characterization of some breeds and hybrids of … · in the present study, phd. thesis, the...
TRANSCRIPT

UNIVERSITY OF AGRICULTURAL SCIENCES AND VETERINARY MEDICINE CLUJ-NAPOCA
DOCTORAL SCHOOL FACULTY OF ANIMAL SCIENCE AND BIOTECHNOLOGIES
Biol. FURDUI EMILIA MARIA
CHARACTERIZATION OF SOME BREEDS AND
HYBRIDS OF BOMBYX MORI L. SILKWORMS
THROUGH MOLECULAR TECHNIQUES (SUMMARY OF PhD THESIS)
SCIENTIFIC COORDINATOR:
Prof. univ. dr. LIVIU ALEXANDRU MǍRGHITAŞ
262011
Cluj-Napoca

CONTENTS Thesis SummaryINTRODUCTION 1 31 PART I. BIBLIOGRAPHIC STUDY 2 CHAPTER I. THE PRESENT STAGE OF RESEARCH AND HISTORY OF SILKWORM BOMBYX MORI L. BREEDING IN WORLDWIDE AND IN ROMANIA
3 31
1.1. IMPORTANCE OF SERICULTURE 3 31 1.2. THE HISTORY OF SILKWORM BREEDING 6 1.3. WORLDWIDE PRESENT STAGE OF SILKWORM BREEDING
8 31
1.4. THE PRESENT STAGE OF SILKWORM BREEDING AND SERICULTURE RESEARCH IN ROMANIA
11 31
1.5. IMPORTANCE OF SILKWORM SCIENTIFIC RESEARCH ON SILKWORM
14 32
1.6. GENETIC STUDY OF SILKWORM BOMBYX MORI 16 1.6.1. Cytogenetic and molecular aspects of the silkworm Bombyx mori
17
1.6.2. Karyotype 18 1.6.3. Basics of chromosomal sex determination 21 1.6.4. The study of mutations occurred in silkworms 22 CHAPTER II. TAXONOMY AND EVOLUTIVE CYCLE OF BOMBYX MORI
26 32
2.1. TAXONOMY OF SILKWORM 26 32 2.2. EVOLUTIVE CYCLE OF BOMBYX MORI L. 28 32 CHAPTER III. METHODS OF GENETIC FINGERPRINTING 30 33 3.1. CLASSIFICATION OF MOLECULAR MARKERS 30 3.2. COMPARISON OF DIFFERENT MOLECULAR MARKING
TECHNIQUES 32
3.3. HIGHLIGHTING OF GENETIC VARIABILITY USING MOLECULAR MARKERS
34 33
3.3.1. PCR technique (Polymerase Chain Reaction) 34 3.3.2. General information regarding RAPD-Random Amplified
Polymorphic DNA 36
3.3.3. General information regarding microsattelites: SSR-Simple Sequence Repeats
40
PART II (PERSONAL RESEARCH) 45 PURPOSE OF RESEARCH 46 30 AIMED OBJECTIVES 47 30 CHAPTER IV. RESEARCH DESIGN OF SILKWORM BREEDING 49 34 4.1. BIOLOGICAL MATERIAL 49 34 4.2. MATERIALS USED IN EXPERIMENTAL BREEDING 59 34 4.3. METHODS USED FOR ORGANIZATION OF BREEDING BATCHES
63 34
4.4. DATA ANALYSIS 68 34
27

CHAPTER V- RESULTS AND DISCUSSIONS REGARDING THE EXPERIMENTAL GROWTH
70 35
5.1.INCUBATION AND HATCHING OF SILKWORM EGGS 70 35 5.2. EVOLUTION OF LARVAE STAGES 76 35 5.3. BIOLOGICAL PARAMETERS OF LARVAE 81 36 5.4. THE WEIGHT OF THE SILK GLANDS 86 5.4. DEVELOPMENT OF COCOON 88 5.6. BIOLOGICAL PARAMETERS OF RAW COCOONS 90 37 5.6.1. The weight of raw cocoons 90 5.6.2. Longitudinal and transversal axis of raw cocoons 92 5.7. THE TECHNOLOGICAL PARAMETERS OF DRY COCOON 94 37 5.7.1. The weight of dry cocoon 94 5.7.2. The longitudinal and transversal axis of dry cocoons 96 5.7.3. The length of silk filament 97 5.8. PRELIMINARY CONCLUSSIONS 100 38 CHAPTER VI. MATERIALS AND METHODS USED FOR RAPD ANALYSIS
103 38
6.1. MATERIALES USED FOR RAPD ANALYSIS 103 38 6.1.1. Biological material 103 6.1.2. Chemical materials 103 6.1.3. Electronic devices and instruments 104 6.2. METHODS 105 38 6.2.1. DNA extraction protocol 105 38 6.2.2. DNA quantification 107 38 6.2.3. PCR amplification protocol 108 6.2.4. Composition of PCR reaction 110 6.2.5. PCR Amplification 112 39 6.2.6. RAPD primers used for DNA amplification 113 6.2.7. Agarose gel electrophoresis 114 39 6.2.8. Electrophoretic migration and DNA samples preparation 115 6.2.9. Image capture 117 39 6.3. DATAS ANALYSIS 117 39 CHAPTER VII. RESULTS REGARDING THE POLYMORPHISM OBTAINED WITH RAPD PRIMERS
123 39
7.1. RESULTS REGARDING DNA EXTRACTION 123 7.1.1. Results regarding DNA extraction from silkworm breeds 123 7.1.2. Results regarding DNA extraction from silkworm hybrids 124 7.2. RESULTS REGARDING RAPD PRODUCTS
AMPLIFICATION AND ELECTROPHORESIS 126 39
7.2.1. Results regarding RAPD products amplification and electrophoresis for silkworm breeds
126
7.2.2. Results regarding RAPD products amplification and electrophoresis for silkworm hybrids
133
7.3. PRELIMINARY CONCLUSSIONS 139 43 CHAPTER VIII. MATERIALS AND METHODS USED FOR SSR ANALYSIS
141 44
28

