
into 3 categories: chemical, physical, and biotechnological modifications. By reducing the
ratio of positively charged to negatively charged surface residues via acetylation or
succinylation, the activity and stability of enzymes can be improved in ILs 202, 292.
Immobilization of laccase from Myceliophthora thermophila (MtL) on glyoxyl-agarose
beads improved enzyme stability in aqueous concentrations (0 – 75%) of [C2C1Im][EtSO4]
198. Single and multiple amino acid mutations have been used to improve the activity of
laccases in alkylimidazolium ILs 265, 293, 294. Recently, several thermotolerant organisms
have been found to produce laccases or laccase-like enzymes. Due to the ability of these
organisms to survive in extreme conditions of extreme temperature, pH, and salinity they
could prove to be useful sources of biocatalysts for industrial applications such as lignin
valorization in aqueous ILs 199. Bacillus subtilis produces a laccase-like enzyme (Topt > 70
ºC) that is capable of oxidizing canonical laccase substrates such as syringaldazine and
ABTS in the presence of several alkylimidazolium chloride ILs 91, 265. The laccase
produced by Thermus thermophilus (Topt > 90 ºC), is the most thermotolerant laccase
identified to date 93. However, there has been no work to characterize the behavior of the
T. thermophilus laccase in ILs.
In this study we sought to characterize the interactions between the IL
[C2C1Im][OAc] and the laccase produced by T. thermophilus (TtL). To do this, we first
screened the biocompatibility of [C2C1Im][OAc] with recombinantly produced TtL. We

88
then used enzyme kinetics and docking simulations to better understand how
[C2C1Im][OAc] interacts with TtL. Finally, we used a rational design approach to make a
series of single amino acid mutations in an effort to improve the activity of TtL in
[C2C1Im][OAc]. This study expands our understanding of the effects ILs can have on
laccases and highlights some of the challenges faced when designing a biological lignin
valorization strategy.
3.3 Materials and Methods
3.3.1 Materials
ABTS tablets were purchased from Thermo Fisher and the IL [C2C1Im][OAc] was
purchased from MilliporeSigma. The TtL sequence was codon optimized for E. coli
expression and inserted into pET32a(+) vector at NcoI and BamHI sites by GenScript
(Piscataway, NJ) to make the pET32aTtL expression plasmid. Mutant primers were
purchased from Eurogentec (Seraing, Belgium). The PfuUltra High-Fidelity DNA
Polymerase, buffer, and dNTP mixture were purchased from Agilent.
3.3.2 Expression of T. thermophilus Laccase
E. coli Rosetta (DE3) cells transformed with pET32aTtL were cultured in 1 L LB
broth containing ampicillin (50 µg/mL) and chloramphenicol (34 µg/ mL) with shaking at
37 ºC until the optical density at 600 nm reached 0.5. The laccase expression was induced
by adding 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) and the culture was
transferred to a 16 ºC shaker incubator overnight. Cells were collected by centrifugation
and resuspended in 40 mL of lysis buffer (50 mM NaH2PO4, 300 mM NaCl, pH 8.0) and
stored at -20 ºC until purification.

89
3.3.3 Purification and copper-induced laccase folding
The frozen cells were thawed and subsequently sonicated on ice. The cell debris
was removed by centrifugation at 15,000 rpm for 50 min at 8 ºC. The supernatant was
heated for 1 h at 60 ºC to precipitate any heat sensitive protein, centrifuged for 30 min at
15,000 rpm, then filtered and loaded onto a column with 2.5 mL cobalt resin equilibrated
with lysis buffer. The bound protein was washed with 15 mL lysis buffer followed by 5
mL wash buffer (50 mM NaH2PO4, 300 mM NaCl, 20 mM imidazole, pH 8.0) to remove
any protein bound to the column by non-specific binding. Finally, the protein was eluted
with 5 mL elution buffer (50 mM NaH2PO4, 300 mM NaCl, 50 mM imidazole, pH 8.0).
After purification, the native, copper-containing enzyme was produced according
to a previously used protocol 295. The eluted protein was dialyzed twice against 1 L of a
copper containing buffer (20 mM sodium acetate, 0.1 mM CuSO4, pH 6.0) for 12 h at 4 ºC.
The protein was then dialyzed twice against 1 L of a copper-free buffer (20 mM sodium
acetate, pH 4.5) for 12 h at 4 ºC to remove any remaining copper from the first dialysis
step. Precipitate was removed by centrifugation at 15,000 rpm for 20 min at 4 ºC. The
final protein concentration was determined by measuring the absorbance at 280 nm and
using the molar absorption coefficient Ɛ280 = 46,065 M-1 cm-1.
3.3.4 Biocompatibility screening
The biocompatibility of [C2C1Im][OAc] in aqueous solution (approximately 1-10%
w/v), was screened with recombinant TtL. To reduce the effect of pH, the IL solution was
adjusted to pH 4.5 using 1 M hydrochloric acid prior to testing. Activity was screened with
20 mM sodium acetate buffer (pH 4.5), 0.1 mM CuSO4, IL (0, 1, 2, 3, 4, or 10% w/v) and

90
2 mM ABTS in quartz cuvettes with a 1 cm path length. Absorbance readings were taken
continuously for 10 min at 61.5 ºC. Oxidation of ABTS in buffer and AILs without laccase
were measured as blanks. The relative activity of TtL in ILs relative to buffer was
calculated using equation 3-1.
𝑅𝑒𝑙𝑎𝑡𝑖𝑣𝑒 𝑎𝑐𝑡𝑖𝑣𝑖𝑡𝑦 = 𝐼𝑛𝑖𝑡𝑖𝑎𝑙 𝑉𝑒𝑙𝑜𝑐𝑖𝑡𝑦 𝑖𝑛 𝐼𝐿
𝐼𝑛𝑖𝑡𝑖𝑎𝑙 𝑉𝑒𝑙𝑜𝑐𝑖𝑡𝑦 𝑖𝑛 𝑏𝑢𝑓𝑓𝑒𝑟 (3-1)
3.3.5 Inhibition kinetics
Michaelis-Menten curves were generated for IL concentrations (0, 1, 2, 3, and 4%
w/v) by varying the concentration of ABTS (0.75 – 2.00 mM) and measuring initial
velocities. ABTS oxidation was measured using the same method as described in the
biocompatibility screening. The oxidized ABTS was quantified by measuring the
absorbance at 420 nm and using the absorbance extinction coefficient Ɛ420 = 36,000 M-1
cm-1 295. An activity unit was defined as 1 µmol ABTS oxidized in 1 min (µmol
ABTS/min). Initial velocity (V) as a function of ABTS concentration ([S]) was fit to the
Michaelis-Menten curve, shown in equation 3-2.
𝑉 = 𝑉𝑚𝑎𝑥∗[𝑆]
𝐾𝑚+[𝑆] (3-2)
The kinetic coefficients were calculated by plotting equation 3-2 in Sigma Plot and
calculating Km and Vmax using the ligand binding curve fitting algorithm.
3.3.6 Docking Simulations
The 3D structures and PDB files of the ligands were prepared in YASARA
Structure (YASARA Biosciences GmbH, Vienna, Austria) using the SMILES strings
obtained from PubChem. The PDBQT files of the ligands and TtL (PDBID: 2XU9) were

91
prepared with AutoDock (version 4.2.6, MGLTools, La Jolla, CA). Charges for the
coppers were added by manually editing the TtL PDBQT file. AutoGrid parameters were
as follows: space value of 0.375Å, (x, y, z) grid centered at (-28.148, -27.636, 14.016), and
grid size of 126 in all directions. AutoDock parameters were as follows: Lamarckian GA,
1,000 genetic algorithm runs, and 25,000,000 max eval size. [C2C1Im]+ and [OAc]- were
docked separately to both unbound TtL and TtL with ABTS bound to the active site. The
structure of TtL with ABTS in the active site was prepared in PyMol by aligning TtL with
B. subtilis CotA containing ABTS in the active site (PDBID: 1OF0).
3.3.7 Site-directed mutagenesis of TtL
Primer design and site-directed mutagenesis of TtL was carried out using the
QuikChange Site-Directed Mutagenesis Kit from Stratagene (La Jolla, CA, USA). Table
S1 lists the primers used in this study. The WT TtL plasmid (pET32aTtL) was used as the
template for all PCR reactions. The PCR reaction mixture (50 uL) contained 5 ng of DNA
template, 125 ng each of forward and reverse primers, 1 uL dNTP mix, and 2.5 U of
polymerase. Reactions were conducted using the following sequence heated for 30 seconds
at 95 ºC followed by 16 cycles of heating (95 ºC, 30 sec), annealing (55 ºC, 1 min), and
elongation (68 ºC, 7.3 min). Following PCR, DpnI restriction enzyme was added to digest
the methylated template plasmid. Plasmid sequences were verified by Eurofins Genomics
(Ebersberg, Germany). Expression, purification, and biocompatibility screening of TtL
mutants were carried out using the previously described protocols in sections 3.3.3 and
3.3.4.

92
3.4 Results
3.4.1 WT TtL Biocompatibility Screening
To determine the biocompatibility of [C2C1Im][OAc], we screened the activity of
TtL in a range of IL concentrations in water. Figure 3-1A shows the activity of TtL in
different concentrations of [C2C1Im][OAc] relative to the activity in buffer. A 50%
reduction of TtL initial velocity in buffer was seen in 2% [C2C1Im][OAc], in contrast with
recent studies of thermophilic enzymes in [C2C1Im][OAc]. A thermophilic cellulase (Topt
= 80 ºC) retained 40% initial activity in 20% [C2C1Im][OAc], while a hyperthermophilic
cellulase (Topt > 95 ºC) retained 90% initial activity in 20% [C2C1Im][OAc] 199. The range
of IL concentrations tested is relevant to previous studies investigating the effect of water
on biomass pretreatment with ILs. When studying the role of water during pretreatment
with [C2C1Im][OAc], it was found that pretreatment efficacy is reduced when more than
50% water is present 171.
Figure 3-1. Activity of the WT TtL in different concentrations of [C2C1Im][OAc] (A).
Michaelis-Menten curves of TtL in different concentrations of [C2C1Im][OAc] (B). Values
shown are mean with error bars representing the SD of 3 experiments.
There are several factors that mediate the IL’s effect on enzyme activity. In the
case of alkylimidazolium ILs, the ability of the anion to disrupt the water shell around the
[C2C1Im][OAc] (%w/v)
0 2 4 6 8 10
Re
lati
ve A
cti
vit
y
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
[ABTS] (mM)
0,0 0,5 1,0 1,5 2,0
Sp
ec
ific
Acti
vit
y (
U m
g-1
)
0,0
0,1
0,2
0,3
0,4
0% [C2C1Im][OAc]
1% [C2C1Im][OAc]
2% [C2C1Im][OAc]
3% [C2C1Im][OAc]
4% [C2C1Im][OAc]
Init
ial
Velo
cit
y (
U m
g-1
)
(A) (B)

93
protein (chaotropicity) and the alkyl chain length of the cation are thought to be largely
responsible for the IL biocompatibility. The activity of TvL was relatively unaffected by
50% 1-ethyl-3-methylimidazolium ethyl sulfate ([C2C1Im][EtSO4]), but was severely
inhibited in ILs with longer alkyl chain length cations ([C4C1Im]+, [C6C1Im]+, and
[C10C1Im]+) and more chaotropic anions (Cl- and Br-) 193. An increase in the alkyl chain
length of the alkylimidazolium chloride ([CnC1Im][Cl]) ILs (C2 -> C4 -> C6) decreased the
activity of MtL, followed by a greater decrease in activity when the alkyl chain length was
further increased (C6 -> C10) 269. Therefore, the short alkyl chain length [C2C1Im]+ cation
does not play a significant role in TtL inhibition. Rather, it is the chaotropic [OAc]- anion
that causes the inhibition in low IL concentrations.
3.4.2 Enzyme Kinetics
To further understand the interaction between [C2C1Im][OAc] and TtL, we carried
out a more in depth investigated of enzyme kinetics using ABTS as a substrate (Figure 3-
1B). With increasing concentrations of [C2C1Im][OAc], Km increased while the kcat
decreased (Table 3-1). This suggests that [C2C1Im][OAc] is a mixed competitive and
noncompetitive inhibitor of TtL. Such behavior could be the result of [C2C1Im][OAc]
interaction remote from the active site that decreases both substrate binding and catalysis,
or the IL could be binding both at the active site and a remote location. As indicated in
Table 3-1, kcat was lowered from 0.42 s-1 in 0% [C2C1Im][OAc] to 0.29 s-1 in 1%
[C2C1Im][OAc]. However, a smaller reduction in kcat was observed between 1%
[C2C1Im][OAc] (0.29 s-1) and 4% [C2C1Im][OAc] (0.20 s-1). In contrast, Km steadily
increased between 0% [C2C1Im][OAc] (0.52 mM ABTS) and 4% [C2C1Im][OAc] (1.90
mM ABTS). The different Km and kcat variations with IL concentration support the

94
presence of 2 major binding sites. The first is a high affinity noncompetitive binding site
that lowers the enzyme activity and is almost fully occupied in 1% [C2C1Im][OAc]. The
second is a low affinity competitive binding site that likely overlaps with the substrate
binding site, which lowers the substrate affinity of the enzyme.
Table 3-1. Kinetic constants of the WT TtL in different concentrations of [C2C1Im][OAc].
Values shown are mean ± standard error (SE) of 3 experiments.
[C2C1Im][OAc]
(%w/v)
Km
(mM ABTS)
Vmax
(U mg TtL-1)
kcat
(s-1)
kcat/Km
(s-1 mM-1)
0% [C2C1Im][OAc] 0.52 ± 0.05 0.34 ± 0.01 0.41 ± 0.01 0.78
1% [C2C1Im][OAc] 0.76 ± 0.08 0.24 ± 0.01 0.29 ± 0.01 0.38
2% [C2C1Im][OAc] 0.96 ± 0.10 0.22 ± 0.01 0.27 ± 0.01 0.28
3% [C2C1Im][OAc] 1.34 ± 0.30 0.20 ± 0.02 0.24 ± 0.03 0.17
4% [C2C1Im][OAc] 1.90 ± 0.33 0.17 ± 0.02 0.20 ± 0.02 0.11
Previous kinetics work with laccases and ILs have shown that ILs can affect both
Km and kcat of several laccases. Several [C2C1Im]+ ILs lowered the activity and increased
the substrate affinity of MtL by uncompetitive inhibition 191. In contrast, the IL
[C2C1Im][Cl] was found to be a competitive inhibitor of the same laccase 269. The activity
of TvL and MtL are increased by the ILs choline dihydrogen phosphate ([Ch][H2PO4]) and
[C2C1Im][EtSO4], respectively; enzyme kinetics suggest these ILs decrease substrate
affinity (increase Km) but increase enzyme activity (increase Vmax) via an uncompetitive
inhibition mechanism 150, 198.
3.4.3 Docking simulations
To gain insight into the possible binding locations of [C2C1Im][OAc] on the surface
of TtL, we performed docking simulations with AutoDock. Figures 3-2A and 3-2B show

95
the most populated IL docking poses to the surfaces of [E] and [E][S], respectively, after
1,000 docking simulations with AutoDock. Neither the cation nor the anion binds close
enough to the coppers to suggest that the IL disrupts the residues that coordinate and
stabilize the coppers. The binding locations of the cation and the anion to the surface of
TtL is affected by the presence of ABTS in the active site. The binding location of the
cation to [E] (Fig. 3-2A) appears to overlap with ABTS bound in the active site, leading to
the displacement of the cation when docked to [E][S] (Fig. 3-2B). This suggests that the
cation directly competes with the ABTS for the active site which leads to the increased Km
seen in the enzyme kinetics. Similarly, there is a shift in the anion binding site when
docked to [E] vs [E][S]; however, the anion binding location does not overlap with the
ABTS. This suggests that the anion plays a role decreasing kcat but may also contribute to
the change in Km, since substrate affinity would be decreased by the cost of displacing the
anion.
Figure 3-2. Top docking locations from AutoDock of [C2C1Im][OAc] to the surface of [E]
(A) and [E][S] (B). The IL is shown as magenta spheres, the coppers are shown as brown
spheres, and ABTS is shown as red sticks.
Docking results from this work are consistent with previous simulations examining
docking locations of several ILs to TvL and docking of alkylimidazolium cations to MtL.
(A) (B)

96
Highly populated binding sites were found close to the T1 copper when [C2C1Im][OAc],
[Ch][Lys], and diethylamine hydrogen sulfate ([DEA][HSO4]) were docked to TvL 291.
When docked to MtL, short alkyl chain length cations (C2 – C6) diffused into the active site
whereas the long chain length cations (C8 – C10) interacted with leucine residues around
the active site 269. However, the activity of MtL is increased in the presence of
[C2C1Im][EtSO4] 198, therefore IL docking close to the T1 copper does not imply the IL
will inhibit laccase activity.
3.4.4 Rational Design of TtL
The mutations were chosen based on several approaches aimed at improving
enzyme activity and stability in ILs. Recent directed evolution and semi-rational design
studies have identified the L1 loop in TvL as a key target for laccase engineering. A triple
mutant targeting the alanines on the L1 loop in TvL, A310D/A312P/A318R, retained 21%
initial activity in 40% [C2C1Im][EtSO4], compared to 0% remaining activity for the WT
laccase 294. The L1 loop in TtL presents 3 alanines (Ala320, Ala332, and Ala341) (Fig. S3-
1A). The mutations of these alanines were chosen based on recent chemical modifications
of enzymes in ILs. Reducing the ratio of positively charged amines to negatively charged
acids on the enzyme surface by acetylation or succinylation improved their activity and
stability in ILs 202, 292. For this reason, the Ala320, Ala332, and Ala341 from TtL L1 loop
were mutated to: A320D/E, A332D/E, and A341D/E. Although we could have selected
additional mutations based on previous studies with TvL (e.g. A320P or A332R), we chose
the mutations to aspartate and glutamate in order to limit our design space. Additionally,
this strategy of engineering the L1 loop is based on studies with a fungal laccase; however,
we felt it appropriate to apply this strategy to the bacterial laccase used in this study. We

97
also made mutations to Glu170 based on previous work improving the activity of a laccase-
like enzyme from B. subtilis in ILs. The catalytic efficiency (kcat/Km) of B. subtilis HR03
mutants E188F and E188Y was increased compared to the WT in several [CnC1Im][Cl] ILs
265. Through sequential alignment, we identified a homologous glutamate residue in TtL,
Glu170 (Fig. S3-1B), which was mutated to both tyrosine (E170Y) and phenylalanine
(E170F).
Aiming to improve the activity of TtL in [C2C1Im][OAc], we made 8 single amino
acid mutations to TtL. With the exception of the E170Y mutant at 0% IL, none of the
mutations significantly improved the activity of TtL in buffer or [C2C1Im][OAc] when
compared to WT (Table 3-2). The A320 mutations significantly decreased the activity in
buffer and some IL concentrations, as did the E170F mutation at 2% [C2C1Im][OAc]. There
were no significant differences from WT when normalizing each variant to its activity in
0% IL, except for the lower activity of the E170Y mutant in 2% [C2C1Im][OAc], indicative
of its generally greater sensitivity to low [C2C1Im][OAc] concentrations.
Table 3-2. Activity of WT and mutants relative to activity in buffer in different
concentrations of [C2C1Im][OAc]. Values shown are mean ± standard deviation (SD) of 3
experiments1.
Relative to Activity in Buffer
IL (%w/v) WT A320D A320E A332D A332E A341D A341E E170F E170Y
0 1.00a 1.00a 1.00a 1.00a 1.00a 1.00a 1.00a 1.00a 1.00a
1 0.70 ± 0.04a 0.72 ± 0.07a 0.75 ± 0.05a 0.70 ± 0.03a 0.64 ± 0.01a 0.72 ± 0.04a 0.70 ± 0.03a 0.71 ± 0.01a 0.58 ± 0.04a
2 0.49 ± 0.03a 0.42 ± 0.05a 0.44 ± 0.05a 0.45 ± 0.02a 0.42 ± 0.04a 0.45 ± 0.05a 0.43 ± 0.03a 0.39 ± 0.02a 0.33 ± 0.03b
3 0.38 ± 0.01a 0.41 ± 0.05a 0.42 ± 0.04a 0.40 ± 0.03a 0.40 ± 0.04a 0.38 ± 0.03a 0.35 ± 0.04a 0.37 ± 0.04a 0.29 ± 0.01a
4 0.28 ± 0.06a 0.26 ± 0.04a 0.31 ± 0.06a 0.32 ± 0.01a 0.31 ± 0.05a 0.28 ± 0.04a 0.28 ± 0.04a 0.32 ± 0.05a 0.22 ± 0.02a
10 0.09 ± 0.02a 0.09 ± 0.02a 0.09 ± 0.02a 0.09 ± 0.00a 0.09 ± 0.03a 0.10 ± 0.01a 0.09 ± 0.01a 0.09 ± 0.02a 0.11 ± 0.00a
Relative to WT Activity in Buffer
0 1.00a 0.85 ± 0.00b 0.73 ± 0.03b 0.96 ± 0.01a 0.94 ± 0.06a 1.00 ± 0.05a 1.07 ± 0.04a 0.94 ± 0.04a 1.23 ± 0.04b
1 0.70 ± 0.04a 0.58 ± 0.05a 0.52 ± 0.06b 0.67 ± 0.02a 0.63 ± 0.04a 0.72 ± 0.04a 0.75 ± 0.03a 0.68 ± 0.05a 0.71 ± 0.04a
2 0.49 ± 0.03a 0.33 ± 0.04b 0.32 ± 0.03b 0.43 ± 0.02a 0.40 ± 0.04a 0.45 ± 0.05a 0.46 ± 0.03a 0.37 ± 0.02b 0.41 ± 0.04a
3 0.38 ± 0.01a 0.33 ± 0.04a 0.30 ± 0.03a 0.38 ± 0.03a 0.37 ± 0.04a 0.38 ± 0.03a 0.38 ± 0.04a 0.35 ± 0.04a 0.36 ± 0.01a
4 0.28 ± 0.06a 0.20 ± 0.03a 0.22 ± 0.04a 0.28 ± 0.04a 0.29 ± 0.04a 0.29 ± 0.04a 0.30 ± 0.04a 0.30 ± 0.05a 0.27 ± 0.02a
10 0.09 ± 0.02a 0.06 ± 0.00a 0.07 ± 0.02a 0.10 ± 0.02a 0.09 ± 0.03a 0.10 ± 0.01a 0.10 ± 0.01a 0.09 ± 0.02a 0.12 ± 0.01a

98
1The superscript letters “ab” indicate the significance levels (α = 0.05) of the means compared to the WT
means
Despite our results showing that engineering the L1 loop of TtL with single amino
acid mutations does not increase activity in ILs, it is important to note that single mutations
can be deleterious as it was observed for the A318V mutant of TvL, which decreased
activity in ILs relative to WT 296. It is only when multiple amino acid mutations are made
to the L1 loop that the laccase activity increased in ILs 294. Additionally, the increase in
the catalytic efficiency of the HR03 mutants was attributed to increased π – π and π – anion
interactions between the mutants and several aspartate and tyrosine residues present on the
enzyme surface 265. Figure S3-2 shows there are relatively fewer aromatic and charged
residues surrounding Glu170 in TtL that could help stabilize the E170Y/F mutants.
3.5 Discussion
In this study we showed that unlike most thermophilic enzymes the activity of the
hyperthermophilic, bacterial laccase from Thermus thermophilus is sensitive to the
presence of the IL [C2C1Im][OAc]. Through in vitro and in silico techniques, we provide
insightful information about 1) the inhibition of TtL by [C2C1Im][OAc] and 2) rational
design of TtL to improve activity in [C2C1Im][OAc]. The biocompatibility screening
showed that >50% initial enzyme velocity is lost in just 2% [C2C1Im][OAc]. The docking
simulations, along with the enzyme kinetics results, suggest that [C2C1Im][OAc] inhibits
the activity of TtL by interfering with ABTS binding to the active site (competitive
inhibition) and interfering with the ABTS oxidation once it is bound (uncompetitive
inhibition). We made several single amino acid mutations to TtL with the aim of improving
the activity in [C2C1Im][OAc]. These mutations were made based on previous studies that
focused on engineering the L1 loop in fungal laccases as well as previous work that

99
improved the activity of a bacterial laccase in ILs. While the E170Y mutant displayed
significantly higher activity in buffer, none of the mutants had significantly higher levels
of activity in [C2C1Im][OAc] when compared to the WT enzyme. We have shown that
thermophilicity alone does not indicate whether or not an enzyme will have high activity
in ILs. Future studies might aim to better understand the mechanism underlying inhibition
of TtL by [C2C1Im][OAc] or other ILs using molecular dynamics (MD) simulations or co-
crystallizing TtL with the IL, along with employing other techniques (e.g. multiple amino
acid mutations, chemical modifications, immobilization, computational design) to improve
TtL activity in ILs.
3.6 Author Contributions
Joseph Stevens, David Rodgers, Claire Dumon, and Jian Shi conceptualized the
work, designed the experiments, and analyzed the data. Joseph Stevens conducted the
experiments and wrote the manuscript.
3.7 Acknowledgements
The authors acknowledge the support from the National Science Foundation under
Cooperative Agreement No. 1355438 and 1632854 (2016 – 2020) and the National
Institute of Food and Agriculture, U.S. Department of Agriculture, Hatch-Multistate
project under accession number 1018315 (2018 – 20203). This material is based upon
research supported by the Chateaubriand Fellowship of the Office for Science &
Technology of the Embassy of France in the United States (2019).

