characterization and solubilization of gonadotropin ... · the journal of biological chemistry vol....
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 249, No. 7, Issue of April 10, pp. 2195-2205, 1974
Printed in U.S.A.
Characterization and Solubilization of Gonadotropin
Receptor of Bovine Corpus Luteum*
(Received for publication, July 6, 1973)
FRANCE HAOUR AND BRIJ B. SAXENA~
From the Cornell University Medical College, New York, Xew York 10021
SUMMARY
Plasma membranes isolated from bovine corpora lutea showed specific binding with 1251-human chorionic gonado- tropin and 1251-human luteinizing hormone; unlabeled hor- mones competitively inhibited the binding. The binding of the 1251-human chorionic gonadotropin to the receptor was a saturable phenomenon and the half-saturation was attained at a concentration of 4 X lo-r0 M. Maximum hormone receptor binding was obtained within 15 min at pH 7.2 and 37”. The rate constants of association and dissociation of human chorionic gonadotropin to the receptor determined at 37” were 2.8 X lo6 M-' s-r and 2.1 X lop5 s-r, respectively. The dissociation constant calculated from these rates was 8.0 X lo-r0 M. At 4’ the rate of association was extremely slow and the hormone receptor complex was stable up to 48 hours. The dissociation constants obtained from equilib- rium data were calculated to be 1.5 X lo-r0 M for human chorionic gonadotropin and the number of binding sites were approximately 6.3 x lo+ per mg of tissue. The binding of 1251-human chorionic gonadotropin to the plasma membranes was optimum at pH 7.2 and was reduced at acidic and basic pH. The binding was also inhibited at high concentrations of CaC12, Mg&, and NaCI, and in the presence of guanidine HCl and urea. The inhibition of binding at pH 5 and 9 and up to 1 M concentration of various ions was reversible. Treatment of plasma membrane with phospholipase C, neuraminidase, glucosidase, trypsin, and cy-chymotrypsin did not decrease the binding of 1251-human chorionic gonado- tropin. Pepsin and phospholipase A inhibited the binding of lZ51-human chorionic gonadotropin to plasma membranes. The plasma membranes were solubilized in detergents. The proteins reprecipitated from the solution by ethanol- ammonium acetate retained the ability to bind 1251-human chorionic gonadotropin. Solubilization of the plasma mem- branes in 6 M guanidine HCI resulted in the separation of protein and lipid components. Fractionation of the protein component was achieved by gel filtration on a column of Sepharose 4B in 6 M guanidine HCl and ion exchange chro- matography on DEAE-cellulose in 3 M urea. The receptor
* This work was supported by Contract NICHD72 2763 and Grants CA-13908 and HI)-06543-from the National Institutes of Health and bv Grant M72-189 from The Pooulation Council. The Rockefeller University, New York, New York.
1 Career Scientist Awardee, Contract I-621, Health Research Council of the City of New York.
activity of the purified fractions could be reconstituted by emulsification with the lipid fraction, indicating the require- ment of lipids to retain the native conformation of the re- ceptor. The molecular weight of the active material esti- mated from Sepharose 4B column in 6 M guanidine HCI and polyacrylamide gel electrophoresis in sodium dodecyl sulfate was between 30,000 to 70,000. This partially purified pro- tein may represent the regulatory subunit of the gonado- tropin receptor.
The presence of specific receptors for gonadotropins (FSH,’ LH, and HCG) in the ovaries (l-12), testes (13-14), and in adrenocortical carcinoma (15) has been demonstrated. The in- teraction of the gonadotropins to the receptors in the plasma membranes of the intact ovary has been shown to activate aden- ylate cyclase and steroidogencsis (16-18). There is, however, recent evidence that ovarian steroidogenesis could occur without the activation of adenylate cyclase and that the hormone may enter the cell to stimulate intracellular kinases involved in gonadal steroidogenesis (4, 19, 20). It is not clear whether the receptor and the adenylate cyclase are the components of the same or different molecules. There is evidence for the presence of multiple hormone receptor sites in the plasma membranes of the fat cells which compete for the same adenylate cyclase (21) and that the hormonal binding to the recept,or and the activation of adenylate cyclase can occur independently (22-24). It is likely that hormonal action may involve more than one protein molecule. Isolation and characterization of the gonadotropin receptor is therefore of fundamental importance to elucidate the mechanism of action of gonadotropic hormones at the molecular level.
Solubilization of the plasma membrane proteins of the target tissue to isolate the receptors for norepinephrine (22), acetyl- choline (25), insulin (26-27), and HCG (28) has been achieved by the use of various ionic and nonionic detergents. In this paper, we describe the characterization and solubilization of re- ceptors for gonadotropins from the plasma membranes of bovine corpora lutea.
1 The abbreviations used are: FSH, follicle-stimulating hor- mone; LH, luteinizing hormone; HCG, human chorionic gonado- tropin; SIX, sodium dodecyl sulfate.
2195
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

2196
MATERIALS AND MIETHOIIS Binding of Labeled Hormones to Plasma Membrane Receptor- Plasma membrane fractions equivalent to 200 rg of proteid were
Preparation of Plasma Membranes-Bovine ovaries of earlv incubated at 37” for 45 min with lz51-HCG or lz51-LH (1 to 1.5 x
pregnancy (first”trimester; fetus of crown to rump length up th 1OVo M; specific activity, 30 to 35 &i per fig) or with [&]FSH (1.5 22 cm) were obtained fresh from the slaughter house in chilled X 10m8 M; specific activity, 0.4 &i per pg) in 300 ~1 of 10 mM Tris- containers. The corpora lutea were dissected and either HCl buffer, pH 7.2, containing O.lyc bovine serum albumin. Sim- processed immediately or stored in liquid nitrogen until used. ilar incubations were performed with an excess of unlabeled LH In a typical case, 20 g of the tissue were homogenized by 15 to 20 and HCG (1 X 1OV M) as well as with unlabeled FSH (1 X 10-b strokes in a glass-Teflon homogenizer in 200 ml of 10 mM Tris-HCl M). The receptor-bound hormone was separated by filtration buffer, pH 7.8, containing 1 mM dithiothreitol and 1 rnM MgC12. through an oxoid membrane of pore size 0.45 pm (Amersham- The plasma membranes were isolated according to the procedure Searle). The hormone-receptor complex retained on the filter described in 1)iagram 1 using the Beckman preparative ultra- was washed with 2 to 3 ml of 1% bovine serum albumin in 0.9% centrifuge (model L265B; swinging bucket rotor, type SW 25-l). NaCl and 1 ml of plasma. Usually less than 1% of the total radio- Fractions obtained from the continuous sucrose density gradient activity was adsorbed on the filter. The filters were transferred were examined for purity by electron microscope and for specific into glass vials and the radioactivity was counted in a Packard binding with *251-labeled HCG as described below. As shown Auto Gamma counter with an efficiencv of 51% for 1251. Reagent in Fig. 1, the fractions containing plasma membranes and showing blank as well as controls were performed, with the homogen&$ the maximum binding with lz51-HCG were pooled and stored in of rat rectus muscle, with each experiment. The specific binding liquid nitrogen in suitable aliquots. Aliquots of ovarian ho- was defined as the difference between the total binding and the mogenate and plasma membrane fraction were solubilized in 1 M binding in the presence of the excess unlabeled hormones (40). NabH containing 0.1% SDS. The protein content was deter- In the case if [3H]FSH, the filters were solubilized in thk ilass mined by the method of Lowry et al. (29) using bovine serum vials by the addition of 1 ~1 of TS-1 (Research Product Inter- albumin as the standard. national Corp.) and counted for radioactivitv in Packard scintilla-
Electron Microscopy-An aliquot of the plasma membrane fraction was fixed for 24 hours in G.25% alutaraldehvde in 0.067 M - cacodylate buffer, pH 7.3. The samples were washed for 5 min in chilled 0.25 M cacodylate or phosphate buffer containing 17; 0~01, pH 7.3, for 2 hours. Subsequently, all samples were de- hydrated by passing through a graded series of alcohol and em- bedded in either Upon or Araldite. Thin sections (0.06 to 0.09 pm) were cut and stained in a 4% aqueous uranyl acetate solution and photographed by a Phillips EM-300 electron microscope.
tained from Dr. It. E. Canfield, Coliege of Physicians and Sur- geons, Columbia University, New York), containing 12,000 i.u. per mg (31) or human LH containing approximately 5,292 i.u. of 2nd IRP human menopausal gonadotropin per mg (32) were
Preparation of Radioactive HCG, LH, and FSH-Radioisotopic labeling with lzsI was performed essentially by the procedure of Hunter and Greenwood (30). Twenty micrograms of HCG (ob-
tion counter with an efficiency of 4070 (Packard Instrument Co.). Molecular weights of 32,000, 28,000, and 40,000 were used in the calculation of molar uptake of FSH, LH, and HCG, respectively.