8.1. MATERIALS USED FOR SSR ANALYSIS 143 44 8.1.1. Biological material 143 8.1.2. Chemical materials 143 8.1.3. Electronic devices and instruments 144 8.2. METHODS 144 8.2.1. DNA Extraction 144 44 8.2.2. Composition of PCR reaction 145 8.2.3. PCR amplification 147 44 8.2.4. Capillary electrophoresis 149 8.3. DATA ANALYSIS 151 44 CHAPTER IX. RESULTS REGARDING THE POLYMORPHISM OBTAINED WITH SSR PRIMERS
155 44
9.1. RESULTS OBTAINED WITH SSR PRIMERS 155 44 9.1.1. SSR alleles sizes generated by primers 156 9.1.2. Allelic structure 159 9.1.3. Distribution of unique alleles and fixed 160 45 9.2. HARDY-WEINBERG STABILITY 171 9.2.1. Hardy-Weinberg stability results for SAT 951 locus 172 9.2.2. Hardy-Weinberg stability results for SAT 1423 locus 174 9.2.3. Hardy-Weinberg equilibrium results for TO1CTA07R locus
176
9.2.4. Hardy-Weinberg Stability Results for SAT346 locus 177 9.2.5. Hardy-Weinberg stability results for CA16G03R locus 178 9.3. GENETIC DIVERSITY COEFFICIENTS 182 9.4. ANALYSIS OF GENETIC VARIATION AND DIFFERENTIATION AMONG BREEDS AND HYBRIDS
196 46
9.5. PRELIMINARY CONCLUSSIONS 202 47 CHAPTER X. GENERAL CONCLUSIONS 205 47 ORIGINAL ISSUES 209 48 LIST OF PUBLISHED PAPERS 211 BIBLIOGRAPHYC REFERENCES 213 49 APPENDIX 1 243 APPENDIX 2 245 APPENDIX 3 246 APPENDIX 4 248
29

PURPOSE OF RESEARCH
In the present study, PhD. thesis, the research purposes was the characterization of
races and hybrids of the silkworm Bombyx mori L. from Romania by phenotypic characterization (internationally recognized methods) and molecular technique (RAPD markers using Random Amplified Polymorphic DNA and SSR-Simple Sequence and repeats).
Research initiated by us, phenotypic and molecular characterization of the silkworm, is the first study of its kind in the field in Romania.
AIMED OBJECTIVES The research objectives were related at the following aspects: Accurate inventory of races and hybrids of the silkworm Bombyx mori by
morphological controls of genetic stock of races eggs, larvae, taking into account of phenotypic breed standards for local genetic from Romania.
The index studied for parental races and hybrids resulted from those are: -Biological parameters of eggs:
Hatching biological parameters Hatching percentage
-Biological parameters of larvae: Larval mass Larval weight Weight of the silk gland
- The biological parameters of the raw cocoon: Weight of raw cocoons Weight of the silk incartament Longitudinal axis of raw cocoons Transversal axis of raw cocoons
- Technical indices of cocoons and fibres: Weight of dry cocoons Weight of cocoons waste Length of silk filament
The objectives of research on molecular characterization are:
Using molecular techniques (RAPD and SSR) in order to identify a profile of races and hybrid studied;
Identification of RAPD primers capable to identifying the polymorphism at the molecular level;
Establishment of DNA extraction protocol; Inference of the phylogeny of Bombyx mori races and hybrids; Choosing and analysis the SSR primers able to discriminate races
and hybrids studied;
30

Extraction of DNA from larvae and highlight genetic profile at races and hybrids studied;
Molecular characterization and genetic diversity of races and hybrids of silkworm larvae studied with SSR markers.
INTRODUCTION
Of all species, which create cocoons from silk thread, 90% is produced by mulberry silkworm, followed by castor-oil plant and oak silkworms; the spread of these species is determined by the areas where the food plant species are growing.
Ever since cocoons fell into a cup of Chinese princess Xi Lingshi, 5000 years ago (Kurin, 2002), the butterfly Bombyx mori L., was intimately connected with humanity. The chinese have retained this secret for hundreds of years, and its disclodure was punishable by deadth. Being the base of sericiculture, the silk butterfly ensured the economic survivial of farmers and textile workers. The silk trade has fortified human activity through art and culture, helping to create an early form of globalization for about 2000 years, during the era of silk road (Kurin, 2002).
Bombyx mori represents the only truly domesticated insect, because is completely dependent for survial and for reproduction by man (Hubbell, 2001).
CHAPTER I. THE PRESENT STAGE OF RESEARCH AND HISTORY OF
SILKWORM BOMBYX MORI L. BREEDING IN WORLDWIDE AND ROMANIA
1.1. IMPORTANCE OF SERICULTURE
During proliferation the silkworm provides the silk manufacturing with raw material, namely cocoons. Natural silk has hight resistance, hight thickness (12-30 microns), high gloss, good touch and less electric charge than synthetic fibres, so that because of these traits it is considered the „luxury fiber textile of industry” (Bura, 1992).
The original country of silk is China. The historical and archaeological researchers found centres around the little city Schengze, from Wuxion region, Jiangsu province, the citizens cultivated mulberry trees and grew silkworms; they knew the pursuit of thread and fabric making.
1.2. WORLDWIDE PRESENT STAGE OF SILKWORM BREEDING
Currently, as reported in 2009 by the member countries of the International Association of Sericulture (I.S.C. Website), the world production is 126.995 tonnes registered cocoons. The first place in silk production is held by China (104.000 tonnes), followed by India (19690 tonnes), Brazil (811 tonnes), Uzbekistan (750 tonnes) etc. 1.3. THE PRESENT STAGE OF SILKWORM BREEDING AND SERICULTURE
RESEARCH IN ROMANIA
Nowadays sericiculture passes through a crisis; production of cocoons being endangered. As potential sericiculture, Romania is ranked second in Europa after France,
31