100
3.8 Supplemental Information
Table S 3-1. List of primer names and sequences for making the mutants in this study.
Mutated nucleotides are underlined.
Oligoname Sequence (5’ -› 3’) Position
A320D_Fo gaa acc ctg ctg tat ctg att gat ccg aaa aac ccg aaa cc 320
A320D_Rev gg ttt cgg gtt ttt cgg atc aat cag ata cag cag ggt ttc 320
A320E_Fo gaa acc ctg ctg tat ctg att gaa ccg aaa aac ccg aaa cc 320
A320E_Rev gg ttt cgg gtt ttt cgg ttc aat cag ata cag cag ggt ttc 320
A332D_Fo ctg ccg ctg ccg aag gat ctg agc ccg ttc cc 332
A332D_Rev gg aaa cgg gct cag atc ttt cgg cag cgg cag 332
A332E_Fo ctg ccg ctg ccg aaa gaa ctg agc ccg ttt cc 332
A332E_Rev gg aaa cgg gct cag ttc ttt cgg cag cgg cag 332
A341D_Fo g ttc ccg acc ctg ccg gat ccg gtg gtg ac 341
A341D_Rev gt cac cac cgg atc cgg cag ggt cgg aaa c 341
A341E_Fo g ttt ccg acc ctg ccg gaa ccg gtg gtg ac 341
A341E_Rev gt cac cac cgg ttc cgg cag ggt cgg aaa c 341
E170F_Fo gaa ctg cgc gaa gcg ttt gaa cat ctg ctg gtg ctg aag 170
E170F_Rev ctt cag cac cag cag atg ttc aaa cgc ttc gcg cag ctc 170
E170Y_Fo gag ctg cgt gaa gcg tat gaa cac ctg ctg gtg c 170
E170Y_Rev g cac cag cag gtg ttc ata cgc ttc acg cag ctc 170

101
Figure S 3-1. Crystal structures of TtL (PDBID: 2XU9, left) and TvL (PDBID: 1GYC,
right) showing domain 1 (green), domain 2 (orange), domain 3 (magenta), and the L1 loop
(cyan) connecting domains 2 and 3 (A). Sequence alignment of TtL and B. subtilis CotA,
homologous Glutamate residues are highlighted (B).
(A)
(B)

102
Figure S 3-2. Aromatic and anionic residues around E170 in TtL (A) and E188 in B. subtilis
CotA (B). Glutamates targeted for mutations are shown in cyan while all other residues
are shown in green. All residues shown are within 5Å of the respective Glutamate.
GLU168
GLU170
GLU171
HIS172 GLU246
TYR250
TYR189
GLU188
ASP187
ASP190(B) (A)

103
Figure S 3-3. Codon optimized sequence of TtL (NCBI Accession Number: I7AL37)

104
CHAPTER 4. MODIFYING SURFACE CHARGES OF A THERMOPHILIC LACCASE TOWARDS
IMPROVING ACTIVITY AND STABILITY IN AQUEOUS IONIC LIQUID
4.1 Abstract
The multicopper oxidase enzyme laccase holds great potential to be used for
biological lignin valorization alongside a biocompatible ionic liquid. However, the ionic
liquid concentrations required for biomass pretreatment severely inhibit laccase activity.
Due to their ability to function in extreme conditions, many thermophilic enzymes have
found use in industrial applications. The thermophilic fungal laccase from Myceliophthora
thermophila was found to retain high levels of activity in [C2C1Im][EtSO4], making it a
desirable biocatalyst to be used for lignin valorization. In contrast to [C2C1Im][EtSO4], the
biocompatibility of [C2C1Im][OAC] with the laccase was markedly lower. Severe
inhibition of laccase activity was observed in 15% [C2C1Im][OAc]. Based on previous
studies, the enzyme surface charges were modified via acetylation, succinylation,
cationization, or neutralization. These modifications did not improve laccase activity or
stability in [C2C1Im][OAc] in contrast with the previous studies, although it is unknown if
all modifications were made to the enzyme. Docking simulations show that the IL docks
close to the T1 catalytic copper, likely interfering with substrate binding. Although
additional docking locations for [OAc]- are observed after making enzyme modifications,
it does not appear that these locations play a role in the inhibition of enzyme activity. The
results of this study could guide future enzyme engineering efforts by showing that the
inhibition mechanism of [C2C1Im][OAc] towards M. thermophila laccase is not dependent
upon the IL interacting with the enzyme surface.

105
4.2 Introduction
Lignin is among the most abundant terrestrial biopolymers that makes up ~30% of
the weight of biomass 40, 244. It is primarily made of 3 phenylpropanoid subunits: p-
hydroxyphenyl (H), guaiacyl (G), and syringyl (S) that can be linked together via several
condensed (C-C) or ether (C-O) bonds, with the phenylcoumaran (β – 5), β – aryl ether (β
– O – 4), and pinoresinol (β – β) being the most common 39, 41, 43, 45. Additionally, the ratio
of subunits present in the lignin network is dependent upon the source of the biomass 44.
As a result of lignin’s structural and monomeric heterogeneity, thermochemical and
biological methods achieve poor selectivity and yield of breakdown products. Lignin is
currently generated in high volumes as a waste product by the paper and pulping industry
and during production of cellulosic biofuels 1, 297. Converting lignin from a waste product
to value added chemicals via new technologies will not only reduce waste lignin
accumulation but also add value to the paper & pulping and future cellulosic biorefineries
3, 4. While the applications of polymeric lignin are limited, the phenolic products obtained
from lignin deconstruction can be used as fuels or precursors for chemicals in the food,
pharmaceutical, or plastic industries; many of these fuels and chemicals are currently
derived from petroleum 248, 249.
Current methods for lignin deconstruction can be divided into thermochemical (e.g.
high temperature pyrolysis, catalytic oxidation, and hydrogenolysis) and biological (e.g.
lignin degrading enzymes [LDEs] or microbes) 6, 279, 280. Much of the research into the
thermochemical methods has focused on improving the catalyst performance at high
temperatures and improving the selectivity of lignin-derived products 255, 298, 299. The use
of LDEs in biological lignin deconstruction strategies could enable lignin deconstruction

106
at lower temperatures and improve product selectivity due to the inherent selectivity and
efficiency of biocatalysts 6. Many LDEs such as the peroxidases (lignin, manganese, and
versatile) and laccases have been discovered in bacteria, plant, and fungi 85, 91, 93.
Additionally, catabolic lignin pathways in soil bacteria, such as Sphingobium sp. SYK-6
that catabolizes lignin using NAD or glutathione dependent enzymes have recently been
identified 281. The application of LDEs for lignin deconstruction has been limited by a
number of factors, chief among these being the high cost of enzyme production and the
low yield of products due to the poor solubility of lignin in solvents biocompatible with
LDE activity 282, 283.
Many of the known lignin solvents (e.g. dimethyl sulfoxide [DMSO] and alkaline
solutions) reduce or altogether eliminate enzyme activity 18, 19. Additionally, these solvents
require high temperatures to facilitate the complete dissolution of plant derived lignin,
leading to a further loss in enzyme activity 284. Therefore, a solvent system that is capable
of solubilizing lignin at low temperatures and mild pH is desirable. Ionic liquids (ILs) are
molten, organic salts liquid at temperatures <100oC, the properties of which can be tuned
by selecting the appropriate cation and anion 127, 128, 131, 144. The alkylimidazolium ILs, such
as 1-ethyl-3-methylimidazolium acetate ([C2C1Im][OAc]) and 1-butyl-3-
methylimidazolium chloride ([C4C1Im][Cl]), have been the focus of numerous studies on
biomass pretreatment at relatively low temperatures 251-253. Due to the high cost associated
with using these ILs, recent efforts have focused on developing low cost ILs from
bioderived cations (e.g. choline and ammonium) and anions (e.g. carboxylic acids and
amino acids) that can be used in low concentrations during pretreatment 167, 285, 300. One

107
example is cholinium lysinate ([Ch][Lys]) which, at only 10% (w/v) in water, was shown
to remove 80% of lignin from genetically engineered switchgrass during pretreatment 254.
Laccases are a member of the multicopper oxidase superfamily of enzymes (E.C.
1.10.3.2). They were first discovered in extract from the Japanese lacquer tree
(Toxicodendron vernicifluum) in 1883 84. Laccases differ from the other group of lignolytic
enzymes, the heme peroxidases, in that they are copper-containing enzymes that do not
require the presence of a strong oxidant, such as hydrogen peroxide, for the reaction
mechanism 71, 86. The substrate is oxidized at the T1 catalytic copper via 4-electron
removal, after which the electrons are shuttled 13Å to the trinuclear copper cluster where
molecular oxygen is reduced to water 301. The T1 catalytic copper and the trinuclear
coppers (a T2 and 2 T3) are coordinated by 4 conserved HXH motifs; X = cysteine in the
case of the T1 copper 86. The T1 copper is also coordinated by a methionine or
leucine/phenylalanine in bacterial/plant and fungal laccases, respectively 302. Figure 4-1
shows the conserved copper coordinating motifs in the sequences of laccases from different
organisms.

108
Figure 4-1. Clustal Omega sequence alignment of laccases from a bacterium (T.
thermophilus – 2XU9), a plant (T. vernicifluum - Plant), and fungi (T. versicolor – 1GYC,
M. thermophila – 6F5K). Conserved HXH motifs and T1-coordinating
methionine/phenylalanine/leucine are highlighted with thick blue bars.
Laccases are also capable of oxidizing nonphenolic lignin compounds when used
alongside a small molecule mediator, such as 1-hydroxybenzotriazole (HBT) or 2,2’-azino-
bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) 97, 256, 289. The relatively mild reaction
conditions required for laccase activity along with the oxidation of mediator compounds
make laccases a desirable biocatalyst for use in many biotechnological applications,
including biological lignin deconstruction strategies 222, 288. Many ILs have been shown to
be biocompatible with the activity of laccases at low concentrations in water. When Galai
et al. investigated the effect of 56 ILs on the activity of T. versicolor laccase for dye
decoloring applications, they found that 13 of the ILs increased the activity of the laccase;
10 mM choline dihydrogen phosphate ([Ch][H2PO4]) increased activity by 451% 150. The

109
initial activity of laccase from the white rot fungus Trametes versicolor was only reduced
by 20% in a reaction mixture containing 50% (v/v) 1-ethyl-3-methylimidazolium
ethylsulfate ([C2C1Im][EtSO4]) 290. However, other alkylimidazolium ILs have been
found to be less biocompatible with laccase activity. The initial activity of T. versicolor
laccase was reduced by 50% in reaction mixtures containing just 3, 20, 40, and 5% (v/v)
of [C2C1Im][OAc], [C4C1Im][Cl], 1-hexyl-3-methylimidazolium bromide ([C6C1Im][Br]),
and 1-decyl-3-methylimidazolium chloride ([C10C1Im][Cl]), respectively 290, 291.
Recent studies have shown that ILs are more biocompatible with thermophilic
enzymes compared to mesophilic enzymes. Hydrolysis of carboxymethylcellulose (CMC)
by hyperthermophilic (Topt > 95oC) and thermophilic (Topt = 80oC) cellulases was
minimally impacted in 0 – 20% (v/v) [C2C1Im][OAc] while a mesophilic cellulase (Topt =
37oC) was unable to hydrolyze CMC in only 10% (v/v) [C2C1Im][OAc] 199. The activity
of laccase from the thermophilic fungi Myceliophthora thermophila (MtL) is increased 3-
fold in 25% (v/v) [C2C1Im][EtSO4] 198. The bacteria Bacillus subtilis produces a laccase-
like spore coat protein (CotA) that can oxidize canonical laccase substrates ABTS and
syringaldazine (SGZ) in the presence of several alkylimidazolium chloride ([CnC1Im][Cl])
ILs; [CnC1Im][Cl] ILs have been shown to severely inhibit the activity of a mesophilic
laccase at low concentrations 91, 265. However, we showed in a previous study that the most
thermophilic laccase identified to date, produced by the hyperthermophilic bacterium
Thermus thermophilus, is highly sensitive to low concentrations (<10% v/v) of
[C2C1Im][OAc], suggesting that enzyme thermophilicity alone does not guarantee high
activity in ILs 93.

110
In this study we sought to determine the biocompatibility of aqueous
[C2C1Im][OAc] with MtL. To do this we first screened the activity of MtL in different
concentrations of [C2C1Im][OAc] up to 50% in water. We also measured the effect of low
concentrations of [C2C1Im][OAc] on the thermostability of MtL. Following the initial
biocompatibility and stability screenings, we made several surface charge modifications
with the aim of improving MtL activity and stability in [C2C1Im][OAc]. We also used
molecular docking simulations to understand how [C2C1Im][OAc] docks to the surface of
MtL and the 4 charge variants we produced. The results of this study expand our
understanding of laccase-IL interactions and the difficulties faced when trying to improve
enzyme activity in ILs.
4.3 Methods and Materials
4.3.1 Materials
Laccase from M. thermophila expressed in Aspergillus oryzae (Batch #
OMN07029) was kindly provided by Novozymes (Bagsværd, Denmark). The 1-Step™
ABTS Substrate Solution was purchased from ThermoFisher Scientific (Waltham, MA).
All other reagents, including the IL [C2C1Im][OAc], were purchased from MilliporeSigma
(St. Louis, MO).
4.3.2 Surface Charge Modifications
MtL was purified by sequentially spin filtering with 100 kDa and 30 kDa MWCO
filters at 8oC prior to surface charge modifications. Surface charges were modified
according to previously reported protocols 202, 292. Cationized MtL was produced by
modification with 0.5 M ethylenediamine hydrochloride in 200 mM MES buffer (pH 4.5)

111
containing a 20:1 molar ratio of the crosslinking reagent N-(3-dimethylaminopropyl)-N’-
ethylcarbodiimide (EDC)-to-acid sites for 2.5 h at room temperature; neutralized MtL was
similarly produced using diethylamine in place of ethylenediamine hydrochloride.
Succinylated MtL was produced by addition of 4 aliquots of succinic anhydride to the
enzyme in 1 M sodium carbonate buffer (pH 8.5) over 3 h at room temperature. Acetylated
MtL was produced by addition of 3 aliquots of acetic anhydride to the enzyme in 100 mM
sodium phosphate buffer (pH 7.0) over 1.5 h at room temperature. The final molar ratio of
acetic or succinic anhydride to enzyme containing primary amines was approximately 30:1.
Excess reagent was removed with 30 kDa MWCO spin filters at 4oC and the enzymes were
stored in 100 mM sodium phosphate buffer (pH 7.0) prior to experiments. Protein content
was measured using the Bradford assay.
4.3.3 Biocompatibility Screening
The biocompatibility of [C2C1Im][OAc] in aqueous solution (approximately 1-50%
w/v), was screened with all MtL variants. To reduce the effect of pH, the IL solution was
adjusted to pH 4.5 using 1 M hydrochloric acid prior to testing. Activity was screened with
50 mM citrate/100 mM phosphate buffer (pH 4.5), IL (0, 1, 2, 3, 4, or 10% w/v) and 50 μL
ABTS solution in clear, flat bottom, 96-well Costar assay plates (Corning Inc., Kennebunk,
ME). Absorbance readings were taken every 15 seconds for 10 min at 40 ºC in a
SpectraMax M2 plate reader (Molecular Devices, Sunnyvale, CA). Plates were shaken for
3 seconds prior to each reading to ensure a homogeneous well solution. Oxidation of ABTS
in buffer and AIL without laccase were measured as blanks. The activity of MtL in ILs
relative to buffer was calculated using equation 4-1.

112
𝑅𝑒𝑙𝑎𝑡𝑖𝑣𝑒 𝑎𝑐𝑡𝑖𝑣𝑖𝑡𝑦 = 𝐼𝑛𝑖𝑡𝑖𝑎𝑙 𝑉𝑒𝑙𝑜𝑐𝑖𝑡𝑦 𝑖𝑛 𝐼𝐿
𝐼𝑛𝑖𝑡𝑖𝑎𝑙 𝑉𝑒𝑙𝑜𝑐𝑖𝑡𝑦 𝑖𝑛 𝑏𝑢𝑓𝑓𝑒𝑟 (4-1)
4.3.4 MtL Thermostability
To measure the thermostability of MtL, all charge variants were incubated at 40oC
in 0, 2.5, or 5% [C2C1Im][OAc] and 50 mM citrate/100 mM phosphate buffer, pH 4.5. At
each time interval (0, 30, 60, 120, 180, and 240 min) aliquots were removed and placed on
ice prior to measuring residual enzyme activity. Activity was screened using the same
method in 4.3.3. Biocompatibility Screening. The residual activity of MtL and the charge
variants after incubation was calculated using equation 4-2.
𝑅𝑒𝑠𝑖𝑑𝑢𝑎𝑙 𝑎𝑐𝑡𝑖𝑣𝑖𝑡𝑦 = 𝐼𝑛𝑖𝑡𝑖𝑎𝑙 𝑉𝑒𝑙𝑜𝑐𝑖𝑡𝑦 𝑎𝑓𝑡𝑒𝑟 𝑖𝑛𝑐𝑢𝑏𝑎𝑡𝑖𝑜𝑛
𝐼𝑛𝑖𝑡𝑖𝑎𝑙 𝑉𝑒𝑙𝑜𝑐𝑖𝑡𝑦 𝑎𝑓𝑡𝑒𝑟 0 min 𝑖𝑛𝑐𝑢𝑏𝑎𝑡𝑖𝑜𝑛 (4-2)
4.3.5 Docking Simulations
The 3D structures and PDB files of the ligands were prepared in YASARA
Structure (YASARA Biosciences GmbH, Vienna, Austria) using the SMILES strings
obtained from PubChem. The PDBQT files of the ligands and MtL (PDBID: 6F5K) were
prepared with AutoDock (version 4.2.6, MGLTools, La Jolla, CA). Charges for the
coppers were added by manually editing the MtL PDBQT file. AutoGrid parameters were
as follows: space value of 0.375Å, (x, y, z) grid centered at (4.840, 28.290, -18.567), and
grid size of 126 in all directions. AutoDock parameters were as follows: Lamarckian GA,
100 genetic algorithm runs, and 25,000,000 max eval size. Docking results were visualized
using PyMol (Schrodinger LLC, New York, NY). To simulate the surface charge
modifications, target surface residues were mutated in PyMol. These mutations are shown
in Table 4-2.

