characterization of protein kinase c in early xenopus...
TRANSCRIPT

Development 110, 461-470 (1990)Printed in Great Britain © The Company of Biologists limited 1990
461
Characterization of protein kinase C in early Xenopus embryogenesis
ARIE P. OTTE, IJSBRAND M. KRAMER, MAURICE MANNESSE, CARO LAMBRECHTS and
ANTONY J. DURSTON
Hubrecht Laboratory, Netherlands Institute for Developmental Biology, Uppsalalaan 8, 3584 CT UTRECHT, The Netherlands
Summary
Recently, we presented evidence that protein kinase C(PKC) is involved in mediating the endogenous signalsthat induce competent Xenopus ectoderm to differentiateto neural tissue. We report here that PKC is alreadystrongly activated in neural-induced ectoderm frommidgastrula embryos and that this activation runsparallel with an increase in the level of inositolphosphates. We further identify several proteins that arephosphorylated, both in natural neural-induced ecto-derm and in TPA-treated ectoderm, suggesting that theyare phosphorylated through the PKC route. We foundno major changes in PKC activity among different
pregastrula stages, including the unfertilized egg.However, PKC isolated from animal, ectodermal cells ishighly sensitive to Ca2+ and can be activated by lowconcentrations, (6-25 /iM) of arachidonic acid, whilePKC isolated from vegetal, endodermal cells is moreinsensitive to Ca2+ and cannot be activated by arachi-donic acid. These results suggest that different PKCisozymes are present in animal and vegetal cells.
Key words: protein kinase C, signal transduction, neuralinduction, Xenopus.
Introduction
Protein kinase C (PKC) plays an important role in themechanisms by which external signals are transmittedto the interior of cells. Several growth factors andhormones that bind to, and activate, receptors on thecell surface, stimulate inositol phospholipid turnovervia phospholipase C activation, thereby producing thesecond messengers diacylglycerol and inositol phos-phates, of which diacylglycerol activates PKC (Nishi-zuka, 1984, 1986; Berridge, 1987; Berridge and Irvine,1989). Several growth-factors and hormones thatstimulate inositol phospholipid turnover, induce trans-location of PKC from the cytosol to the membrane(Niedel and Blackshear, 1986). PKC also serves as thereceptor for phorbol esters, a class of tumour promoters(Castagna et al. 1982). One of them, 12-0-tetradeca-noyl phorbol-13-acetate (TPA), activates PKC ef-ficiently and causes PKC to translocate from the cytosolto the membrane.
There are clues suggesting that the PKC/inositolphosphate pathway plays an essential role in earlyembryogenesis. Fertilization increases the polyphos-phoinositide content of sea urchin eggs (Turner et al.1984) and induces changes in phosphatidylinositolturnover (Dworkin and Dworkin-Rastl, 1989). Ionto-phoresis of inositol trisphosphate activates Xenopus(Busa et al. 1985) and sea urchin eggs (Whitaker andIrvine, 1984). Also, TPA activates mouse oocytes(Cuthbertson and Cobbold, 1985), sea urchin eggs
(Swann and Whitaker, 1985) and Xenopus eggs(Bement and Capco, 1989). In addition, TPA caused acortical contraction when applied to unfertilized Ranapipiens eggs, suggesting that it causes Ca2+ release(Zimmerman, 1985). In Xenopus oocytes, meioticmaturation is induced by TPA (Stith and Mailer, 1987;Smith, 1989) and facilitated by injection of proteinkinase C (Kamata and Kung, 1990). These studiesindicate a role for PKC and phosphoinositol phosphateturnover in the maturation of oocytes and in fertiliz-ation.
Another clue about the roles of PKC and the inositolphosphates in early embryogenesis comes from exper-iments with Li+. This ion is thought to block theprocessing of inositol phosphates, resulting in depletionof the phosphatidylinositol phosphate pools (reviewedin Berridge et al. 1989). Short treatments of 32- to 64-cell Xenopus embryos with Li+ dorsalizes theseembryos (Kao et al. 1986). Injection of Li+ into aventral blastomere also leads to dorsalization. Import-antly, these teratogenic effects can be annihilated whenmyo-inositol is coinjected with Li+ into a ventralblastomere (Busa and Gimlich, 1989; for an excellentreview on this subject see Berridge et al. 1989). Busaand Gimlich also found that activation of PKC by TPAoverrules the effect of Li+, indicating a role for PKC inestablishing the dorsoventral polarity of the embryo.
Another clue about the role of PKC in earlyembryogenesis comes from our own work. We foundthat PKC is translocated from the cytosol to the

462 A. P. Otte and others
membrane in neural-induced ectoderm, but not in non-induced ectoderm. We also found that TPA causestranslocation of Xenopus PKC and induces competentectoderm to differentiate to neural tissue (Otte et al.1988).
In this paper, we show that there are several proteinsthat become strongly phosphorylated in neural-inducedectoderm, either after artificial PKC activation orduring natural neural induction. An increased level ofinositol phosphates was also detected in neural-inducedectoderm. We show further that PKC, which is presentin animal, ectodermal cells, is highly sensitive to Ca2+
and can be activated by low concentrations of arachi-donic acid. PKC, which is present in the vegetal,presumptive endodermal cells, is more independent ofCa2+ for its activation and cannot be activated byarachidonic acid. These observations suggest that thereare different PKC isozymes present in early embryo-genesis.
Materials and methods
Culture conditionsEmbryos were obtained by natural fertilization via standardprocedures. Embryonic stages were determined according toNieuwkoop and Faber (1967). Embryos were dejellied in 2 %cysteine-hydrochloride (Sigma) (pH8.0). Explants werecultured in 100% Flickinger solution (NaCl, 58 mM; KC1,IITIM; NaHCO3, 0.24 HIM; Na2HPO4, IIUM; KH2PO4)
0.2mM, CaCl2) 0.5HIM; MgSO4, IITIM) (pH7.5).
