characterization of proteinase inhibitor-ii gene under
TRANSCRIPT

494
http://journals.tubitak.gov.tr/biology/
Turkish Journal of Biology Turk J Biol(2017) 41: 494-502© TÜBİTAKdoi:10.3906/biy-1609-42
Characterization of proteinase inhibitor-II gene under OsRGLP2promoter for salt stress in transgenic Nicotiana benthamiana
Shazia REHMAN1, Bodil JØRGENSEN2, Søren Kjærsgaard RASMUSSEN2, Tariq MAHMOOD1,*1Department of Plant Sciences, Quaid-i-Azam University, Islamabad, Pakistan
2Department of Plant and Environmental Sciences, Faculty of Sciences, University of Copenhagen, Frederiksberg, Denmark
* Correspondence: [email protected]
1. IntroductionIn natural environments, plants are confronted with multiple stresses including both biotic and abiotic stresses. Under such conditions, plants have evolved sophisticated mechanisms for upregulation of genes that can eliminate or reduce the effects of these stresses. Plant proteinase inhibitors (PIs) are among the promising defensive proteins that help plants resist different kinds of environmental stresses (biotic and abiotic) (Ryan, 1990; Fujita et al., 2006; Srinivasan et al., 2009). In plants, PIs inhibit the proteolytic activity of their target proteases under stress conditions, and thus the stability of vital cellular proteins and enzymes increases, leading to enhanced stress tolerance. Plant hormones like jasmonic acid (JA) and its derivatives, and ethylene (ET) and salicylic acid (SA), are particularly known to be involved in upregulation of PIs as a part of biotic stress responses (Pearce et al., 1993; Fujita et al., 2006), while abscisic acid (ABA) is induced during abiotic stress conditions. However, all these stress hormones work synergistically or antagonistically with each other, suggesting cross-talk between biotic and abiotic stress responses (Fujita et al., 2006; Cao et al., 2011).
PIs have been categorized into four classes based on their catalytic mechanisms or by the type of protease they
inhibit, i.e. serine, metallo, cysteine, and aspartyl PIs. Serine proteinase inhibitors appear to be the largest family, which has been extensively studied and characterized from various plant sources (Mello et al., 2001; Haq and Khan, 2003). They are most abundant in storage organs (seeds and tubers) and also occur in other vegetative tissues in low concentration.
Proteinase inhibitor II (PI-II) proteins are members of the serine proteinase inhibitors that have been isolated and characterized from several members of the family Solanaceae including Solanum lycopersicum (tomato) (Gustafson and Ryan, 1976), Solanum tuberosum (potato) (Bryant et al., 1976), Solanum melongena (eggplant) (Richardson, 1979), Nicotiana tabacum (tobacco) (Pearce et al., 1993), Solanum americanum (nightshade) (Xu et al., 2001; Sin and Chye, 2004), and Capsicum annuum (pepper) (Mishra et al., 2012). Endogenously, serine PIs have been associated in modulating programmed cell death events necessary for normal development and cellular defense responses (Solomon et al., 1999; Park et al., 2001). In recent years, expression of proteinase inhibitors in response to multiple stresses has been studied by various groups. PI genes from chestnut, rice, and barley were upregulated by salt and drought stresses (Gaddour et al.,
Abstract: Proteinase inhibitors in plants are mainly involved in conferring resistance against herbivores and pathogens. However, their role in abiotic stress resistance has also been reported. Therefore, transgenic tobacco plants were generated constitutively expressing the tomato proteinase inhibitor-II (PI-II) under control of the rice root germin-like protein 2 (OsRGLP2) promoter using Agrobacterium tumefaciens. T1 transgenic seedlings were subjected to stress treatments with different concentrations of sodium chloride (NaCl) (100 mM, 200 mM, 300 mM). Quantitative real-time PCR revealed considerable levels of expression and upregulation of PI-II in transgenic lines under salt stress. Considering the phenotypic effects, the transgenic seedlings showed better growth, higher chlorophyll content, higher proline contents, and higher average fresh weight when compared to the wild type. This comparative analysis of transgenic and wild-type seedlings allows us to suggest that PI-II gene activity can be upregulated via the OsRGLP2 promoter under osmotic stress conditions.