113
Table 4-1. Summary of mutations to used mimic surface charge modifications in PyMol
Charge
Variant
Modification
Chemistry
Target
Residues
PyMol
Mutations
Unmodified
N/A
N/A N/A
Acetylated
Primary
Amines
(Lys)
Lys ->
Ala
Neutralized
Carboxylic
Acid
(Asp or
Glu)
Asp ->
Ala
Glu ->
Ala
Succinylated
Primary
Amines
(Lys)
Lys ->
Asp
Cationized
Carboxylic
Acid
(Asp or
Glu)
Asp ->
Lys
Glu ->
Lys
4.4 Results and Discussion
4.4.1 Biocompatibility Screening
To understand the biocompatibility of [C2C1Im][OAc], we measured the activity of
MtL and all of the charged variants in aqueous solutions (0 – 50% w/v) of [C2C1Im][OAc]
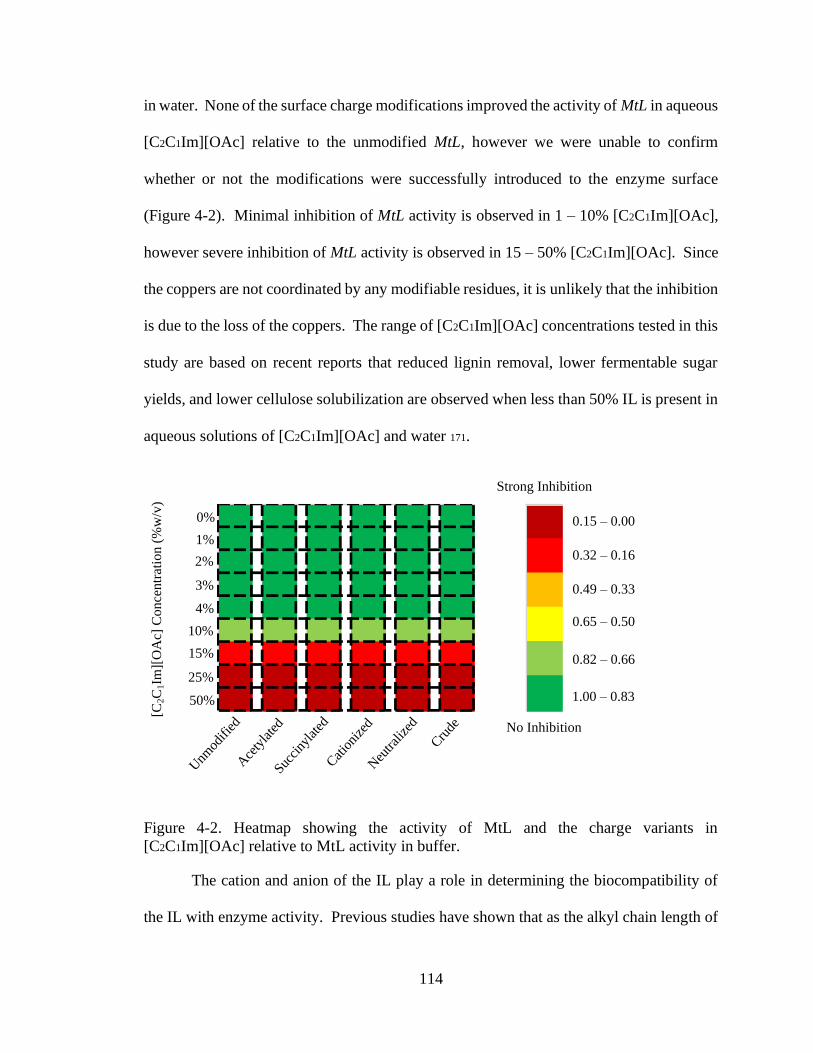
114
in water. None of the surface charge modifications improved the activity of MtL in aqueous
[C2C1Im][OAc] relative to the unmodified MtL, however we were unable to confirm
whether or not the modifications were successfully introduced to the enzyme surface
(Figure 4-2). Minimal inhibition of MtL activity is observed in 1 – 10% [C2C1Im][OAc],
however severe inhibition of MtL activity is observed in 15 – 50% [C2C1Im][OAc]. Since
the coppers are not coordinated by any modifiable residues, it is unlikely that the inhibition
is due to the loss of the coppers. The range of [C2C1Im][OAc] concentrations tested in this
study are based on recent reports that reduced lignin removal, lower fermentable sugar
yields, and lower cellulose solubilization are observed when less than 50% IL is present in
aqueous solutions of [C2C1Im][OAc] and water 171.
Figure 4-2. Heatmap showing the activity of MtL and the charge variants in
[C2C1Im][OAc] relative to MtL activity in buffer.
The cation and anion of the IL play a role in determining the biocompatibility of
the IL with enzyme activity. Previous studies have shown that as the alkyl chain length of
0%
1%
2%
3%
4%
10%
15%
25%
50%
Unm
odifi
edA
cety
late
dSuc
ciny
late
dCat
ioni
zed
Neu
traliz
ed
Cru
de
[C2C
1Im
][O
Ac]
Con
centr
atio
n (
%w
/v)
0.15 – 0.00
0.32 – 0.16
0.49 – 0.33
0.65 – 0.50
0.82 – 0.66
1.00 – 0.83
Strong Inhibition
No Inhibition

115
the cation increases, alkylimidazolium ILs become more inhibitory 265, 290. The inhibitory
effect of the anion has been shown to follow the Hofmeister series 133, 303. ILs with
chaotropic anions (e.g. NTf2- and Br-) more strongly destabilize proteins when compared
to kosmotropic anions (e.g. SO42- and PO43-). For example, the activity of T. versicolor
laccase is increased 451% in 10 mM choline dihydrogen phosphate ([Ch][H2PO4]), an IL
with a kosmotropic anion, but is not affected by the same concentration of choline
bis([trifluoromethyl]sulfonyl)imide ([Ch][NTf2]), an IL with a chaotropic anion 150.
Nordwald et al. previously showed that acetylation and succinylation improves enzyme
activity and stability in 1-butyl-3-methylimidazolium chloride ([C4C1Im][Cl]) by reducing
the interaction of the chaotropic Cl- anion with the enzyme surface. Additionally, low
enzyme activity in higher concentrations of [C2C1Im][OAc] can be attributed to the
buffering capacity of [C2C1Im][OAc]. Optimal ABTS oxidation occurs at pH < 5 for most
fungal and baterial laccases, but [C2C1Im][OAc] buffers at a pH well above this range
thereby lowering MtL activity in high concentrations of [C2C1Im][OAc] 304.
4.4.2 Stability [C2C1Im][OAc]
Although the surface charge modifications did not improve the activity of MtL, we
sought to understand if the modifications improved the stability of MtL in [C2C1Im][OAc].
All charge variants of MtL were incubated in 50 mM citrate/100 mM phosphate buffer (pH
4.5) with 0%, 2.5%, and 5% [C2C1Im][OAc] at 40oC and the activity was measured at
different time intervals up to 4 hours. The surface charge modifications, assuming they
were successfully introduced to the enzyme surface, did not improve the stability of MtL
in low concentrations of [C2C1Im][OAc] (Figure 4-3). Minimal activity loss was observed
after 4 h in buffer for all charge variants. When incubated in 2.5% [C2C1Im][OAc] >50%

116
initial activity is lost after 2 h, while >50% initial activity is lost after only 1 h in 5%
[C2C1Im][OAc]. The IL concentrations tested are based on the results of the
biocompatibility screening: 2.5 and 5% [C2C1Im][OAc] represent ranges of IL
concentrations in which no or minimal activity loss is observed, respectively.
Figure 4-3. Heatmaps showing the residual activity of MtL charge variants after incubation
in 50 mM citrate/100 mM phosphate buffer and 2.5 or 5% [C2C1Im][OAc] at pH 4.5 and
40oC.
Previously, enzymes with surface charge modifications in ILs were shown to have
a higher or lower half-life relative to the wild type in the IL. Both succinylation and
0% [C2C1Im][OAc]
2.5% [C2C1Im][OAc]
5% [C2C1Im][OAc]
0 min30 min
60 min
120 min
180 min
240 min
Unm
odifi
edA
cety
late
dSuc
ciny
late
dCat
ioni
zed
Neu
traliz
ed
0 min30 min
60 min
120 min
180 min
240 min
0 min30 min
60 min
120 min
180 min
240 min
Incu
bat
ion T
ime 0.15 – 0.00
0.32 – 0.16
0.49 – 0.33
0.65 – 0.50
0.82 – 0.66
0.83 – 1.00
Low Residual Activity
High Residual Activity

117
acetylation improved the half-life of lipase and α - chymotrypsin in 40% [C4C1Im][Cl] and
55% [C2C1Im][EtSO4], respectively, while acetylation improved the half-life of papain in
30% [C4C1Im][Cl]; however, neutralization and cationization reduced the half-life of lipase
and α - chymotrypsin in IL, respectively 292. Succinylated and acetylated enzymes are
thought to be more stable in ILs due to 2 factors: their similarity to halophilic enzymes and
the formation of salt bridges. Halophilic enzymes have a large number of acidic surface
residues which attract water molecules, increasing protein hydration and preventing the
enzyme from aggregating, while the salt bridges further increase enzyme stability in saline
environments 94, 305, 306.
4.4.3 Docking Simulations
Due to the unknown concentration of ABTS in the 1-Step solution used in the
biocompatibility screening, we were unable to use enzyme kinetics analyses to understand
the inhibition of MtL in [C2C1Im][OAc]. Therefore, docking simulations were used to
better understand the effect of the surface charge modifications on the major binding
locations of [C2C1Im][OAc] to the enzyme surface. The results of the docking simulations
show that the modification chemistry affects the major docking locations of the IL to the
surface of MtL (Figure 4-4). The major docking location of the IL to the surface of the
unmodified MtL was limited to a region close to the T1 copper (Figure 4-4a). When MtL
was acetylated or succinylated the IL docked close to the T1 copper in addition to a new
[OAc]- binding location away from the T1 copper (Figure 4-4b). When MtL was cationized
or neutralized [C2C1Im]+ docked close to the T1 copper while [OAc]- docked further from
the T1 copper but still close to the active site entrance (cationized) or away from the active
site (cationized and neutralized) (Figure 4-4c).

118
Figure 4-4. Major docking locations of [C2C1Im][OAc] to the surface of unmodified (a),
succinylated/acetylated (b), and cationized/neutralized MtL (c). [C2C1Im]+ is shown as
magenta spheres, [OAc]- is shown as red spheres, and coppers are shown as brown spheres
A closer inspection of the [C2C1Im]+ docking location reveals the presence of
several aromatic residues that would not be affected by any of the surface charge
modifications. These aromatic residues would be able to stabilize [C2C1Im]+ via π – π or
π – cation interactions in all charge variants. While these results are consistent with a
previous report that [C2C1Im]+ docks to MtL in the presence of aromatic residues, we did
not observe [C2C1Im]+ entering the active site 269. This study did not restrict the possible
[C2C1Im]+ binding locations, whereas the previous study focused the docking around the
MtL active site which could affect the degree to which the cation entered into the active
site. Arg456 and His457 stabilize [OAc]- close to the active site of unmodified,
succinylated, and acetylated MtL. When docking to succinylated and acetylated MtL,
[OAc]- is stabilized by Arg226 and Arg228 at the 2nd [OAc]- binding site. The major
[OAc]- docking location close to the active site of cationized MtL contains 2 modified
residues (Asp360 and Glu420), in addition to several lysines. The 2nd [OAc]- binding site
to cationized and neutralized MtL contains several arginines, lysines, and modified
residues.
a) b) c)

119
The shift in [OAc]- binding is determined by the modification chemistry.
Acetylation and succinylation reduce the positive surface charges that stabilize [OAc]-,
driving the anion to the arginine-rich binding pocket. Neutralization and cationization
further increase the favorability of anion binding to an arginine-rich pocket containing
modified glutamate and aspartate. The previous work on modifying surface charges
suggest that acetylation and succinylation discourage the chaotropic anion (e.g. Cl- or
[OAc]-) from interacting with the enzyme surface, thereby improving enzyme activity and
stability in IL 202, 292. Our docking results show that all surface charge modifications
encourage [OAc]- binding away from the active site, however neither MtL activity nor
stability in [C2C1Im][OAc] are affected by the modifications. Laccase activity is dependent
upon the presence of a T1 copper in the active site, a trinuclear copper cluster, and a Cys-
His pathway between the T1 copper and the trinuclear cluster. If the surface charge
modifications are confirmed, then the following mechanism can be proposed: the loss of
MtL activity and stability in [C2C1Im][OAc] is likely due to destabilization of the coppers
and the Cys-His pathway by the IL via a mechanism that cannot be influenced by
modifying the enzyme surface charges. Without confirming the presence of the enzyme
surface modifications this mechanism cannot be supported by the current data.
4.5 Conclusions and Future Work
In this study we examined the biocompatibility of aqueous [C2C1Im][OAc] with the
activity of the thermophilic fungal laccase from Myceliophthora thermophila. Results
show severe inhibition of MtL activity in only 15% (w/v) [C2C1Im][OAc], far below the IL
concentration necessary for effective biomass pretreatment. In an effort to reduce this
inhibition of MtL activity, we made several surface charge modifications to increase the

120
acid or amine surface residues. Although previous studies have shown that surface charge
modifications can increase enzyme activity and stability in aqueous ILs, the stability and
activity of all MtL charge variants in [C2C1Im][OAc] were unchanged compared to the
unmodified enzyme. However, it is not known at this time if the modifications were
successfully introduced to the enzyme surface. Docking simulations show that the IL
docks close to the T1 copper on all charge variants, with some additional [OAc]- docking
locations on the surface of modified enzymes. Further work might seek to 1) confirm the
presence of the surface charge modifications, 2) better understand the mechanism by which
the IL inhibits MtL activity through the use of enzyme kinetics and molecular dynamics
(MD) simulations and 3) employ additional techniques (e.g. rational design or
immobilization) to improve MtL activity in [C2C1Im][OAc].
Methods used to assess the effects of ILs on enzymes can be divided into in vitro
(e.g. activity assays, stability assays, and enzyme kinetics) and in silico (e.g. docking
simulations and molecular dynamics [MD] simulations) techniques. In vitro techniques
can be used to provide a broad overview of how the ILs are affecting the enzyme activity,
stability, and substrate affinity. By measuring the activity of TvL in ILs, past studies have
shown that [C2C1Im][EtSO4] is the most biocompatible alkylimidazolium IL, suggesting
that this laccase-IL combination would be best suited for lignin deconstruction applications
198, 290. However, these activity assays are done on a short time scale which does not
provide any information about the stability of the laccase in the IL. Although the initial
activity of MtL immobilized on glyoxyl-agarose beads is lower in 25% [C2C1Im][EtSO4]
than the free laccase initial activity, the immobilized laccase is far more stable in 75% IL
than the free laccase over a 7 day period 198. Enzyme kinetics studies, performed by

121
measuring the initial velocity of enzymes in different substrate concentrations, can further
indicate if the loss in activity is due to the IL affecting the enzyme activity or substrate
binding. Kinetic studies of MtL in [C2C1Im][Cl] showed that the IL is a competitive
inhibitor that competes with the substrate for binding in the active site 269.
[C2C1Im][EtSO4] was found to interact with MtL as an uncompetitive inhibitor that does
not affect substrate binding, but rather affects substrate oxidation once it has bound 198.
Some ILs are also able to affect both substrate binding and oxidation, for example
[C2C1Im][OAc] and [Ch][Lys], which were found to be mixed inhibitors that decreased
substrate affinity and activity of TvL 291 .
To further understand the laccase-IL interactions seen with kinetic studies, recent
studies have used docking simulations with ILs and static protein models. Docking
simulations showed that the diffusion of [CnC1Im]+ cations (n = 2, 4, 6, 8, 10) into the
active site of MtL is determined by the alkyl chain length 269. Similarly, [C2C1Im][OAc]
and [Ch][Lys] were found to dock close to the T1 copper of TvL 291. In both of these cases
the docking simulations supported the previous kinetics results showing the ILs can affect
substrate binding and oxidation. MD simulations can also be used to better capture the
dynamic nature of enzyme-IL interactions. After in vitro experiments showed
[C2C1Im][OAc] is more biocompatible with thermophilic cellulases than a mesophilic
cellulase, MD simulations revealed the IL effect is limited to local disturbances of
thermophilic cellulases or secondary structure unfolding of a mesophilic cellulase 199, 307.
MD simulations also provided further insight into enzyme-IL interactions that increase the
thermostability of a Fusarium solani serine protease cutinase in 1-butyl-3-
methylimidazolium hexafluorophosphate ([C4C1Im][PF6]) 308. Further insight into the

122
effect of the IL on enzymes can be gained by applying different methods to understand the
binding free energy of the IL or substrate to the enzyme surface or active site 309.
In this study we applied a single enzyme engineering approach to try and improve
MtL activity and stability in [C2C1Im][OAc]. However, previous studies have applied other
enzyme engineering strategies to achieve the same goal (Table 4-1). The enzyme used in
this study was not produced recombinantly, therefore we were unable to apply and
biotechnological (e.g. directed evolution or rational design) methods to improve MtL
activity and stability. Future studies with MtL in [C2C1Im][OAc] might apply
immobilization techniques similar to those previously used to improve stability in
[C2C1Im][EtSO4] 198. Additionally, recombinant expression of MtL would enable future
researchers to apply other enzyme engineering techniques. The L1 loop can be targeted
for mutations with a directed evolution or computationally assisted strategy, similar to
previous studies with TvL 293, 294. Additionally, novel targets for mutations can be
identified using a computationally assisted approach like that used to identify the Glu170
target in B. subtilis CotA 265. The application of more robust methods for analyzing
laccase-IL interactions alongside targeted enzyme engineering strategies will facilitate the
identification of laccase-IL combinations suited for use in future lignin valorization
systems.

123
Table 4-2. Previous studies using various methods to improve enzyme activity in aqueous
ILs
Method Enzyme Outcome Citation
Computationally assisted
protein engineering
T. versicolor laccase Triple and quadruple mutants with
increased activity in [C2C1Im][EtSO4]
294
Directed evolution T. versicolor laccase Double mutant with increased activity
in [C2C1Im][EtSO4]
293
Computationally assisted
protein engineering
B. subtilis laccase Single mutants with increased catalytic
efficiency (kcat/Km) in 3 [CnC1Im][Cl]
ILs
265
Surface charge
modification
Bovine pancreas α-
chymotrypsin, Carica
papaya papain, Candida
rugosa lipase
Succinylation and acetylation
improved activity and stability in
[C4C1Im][Cl] and [C2C1Im][EtSO4]
292
Surface charge
modification
Trichoderma reesei
cellulase
Succinylation improved cellulose
hydrolysis in [C4C1Im][Cl]
202
Immobilization M. thermophila laccase Immobilization on glyoxyl-agarose
beads improved stability in
[C2C1Im][EtSO4]
198
4.6 Author Contributions
Joseph Stevens and Jian Shi conceptualized the work, designed the experiments,
and analyzed the data. Joseph Stevens conducted the experiments and wrote the
manuscript.
4.7 Acknowledgements
The authors acknowledge the support from the National Science Foundation under
Cooperative Agreement No. 1355438 and 1632854 (2016 – 2020) and the National

124
Institute of Food and Agriculture, U.S. Department of Agriculture, Hatch-Multistate
project under accession number 1018315 (2018 – 2023).

125
CHAPTER 5. A MIXED TECHNIQUE APPROACH TOWARDS UNDERSTANDING IONIC LIQUID
INHIBITION OF A PLANT LACCASE
5.1 Abstract
High concentrations of alkylimidazolium ionic liquids are required for biomass
pretreatment, increasing their cost of use and lowering their biocompatibility with the
lignolytic enzyme laccase. Due to the low concentration of [Ch][Lys] required for biomass
pretreatment, it is an attractive target to be used alongside laccases in a low-cost biological
lignin valorization system. We investigated the biocompatibility of [Ch][Lys] with a plant
laccase from Toxicodendron vernicifluum. Severe inhibition of laccase activity was seen
in only 3% [Ch][Lys]. Enzyme kinetics and docking simulations suggest the [Ch][Lys]
inhibits via a mixed competitive and noncompetitive mechanism. [Ch]+ acts as the
competitive inhibitor, directly interfering with DMP binding the active site, while [Lys]+
acts as the noncompetitive inhibitor that interferes with DMP catalysis once it is bound.
Subsequent molecular dynamics simulations were used to better capture the dynamic
nature of the laccase-IL interactions, the first time these simulations have been used with a
laccase. These simulations revealed that [Ch][Lys] does not perturb the enzyme structure,
but rather lowers the stability of DMP in the active site. These results show for the first
time that IL inhibition of laccase activity could be due to substrate-IL interactions in
addition to the laccase-IL interactions seen in previous studies.

126
5.2 Introduction
Laccases are a member of the multicopper oxidase superfamily of enzymes (E.C.
1.10.3.2). First identified in extract from the Japanese lacquer tree Toxicodendron
vernicifluum, several laccases and laccase-like enzymes have been discovered in bacteria,
fungi, and archaea 84, 85, 90, 91, 93. Laccases differ from the other lignolytic enzymes, the
heme peroxidases, in that they are copper containing enzymes that require molecular
oxygen, as opposed to hydrogen peroxide, for their oxidation mechanisms 71, 80, 310.
Laccases are also capable of oxidizing non-phenolic lignin compounds when coupled with
a small molecule mediator such as 2,2’-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid)
(ABTS) or 1-hydroxybenzotriazole (HBT) 97, 256, 289. As a result laccases have been the
target of numerous biotechnological applications, including lignin valorization 288.
Lignin is among the most abundant biopolymers on earth, making up about 30% of
the weight of terrestrial biomass 40, 244. It is primarily made of 3 phenylpropanoid subunits:
p-hydroxyphenyl (H-lignin), guaiacyl (G-lignin), and syringyl (S-lignin) 41, 43. These
subunits are linked by a complex network of condensed (e.g. β – β or β – 5) and ether (e.g.
β – O – 4 or β – O – 5) bonds 39, 45. The ratio of subunits present in the lignin network is
also dependent upon the biomass source 44. As a result of its monomeric and structural
heterogeneity, lignin is very recalcitrant towards thermochemical and biological
depolymerization methods for production of chemicals at high selectivity. Lignin is
generated in large volumes as a waste product by the paper and pulping industry and during
the production of cellulosic biofuels 1, 247 . Developing technologies that convert lignin
from waste to value-added products will not only reduce waste but add value to the paper
and pulping industry and cellulosic biorefineries 3, 311.

127
Numerous lignolytic enzymes have been identified in bacteria and fungi, most
notably the heme peroxidases (manganese, versatile, and lignin) and laccase; lignin
breakdown pathways have also been identified in the soil bacterium Sphingobium sp. SYK-
6 which metabolizes aromatic compounds via NADP and glutathione dependent enzymes
13, 15, 281. The use of enzymes in biological lignin valorization can be beneficial from both
an operating cost and safety standpoint as a result of the mild operating conditions (low
temperature, low pressure, mild pH) and improved yield of lignin breakdown products due
to the efficiency of biocatalysts 6, 12. These monomeric lignin breakdown products can be
used in place of petroleum-derived precursors for chemicals used in the pharmaceutical,
food, and plastic industries 248, 249. However, many of the known lignin solvents (e.g.
dimethylsulfoxide [DMSO], acetone, and alkaline solutions) reduce or altogether eliminate
enzyme activity 18, 19. These solvents also require the use of high temperatures for complete
dissolution of native plant lignin, further eliminating enzyme activity in the process 284.
The potential for biological lignin valorization strategies is great, however many challenges
(e.g. high cost of enzyme production and poor monomer yield due to low lignin solubility)
must be overcome by using a low-cost biocompatible solvent that is capable of dissolving
lignin at low temperatures 16, 117.
Ionic liquids (ILs) are molten salts that are liquid at temperatures below 100oC.
Considered to be “green solvents” due to their relatively low toxicity and vapor pressure,
the properties of ILs can be tuned by selecting the appropriate cation and anion 127, 128, 131,
133. Alkylimidazolium ILs, for example 1-ethyl-3-methylimidazolium acetate
([C2C1Im][OAc]), have been studied for their ability to reduce cellulose crystallinity and
extract lignin during relatively low temperature biomass pretreatment 251-253. Many

128
alkylimidazolium ILs have been shown to be biocompatible with the activity of laccase
from the white rot fungi Trametes versicolor at different concentrations in water; >80%
initial activity remained in 50% (v/v) 1-ethyl-3-methylimidazolium ethylsulfate
([C2C1Im][EtSO4]) 290. Due to the high cost associated with using [C2C1Im][OAc], recent
efforts have focused on developing ILs for biomass pretreatment using low-cost, bio-
derived cations (e.g. choline and ammonium) and anions (e.g. amino acids and carboxylic
acids) that can be used at low concentrations in water 164, 167, 243. For example, when
switchgrass was pretreated with 10% [Ch][Lys] in water, greater than 80% lignin removal
was observed 254. The low concentration of [Ch][Lys] required for effective biomass
pretreatment could improve its biocompatibility since enzyme activity in ILs is strongly
dependent upon IL concentration 290, 293. However, we previously found that T. versicolor
laccase is severely inhibited in just 1% [Ch][Lys], far below the concentration required for
effective biomass pretreatment 291. The tunability and biocompatibility of ILs makes them
attractive solvents to be used with laccases in biological lignin valorization strategies,
however relatively little is known of laccase-IL interactions beyond in vitro experiments
Molecular dynamics (MD) simulations can be used to understand the interactions
between enzymes and ILs at an atomistic resolution. After initial in vitro experiments
determined that [C2C1Im][OAc] is more biocompatible with thermophilic cellulases than a
mesophilic cellulase, subsequent MD simulations revealed that the main effect of the IL
was limited to local disturbances, in the case of the thermophilic cellulases, or complete
unfolding of the enzyme secondary structure, in the case of the mesophilic cellulase 199, 307.
MD simulations of a serine protease cutinase from Fusarium solani in 1-butyl-3-
methylimidazolium hexafluorophosphate ([C4C1Im][PF6]) found that the IL stabilizes the