Protein kinase C assayEmbryos or explants were washed and homogenized in ice-cold lysis buffer (20 mM Tris pH7.5, 2mM EDTA, 0.5 mMEGTA, 5mM /3-mercaptoethanol, 2mM phenylmethylsul-phonyl fluoride and 0.01 mgml"1 leupeptin). The homogen-ate was centrifuged at lOOOOg for 15s, the pellet washomogenized again, centrifuged and the supernatants werecombined. The pellet, containing yolk platelets and organ-elles, was discarded, as it contained no PKC activity. Thesupernatant was spun at 100000g for lh. The 100000gsupernatant, considered to be the cytosolic fraction, wasremoved. The pellet was used for the in vitro phosphorylationexperiments described in Fig. 3. To isolate membrane-boundPKC, the pellet was resuspended and shaken regularly for45 min in lysis buffer containing 1 % Nonidet-40 (Sigma). Themixture was centrifuged for 15 min at 10 000 £ and the pelletdiscarded. The supernatant was considered to be thepaniculate or membrane fraction. When total (soluble andpaniculate) PKC was isolated from tissue, the tissue washomogenized in 1 ml ice-cold lysis buffer containing 1 %Nonidet-40 and shaken regularly for 45 min. After that, thehomogenate was spun at lOOOOg for lOmin. The supernatantwas loaded onto a DEAE column. Partial purification of PKCwith DEAE-cellulose was performed exactly as described(Otte et al. 1988). The PKC assay, in which histone III S wasused as a substrate, was performed exactly as describedpreviously (Otte et al. 1988). The PKC assay in which anonapeptide, derived from the EGF receptor (Val-Arg-Lys-Arg-Thr-Leu-Arg-Arg-Leu-NH2) (Auspep, South Mel-bourne, Australia) was used as substrate, was essentiallyperformed as described by House et al. (1987) and House andKemp (1987). 25/il of DEAE PKC eluate was added to 60^1of reaction mix containing (as the final concentration) 20 mM
Tris pH7.5, 7.5 mM MgAc, lmgml l leupeptin, 10 ^M EGFreceptor peptide, 0.25 mg ml"1 BSA, 0.2 mM ATP,l.SxlO^ctsmin"1 [32P]>^ATP (New England Nuclear) andeither imM EGTA (background) or lmin Ca2+, 8/igml"1
phosphatidylserine (PS) (Spinal cord, Lipid Products),0.8/igml"1 diacylglycerol (Diolein, Sigma) (assay). Lipidswere stored under nitrogen at — 20°C in chloroform/methanol3:1. They were dried using a flow of nitrogen, suspended in20 mM Tris pH7.5 and sonicated for 3 min to obtainliposomes. Different Ca2+-concentrations were obtained byusing Ca2+-EGTAbuffers (Bartfai, 1979). The reactions werestarted by adding 25 /A PKC eluate to 60 /A reaction mix. Theincorporation of 32PO4 at 30°C was linear with time up to60min. After 15 min, the reactions were stopped by adding25/A 25 % TCA. After 20min on ice, the samples were spun(4°C) at lOOOOg for lOmin and 90 fA of the supernatant wasthen spotted onto a Whatman P-81 phosphocellulose filter andthis was immediately transferred to 400 ml 75 mM phosphoricacid solution in which a magnetic stirrer rotated slowly. Afterchanging the phosphoric acid solution 4 times (10-15 minrotation each), the filters were dried and counted. We foundidentical results using either histone HI S or the EGF-receptornonapeptide as substrate. Only the absolute value of thespecific activities differed (compare with Otte et al. 1988), dueto different numbers of phosphorylation sites in histone III Sand the nonapeptide. Using the nonapeptide as a substratehas the advantage that lower backgrounds can be obtained,leading to a better signal to noise ratio.
Substrate phosphorylation and gel electrophoresisExplants were incubated for 3h in 5 ml Holtfreter solution(59mM NaCl, 0.7 mM KC1, 0.9 mM CaCl2) containing 80/*Ci[32P]orthophosphate. After incubation, the explants werewashed and homogenized in sample buffer consisting of 8 Murea, 2 % Nonidet P40 and 0.1 % ampholytes pH3-10. Two-dimensional electrophoresis was carried out according toO'Farrell (1975), using isoelectric focusing (2% ampholytespH3-10, Sigma) in the first dimension and SDS-polyacryla-mide gel electrophoresis (10 % polyacrylamide) in the seconddimension.
Measurement of inositolphosphates andphosphatidylinositolphosphatesDetermination of total inositol phosphates was performedessentially according to methods described by Tilly et al.(1987) and by Dworkin and Dworkin-Rastl (1989). Determi-nation of the phosphatidylinositol phosphates was performedessentially according to methods described by Lacal et al.(1987) and by Dworkin and Dworkin-Rastl (1989). Fertilizedeggs were microinjected with 40 nl of 20 mM Tris (pH7.2)containing 2>yC\ of [3H]myoinositol (Amersham, 3.5 Cimmol"1). The eggs were cultured to stage 10 and entireectoderm explants were dissected, either with or without asmall piece (<20%) of dorsal mesoderm left attached. Theexplants were cultured for as long as it took control stage 10embryos to reach stage Hi. The explants were thenhomogenized in 2 ml chloroform :methanol: 12 N HC1(200:100:0.75). After a lOmin extraction (RT) 8/d Folch II(Sigma) was added and the mixture was vortexed. 500 /A 0.6 NHC1 was added and the mixture was centrifuged at 3000 g for5 min. The upper water phase was removed and the lowerphase was washed twice with 1 ml chloroform:methanol:0.6NHC1 (3:48:47). The water phases from these sequential washeswere combined. The remaining chloroform and methanol wasremoved by evaporation under a flow of nitrogen. An equalvolume of 10 % TCA was added to the water phase to removeproteins. The mixture was centrifuged at 4000g for 10 min.
Protein kinase C in early Xenopus embryogenesis 463
The supernatant was washed three times with an equalvolume (±3 ml) of ether. The remaining water phase wasneutralized by adding 0.5 ml 1M Tris (pH5=10). The volumewas determined and 100/J was counted by scintillationcounting to determine the input of [3H]inositol. The waterphase was loaded on a 0.5 ml AG-1X8 (Biorad) 100-200 meshformate form column and washed with 2x2 ml H2O and4x2 ml 60mM ammonium formate/5 mM Borax buffer.
Total IPn was eluted with 4x2ml 1M ammonium formate/0.1 M formic acid buffer. 4x0.5 ml of each eluate was countedafter addition of 100 iA 1 M Tris pH 8.5 to neutralize the eluate.The lipid phase was dried under a flow of nitrogen and kept at-20°C until use. The dried lipids were then taken up in lmlchloroform:methanol:H2O (75/25/2) and dried again under aflow of nitrogen. The dried lipids were taken up in 25 jA ofchloroform:methanol:H2O (75:25:2) of -20°C and spotted onpre-coated TLC plates, Silica Gel 60 (Merck). The plateswere activated by spraying with 1.2% potassium-oxalate inH2O:methanol (60:40) and heating at 120°C for lh . Afteractivation, the plates were used within 30min. Plates weredeveloped in a saturated atmosphere containing chloroform:methanol:acetone:acetic acid:H2O (40:15:15:12:8). Phospho-lipid standards (phosphatidylinosites, phosphatidic acid fromSigma) were used and localized by I2 vapour. The spotscontaining PI, PIP and PIP2 were scraped and shakenovernight in 0.6 ml 0.6 N HCl:methanol (60:40) and counted.
We found no differences either in inositol phosphate orphosphatidylinositol phosphate levels between stage 8,10, Hiand 13 ectoderm explants, indicating that the phosphatidyl-inositol phosphate pool was labelled to equilibrium.
Results
The timing of protein kinase C translocation duringneural inductionRecently, we showed that contact with dorsal meso-derm causes a specific translocation of protein kinase C(PKC) to the plasma-membrane in the induced neuro-ectoderm during neural induction (Otte etal. 1988). Wethen compared two developmental stages: the earlygastrula (stage 10) and the early neurula (stage 13)(Nieuwkoop and Faber, 1967). It has been shown,however, that the expression of the neural cell adhesionmolecule, N-CAM, starts in response to neural induc-tion between stages 10 and 12 and increases stronglybetween stages 12 and 14 (Kintner and Melton, 1987).We also showed that cAMP concentration and adeny-late cyclase activity increase strongly and specifically inthe induced neuroectoderm, from stage 12 onwards(Otte et al. 1989). We therefore wanted to know when,during neural induction, PKC is translocated from thecytoplasm to the plasma membrane, in neural-inducedectoderm.