Key words: Expression, osmotic stress, phenotypic effects, proteinase inhibitor-II, sodium chloride
Received: 19.09.2016 Accepted/Published Online: 12.01.2017 Final Version: 14.06.2017
Research Article

REHMAN et al. / Turk J Biol
495
2001; Huang et al., 2007). Proteinase inhibitors belonging to the Kunitz family were upregulated in NaCl-treated radish and drought-stressed Arabidopsis thaliana (Lopez et al., 1994; Gosti et al., 1995). Similarly, Dombrowski (2003) observed that salt stress causes the activation of wound-inducible genes and accumulation of serine PI (II) in tomato plants. In rice, OCPI1-overexpressing transgenic plants showed improved resistance against drought (Huang et al., 2007). Previously, a trypsin PI gene from tobacco (NtPI) was constitutively expressed under the CAM35S promoter to confer tolerance under multiple stress conditions like drought, salinity, and pH (Srinivasin et al., 2009). Many stress-related genes have been expressed in transgenic plants using either constitutive or stress-inducible promoters. The OsRGLP2 promoter (plant-derived wound-inducible promoter) is a robust promoter capable of driving the expression of downstream genes under certain stress conditions, including wounding, salt, dehydration, and pathogenic infection (Mahmood et al., 2013; Munir et al., 2015). Therefore, expression of the PI-II gene under the OsRGLP2 promoter was investigated in salt-stressed transgenic Nicotiana benthamiana. To our best knowledge, this is the first report showing expression of the tomato PI-II gene under the OsRGLP2 promoter conferring tolerance against salt stress.
2. Materials and methods2.1. Plant material and transformation vectorThe tobacco variety Nicotiana benthamiana was selected as experimental material for Agrobacterium-mediated transformation. Greenhouse-grown tobacco plants were kindly provided by the Department of Plant and Environmental Sciences, Copenhagen University, Denmark. The PCAMBIA1391Z vector having the Solanum lycopersicum proteinase inhibitor-II gene (Accession No. AY007240) fused with the GUS under the control of the OsRGLP2 promoter (Mahmood et al., 2013; Figure 1) in Agrobacterium strain EHA101 was used to produce transgenic plants using the leaf disk method (Horsch et al., 1985).
2.2. Agrobacterium-mediated transformation and confirmation of integrationThe transformation of tobacco plants with Agrobacterium was performed following the procedure of Horsch et al. (1985) and selection of transformants was done using 50 mg/L hygromycin. The integration of the transgene in putative transgenic N. benthamiana was confirmed by PCR. Total genomic DNA was extracted from leaves of transgenic plants using the DNeasy Plant Mini Kit (QIAGEN). PCR of transgenic plants were carried out with PI-II and hygromycin-resistant gene primers. The sequences of these primers are PI-IIF: 5’TATCCATCATGGCTGTCCAC3’, PI-IIR: 5’AACACACAACTTGATCCCCACA3’, and HygroF: 5’GCTCCATACAAGCCAACCAC3’, HygroR: 5’CGAAAAGTTCGACAGCGTCTC3’. For amplification, 25 µL of amplification reaction was prepared containing 45 ng/µL genomic DNA, 2.5 µL of 10X PCR buffer, 1 µL (25 pmol) of forward and reverse primer, 1.5 µL (25 mM) of MgCl2, 1.5 µL (2.0 mM) of dNTPs, and 1.5 U of Taq polymerase. The PCR conditions for amplification were as follows: preamplification denaturation at 94 °C for 5 min; then 35 cycles of denaturing at 94 °C for 40 s, 55 °C for 40 s, and 72 °C for 45 s; and a final extension step at 72 °C for 20 min. 2.3. Salt stress assays Seeds from T0 transgenic lines (1, 2, and 3) and the wild type (WT ) were surface-sterilized with 3 % (v/v) sodium hypochlorite for 5 min, followed by washing 3–5 times with autoclaved distilled water. T1 transgenic seedlings were selected on the basis of hygromycin resistance. Stress tolerance of transgenic plants was evaluated at the seedling stage. For this, 10-day-old seedlings of WT and transgenic lines (lines 1, 2, and 3) were transferred to MS-agar media with different concentrations of NaCl (100 mM, 200 mM, and 300 mM NaCl). After 14 days of stress treatment, salt-stressed assays were performed, which included analysis of seedling growth response, measurement of average fresh weight, and estimation of chlorophyll contents and proline contents.
Figure 1. A schematic representation of the pCAMBIA1391Z_OsRGLP2 construct. The proteinase inhibitor gene (PI-II) was cloned in pCAMBIA1391Z_OsRGLP2 at the EcoR1 restriction site.