129
protein structure at high temperatures, a result already obtained from in vitro lab
experiments 308. Previous studies on laccase-IL interactions have used docking simulations
to identify docking locations of the IL on the enzyme surface, for example T. versicolor
laccase with [C2C1Im][OAc], [Ch][Lys], and [DEA][HSO4] or M. thermophila laccase
with several alkylimidazolium cations 269, 291. In both of these cases the docking results
supported the kinetics data, however the ILs were docked to a static model of the laccases
which fails to capture the dynamic nature of the laccase-IL interactions. Additionally,
previous kinetics data suggest that ILs not only interfere with substrate binding but also
affect oxidation once the substrate is bound 150, 198, 291; an in-depth investigation of laccase-
IL interactions would greatly benefit from the atomistic resolution of MD simulations.
5.3 Methods and Materials
5.3.1 Materials
Laccase from T. vernicifluum (RvL, ≥50 units/mg solid), 2,6-dimethoxyphenol
(DMP), choline hydroxide, L-lysine, and all other reagents were purchased from
MilliporeSigma (St. Louis, MO, USA). Cholinium lysinate ([Ch][Lys]) was synthesized
using the anion and cation precursors according to previously developed protocols 167.
5.3.2 Plant laccase purification
RvL was purified from the crude acetone extract using the method previously
published 312. First, 35 mg of the crude extract was mixed with 10 mL buffer (0.1 M
potassium phosphate, pH 6.0) at room temperature for 16-20 h. After overnight mixing,
any undissolved material was filtered using 0.45 and 0.2 um syringe filters. The filtered
material (10 mL) was then mixed with 3g of (NH4)2SO4 then centrifuged for 20 min at

130
15000g to remove any precipitated material. Finally, the supernatant was applied at a rate
of 0.5 mL min-1 to a column (11 mm diameter x 10.4 cm height) of phenyl-Sepharose
(Tosoh Bioscience, Tokyo, Japan) equilibrated with 2.67 M (NH4)2SO4 in 0.01 M
potassium phosphate buffer (pH 6.0). The protein was eluted off the column by reducing
the concentration of (NH4)2SO4 at a gradient from 2.67 M to a final concentration of 0.27
M. Protein content was determined using the Bradford method.
5.3.3 Biocompatibility screening
The biocompatibility of [Ch][Lys] in aqueous solution (approximately 1-10% w/v),
was screened with purified RvL. To reduce the effect of pH, the IL solution was adjusted
to pH 8.0 using 1 M hydrochloric acid prior to testing. Activity was screened with 25 mM
MES, HEPES, glycine, and acetic acid buffer (pH 8.0); IL (0, 1, 2, 3, 4, or 10% w/v); and
25 mM DMP into a clear, flat-bottom, 96-well Costar assay plate. Absorbance readings
were taken continuously for 10 min at 25 ºC using a SpectraMax M3 Microplate reader
(Molecular Devices, Sunnyvale, CA). Oxidation of DMP in buffer and AILs without
laccase were measured as blanks. The relative activity of RvL in ILs relative to buffer was
calculated using equation 5-1.
𝑅𝑒𝑙𝑎𝑡𝑖𝑣𝑒 𝑎𝑐𝑡𝑖𝑣𝑖𝑡𝑦 = 𝐴𝑐𝑡𝑖𝑣𝑖𝑡𝑦 𝑖𝑛 𝐼𝐿
𝐴𝑐𝑡𝑖𝑣𝑖𝑡𝑦 𝑖𝑛 𝑏𝑢𝑓𝑓𝑒𝑟 (5-1)
5.3.4 Enzyme Kinetics
Michaelis-Menten curves were generated for IL concentrations (0, 2, and 4% w/v)
by varying the concentration of DMP (2 – 25 mM) and measuring initial velocities. DMP
oxidation was measured using the same method as described in the biocompatibility
screening. The oxidized DMP was quantified by measuring the absorbance at 469 nm and

131
using the absorbance extinction coefficient Ɛ469 = 35,645 M-1 cm-1. An activity unit was
defined as 1 µmol DMP oxidized in 1 min (µmol DMP/min). Initial velocity (V) as a
function of DMP concentration ([S]) was fit to the Michaelis-Menten curve, shown in
equation 5-2.
V = Vmax×[S]
Km+[S] (5-2)
The kinetic coefficients were calculated by plotting equation 5-2 in Sigma Plot and
calculating Km and Vmax using the ligand binding curve fitting algorithm.
5.3.5 Plant laccase modelling and copper parameterization
The 3D model of plant laccase was built using the Modeller Software, version 9.23
313. The sequences of Zea mays laccase 3 (PDBID: 6KLG, sequence id: 48.91%) and
Cucurbita pepo ascorbate oxidase (PDBID: 1AOZ, sequence id: 31.80%) were used as
templates in Modeller. Multiple sequence alignment was performed with MAFFT
Software, version 7.310 314. The 100 models generated by Modeller were ranked based on
their Discrete Optimized Protein Energy (DOPE) score 315. The model with the lowest
DOPE score was selected as the best model. The parameters (force constants, geometry,
and charges) of the coppers and their coordinating residues were calculated using
MCPB.py, a python based metal center parameter builder 316. All quantum calculations
were computed using Gaussian 09 D.01 with B3LYP-631G* functionality 317.

132
5.3.6 Docking Simulations
The 3D structures and PDB files of the ligands were prepared in YASARA
Structure (YASARA Biosciences GmbH, Vienna, Austria) using the SMILES strings
obtained from PubChem. The PDBQT files of the ligands and the RvL model were
prepared with AutoDock (version 4.2.6, MGLTools, La Jolla, CA). Charges for the
coppers were added by manually editing the RvL PDBQT file. AutoGrid parameters were
as follows: space value of 0.375Å, (x, y, z) grid centered at (80.297, 66.064, 8.193), and
grid size of 126 in all directions. AutoDock parameters were as follows: Lamarckian GA;
100 genetic algorithm runs; 0.5Å RMS tolerance; and 25,000,000 max eval size. [Ch]+ and
[Lys]+ were docked separately to both unbound RvL and RvL with DMP bound to the active
site. The structure of RvL with DMP in the active site was prepared in PyMol by aligning
the RvL model with Melanocarpus albomyces laccase containing DMP in the active site
(PDBID: 3FU8).
5.4 Results and Discussion
5.4.1 Biocompatibility of AILs with laccase activity
To determine the biocompatibility of [Ch][Lys] with RvL, the activity of RvL was
measured in aqueous concentrations of [Ch][Lys] (1-10%) in water. The activity of RvL
in different concentrations of [Ch][Lys] relative to the activity in buffer is shown in Figure
5-1. RvL is strongly inhibited by [Ch][Lys], only 50% initial activity is remaining in 3%
[Ch][Lys]. As the concentration of [Ch][Lys] continues to increase, the decrease in RvL
activity is not as severe although only 20% initial activity remains in 10% [Ch][Lys]. The
range of [Ch][Lys] tested is relevant to recent work investigating the effect of water during

133
IL pretreatment of biomass. The pretreatment efficacy of [C2C1Im][OAc] is severely
reduced at a concentration of 50% in water 171. High sugar yields were obtained after
pretreating switchgrass at 180oC for 15 min using triethylammonium hydrogen sulfate
([TEA][HSO4]) in 20% water 285. In contrast, 10% [Ch][Lys] in water removes 80% of
lignin from genetically engineered switchgrass during pretreatment 254.
Figure 5-1. Effect of aqueous [Ch][Lys] on the activity of RvL. Values shown are the mean
± SD of 3 experiments
Both the cation and the anion play a role in enzyme inhibition. As the alkyl chain
length increased (C2 -> C10) the activity of Myceliophthora thermophila laccase decreased
in several alkylimidazolium chloride ([CnC1Im][Cl]) ILs 269. A decrease in the activity of
Trametes versicolor laccase was seen as the alkyl chain length increased (C2 -> C10) and
the chaotropicity of the anion increased 290. Chaotropic anions, e.g.
bis(trifluoromethylsolfonyl) imide ([NTf2]-) and bromide (Br-), are more inhibitory than
less chaotropic anions, e.g. chloride (Cl-) and phosphate (PO43-), closely following the
[Ch][Lys] (%w/v)
0 2 4 6 8 10
Rela
tive A
cti
vit
y
0.0
0.2
0.4
0.6
0.8
1.0

134
Hofmesiter series 148. Previous experiments have shown that the activity of T. versicolor
laccase was increased 451% by 10 mM choline dihydrogenphosphate ([Ch][H2PO4]), not
affected by 10 mM ([Ch][NTf2]), and severely inhibited by 1% [Ch][Lys] 150, 291. Our
results, along with the results of the previous studies indicate that lysine is not
biocompatible with laccase activity.
5.4.2 Inhibition Mechanisms
We used enzyme kinetics to elucidate the inhibition mechanisms of [Ch][Lys]
towards RvL activity using DMP as a substrate. We chose to investigate the kinetics of
DMP oxidation at 0, 2, and 4% [Ch][Lys] based on the results of the biocompatibility
screening. The changes in kcat and Km with increasing [Ch][Lys] concentration suggest the
presence of 2 binding sites (Table 5-1). A high affinity competitive binding site that is
completely occupied in 2% [Ch][Lys] increases Km while a low affinity non-competitive
binding site lowers kcat in all concentrations of [Ch][Lys].
Table 5-1. Kinetic constants of RvL in different concentrations of [Ch][Lys]. Values shown
are mean ± standard error of 3 experiments
[Ch][Lys]
(%w/v)
Km
(mM DMP)
kcat
(s-1)
kcat/Km
(s-1 mM-1)
0% [Ch][Lys] 28.88 ± 3.61 3.15 ± 0.15 0.11
2% [Ch][Lys] 39.74 ± 2.88 2.75 ± 0.05 0.07
4% [Ch][Lys] 40.58 ± 3.14 2.10 ± 0.03 0.05
Previous kinetics work has shown that several ILs are capable of acting as
competitive, noncompetitive, or uncompetitive inhibitors. Aqueous solutions of

135
[C2C1Im][OAc] and 1-ethyl-3-methylimidazolium sulfate ([C2C1Im][EtSO4]) acted as
competitive (increased Km) and noncompetitive (decreased Vmax) inhibitors of T. versicolor
laccase 183. [Ch][H2PO4] and [C2C1Im][EtSO4] increased Km and kcat of T. versicolor and
M. thermophila laccase, respectively, by binding to [E][S] as uncompetitive inhibitors 150,
290.
5.4.3 Copper parameterization and stabilization during simulations
The calculated charges of the coppers and hydroxyl groups bounded to the coppers
are shown in Figure S5-2. The distance between the coppers and their coordinating
residues were recorded during a 100 ns simulation to ensure the stability of the coppers.
As seen in Figure 5-2, the distances between the coppers and their coordinating residues
are around 2Å, consistent with the distances seen in laccase crystal structures 86, 318.

136
Figure 5-2. Distances between (top) T1 copper, (middle) T2 copper, and (bottom) T3
coppers and coordinating residues during 100 ns simulations
5.4.4 IL Docking
To better understand where [Ch][Lys] binds to the surface of RvL, we performed
docking simulations using AutoDock. Major docking poses of [Ch][Lys] are shown in
Figure 5-3. Three major docking locations were identified on both [E] and [E][S]. [Ch]+
docks close to the T1 copper and the trinuclear copper cluster in a pocket close to the C-

137
terminus. [Lys]+ binds to the same C-terminus pocket and a 3rd location close to the
trinuclear copper cluster. These results suggest that [Ch]+ acts as both a competitive and
noncompetitive inhibitor, while [Lys]+ only acts as a noncompetitive inhibitor.
Figure 5-3. Major docking locations of [Ch][Lys] to the surface of [E] (panel a, green) and
[E][S] (panel b, cyan). [Ch]+ is shown as red spheres, [Lys]+ is shown as magenta spheres,
DMP is shown as yellow spheres, and coppers are shown as brown spheres.
Previous results docking ILs to laccases have shown that ILs are capable of docking
close to or within the active site. When alkylimidazolium cations were docked to M.
thermophila laccase, it was found that short chain length cations (C2 – C6) were able to
diffuse into the active site, whereas the longer chain length cations (C8 – C10) were limited
to interacting with leucines surrounding the active site 269. We showed in a previous study
that [C2C1Im][OAc] and [Ch][Lys] dock with high probability to a region close to the
active site of T. versicolor laccase 291.
5.4.5 Steered Molecular Dynamics of DMP in [Ch][Lys]
To better understand the effect of [Ch][Lys] on the stability of DMP in the active
site of RvL, we analyzed the free energy profile of DMP using steered molecular dynamics
(a) (b)

138
(SMD) analysis. The free energy of DMP was calculated at a range of distances between
the hydroxyl group of DMP and the T1 copper of RvL. Figure 5-4 below shows the free
energy profile of DMP in 0, 2, and 4% [Ch][Lys] at different distances from the T1 copper.
The lower free energy 6 – 8 Å away from the T1 copper is consistent with the location of
DMP when docked into the active site of RvL from the M. albomyces crystal structure.
Additionally, the free energy within this range becomes less negative as the concentration
of [Ch][Lys] increases, suggesting that [Ch][Lys] lowers the stability of DMP in the active
site. These results are consistent with the kinetics, which showed that [Ch][Lys] disrupts
substrate binding and oxidation, and the docking simulations, which showed that [Ch][Lys]
can dock close to the T1 copper in close proximity to DMP.
Figure 5-4. Free energy profile of DMP in 0 (blue line), 2 (green line), and 4% (red line)
[Ch][Lys]. The measurement of the distance between the T1 copper of DMP hydroxyl
group is also shown for reference.
5.5 Author Contributions
Joseph Stevens, Jérémy Esque, Isabelle André, David Rodgers, and Jian Shi
conceptualized the work, designed the experiments, and analyzed the data. Jérémy Esque
performed the RvL modelling, copper parameterization, and MD simulations. Joseph
Stevens conducted the all other experiments and wrote the manuscript.
6.8Å

139
5.6 Acknowledgements
The authors acknowledge the support from the National Science Foundation under
Cooperative Agreement No. 1355438 and 1632854 (2016 – 2020) and the National
Institute of Food and Agriculture, U.S. Department of Agriculture, Hatch-Multistate
project under accession number 1018315 (2018 – 2023). This material is based upon
research supported by the Chateaubriand Fellowship of the Office for Science &
Technology of the Embassy of France in the United States (2019).

140
5.7 Supplemental Information
Figure S 5-1. Michaelis-Menten curves of RvL in aqueous [Ch][Lys]. Data were plotted
and models were fit in Sigma-Plot. Values plotted are mean ± SD of 3 experiments.
[DMP] (mM)
0 5 10 15 20 25
Sp
ecif
ic A
cti
vit
y (
U *
mg
-1)
0.0
0.2
0.4
0.6
0.8
0% [Ch][Lys]2% [Ch][Lys]4% [Ch][Lys]

141
Figure S 5-2. Final charges and geometry of coppers and coordinating residues

142
CHAPTER 6. CONCLUSIONS AND FUTURE WORK
6.1 Conclusions
The aim of this body of work was to create a biological lignin valorization system
using a biocompatible IL and a laccase. Initial studies showed that the selected ILs, 1-
ethyl-3-methylimidazolium acetate ([C2C1Im][OAc]), cholinium lysinate ([Ch][Lys]), and
diethylamine hydrogen sulfate ([DEA][HSO4]), were biocompatible with a mesophilic
fungal laccase from Trametes versicolor in low concentrations. When Kraft lignin was
treated with the laccase and ILs, few overall changes were made to the lignin network and
product yields were low. These results prompted the search for a more robust biocatalyst
to be used alongside the ILs.
Prior studies showing improved biocompatibility of [C2C1Im][OAc] with
thermophilic cellulases lead to the investigation of thermophilic laccases as biocatalysts
for lignin valorization. The first of these laccases was from the hyperthermophilic
bacterium Thermus thermophilus. Despite this being the most thermotolerant laccase
identified to date (Topt = 92oC), severe loss of initial activity was observed in only 2%
[C2C1Im][OAC]. A rational design approach to improve activity in [C2C1Im][OAc]
subsequently failed to produce any mutants with improved activity in the IL, highlighting
the challenging nature of designing a lignin valorization system with laccases and ILs.
To continue the search for a suitable biocatalyst, the biocompatibility of
[C2C1Im][OAc] was screened with a thermophilic fungal laccase from Myceliophthora
thermophila. Although this laccase was not as thermotolerant as the bacterial laccase,
previous studies have shown it to be active and stable in high concentrations of 1-ethyl-3-
methylimidazolium ethylsulfate ([C2C1Im][EtSO4]). Minimal activity loss was observed

143
in 10% [C2C1Im][OAc], however severe inhibition was observed in concentrations greater
than 15%. Previous studies have shown that modifying the surface charges of enzymes to
increase the number of acid residues improves enzyme activity and stability in ILs. Due
to the ubiquity of this approach, the surface charges of M. thermophila laccase were
modified to improve activity and stability in [C2C1Im][OAc]. These modifications did not
improve the activity or stability of M. thermophila laccase in the IL, however docking
simulations revealed that they do affect the binding of [OAc]- to the enzyme surface. These
results suggest that the laccase-IL interactions leading to M. thermophila laccase inhibition
cannot be mediated by changing the interaction of the IL with the enzyme surface.
The previous studies failed to produce a laccase with improved activity and stability
in [C2C1Im][OAc] by applying methods previously shown to do just that. These studies
had also used enzyme kinetics and molecular docking simulations to understand where the
IL is docking to the enzyme surface, however these simulations fail to accurately capture
the dynamic nature of enzyme-IL interactions at high resolution. Therefore, we used
molecular dynamics (MD) simulations to gain insight into the interactions between the IL
cholinium lysinate ([Ch][Lys]) and a plant laccase from Toxicodendrom vernicifluum.
These simulations showed that the IL does not affect the laccase structure, but rather
decreases the stability of the substrate in the active site. The initial goal of designing a
biological lignin valorization system was unmet, however we gained great insight into
laccase-IL interactions and the challenges of using enzyme engineering methods to
improve the performance of a biocatalyst for an industrial application. The content of these
studies can provide a good foundation for future work investigating and engineering
laccase-IL interactions for lignin valorization.

144
6.2 Future Work
This body of work lays the foundation for many possible avenues of investigating
laccase-IL interactions in the future. Perhaps most obvious are future enzyme engineering
efforts to improve the activity and stability of T. thermophilus laccase in [C2C1Im][OAc].
Our approach was limited in scope and would have benefited from a directed evolution or
computationally guided approach, rather than relying on previous research on fungal
laccases to produce many of the mutants screened in that study. The MD simulations in
Chapter 5 provided useful insight into the nature of laccase-IL, or rather substrate-IL,
interactions. This approach could be used with other laccase-IL combinations to
understand these interactions with atomistic resolution. These results can then be used to
guide enzyme engineering efforts by identifying key residues that mediate the IL
interactions on the enzyme surface. Similar to T. thermophilus laccase, a more thorough
analysis of the interactions between [C2C1Im][OAc] and M. thermophila laccase could
guide enzyme engineering efforts to further improve activity and stability in the IL.
This groundwork could also be applied to systems other than laccases and ILs. The
next generation of designer solvents is arriving with the advent of deep eutectic solvents
(DES). Many studies have already been published that show DESs to be cheaper and easier
to synthesize than ILs. These same DESs are comparable to ILs when studied for their
ability to fractionate biomass during pretreatment. Future studies may investigate laccase-
DES interactions with many of the same methods used in this study. These results may
guide enzyme engineering efforts to improve laccase activity in DES for a cost-effective
and sustainable biological lignin valorization system.

145
BIBLIOGRAPHY
1. Tilman, D.; Socolow, R.; Foley, J. A.; Hill, J.; Larson, E.; Lynd, L.; Pacala, S.;
Reilly, J.; Searchinger, T.; Somerville, C.; Williams, R., Beneficial Biofuels—The Food,
Energy, and Environment Trilemma. Science 2009, 325 (5938), 270-271.
2. Pettersson, K. Black Liquor Gasification-Based Biorefineries – Determining
Factors for Economic Performance and CO2 Emission Balances. Chalmers University of
Technology, 2011.
3. Mottiar, Y.; Vanholme, R.; Boerjan, W.; Ralph, J.; Mansfield, S. D., Designer
lignins: harnessing the plasticity of lignification. Current Opinion in Biotechnology 2016,
37 (Supplement C), 190-200.
4. Ragauskas, A. J.; Beckham, G. T.; Biddy, M. J.; Chandra, R.; Chen, F.; Davis,
M. F.; Davison, B. H.; Dixon, R. A.; Gilna, P.; Keller, M.; Langan, P.; Naskar, A. K.;
Saddler, J. N.; Tschaplinski, T. J.; Tuskan, G. A.; Wyman, C. E., Lignin Valorization:
Improving Lignin Processing in the Biorefinery. Science 2014, 344 (6185).
5. Laskar, D. D.; Yang, B.; Wang, H.; Lee, J., Pathways for biomass‐derived lignin
to hydrocarbon fuels. Biofuels, Bioproducts and Biorefining 2013, 7 (5), 602-626.
6. Beckham, G. T.; Johnson, C. W.; Karp, E. M.; Salvachúa, D.; Vardon, D. R.,
Opportunities and challenges in biological lignin valorization. Current Opinion in
Biotechnology 2016, 42, 40-53.
7. Mu, W.; Ben, H.; Ragauskas, A.; Deng, Y., Lignin pyrolysis components and
upgrading—technology review. BioEnergy Research 2013, 6 (4), 1183-1204.
8. De Wild, P. J.; Huijgen, W. J.; Gosselink, R. J., Lignin pyrolysis for profitable
lignocellulosic biorefineries. Biofuels, Bioproducts and Biorefining 2014, 8 (5), 645-657.
9. Zakzeski, J.; Bruijnincx, P. C.; Jongerius, A. L.; Weckhuysen, B. M., The catalytic
valorization of lignin for the production of renewable chemicals. Chemical reviews 2010,
110 (6), 3552-3599.
10. Chatel, G.; Rogers, R. D., Review: Oxidation of lignin using ionic liquids An
innovative strategy to produce renewable chemicals. ACS Sustainable Chemistry &
Engineering 2013, 2 (3), 322-339.
11. Rinaldi, R.; Jastrzebski, R.; Clough, M. T.; Ralph, J.; Kennema, M.; Bruijnincx,
P. C.; Weckhuysen, B. M., Paving the way for lignin valorisation: recent advances in
bioengineering, biorefining and catalysis. Angewandte Chemie International Edition 2016,
55 (29), 8164-8215.
12. Linger, J. G.; Vardon, D. R.; Guarnieri, M. T.; Karp, E. M.; Hunsinger, G. B.;
Franden, M. A.; Johnson, C. W.; Chupka, G.; Strathmann, T. J.; Pienkos, P. T.; Beckham,
G. T., Lignin valorization through integrated biological funneling and chemical catalysis.
Proceedings of the National Academy of Sciences 2014, 111 (33), 12013-12018.
13. Munk, L.; Sitarz, A. K.; Kalyani, D. C.; Mikkelsen, J. D.; Meyer, A. S., Can
laccases catalyze bond cleavage in lignin? Biotechnology Advances 2015, 33 (1), 13-24.
14. Varman, A. M.; He, L.; Follenfant, R.; Wu, W.; Wemmer, S.; Wrobel, S. A.;
Tang, Y. J.; Singh, S., Decoding how a soil bacterium extracts building blocks and
metabolic energy from ligninolysis provides road map for lignin valorization. Proceedings
of the National Academy of Sciences 2016, 113 (40), E5802-E5811.
15. Shi, J.; Sharma-Shivappa, R. R.; Chinn, M. S., Interactions between fungal growth,
substrate utilization and enzyme production during shallow stationary cultivation of