We dissected entire stage 10 ectoderm explants with asmall piece (<20%) of the neural-inducing dorsalmarginal zone mesoderm left attached and culturedthese, for as long as it took control stage 10 embryos toreach stages 11, 13 and 16. We routinely cultured someof these explants till stage 38-40. These explantscontained invariably neural tissue as determined viaclassical histology and using a monoclonal antibodyagainst neural tissue (Jones and Woodland, 1989). As acontrol, we also cultured stage 10 ectoderm explants
_r loo-Ofi
oo.
u
IStage 10 10 11 13 16 16
hrs 9.5 9.5 12.5 16 22 22
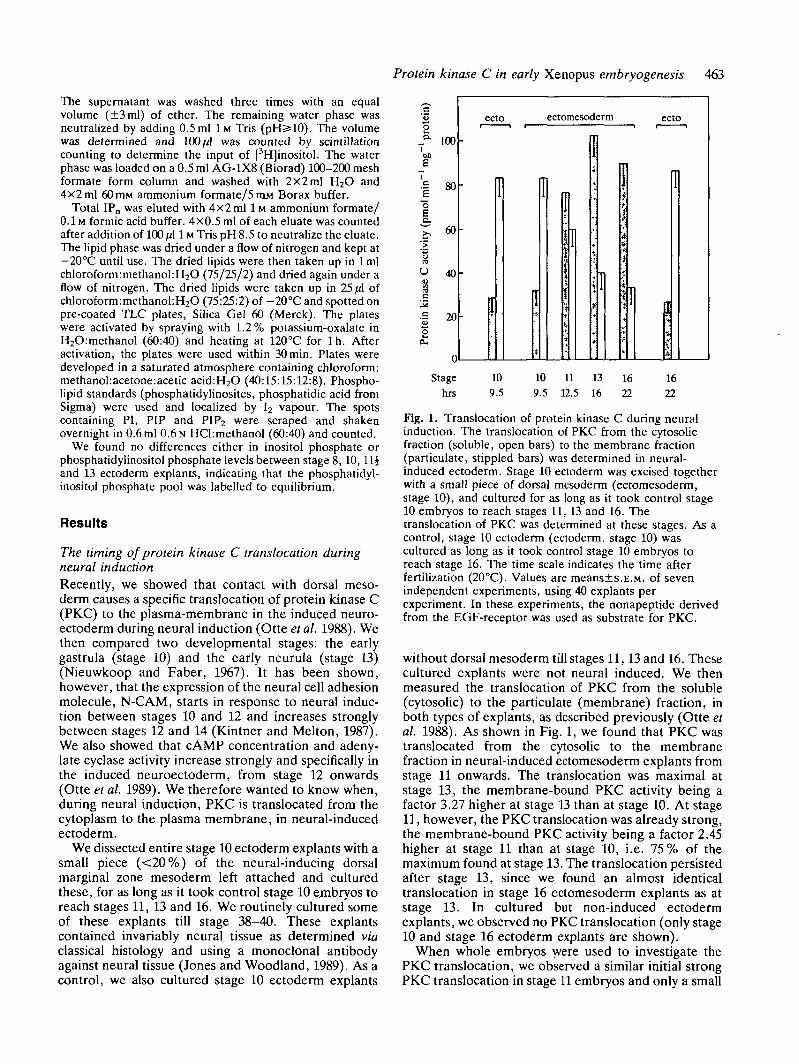
Fig. 1. Translocation of protein kinase C during neuralinduction. The translocation of PKC from the cytosolicfraction (soluble, open bars) to the membrane fraction(particulate, stippled bars) was determined in neural-induced ectoderm. Stage 10 ectoderm was excised togetherwith a small piece of dorsal mesoderm (ectomesoderm,stage 10), and cultured for as long as it took control stage10 embryos to reach stages 11, 13 and 16. Thetranslocation of PKC was determined at these stages. As acontrol, stage 10 ectoderm (ectoderm, stage 10) wascultured as long as it took control stage 10 embryos toreach stage 16. The time scale indicates the time afterfertilization (20°C). Values are means±s.E.M. of sevenindependent experiments, using 40 explants perexperiment. In these experiments, the nonapeptide derivedfrom the EGF-receptor was used as substrate for PKC.
without dorsal mesoderm till stages 11,13 and 16. Thesecultured explants were not neural induced. We thenmeasured the translocation of PKC from the soluble(cytosolic) to the particulate (membrane) fraction, inboth types of explants, as described previously (Otte etal. 1988). As shown in Fig. 1, we found that PKC wastranslocated from the cytosolic to the membranefraction in neural-induced ectomesoderm explants fromstage 11 onwards. The translocation was maximal atstage 13, the membrane-bound PKC activity being afactor 3.27 higher at stage 13 than at stage 10. At stage11, however, the PKC translocation was already strong,the membrane-bound PKC activity being a factor 2.45higher at stage 11 than at stage 10, i.e. 75 % of themaximum found at stage 13. The translocation persistedafter stage 13, since we found an almost identicaltranslocation in stage 16 ectomesoderm explants as atstage 13. In cultured but non-induced ectodermexplants, we observed no PKC translocation (only stage10 and stage 16 ectoderm explants are shown).
When whole embryos were used to investigate thePKC translocation, we observed a similar initial strongPKC translocation in stage 11 embryos and only a small
464 A. P. Otte and others
further increase in PKC translocation at stage 13(results not shown). We conclude that PKC is translo-cated from the cytoplasm to the plasma membraneduring an early phase in neural induction.
Protein phosphorylation in neural induced ectoderm .We next investigated the phosphorylation of proteins inneural-induced ectoderm. Entire stage 10 ectodermexplants were incubated in [32P]orthophosphate-containing medium for 3h; this is as long as it tookcontrol stage 10 embryos to reach stage 11, the periodduring which PKC is almost maximally translocated(see Fig. 1). We also incubated ectoderm under thesame conditions, for the same period, but in thepresence of 350nM TPA or with a small piece (<20%)of the dorsal mesoderm left attached.
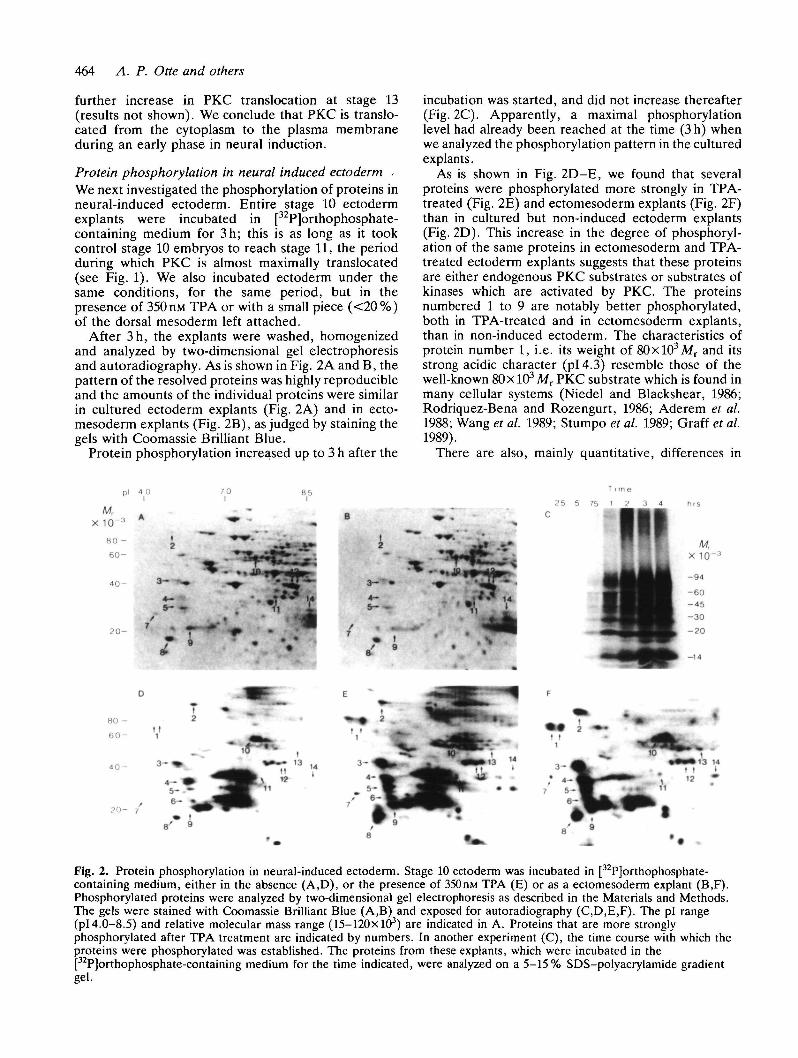
After 3h, the explants were washed, homogenizedand analyzed by two-dimensional gel electrophoresisand autoradiogTaphy. As is shown in Fig. 2A and B, thepattern of the resolved proteins was highly reproducibleand the amounts of the individual proteins were similarin cultured ectoderm explants (Fig. 2A) and in ecto-mesoderm explants (Fig. 2B), as judged by staining thegels with Coomassie Brilliant Blue.