Characterization of Plasma-Membrane Receptor-To establish optimal conditions, effect of various pH values, temperatures, and ions (sodium, calcium, and magnesium) or hormone-receptor binding was examined (Figs. 2, 3, and 9).
estimate the number of receptor sites. The specificity of the hormone-receptor binding was established by competition anal- ysis with increasing quantities of the unlabeled hormones (Fig.
7).
Binding Constants of Hormone-Receptor Interaction (Table Z)- Association constant (K,), dissociation constant (Kd), and K of hormone-receptor binding were determined assuming hormone- receptor binding as an over-all second order reaction. Scatchard analvsis (41) of data obtained from saturation curves was used to
dissolved in 20 ~1 of lO.mM Tris-HCl buffer, pH 7.2, containing 0.9%, NaCl in the reaction vial. To the vial. 50 ul of 0.5 M Dhos- phate buffer (pH 7.4), 1.5 mCi of lz61 (Cambridgk Nuclear, cam- bridge, Mass.), and 70 pg of chloramine-T in 20 ~1 of 0.05 M phos- phate buffer, pH 7.4, were added and mixed. After 10 s the reac- tion was stopped by the addition of 24Opg of sodium metabisulfite in 100 ~1 of phosphate buffer. Two hundred milligrams of iobeads (Hycel Reagents, Houston, Texas) were added to the crude reac- tion mixture to adsorb unreacted lzsI (33). The crude reaction mixture was purified by gel filtration through a column of
Per cent damage and specific activity were determined as de-
Seohadex G-100 (1 X 30 cm) eauilibrated in and eluted with 0.9%
scribed earlier (34).
NLCl containing‘ l@i bovink s&urn albumin. The purity of tie labeled hormone was tested by chromatoelectrophoresis (34).
LH, HCG, and LH-8 were also labeled with lz51 by lactoperoxidase (35) prepared from milk (RZ = 0.78; Sigma
Enzymatic Treatment of Plasma Membranes (Table ZZ)-Phos- pholipase C (Clostridium welchii) and phospholipase A (bee venom) were obtained from Sigma. St. Louis. MO. Plasma mem- branes were incubated with ph&pgolipase A and C in 10 mM Tris-. HCl buffer, with neuraminidase (Worthington Biochemical Corp., Freehold, N.J.), glucosidase (Almond Emulsin, National Bio- chemicals Corp., Cleveland, Ohio), pepsin, trypsin, and or-chymo- trypsin (Mann Research Lab., Orangeburg, N.Y.) in 0.1 M sodium acetate buffer containing 1 mM MgC12, 1 mM MnC12, and 1 mM CaClz for 1 hour at 37”. In another experiment, 1 mg of plasma membrane protein was also incubated with 0.1 to 1Omg
with excess of 10 mM Tris-HCl buffer, pH 7.2, containing 1% bo-
of phospholipase C under conditions described above (Fig. 10). After incubation, the plasma membranes were washed three times
vine serum albumin. Soybean trypsin inhibitor (Sigma, St. Louis, MO.) in lo-fold excess was added to stop the action of
Chemical Cb., St. Louis, MO. i. The specific activity of thela- beled HCG, LH, and LH-8 were 20 to 35 pCi per pg as determined by trichloroacetic acid precipitation. Five microliters of the crude reaction mixture were diluted to 5 ml with 0.05 M phosphate buffer. DH 7.5. containing O.l%, bovine serum albumin. To a 200~pl’afiquot df this solution w&e added 400 ~1 of trichloroacetic acid to a final concentration of 10% trichloroacetic acid. The precipitated proteins were recovered by centrifugation. The radioactivity with the trichloroacetic acid-precipitable material was considered as protein bound and was used in the calculation of the specific activity of the labeled hormone. The biological activity of the labeled LH and HCG was 3,881 i.u. (95yo confidence limits; 2,317 to 4,684 i.u.) and 8,923 i.u. (95yc confidence limits;
proteolytic enzymes. The hormonal binding OF the 200 fig of protein equivalent of treated membrane was tested as described earlier.
Solubilization of Plasma Membranes-Solubilization of the plasma membranes was attempted at 37 and 4” in various ionic detergents like sodium deoxycholate (Schwarz-Mann) and SDS (Pierce Chemicals), as well as non-ionic detergents like Brij 35 (Mann Research Laboratories), Triton Xl00 (Rohm & Haas), Lubrol WX (General Biochemicals), Igepal 630 (General Aniline and Film Corp.) and Tween 80 (Schwarz-Mann). The solubilized membranes were centrifuged at 100,000 X g in a Beckman ultra- centrifuge L265B of rotor twe 50 for 1 hour and at 300.000 X o in rotorType SW 50 for 2 ho&. The absence of nonsedi&ent&le
5,826 to 12,250 i.u.), respectively, as determined by ovarian ascor- material was considered as the evidence for the solubilization of bit acid depletion test (36). Human pituitary FSH containing most of the plasma membranes. The solubilized membranes 5,677 i.u. of 2nd IRP human menopausal gonadotropin (37) was were also not retained during filtration on the oxoid filter of pore also labeled with “H ([aH]KBH, 6 Ci per mmole, Amersham-Searle) size 0.45 pm. The solubilized receptor was precipitated at 4” by according to Vaitukaitis et al. (38). The biological activity of making the solution 10% in ammonium acetate (w/v) and 30yo the [3H]FSH was 3,258 i.u. (95oj, confidence limits; 2,029 to 3,858 in ethanol (v/v). The precipitate was washed three times with i.u.) as determined by Steelman-Pohley bioassay (39). The 10 mM Tris-HCl, pH 7.2. The hormone-binding ability of the specific activity of the [3H]FSH was determined by trichloroacetic precipitate was determined in the same manner as for the intact acid precipitation as described for LH and HCG. plasma membranes. The assay of the receptor protein in insolu-
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

2197
ble form was also applicable to various fractions obtained follow- ing the fractionation of the receptor proteins in the presence of various types and concentrations of the solubilizing agents. The plasma membrane proteins were also solubilized in various con- centrations of guanidine HCl and urea as shown in Fig. 14. From the data presented in Fig. 12 it was obvious that the plasma mem- branes were soluble in higher concentrations of guanidine HCl and urea than of Triton X300, Lubrol WX, sodiumdeoxycholate, and SDS. However. to avoid the formation of micelles (42) and consequent difficulties in removing the detergents, the plasma membranes were solubilized in 6 M guanidine HCI for further purification on a column of Sepharose 4B.
Purification of Plasma Membrane Proteins on Sepharose 4B- Prior to use, guanidine HCl (Heico, Inc., Delaware Water Gap, Pa.) was purified by the addition of 1 g of charcoal (Norit A, Eastman Kodak) to 100 ml of a 6 M guanidine HCl solution. The charcoal was removed by filtration. A column of Sepharose 4B (2.5 X 100 cm) (Pharmacia, Piscataway, N.J.) was equilibrated at 4’ in the 6 M guanidine HCI, pH 5.5. Proteins of known mo- lecular weights, for example bovine serum albumin (mol wt 69,000), egg%valbumin (moi wt 43,000), lysozyme (mol wt 14,300), and insulin (mol wt 6.000). were passed through the column. The elution volume (V,) of the known proteins w& determined in re- lation to the elution of the dextran blue as the reference. Seventy- five milligrams of the plasma membrane protein were solubilized in 1.5 ml of G M guanidine HCl solution. The solution was cen- trifuged at 100,000 X g (Beckman model L265B; rotor type 50) for 1 hour. The solubilized material was separated into a clear por- tion at the bottom and a viscous lipid layer at the top of the tube. The clear solution was aspirated-and concentrated to 5 ml by ultrafiltration (PM-lo. Amicon Corn.. Lexington. Mass.). The concentrated material &as applied to-the column of Sepharose 4B. The column was eluted with 6 M guanidine HCl and 2.ml fractions were collected per 15 min. The eluate was divided into six frac- tions as shown in Fig. 15. Each of these fractions was dialyzed to eliminate the guanidine HCl to recover the receptor protein as a precipitate. These precipitates were tested for hormonal binding. The protein fractions were also emulsified (Bronson sonicator W 185; three pulses of 20 s at 50 watts) with lipid frac- tions isolated during the solubilization of the plasma membranes with euanidine HCl as well as lipids obtained in the top layer of the sAcrose density gradient used-for the purification of the plasma membrane (protein to lipid ratio, 5:1, v/v; 1 ml of fraction con- tained 400 to 500 pg of protein). Phospholipids, such as phospha- tidylethanolamine, phosphatidylcholine, and phosphatidylserine, in concentrations of 1 to 10 mg per ml, also were emulsified with the nrotein Fraction III from the column of Sepharose 4B. The protkin-lipid complexes were examined for binding with lz51-HCG.