146
Phanerochaete chrysosporium on cotton stalks. Enzyme and Microbial Technology 2012,
51 (1), 1-8.
16. Brown, M. E.; Chang, M. C., Exploring bacterial lignin degradation. Current
opinion in chemical biology 2014, 19, 1-7.
17. Bugg, T. D.; Ahmad, M.; Hardiman, E. M.; Rahmanpour, R., Pathways for
degradation of lignin in bacteria and fungi. Natural product reports 2011, 28 (12), 1883-
1896.
18. Klibanov, A. M., Improving enzymes by using them in organic solvents. Nature
2001, 409 (6817), 241-246.
19. MOZHAEV, V. V.; KHMELNITSKY, Y. L.; SERGEEVA, M. V.; BELOVA, A.
B.; KLYACHKO, N. L.; LEVASHOV, A. V.; MARTINEK, K., Catalytic activity and
denaturation of enzymes in water/organic cosolvent mixtures. European Journal of
Biochemistry 1989, 184 (3), 597-602.
20. Park, S.; Kazlauskas, R. J., Biocatalysis in ionic liquids–advantages beyond green
technology. Current Opinion in Biotechnology 2003, 14 (4), 432-437.
21. Liu, E.; Das, L.; Zhao, B.; Crocker, M.; Shi, J., Impact of dilute sulfuric acid,
ammonium hydroxide, and ionic liquid pretreatments on the fractionation and
characterization of engineered switchgrass. Bioenergy Research 2017, 10 (4), 1079-1093.
22. Xia, S.; Baker, G. A.; Li, H.; Ravula, S.; Zhao, H., Aqueous ionic liquids and deep
eutectic solvents for cellulosic biomass pretreatment and saccharification. RSC Advances
2014, 4 (21), 10586-10596.
23. Dutta, T.; Shi, J.; Sun, J.; Zhang, X.; Cheng, G.; Simmons, B. A.; Singh, S.,
CHAPTER 3 Ionic Liquid Pretreatment of Lignocellulosic Biomass for Biofuels and
Chemicals. In Ionic Liquids in the Biorefinery Concept: Challenges and Perspectives, The
Royal Society of Chemistry: 2016; pp 65-94.
24. Das, L.; Liu, E.; Saeed, A.; Williams, D. W.; Hu, H.; Li, C.; Ray, A. E.; Shi, J.,
Industrial hemp as a potential bioenergy crop in comparison with kenaf, switchgrass and
biomass sorghum. Bioresource technology 2017, 244, 641-649.
25. Zakzeski, J.; Jongerius, A. L.; Weckhuysen, B. M., Transition metal catalyzed
oxidation of Alcell lignin, soda lignin, and lignin model compounds in ionic liquids. Green
chemistry 2010, 12 (7), 1225-1236.
26. Zakzeski, J.; Bruijnincx, P. C.; Weckhuysen, B. M., In situ spectroscopic
investigation of the cobalt-catalyzed oxidation of lignin model compounds in ionic liquids.
Green chemistry 2011, 13 (3), 671-680.
27. Behling, R.; Valange, S.; Chatel, G., Heterogeneous catalytic oxidation for lignin
valorization into valuable chemicals: what results? What limitations? What trends? Green
Chemistry 2016, 18 (7), 1839-1854.
28. Xu, C.; Arancon, R. A. D.; Labidi, J.; Luque, R., Lignin depolymerisation
strategies: towards valuable chemicals and fuels. Chemical Society Reviews 2014, 43 (22),
7485-7500.
29. Klahn, M.; Lim, G. S.; Seduraman, A.; Wu, P., On the different roles of anions and
cations in the solvation of enzymes in ionic liquids. Physical Chemistry Chemical Physics
2011, 13 (4), 1649-1662.
30. Zhao, H., Methods for stabilizing and activating enzymes in ionic liquids—a
review. Journal of Chemical Technology & Biotechnology 2010, 85 (7), 891-907.
31. Naushad, M.; Alothman, Z. A.; Khan, A. B.; Ali, M., Effect of ionic liquid on

147
activity, stability, and structure of enzymes: A review. International Journal of Biological
Macromolecules 2012, 51 (4), 555-560.
32. Moniruzzaman, M.; Kamiya, N.; Goto, M., Activation and stabilization of enzymes
in ionic liquids. Organic & Biomolecular Chemistry 2010, 8 (13), 2887-2899.
33. Zhao, H.; Olubajo, O.; Song, Z.; Sims, A. L.; Person, T. E.; Lawal, R. A.; Holley,
L. A., Effect of kosmotropicity of ionic liquids on the enzyme stability in aqueous
solutions. Bioorganic Chemistry 2006, 34 (1), 15-25.
34. Kaar, J. L.; Jesionowski, A. M.; Berberich, J. A.; Moulton, R.; Russell, A. J.,
Impact of Ionic Liquid Physical Properties on Lipase Activity and Stability. Journal of the
American Chemical Society 2003, 125 (14), 4125-4131.
35. Park, S.; Kazlauskas, R. J., Improved Preparation and Use of Room-Temperature
Ionic Liquids in Lipase-Catalyzed Enantio- and Regioselective Acylations. The Journal of
Organic Chemistry 2001, 66 (25), 8395-8401.
36. Forsyth, S. A.; MacFarlane, D. R.; Thomson, R. J.; von Itzstein, M., Rapid, clean,
and mild O-acetylation of alcohols and carbohydrates in an ionic liquid. Chemical
Communications 2002, (7), 714-715.
37. Weingartner, H.; Cabrele, C.; Herrmann, C., How ionic liquids can help to stabilize
native proteins. Physical Chemistry Chemical Physics 2012, 14 (2), 415-426.
38. Hofrichter, M., Review: lignin conversion by manganese peroxidase (MnP).
Enzyme and Microbial Technology 2002, 30 (4), 454-466.
39. Boerjan, W.; Ralph, J.; Baucher, M., Lignin biosynthesis. Annual Review of Plant
Biology 2003, 54, 33.
40. Weng, J.-K.; Chapple, C., The origin and evolution of lignin biosynthesis. New
Phytologist 2010, 187 (2), 273-285.
41. Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W., Lignin
Biosynthesis and Structure. Plant Physiology 2010, 153 (3), 895-905.
42. Stewart, J. J.; Akiyama, T.; Chapple, C.; Ralph, J.; Mansfield, S. D., The effects
on lignin structure of overexpression of ferulate 5-hydroxylase in hybrid poplar1. Plant
physiology 2009, 150 (2), 621-635.
43. Sette, M.; Wechselberger, R.; Crestini, C., Elucidation of Lignin Structure by
Quantitative 2D NMR. Chemistry – A European Journal 2011, 17 (34), 9529-9535.
44. Suhas; Carrott, P. J. M.; Ribeiro Carrott, M. M. L., Lignin – from natural adsorbent
to activated carbon: A review. Bioresource Technology 2007, 98 (12), 2301-2312.
45. Zeng, J.; Helms, G. L.; Gao, X.; Chen, S., Quantification of Wheat Straw Lignin
Structure by Comprehensive NMR Analysis. Journal of Agricultural and Food Chemistry
2013, 61 (46), 10848-10857.
46. Wilkerson, C. G.; Mansfield, S. D.; Lu, F.; Withers, S.; Park, J.-Y.; Karlen, S.
D.; Gonzales-Vigil, E.; Padmakshan, D.; Unda, F.; Rencoret, J.; Ralph, J., Monolignol
Ferulate Transferase Introduces Chemically Labile Linkages into the Lignin Backbone.
Science 2014, 344 (6179), 90-93.
47. Parsell, T.; Yohe, S.; Degenstein, J.; Jarrell, T.; Klein, I.; Gencer, E.; Hewetson,
B.; Hurt, M.; Kim, J. I.; Choudhari, H.; Saha, B.; Meilan, R.; Mosier, N.; Ribeiro, F.;
Delgass, W. N.; Chapple, C.; Kenttamaa, H. I.; Agrawal, R.; Abu-Omar, M. M., A
synergistic biorefinery based on catalytic conversion of lignin prior to cellulose starting
from lignocellulosic biomass. Green Chemistry 2015, 17 (3), 1492-1499.
48. Li, X.; Bonawitz, N. D.; Weng, J.-K.; Chapple, C., The Growth Reduction

148
Associated with Repressed Lignin Biosynthesis in <em>Arabidopsis thaliana</em> Is
Independent of Flavonoids. The Plant Cell 2010, 22 (5), 1620-1632.
49. Bonawitz, N. D.; Kim, J. I.; Tobimatsu, Y.; Ciesielski, P. N.; Anderson, N. A.;
Ximenes, E.; Maeda, J.; Ralph, J.; Donohoe, B. S.; Ladisch, M.; Chapple, C., Disruption
of Mediator rescues the stunted growth of a lignin-deficient Arabidopsis mutant. Nature
2014, 509 (7500), 376-380.
50. Shi, J.; Pattathil, S.; Parthasarathi, R.; Anderson, N. A.; Kim, J. I.;
Venketachalam, S.; Hahn, M. G.; Chapple, C.; Simmons, B. A.; Singh, S., Impact of
engineered lignin composition on biomass recalcitrance and ionic liquid pretreatment
efficiency. Green Chemistry 2016, 18 (18), 4884-4895.
51. Simmons, B. A.; Loqué, D.; Ralph, J., Advances in modifying lignin for enhanced
biofuel production. Current Opinion in Plant Biology 2010, 13 (3), 312-319.
52. Lora, J. H.; Glasser, W. G., Recent Industrial Applications of Lignin: A Sustainable
Alternative to Nonrenewable Materials. Journal of Polymers and the Environment 2002,
10 (1), 39-48.
53. Yang, H.; Xie, Y.; Zheng, X.; Pu, Y.; Huang, F.; Meng, X.; Wu, W.; Ragauskas,
A.; Yao, L., Comparative study of lignin characteristics from wheat straw obtained by soda-
AQ and kraft pretreatment and effect on the following enzymatic hydrolysis process.
Bioresource Technology 2016, 207, 361-369.
54. Li, C.; Zhao, X.; Wang, A.; Huber, G. W.; Zhang, T., Catalytic Transformation of
Lignin for the Production of Chemicals and Fuels. Chemical Reviews 2015, 115 (21),
11559-11624.
55. Rahimi, A.; Ulbrich, A.; Coon, J. J.; Stahl, S. S., Formic-acid-induced
depolymerization of oxidized lignin to aromatics. Nature 2014, 515 (7526), 249-252.
56. Molinari, V.; Clavel, G.; Graglia, M.; Antonietti, M.; Esposito, D., Mild
Continuous hydrogenolysis of kraft lignin over titanium nitride–nickel catalyst. ACS
Catalysis 2016, 6 (3), 1663-1670.
57. Deng, W.; Zhang, H.; Wu, X.; Li, R.; Zhang, Q.; Wang, Y., Oxidative conversion
of lignin and lignin model compounds catalyzed by CeO2-supported Pd nanoparticles.
Green Chemistry 2015, 17 (11), 5009-5018.
58. Barta, K.; Matson, T. D.; Fettig, M. L.; Scott, S. L.; Iretskii, A. V.; Ford, P. C.,
Catalytic disassembly of an organosolv lignin via hydrogen transfer from supercritical
methanol. Green Chemistry 2010, 12 (9), 1640-1647.
59. Regmi, Y. N.; Mann, J. K.; McBride, J. R.; Tao, J.; Barnes, C. E.; Labbé, N.;
Chmely, S. C., Catalytic transfer hydrogenolysis of organosolv lignin using B-containing
FeNi alloyed catalysts. Catalysis Today 2017.
60. Kim, K. H.; Simmons, B. A.; Singh, S., Catalytic transfer hydrogenolysis of ionic
liquid processed biorefinery lignin to phenolic compounds. Green Chemistry 2017, 19 (1),
215-224.
61. Yang, H.; Yan, R.; Chen, H.; Lee, D. H.; Zheng, C., Characteristics of
hemicellulose, cellulose and lignin pyrolysis. Fuel 2007, 86 (12-13), 1781-1788.
62. Khachatryan, L.; Barekati-Goudarzi, M.; Kekejian, D.; Aguilar, G.; Asatryan, R.;
Stanley, G. G.; Boldor, D., Pyrolysis of Lignin in Gas-Phase Isothermal and cw-CO2 Laser
Powered Non-Isothermal Reactors. Energy & Fuels 2018, 32 (12), 12597-12606.
63. Barekati-Goudarzi, M.; Boldor, D.; Marculescu, C.; Khachatryan, L., Peculiarities
of pyrolysis of hydrolytic lignin in dispersed gas phase and in solid state. Energy & Fuels

149
2017, 31 (11), 12156-12167.
64. Mullen, C. A.; Boateng, A. A.; Goldberg, N. M.; Lima, I. M.; Laird, D. A.; Hicks,
K. B., Bio-oil and bio-char production from corn cobs and stover by fast pyrolysis. Biomass
and bioenergy 2010, 34 (1), 67-74.
65. Martı́nez, A. T., Molecular biology and structure-function of lignin-degrading heme
peroxidases. Enzyme and Microbial Technology 2002, 30 (4), 425-444.
66. Dong, S.; Mao, L.; Luo, S.; Zhou, L.; Feng, Y.; Gao, S., Comparison of lignin
peroxidase and horseradish peroxidase for catalyzing the removal of nonylphenol from
water. Environmental Science and Pollution Research 2014, 21 (3), 2358-2366.
67. Nousiainen, P.; Kontro, J.; Manner, H.; Hatakka, A.; Sipilä, J., Phenolic mediators
enhance the manganese peroxidase catalyzed oxidation of recalcitrant lignin model
compounds and synthetic lignin. Fungal Genetics and Biology 2014, 72, 137-149.
68. Sahadevan, L. D. M.; Misra, C. S.; Thankamani, V., Characterization of lignin-
degrading enzymes (LDEs) from a dimorphic novel fungus and identification of products
of enzymatic breakdown of lignin. 3 Biotech 2016, 6 (1), 16.
69. Johansson, T.; Nyman, P. O., Isozymes of Lignin Peroxidase and Manganese(II)
Peroxidase from the White-Rot Basidiomycete Trametes versicolor. Archives of
Biochemistry and Biophysics 1993, 300 (1), 49-56.
70. Camarero, S.; Sarkar, S.; Ruiz-Dueñas, F. J.; Martı́nez, M. a. J.; Martı́nez, Á. T.,
Description of a Versatile Peroxidase Involved in the Natural Degradation of Lignin That
Has Both Manganese Peroxidase and Lignin Peroxidase Substrate Interaction Sites.
Journal of Biological Chemistry 1999, 274 (15), 10324-10330.
71. Glenn, J. K.; Morgan, M. A.; Mayfield, M. B.; Kuwahara, M.; Gold, M. H., An
extracellular H2O2-requiring enzyme preparation involved in lignin biodegradation by the
white rot basidiomycete Phanerochaete chrysosporium. Biochemical and Biophysical
Research Communications 1983, 114 (3), 1077-1083.
72. Blodig, W.; Smith, A. T.; Doyle, W. A.; Piontek, K., Crystal structures of pristine
and oxidatively processed lignin peroxidase expressed in Escherichia coli and of the
W171F variant that eliminates the redox active tryptophan 171. Implications for the
reaction mechanism1 1Edited by R. Huber. Journal of Molecular Biology 2001, 305 (4),
851-861.
73. Sundaramoorthy, M.; Kishi, K.; Gold, M. H.; Poulos, T. L., The crystal structure
of manganese peroxidase from Phanerochaete chrysosporium at 2.06-A resolution. Journal
of Biological Chemistry 1994, 269 (52), 32759-32767.
74. Perez-Boada, M.; Ruiz-Duenas, F. J.; Pogni, R.; Basosi, R.; Choinowski, T.;
Martínez, M. J.; Piontek, K.; Martínez, A. T., Versatile peroxidase oxidation of high redox
potential aromatic compounds: site-directed mutagenesis, spectroscopic and
crystallographic investigation of three long-range electron transfer pathways. Journal of
molecular biology 2005, 354 (2), 385-402.
75. Berman, H. M.; Battistuz, T.; Bhat, T. N.; Bluhm, W. F.; Bourne, P. E.; Burkhardt,
K.; Feng, Z.; Gilliland, G. L.; Iype, L.; Jain, S.; Fagan, P.; Marvin, J.; Padilla, D.;
Ravichandran, V.; Schneider, B.; Thanki, N.; Weissig, H.; Westbrook, J. D.; Zardecki,
C., The Protein Data Bank. Acta Crystallographica Section D 2002, 58 (6 Part 1), 899-907.
76. Fernández-Fueyo, E.; Ruiz-Dueñas, F. J.; Ferreira, P.; Floudas, D.; Hibbett, D.
S.; Canessa, P.; Larrondo, L.; James, T. Y.; Seelenfreund, D.; Lobos, S.; Polanco, R.;
Tello, M.; Honda, Y.; Watanabe, T.; Watanabe, T.; Ryu, J. S.; Kubicek, C. P.; Schmoll,

150
M.; Gaskell, J.; Hammel, K. E.; St. John, F. J.; Vanden Wymelenberg, A.; Sabat, G.;
Bondurant, S. S.; Syed, K.; Yadav, J.; Doddapaneni, H.; Subramanian, V.; Lavín, J. L.;
Oguiza, J. A., Comparative genomics of Ceriporiopisis subvermispora and Phanerochaete
chrysosporium provide insight into selective ligninolysis. Proc Natl Acad Sci U S A 2012,
109.
77. Coconi-Linares, N.; Magaña-Ortíz, D.; Guzmán-Ortiz, D. A.; Fernández, F.;
Loske, A. M.; Gómez-Lim, M. A., High-yield production of manganese peroxidase, lignin
peroxidase, and versatile peroxidase in Phanerochaete chrysosporium. Applied
Microbiology and Biotechnology 2014, 98 (22), 9283-9294.
78. Fujii, K.; Uemura, M.; Hayakawa, C.; Funakawa, S.; Kosaki, T., Environmental
control of lignin peroxidase, manganese peroxidase, and laccase activities in forest floor
layers in humid Asia. Soil Biology and Biochemistry 2013, 57, 109-115.
79. Min, K.; Gong, G.; Woo, H. M.; Kim, Y.; Um, Y., A dye-decolorizing peroxidase
from Bacillus subtilis exhibiting substrate-dependent optimum temperature for dyes and β-
ether lignin dimer. 2015, 5, 8245.
80. Semba, Y.; Ishida, M.; Yokobori, S.-i.; Yamagishi, A., Ancestral amino acid
substitution improves the thermal stability of recombinant lignin-peroxidase from white-
rot fungi, Phanerochaete chrysosporium strain UAMH 3641. Protein Engineering, Design
and Selection 2015, 28 (7), 221-230.
81. Tien, M.; Kirk, T. K., Lignin-degrading enzyme from the hymenomycete
Phanerochaete chrysosporium Burds. Science 1983, 221.
82. Huang, Z.; Liers, C.; Ullrich, R.; Hofrichter, M.; Urynowicz, M. A.,
Depolymerization and solubilization of chemically pretreated powder river basin
subbituminous coal by manganese peroxidase (MnP) from Bjerkandera adusta. Fuel 2013,
112, 295-301.
83. Bilal, M.; Asgher, M.; Iqbal, H. M. N.; Hu, H.; Zhang, X., Gelatin-Immobilized
Manganese Peroxidase with Novel Catalytic Characteristics and Its Industrial Exploitation
for Fruit Juice Clarification Purposes. Catalysis Letters 2016, 146 (11), 2221-2228.
84. Yoshida, H., LXIII.-Chemistry of lacquer (Urushi). Part I. Communication from the
Chemical Society of Tokio. Journal of the Chemical Society, Transactions 1883, 43 (0),
472-486.
85. An, H.; Xiao, T.; Fan, H.; Wei, D., Molecular characterization of a novel
thermostable laccase PPLCC2 from the brown rot fungus Postia placenta MAD-698-R.
Electronic Journal of Biotechnology 2015, 18 (6), 451-458.
86. Piontek, K.; Antorini, M.; Choinowski, T., Crystal Structure of a Laccase from the
FungusTrametes versicolor at 1.90-Å Resolution Containing a Full Complement of
Coppers. Journal of Biological Chemistry 2002, 277 (40), 37663-37669.
87. Thurston, C. F., The structure and function of fungal laccases. Microbiology 1994,
140 (1), 19-26.
88. Kurniawati, S.; Nicell, J. A., Characterization of Trametes versicolor laccase for the
transformation of aqueous phenol. Bioresource Technology 2008, 99 (16), 7825-7834.
89. Bourbonnais, R.; Paice, M.; Reid, I.; Lanthier, P.; Yaguchi, M., Lignin oxidation
by laccase isozymes from Trametes versicolor and role of the mediator 2, 2'-azinobis (3-
ethylbenzthiazoline-6-sulfonate) in kraft lignin depolymerization. Applied and
Environmental Microbiology 1995, 61 (5), 1876-1880.
90. Kiiskinen, L.-L.; Kruus, K.; Bailey, M.; Ylösmäki, E.; Siika-aho, M.; Saloheimo,

151
M., Expression of Melanocarpus albomyces laccase in Trichoderma reesei and
characterization of the purified enzyme. Microbiology 2004, 150 (9), 3065-3074.
91. Hullo, M.-F.; Moszer, I.; Danchin, A.; Martin-Verstraete, I., CotA of Bacillus
subtilis Is a Copper-Dependent Laccase. Journal of Bacteriology 2001, 183 (18), 5426-
5430.
92. Martins, L. O.; Soares, C. M.; Pereira, M. M.; Teixeira, M.; Costa, T.; Jones, G.
H.; Henriques, A. O., Molecular and biochemical characterization of a highly stable
bacterial laccase that occurs as a structural component of theBacillus subtilis endospore
coat. Journal of Biological Chemistry 2002, 277 (21), 18849-18859.
93. Miyazaki, K., A hyperthermophilic laccase from Thermus thermophilus HB27.
Extremophiles 2005, 9 (6), 415-425.
94. Uthandi, S.; Saad, B.; Humbard, M. A.; Maupin-Furlow, J. A., LccA, an Archaeal
Laccase Secreted as a Highly Stable Glycoprotein into the Extracellular Medium by
Haloferax volcanii. Applied and Environmental Microbiology 2010, 76 (3), 733-743.
95. Couto, R.; Herrera, S. T.; Luis, J., Industrial and biotechnological applications of
laccases: A review. Biotechnology Advances 2006, 24 (5), 500-513.
96. Muthukumarasamy, N. P.; Murugan, S., Production, Purification and Application
of Bacterial Laccase: A Review. Biotechnology 2014, 13 (5), 11.
97. Gamelas, J. A. F.; Tavares, A. P. M.; Evtuguin, D. V.; Xavier, A. M. B., Oxygen
bleaching of kraft pulp with polyoxometalates and laccase applying a novel multi-stage
process. Journal of Molecular Catalysis B: Enzymatic 2005, 33 (3), 57-64.
98. Campos, R.; Kandelbauer, A.; Robra, K. H.; Cavaco-Paulo, A.; Gübitz, G. M.,
Indigo degradation with purified laccases from Trametes hirsuta and Sclerotium rolfsii.
Journal of Biotechnology 2001, 89 (2), 131-139.
99. Pazarlıoǧlu, N. K.; Sariişik, M.; Telefoncu, A., Laccase: production by Trametes
versicolor and application to denim washing. Process Biochemistry 2005, 40 (5), 1673-
1678.
100. Lantto, R.; Schönberg, C.; Buchert, J.; Heine, E., Effects of Laccase-Mediator
Combinations on Wool. Textile Research Journal 2004, 74 (8), 713-717.
101. Basto, C.; Tzanov, T.; Cavaco-Paulo, A., Combined ultrasound-laccase assisted
bleaching of cotton. Ultrasonics Sonochemistry 2007, 14 (3), 350-354.
102. Novotný, Č.; Svobodová, K.; Kasinath, A.; Erbanová, P., Biodegradation of
synthetic dyes by Irpex lacteus under various growth conditions. International
Biodeterioration & Biodegradation 2004, 54 (2), 215-223.
103. Pointing, S., Feasibility of bioremediation by white-rot fungi. Applied
Microbiology and Biotechnology 2001, 57 (1), 20-33.
104. Anastasi, A.; Coppola, T.; Prigione, V.; Varese, G. C., Pyrene degradation and
detoxification in soil by a consortium of basidiomycetes isolated from compost: Role of
laccases and peroxidases. Journal of Hazardous Materials 2009, 165 (1), 1229-1233.
105. Niku-Paavola, M. L.; Viikari, L., Enzymatic oxidation of alkenes. Journal of
Molecular Catalysis B: Enzymatic 2000, 10 (4), 435-444.
106. Alcalde, M.; Ferrer, M.; Plou, F. J.; Ballesteros, A., Environmental biocatalysis:
from remediation with enzymes to novel green processes. Trends in Biotechnology 2006,
24 (6), 281-287.
107. Alper, N.; Acar, J., Removal of phenolic compounds in pomegranate juices using
ultrafiltration and laccase-ultrafiltration combinations. Food / Nahrung 2004, 48 (3), 184-