Protein phosphorylation increased up to 3 h after the
incubation was started, and did not increase thereafter(Fig. 2C). Apparently, a maximal phosphorylationlevel had already been reached at the time (3 h) whenwe analyzed the phosphorylation pattern in the culturedexplants.
As is shown in Fig. 2D-E, we found that severalproteins were phosphorylated more strongly in TPA-treated (Fig. 2E) and ectomesoderm explants (Fig. 2F)than in cultured but non-induced ectoderm explants(Fig. 2D). This increase in the degree of phosphoryl-ation of the same proteins in ectomesoderm and TPA-treated ectoderm explants suggests that these proteinsare either endogenous PKC substrates or substrates ofkinases which are activated by PKC. The proteinsnumbered 1 to 9 are notably better phosphorylated,both in TPA-treated and in ectomesoderm explants,than in non-induced ectoderm. The characteristics ofprotein number 1, i.e. its weight of 80xl03Mr and itsstrong acidic character (pi 4.3) resemble those of thewell-known 80 x 103 Mr PKC substrate which is found inmany cellular systems (Niedel and Blackshear, 1986;Rodriquez-Bena and Rozengurt, 1986; Aderem et al.1988; Wang et al. 1989; Stumpo et al. 1989; Graff et al.1989).
There are also, mainly quantitative, differences in
pi 4 0 70I 85
M,X10"3
80 -
6 0 -
4 0 -
2 0 -
5 - —
3 - " •
5
•••««»
80 -
6 0 -
4 0 -
t l
I13
4— I5 -
e' 9
n
f —
Fig. 2. Protein phosphorylation in neural-induced ectoderm. Stage 10 ectoderm was incubated in [32P]orthophosphate-containing medium, either in the absence (A,D), or the presence of 350niu TPA (E) or as a ectomesoderm explant (B,F).Phosphorylated proteins were analyzed by two-dimensional gel electrophoresis as described in the Materials and Methods.The gels were stained with Coomassie Brilliant Blue (A,B) and exposed for autoradiography (C,D,E,F). The pi range(pi 4.0-8.5) and relative molecular mass range (15-120X103) are indicated in A. Proteins that are more stronglyphosphorylated after TPA treatment are indicated by numbers. In another experiment (C), the time course with which theproteins were phosphorylated was established. The proteins from these explants, which were incubated in thepPJorthophosphate-containing medium for the time indicated, were analyzed on a 5-15% SDS-polyacrylamide gradientgel.

Protein kinase C in early Xenopus embryogenesis 465
protein phosphorylation between TPA-treated(Fig. 2E) and ectomesoderm explants (Fig. 2F). No-tably proteins 2, 3, 4, 5, 9 and 14 are more stronglyphosphorylated in ectomesoderm than in TPA-treatedexplants.
Some phosphorylated proteins are localized in themembranes of neural-induced ectodermIt is known of several PKC substrates that they arepresent in, or become translocated to, the plasmamembrane (Aderem et al. 1988; Wang et al. 1989). Thislocalization would facilitate phosphorylation by thetranslocated and activated membrane-bound PKC. Weinvestigated whether the same is true for the phos-phorylated proteins that we identified (above), inneural-induced ectoderm. We therefore isolated mem-branes from ectoderm and ectomesoderm explantswhich were cultured as long as it took control stage 10embryos to reach stage 11. We also isolated membranesfrom stage 10 ectoderm explants, which had beentreated with 500nM TPA for 30min, a treatment thatcauses a maximal PKC translocation (Otte et al. 1988).These membranes were prepared exactly as when thetranslocation of PKC was measured, i.e. homogenizedtissue was centrifuged at 100000g for lh . Themembranes were then homogenized and lmM Ca2+,diacylglycerol/phosphatidylserine liposomes, and[32P]y-ATP were then added. After 20 min incubation at30cC, phosphorylation of the membrane proteins wasanalyzed by two-dimensional gel electrophoresis andautoradiography.
As is shown in Fig. 3, we found that some of thephosphorylated proteins that were identified in intactcells (Fig. 2) are present and strongly phosphorylated inmembranes from TPA-treated ectoderm and ectomeso-derm explants (Fig. 3B and C) but are much lessphosphorylated in membranes from cultured, but non-treated ectoderm explants (Fig. 3A). The more basicproteins (numbers 10 to 14), that are not the most
strongly phosphorylated in vivo, (compare with Fig. 2),are notably well phosphorylated in membranes thatcontain translocated and activated PKC (Fig. 3B andC). The most-strongly phosphorylated protein in thesemembranes is the 80xl(^MT protein (protein number1), which is only poorly phosphorylated in membranesfrom non-induced ectoderm explants (Fig. 3A).
These results show that some of the major proteinsthat are phosphorylated in vivo are present in mem-branes from neural-induced ectoderm. This ectodermcontains increased PKC activity in its membranes, dueto PKC translocation during neural induction, and it ispossible that these proteins are translocated to themembrane, concomitantly with PKC. We cannot,however, exclude from these experiments that theseproteins were already present in the membrane.
Neural-induced ectoderm contains a higher level ofinositol phosphatesCell-surface receptor mediated hydrolysis of phosphati-dylinositol bisphosphate via phospholipase C raises thelevels of diacylglycerol and of inositol phosphates. Wetested whether phospholipase C is activated concomi-tantly with PKC activation, during neural induction,leading to an increase in the level of inositol phos-phates. Fertilized eggs were injected with 3^Ci[3H]myoinositol per egg and cultured to stage 10. Thisperiod has been shown to be long enough to label theendogenous phosphatidylinositol lipid pools in Xeno-pus embryos to equilibrium (Lacal et al. 1987; seeMaterials and methods). At stage 10, ectoderm andectomesoderm explants were excised and cultured aslong as it took control stage 10 embryos to reach stage11. We then measured the levels of inositol phosphatesand phosphatidylinositol phosphates in neural-inducedand non-induced ectoderm at stage 11, since PKC is stillin the process of being activated during neuralinduction at this stage (Fig. 1).
The stage 11 explants were homogenized and total
1fO13 V»
f c
Fig. 3. Some of the in vivo phosphorylated proteins are present in the membranes of neural-induced ectoderm. Stage 10ectoderm was excised without dorsal mesoderm (A and B), and either treated with 500 nM TPA for 30 min (B) or not (A)or else was excised with the dorsal mesoderm left attached (C). The non-treated ectoderm explants and ectomesodermexplants were then cultured as long as it took control stage 10 embryos to reach stage 16. Membrane preparations wereisolated from the explants and incubated in the presence of [32P]y-ATP, diacylglycerol/phospatidylserine liposomes andlmM Ca2+ at 30°C for 20min. After incubation, the proteins were analyzed by two-dimensional gel electrophoresis andautoradiography. The pi and molecular weight range is as indicated as in Fig. 2A. The phosphorylated proteins, which arenumbered, correspond to the phosphorylated proteins in Fig. 2.