Ion Exchange Chromatography on DEAE-cellulose-Fraction III from the Sepharose 4B column was equilibrated and concentrated bv ultrafiltration using PM-10 membrane (Amicon) in 10 rnrvs T”ris-HCl buffer, pH 7.2; containing 3 M urea, 1 mM MgC12, and 1 mM CaCI. and was annlied to a column of DEAE-cellulose (2 X 30 cm) (Cellex-D, Olid meq per g, Bio-Iiad Laboratories, Calif.) equilibrated in the same buffer. The column was eluted by a combination of a stepwise and gradient elution with Tris-HCl buffer containing various molarities of NaCl. The elution pro- file is presented in Fig. 17. Fractions were pooled and an aliquot of each fraction was dialyzed to recover the protein in the insolu- ble form. The insoluble proteins from various fractions were emulsified with the lipid fraction and examined for binding with 12SI-labeled HCG to locate the fraction containing receptor pro- tein.
Isoelectric Focusing-Fractions 72 to 83 from DEAE-cellulose which showed hormonal binding ability (Fig. 17) were pooled and further fractionated by isoelectric focusing in continuous sucrose gradient containing carrier ampholines in the pH range of 3 to 10 in 3 M urea at 10” (43). At the end of the isoelectric focusing, 2-ml fractions were collected via an automatic UV-Cord (LKB Instruments, Stockholm, Sweden) to record the absorption at 280 nm. Fractions were extensively dialyzed first against water to remove sucrose, urea, and carrier ampholines and then against 10 mM Tris-HCI buffer, pH 7.2, containing 1 mM CaCln and 1 mM MgClz to precipitate the proteins. The precipitated proteins were recovered by centrifugation, washed with Tris-HCl buffer, tested for hormonal binding, and analyzed by SDS gel electropho-
v fraction number
rests.
FIG. 1. Specific binding of various fractions to lZ51-HCG from bovine corpora lutea by continuous sucrose density gradient.
Polyacrylamide Gel Electrophoresis in SDS-Plasma membranes, guanidine HCl-soluble material, and various fractions obtained from Senharose 4B. DEAE-cellulose. and isoelectric focusinp columns-were analyzkd by disc electrophoresis in 6.5%~ polyacryla- mide gels containing 1% SDS according to the method of Fair- banks (44). Protein bands were visualized by staining with Coomassie blue and glycoprotein bands were stained with Schiff’s reagent (44). Molecular weight of various protein components was estimated from the mobility of proteins of known molecular weights, for example, bovine serum albumin (mol wt 69,000), egg ovalbumin (mol wt 43,000), and lysozyme (mol wt 14,300), deter- mined simultaneously with those of the receptor protein fractions.
RESULTS
Isolation and Characterization of Plasma Membranes of Bovine Corpora Lufea-Using the procedure described in I>iagram 1, 1 g
of corpora lutea yielded approximately 7 to 10 mg of plasma
membrane protein. The hormonal binding capacity of the
purified plasma membranes was usually 70 to 80% of the total
binding capacity of the corresponding corpora lutea homogenate,
suggesting a small loss of plasma membranes in other subcellular
fractions. Fractions I6 to 22 showed maximum hormone-bind-
ing activity (Fig. 1) and were used in all subsequent sbudies.
The electron photomicrographs of these fractions showed pre-
dominently plasma membranes. Contamination with mito-
chondria was in the range of 15 to 20%. Microsomal contam-
ination was insignificant.
209 corpora lutea, homogenized
in *ym’ Of buffer tris-f.jC( pH7.8--1OmM
centrTfugation, dithiothreitol --- 1 mM
4809 for 20min Mg& -------___ l,,,M
WI. sucrose -------- 0.25 mM
pellk (PI) supeTnatant fS1 J
4 centrifugation,
centrifugation, 55,ooOg for 2 hr on sucrose density gradient fO.5- 1.5M)
4 plasma membranes
I~IAGRAM 1. Preparation of plasma membrane from bovine corpora lutea.
Top 0.5 M wtr~se
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from
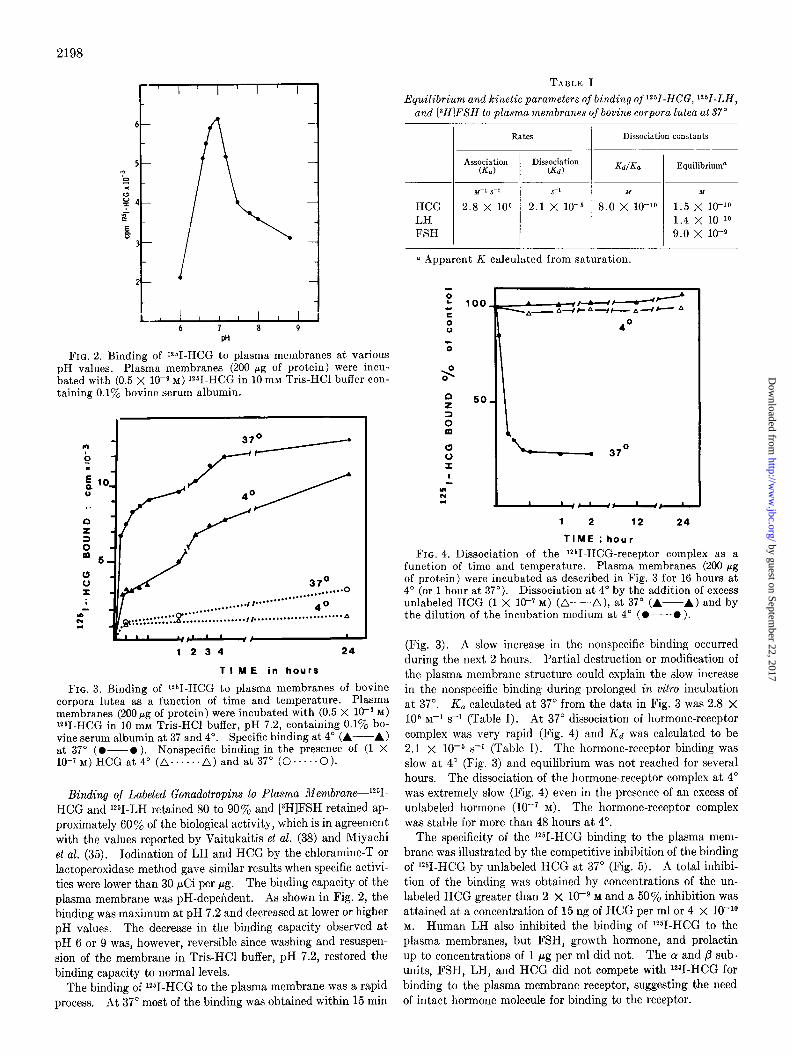
2198
TABLE I
Equilibrium and kinetic parameters of binding of ‘25I-HCG, 12SI-LH, and [SHIFSH to plasma membranes of bovine corpora lutea at 87”
I Lo 1’ 1 ’ 1 ’ 1 I
6 7 8 9 PH
FIG. 2. Binding of i251-HCG to plasma membranes at various pH values. Plasma membranes (200 fig of protein) were incu- bated with (0.5 X 10Wg M) iz51-HCG in 10 mM Tris-HCl buffer con- taining 0.1% bovine serum albumin.