and as production in our country annually produces less than four tonnes of cocoons, annually (Paşca et al., 2008b), figure 1.
Due to favorable climatic condition mulberry cultivation has developed very rapidy in our country.
00.5
11.5
22.5
33.5
4
tone
/tons
2001 2002 2003 2004 2005 2006
Anul productiei/Production Year
Figure 1. Cocoons production between 2001-2006
1.4. IMPORTANCE OF SILKWORM SCIENTIFIC RESEARCH
The study of silkworms was initiated in Japan, under the impuls of developement of industry sericulture silk and quickly became a valuable model for fundamental studies. Nowadays silkworm has an important role in three areas: research, biotechnology and sericulture (Goldsmith et al., 2005); In scientific research, Bombyx mori were a privileged model for some discoveries in biology, genetics and medicine, such as analysis of genetic regulation of protein biosynthesis silk morphogenetic development, study of poikiloterme related adjustments, being premiers. Studies and research have taken place in other areas: endocrinology, toxicology, radiology, virology and biotechnology.
CHAPTER II. TAXONOMY AND EVOLUTIVE CYCLE OF BOMBYX MORI L.
2.1. TAXONOMY OF SILKWORMS: The mulberry silkworm take parts of:
Phylum: Artropoda; Subphylum: Mandibulata; Classa: Insecta; Subclass: Pterigota; Order Lepidoptere; Series: Lepidoptere nocturne; Family: Bombycidae; Gens: Bombyx Species: Bombyx mori
2.2. EVOLUTIVE CYCLE OF BOMBYX MORI L.
The mulberry silkworm or silkworm butterfly (Bombyx mori L.) is parts of the group of insects, which has a complete metamorphosis, in its developmental cycle
32

passing through 4 different stages: egg (or seed), larvae (silkworm), chrysalides (nymph or pupae) and butterfly (or adult), having one (monovoltine race) or more generations (polivoltine races) per year.
After fertilization the embryogenetic process lasts place for three days, then diapauses started. These are during function of race, and function of genetics’ and environmental factors, and at the newly formed races this take place for 9-10 months, generally from July to April. The eggs maintained are incubated for 10-14 days in optimal conditions, and this process ends whit the larva’s hatching.
The larval stage varies function of race, being between 25 and 35 days, and the larvae’s are passing thought five instars, delimitated by resting periods, called „sleep”. During these sleeping periods the larvae doesn’t eat and the moulting take places.
CHAPTER III. METHODS OF GENETIC FINGERPRINTING
3.1. HIGHLIGHTING OF GENETIC VARIABILITY USING MOLECULAR
MARKERS
RAPD markers (Random amplified Polymorphic DNA)-are product from PCR amplification of unknown genomic DNA segments with a random decamere primers (Williams et al., 1990). It is intensely in: gene identification (Aufauvre et al., 1992, Ballinger-Crabtree, 1992), variability of genetic population (Lehmann, 1992, Chatterjee et al., 2004), chromosomal maps and phylogenetic relationships (Kazan et al., 1993 ). The products of amplification are separated by agarose gel electrophoresis and made visible by UV light after prior staining with a specific dye. The image obtained is called the fingerprint. RAPD markers act as dominant markers when looking at the descent of individuals..
Genetic diversity and relationships of relationship between individuals are evaluated based on the presence or absence of bands. The advantages of RAPD analysis: highlighting a large number of polymorphisms, low cost, using fluorescent visualization methods instead of radioactivity, analyzing a large number of samples in one day, one involving a high degree of training of laboratory staff;
Disadvantages: RAPD markers dominant behavior and increased sensitivity of the technique.
Microsatellites (highly repetitive DNA) or SSR (Simple Sequence repeats) are short sequences highly repeated, usually containing 2-5 base pairs repeated in tandem, flanking the unique sequence DNA (Field et al., 1996).
Microsatellites have become widely used in many applications such as mapping the human genome (Dib et al., 1996), dogs (Ostander et al., 1993), the mice (Dietrich et al., 1996) of chicken (Crooijmans et al., 1996) of the silkworm Bombyx mori (Prasad et al., 2005a) but also of many plant species (Akkaya et al., 1995), phylogenetics (Morin et al., 1994), the genetic structure of populations (Bruford et al., 1993), the genetic conservation (Gotelli et al., 1994) and forensics (Jung, 1989, Jeffreys et al., 1991).
Polymorphism is due to variation in the number of repetitions in tandem PCR amplification with primers from the complementary sequences flanking the microsatellites DNA (sequences conserved within species, and sometimes genera). SSR markers are given advantages: high polymorphism, their uniform distribution in the genome, high reproducibility and co- dominance.
33