466 A. P. Otte and others
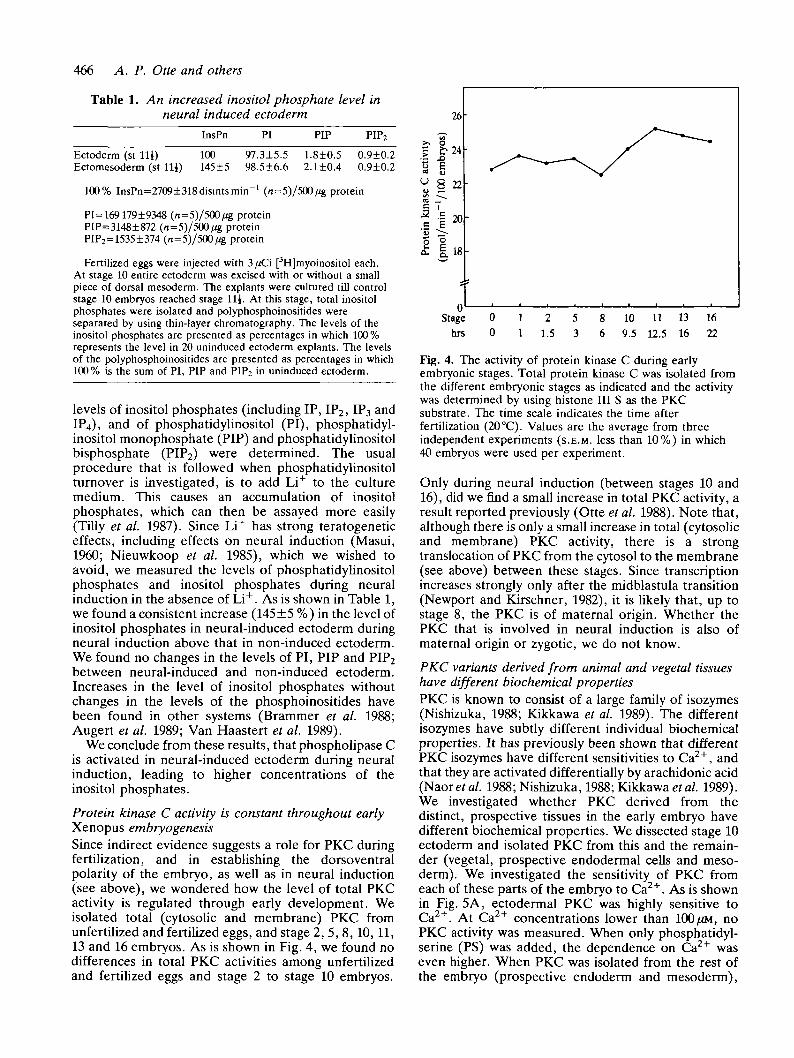
Table 1. An increased inositol phosphate level inneural induced ectoderm
InsPn PI PIP PIP2
Ectoderm (st Hi) 100 97.3±5.5 1.8±0.5 0.9±0.2Ectomesoderm (st Hi) 145±5 98.5±6.6 2.1±0.4 0.9±0.2
100% InsPn=2709±318disintsmin"1 (n=5)/500/ig protein
PI=169179±9348 (n=5)/5O0/ig proteinPIP=3148±872 (n=5)/500/*g proteinPrP2=1535±374 (n=5)/500/ig protein
Fertilized eggs were injected with 3/iCi [3H]myoinositol each.At stage 10 entire ectoderm was excised with or without a smallpiece of dorsal mesoderm. The explants were cultured till controlstage 10 embryos reached stage Hi. At this stage, total inositolphosphates were isolated and polyphosphoinositides wereseparated by using thin-layer chromatography. The levels of theinositol phosphates are presented as percentages in which 100 %represents the level in 20 uninduced ectoderm explants. The levelsof the polyphosphoinositides are presented as percentages in which100% is the sum of PI, PIP and PIP2 in uninduced ectoderm.
levels of inositol phosphates (including IP, IP2, IP3 andIP4), and of phosphatidylinositol (PI), phosphatidyl-inositol monophosphate (PIP) and phosphatidylinositolbisphosphate (PIP2) were determined. The usualprocedure that is followed when phosphatidylinositolturnover is investigated, is to add Li+ to the culturemedium. This causes an accumulation of inositolphosphates, which can then be assayed more easily(Tilly et al. 1987). Since Li+ has strong teratogeneticeffects, including effects on neural induction (Masui,1960; Nieuwkoop et al. 1985), which we wished toavoid, we measured the levels of phosphatidylinositolphosphates and inositol phosphates during neuralinduction in the absence of Li+. As is shown in Table 1,we found a consistent increase (145 ±5 %) in the level ofinositol phosphates in neural-induced ectoderm duringneural induction above that in non-induced ectoderm.We found no changes in the levels of PI, PIP and PIP2between neural-induced and non-induced ectoderm.Increases in the level of inositol phosphates withoutchanges in the levels of the phosphoinositides havebeen found in other systems (Brammer et al. 1988;Augert et al. 1989; Van Haastert et al. 1989).
We conclude from these results, that phospholipase Cis activated in neural-induced ectoderm during neuralinduction, leading to higher concentrations of theinositol phosphates.
Protein kinase C activity is constant throughout earlyXenopus embryogenesisSince indirect evidence suggests a role for PKC duringfertilization, and in establishing the dorsoventralpolarity of the embryo, as well as in neural induction(see above), we wondered how the level of total PKCactivity is regulated through early development. Weisolated total (cytosolic and membrane) PKC fromunfertilized and fertilized eggs, and stage 2, 5, 8,10,11,13 and 16 embryos. As is shown in Fig. 4, we found nodifferences in total PKC activities among unfertilizedand fertilized eggs and stage 2 to stage 10 embryos.
Stagehis
00
11
21.5
53
86
109.5
1112.5
1316
1622
Fig. 4. The activity of protein kinase C during earlyembryonic stages. Total protein kinase C was isolated fromthe different embryonic stages as indicated and the activitywas determined by using histone III S as the PKCsubstrate. The time scale indicates the time afterfertilization (20 °C). Values are the average from threeindependent experiments (S.E.M. less than 10%) in which40 embryos were used per experiment.
Only during neural induction (between stages 10 and16), did we find a small increase in total PKC activity, aresult reported previously (Otte et al. 1988). Note that,although there is only a small increase in total (cytosolicand membrane) PKC activity, there is a strongtranslocation of PKC from the cytosol to the membrane(see above) between these stages. Since transcriptionincreases strongly only after the midblastula transition(Newport and Kirschner, 1982), it is likely that, up tostage 8, the PKC is of maternal origin. Whether thePKC that is involved in neural induction is also ofmaternal origin or zygotic, we do not know.