I
n b - ; $ 10, ”
. .
0 2 -
: -
ID Is-
0 - 0 I -
,” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A LI
I I 1 w-r-
1234 24
T I ME in hours
FIG. 3. Binding of i251-HCG to plasma membranes of bovine corpora lutea as a function of time and temperature. Plasma membranes (200 fig of protein) were incubated with (0.5 X 10e9 M) lz51-HCG in 10 mM Tris-HCl buffer, pH 7.2, containing 0.1% bo- vine serum albumin at 37 and 4”. Specific binding at 4” (A--A) at 37” (O-O). Nonspecific binding in the presence of (1 X 10+~)HCGat4” (n......A)andat37’(0*....0).
Binding of Labeled Gonadotropins to Plasma iilembrane--‘2SI- HCG and i*51-LH retained 80 to 90% and [3H]FSH retained ap- proximately 60% of the biological activity, which is in agreement with the values reported by Vaitukaitis et al. (38) and Miyachi et al. (35). Iodination of LH and HCG by the chloramine-T or lactoperoxidase method gave similar results when specific activi- ties were lower than 30 $Zi per pg. The binding capacity of the plasma membrane was pH-dependent. As shown in Fig. 2, the binding was maximum at pH 7.2 and decreased at lower or higher pH values. The decrease in the binding capacity observed at pH 6 or 9 was, however, reversible since washing and resuspen- sion of the membrane in Tris-HCl buffer, pH 7.2, restored the binding capacity to normal levels.
The binding of ‘*51-HCG to the plasma membrane was a rapid process. At 37” most of the binding was obtained within 15 min
Rates Dissociation constants
Association Dissociation VU (Kd) KiIKz Equilibriuma
x-1 s-1 s-1 M M
HCG 2.8 x 106 2.1 x 10-E 8.0 x 10-10 1.5 x 10-10 LH 1.4 x IO-‘0 FSH 9.0 x 10-g
a Apparent K calculated from saturation
II, L 37O
-u-I
1 2 12 24
TIME ; hour
FIG. 4. Dissociation of the iz51-HCG-receptor complex as a function of time and temperature. Plasma membranes (200 Kg of protein) were incubated as described in Fig. 3 for 16 hours at 4” (or 1 hour at 37”). Dissociation at 4” by the addition of excess unlabeled HCG (1 X 1OW M) (A-A), at 37” (A-A) and by the dilution of the incubation medium at 4” (O--O).
(Fig. 3). A slow increase in the nonspecific binding occurred during the next 2 hours. partial destruction or modification of the plasma membrane structure could explain the slow increase in the nonspecific binding during prolonged in vitro incubation at 37”. K, calculated at 37” from the data in Fig. 3 was 2.8 x lo6 M-I s-l (Table I). At 37” dissociation of hormone-receptor complex was very rapid (Fig. 4) and Kd was calculated to be 2.1 x 10m5 s-l (Table I). The hormone-receptor binding was slow at 4” (Fig. 3) and equilibrium was not reached for several hours. The dissociation of the hormone-receptor complex at 4” was extremely slow (Fig. 4) even in the presence of an excess of unlabeled hormone (lo-’ M). The hormone-receptor complex was stable for more than 48 hours at 4”.
The specificity of the iz51-HCG binding to the plasma mem- brane was illustrated by the competitive inhibition of the binding of ‘*51-HCG by unlabeled HCG at 37” (Fig. 5). A total inhibi- tion of the binding was obtained by concentrations of the un- labeled HCG greater than 2 x 10mg M and a 50% inhibition was attained at a concentration of 15 ng of HCG per ml or 4 x lo-i0 M. Human LH also inhibited the binding of lz51-HCG to the plasma membranes, but FSH, growth hormone, and prolactin up to concentrations of 1 pg per ml did not. The Q! and p sub- units, FSH, LH, and HCG did not compete with 1251-HCG for binding to the plasma membrane receptor, suggesting the need of intact hormone molecule for binding to the receptor.
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

2199
-10 10 M
. I
1 I I 5 10 20 40 60 160
HCG : rig/ml
FIG. 5. Competitive inhibition by unlabeled HCG on the bind- ing of 1261-HCG to plasma membranes of bovine corpora lutea. Incubates containing 250 rg of plasma membrane protein were incubated with 1 X lo-10 M lzSI-HCG in the presence of increasing amounts of unlabeled HCG.
Characterization of Binding of lz51-HCG, lz51-LH, and [3H]F~H to Plasma Membranes-The binding of the hormone to the re- ceptor was a saturable phenomenon (Fig. 6). With increasing concentrations of i*5I-HCG, the saturation was attained between 10-g to lop8 M. Scatchard plot of the equilibrium data indicated a Kd of 1.5 x lo-i0 M (Fig. 7 and Table 1). The X intercept of the Scatchard plot yielded an estimate of the number of binding sites to be in the order of 2.5 to 10 x lo-i3 moles per mg of pIasma membrane protein, which are 100.foId greater than the number of sites in the fresh corpora lutea homogenate (2.5 to 10 X lo+ moles per mg of tissue).
Biologically active [3H]FSH bound specifically to plasma mem- brane of cow corpus luteum (Fig. 8) ; however, very high non- specific binding was noticed. The binding capacity of mem- brane derived from the equilibrium data was evaluated to a value of 1 X lo-i3 moles per mg of protein. The value of Kd derived from this data is 9.0 x 10m9 M (Table I).
Effect oj Various Treatments on Plasma il1embranes-Sonica- tion did not destroy the ability of the purified plasma membrane to bind i*51-HCG. The competitive inhibition curve with un- labeled HCG was identical to that of the intact membranes. However, the sonicated plasma membranes showed a greater tendency to aggregate, which may explain a slight decrease in the binding capacity of the preparation.
As shown in Fig. 9, an increase in the binding was observed with concentrations of CaC& up to 10 mM. Such an effect was not seen in the case of MgC12, perhaps due to the fact that mem- branes were prepared in Tris-HCl buffer already containing 10 mM MgClz and the incubation medium also contained 0.2 to 0.5 mM MgC&. An increase in the concentrations of various ions, however, inhibited the binding of ‘*51-HCG to the receptor. In the case of NaCl, a 50% inhibition in the binding was observed at a concentration of 0.35 M. A similar inhibition in the binding of the hormone to sonicated plasma membranes was observed
20 /
Binding
Specific Specific
10 -10
10-J 10 -9
125 I-HCG (M)
FIG. 6. Binding of n51-HCG to bovine corpus luteum mem- branes as a function of the concentration of labeled HCG. Plasma membranes (400 pg of protein) were incubated with increasing amounts of lzSI-HCG in 10 rnM Tris-HCl buffer, pH 7.2, containing 0.14” bovine serum albumin. Total binding, O---O ; nonspecific binding in the presence of 1 X lo+ M unlabeled HCG, O-O; blank representing radioactivity adsorbed on the oxoid filter, A---A; specific binding, 0 . . . . . l
.7 .8 .9 1
HCG BOUND (MxlO 13
)
FIG. 7. Scatchard plot analysis of the data presented in Fig. 6. Molar concentration of HCG bound is given for 400 rg of mem- brane protein.
at NaCl concentrations greater than 1 M. In the case of CaClz and h1gC12, 50% inhibition was observed at a concentration of 0.12 M. It is interesting to note that the inhibition of the binding at salt concentrations up to 1 M was fully reversible after washing and resuspension of the plasma membranes in the incubation buffer. These observations indicate that low concentrations of divalent ions like Mgzf and Ca2+ facilitate the binding.
Phospholipase A suppressed the ability of the intact and soni- cated plasma membranes to bind with iz51-HCG, whereas phos-
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

2200
9 1 Binding
Binding
I 1 I I
10 -9
10 -6
3H-FSH (MI
FIG. 8. Binding of 13H]FSH to plasma membrane of bovine corpora lutea. Plasma membranes (200 pg of protein) were in- cubated with increasing concentrations of [3H]FSH in 300 ~1 of 10 mM Tris-HCl buffer, pH 7.2, containing 0.1% bovine serum albumin at 37” for 45 min. Total binding, O--O; nonspecific binding in the presence of 1 X 1O-5 M FSH, O--O; blank or radioactivity adsorbed on the oxoid filter, A---A.