REHMAN et al. / Turk J Biol
496
2.4. Estimation of chlorophyll content and average fresh weightIn order to define the salt tolerance or sensitivity of seedlings, fresh weight and chlorophyll contents of salt-stressed seedlings were examined at 14 days of stress treatment in the presence of various NaCl levels. For estimation of chlorophyll content, about 0.2 g of NaCl-treated seedlings of the transgenic and WT lines were used. Chlorophyll was extracted from fresh material with 80% acetone. Total chlorophyll content was then estimated spectrophotometrically by measuring the optical density at 645 and 663 nm according to the method described by Arnon (1949) using the following formula, and results were expressed as µg g–1 of fresh tissue:Total chlorophyll = [{(20.2 × A645) + (8.02 × A663)}/1000 × W] × V, where V = extracted volume (mL) and W = fresh weight of the material (g).2.5. Analysis of proline contentsProline contents of transgenic and WT tobacco seedlings were measured spectrophotometrically according to Bates et al. (1973). For this purpose, plant tissues (0.3 g) were homogenized with 3% (w/v) sulfosalicylic acid and then centrifuged at 1000 rpm for 5 min. The supernatant (2 mL) was incubated for 1 h at 100 °C after adding 2 mL of acid-ninhydrin and 2 mL of glacial acetic acid. After cooling, the mixture was extracted with 2 mL of toluene and again centrifuged at 1000 rpm for 5 min. The upper aqueous phase (containing proline) was collected and its absorbance was read at 520 nm using toluene as a blank. Proline concentration was quantified by referring to a standard curve of proline.2.6. RNA extraction and cDNA synthesisFor expression analysis, 15-day-old NaCl-treated seedlings of WT and transgenic lines at different concentrations were collected after 24 h and ground with a mortar and pestle in the presence of liquid nitrogen. Extraction of total RNA was carried out using the TRIzol reagent (Invitrogen). RNA samples were purified from contaminated DNA after Turbo DNA-free DNase treatment. The concentration of purified RNA was measured with a NanoDrop 2000 (Thermo Scientific). The RNA quality and integrity were determined by 2% agarose gel electrophoresis. From 1 µg of RNA, first-strand cDNA was synthesized using oligo (dT) primers and M-MuLV reverse transcriptase in a total volume of 20 µL. The reaction mixture was incubated for 1 h at 42 °C followed by 70 °C for 10 min to stop the reaction. Finally, the cDNA was stored at –20 °C until further use.2.7. qPCR Quantitative real-time PCR (qPCR) was performed in 96-well reaction plates using the Stratagene Mx3005P QPCR System. The PCR mixture was composed of 4 µL of cDNA (1:10 dilution), 0.6 µL of each forward and
reverse primer (5 µM), 4 µL of 1X EvaGreen master mix, and 10.8 µL of nuclease-free water. RT-PCR reaction was performed using the following gene specific primers: 5’TTCGGGATATGCCCACGTTC3’ (forward) and 5’AGGTGCAAGCATTTGGCCTT3’ (reverse). As an internal control, N. benthamiana actin primers 5’GATGAAGATACTCACAGAAAGA3’ (forward) and 5’GTGGTTTCATGAATGCCAGCA3’ (reverse) were used. The PCR program was as follows: predenaturing at 95 °C for 10 min, and 40 cycles of denaturing at 95 °C for 30 s, annealing at 56 °C for 1 min, and extension at 72 °C for 10 s. The relative expression level was calculated according to the formula 2–ΔΔCt.2.8. Statistical analysisAll assays were repeated at least three times. The PROC GLM procedure was used to study the analysis of variance (ANOVA) using SAS 9.4. The means within each treatment were compared using the least significant difference (LSD) test with a threshold probability of P < 0.01.
3. Results and discussion3.1. Transformation and regeneration of transgenicsUsing Agrobacterium-mediated transformation, the PI-II gene was introduced into N. benthamiana. After cocultivation the putative transformed leaf disks developed calli from the cut edges of the explants after 10–15 days on hygromycin selection media. Adventitious shoots started to emerge after approximately 2–4 weeks. Regenerated shoots were rooted on MS media and later transferred to soil and maintained in a greenhouse to obtain seeds. These T0 plants were then self-pollinated to harvest T1 transgenic N. benthamiana seeds.3.2. Molecular analysis To verify the insertion and expression of the transgene in the putative transgenic plants, PCR analysis was performed with gene-specific and resistant gene primers. Figh-quality PCR products of approximately 684 bp in size were obtained using PI-IIF and PI-IIR primer sets (Figure 2). Further confirmation was done with HygF and HygR primers that resulted in the amplification of a fragment of about 700 bp (Figure 3). Later the amplified products were run on 1% agarose gel. A total of 8 transgenic lines (T0) were identified through PCR amplification, of which three independent transgenic lines (TL1, TL2, and TL3) were randomly selected for further analysis. To study the effect of salt stress, the transcript levels of this gene were measured through qPCR after exposing the 15-day-old putative transgenic T1 seedlings (TL1, TL2, and TL3) and WT to NaCl stress (100 mM, 200 mM, and 300 mM) for 24 h. As shown in Figure 4, a significant difference in transgene expression (P < 0.01) was noticed among the different transgenic lines and WT at all NaCl

REHMAN et al. / Turk J Biol
497
concentrations. With a low concentration of NaCl (100 mM), the relative expression level of all transgenic lines was comparatively higher (7.4, 5.6, and 5.1 respectively) than that of the WT. Among the different transgenic lines tested, TL1 showed significantly higher expression under different salt stress conditions. Furthermore, salt stress also induced the expression of the PI-II gene at a higher concentration in transgenic lines but the induced levels were lower than at 100 mM NaCl. These results indicated that overexpression of the PI-II gene under the OsRGLP2 promoter leads to a significant increase in salt tolerance in transgenic plants at different levels of salinity stress. In a similar study, the transcript level of a trypsin inhibitor gene of Nicotiana benthamiana (NtPI) was found to increase at elevated salt concentrations in three transgenic lines when constitutively expressed under the transcriptional control of the CaMV35S promoter. Recently, Li et al. (2015) reported that overexpression of a cysteine proteinase inhibitor from Jatropha curcas (JcCPI) using the 2XCaMV35S promoter increased the transcript level of the transgene in three transgenic tobacco lines
Figure 2. PCR amplification of the PI-II gene from transgenic plants. L1 and L2: 1-kb plus ladder, lanes 1 to 8: amplified products of PI-II gene, lane 9: negative control (WT), lane 10: positive control.