Among the main disadvantages is the cost constraint and the need to develop knowledge primers for the DNA sequence of interest.
CHAPTER IV. RESEARCH DESIGN OF SILKWORM BREEDING
4.1. BIOLOGICAL MATERIAL In the experiments performed in 2009, the biological material consisted of: -7 races of silkworms: RG 90; AC/T (Alb de Cislău); AB (Alb de Băneasa);
IBV; AC 29/T (Alb Chinezesc); B1 (Băneasa1); S8; -8 consecrated hybrids: S8 X AC29/T (Ana 1); AC 29/T X S8 (Ana 2); AC x B1
(Cislău 1); B1 X AC (Cislău 2); B1 X SVILA2 (Bulgaria); BB1 X HESA2 (Bulgaria); HESA1 X SVILA 2 (Bulgaria); VRATZA 35 X SVILA 2 (Bulgaria); 4.2. MATERIALS USED IN EXPERIMENTAL BREEDING
The materials used in the method growth of silkworm are: plantation mulberry and mulberry leaves taken, the space for growth and experimental series of cocoons development materials used. 4.3. METHODS USED FOR ORGANIZATION OF BREEDING BATCHES
• Incubation of silkworm eggs – gradually raising the temperature; • Conditions for growth and development of larvae, T: 26-27 ° C, 25-26 ° C, 23-24 °
C, H: 80-85%, 75%, 65-70%; • Cocoon development: T-22 and C, H-65%; • Cocoon stove; • Reeling basin.
4.4. DATA ANALYSIS
Data analysis: • Software: PAST 2.04; The coefficient of variation (V%);
With regard to the objectives determinations were performed as follows: The number of individuals studied was different depending on the characteristics analyzed. Thus the biological parameters (prolificity, and percentage of hatching) expresses the average results of four different variants both races and hybrids studied;
Larval weight was studied for 20 individuals, the length of larvae for 20, silk-secreting gland mass was determined glands extracted from 20 larvae, cocoons raw and dry mass, incartment silky, and transverse axis of raw and dried cocoons were determined by weighing -20 cocoons, filament length and the waste of about 10 cocoon.
34
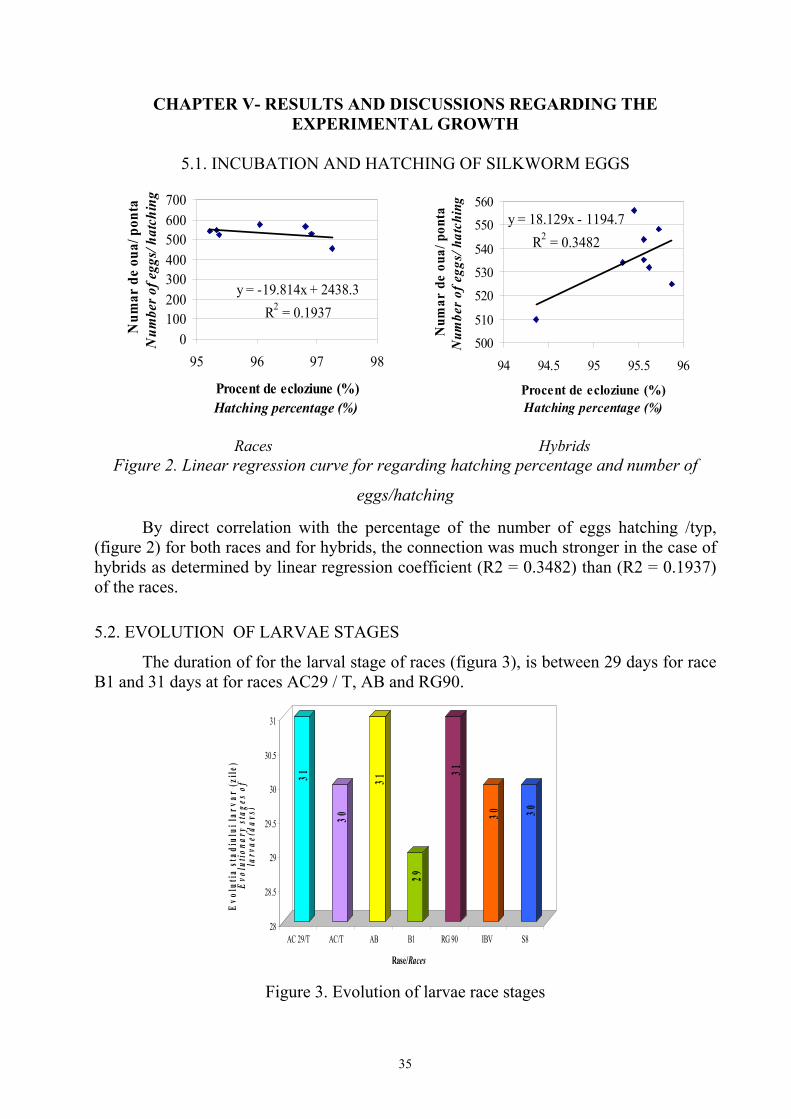
CHAPTER V- RESULTS AND DISCUSSIONS REGARDING THE EXPERIMENTAL GROWTH
5.1. INCUBATION AND HATCHING OF SILKWORM EGGS
y = -19.814x + 2438.3R2 = 0.1937
0100200300400500600700
95 96 97 98
Procent de ecloziune (%) Hatching percentage (%)
Num
ar d
e ou
a/ p
onta
N
umbe
r of e
ggs/
hatc
hing
y = 18.129x - 1194.7R2 = 0.3482
500
510
520
530
540
550
560
94 94.5 95 95.5 96
Procent de ecloziune (%) Hatching percentage (%)
Num
ar d
e ou
a/ p
onta
N
umbe
r of
egg
s/ h
atch
ing
Races Hybrids
Figure 2. Linear regression curve for regarding hatching percentage and number of
eggs/hatching
By direct correlation with the percentage of the number of eggs hatching /typ, (figure 2) for both races and for hybrids, the connection was much stronger in the case of hybrids as determined by linear regression coefficient (R2 = 0.3482) than (R2 = 0.1937) of the races.
5.2. EVOLUTION OF LARVAE STAGES
The duration of for the larval stage of races (figura 3), is between 29 days for race B1 and 31 days at for races AC29 / T, AB and RG90.
31
30
31
29
31
30 30
28
28.5
29
29.5
30
30.5
31
AC 29/T AC/T AB B1 RG 90 IBV S8
Rase/Races
Evol
utia
stad
iulu
i lar
var
(zile
)Ev
olut
iona
ry st
ages
of
larv
ae(d
ays)
Figure 3. Evolution of larvae race stages
35