NaCl u
MgC12 o-o
CaCl* -
1 b.................... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ..A
/’ ” ” ” ” ’ .I .2 .3 .I .5 .6 .7 .a .s 1
Molarity
FIG. 9. Effect of various ionic concentrations on the binding of lz51-HCG to plasma membranes of bovine corpora lutea. Plasma membrane (250 rg of protein) were incubated with in- creasing concentrations of various iolls in 10 mM Tris-HCl buffer, pH 7.2, containing 0.17c bovine serum albumin at 37” for 45 min. NaCl, .---0;MgC12, O-O; CaC12, w-4; blank, A.....A.
pholipase C siguificantly iucreased the binding ability. As shown in Table II and Fig. 10, the binding ability of the sonicated plasma membranes was increased 2.4.fold by treatment with phospholipase C ; however, at higher coneent,rations the binding ability was iuhibited. It is likely that at lower concen- trations phospholipase C deaggregated the plasma membrane to unmask additional receptor sites, whereas at higher conceu trations of phospholipase C, the plasma membraue structure was disrupted. As showu in Table II, hormone-binding ability of
TABLE II
Effect of enzymatic treatmerd 011 plasma membralzes
a-Chymotrypsin., Trypsin. Pepsin. Emulsin Neuraminidase.. Phospholipase A. Phospholipase C Phospholipase C
Specific binding
70 of conirol~
109 (100-117) 95 (866100) 3 (@8)
203 (193-239) 77 (63-90) 10 (512)
112 (100-119) 240 (23(f258)
Incubation
PH
8 8 5 5 5.6 7.2 7.2 7.2
a Average and range of the triplicate determination * Protein. c Sonicat,ed plasma membranes.
E:S
W/Wb
1:60 1:30 1:60 1:3 1:30 1:lO 1:10 1:1oc
0 .I5 .c 2.5 10
PHOSPHOLIPASE C; mg/ml
FIG. 10. Effect of phospholipase C on the binding of lZ61-HCG to plasma membranes. Sonicated plasma membranes (200 /*g of protein) were incubated with increasing amounts of phospholipase C at 37” for 1 hour.
the plasma membranes was stable to neuraminidase; however glucosidase treatment increased the binding of lZ51-HCG. Tryp- siu and a-chymotrypsin did not affect, whereas pepsin com- pletely suppressed, the binding of the hormone to the plasma membranes. It may be suggested that the complex lipid-protein nature of the plasma membrane receptor may be a steric hin- drance for the enzymes to attack specific sites.
Solubilization of Plasma Jlembranes-Ionic detergents (deoxy- cholate, SDS) and nonionic detergents (Tritou X-100, Lubrol WX, Igepal 630, Brij 35, and Tween 80) have been used to solu- bilize plasma membranes. As showu in Fig. 11, the concentra- tion of the detergeut necessary to solubilize the membrane re- ceptor is temperature-dependent. Attempts to solubilize the receptor at room temperature or at 37” adversely affected the hormone-binding ability of the receptor. Solubilization was therefore performed at 4”. At this temperature more than lo- fold detergent was required to totally solubilize 1 mg of purified plasma membrane. Effectiveness of various detergents, as measured by their ability to decrease the activity of labeled plasma membranes retaiued on the osoid filter, was in the fol-
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from
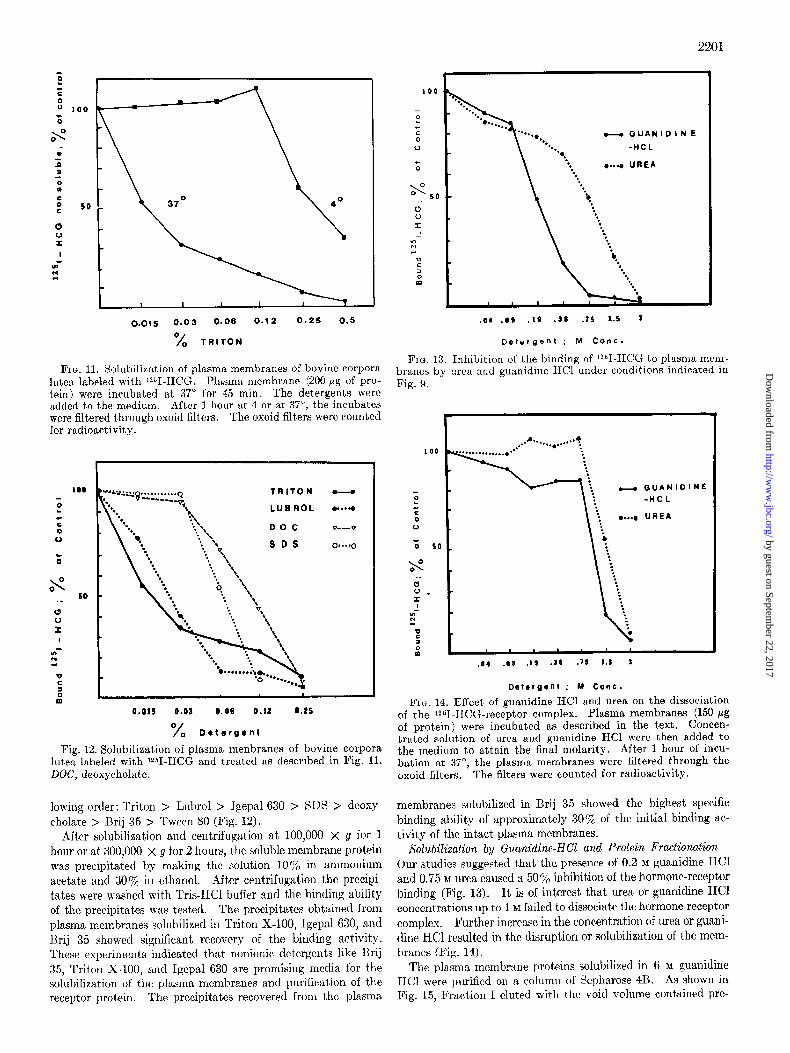
0.015 0.03 0.06 0.12 0.25 0.5 .04 -09 .I9 .38 l ts 1.5 3
0 / 0 TRITON Detergent ; M Cont.
FIG. Il. Solubilization of plasma membranes of bovine corpora lutea labeled with 12gI-HCG. Plasma membrane (200 pg of pro- tein) were incubated at 37” for 45 min. The detergents were added to the medium. After 1 hour at 4 or at 37”, the incubates were filtered through oxoid filters. The oxoid filters were counted for radioactivity.
100
eo L CI
E 0
m’ CII CI
TRITON
LU!ROL
I
r
0.01s 0.03 0.06 0.12 0.25
0 / 0 Detergent
Fig. 12. Solubilization of plasma menbranes of bovine corpora lutea labeled with W-HCG and treated as described in Fig. 11. DOG, deoxycholate.
lowing order: Triton > Lubrol > Igepal 630 > SDS > deoxy- cholate > I3rij 35 > Tween 80 (Fig. 12).
After solubilization and centrifugation at 100,000 x g for 1 hour or at 300,000 x g for 2 hours, the soluble membrane protein was precipitated by making the solution 10% in ammonium acetate and 30y0 in ethanol. After centrifugation the precipi- tates were washed with Tris-HCl buffer and the binding ability of the precipitates was tested. The precipitates obtained from plasma membranes solubilized in Triton X-100, Igepal 630, and I3rij 35 showed significant recovery of the binding activity. These experiments indicated that nonionic detergents like Brij 35, Triton X-100, and Igepal 630 are promising media for the solubilization of the plasma membranes and purification of t,he receptor prot,ein. The precipitates recovered from the plasma
‘a T GUANIDINE ‘0
l * l * -HCL
l g. . . . l .**e UREA . . .
FIG. 13. Inhibition of the binding of 1261-HCG to plasma mem- branes by urea and guanidine HCl under conditions indicated in Fig. 9.
100
GUANIDINE
2 -HC L Y e 0 UREA v
l 04 l 09 .19 .38 .7S 1.5 3
Detergent ; M Cont.