Figure 3. PCR amplification of hygromycin gene from transgenic plants. L1 and L2: 1-kb plus ladder, lanes 1 to 8: amplified products of hygromycin gene, lane 9: negative control (WT), lane 10: positive control.
Figure 4. Expression profile of PI-II in transgenic lines under salt stress. The data presented are the means ± SE of three replicates (n = 3). Letters on each bar within each treatment indicate significant differences at P < 0.01.

REHMAN et al. / Turk J Biol
498
when compared to the WT under salt stress conditions. Previously, Kim et al. (2001) observed elevated transcript levels of a serine proteinase inhibitor (CaPI-2) in red pepper under salt stress. Similarly, a Bowman–Birk type proteinase inhibitor (BBI) was shown to be responsible for increasing salt tolerance in wheat (Shan et al., 2008). Moreover, overexpression of rice OCPI2 was found to enhance the salt tolerance in transgenic Arabidopsis under the CAM35S promoter (Tiwari et al., 2015). Our results are consistent with the above reports, in which expression of the PI gene was shown to be upregulated under a constitutive promoter in salt-stressed conditions. It thus appears that the PI-II gene is responsive to osmotic stress and might play a pivotal role in abiotic stresses under the control of the OsRGLP2 promoter.
Although previous studies revealed the functions of proteinase inhibitors under constitutive promoters for providing resistance against insects and pathogens, there are few studies about the proteinase inhibitors in conferring resistance against abiotic stresses including NaCl stress. Many abiotic stress conditions are involved in the induction of PI genes and plants respond to these stresses through a network of signaling pathways like ABA, JA, and ET (Pena-Cortes et al., 1995; Fujita et al., 2006). Exposure to UV radiation (Conconi et al., 1996), salt stress (Dombrowski et al., 2003; Huang et al., 2007), and drought stress (Fujita et al., 2004) has resulted in the JA-mediated induction of proteinase inhibitors. It is explained that, under these conditions, PI genes are activated and JA is known to be a potent inducer of PIs. The OsRGLP2 promoter contains several putative stress-responsive regulatory regions (Mahmood et al., 2013; Munir et al., 2015), which might play significant roles in
upregulating the stress-responsive genes including the PI-II genes. 3.3. Response of T1 progenies to salt stressSalt stress induces several biochemical and physiological responses in plants depending on the severity and duration of stress (Nemoto and Sasakuma, 2002; Megdiche et al., 2007). Growth inhibition is a common response to salinity and many studies confirmed the inhibitory effects of salinity on overall plant growth and productivity (Parida and Das, 2005; Voigt et al. 2009). In the present report, the phenotypic differences were prominent between transgenic lines and the WT when grown under salt stress conditions. The growth response of PI-II-expressing transgenics was better than that of the WT at the lower salt concentration (100 mM NaCl). Unlike WT seedlings, the majority of the transgenic seedlings survived higher salinity stress (200 and 300 mM NaCl) (Figure 5). However, transgenics at lower concentrations showed earlier growth than at higher concentrations. A similar phenotypic response was noted in transgenic plants overexpressing the rice chymotrypsin proteinase inhibitor (OCPI2) (Tiwari et al., 2015) and a tobacco PI (NtPI) (Srinivasan et al., 2009).
Chlorophyll content is considered as one of the few physiological parameters that can show good correlation with salinity tolerance (Srivastava et al., 1988; Ali et al., 2004). Salt stress-induced damage was reflected by the degree of loss of chlorophyll contents under elevated salinity conditions in transgenics and control seedlings (Figure 6). There was a clear difference in chlorophyll content after subjecting the seedlings to different concentrations of NaCl. With respect to the WT, the effect of NaCl on chlorophyll content was lower on transgenic plants and differed significantly between transgenics and
Figure 5. Phenotypic response of transgenic seedlings and the WT under salt stress: A) Seedling growth of transgenic lines and WT at 100 mM NaCl. B) Seedling growth of transgenic lines and WT at 200 mM NaCl. C) Seedling growth of transgenic lines and WT at 300 mM NaCl.