28 28
29 29
28 28
29
30
27
27.5
28
28.5
29
29.5
30
B1X
AC
ACX
B1
AC2
9/TX
S8
S8X
AC2
9/T
HES
A1X
SVIL
A2
B1X
SVIL
A2
B1X
HES
A2
VRA
TZA
35X
SVIL
A2
Hibrizi/Hybrids
Evo
lutia
stad
iulu
i lar
var
(zile
)Ev
olut
iona
ry st
ages
of l
arva
e(da
ys)
Figure 4. Evolution of larvae hybrids stages
Duration of larval stages for hybrids (figure 4), is between 28 days for hybrids B1 X AC, AC x B1, HESA1 x SVILA2, B1 x SVILA2 and 30 days for VRATZA35 x SVILA2.
5.3. BIOLOGICAL PARAMETERS OF LARVAE
y = 2.3741x - 12.479R2 = 0.9458
0123456
6 6.5 7 7
Lungimea larvelor (cm) Larvae length (cm)
Mas
a la
rvel
or (g
)
Larv
ae w
eigt
h (g
)
.5
y = 0.9643x - 2.5281R2 = 0.2204
4.2
4.4
4.6
4.8
5
5.2
7.2 7.4 7.6 7.8 8
Lungimea larvelor (cm) Larvae length (cm)
Mas
a la
rvel
or (g
)
Larv
ae w
eigt
h (g
)
Races Hybrids Figure 5. Linear regression curve regarding larvae lenght and weight -races and hybrids
In case silkworm races by direct correlation of length and mass of larvae, figure 5, one can observe a positive correlation, estimated by linear regression coefficient (R2 = 0.9458), which means that the length of the larvae is correlated almost perfectly with larval mass. In the case of hybrids, the linear regression coefficient (R2 = 0.2204), the
36
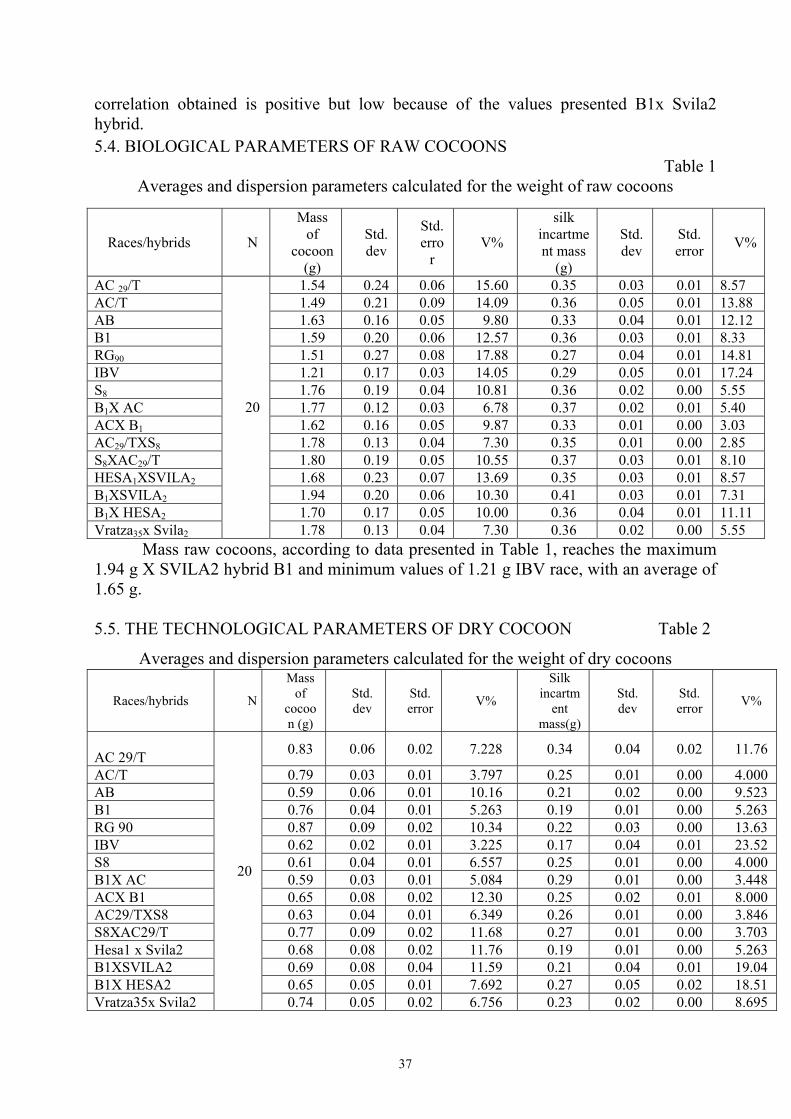
correlation obtained is positive but low because of the values presented B1x Svila2 hybrid. 5.4. BIOLOGICAL PARAMETERS OF RAW COCOONS
Table 1 Averages and dispersion parameters calculated for the weight of raw cocoons
Mass raw cocoons, according to data presented in Table 1, reaches the maximum 1.94 g X SVILA2 hybrid B1 and minimum values of 1.21 g IBV race, with an average of 1.65 g.
5.5. THE TECHNOLOGICAL PARAMETERS OF DRY COCOON Table 2
Averages and dispersion parameters calculated for the weight of dry cocoons
Races/hybrids N
Mass of
cocoon (g)
Std. dev
Std. erro
r V%
silk incartment mass
(g)
Std. dev
Std. error V%
AC 29/T 1.54 0.24 0.06 15.60 0.35 0.03 0.01 8.57 AC/T 1.49 0.21 0.09 14.09 0.36 0.05 0.01 13.88 AB 1.63 0.16 0.05 9.80 0.33 0.04 0.01 12.12 B1 1.59 0.20 0.06 12.57 0.36 0.03 0.01 8.33 RG90 1.51 0.27 0.08 17.88 0.27 0.04 0.01 14.81 IBV 1.21 0.17 0.03 14.05 0.29 0.05 0.01 17.24 S8 1.76 0.19 0.04 10.81 0.36 0.02 0.00 5.55 BB1X AC 1.77 0.12 0.03 6.78 0.37 0.02 0.01 5.40 ACX B1 1.62 0.16 0.05 9.87 0.33 0.01 0.00 3.03 AC29/TXS8 1.78 0.13 0.04 7.30 0.35 0.01 0.00 2.85 S8XAC29/T 1.80 0.19 0.05 10.55 0.37 0.03 0.01 8.10 HESA1XSVILA2 1.68 0.23 0.07 13.69 0.35 0.03 0.01 8.57 BB1XSVILA2 1.94 0.20 0.06 10.30 0.41 0.03 0.01 7.31 BB1X HESA2 1.70 0.17 0.05 10.00 0.36 0.04 0.01 11.11 Vratza35x Svila2
20
1.78 0.13 0.04 7.30 0.36 0.02 0.00 5.55
Races/hybrids N
Mass of
cocoon (g)
Std. dev
Std. error V%
Silk incartm
ent mass(g)
Std. dev
Std. error V%
AC 29/T 0.83 0.06 0.02 7.228 0.34 0.04 0.02 11.76
AC/T 0.79 0.03 0.01 3.797 0.25 0.01 0.00 4.000 AB 0.59 0.06 0.01 10.16 0.21 0.02 0.00 9.523 B1 0.76 0.04 0.01 5.263 0.19 0.01 0.00 5.263 RG 90 0.87 0.09 0.02 10.34 0.22 0.03 0.00 13.63 IBV 0.62 0.02 0.01 3.225 0.17 0.04 0.01 23.52 S8 0.61 0.04 0.01 6.557 0.25 0.01 0.00 4.000 B1X AC 0.59 0.03 0.01 5.084 0.29 0.01 0.00 3.448 ACX B1 0.65 0.08 0.02 12.30 0.25 0.02 0.01 8.000 AC29/TXS8 0.63 0.04 0.01 6.349 0.26 0.01 0.00 3.846 S8XAC29/T 0.77 0.09 0.02 11.68 0.27 0.01 0.00 3.703 Hesa1 x Svila2 0.68 0.08 0.02 11.76 0.19 0.01 0.00 5.263 B1XSVILA2 0.69 0.08 0.04 11.59 0.21 0.04 0.01 19.04 B1X HESA2 0.65 0.05 0.01 7.692 0.27 0.05 0.02 18.51 Vratza35x Svila2
20
0.74 0.05 0.02 6.756 0.23 0.02 0.00 8.695
37