FIG. 14. Effect of guanidine HCl and urea on the dissociation of the ?I-HCG-receptor complex. Plasma membranes (150 pg of protein) were incubated as described in the text. Concen- trated solution of urea and guanidine HCl were then added to the medium to attain the final molarity. After 1 hour of incu- bation at 37”, the plasma membranes were filtered through the oxoid filters. The filters were counted for radioactivity.
membranes solubilized in Brij 35 showed the highest specific binding ability of approximately 300j0 of the initial binding ac- tivity of the intact plasma membranes.
Solubilization by Guanidine-NC1 and Protein Fractionation- Our studies suggested that the, presence of 0.2 M guanidine HC1 and 0.75 M urea caused a 50% inhibition of the hormone-receptor binding (Fig. 13). It is of interest that urea or guanidine HCI concentrations up to 1 M failed to dissociate the hormone-receptor complex. Further increase in the concentration of urea or guani- dine HCl resulted in the disruption or solubilization of the mem- branes (Fig. 14).
The plasma membrane proteins solubilized in 6 M guanidine HCl were purified on a column of Sepharose 4B. As shown in Fig. 15, Fraction I eluted with the void volume contained pre-
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

0 L ii OUr 20 30 40 50 60 70 80 Tube number
FIG. 15. Fractionation of 50 mg of plasma membrane proteins of bovine corpora lutea solubilized in 6 M guanidine HCI on a column of Sepharose 4B (2.5 X 100 cm) equilibrated with ci M
guanidine HCl, pH 5.5, at 4”. BSA, bovine serum albumin.
dominantly high molecular weight material. Fractions II, III, and IV were rich in prot’eins, whereas Fractions I and VI con- t,ained residual lipids and pigments. These fractions were dialyzed against water followed by 10 mM Tris-IX1 buffer, pi-I 7.2, to eliminate the guanidine HCl ard to recover the protein in the precipitated form. These precipitates showed no specific binding ability with 12%HCG. It may be pointed out that Fractions II to V were essentially free of lipids, since most of the lipids were removed during centrifugation following the solubili- zation in 6 M guanidine IX1 and t(he filtration on t’he column of Sepharose 4B. An examination of the binding activity of the protein-lipid complexes revealed that lipids in the top layer of the guanidine IICl-solubilized membrane did not restore the binding ability of t,hese fractions; however, the crude lipids in the top layer of the sucrose density gradient recovered during the initial purification of the plasma membranes (Diagram 1) restored the hormone-binding activity of Fractions II, III, and IV. It is of interest that the lipid fractions did not show any binding ability by themselves. The lack of restoration of the binding activity of t,he Fractions II, III, and IV, with the lipids obtained after the solubilization of the plasma membrane, was due to the presence of guanidine HCl. After remova, of guani- dine IX1 by dialysis alternate with sonication, the lipid fraction was able to restore the binding activity of Fractions II, III, and IV in the same way as the crude lipids.
These observations suggested that solubilization of plasma membranes resulted in (a) the separation of a major protein component and a minor but vital lipid component of the gonado- tropin receptor, (b) the fractionation of the protein component on a column of Sepharose 41~ resulted in t’he purification of the receptor prot,ein, and (c) it was possible to recon&tute the re- ceptor by mising prot.ein and lipid components in vitro. As shown in Fig. 15, the hatched area indicates the fractions which showed specific binding. Fractions I and VI showed no binding. Fractions II, III, and IV showed specific binding, The maxi- mum binding was present in Fraction III.
Att’empts to reconstitute the binding with phosphatidyleth- anolamine, phosphatidylcholine, or phosphatidylserine at con- centrations between 1 and 10 mg per ml have not as yet been successful. Characterization of the lipid fraction should permit
the identification of the essential lipid involved in t,he receptor structure.
Ion exchange chromatography of Fraction III on DEAE-cellu- lose resulted in further purification of the receptor protein. The fraction eluted between 0.04 M and 0,2 M NaCl, as indicated by the hatched area in Fig. 16, showed specific binding with 1251-HCG after reconstitution with the lipids; however, this fraction was not homogeneous when examined by polyacrylamide gel electro- phoresis in SDS. Efforts toward further purification of the re- ceptor protein by isoelectric focusing in 2 M urea resulted in the separation of several protein bands. Due to the precipitation of these bands on the column, a discrete isolation of the protein associated with the receptor has not been achieved. The iso- electric point of the fraction containing receptor activity was ap- proximately 4.5. On the basis of the recovery of the protein, it was estimated that at least lOOO-fold more purification was neces- sary to obtain a homogeneous protein with receptor activit’y.
YoEyacrylamide Gel Rlectrophoresis in SDS-Disc electro- phoretic pattern of intact plasma membrane (Fig. 17) indicated the presence of at least 20 major protein bands, whereas Schiff’s reagent indicated the presence of four glycoproteins in the molec- ular weight range of 100,000, 80,000, 45,000, and 10,000. From the Sepharose column, Fraction I contained highly aggregated material which did not enter the gel. Fractions II, IV, and V showed multiple protein bands in the molecular weight range 10,000 to 100,000. A little receptor activity present in these fractions was due to overlap with Fraction III which contained most of the receptor protein activity. Fraction III showed pro- t,ein bands of molecular weight range between 30,000 and 70,000. This estimate was close to the molecular weight of Fraction III estimated from the Sepharose 413 column.
DISCUSSION
The general charact,erist,ics of the binding of 1251-HCG to the plasma membranes of bovine corpora lutea are comparable lcvith those reported for gonadal tissue. RIost of the specific and high affinity binding was demonstrated by the plasma membranes. The binding phenomenon was pH- and temperature-dependent. and indicated the requirement of divalent ions (Mg2+ or Ca2+) in the incubation medium for optimum binding. The estimates of Kd of the binding of Y-HCG to the plasma membranes calcu- lated from equilibrium data at 37’ and from the ratio of rate constants were 8.0 and 1.5 x lo-lo M as compared to 3 and 1.1 x 10Lg M reported for the binding of bovine LH to plasma membranes of bovine corpus luteum (11) l These values can also be compared with those obtained for the plasma membrane of rat superovulated ovaries (3 X lo-l2 M (4)), for rat testes homogenate (3 X lo-lo M (44)), and for human corpus luteum (8.6 X lo-l1 M (45)). 0 varian slices yielded a lower value of 1.5 x 10mL8 M (8). It seems that the apparent affinity of the hormone binding increases in purified plasma membranes. The lower Kd in crude receptor preparat,ions may partly be due to nonspecific adsorption or enzymat,ic destruction of the hormone during incubation. It is interesting to note that HCG shows similar binding ability for the receptor sites of rat t,estis, super- ovulated rat ovary, or pregnant cow and human corpora lutea, suggesting lack of species specificity of hormone-receptor bind- ing; the possibility of subtle quantitative differences, however, are not ruled out. The number of binding sites per mg of fresh bovine ovarian tissue was estimated to be 5 x IO-l5 M, a value which is close to the number reported for LH in human corpus luteum (1.8 x lo-i4 M (8)) and for HCG in superovulated rat
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

2203
FIG. 16. Disc electrophoresis in 6.6% polyacrylamide gel in lyc SDS. TWCI hundred micrograms of proteins of plasma membranes or fractions from the column of Sepharose 4B (Fig. 15) were analyzed. The gels were stained with Coomassie blue (CB) or with Schiff’s reagent (PAS). On the left are shown the molarities of proteins of known molecular weights (see text) analyzed simul- taneously.
DEAE CELLULOSE IN TRIS-HCL EUFFER.PHT,ImY MgCl2,3M UREA
FIG. 17. Fractionation of plasma membrane protein (Fraction IZZ
column, Fig. 15) from Sepharose 4B on DEAE-cellulose.
0.02L) 0.04 u
I I
NaCI
GRADIENT 0.04 M
0.2 Y
.I
10 20 30 40 so 70 00 00 100 110 **0 120 130 140
TUBE NUMBERS
0.5 m I.0 H
I 1
ovary (1.8 X lo-r4 M (4)). It is interesting to note that the affinity of the gonadotropin receptor for HCG was comparable to the values obtained in the case of insulin-receptor interaction in the fat cells (27-47). The weight of receptor present per mg of membrane protein can be calculated from the number of binding sites (6.3 x lo-l3 moles per mg) and from a postulated molecular weight of the receptor protein of 70,009. Thus, ap- proximately 60 ng of receptor protein are present in 1 mg of plasma membrane proteins. Hence, approximately 20,000-fold purification of the purified plasma membranes was necessary to obtain chemically pure receptor.