REHMAN et al. / Turk J Biol
499
the WT under different salt concentrations (P < 0.01; Figure 6), thus documenting the better metabolic status of transgenic lines over the nontransgenic control under salt stress. However, significant variation for total chlorophyll was observed among the three independent transgenic plants and the WT at different levels of salinity stress. At 100 mM NaCl concentration, TL1 retained the highest chlorophyll content compared to other transgenic lines and WT plants (Figure 6). Moreover, no significant difference in total chlorophyll content was observed between transgenic lines (TL2 and TL3) at 200 mM NaCl. Similarly, in response to 300 mM NaCl, TL1 and TL2 did not show any significant difference for total chlorophyll contents. Additionally, at higher NaCl concentrations (200 and 300 mM), chlorophyll reduction appeared in both WT and transgenic seedlings. However, the seedlings from the WT presented higher chlorophyll reduction at concentrations higher than 100 mM NaCl, while transgenics survived the stress and remained green for longer periods at elevated salt stress. Overall, the total chlorophyll reduction of the WT was 84% after increasing the NaCl concentration from 100 to 300 mM. However, TL1, TL2, and TL3 displayed 63%, 73%, and 77% reduction in chlorophyll content, respectively (Figure 6). In a recent study, Yadav et al. (2016) developed transgenic tobacco plants overexpressing the Arabidopsis annexin gene (AnnAt8) and demonstrated higher chlorophyll retention in transgenic plants than the WT after treatment with 300 mM and 400 mM NaCl. Tan et al. (2015) also demonstrated higher chlorophyll content in transgenic Arabidopsis plants overexpressing a novel cystatin gene from Malus prunifolia (MpCYS5) when compared with the WT in the presence of 200 mM NaCl.
In a related study, similar increased chlorophyll retention was observed in transgenic plants expressing the stress-responsive gene VvRD22 with respect to the WT upon exposure to 400 mM NaCl (Jardak-Jamoussi et al., 2014).
Regarding the fresh weight, increased salt tolerance was manifested by comparing the total fresh weight of transgenics along with the WT at elevated NaCl concentrations. After salt treatment, the difference in average fresh weight of transgenics versus WT was significant for all NaCl concentrations tested (P < 0.01; Figure 7), indicating better growth of transgenic lines than WT plants under salt stress conditions. In the presence of 100 mM NaCl, no notable difference in average fresh weight was seen between TL1 and TL2. Additionally, the same nonsignificant difference for average fresh weight was observed for TL2 and TL3 at the 300 mM NaCl concentration. In the presence of 200 and 300 mM NaCl, average fresh weight decreased gradually with increasing NaCl concentration. However, this reduction in fresh weight in transgenic lines was comparatively lower than in the WT plants (Figure 7). Furthermore, an average decline of 48%, 50%, 44%, and 61% in fresh weight was registered in TL1, TL2, TL3, and WT plants, respectively, with an increase in NaCl concentrations (100 to 300 mM NaCl). Collectively, these results indicated that transgenic plants constitutively expressing the PI-II gene under the OsRGLP2 promoter were more resistant to osmotic stress. Srinivasan et al. (2009) reported similar results, indicating that transgenic lines overexpressing NtPI exhibited higher fresh weight than nontransgenic plants under osmotic stress. Likewise, Zhang et al. (2008) observed better
Figure 6. Chlorophyll content of WT and transgenic lines under salt stress after 14 days of stress treatment. The data are the means ± SE of three replicates (n = 3). Letters on each bar within each treatment indicate significant differences at P < 0.01, and bars sharing a common letter are not significantly different.
Figure 7. Average fresh weight of WT and transgenic lines under salt stress after 14 days of stress treatment. The data are the means ± SE of three replicates (n = 3). Letters on each bar within each treatment indicate significant differences at P < 0.01, and bars sharing a common letter are not significantly different.
REHMAN et al. / Turk J Biol
500
growth and greater fresh weight in transgenic Arabidopsis overexpressed with two cystatins, AtCYSa and AtCYSb, when compared with the WT. These findings have strong correlations with previous studies under salt stress conditions (Dombrowski, 2003; Huang et al., 2007).