As a result of determinations we can could see that the average value of 0.87 g
was recorded at RG race 90, and the lowest average of 0.59 g to hybrid B1X AC and race AC, the average mass of dry cocoons races and hybrids is 0.69 g, the mean minimum of incartment was registered for race IBV 0.17 g and a maximum of 0.34g for race AC29/T.
Coefficients of variation obtained for dry mass and mass of silk incartment are in all cases below 15%, which shows a great homogeneity of races and hybrids analyzed.
In terms of filament length, the average maximum was recorded at 1256 m AC29/TX S8 hybrid, and the minimum 1008 m B1X HESA2 hybrid, races are found between hybrids, with maximum values of 1210 m to race AC/T and minimum values of 1038 m on IBV race.
5.6. PRELIMINARY CONCLUSIONS
For the larval mass parameters the biological coefficient of variation shows an average of 8.04%, you can see a great homogeneity of the races studied. By direct correlation between length and mass of larvae in races, one can observe a positive correlation, estimated by linear regression coefficient (R2 = 0.9458), which means that larvae length is almost perfectly correlated with larval mass.
CAPITOLUL VI. MATERIALS AND METHODS USED FOR RAPD ANALYSIS
6.1. MATERIALES USED FOR RAPD ANALYSIS
Biological material used in the first stage: 6 rase: AB, IBV, B1, RG90, AC29/T, AC/T and 2 hybrids: H1 = ♂ AC/T x ♀S8XAc29/T; H2 = ♂ AC/T x ♀ AC29/T;
- And in the second stage 8 of hybrids hybrids consecrated: S8 x AC 29/T, AC 29/T x S8, AC x B1, B1 x AC, Hesa1 x Svila2, B1 x Svila2, B1 x Hesa2, Vratza35 x Svila2 and one race S8;
6.2. METHODS 6.2.1. DNA extraction protocol
In order to obtain a high quality DNA that can be used for the RAPD technique, as a first step genomic DNA was isolated from silkworm eggs from SC Sericarom Baneasa.
In the second stage DNA isolation was carried out from eggs (Hesa1 x Svila2, B1 x Svila2, B1 x Hesa2, Vratza35 x Svila2) and from silk glands of silkworm Bombyx mori L. (instars five) (S8 x AC 29/T, AC 29/T x S8, AC x B1, B1 x AC, S8).
Silkworm larvae were reared in family module at Beekeeping and Sericiculture laboratory UASVM Cluj-Napoca, and sampled in instar five and then kept at -20 ° C until used. Isolation of the biological material was performed by grinding and mixing in liquid nitrogen according to the protocol described by Suzuki et al., in 1972. DNA extraction continued with the Wizard Genomic DNA purification kit [Promega] - (WGDP) - which is a kit that provides DNA extraction of high purity. 6.2.2. DNA quantification The usual method for assessing the purity of DNA is the spectrophotometry, to quantify the extracted DNA the NanoDrop ND-1000 was used.
38