At low temperature (4”), the hormone-receptor complex was stable for more than 48 hours, even in the presence of excess of
unlabeled HCG which is in agreement with similar observations obtained with rat ovarian membranes (4), homogenates (48), and intact granulosa cells (49). It is logical to assume that varying amounts of endogenous LH may remain bound to the receptor in plasma membranes prepared from the ovaries at different stages of the reproductive cycle. Further support of this conjecture may be derived from the fact that the number of receptor sites to human LH in human ovary vary at various states of the reproductive (50) and menstrual cycle (9). Chan- ning and Kammerman (49) have recently suggested that the endogenous hormone bound to the ovarian receptor may interfere in the measurement of the binding of r*“I-HCG to the ovarian cells. It may be mentioned that our preliminarv experiments
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

2204
have revealed that the number of binding sites actually increased in the plasma membranes after the dissociation of endogenously bound hormone under suitable conditions of pH, temperature, and the composition of the incubation misture. The dissociated hormone exhibited immunological identity with t,he native hor- mone in t’he radioimmunoassay. The details of these esperi- merits will be published elsewhere. The high variability ob- served in the determination of the number of hormone binding sites in the target organs may also be due to the ratio of high and low affinity receptors (50). One would therefore invoke a con-
cept of apparent and total binding sites in the evaluation of hormone-receptor binding phenomenon.
The results obtained in our studies have indicated that divalent ions, at low concentrations, facilitate the binding of HCG to plasma membranes. However, concentrations great’er t,han 10 mM CaC&, and R4gC12 produced an inhibiting effect similar t,o the inhibit,ion of glucagon binding to fat cells (47) and thyroid- stimulating hormone binding to thyroid plasma membranes (51). These observations are in variance with those obtained for insulin binding to the liver and fat cells (27) and for HCG binding t’o the plasma membranes of superovulated rat ovaries (4). In t’hese studies, an increase in the binding ability of t’he plasma membrane with increasing NaCl concentration up to 2 M was report,ed. It is interesting to note that the suppression of binding caused by concentrations of NaCl, Mg&, and CaC12 up to 1 M was fully reversible, suggesting that at these concentrations, no irreversible conformational changes occurred in t,he plasma membrane struc- ture. The supernatants recovered following t,reatment with higher salt concentration (greater than 1 M) showed a precipitate after the removal of salts by dialysis which contained specific binding with 1251-HCG suggesting that some receptor was solu- bilized and may account for the loss of binding activity at higher salt concentrations.
Similarly, the inhibition of the binding caused by urea or guanidine HCl up to the concentrations of 1 M was fully reversible, indicating no significant destruction of the receptor (Fig. 13). The stability of the hormone-receptor complex was demon- strated when concentrations upto 1 M urea or guanidine HCl failed to dissociate the hormone-receptor complex (Fig. 14). These observations confirm the high stability of the noncovalent linkage between hormone and receptor molecules.
The protein nature of the plasma membrane receptors has been amply demonstrated (47,52). In view of the highly hydrophobic nature of the plasma membrane structure, the results of the proteolytic attack should be interpret,ed with caution. Our results indicate that0 the receptor was stable to the proteolytic enzymes like trypsin and chymotrypsin due to st,eric hindrance for these enzvmes to reach the specific peptide linkages; however, pepsin treatment at pH 5.0 destroyed the receptor activity. Neuraminidase and glucosidases also do not suppress the binding ability, indicating that, if any, a minor carbohydrate moiety might be involved in the receptor structure. An increase in the hormone-binding ability following t,reatment with phospholipase in the case of insulin receptor (47) and HCG receptor (4) has been reported. In the case of glucagon receptor there was a decrease in the binding (52). In the case of intact plasma mem- branes, our studies did not show any significant increase in the binding. A very consistent increase, however, was observed in the case of sonicated membranes. These considerations are important for the explanation of the general physiological role of receptor sites. If the number of receptor sites can be modified under various pathological or physiological (9, 46) condit,ions, one may speculate that either existing receptors are unmasked or
that new receptors are inserted in the membrane structure. Evidence of a change in t,he number of binding sites has been obtained in the case of insulin receptors (46). Our data would support the hypothesis that there are no hidden binding sites in the case of HCG receptor and that the apparent change in the number of sites may be a result of the degree of binding sites occupied by the endogenous hormone,
Tritiated FSI3I was specifically concent,rated by the ovary of immature or pseudo-pregnant rat (38). Our data confirms t,he presence of a specific binding site for FSH in the plasma mem- branes of the corpus luteum of pregnant cow. The Kd of the binding was estimated to be 9.0 x 10mg M and can be compared to the value of 7 x 10Bg M reported in the case of rat testis (14). Since FSH did not compet,e kth lz51-HCG for binding, it may be postulated that the FSH is bound to t,he sites dist’inct from those for HCG.
Solubilization of the plasma membranes at t,emperatures 28” and 37’ did not result in an active form of the receptor. This is in contrast to the insulin receptor. Possibly at higher tempera- t,ure there was a greater proteolytic destruction. Solubiliza,tion of the receptor by guanidine HCl and the recovery of binding activity of the protein moiety after reconstitution with the lipid fraction suggested that the receptor proteins were not, irreversibly denaturated by guanidine HCl and that the reconstitution of the native structure requires lipids. It is of interest t,o note that dissociation of LH into subunit’s by guanidine HCl and their reconst,itution after removal of guanidine HCl has been achieved (53). Denaturation of human serum albumin by the same chaotropic agent has been shown to be a reversible phenomenon (54). Separation of membrane proteins has been successfully achieved in the presence of guanidine HCl. It has also been demonstrated t,hat the phospholipids were able to restore hor- mone stimulat,ing act’ivity of cat myocardial adenyla’te cyclase (22) as we31 a,s glucose &phosphatase activity of the liver micro- somes (56). The data presented here showed that the gonado- tropin receptor could be separated into protein and lipid moieties by solubilization in guanidine HCl and that the receptor could be reconstituted i?z vitro by combining the lipid and protein moieties. The reconstituted receptor eshibit’ed restoration of t’he specific binding with 1251-HCG.
The molecular weight of t,he protein moiet,y of the receptor determined in the presence of 6 M guanidine HCl and SIX was in the range of 30,000 to 70,000. These est.imates are lower than the values obtained in detergents for the intact receptor protein for insulin (300,000) (27) or for acetylcholine (360,000) (25). The cholinergic recept,or has also been reported t’o be constituted of subunits of the molecular weight of 40,000 to 50,000 and a p adrenergic receptor was recent’ly described iI1 two molecular forms of molecular weight,s 40,000 and 160,000 (57). According to the concept of Sutherland et al. (58) a hormone recept’or may involve a regulatory subunit, a catalytic subunit (adenylate cyclase), and, perhaps, intermediary couplers. The partially purified protein moiety of a molecular weight between 30,000 to 70,000 described here may represent t,he regulatory subunit of the gonadotropin receptor involved in the recognition of the hormone. It is also concluded that the lipid moiety is essential for the native conformation of the receptor structure. Further charac- terization of the protein and lipid moieties of the receptor, which appears promising by using detergents and guanidine HCl as solubilization agents, would permit a precise elucidation of the structure-fun&on relationship of the gonadotropin receptor.
Acknowledgments-We are grateful to Drs. R. Canfield and
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

2205
0. P. 13ahl for HCG and their subunits, to Dr. P, Rathnam for human pituitary FSH, LH, and their subunits, to Dr. E. Nunez for electron microscopic studies, to Ms. Nancy Moore for techni- cal assistance, and to Ms. Sharon Wardlaw for excellent help during the course of these studies.
Addendum-Since this manuscript was submitted for publica- tion, Gospodarowicz (59) has described the binding of bovine LH to bovine corpora lutea and determined a Kd of 3 x 10Pg M.