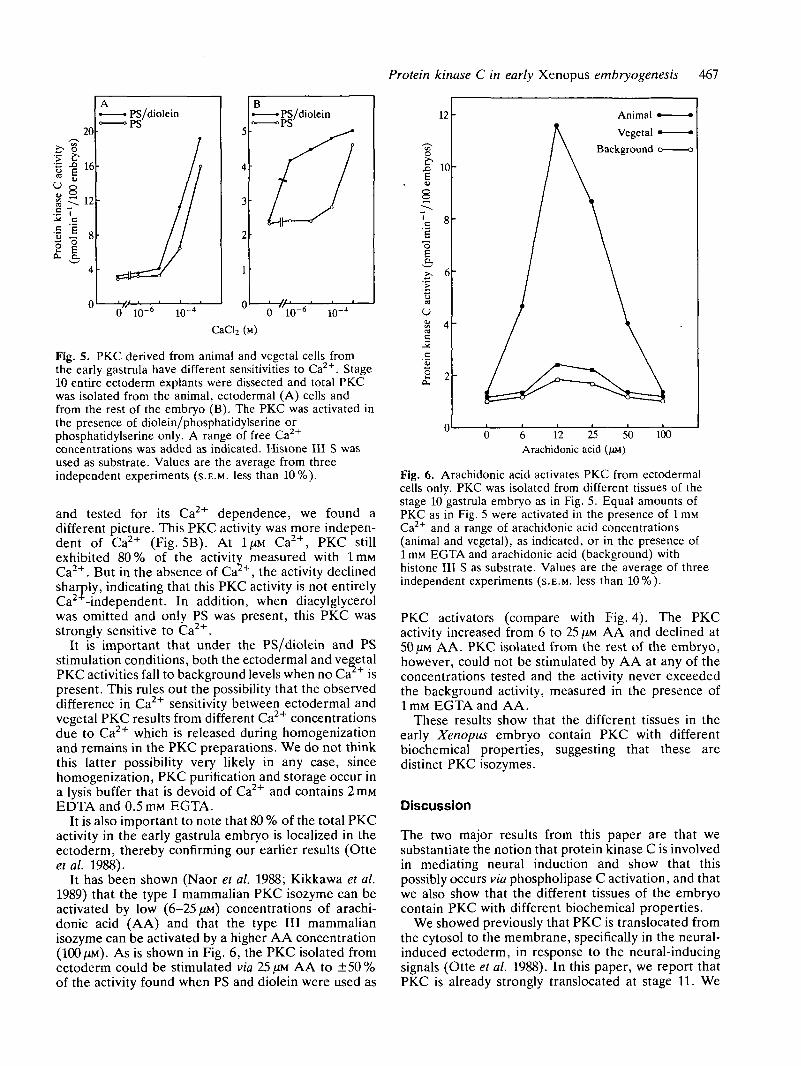
PKC variants derived from animal and vegetal tissueshave different biochemical propertiesPKC is known to consist of a large family of isozymes(Nishizuka, 1988; Kikkawa et al. 1989). The differentisozymes have subtly different individual biochemicalproperties. It has previously been shown that differentPKC isozymes have different sensitivities to Ca2+, andthat they are activated differentially by arachidonic acid(Naorera/. 1988; Nishizuka, 1988; Kikkawa et al. 1989).We investigated whether PKC derived from thedistinct, prospective tissues in the early embryo havedifferent biochemical properties. We dissected stage 10ectoderm and isolated PKC from this and the remain-der (vegetal, prospective endodermal cells and meso-derm). We investigated the sensitivity of PKC fromeach of these parts of the embryo to Ca2+. As is shownin Fig. 5A, ectodermal PKC was highly sensitive toCa2+. At Ca2+ concentrations lower than 100(JM, noPKC activity was measured. When only phosphatidyl-serine (PS) was added, the dependence on Ca2+ waseven higher. When PKC was isolated from the rest ofthe embryo (prospective endoderm and mesoderm),
Protein kinase C in early Xenopus embryogenesis 467
0 10- 1(T4 " 0 10"6 10"4
CaCl2 (M)
Fig. 5. PKC derived from animal and vegetal cells fromthe early gastrula have different sensitivities to Ca2+. Stage10 entire ectoderm explants were dissected and total PKCwas isolated from the animal, ectodermal (A) cells andfrom the rest of the embryo (B). The PKC was activated inthe presence of diolein/phosphatidylserine orphosphatidylserine only. A range of free Ca2+
concentrations was added as indicated. Histone III S wasused as substrate. Values are the average from threeindependent experiments (S.E.M. less than 10%).
and tested for its Ca2+ dependence, we found adifferent picture. This PKC activity was more indepen-dent of Ca2+ (Fig. 5B). At 1/XM Ca2+, PKC stillexhibited 80% of the activity measured with lmMCa2+. But in the absence of Ca , the activity declinedsharply, indicating that this PKC activity is not entirelyCa -independent. In addition, when diacylglycerolwas omitted and only PS was present, this PKC wasstrongly sensitive to Ca2+.
It is important that under the PS/diolein and PSstimulation conditions, both the ectodermal and vegetalPKC activities fall to background levels when no Ca + ispresent. This rules out the possibility that the observeddifference in Ca2+ sensitivity between ectodermal andvegetal PKC results from different Ca2+ concentrationsdue to Ca2+ which is released during homogenizationand remains in the PKC preparations. We do not thinkthis latter possibility very likely in any case, sincehomogenization, PKC purification and storage occur ina lysis buffer that is devoid of Ca2+ and contains 2ITIMEDTA and 0.5mM EGTA.
It is also important to note that 80 % of the total PKCactivity in the early gastrula embryo is localized in theectoderm, thereby confirming our earlier results (Otteetal. 1988).
It has been shown (Naor et al. 1988; Kikkawa et al.1989) that the type I mammalian PKC isozyme can beactivated by low (6-25/ZM) concentrations of arachi-donic acid (AA) and that the type III mammalianisozyme can be activated by a higher AA concentration(100 JXM). As is shown in Fig. 6, the PKC isolated fromectoderm could be stimulated via 25 (JM AA to ±50 %of the activity found when PS and diolein were used as
12
.n 10
8
o
a.
U
£ 2
Animal <
Vegetal
Background
6 12 25Arachidonic acid
50 100
Fig. 6. Arachidonic acid activates PKC from ectodermalcells only. PKC was isolated from different tissues of thestage 10 gastrula embryo as in Fig. 5. Equal amounts ofPKC as in Fig. 5 were activated in the presence of 1 mMCa2+ and a range of arachidonic acid concentrations(animal and vegetal), as indicated, or in the presence of1 mM EGTA and arachidonic acid (background) withhistone III S as substrate. Values are the average of threeindependent experiments (S.E.M. less than 10%).
PKC activators (compare with Fig. 4). The PKCactivity increased from 6 to 25 JIM AA and declined at50 /JM AA. PKC isolated from the rest of the embryo,however, could not be stimulated by AA at any of theconcentrations tested and the activity never exceededthe background activity, measured in the presence oflmM EGTA and AA.
These results show that the different tissues in theearly Xenopus embryo contain PKC with differentbiochemical properties, suggesting that these aredistinct PKC isozymes.
Discussion
The two major results from this paper are that wesubstantiate the notion that protein kinase C is involvedin mediating neural induction and show that thispossibly occurs via phospholipase C activation, and thatwe also show that the different tissues of the embryocontain PKC with different biochemical properties.
We showed previously that PKC is translocated fromthe cytosol to the membrane, specifically in the neural-induced ectoderm, in response to the neural-inducingsignals (Otte et al. 1988). In this paper, we report thatPKC is already strongly translocated at stage 11. We

468 A. P. Otte and others
found earlier that the levels of cAMP and adenylatecyclase (AC) increase from stage 12 onward (Otte et al.1989). We also found that the increase in AC activity isdependent on PKC activation, indicating cross-talkbetween these pathways (Otte et al. 1989). The resultshown here, that PKC is activated at an earlier stagethan the cAMP pathway, fits our model that PKC needsto be activated first, before the cAMP pathway can beactivated (Otte et al. 1989).
It is important to note that PKC translocation lasted,even after the involution of the neural inducingmesoderm was complete and the area which becomesneural tissue had been established (stage 13) (Nieuw-koop and Faber, 1967). This might mean that thistranslocated PKC is still active, but it might also bepresent in an inactive form. This last possibility isproposed in a recent model, which postulates a role forsustained PKC translocation in cellular memory (Alkonand Rasmussen, 1988). This quasi-permanently translo-cated PKC might become reactivated whenever theintracellular Ca concentration is raised.
Two of our present observations strengthen ourprevious suggestion that PKC is involved in mediatingneural induction:
(i) The neural-induced ectoderm contains a higherlevel of inositol phosphates, indicating phospholipase Cactivation. We have not tested though, whetherphospholipase C activation also leads to the productionof diacylglycerol and PKC activation in Xenopusectoderm. Since we find PKC activation at a similartime, this appears, however, to be a very likelypossibility.