6.2.3. PCR Amplification
PCR amplification of genomic DNA was performed by 44 RAPD primers, amplification protocol described by Williams et al., 1990, as amended by Nagaraja and et al., in 1995, this amplification protocol was optimized depending on the used termocycler 2009 (BioRad, iCycler), the temperature used in the present experiment was 1-2°C lower, and for the extension time was reduced by half. To reflect polymorphisms between races and silkworm hybrids studied, 44 RAPD primers were chosen RAPD primers, according to the literature. 6.2.4. Electrophoretic migration – in 2% agarose gel. 6.2.5. Image capture
Product amplification was viewed by electrophoresis under UV light stands, image capture is performed using the UVP system (AC BioSpectrum Imaging System) Biochemie HR Camera (Made in Japan), and SYBR Gold filter (485-655nm). The image obtained is called the fingerprint. 6.3. DATA ANALYSYS
The first step: bioinformatics data analysis was performed by Neighbor Joining, Nei-Li coefficient with 1000 Bootstrap repeats.
In the second stage: interpretation of results was performed by Tree Free 0.9.1.50 program using Jaccard coefficient for genetic distances, and UPGMA method;
CHAPTER VII. RESULTS REGARDING THE POLYMORPHISM OBTAINED
WITH RAPD PRIMERS
7.1. BIOLOGICAL MATERIAL DNA extraction (Bombyx mori) showed that the liquid nitrogen used according to
(Suzuki, 1972) resulted in better grinding and mixing of samples. DNA fragments were amplified using PVP 2% without any problem the reaction mixture.
7.2. RESULTS REGARDING RAPD PRODUCTS AMPLIFICATION AND
ELECTROPHORESIS 44 RAPD primers tested were used to assess genetic diversity on all hybrids, 35. RAPD reactions by primers generated a total of 876 polymorphic fragments with
an average of 8.9 fragments/primer. These 35 primers were chosen based on the number of bands polymorphic amplifications data and repeatability.
39
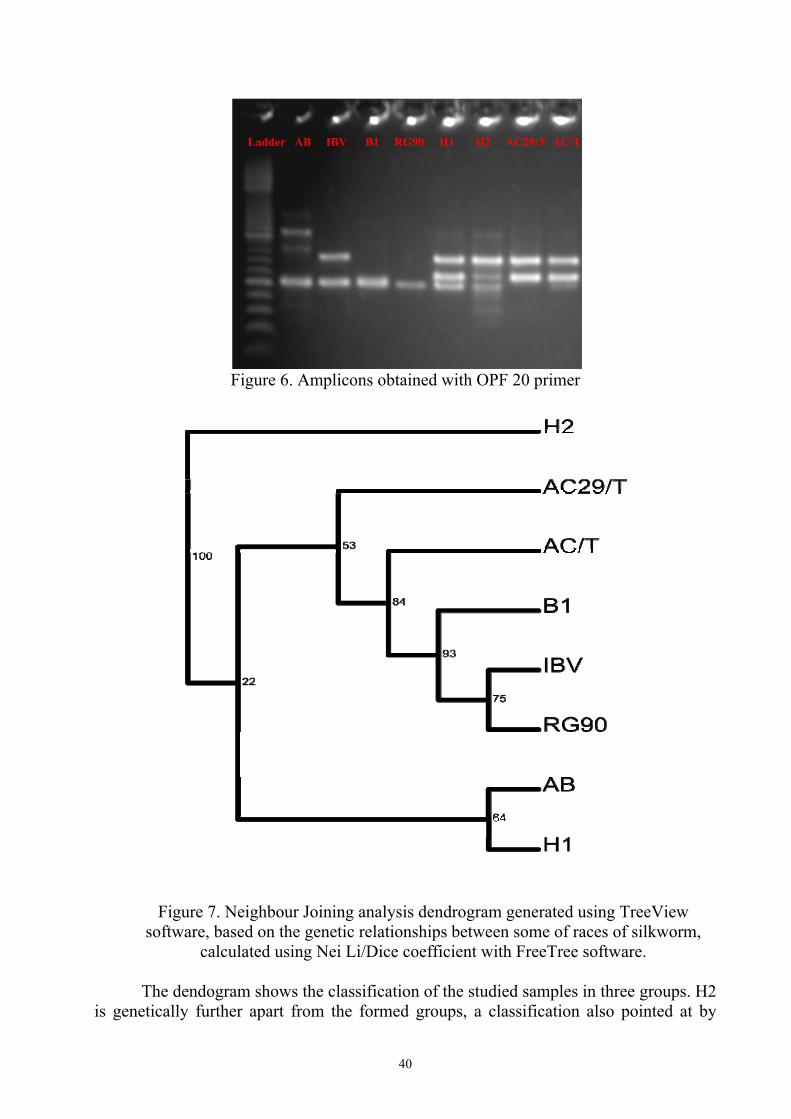
Ladder AB IBV B1 RG90 H1 H2 AC29/T AC/T
Figure 6. Amplicons obtained with OPF 20 primer
Figure 7. Neighbour Joining analysis dendrogram generated using TreeView software, based on the genetic relationships between some of races of silkworm,
calculated using Nei Li/Dice coefficient with FreeTree software.
The dendogram shows the classification of the studied samples in three groups. H2 is genetically further apart from the formed groups, a classification also pointed at by
40