28. DUFAU, M. I,., AND CATT, K. J. (1973) Nature New BioZ. 242, 246
29. LOWRY, 0. H., ROSEHROUGH, N. J., FARR, A. L., AND RAN- D~ZLL, R. J. (1951) J. BioL. Chem. 193, 265
30. HuNTII;R, W. M., AND GREENWOOD, F. C. (1962) Nalure 194, 495
31. CANFIELD, R. E., MORWN, F. J., KAMMERMAN, S., BI~xL, J. J,, AND AGOSTO, G. M. (1971) Recent Progr. Hormone Res. 27, 121
32. RATHNAM, P., AND SAXE:NA, B. B. (1971) J. Biol. Chem. 246,
REFERENCES
1. EXPELAND, D. H., NAFTOLIN, F., ,~ND PAULSEN, C. A. (1968) in Gonadotropins (ROSEMBERG, E., ed) p. 179, Geron-X, Inc., Los Altos, Calif.
2. RAJANIEMI, H., AND VANHA-PERTTULA, T. (1972) Endocri- nology 90, 1
3. KAMMERMAN, S., AND CANFIE:LD, R. E. (1972) Endocrinology 90, 384
4. RAO, CH. V., AND SAXENA, B. B. (1973) Biochim. Biophys. Acta 313, 372
5. COULSON, P., LIU, T. C., MORRIS, P., AND GORSKI, J. (1972) in Gonadotropins (SAXF;NA, B. B., BELINC, C. G., AND
GANDY, H. M., eds), p. 227, John Wiley and Sons, New York 6. DANZO, B. J,, MIDGLEY, A. R., JR., AND KLEINSMITH, L. J.
(1972) Proc. Sot. Exp. Biol. Med. 139, 88 7. TSURUHARA, T., VAN HALL, E. V., DUFAU, M. I;., AND CATT,
K. J. (1972) Endocrinology 91, 463 8. LEE, C. Y., AND RYAN, IL J. (1970) Endocrinology 89, 1515 9. LEE, C. Y., COULAM, C. B., JUNG, N. S., AND RYAN, R. J.
(1973) J. Clin. Endocrinol. Metab. 36, 148 10. HAOUR, F., RATHNAM, P., WARDLAW, S., AND SAXENA, B, B.
(1973) Fed. Proc. 32, Abstr. 1973 11. GOSPODAROWICZ, D. (1973) Fed. Proc. 32, Abstr. 373 12. CATT, K. J., T~UFAU, M, L., AND TSURUHARA, T. (1972) J.
Clin. Endocrinol 34, 123 13. CATT, K. J,, DUFAU, M. L., TSURUHARA, T. (1971) J. Chin.
Endocrinol. Metab. 32, 860 14. MEANS, A. XL., AND VAITUKAITIS, J. (1972) Endocrinology 90,39 15. SCHORR, I., RATHNAM, P., SAXB:NA, B. B., AND NEY, IL L.
(1971) J. Biol. Chem, 246, 5806 16. MARSH, J. M., AND SAVARD, K. (1966) Steroids 8, 133 17. MARSH, J. M. (1970) J. Biol. Chem. 245, 1596 18. SAVARD, K., MARSH, J. M., AND RICE, B. F. (1965) Rec. Progr.
Hormone Res, 21, 285 19. DUFAU, M. L., TSURUHARA, T., WATANABE, K., AND C~TT,
K. J. (1973) Abstracts of the 55th Annual Meeting of the Endocrinology Society, Chicago Abstr. 157, p. Al27
20. FABRIANI, A. (1973) Workshop Conference on Gunadotropins and Gonadal Steroids, Milan, May, 1973, Academic Press, New York, in press
21. BIRNBAUMER, L., AND RODBELL, M. (1969) J. Biol. Chem. 244, 3477
22. LEVEY, G. S. (1971) J. Biol. Chem. 246, 7405 23. POHL, S. L., BIRNBAUMER, L., AND RODBELL, M. (1971) J.
Biol. Chem. 246, 1849 24. JARETT, L., REUTER, M,, MCKEEL, D. W., AND SMITH, R. M.
(1971) Endocrinology 89, 1186 25. MEUNIER, J. C., OLSEN, R. W., AND CHANGEUX, J. P. (1972)
Fed. Eur. Biochem. Sot. 24, 63 26. CUATRECASAS, P. (1972) Proc. Nat. Acad. Sci. U. S. A. 69,318 27. CUATRF:CASAS, P. (1972) J. Biol. Chem. 247, 1980
33. SAXENA, B. B., LEYENDECRER, G., CHFN, W., GANDY, H. M., AND PETERSON, IL E. (1969) Acta Endocrinol. 142 (suppl), 185
34. SAXENA, B. B., DEMURER, H., GANDY, H. M., AND P~;TERSON, IL E. (1968) J. CZ in, Endocrinol. Metab. 28, 519
35. MIYACHI, Y., VAITUICAITIS, J. L., NII~:XCHLAG, E., AND LIP- SXTT, M. B. (1972) J. Clin. Endocrinol. Metab. 34, 23
36. PARLOW, A. F. (1961) in Human Pituitary Gonadotropins (ALI~II:RT, A., ed), p- 300, Charles C Thomas, Publisher, Springfield, 111
37. SAXII:NA, B. B., AND RATHNAM, P. (1967) J. Biol. Chem. 242, 3769
38. VAITUICAITIX, J. L., SHERINS, IL., Ross, G. T., HICRMAN, J., AND ASHWI~:LL, G. (1971) Endocrinology 89, 1356
39. STII:EI~M~N, S. I;., AND POHLXY, F. M. (1953) Endocrinology 63, 604
40. LIGILI IXNBICRGICR, F., 11~~ I~XICHII:RT, L. E., JR. (1972) DfLdo- crinotogy 91, 135
41. SCATCHAI~D, CL (1949) Ann. N. Y. Acad. Sci, fil, 660 42. DWIGGINS, C. W., JR., BOLEN, II. J., AND DUNNING, H. N.
(1960) J. Physiol. Chem. 64, 1175 43. RATHNAM, P., AND SAXENA, B. B. (1970) J. Biol. Chem. 246,
3725 44. FAIRBANKS, G., STXCIC, T. L., AND WALLACH, I). F. H. (1971)
Biochemistry 10, 2606 45. COLE, F, E., AND RICJI:, B. F, (1973) Abstracts of the 55th An-
nual Meeting of the Endocrinology Society, Chicago, Abstr. 315, p. A206
46. KAHN, C. R., NEVILLE, D. M., JR., AND ROTH, J. (1973) J. Biol. Chem. 240, 244
47. CUATRECASAS, P. (1971) J. BioL. Chem. 246,7265 48. RAO, CH. V., SAXENA, B. B., AND GANDY, H. M. (1972) in
Gonadotropins (SAXII:NA, B. B., BELING, C. G., AND GANDY, H. M., eds), p. 261, John Wiley and Sons, New York
49. CHANNING, C. P., AND KAMMERMIZN, S. (1962) EndocrinoEogy 92,531
50. SAVARD, K. (1973) Biol, Reprod. 8, 183 51. AMIR, S, MI., CARRAWAY, T. F., JR., KOHN, L. D., AND WI-
NAND, R. J. (1973) J. Biol. Chem. 248, 4092 52. RODT~I!;LL, M., KRANS, H. M. J., POHL, S. L., AND BIRNRAUMER,
L. (1971) J. BioZ. Chem. 246, 1861 53. DE LA LLOSA, P., AND JUSTISZ, M. (1969) Biochim. Biophys.
Acta 181, 426 54. WALLEWIIC, K. (1973) J. Biol. Chem. 248, 2650 55. GWYNNI~, J. T., AND TANFORD, C. (1970) J. Biol. Chem. 246,
3269 56. GARLAND, R. C., AND CORI, C. F. (1972) Biochemistry 11,4712 57. LI~;FKOWITZ, R. J., HABER, E., AND O’HARA, I>. (1972) PTOC.
Nat. Acad. Sci. U, S. A. 69, 2828 58. SUTHERLAND, E. W., ROI~ISON, G. A., AND BUTCHER, R. W.
(1968) Circulation 37, 279 59. GOSPODAROWXZ, D. (1973) J. Biol. Chem. 248, 5050
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from

France Haour and Brij B. SaxenaLuteum
Characterization and Solubilization of Gonadotropin Receptor of Bovine Corpus
1974, 249:2195-2205.J. Biol. Chem.
http://www.jbc.org/content/249/7/2195Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/249/7/2195.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on September 22, 2017
http://ww
w.jbc.org/
Dow
nloaded from