152
187.
108. Selinheimo, E.; Kruus, K.; Buchert, J.; Hopia, A.; Autio, K., Effects of laccase,
xylanase and their combination on the rheological properties of wheat doughs. Journal of
Cereal Science 2006, 43 (2), 152-159.
109. Minussi, R. C.; Pastore, G. M.; Durán, N., Laccase induction in fungi and
laccase/N–OH mediator systems applied in paper mill effluent. Bioresource Technology
2007, 98 (1), 158-164.
110. Nicotra, S.; Cramarossa, M. R.; Mucci, A.; Pagnoni, U. M.; Riva, S.; Forti, L.,
Biotransformation of resveratrol: synthesis of trans-dehydrodimers catalyzed by laccases
from Myceliophtora thermophyla and from Trametes pubescens. Tetrahedron 2004, 60 (3),
595-600.
111. Harris, Z. L.; Davis-Kaplan, S. R.; Gitlin, J. D.; Kaplan, J., A fungal multicopper
oxidase restores iron homeostasis in aceruloplasminemia. Blood 2004, 103 (12), 4672-
4673.
112. Chen, Q.; Marshall, M. N.; Geib, S. M.; Tien, M.; Richard, T. L., Effects of laccase
on lignin depolymerization and enzymatic hydrolysis of ensiled corn stover. Bioresource
Technology 2012, 117, 186-192.
113. Li, Q.; Xie, S.; Serem, W. K.; Naik, M. T.; Liu, L.; Yuan, J. S., Quality carbon
fibers from fractionated lignin. Green Chemistry 2017, 19 (7), 1628-1634.
114. Zhao, C.; Xie, S.; Pu, Y.; Zhang, R.; Huang, F.; Ragauskas, A. J.; Yuan, J. S.,
Synergistic enzymatic and microbial lignin conversion. Green Chemistry 2016, 18 (5),
1306-1312.
115. Mani, P.; Keshavarz, T.; Chandra, T. S.; Kyazze, G., Decolourisation of Acid
orange 7 in a microbial fuel cell with a laccase-based biocathode: Influence of mitigating
pH changes in the cathode chamber. Enzyme and Microbial Technology 2017, 96, 170-176.
116. Le Goff, A.; Holzinger, M.; Cosnier, S., Recent progress in oxygen-reducing
laccase biocathodes for enzymatic biofuel cells. Cellular and Molecular Life Sciences
2015, 72 (5), 941-952.
117. Bugg, T. D.; Ahmad, M.; Hardiman, E. M.; Singh, R., The emerging role for
bacteria in lignin degradation and bio-product formation. Current opinion in biotechnology
2011, 22 (3), 394-400.
118. de Gonzalo, G.; Colpa, D. I.; Habib, M. H.; Fraaije, M. W., Bacterial enzymes
involved in lignin degradation. Journal of biotechnology 2016, 236, 110-119.
119. Masai, E.; Katayama, Y.; Fukuda, M., Genetic and Biochemical Investigations on
Bacterial Catabolic Pathways for Lignin-Derived Aromatic Compounds. Bioscience,
Biotechnology, and Biochemistry 2007, 71 (1), 1-15.
120. Masai, E.; Ichimura, A.; Sato, Y.; Miyauchi, K.; Katayama, Y.; Fukuda, M., Roles
of the Enantioselective Glutathione S-Transferases in Cleavage of β-Aryl Ether. Journal of
Bacteriology 2003, 185 (6), 1768-1775.
121. Pereira, J. H.; Heins, R. A.; Gall, D. L.; McAndrew, R. P.; Deng, K.; Holland,
K. C.; Donohue, T. J.; Noguera, D. R.; Simmons, B. A.; Sale, K. L.; Ralph, J.; Adams,
P. D., Structural and Biochemical Characterization of the Early and Late Enzymes in the
Lignin β-Aryl Ether Cleavage Pathway from Sphingobium sp. SYK-6. Journal of
Biological Chemistry 2016, 291 (19), 10228-10238.
122. Masai, E.; Kamimura, N.; Kasai, D.; Oguchi, A.; Ankai, A.; Fukui, S.; Takahashi,
M.; Yashiro, I.; Sasaki, H.; Harada, T.; Nakamura, S.; Katano, Y.; Narita-Yamada, S.;

153
Nakazawa, H.; Hara, H.; Katayama, Y.; Fukuda, M.; Yamazaki, S.; Fujita, N., Complete
Genome Sequence of Sphingobium sp. Strain SYK-6, a Degrader of Lignin-Derived
Biaryls and Monoaryls. Journal of Bacteriology 2012, 194 (2), 534-535.
123. Mnich, E.; Vanholme, R.; Oyarce, P.; Liu, S.; Lu, F.; Goeminne, G.; Jørgensen,
B.; Motawie, M. S.; Boerjan, W.; Ralph, J.; Ulvskov, P.; Møller, B. L.; Bjarnholt, N.;
Harholt, J., Degradation of lignin β-aryl ether units in Arabidopsis thaliana expressing
LigD, LigF and LigG from Sphingomonas paucimobilis SYK-6. Plant Biotechnology
Journal 2017, 15 (5), 581-593.
124. Wu, W.; Dutta, T.; Varman, A. M.; Eudes, A.; Manalansan, B.; Loqué, D.; Singh,
S., Lignin Valorization: Two Hybrid Biochemical Routes for the Conversion of Polymeric
Lignin into Value-added Chemicals. Scientific Reports 2017, 7 (1), 8420.
125. Wu, W.; Liu, F.; Singh, S., Toward engineering <em>E. coli</em> with
an autoregulatory system for lignin valorization. Proceedings of the National Academy of
Sciences 2018.
126. Freemantle, M., Ionic liquids may boost clean technology development. Chemical
& Engineering News Archive 1998, 76 (13), 32-37.
127. Rogers, R. D.; Seddon, K. R., Ionic Liquids--Solvents of the Future? Science 2003,
302 (5646), 792-793.
128. Brennecke, J. F.; Maginn, E. J., Ionic liquids: Innovative fluids for chemical
processing. AIChE Journal 2001, 47 (11), 2384-2389.
129. Cvjetko Bubalo, M.; Hanousek, K.; Radošević, K.; Gaurina Srček, V.;
Jakovljević, T.; Radojčić Redovniković, I., Imidiazolium based ionic liquids: Effects of
different anions and alkyl chains lengths on the barley seedlings. Ecotoxicology and
Environmental Safety 2014, 101, 116-123.
130. Smiglak, M.; Reichert, W. M.; Holbrey, J. D.; Wilkes, J. S.; Sun, L.; Thrasher,
J. S.; Kirichenko, K.; Singh, S.; Katritzky, A. R.; Rogers, R. D., Combustible ionic liquids
by design: is laboratory safety another ionic liquid myth? Chemical Communications 2006,
(24), 2554-2556.
131. Gutowski, K. E.; Broker, G. A.; Willauer, H. D.; Huddleston, J. G.; Swatloski, R.
P.; Holbrey, J. D.; Rogers, R. D., Controlling the Aqueous Miscibility of Ionic Liquids:
Aqueous Biphasic Systems of Water-Miscible Ionic Liquids and Water-Structuring Salts
for Recycle, Metathesis, and Separations. Journal of the American Chemical Society 2003,
125 (22), 6632-6633.
132. van Rantwijk, F.; Sheldon, R. A., Biocatalysis in Ionic Liquids. Chemical Reviews
2007, 107 (6), 2757-2785.
133. Patel, R.; Kumari, M.; Khan, A. B., Recent Advances in the Applications of Ionic
Liquids in Protein Stability and Activity: A Review. Applied Biochemistry and
Biotechnology 2014, 172 (8), 3701-3720.
134. Kamlet, M. J.; Taft, R., The solvatochromic comparison method. I. The. beta.-scale
of solvent hydrogen-bond acceptor (HBA) basicities. Journal of the American Chemical
Society 1976, 98 (2), 377-383.
135. Reichardt, C., Pyridinium N-phenolate betaine dyes as empirical indicators of
solvent polarity: Some new findings. Pure and applied chemistry 2004, 76 (10), 1903-
1919.
136. Brandt, A.; Ray, M. J.; To, T. Q.; Leak, D. J.; Murphy, R. J.; Welton, T., Ionic
liquid pretreatment of lignocellulosic biomass with ionic liquid-water mixtures. Green

154
Chemistry 2011, 13 (9), 2489-2499.
137. Ab Rani, M.; Brant, A.; Crowhurst, L.; Dolan, A.; Lui, M.; Hassan, N.; Hallett,
J.; Hunt, P.; Niedermeyer, H.; Perez-Arlandis, J., Understanding the polarity of ionic
liquids. Physical Chemistry Chemical Physics 2011, 13 (37), 16831-16840.
138. Xu, A.; Wang, J.; Wang, H., Effects of anionic structure and lithium salts addition
on the dissolution of cellulose in 1-butyl-3-methylimidazolium-based ionic liquid solvent
systems. Green Chemistry 2010, 12 (2), 268-275.
139. Doherty, T. V.; Mora-Pale, M.; Foley, S. E.; Linhardt, R. J.; Dordick, J. S., Ionic
liquid solvent properties as predictors of lignocellulose pretreatment efficacy. Green Chem.
2010, 12 (11), 1967-1975.
140. Sun, N.; Parthasarathi, R.; Socha, A. M.; Shi, J.; Zhang, S.; Stavila, V.; Sale, K.
L.; Simmons, B. A.; Singh, S., Understanding pretreatment efficacy of four cholinium and
imidazolium ionic liquids by chemistry and computation. Green Chemistry 2014.
141. Hauru, L. K. J.; Hummel, M.; King, A. W. T.; Kilpeläinen, I. A.; Sixta, H., Role
of solvent parameters in the regeneration of cellulose from ionic liquid solutions.
Biomacromolecules 2012.
142. Kahlen, J.; Leonhard, K., Modelling cellulose solubilities in ionic liquids using
COSMO-RS. Green Chemistry 2010, 12 (12), 2172-2181.
143. Freire, M. G.; Santos, L. M. N. B. F.; Fernandes, A. M.; Coutinho, J. A. P.;
Marrucho, I. M., An overview of the mutual solubilities of water–imidazolium-based ionic
liquids systems. Fluid Phase Equilibria 2007, 261 (1), 449-454.
144. Hajime, M.; Masahiro, Y.; Kazumi, T.; Masakatsu, N.; Yukiko, K.; Yoshinori, M.,
Highly Conductive Room Temperature Molten Salts Based on Small
Trimethylalkylammonium Cations and Bis(trifluoromethylsulfonyl)imide. Chemistry
Letters 2000, 29 (8), 922-923.
145. Bhatt, A. I.; May, I.; Volkovich, V. A.; Hetherington, M. E.; Lewin, B.; Thied,
R. C.; Ertok, N., Group 15 quaternary alkyl bistriflimides: ionic liquids with potential
application in electropositive metal deposition and as supporting electrolytes. Journal of
the Chemical Society, Dalton Transactions 2002, (24), 4532-4534.
146. Howlett, P. C.; MacFarlane, D. R.; Hollenkamp, A. F., High Lithium Metal Cycling
Efficiency in a Room-Temperature Ionic Liquid. Electrochemical and Solid-State Letters
2004, 7 (5), A97-A101.
147. MacFarlane, D. R.; Tachikawa, N.; Forsyth, M.; Pringle, J. M.; Howlett, P. C.;
Elliott, G. D.; Davis, J. H.; Watanabe, M.; Simon, P.; Angell, C. A., Energy applications
of ionic liquids. Energy & Environmental Science 2014, 7 (1), 232-250.
148. Liu, H.; Wu, X.; Sun, J.; Chen, S., Stimulation of Laccase Biocatalysis in Ionic
Liquids: A Review on Recent Progress. Current Protein and Peptide Science 2018, 19 (1),
100-111.
149. Rehmann, L.; Ivanova, E.; Ferguson, J. L.; Gunaratne, H. Q. N.; Seddon, K. R.;
Stephens, G. M., Measuring the effect of ionic liquids on laccase activity using a simple,
parallel method. Green Chemistry 2012, 14 (3), 725-733.
150. Galai, S.; P. de los Rios, A.; Hernandez-Fernandez, F. J.; Haj Kacem, S.; Tomas-
Alonso, F., Over-activity and stability of laccase using ionic liquids: screening and
application in dye decolorization. RSC Advances 2015, 5 (21), 16173-16189.
151. Burney, P. R.; Pfaendtner, J., Structural and dynamic features of Candida rugosa
lipase 1 in water, octane, toluene, and ionic liquids BMIM-PF6 and BMIM-NO3. The

155
Journal of Physical Chemistry B 2013, 117 (9), 2662-2670.
152. Ranke, J.; Müller, A.; Bottin-Weber, U.; Stock, F.; Stolte, S.; Arning, J.;
Störmann, R.; Jastorff, B., Lipophilicity parameters for ionic liquid cations and their
correlation to in vitro cytotoxicity. Ecotoxicology and environmental safety 2007, 67 (3),
430-438.
153. Stolte, S.; Arning, J.; Bottin-Weber, U.; Matzke, M.; Stock, F.; Thiele, K.;
Uerdingen, M.; Welz-Biermann, U.; Jastorff, B.; Ranke, J., Anion effects on the
cytotoxicity of ionic liquids. Green Chemistry 2006, 8 (7), 621-629.
154. Couling, D. J.; Bernot, R. J.; Docherty, K. M.; Dixon, J. K.; Maginn, E. J.,
Assessing the factors responsible for ionic liquid toxicity to aquatic organisms via
quantitative structure–property relationship modeling. Green Chemistry 2006, 8 (1), 82-
90.
155. Yoo, B.; Jing, B.; Jones, S. E.; Lamberti, G. A.; Zhu, Y.; Shah, J. K.; Maginn, E.
J., Molecular mechanisms of ionic liquid cytotoxicity probed by an integrated experimental
and computational approach. Scientific Reports 2016, 6, 19889.
156. Achinivu, E. C.; Howard, R. M.; Li, G.; Gracz, H.; Henderson, W. A., Lignin
extraction from biomass with protic ionic liquids. Green Chemistry 2014, 16 (3), 1114-
1119.
157. Haghighi Mood, S.; Hossein Golfeshan, A.; Tabatabaei, M.; Salehi Jouzani, G.;
Najafi, G. H.; Gholami, M.; Ardjmand, M., Lignocellulosic biomass to bioethanol, a
comprehensive review with a focus on pretreatment. Renewable and Sustainable Energy
Reviews 2013, 27, 77-93.
158. Alvira, P.; Tomas-Pejo, E.; Ballesteros, M.; Negro, M. J., Pretreatment
technologies for an efficient bioethanol production process based on enzymatic hydrolysis:
a review. Bioresour Technol 2010, 101.
159. Casas, A.; Omar, S.; Palomar, J.; Oliet, M.; Alonso, M. V.; Rodriguez, F., Relation
between differential solubility of cellulose and lignin in ionic liquids and activity
coefficients. RSC Advances 2013, 3 (10), 3453-3460.
160. Pinkert, A.; Marsh, K. N.; Pang, S., Reflections on the Solubility of Cellulose.
Industrial & Engineering Chemistry Research 2010, 49 (22), 11121-11130.
161. Kosan, B.; Schwikal, K.; Meister, F., Solution states of cellulose in selected direct
dissolution agents. Cellulose 2010, 17 (3), 495-506.
162. Liu, Q.-P.; Hou, X.-D.; Li, N.; Zong, M.-H., Ionic liquids from renewable
biomaterials: synthesis, characterization and application in the pretreatment of biomass.
Green Chemistry 2012, 14 (2), 304-307.
163. Hou, X.-D.; Li, N.; Zong, M.-H., Renewable bio ionic liquids-water mixtures-
mediated selective removal of lignin from rice straw: Visualization of changes in
composition and cell wall structure. Biotechnology and Bioengineering 2013, 110 (7),
1895-1902.
164. Hou, X.-D.; Xu, J.; Li, N.; Zong, M.-H., Effect of anion structures on cholinium
ionic liquids pretreatment of rice straw and the subsequent enzymatic hydrolysis.
Biotechnology and Bioengineering 2015, 112 (1), 65-73.
165. da Costa Lopes, A. M.; João, K. G.; Rubik, D. F.; Bogel-Łukasik, E.; Duarte, L.
C.; Andreaus, J.; Bogel-Łukasik, R., Pre-treatment of lignocellulosic biomass using ionic
liquids: Wheat straw fractionation. Bioresource Technology 2013, 142, 198-208.
166. Lee, S. H.; Doherty, T. V.; Linhardt, R. J.; Dordick, J. S., Ionic liquid-mediated

156
selective extraction of lignin from wood leading to enhanced enzymatic cellulose
hydrolysis. Biotechnology and Bioengineering 2009, 102 (5), 1368-1376.
167. George, A.; Brandt, A.; Tran, K.; Zahari, S. M. N. S.; Klein-Marcuschamer, D.;
Sun, N.; Sathitsuksanoh, N.; Shi, J.; Stavila, V.; Parthasarathi, R., Design of low-cost
ionic liquids for lignocellulosic biomass pretreatment. Green Chemistry 2015, 17 (3), 1728-
1734.
168. Domínguez de María, P., Recent trends in (ligno)cellulose dissolution using
neoteric solvents: switchable, distillable and bio-based ionic liquids. Journal of Chemical
Technology & Biotechnology 2014, 89 (1), 11-18.
169. Hou, X.-D.; Smith, T. J.; Li, N.; Zong, M.-H., Novel renewable ionic liquids as
highly effective solvents for pretreatment of rice straw biomass by selective removal of
lignin. Biotechnology and Bioengineering 2012, 109 (10), 2484-2493.
170. Sun, J.; Konda, N. M.; Shi, J.; Parthasarathi, R.; Dutta, T.; Xu, F.; Scown, C.
D.; Simmons, B. A.; Singh, S., CO 2 enabled process integration for the production of
cellulosic ethanol using bionic liquids. Energy & Environmental Science 2016, 9 (9), 2822-
2834.
171. Shi, J.; Balamurugan, K.; Parthasarathi, R.; Sathitsuksanoh, N.; Zhang, S.;
Stavila, V.; Subramanian, V.; Simmons, B. A.; Singh, S., Understanding the role of water
during ionic liquid pretreatment of lignocellulose: co-solvent or anti-solvent? Green
Chemistry 2014, 16 (8), 3830-3840.
172. Liu, E.; Li, M.; Das, L.; Pu, Y.; Frazier, T.; Zhao, B.; Crocker, M.; Ragauskas,
A. J.; Shi, J., Understanding Lignin Fractionation and Characterization from Engineered
Switchgrass Treated by an Aqueous Ionic Liquid. ACS Sustainable Chemistry &
Engineering 2018, 6 (5), 6612-6623.
173. Mai, N. L.; Ahn, K.; Koo, Y. M., Methods for recovery of ionic liquids-A review.
Process Biochemistry 2014, 49 (5), 872-881.
174. Wang, X. L.; Nie, Y.; Zhang, X. P.; Zhang, S. J.; Li, J. W., Recovery of ionic
liquids from dilute aqueous solutions by electrodialysis. Desalination 2012, 285, 205-212.
175. Sun, J.; Shi, J.; Konda, N. M.; Campos, D.; Liu, D.; Nemser, S.; Shamshina, J.;
Dutta, T.; Berton, P.; Gurau, G., Efficient dehydration and recovery of ionic liquid after
lignocellulosic processing using pervaporation. Biotechnology for biofuels 2017, 10 (1),
154.
176. Singh, S.; Simmons, B. A., Ionic Liquid Pretreatment: Mechanism, Performance,
and Challenges. In Aqueous Pretreatment of Plant Biomass for Biological and Chemical
Conversion to Fuels and Chemicals, John Wiley & Sons, Ltd: 2013; pp 223-238.
177. Klein-Marcuschamer, D.; Simmons, B. A.; Blanch, H. W., Techno-economic
analysis of a lignocellulosic ethanol biorefinery with ionic liquid pre-treatment. Biofuels,
Bioproducts and Biorefining 2011, 5 (5), 562-569.
178. Konda, N. M.; Shi, J.; Singh, S.; Blanch, H. W.; Simmons, B. A.; Klein-
Marcuschamer, D., Understanding cost drivers and economic potential of two variants of
ionic liquid pretreatment for cellulosic biofuel production. Biotechnology for Biofuels
2014, 7 (1), 86.
179. Wen, J.-L.; Yuan, T.-Q.; Sun, S.-L.; Xu, F.; Sun, R.-C., Understanding the
chemical transformations of lignin during ionic liquid pretreatment. Green Chemistry 2014,
16 (1), 181-190.
180. Cox, B. J.; Jia, S.; Zhang, Z. C.; Ekerdt, J. G., Catalytic degradation of lignin

157
model compounds in acidic imidazolium based ionic liquids: Hammett acidity and anion
effects. Polymer Degradation and Stability 2011, 96 (4), 426-431.
181. Bose, S.; Armstrong, D. W.; Petrich, J. W., Enzyme-Catalyzed Hydrolysis of
Cellulose in Ionic Liquids: A Green Approach Toward the Production of Biofuels. The
Journal of Physical Chemistry B 2010, 114 (24), 8221-8227.
182. Alberto Domínguez, O. R., Ana Paula M. Tavares, Eugenia A. Macedo, Ma.
Asunción Longo, Ma. Ángeles Sanromán, Studies of laccase from Trametes versicolor in
aqueous solutions of several methylimidazolium ionic liquids. Bioresource Technology
2011, 102 (16), 6.
183. Harwardt, N.; Stripling, N.; Roth, S.; Liu, H.; Schwaneberg, U.; Spiess, A. C.,
Effects of ionic liquids on the reaction kinetics of a laccase–mediator system. RSC
Advances 2014, 4, 8.
184. Lozano, P.; de Diego, T.; Guegan, J.-P.; Vaultier, M.; Iborra, J. L., Stabilization of
α-chymotrypsin by ionic liquids in transesterification reactions. Biotechnology and
Bioengineering 2001, 75 (5), 563-569.
185. Lai, J.-Q.; Hu, Z.-L.; Wang, P.-W.; Yang, Z., Enzymatic production of microalgal
biodiesel in ionic liquid [BMIm][PF6]. Fuel 2012, 95, 329-333.
186. Attri, P.; Venkatesu, P.; Kumar, A., Activity and stability of [small alpha]-
chymotrypsin in biocompatible ionic liquids: enzyme refolding by triethyl ammonium
acetate. Physical Chemistry Chemical Physics 2011, 13 (7), 2788-2796.
187. Arai, S.; Nakashima, K.; Tanino, T.; Ogino, C.; Kondo, A.; Fukuda, H.,
Production of biodiesel fuel from soybean oil catalyzed by fungus whole-cell biocatalysts
in ionic liquids. Enzyme and Microbial Technology 2010, 46 (1), 51-55.
188. Hekmat, D.; Hebel, D.; Joswig, S.; Schmidt, M.; Weuster-Botz, D., Advanced
protein crystallization using water-soluble ionic liquids as crystallization additives.
Biotechnology Letters 2007, 29 (11), 1703-1711.
189. S. Galai, A. P. d. l. R., F. J. Hernandez-Fernandez, S. Haj Kacemb and F. Tomas-
Alonso, Over-activity and stability of laccase using ionic liquids: screening and application
in dye decolorization. RSC Advances 2015, 5, 17.
190. Wang, Y.; Radosevich, M.; Hayes, D.; Labbé, N., Compatible Ionic liquid-
cellulases system for hydrolysis of lignocellulosic biomass. Biotechnology and
Bioengineering 2011, 108 (5), 1042-1048.
191. Tavares, A. P. M.; Rodriguez, O.; Macedo, E. A., Ionic Liquids as Alternative Co-
Solvents for Laccase: Study of Enzyme Activity and Stability. Biotechnology and
Bioengineering 2008, 101 (1), 7.
192. Constatinescu, D.; Herrmann, C.; Weingartner, H., Patterns of protein unfolding
and protein aggregation in ionic liquids. Physical Chemistry Chemical Physics 2010, 12
(8), 1756-1763.
193. Domínguez, A.; Rodríguez, O.; Tavares, A. P. M.; Macedo, E. A.; Longo, M. A.;
Sanromán, M. Á., Studies of laccase from Trametes versicolor in aqueous solutions of
several methylimidazolium ionic liquids. Bioresource technology 2011, 102 (16), 7494-
7499.
194. Zhao, H.; Song, Z., Nuclear magnetic relaxation of water in ionic-liquid solutions:
determining the kosmotropicity of ionic liquids and its relationship with the enzyme
enantioselectivity. Journal of Chemical Technology & Biotechnology 2007, 82 (3), 304-
312.