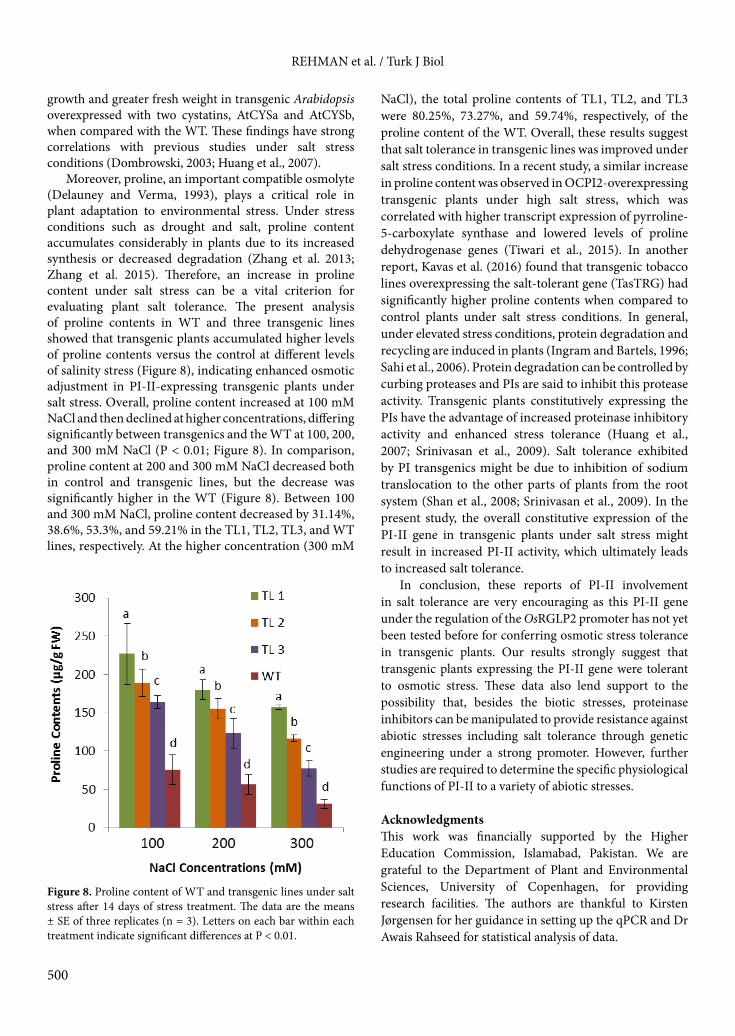
Moreover, proline, an important compatible osmolyte (Delauney and Verma, 1993), plays a critical role in plant adaptation to environmental stress. Under stress conditions such as drought and salt, proline content accumulates considerably in plants due to its increased synthesis or decreased degradation (Zhang et al. 2013; Zhang et al. 2015). Therefore, an increase in proline content under salt stress can be a vital criterion for evaluating plant salt tolerance. The present analysis of proline contents in WT and three transgenic lines showed that transgenic plants accumulated higher levels of proline contents versus the control at different levels of salinity stress (Figure 8), indicating enhanced osmotic adjustment in PI-II-expressing transgenic plants under salt stress. Overall, proline content increased at 100 mM NaCl and then declined at higher concentrations, differing significantly between transgenics and the WT at 100, 200, and 300 mM NaCl (P < 0.01; Figure 8). In comparison, proline content at 200 and 300 mM NaCl decreased both in control and transgenic lines, but the decrease was significantly higher in the WT (Figure 8). Between 100 and 300 mM NaCl, proline content decreased by 31.14%, 38.6%, 53.3%, and 59.21% in the TL1, TL2, TL3, and WT lines, respectively. At the higher concentration (300 mM
NaCl), the total proline contents of TL1, TL2, and TL3 were 80.25%, 73.27%, and 59.74%, respectively, of the proline content of the WT. Overall, these results suggest that salt tolerance in transgenic lines was improved under salt stress conditions. In a recent study, a similar increase in proline content was observed in OCPI2-overexpressing transgenic plants under high salt stress, which was correlated with higher transcript expression of pyrroline-5-carboxylate synthase and lowered levels of proline dehydrogenase genes (Tiwari et al., 2015). In another report, Kavas et al. (2016) found that transgenic tobacco lines overexpressing the salt-tolerant gene (TasTRG) had significantly higher proline contents when compared to control plants under salt stress conditions. In general, under elevated stress conditions, protein degradation and recycling are induced in plants (Ingram and Bartels, 1996; Sahi et al., 2006). Protein degradation can be controlled by curbing proteases and PIs are said to inhibit this protease activity. Transgenic plants constitutively expressing the PIs have the advantage of increased proteinase inhibitory activity and enhanced stress tolerance (Huang et al., 2007; Srinivasan et al., 2009). Salt tolerance exhibited by PI transgenics might be due to inhibition of sodium translocation to the other parts of plants from the root system (Shan et al., 2008; Srinivasan et al., 2009). In the present study, the overall constitutive expression of the PI-II gene in transgenic plants under salt stress might result in increased PI-II activity, which ultimately leads to increased salt tolerance.
In conclusion, these reports of PI-II involvement in salt tolerance are very encouraging as this PI-II gene under the regulation of the OsRGLP2 promoter has not yet been tested before for conferring osmotic stress tolerance in transgenic plants. Our results strongly suggest that transgenic plants expressing the PI-II gene were tolerant to osmotic stress. These data also lend support to the possibility that, besides the biotic stresses, proteinase inhibitors can be manipulated to provide resistance against abiotic stresses including salt tolerance through genetic engineering under a strong promoter. However, further studies are required to determine the specific physiological functions of PI-II to a variety of abiotic stresses.
Acknowledgments This work was financially supported by the Higher Education Commission, Islamabad, Pakistan. We are grateful to the Department of Plant and Environmental Sciences, University of Copenhagen, for providing research facilities. The authors are thankful to Kirsten Jørgensen for her guidance in setting up the qPCR and Dr Awais Rahseed for statistical analysis of data.