(ii) The existence of a set of proteins which becomestrongly phosphorylated in induced neurectoderm, andwhich also become phosphorylated after PKC acti-vation by TPA, makes it likely that PKC activation isthe causal event in both of these situations. Whetherthese phosphorylated proteins are actual direct PKCsubstrates is difficult to determine. It is quite possiblethat PKC activation leads to the activation of otherkinases and that the phosphorylated proteins aresubstrates of these kinases. We can also not exclude thepossibility that natural neural induction is mediated bykinases other than PKC that phosphorylate the same setof proteins as those phosphorylated via TPA activatedPKC. We feel, however, that, considering the closesimilarity between the phosphorylation patterns inTPA-treated ectoderm and in ectomesoderm explantsand the observation that both TPA (Otte et al. 1988)and dorsal mesoderm induce PKC translocation (seeabove), these results at least indicate that PKC isinvolved in mediating natural neural induction.
One of these phosphorylated proteins resembles thewell-known 80 x 103 Mr PKC substrate (Rodriguez-Penaand Rozengurt, 1986; Niedel and Blackshear, 1988).We base this suggestion on the molecular weight of theprotein and on its highly acidic character (pi 4.3). Wefound further that some of these phosphorylatedproteins are present in the membranes of neural-induced ectoderm. The SOxlO3 MT substrate was es-pecially abundant. It was long thought that this
substrate needs to be present in the membrane in orderto become phosphorylated (Aderam etal. 1988; Stumpoet al. 1989; Wang et al. 1989), although recentexperiments now challenge this idea (Graff et al. 1989).In our case, however, some, but certainly not all of theproteins that are phosphorylated in vivo appear to bepresent in the membrane when they are phosphoryl-ated. Since we find these proteins in membranes after ashort (30min) TPA treatment, they are likely to beactual, direct PKC substrates. This leaves open thepossibility that the other, in vivo phosphorylatedproteins are substrates of kinases that are activated byPKC, or that they are PKC substrates that have notbeen translocated to the membrane.
The observation that the PKC types derived fromprospective ectoderm and from vegetal tissue, respect-ively, have different sensitivities to Ca2+ and arachi-donic acid suggest that these are different PKCisozymes. The different PKC isozymes exhibit distinctpatterns of tissue-specific expression (Kikkawa et al.1989; Hidaka et al. 1988; Huang et al. 1987; Brandt et al.1987), suggesting important, different, but as yetunknown, physiological functions. In a recent studyNoar and coworkers (1988) showed that PKC type I ory-PKC is highly dependent on Ca2+ for its activationand that it can be activated by 12-25 JIM arachidonicacid. It is therefore tempting to speculate that the PKCisolated from ectodermal cells represents the mam-malian type I PKC isozyme. This is all the moreinteresting' because this isozyme has been foundexclusively in neural tissue (Kikkawa et al. 1989), whichis derived from ectoderm. As long as there are nonucleotide sequence data available from Xenopus PKCgenes, this point remains speculative, however.
The presence of different PKC isozymes withdifferent biochemical properties might throw an inter-esting light on the various putative roles of PKC in earlyembryonic events like fertilization (Turner et al. 1984;Whitaker and Irvine, 1984; Busa et al. 1985; Dworkinand Dworkin-Rastl, 1989), the establishment of thedorsoventral polarity of the embryo (Kao et al. 1987;Busa and Gimlich, 1989; Berridge et al. 1989) andneural induction (Otte et al. 1988 and this presentwork). Since the establishment of the dorsoventralpolarity of the embryo occurs in the vegetal cells(Boterenbrood and Nieuwkoop, 1973; Gimlich andGerhart, 1984) and neural induction in the ectodermalcells, it is tempting to speculate that different PKCisozymes function in these two distinct, importantembryological events. More detailed studies will,however, be needed to substantiate this idea.
We thank Drs R. Van Driel and S. de Laat for criticallyreading this manuscript.
References
ADEREM, A. A., ALBERT, K. A., KEUM, M. H., WANG, J. K. T.,GREENGARD, P. AND COHN, Z. A. (1988). Stimulus-dependentmyristoylation of a major substrate for protein kinase C. Nature332, 362-364.

Protein kinase C in early Xenopus embryogenesis 469
ALKON, D. L. AND RASMUSSEN, H. (1988). A spatial-temporalmodel of cell activation. Science 239, 998-1005.
AUGERT, G., BLACKMORE, P. F. AND EXTON, J. H. (1989). Changes
in the concentration and fatty acid composition ofphosphoinositides induced by hormones in hepatocytes. J. biol.Chem. 264, 2574-2580.
BARTFAJ, T. (1979). Preparation of metal-chelate complexes andthe design of steady-state kinetic experiments involving metalnucleotide complexes. Adv. Cyclic Nucleotide Res. 10, 219-241.
BEMENT, W. M. AND CAPCO, D. G. (1989). Activators of proteinkinase C trigger cortical granule exocytosis, cortical contraction,and cleavage furrow formation in Xenopus laevis oocytes andeggs. J. Cell Biol. 108, 885-892.
BERRIDGE, M. J. (1987). Inositol triphosphate and diacylglycerol:two interacting second messengers. A. Rev. Biochem. 56,159-193.
BERRIDGE, M. J., DOWNES, C. P. AND HANLEY, M. R. (1989).
Neural and developmental actions of lithium: a unifyinghypothesis. Cell 59, 411-419.
BERRIDGE, M. J. AND IRVINE, R. F. (1989). Inositol phosphates andcell signalling. Nature 341, 197-205.
BOTERENBROOD, E. C. AND NIEUWKOOP, P. D. (1973). The
formation of the mesoderm in urodelean amphibians. WilhelmRoux' Arch. EntwMech. 173, 319-332.
BRAMMER, M. J., HAJIMOHAMMADREZA, I., SARDIWAL, S. AND
WEAVER, K. (1988). Is inositol bisphosphate the product ofA23187 and carbachol-mediated polyphosphoinositidebreakdown in synaptosomes? J. Neurochem. 51, 514-521.
BRANDT, S. J., NIEDEL, J. E., BELL, R. M. AND SCOTT YOUNG, W.
(1987). Distinct patterns of expression of different protein kinaseC mRNAs in rat tissues. Cell 49, 57-63.
BUSA, W. B., FERGUSON, J. E., JOSEPH, S. K., WILLIAMSON, J. R.
AND NUCCTTELLI, R. (1985). Activation of frog (Xenopus laevis)eggs by inositol triphosphate. I. Characterization of Ca2+ releasefrom intracellular stores. J. Cell Biol. 101, 677-682.
BUSA, W. B. AND GIMUCH, R. L. (1989). Lithium-inducedteratogenesis in frog embryo prevented by apolyphosphoinositide cycle intermediate or a diacylglycerolanalog. Devi Biol. 132, 315-324.
CASTAGNA, M., TAKAI, Y., KAIBUCHI, K., SANO, K., KIKKAWA, V.
AND NISHIZUKA, Y. (1982). Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J. biol. Chem. 257, 7847-7851.
CUTHBERTSON, K. S. AND COBBOLD, P. H. (1985). Phorbol esterand sperm activate mouse oocytes by inducing sustainedoscillations in cell Ca2 + . Nature 316, 541-542.
DWORKIN, M. B. AND DWORHN-RASTL, E. (1989). Metabolicregulation during early frog development: flow of glycolyticcarbon into phospholipids in Xenopus oocytes and fertilizedeggs. Devi Biol. 132, 524-528.
GIMLICH, R. L. AND GERHART, J. C. (1984). Early cellularinteractions promote embryonic axis formation in Xenopuslaevis. Devi Biol. 104, 117-130.