158
195. de los Ríos, A. P.; Hernández Fernández, F. J.; Gómez, D.; Rubio, M.; Víllora, G.,
Biocatalytic transesterification of sunflower and waste cooking oils in ionic liquid media.
Process Biochemistry 2011, 46 (7), 1475-1480.
196. Lai, J.-Q.; Li, Z.; Lü, Y.-H.; Yang, Z., Specific ion effects of ionic liquids on
enzyme activity and stability. Green Chemistry 2011, 13 (7), 1860-1868.
197. Bickerstaff, G. F., Immobilization of enzymes and cells. In Immobilization of
enzymes and cells, Springer: 1997; pp 1-11.
198. Fernández-Fernández, M.; Moldes, D.; Domínguez, A.; Sanromán, M. Á.;
Tavares, A. P. M.; Rodríguez, O.; Macedo, E. A., Stability and kinetic behavior of
immobilized laccase from Myceliophthora thermophila in the presence of the ionic liquid
1-ethyl-3-methylimidazolium ethylsulfate. Biotechnology Progress 2014, 30 (4), 790-796.
199. Datta, S.; Holmes, B.; Park, J. I.; Chen, Z.; Dibble, D. C.; Hadi, M.; Blanch, H.
W.; Simmons, B. A.; Sapra, R., Ionic liquid tolerant hyperthermophilic cellulases for
biomass pretreatment and hydrolysis. Green Chemistry 2010, 12 (2), 338-345.
200. Zhang, T.; Datta, S.; Eichler, J.; Ivanova, N.; Axen, S. D.; Kerfeld, C. A.; Chen,
F.; Kyrpides, N.; Hugenholtz, P.; Cheng, J.-F., Identification of a haloalkaliphilic and
thermostable cellulase with improved ionic liquid tolerance. Green Chemistry 2011, 13 (8),
2083-2090.
201. Gunny, A. A. N.; Arbain, D.; Edwin Gumba, R.; Jong, B. C.; Jamal, P., Potential
halophilic cellulases for in situ enzymatic saccharification of ionic liquids pretreated
lignocelluloses. Bioresource Technology 2014, 155, 177-181.
202. Nordwald, E. M.; Brunecky, R.; Himmel, M. E.; Beckham, G. T.; Kaar, J. L.,
Charge engineering of cellulases improves ionic liquid tolerance and reduces lignin
inhibition. Biotechnology and Bioengineering 2014, 111 (8), 1541-1549.
203. Burney, P. R.; Nordwald, E. M.; Hickman, K.; Kaar, J. L.; Pfaendtner, J.,
Molecular dynamics investigation of the ionic liquid/enzyme interface: Application to
engineering enzyme surface charge. Proteins: Structure, Function, and Bioinformatics
2015, 83 (4), 670-680.
204. Brogan, A. P.; Hallett, J. P., Solubilizing and stabilizing proteins in anhydrous ionic
liquids through formation of protein–polymer surfactant nanoconstructs. Journal of the
American Chemical Society 2016, 138 (13), 4494-4501.
205. Brogan, A. P.; Bui-Le, L.; Hallett, J. P., Non-aqueous homogenous biocatalytic
conversion of polysaccharides in ionic liquids using chemically modified glucosidase.
Nature Chemistry 2018, 10 (8), 859.
206. Dwivedi, U. N.; Singh, P.; Pandey, V. P.; Kumar, A., Structure–function
relationship among bacterial, fungal and plant laccases. Journal of Molecular Catalysis B:
Enzymatic 2011, 68 (2), 117-128.
207. Hildén, K.; Hakala, T. K.; Lundell, T., Thermotolerant and thermostable laccases.
Biotechnology Letters 2009, 31 (8), 1117.
208. Sollewijn Gelpke, M. D.; Lee, J.; Gold, M. H., Lignin Peroxidase Oxidation of
Veratryl Alcohol: Effects of the Mutants H82A, Q222A, W171A, and F267L.
Biochemistry 2002, 41 (10), 3498-3506.
209. Ohtsubo, M.; Noguchi, S.; Takeda, K.; Morohashi, M.; Kawamura, M., Site-
directed mutagenesis of Asp-376, the catalytic phosphorylation site, and Lys-507, the
putative ATP-binding site, of the α-subunit of Torpedo californicaNa+ K+-ATPase.
Biochimica et Biophysica Acta (BBA)-Biomembranes 1990, 1021 (2), 157-160.

159
210. Prokop, J. W.; Duff, R. J.; Ball, H. C.; Copeland, D. L.; Londraville, R. L., Leptin
and leptin receptor: Analysis of a structure to function relationship in interaction and
evolution from humans to fish. Peptides 2012, 38 (2), 326-336.
211. Chao, H.; DeLuca, C. I.; Davies, P. L.; Sykes, B. D.; Sönnichsen, F. D., Structure-
function relationship in the globular type III antifreeze protein: Identification of a cluster
of surface residues required for binding to ice. Protein Science 1994, 3 (10), 1760-1769.
212. Smith, A. T.; Doyle, W. A.; Dorlet, P.; Ivancich, A., Spectroscopic evidence for an
engineered, catalytically active Trp radical that creates the unique reactivity of lignin
peroxidase. Proceedings of the National Academy of Sciences 2009, 106 (38), 16084-
16089.
213. Fukuda, K.; Morioka, H.; Imajou, S.; Ikeda, S.; Ohtsuka, E.; Tsurimoto, T.,
Structure-Function Relationship of the Eukaryotic DNA Replication Factor, Proliferating
Cell Nuclear Antigen. Journal of Biological Chemistry 1995, 270 (38), 22527-22534.
214. Owsianik, G.; D'hoedt, D.; Voets, T.; Nilius, B., Structure–function relationship of
the TRP channel superfamily. In Reviews of Physiology, Biochemistry and Pharmacology,
Amara, S. G.; Bamberg, E.; Grinstein, S.; Hebert, S. C.; Jahn, R.; Lederer, W. J.; Lill,
R.; Miyajima, A.; Murer, H.; Offermanns, S.; Schultz, G.; Schweiger, M., Eds. Springer
Berlin Heidelberg: Berlin, Heidelberg, 2006; pp 61-90.
215. Culver, J. N.; Stubbs, G.; Dawson, W. O., Structure-function Relationship Between
Tobacco Mosaic Virus Coat Protein and Hypersensitivity in Nicotiana sylvestris. Journal
of Molecular Biology 1994, 242 (2), 130-138.
216. Jarosch, M.; Egelseer, E. M.; Huber, C.; Moll, D.; Mattanovich, D.; Sleytr, U.
B.; Sára, M., Analysis of the structure–function relationship of the S-layer protein SbsC of
Bacillus stearothermophilus ATCC 12980 by producing truncated forms. Microbiology
2001, 147 (5), 1353-1363.
217. Katritch, V.; Cherezov, V.; Stevens, R. C., Structure-Function of the G Protein-
Coupled Receptor Superfamily. Annual Review of Pharmacology and Toxicology 2013, 53,
6.
218. Warner, L.; Gjersing, E.; Follett, S. E.; Elliott, K. W.; Dzyuba, S. V.; Varga, K.,
The effects of high concentrations of ionic liquid on GB1 protein structure and dynamics
probed by high-resolution magic-angle-spinning NMR spectroscopy. Biochemistry and
biophysics reports 2016, 8, 75-80.
219. Bornscheuer, U. T.; Pohl, M., Improved biocatalysts by directed evolution and
rational protein design. Current Opinion in Chemical Biology 2001, 5 (2), 137-143.
220. Arnold, F. H., Design by directed evolution. Accounts of chemical research 1998,
31 (3), 125-131.
221. Martin, A.; Sieber, V.; Schmid, F. X., In-vitro Selection of Highly Stabilized
Protein Variants with Optimized Surface. Journal of Molecular Biology 2001, 309 (3), 717-
726.
222. Mate, D. M.; Alcalde, M., Laccase engineering: From rational design to directed
evolution. Biotechnology Advances 2015, 33 (1), 25-40.
223. Fernández-Fueyo, E.; Ruiz-Dueñas, F. J.; Martínez, A. T., Engineering a fungal
peroxidase that degrades lignin at very acidic pH. Biotechnology for Biofuels 2014, 7 (1),
114.
224. Khodakarami, A.; Goodarzi, N.; Hoseinzadehdehkordi, M.; Amani, F.;
Khodaverdian, S.; Khajeh, K.; Ghazi, F.; Ranjbar, B.; Amanlou, M.; Dabirmanesh, B.,

160
Rational design toward developing a more efficient laccase: Catalytic efficiency and
selectivity. International journal of biological macromolecules 2018, 112, 775-779.
225. Wallraf, A.-M.; Liu, H.; Zhu, L.; Khalfallah, G.; Simons, C.; Alibiglou, H.;
Davari, M. D.; Schwaneberg, U., A loop engineering strategy improves laccase lcc2
activity in ionic liquid and aqueous solution. Green Chemistry 2018.
226. Judge, R. A.; Takahashi, S.; Longenecker, K. L.; Fry, E. H.; Abad-Zapatero, C.;
Chiu, M. L., The effect of ionic liquids on protein crystallization and X-ray diffraction
resolution. Crystal Growth and Design 2009, 9 (8), 3463-3469.
227. Kennedy, D. F.; Drummond, C. J.; Peat, T. S.; Newman, J., Evaluating Protic Ionic
Liquids as Protein Crystallization Additives. Crystal Growth & Design 2011, 11 (5), 1777-
1785.
228. Anbarasan, S.; Wahlström, R.; Hummel, M.; Ojamo, H.; Sixta, H.; Turunen, O.,
High stability and low competitive inhibition of thermophilic Thermopolyspora flexuosa
GH10 xylanase in biomass-dissolving ionic liquids. Applied microbiology and
biotechnology 2017, 101 (4), 1487-1498.
229. Arora, D. S.; Sharma, R. K., Ligninolytic fungal laccases and their biotechnological
applications. Applied biochemistry and biotechnology 2010, 160 (6), 1760-1788.
230. d'Acunzo, F.; Galli, C.; Gentili, P.; Sergi, F., Mechanistic and steric issues in the
oxidation of phenolic and non-phenolic compounds by laccase or laccase-mediator
systems. The case of bifunctional substrates. New Journal of Chemistry 2006, 30 (4), 583-
591.
231. Jensen, K. A.; Houtman, C. J.; Ryan, Z. C.; Hammel, K. E., Pathways for
Extracellular Fenton Chemistry in the Brown Rot Basidiomycete Gloeophyllum trabeum.
Applied and Environmental Microbiology 2001, 67 (6), 2705-2711.
232. Vanden Wymelenberg, A.; Gaskell, J.; Mozuch, M.; Sabat, G.; Ralph, J.; Skyba,
O.; Mansfield, S. D.; Blanchette, R. A.; Martinez, D.; Grigoriev, I.; Kersten, P. J.; Cullen,
D., Comparative Transcriptome and Secretome Analysis of Wood Decay Fungi Postia
placenta and Phanerochaete chrysosporium. Applied and Environmental Microbiology
2010, 76 (11), 3599-3610.
233. McAndrew, R. P.; Sathitsuksanoh, N.; Mbughuni, M. M.; Heins, R. A.; Pereira,
J. H.; George, A.; Sale, K. L.; Fox, B. G.; Simmons, B. A.; Adams, P. D., Structure and
mechanism of NOV1, a resveratrol-cleaving dioxygenase. Proceedings of the National
Academy of Sciences 2016, 113 (50), 14324-14329.
234. Guengerich, F. P.; Yoshimoto, F. K., Formation and Cleavage of C–C Bonds by
Enzymatic Oxidation–Reduction Reactions. Chemical Reviews 2018, 118 (14), 6573-6655.
235. Dutta, T.; Isern, N. G.; Sun, J.; Wang, E.; Hull, S.; Cort, J. R.; Simmons, B. A.;
Singh, S., Survey of Lignin-Structure Changes and Depolymerization during Ionic Liquid
Pretreatment. ACS Sustainable Chemistry & Engineering 2017, 5 (11), 10116-10127.
236. Goldfeder, M.; Egozy, M.; Shuster Ben-Yosef, V.; Adir, N.; Fishman, A., Changes
in tyrosinase specificity by ionic liquids and sodium dodecyl sulfate. Applied Microbiology
and Biotechnology 2013, 97 (5), 1953-1961.
237. Akanbi, T. O.; Barrow, C. J.; Byrne, N., Increased hydrolysis by Thermomyces
lanuginosus lipase for omega-3 fatty acids in the presence of a protic ionic liquid. Catalysis
Science & Technology 2012, 2 (9), 1839-1841.
238. Zeuner, B.; Riisager, A.; Mikkelsen, J. D.; Meyer, A. S., Improvement of trans-
sialylation versus hydrolysis activity of an engineered sialidase from Trypanosoma rangeli

161
by use of co-solvents. Biotechnology Letters 2014, 36 (6), 1315-1320.
239. Toledano, A.; Serrano, L.; Labidi, J., Organosolv lignin depolymerization with
different base catalysts. Journal of Chemical Technology & Biotechnology 2012, 87 (11),
1593-1599.
240. Lahtinen, M.; Kruus, K.; Heinonen, P.; Sipila, J., On the reactions of two fungal
laccases differing in their redox potential with lignin model compounds: products and their
rate of formation. Journal of agricultural and food chemistry 2009, 57 (18), 8357-8365.
241. Toledano, A.; Serrano, L.; Labidi, J., Improving base catalyzed lignin
depolymerization by avoiding lignin repolymerization. Fuel 2014, 116, 617-624.
242. Shuai, L.; Amiri, M. T.; Questell-Santiago, Y. M.; Héroguel, F.; Li, Y.; Kim, H.;
Meilan, R.; Chapple, C.; Ralph, J.; Luterbacher, J. S., Formaldehyde stabilization
facilitates lignin monomer production during biomass depolymerization. Science 2016,
354 (6310), 329-333.
243. Gschwend, F. J.; Malaret, F.; Shinde, S.; Brandt-Talbot, A.; Hallett, J. P., Rapid
pretreatment of Miscanthus using the low-cost ionic liquid triethylammonium hydrogen
sulfate at elevated temperatures. Green Chemistry 2018, 20 (15), 3486-3498.
244. Doherty, W. O. S.; Mousavioun, P.; Fellows, C. M., Value-adding to cellulosic
ethanol: Lignin polymers. Industrial Crops and Products 2011, 33 (2), 259-276.
245. Chatel, G.; Rogers, R. D., Review: Oxidation of Lignin Using Ionic Liquids—An
Innovative Strategy To Produce Renewable Chemicals. ACS Sustainable Chemistry &
Engineering 2014, 2 (3), 322-339.
246. Mamman, A. S.; Lee, J.-M.; Kim, Y.-C.; Hwang, I. T.; Park, N.-J.; Hwang, Y.
K.; Chang, J.-S.; Hwang, J.-S., Furfural: Hemicellulose/xylosederived biochemical.
Biofuels, Bioproducts and Biorefining 2008, 2 (5), 438-454.
247. Sciences, N. A. o.; Engineering, N. A. o.; Council, N. R., Liquid Transportation
Fuels from Coal and Biomass: Technological Status, Costs, and Environmental Impacts.
The National Academies Press: Washington, DC, 2009; p 388.
248. Kleinert, M.; Barth, T., Towards a Lignincellulosic Biorefinery: Direct One-Step
Conversion of Lignin to Hydrogen-Enriched Biofuel. Energy & Fuels 2008, 22 (2), 1371-
1379.
249. Cotoruelo, L. M.; Marqués, M. D.; Leiva, A.; Rodríguez-Mirasol, J.; Cordero, T.,
Adsorption of oxygen-containing aromatics used in petrochemical, pharmaceutical and
food industries by means of lignin based active carbons. Adsorption 2011, 17 (3), 539-550.
250. Penkina, A.; Hakola, M.; Paaver, U.; Vuorinen, S.; Kirsimäe, K.; Kogermann,
K.; Veski, P.; Yliruusi, J.; Repo, T.; Heinämäki, J., Solid-state properties of softwood
lignin and cellulose isolated by a new acid precipitation method. International Journal of
Biological Macromolecules 2012, 51 (5), 939-945.
251. Verdia, P.; Brandt, A.; Hallett, J. P.; Ray, M. J.; Welton, T., Fractionation of
lignocellulosic biomass with the ionic liquid 1-butylimidazolium hydrogen sulfate. Green
Chemistry 2014, 16 (3), 1617-1627.
252. Fort, D. A.; Remsing, R. C.; Swatloski, R. P.; Moyna, P.; Moyna, G.; Rogers, R.
D., Can ionic liquids dissolve wood? Processing and analysis of lignocellulosic materials
with 1-n-butyl-3-methylimidazolium chloride. Green Chemistry 2007, 9 (1), 63-69.
253. Kilpeläinen, I.; Xie, H.; King, A.; Granstrom, M.; Heikkinen, S.; Argyropoulos,
D. S., Dissolution of Wood in Ionic Liquids. Journal of Agricultural and Food Chemistry
2007, 55 (22), 9142-9148.

162
254. Liu, E.; Das, L.; Zhao, B.; Crocker, M.; Shi, J., Impact of Dilute Sulfuric Acid,
Ammonium Hydroxide, and Ionic Liquid Pretreatments on the Fractionation and
Characterization of Engineered Switchgrass. BioEnergy Research 2017.
255. Das, L.; Xu, S.; Shi, J., Catalytic Oxidation and Depolymerization of Lignin in
Aqueous Ionic Liquid. Frontiers in Energy Research 2017, 5 (21).
256. Rico, A.; Rencoret, J.; del Río, J. C.; Martínez, A. T.; Gutiérrez, A., Pretreatment
with laccase and a phenolic mediator degrades lignin and enhances saccharification of
Eucalyptus feedstock. Biotechnology for Biofuels 2014, 7 (1), 6.
257. Du, J.; Shao, Z.; Zhao, H., Engineering microbial factories for synthesis of value-
added products. J Ind Microbiol Biotechnol 2011, 38.
258. Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C.,
Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free
radical biology and medicine 1999, 26 (9-10), 1231-1237.
259. Trott, O.; Olson, A. J., AutoDock Vina: Improving the speed and accuracy of
docking with a new scoring function, efficient optimization, and multithreading. Journal
of Computational Chemistry 2010, 31 (2), 455-461.
260. Das, A.; Rahimi, A.; Ulbrich, A.; Alherech, M.; Motagamwala, A. H.; Bhalla,
A.; da Costa Sousa, L.; Balan, V.; Dumesic, J. A.; Hegg, E. L., Lignin conversion to low-
molecular-weight aromatics via an aerobic oxidation-hydrolysis sequence: comparison of
different lignin sources. ACS Sustainable Chemistry & Engineering 2018, 6 (3), 3367-
3374.
261. Prado, R.; Brandt, A.; Erdocia, X.; Hallet, J.; Welton, T.; Labidi, J., Lignin
oxidation and depolymerisation in ionic liquids. Green Chemistry 2016, 18 (3), 834-841.
262. Guerra, A.; Lucia Lucian, A.; Argyropoulos Dimitris, S., Isolation and
characterization of lignins from Eucalyptus grandis Hill ex Maiden and Eucalyptus
globulus Labill. by enzymatic mild acidolysis (EMAL). In Holzforschung, 2008; Vol. 62,
p 24.
263. Rahimi, A.; Azarpira, A.; Kim, H.; Ralph, J.; Stahl, S. S., Chemoselective Metal-
Free Aerobic Alcohol Oxidation in Lignin. Journal of the American Chemical Society
2013, 135 (17), 6415-6418.
264. Yuan, X.; Singh, S.; Simmons, B. A.; Cheng, G., Biomass Pretreatment Using
Dilute Aqueous Ionic Liquid (IL) Solutions with Dynamically Varying IL Concentration
and Its Impact on IL Recycling. ACS Sustainable Chemistry & Engineering 2017, 5 (5),
4408-4413.
265. Dabirmanesh, B.; Khajeh, K.; Ghazi, F.; Ranjbar, B.; Etezad, S.-M., A semi-
rational approach to obtain an ionic liquid tolerant bacterial laccase through π-type
interactions. International journal of biological macromolecules 2015, 79, 822-829.
266. Shi, J.; Gladden, J. M.; Sathitsuksanoh, N.; Kambam, P.; Sandoval, L.; Mitra,
D.; Zhang, S.; George, A.; Singer, S. W.; Simmons, B. A., One-pot ionic liquid
pretreatment and saccharification of switchgrass. Green Chemistry 2013, 15 (9), 2579-
2589.
267. Xu, F.; Sun, J.; Wehrs, M.; Kim, K. H.; Rau, S. S.; Chan, A. M.; Simmons, B.
A.; Mukhopadhyay, A.; Singh, S., Biocompatible Choline-Based Deep Eutectic Solvents
Enable One-Pot Production of Cellulosic Ethanol. ACS Sustainable Chemistry &
Engineering 2018, 6 (7), 8914-8919.
268. Fernández-Fernández M, M. D., Domínguez A, Sanromán MÁ, Tavares AP,