Figure 8. Proline content of WT and transgenic lines under salt stress after 14 days of stress treatment. The data are the means ± SE of three replicates (n = 3). Letters on each bar within each treatment indicate significant differences at P < 0.01.

REHMAN et al. / Turk J Biol
501
References
Ali Y, Aslam Z, Ashraf MY, Tahir GR (2004). Effect of salinity on chlorophyll concentration, leaf area, yield and yield components of rice genotypes grown under saline environment. Int J Environ Sci Technol 1: 221-225.
Arnon DI (1949). Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol 24: 1-15.
Bates LS, Waldren RP, Teare ID (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39: 205-207.
Bryant J, Green TR, Gurusaddaiah T, Ryan CA (1976). Proteinase inhibitor II from potatoes: isolation and characterization of its protomer components. Biochemistry 15: 3418-3424.
Cao FY, Yoshioka K, Desveaux D (2011). The roles of ABA in plant-pathogen interactions. J Plant Res 124: 489-499.
Delauney AJ, Verma DPS (1993). Proline biosynthesis and osmoregulation in plants. Plant J 4: 215-223.
Dombrowski JE (2003). Salt stress activation of wound-related genes in tomato plants. Plant Physiol 132: 2098-2107.
Fujita M, Fujita Y, Maruyama K, Seki M, Hiratsu K, Ohme-Takagi M, Tran LS, Yamaguchi-Shinozaki K, Shinozaki K (2004). A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J 39: 863-876.
Fujita M, Fujita Y, Noutoshi Y, Takahashi F, Narusaka Y, Yamaguchi-Shinozaki K, Shinozaki K (2006). Crosstalk between abiotic and biotic stress responses: a current view from the points of convergence in the stress signaling networks. Curr Opin Plant Biol 9: 436-442.
Gaddour K, Vicente-Carbajosa J, Lara P, Isabel-Lamoneda I, Díaz I, Carbonero P (2001). A constitutive cystatin-encoding gene from barley (Icy) responds differentially to abiotic stimuli. Plant Mol Biol 45: 599-608.
Gosti F, Bertauche N, Vartanian N, Giraudat J (1995). Abscisic acid-dependent and-independent regulation of gene expression by progressive drought in Arabidopsis thaliana. Mol Gen Genet 246: 10-18.
Gustafson G, Ryan CA (1976). Specificity of protein turnover in tomato leaves. Accumulation of proteinase inhibitors, induced with the wound hormone, PIIF. J Biol Chem 251: 7004-7010.
Haq SK, Khan RH (2003). Characterization of a proteinase inhibitor from Cajanus cajan (L.). J Protein Chem 22: 543-554.
Horsch RB, Fry JE, Hoffmann NL, Eichholtz D, Rogers SG, Fraley RT (1985). A simple and general method for transferring genes into plants. Science 227: 1229-1231.
Huang Y, Xiao B, Xiong L (2007). Characterization of a stress responsive proteinase inhibitor gene with positive effect in improving drought resistance in rice. Planta 226: 73-85.
Jardak-Jamoussi R, Elabbassi MM, Ben Jouira H, Hanana M, Zoghlami N, Ghorbel A, Mliki A (2014). Physiological responses of transgenic tobacco plants expressing the dehydration- responsive RD22 gene of Vitis vinifera to salt stress. Turk J Bot 38: 268-280.
Kavas M, Baloğlu MC, Yücel AM, Öktem HA (2016). Enhanced salt tolerance of transgenic tobacco expressing a wheat salt tolerance gene. Turk J Biol 40: 727-735.
Kim JY, Park SC, Hwang I, Cheong H, Nah JW, Hahm KS, Park Y (2009). Protease inhibitors from plants with antimicrobial activity. Int J Mol Sci 10: 2860-2872.
Kim S, Hong YN, An CS, Lee KW (2001). Expression characteristics of serine proteinase inhibitor under variable environmental stresses in hot pepper. Plant Sci 161: 27-33.
Li R, Wang W, Wang W, Li F, Wang Q, Xu Y, Wang S (2015). Overexpression of a cysteine proteinase inhibitor gene from Jatropha curcas confers enhanced tolerance to salinity stress. Electron J Biotechnol 18: 368-375.
Lopez F, Vansuyt G, Derancourt J, Fourcroy P, Casse-Delbart F (1994). Identification by 2D-PAGE analysis of salt-stress induced proteins in radish (Raphanus sativus). Cell Mol Biol 40: 85-90.
Mahmood T, Yasmin T, Haque MI, Naqvi SMS (2013). Characterization of a rice germin-like protein gene promoter. Genet Mol Res 12: 360-369.
Megdiche W, Ben Amor N, Debez A, Hessini K, Ksouri R, Zuily-Fodil Y, Abdelly C (2007). Salt tolerance of the annual halophyte Cakile maritima as affected by the provenance and the developmental stage. Acta Physiol Plant 29: 375-384.