GRAFF, J. M., GORDON, J. I. AND BLACKSHEAR, P. J. (1989).
Myristoylated and nonmyristoylated forms of a protein arephosphorylated by protein kinase C. Science 246, 503-506.
HIDAKA, H., TANAKA, T., ONODA, K., HIGIWARA, M., WATANABE,
M., OHTA, H., ITO, Y., TSURUDOME, M. AND YOSHIDA, T.
(1988). Celltype-specific expression of protein kinase C isozymesin the rabbit cerebellum. / . biol. Chem. 263, 4523-4526.
HOUSE, C. AND KEMP, B. E. (1987). Protein kinase C contains apseudosubstrate prototype in its regulatory domain. Science 238,1726-1728.
HOUSE, C , WETTENHALL, R. E. H. AND KEMP, B. E. (1987). The
influence of basic residues on the substrate specificity of proteinkinase C. J. biol. Chem. 262, 772-777.
HUANG, F. L., YOSHIDA, Y., NAKABAYASHI, H. AND HUANG, K.-P.
(1987). Differential distribution of protein kinase C isozymes inthe various regions of brain. J. biol. Chem. 262, 15714-15720.
JONES, E. A. AND WOODLAND, H. R. (1989). Spatial effects ofneural induction in Xenopus laevis. Development 107, 785-791.
KAMATA, T. AND KUNG, H.-F. (1990). Modulation and ribosomalS6 phosphorylation in Xenopus oocytes by microinjection of
oncogenic ras and protein kinase C. Mol. cell. Biol. 10,880-886.
KAO, K. R., MASUI, Y. AND ELINSON, R. P. (1986). Lithium-induced respecification of pattern in Xenopus laevis embryos.Nature 322, 371-373.
KIKKAWA, U., KISHIMOTO, A. AND NISHIZUKA, Y. (1989). The
protein kinase C family: heterogeneity and its implications.A. Rev. Biochem. 58, 31-44.
KIKKAWA, U. AND NISHIZUKA, Y. (1986). The role of proteinkinase C in transmembrane signalling. A. Rev. Cell Biol. 2,149-178.
KINTNER, C. R. AND MELTON, D. A. (1987). Expression ofXenopus N-CAM RNA in ectoderm is an early resporise toneural induction. Development 99, 311-325.
LACAL, J. C , D E LA PENA, P., MOSCAT, J., GARCIA-BARRENO, P.,
ANDERSON, P. S. AND AARONSON, S. A. (1987). Rapidstimulation of diacylglycerol production in Xenopus oocytes bymicroinjection of H-ras p21. Science 238, 533-536.
MASUI, Y. (1960). Effects of lithium chloride upon differentiationof the neural-plate ectoderm of head area. Memoirs of theKonan University 4, 103-114.
NAOR, Z., SHEARMAN, M. S., KISHIMOTO, A. AND NISHIZUHA, Y.
(1988). Calcium-independent activation of hypothalamic type Iprotein kinase C by unsaturated fatty acids. Endocrinology 2,1043-1048.
NEWPORT, J. AND KKSCHNER, M. W. (1982). A majordevelopmental transition in early Xenopus embryos. I.Characterization and timing of cellular changes at themidblastula stage. Cell 30, 675-686.
NIEDEL, J. E. AND BLACKSHEAR, P. J. (1986). Protein kinase C. InPhosphoinositides and Receptor Mechanisms (ed. Putney, J. W.)pp. 47-88 (New York: Allen R Liss).
NIEUWKOOP, P. D. AND FABER, J. (1967). The Normal Table ofXenopus laevis (Daudin). Second edition. Amsterdam: NorthHolland Publishing Co.
NIEUWKOOP, P. D., JOHNEN, A. G. AND ALBERS, B. (1985). The
Epigenetic Nature of Early Chordate Development. InductiveInteraction and Competence. Cambridge University Press.Cambridge, England.
NISHIZUHA, Y. (1984). The role of protein kinase C in cell surfacesignal transduction and tumour promotion. Nature 308, 693-698.
NISHIZUKA, Y. (1986). Studies and perspectives of protein kinaseC. Science 233, 305-312.
NISHIZUKA, Y. (1988). The molecular heterogeneity of proteinkinase C and its implications for cellular regulation. Nature 334,661-665.
O'FARRELL, P. H. (1975). High resolution two-dimensionalelectrophoresis of proteins. J. biol. Chem. 250, 4007-4021.
OTTE, A. P., KOSTER, C. H., SNOEK, G. T. AND DURSTON, A. J.
(1988). Protein kinase C mediates neural induction in Xenopuslaevis. Nature 334, 618-620.
OTTE, A. P., VAN RUN, P., HEIDEVELD, M., VAN DRIEL, R. AND
DURSTON, A. J. (1989). Neural induction is mediated by cross-talk between the protein kinase C and cyclic AMP pathways.Cell 58, 641-648.
RODRIGUEZ-PENA, A. AND ROZENGURT, E. (1986). Phosphorylationof an acidic mol. wt. 80.000 cellular protein in a cell-free systemand intact Swiss 3T3 cells: a specific marker of protein kinase Cactivity. EMBO J. 5, 77-83.
SMITH, L. D. (1989). The induction of oocyte maturation:transmembrane signaling events and regulation of the cell cycle.Development 107, 685-699.
STTTH, B. J. AND MALLER, J. L. (1987). Induction of meioticmaturation in Xenopus oocytes by 12-O-tetradecanoylphorbol-13-acetate. Expl Cell Res. 169, 514-523.
STUMPO, D. J., GRAFF, J. M., ALBERT, K. A., GREENGARD, P. AND
BLACKSHEAR, P. J. (1989). Molecular cloning, characterization,and expression of a cDNA encoding the '80-to-87-kDa'myristoylated alanine-rich C kinase substrate: a major cellularsubstrate for protein kinase C. Proc. natn. Acad. Sci. U.S.A. 86,4012^1016.
SWANN, K. AND WHITAKER, M. (1985). Stimulation of the Na/Hexchanger of sea urchin eggs by phorbol ester. Nature 314,274-277.

470 A. P. Otte and others
TILLY, B. C, VAN PARIDON, P. A., VERLAAN, I., Wnrrz, K. W.A., DE LAAT, S. W. AND MOOLENAAR, W. H. (1987). Inositolphosphate metabolism in bradykinin-stimulated human A431carcinoma cells. Biochem. J. 244, 129-135.
TURNER, P. R., SHEETZ, M. P. AND JAFFE, L. A. (1984).Fertilization increases the polyphosphoinositide content of seaurchin eggs. Nature 310, 414-415.
VAN HAASTERT, P. J. M., DE VRIES, M. J., PENNING, L. C ,ROOVERS, E., VAN DER KAAY, J., ERNEUX, C. AND VANLOOKEREN CAMPAGNE, M. M. (1989). Chemoattractant andguanosine 5'-[3-thio]triphosphate induce the accumulation ofinositol 1,4,5-triphosphate in Dictyostelium cells that are labelledwith [3H]inositol by electroporation. Biochem. J. 258, 577-586.
WANG, J. K. T., WALAAS, S. I., SIHRA, T. S., ADEREM, A. ANDGREENGARD, P. (1989). Phosphorylation and associatedtranslocation of the 87-kDa protein, a major protein kinase C,in isolated nerve terminals. Proc. natn. Acad. Sci. U.S.A. 86,2253-2256,
WHTTAKER, M. AND IRVINE, R. F. (1984). Inositol 1,4,5-triphosphate microinjection activates sea urchin eggs. Nature312, 636-639.
ZIMMERMAN, I. D. (1985). Induced symmetry breaking in the eggof the frog Rana pipiens. Differentiation 30, 98-101.
(Accepted 26 June 1990)