163
Rodríguez O, Macedo EA., Stability and kinetic behavior of immobilized laccase from
Myceliophthora thermophila in the presence of the ionic liquid 1-ethyl-3-
methylimidazolium ethylsulfate. Biotechnology Progress 2014, 30 (4), 7.
269. Sun, J.; Liu, H.; Yang, W.; Chen, S.; Fu, S., Molecular Mechanisms Underlying
Inhibitory Binding of Alkylimidazolium Ionic Liquids to Laccase. Molecules 2017, 22 (8),
1353.
270. Yang, Z., Hofmeister effects: an explanation for the impact of ionic liquids on
biocatalysis. Journal of biotechnology 2009, 144 (1), 12-22.
271. Constantinescu, D.; Weingärtner, H.; Herrmann, C., Protein denaturation by ionic
liquids and the Hofmeister series: a case study of aqueous solutions of ribonuclease A.
Angewandte Chemie International Edition 2007, 46 (46), 8887-8889.
272. Jia, S.; Cox, B. J.; Guo, X.; Zhang, Z. C.; Ekerdt, J. G., Cleaving the β O 4
Bonds of Lignin Model Compounds in an Acidic Ionic Liquid, 1-H-3-Methylimidazolium
Chloride: An Optional Strategy for the Degradation of Lignin. ChemSusChem 2010, 3 (9),
1078-1084.
273. Rittstieg, K.; Suurnäkki, A.; Suortti, T.; Kruus, K.; Guebitz, G. M.; Buchert, J.,
Polymerization of Guaiacol and a Phenolic β-O-4-Substructure by Trameteshirsuta
Laccase in the Presence of ABTS. Biotechnology Progress 2003, 19 (5), 1505-1509.
274. Das, L.; Kolar, P.; Sharma-Shivappa, R., Heterogeneous catalytic oxidation of
lignin into value-added chemicals. Biofuels 2012, 3 (2), 155-166.
275. Tian, M.; McCormick, R. L.; Ratcliff, M. A.; Luecke, J.; Yanowitz, J.; Glaude,
P.-A.; Cuijpers, M.; Boot, M. D., Performance of lignin derived compounds as octane
boosters. Fuel 2017, 189, 284-292.
276. Yao, S. G.; Mobley, J. K.; Ralph, J.; Crocker, M.; Parkin, S.; Selegue, J. P.;
Meier, M. S., Mechanochemical treatment facilitates two-step oxidative depolymerization
of kraft lignin. ACS Sustainable Chemistry & Engineering 2018, 6 (5), 5990-5998.
277. Crestini, C.; Lange, H.; Sette, M.; Argyropoulos, D. S., On the structure of
softwood kraft lignin. Green Chemistry 2017, 19 (17), 4104-4121.
278. Boerjan, W.; Ralph, J.; Baucher, M., Lignin Biosynthesis. Annual Review of Plant
Biology 2003, 54 (1), 519-546.
279. Laskar, D. D.; Yang, B.; Wang, H.; Lee, J., Pathways for biomass-derived lignin
to hydrocarbon fuels. Biofuels, Bioproducts and Biorefining 2013, 7 (5), 602-626.
280. De Wild, P. J.; Huijgen, W. J. J.; Gosselink, R. J. A., Lignin pyrolysis for profitable
lignocellulosic biorefineries. Biofuels, Bioproducts and Biorefining 2014, 8 (5), 645-657.
281. Varmana, A. M.; Heb, L.; Follenfanta, R.; Wua, W.; Wemmera, S.; Wrobela, S.
A.; Tang, Y. J.; Singh, S., Decoding how a soil bacterium extracts building blocks and
metabolic energy from ligninolysis provides road map for lignin valorization. Proceedings
of the National Academy of Science 2016, 10.
282. Brown, M. E.; Chang, M. C. Y., Exploring bacterial lignin degradation. Current
Opinion in Chemical Biology 2014, 19, 1-7.
283. Bugg, T. D. H.; Ahmad, M.; Hardiman, E. M.; Rahmanpour, R., Pathways for
degradation of lignin in bacteria and fungi. Natural Product Reports 2011, 28 (12), 1883-
1896.
284. Park, S.; Kazlauskas, R. J., Biocatalysis in ionic liquids – advantages beyond green
technology. Current Opinion in Biotechnology 2003, 14 (4), 432-437.
285. Gschwend, F. J. V.; Malaret, F.; Shinde, S.; Brandt-Talbot, A.; Hallett, J. P., Rapid

164
pretreatment of Miscanthus using the low-cost ionic liquid triethylammonium hydrogen
sulfate at elevated temperatures. Green Chemistry 2018, 20 (15), 3486-3498.
286. Brandt, A.; Grasvik, J.; Hallett, J. P.; Welton, T., Deconstruction of lignocellulosic
biomass with ionic liquids. Green Chemistry 2013, 15 (3), 550-583.
287. Bhatia, S. K.; Jagtap, S. S.; Bedekar, A. A.; Bhatia, R. K.; Patel, A. K.; Pant, D.;
Rajesh Banu, J.; Rao, C. V.; Kim, Y.-G.; Yang, Y.-H., Recent developments in pretreatment
technologies on lignocellulosic biomass: Effect of key parameters, technological
improvements, and challenges. Bioresource Technology 2020, 300, 122724.
288. Singh Arora, D.; Kumar Sharma, R., Ligninolytic Fungal Laccases and Their
Biotechnological Applications. Applied Biochemistry and Biotechnology 2010, 160 (6),
1760-1788.
289. d’Acunzo, F.; Galli, C.; Gentili, P.; Sergi, F., Mechanistic and steric issues in the
oxidation of phenolic and non-phenolic compounds by laccase or laccase-mediator
systems. The case of bifunctional substrates. New Journal of Chemistry 2006, 30 (4), 583-
591.
290. Domínguez, A.; Rodríguez, O.; Tavares, A. P. M.; Macedo, E. A.; Asunción
Longo, M.; Ángeles Sanromán, M., Studies of laccase from Trametes versicolor in aqueous
solutions of several methylimidazolium ionic liquids. Bioresource Technology 2011, 102
(16), 7494-7499.
291. Stevens, J. C.; Das, L.; Mobley, J. K.; Asare, S. O.; Lynn, B. C.; Rodgers, D. W.;
Shi, J., Understanding Laccase–Ionic Liquid Interactions toward Biocatalytic Lignin
Conversion in Aqueous Ionic Liquids. ACS Sustainable Chemistry & Engineering 2019, 7
(19), 15928-15938.
292. Erik M. Nordwald, J. L. K., Stabilization of Enzymes in Ionic Liquids Via
Modification of Enzyme Charge. Biotechnology and Bioengineering 2013, 110 (9), 9.
293. Liu, H.; Zhu, L.; Bocola, M.; Chen, N.; Spiess, A. C.; Schwaneberg, U., Directed
laccase evolution for improved ionic liquid resistance. Green Chemistry 2013, 15 (5),
1348-1355.
294. Wallraf, A.-M.; Liu, H.; Zhu, L.; Khalfallah, G.; Simons, C.; Alibiglou, H.;
Davari, M. D.; Schwaneberg, U., A loop engineering strategy improves laccase lcc2
activity in ionic liquid and aqueous solution. Green Chemistry 2018, 20 (12), 2801-2812.
295. Liu, X.; Gillespie, M.; Ozel, A. D.; Dikici, E.; Daunert, S.; Bachas, L. G.,
Electrochemical properties and temperature dependence of a recombinant laccase from
Thermus thermophilus. Analytical and Bioanalytical Chemistry 2011, 399 (1), 361-366.
296. Haifeng Liu, L. Z., Marco Bocola, Nora Chen, Antje C. Spiessb, and Ulrich
Schwaneberg, Directed laccase evolution for improved ionic liquid resistance. Green
Chemistry 2013, 15, 8.
297. Sciences, N. A. o.; Engineering, N. A. o.; Council, N. R., Liquid Transportation
Fuels from Coal and Biomass. 2009.
298. Das, L.; Li, M.; Stevens, J.; Li, W.; Pu, Y.; Ragauskas, A. J.; Shi, J.,
Characterization and Catalytic Transfer Hydrogenolysis of Deep Eutectic Solvent
Extracted Sorghum Lignin to Phenolic Compounds. ACS Sustainable Chemistry &
Engineering 2018, 6 (8), 10408-10420.
299. Li, W.; Zhou, S.; Xue, Y.; Lee, Y.-J.; Smith, R.; Bai, X., Understanding Low-
Pressure Hydropyrolysis of Lignin Using Deuterated Sodium Formate. ACS Sustainable
Chemistry & Engineering 2017, 5 (10), 8939-8950.

165
300. Socha, A. M.; Parthasarathi, R.; Shi, J.; Pattathil, S.; Whyte, D.; Bergeron, M.;
George, A.; Tran, K.; Stavila, V.; Venkatachalam, S.; Hahn, M. G.; Simmons, B. A.;
Singh, S., Efficient biomass pretreatment using ionic liquids derived from lignin and
hemicellulose. Proceedings of the National Academy of Sciences 2014, 111 (35), E3587-
E3595.
301. Jones, S. M.; Solomon, E. I., Electron transfer and reaction mechanism of laccases.
Cellular and Molecular Life Sciences 2015, 72 (5), 869-883.
302. Claus, H., Laccases: structure, reactions, distribution. Micron 2004, 35 (1), 93-96.
303. Weingärtner, H.; Cabrele, C.; Herrmann, C., How ionic liquids can help to stabilize
native proteins. Physical Chemistry Chemical Physics 2012, 14 (2), 415-426.
304. Lele, V. M. a. S. S., Laccase: Properties and Applications. BioResources 2009, 4
(4), 24.
305. Elcock, A. H.; McCammon, J. A., Electrostatic contributions to the stability of
halophilic proteins11Edited by B. Honig. Journal of Molecular Biology 1998, 280 (4), 731-
748.
306. Dym, O.; Mevarech, M.; Sussman, J. L., Structural Features That Stabilize
Halophilic Malate Dehydrogenase from an Archaebacterium. Science 1995, 267 (5202),
1344-1346.
307. Jaeger, V.; Burney, P.; Pfaendtner, J., Comparison of Three Ionic Liquid-Tolerant
Cellulases by Molecular Dynamics. Biophysical Journal 2015, 108 (4), 880-892.
308. Micaêlo, N. M.; Soares, C. M., Protein Structure and Dynamics in Ionic Liquids.
Insights from Molecular Dynamics Simulation Studies. The Journal of Physical Chemistry
B 2008, 112 (9), 2566-2572.
309. Limongelli, V., Ligand binding free energy and kinetics calculation in 2020. WIREs
Computational Molecular Science 2020, 10 (4), e1455.
310. Kuwahara, M.; Glenn, J. K.; Morgan, M. A.; Gold, M. H., Separation and
characterization of two extracellular H2O2-dependent oxidases from ligninolytic cultures
of Phanerochaete chrysosporium. FEBS Lett 1984, 169.
311. Ragauskas, A. J.; Williams, C. K.; Davison, B. H.; Britovsek, G.; Cairney, J.;
Eckert, C. A.; Frederick, W. J.; Hallett, J. P.; Leak, D. J.; Liotta, C. L.; Mielenz, J. R.;
Murphy, R.; Templer, R.; Tschaplinski, T., The path forward for biofuels and biomaterials.
Science 2006, 311.
312. Johnson, D. L.; Thompson, J. L.; Brinkmann, S. M.; Schuller, K. A.; Martin, L.
L., Electrochemical Characterization of Purified Rhus vernicifera Laccase: Voltammetric
Evidence for a Sequential Four-Electron Transfer. Biochemistry 2003, 42 (34), 10229-
10237.
313. Eswar, N.; Webb, B.; Marti-Renom, M. A.; Madhusudhan, M. S.; Eramian, D.;
Shen, M.-y.; Pieper, U.; Sali, A., Comparative Protein Structure Modeling Using Modeller.
Current Protocols in Bioinformatics 2006, 15 (1), 5.6.1-5.6.30.
314. Katoh, K.; Standley, D. M., MAFFT Multiple Sequence Alignment Software
Version 7: Improvements in Performance and Usability. Molecular Biology and Evolution
2013, 30 (4), 772-780.
315. Shen, M.-y.; Sali, A., Statistical potential for assessment and prediction of protein
structures. Protein Science 2006, 15 (11), 2507-2524.
316. Li, P.; Merz, K. M., MCPB.py: A Python Based Metal Center Parameter Builder.
Journal of Chemical Information and Modeling 2016, 56 (4), 599-604.

166
317. Frisch, M. J.; Trucks, G. W.; Schlegel, H. B.; Scuseria, G. E.; Robb, M. A.;
Cheeseman, J. R.; Scalmani, G.; Barone, V.; Petersson, G. A.; Nakatsuji, H.; Li, X.;
Caricato, M.; Marenich, A. V.; Bloino, J.; Janesko, B. G.; Gomperts, R.; Mennucci, B.;
Hratchian, H. P.; Ortiz, J. V.; Izmaylov, A. F.; Sonnenberg, J. L.; Williams; Ding, F.;
Lipparini, F.; Egidi, F.; Goings, J.; Peng, B.; Petrone, A.; Henderson, T.; Ranasinghe,
D.; Zakrzewski, V. G.; Gao, J.; Rega, N.; Zheng, G.; Liang, W.; Hada, M.; Ehara, M.;
Toyota, K.; Fukuda, R.; Hasegawa, J.; Ishida, M.; Nakajima, T.; Honda, Y.; Kitao, O.;
Nakai, H.; Vreven, T.; Throssell, K.; Montgomery Jr., J. A.; Peralta, J. E.; Ogliaro, F.;
Bearpark, M. J.; Heyd, J. J.; Brothers, E. N.; Kudin, K. N.; Staroverov, V. N.; Keith, T.
A.; Kobayashi, R.; Normand, J.; Raghavachari, K.; Rendell, A. P.; Burant, J. C.; Iyengar,
S. S.; Tomasi, J.; Cossi, M.; Millam, J. M.; Klene, M.; Adamo, C.; Cammi, R.;
Ochterski, J. W.; Martin, R. L.; Morokuma, K.; Farkas, O.; Foresman, J. B.; Fox, D. J.
Gaussian 16 Rev. C.01, Wallingford, CT, 2016.
318. Serrano-Posada, H.; Centeno-Leija, S.; Rojas-Trejo, S. P.; Rodríguez-Almazán,
C.; Stojanoff, V.; Rudiño-Piñera, E., X-ray-induced catalytic active-site reduction of a
multicopper oxidase: structural insights into the proton-relay mechanism and O2-reduction
states. Acta Crystallogr D Biol Crystallogr 2015, 71 (Pt 12), 2396-2411.

167
VITA
Joseph Stevens
EDUCATION
Ph.D., Biosystems and Agricultural Engineering, University of Kentucky
Expected August 2020
B.S., Biological Systems Engineering, Virginia Tech
Graduated May 2016
PROFESSIONAL EXPERIENCE
STEM Chateaubriand Fellow, Toulouse Biotechnology Institute
January to October 2019
OTC Graduate Fellow, UK Office of Technology Commercialization
January to December 2018
Graduate Research Assistant, UK Biosystems and Agricultural Engineering
May 2016 to Present
Undergraduate Researcher, Virginia Tech Biomolecular Engineering Lab
January 2015 to May 2016
Undergraduate Researcher, Virginia Tech Bioenergy and Biotechnology Laboratory
June to December 2014
RESEARCH SUMMARY
Journal Articles
1. Stevens J, Rodgers D, Dumon C, Shi J. “Characterization and Enzyme Engineering of
a Hyperthermophilic Laccase toward Improving Its Activity in Ionic Liquid” Front
Energy Res. 2020
2. Das L, Li W, Dodge L, Stevens J, Williams D, Hu H, Li C, Ray A, Shi J. “Comparative
Evaluation of Industrial Hemp Cultivars: Agronomical Practices, Feedstock
Characterization and Potential for Biofuels and Bioproducts” ACS Sus Chem. 2020
3. Stevens J, Das L, Mobley J, Asare S, Lynn B, Rodgers D, Shi J. “Towards biocatalytic
lignin valorization in aqueous ionic liquids: inhibition mechanisms and product
profiling” ACS Sus Chem. 2019
4. Stevens J, Shi J. “Biocatalysis in Ionic Liquids for Lignin Valorization: Opportunities
and Recent Developments” Biotech Adv. 2019
5. Muley P, Mobley J, Tong X, Novak B, Stevens J, Moldovan D, Shi J, and Boldor D.
“Rapid microwave-assisted biomass delignification and lignin depolymerization in
deep eutectic solvents” En Conv Manag. 2019

168
6. Das L, Li M, Stevens J, Li W, Pu Y, Ragauskas A, Shi J. “Characterization and
catalytic transfer hydrogenolysis of deep eutectic solvent extracted sorghum lignin to
phenolic compounds” ACS Sus Chem. 2018
7. Sheng J, Stevens J, Feng X. “Pathway compartmentalization in peroxisome of
Saccharomyces cerevisiae to produce versatile medium chain fatty alcohols” Sci Rep.
2016
8. Chen Y, Sheng J, Jiang T, Stevens J, Feng X, Wei N. “Transcriptional profiling reveals
molecular basis and novel genetic targets for improved resistance to multiple
fermentation inhibitors in Saccharomyces cerevisiae” Biotechnol Biofuels. 2016
Conference Presentations and Posters
1. Stevens, J., Hunter, J., Dumon, C., Rodgers, D., and Shi, J. (2019, November).
“Improving resistance of hyperthermophilic laccase to ionic liquids for in situ lignin
valorization” Paper presented at the 2019 AIChE Annual Meeting, Orlando, FL.
2. Vin-Nnajiofor, M., Shi, J., and Debolt, S. (2019, November). “Evaluating endocarp
biomass as potential feedstocks for biorefinery” Paper presented for author at the 2019
AIChE Annual Meeting, Orlando, FL.
3. Stevens, J., Hunter, J., Esque, J., André, I., Dumon, C., Rodgers, D., and Shi, J. (2019,
October). “A mixed technique approach to better understand interactions between
laccases and ionic liquids” Poster presented at the 3rd Annual Commonwealth
Computational Summit, Lexington, KY.
4. Stevens, J., Hunter, J., Dumon, C., Rodgers, D., and Shi, J. (2019, July). “Towards
biocatalytic lignin valorization: Interactions between laccase and aqueous ionic
liquids” Paper presented at the 15th International Conference on Renewable Resources
and Biorefineries, Toulouse, France.
5. Li, W., Cheng, Y-T., and Shi, J. (2019, July). “Lignin derived carbon-silicon
nanocomposite materials for energy storage applications” Paper presented for author at
the 15th International Conference on Renewable Resources and Biorefineries,
Toulouse, France.
6. Kalinoski, R., Stevens, J., Dodge, L., Das, L., Muley, P., Boldor, D., Nokes, S., and
Shi, J. (2018, October). “Exploring Antimicrobial Properties of Lignin Derived
Compounds and Materials” Paper presented at the AIChE Annual Meeting, Pittsburgh,
PA.
7. Stevens, J., Mobley, J., Das, L., Rodgers, D., and Shi, J. (2018, October). “Towards
Biocatalytic Lignin Valorization: Exploring Laccase-Ionic Liquid Interactions” Paper
presented at the AIChE Annual Meeting, Pittsburgh, PA.
8. Stevens, J., Mobley, J., Das, L., Rodgers, D., and Shi, J. (2018, July). “Towards
Biocatalytic Lignin Valorization: Exploring Laccase-Ionic Liquid Interactions” Poster
presented at the Biocatalysis Gordon Research Conference, Biddeford, ME.
9. Stevens, J., Mobley, J., Das, L., Rodgers, D., and Shi, J. (2018, July). “Towards
Biocatalytic Lignin Valorization: Exploring Laccase-Ionic Liquid Interactions” Poster
presented at the Biocatalysis Gordon Research Seminar, Biddeford, ME.
10. Stevens, J., Mobley, J., Das, L., Rodgers, D., and Shi, J. (2018, March). “Lignin
valorization via biocatalysis in aqueous ionic liquids” Poster presented at the 2018
EPSCoR Interjurisdictional Conference, Baton Rouge, LA.

169
11. Stevens, J., Das, L., and Shi, J. (2017, December). “Directing product formation from
lignin decomposition via biocatalysis in aqueous ionic liquids” Poster presented at the
7th Annual Sustainability Forum, Lexington, KY.
12. Stevens, J., Das, L., and Shi, J. (2017, November). “Exploring Biocatalysis of Laccase
in Ionic Liquids for Selective Decomposition of Lignin” Poster presented at the 25th
NSF EPSCoR National Conference, Missoula, MT.
13. Stevens, J., Liu, E., Das, L., and Shi, J. (2017, August). “Exploring Biocatalysis of
Laccase in Ionic Liquids for Selective Decomposition of Lignin” Poster presented at
the 2017 EPSCoR Interjurisdictional Conference, Lexington, KY.
14. Stevens, J. and Shi, J. (2017, June). “Exploring Biocatalysis of Laccase in Ionic
Liquids for Selective Decomposition of Lignin” Paper presented at the 2017 KY NSF
EPSCoR PechaKucha Challenge, Lexington, KY.
15. Stevens, J., Kalinoski, R., Das, L., and Shi, J. (2017, May). “Exploring Biocatalysis of
Laccase in Ionic Liquids for Selective Decomposition of Lignin” Paper presented at
the 2017 Renewable Energy & Energy Efficiency Workshop, Louisville, KY.
16. Stevens, J., Kalinoski, R., Das, L., and Shi, J. (2017, May). “Exploring Biocatalysis of
Laccase in Ionic Liquids for Selective Decomposition of Lignin” Poster presented at
the 2017 Renewable Energy & Energy Efficiency Workshop, Louisville, KY.
17. Stevens, J., Kalinoski, R., Das, L., and Shi, J. (2017, May). “Exploring Biocatalysis of
Laccase in Ionic Liquids for Selective Decomposition of Lignin” Poster presented at
the 2017 Symposium on Biotechnology for Fuels and Chemicals, San Francisco, CA.
18. Stevens, J., Sheng, J., and Feng, X. (2016, July). “Metabolic Engineering of
Saccharomyces cerevisiae to produce long chain and medium chain fatty alcohols”
Paper presented at the 2016 International ASABE Meeting, Orlando, FL.
PROFESSIONAL SERVICE
• UK BAE Student Recruitment and Outreach Committee, 2019 - 2020
• UK BAE External Review Committee, 2018
AWARDS AND HONORS
• University of Kentucky
o 1st Place, 3rd Annual Commonwealth Computational Summit Poster Competition,
2019
o UK BAE Outstanding Ph.D. Student, 2019
o Chateaubriand STEM Fellowship, Embassy of France in the US, 2018-2019
o OTC Graduate Fellowship, Office of Technology Commercialization, 2018
o 1st Place, KY NSF EPSCoR PechaKucha Challenge, 2017
o 1st Place, BAE Graduate Student Poster Competition, 2017
• Virginia Tech
o Kentuckiana Alumni Chapter Scholarship, Kentuckiana Alumni Chapter, 2012-
2013
o Distinguished University Scholar, Virginia Tech, 2012-2013
o J. Lawrence and Lucille G. Calhoun Endowment, Biosystems Engineering, 2013-
2016
o Peter H. Fitzgerald Endowment, Biosystems Engineering, 2014-2015