Mello GC, Oliva MLV, Sumikawa JT, Machado OL, Marangoni S, Novello JC, Macedo MLR (2001). Purification and characterization of a new trypsin inhibitor from Dimorphandra mollis seeds. J Protein Chem 20: 625-632.
Mishra M, Mahajan N, Tamhane VA, Kulkarni MJ, Baldwin IT, Gupta VS, Giri AP (2012). Stress inducible proteinase inhibitor diversity in Capsicum annuum. BMC Plant Biol 12: 217.
Munir F, Hayashi S, Batley J, Naqvi SMS, Mahmood T (2015). Germin-like protein 2 gene promoter from rice is responsive to fungal pathogens in transgenic potato plants. Funct Integr Genomics 16: 19-27.
Nemoto Y, Sasakuma T (2002). Differential stress responses of early salt-stress responding genes in common wheat. Phytochemistry 61: 129-133.
Parida AK, Das AB (2005). Salt tolerance and salinity effect on plants: a review. Ecotoxicol Environ Saf 60: 324-349.
Park DS, Graham MY, Graham TL (2001). Identification of soybean elicitation competency factor CF-1 as soybean Kunitz trypsin inhibitor. Physiol Mol Plant Pathol 59: 265-273.
Pearce G, Johnson S, Ryan CA (1993). Purification and characterization from tobacco (Nicotiana tabacum) leaves of six small, wound-inducible, proteinase isoinhibitors of the potato inhibitor II family. Plant Physiol 102: 639-644.
Richardson M (1979). The complete amino acid sequence and the trypsin reactive (inhibitory) site of the major proteinase inhibitor from the fruits of aubergine (Solanum melongena L.). FEBS Lett 104: 322-326.

REHMAN et al. / Turk J Biol
502
Ryan CA (1990). Protease inhibitors in plants: genes for improving defenses against insects and pathogens. Annu Rev Phytopathol 28: 425-449.
Shan L, Li C, Chen F, Zhao S, Xia G (2008). A Bowman–Birk type protease inhibitor is involved in the tolerance to salt stress in wheat. Plant Cell Environ 31: 1128-1137.
Sin SF, Chye ML (2004). Expression of proteinase inhibitor II proteins during floral development in Solanum americanum. Planta 219: 1010-1022.
Solomon M, Belenghi B, Delledonne M, Menachem E, Levine A (1999). The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 11: 431-443.
Srinivasan T, Kumar KRR, Kirti PB (2009). Constitutive expression of a trypsin protease inhibitor confers multiple stress tolerance in transgenic tobacco. Plant Cell Physiol 50: 541-553.
Srivastava TP, Gupta SC, Lal P, Muralia PN, Kumar A (1988). Effect of salt stress on physiological and biochemical parameters of wheat. Ann Arid Zone 27: 197-204.
Tan Y, Li M, Ma F (2015). Overexpression of MpCYS2, a phytocystatin gene from Malus prunifolia (Willd.) Borkh., confers drought tolerance and protects against oxidative stress in Arabidopsis. Plant Cell Tiss Organ Cult 123: 15.
Tiwari LD, Mittal D, Chandra Mishra R, Grover A (2015). Constitutive overexpression of rice chymotrypsin protease inhibitor gene OCPI2 results in enhanced growth, salinity and osmotic stress tolerance of the transgenic Arabidopsis plants. Plant Physiol Biochem 92: 48-55.
Voigt EL, Almeida TD, Chagas RM, Ponte LF, Viégas RA, Silveira JA (2009). Source–sink regulation of cotyledonary reserve mobilization during cashew (Anacardium occidentale) seedling establishment under NaCl salinity. J Plant Physiol 166: 80-89.
Xu ZF, Qi WQ, Ouyang XZ, Yeung E, Chye ML (2001). A proteinase inhibitor II of Solanum americanum is expressed in phloem. Plant Mol Biol 47: 727-738.
Yadav D, Ahmed I, Shukla P, Boyidi P, Kirti PB (2016). Overexpression of Arabidopsis AnnAt8 alleviates abiotic stress in transgenic Arabidopsis and tobacco. Plants (Basel) 5: E18.
Zhang F, Li SF, Yang SM, Wang L, Guo W (2015). Overexpression of a cotton annexin gene, GhAnn1, enhances drought and salt stress tolerance in transgenic cotton. Plant Mo Biol 87: 47-67.
Zhang X, Liu S, Takano T (2008). Two cysteine proteinase inhibitors from Arabidopsis thaliana, AtCYSa and AtCYSb, increasing the salt, drought, oxidation and cold tolerance. Plant Mol Biol 68: 131-143.
Zhang XX, Tang YJ, Ma QB, Yang CY, Mu YH, Suo HC, Luo LH, Nian H (2013). OsDREB2A, a rice transcription factor, significantly affects salt tolerance in transgenic soybean. PLoS One 8: e83011.