bootstrap maximum value, (100). It is natural for this to happen because H2 is a hybrid. AB and H1 have formed a group and the other is made up from AC29/T, AC/T, B1, IBV and RG90. Bootstrap values are pointed out in the dendogram at nodes, with values above 50%, the only exception being 22% value, which confirms the two groups: AB and H1, respectively AC29/T, AC/T, B1, IBV and RG 90.
Table 3
Analysis of the calculated values of genetic similarity indices related to races and hybrids
analyzed
Rasa/ Hibridul AB IBV B1 RG90 H1 H2 AC29/T AC/T
AB IBV 0,42 B1 0,58 0,62 RG90 0.21 0,43 0,47 H1 0.75 0,50 0,63 0,27 H2 0,65 0,46 0,50 0,25 0,69 AC29/T 0,62 0,48 0,59 0,24 0,69 0,64 AC/T 0,57 0,56 0,57 0,38 0,62 0,59 0,65
H1 = ♂ AC/T x ♀ S8xAC29/T; H2 = ♂ AC/T x ♀ AC29/T.
The highest genetic proximity (0.75) was registered between AB and H1 samples, and the smallest (0.21) between AB and RG 90. The average genetic distance among individuals was 0.48.
In the second stage 21 of a total of 35 primers using primers generated 921 polymorphic bands that could enter the mathematical-statistical analysis of the interpretation of relations phylogeny Total Lab 120, UVP software.
The UPGMA cluster analysis separated the silkworms into 3 main groups, one consisting of hybrids Hesa 1 x Svila 2, which formed a separate group, reinforced by a very high bootstrap value, equal to 100. The second group consisted of S8 race, S8 hybrids and AC hybrids. The third group consisted of B1 and Svila hybrids.
The separate clustering of Hesa 1 x Svila 2 hybrids can be due to the presence of non-parental bands generated by RAPD markers, the cause of this phenomenon can be explained by more hypotheses: the formation of heteroduplex molecules between the alleles, mutations or recombination at the primer binding site or inside the amplified fragments, competition for primer binding sites or somatic rearrangements as in perennial plants (Nicese et al., 1998).
The samples AC x B1 and B1 x AC clustered together (bootstrap value 70), as well as the samples B1 x Svila2 and Vratza 35 x Svila2 (bootstrap value 90) and also the samples AC28/TxS8 and S8xAC29/T.
41
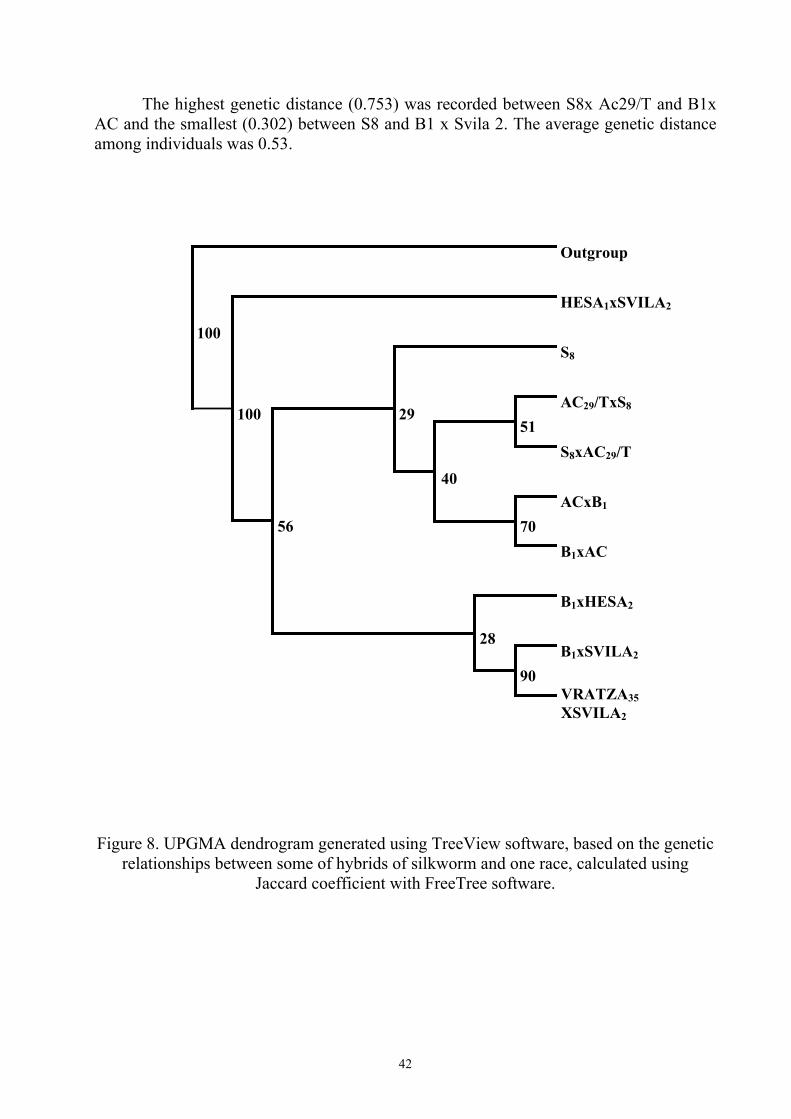
The highest genetic distance (0.753) was recorded between S8x Ac29/T and B1x AC and the smallest (0.302) between S8 and B1 x Svila 2. The average genetic distance among individuals was 0.53.
Outgroup
HESA1xSVILA2
S8
AC29/TxS8
S8xAC29/T 51
ACxB1
B1xAC 70
40
29
B1xHESA2
B1xSVILA2
VRATZA35 XSVILA2
90
28
56
100
100
Figure 8. UPGMA dendrogram generated using TreeView software, based on the genetic relationships between some of hybrids of silkworm and one race, calculated using
Jaccard coefficient with FreeTree software.
42

Table 4
Analysis of the calculated values of genetic similarity indices related to samples analyzed
Race/ Hybrid
Vratza3
5x Svila2
BB1x Svila2
Hesa1x Svila2
S8xAC29/T
BB1x Hesa2
ACxBB1 S8AC29/TxS8
BB1xAC
Vratza35 x Svila2 BB1 X Svila2 0.557 Hesa1 X Svila2 0.689 0.481 S8XAC29/T 0.639 0.479 0.553 BB1X Hesa2 0.444 0.438 0.532 0.527 ACXBB1 0.582 0.365 0.352 0.503 0.436 S8 0.609 0.302 0.494 0.434 0.440 0.376 AC29/TXS8 0.393 0.538 0.591 0.599 0.452 0.516 0.523 BB1 X AC 0.590 0.664 0.672 0.753 0.652 0.687 0.708 0.495
7.3. PRELIMINARY CONCLUSIONS
• The first step: DNA extractions from Bombyx mori eggs showed that using liquid nitrogen results in better crushing and mixing of samples;
• 34 from 44 primers generated polymorphic bands; • The average genetic distance was 0,48; • Significant differences are pointed out on genotype level between studied
breeds, which shows that Bombyx mori posses a well established genetic profile. Its inter-population genetic rate leads to the differentiation of co-adapted genes. The native Bombyx mori races can be used as initial amelioration material, characterized by genotype stability and expressing distinguished phenotypes features.
• In the second stage, and DNA extractions were made from silkglands (larvae of silkworm in fifth instar) and from eggs, it was observed that the highest purity was obtained from samples containing silkglands;
• 21 from 35 primers generated polymorphic bands; • The average genetic distance between the samples was 0.53. The average
genetic distance from analyzed samples proved to be relatively high, which can be due to the fact that hybrids are from two different races and also to the distant origin of these races
43

CHAPTER VIII. MATERIALS AND METHODS USED FOR SSR ANALYSIS 8.1. MATERIALS USED FOR SSR ANALYSIS
8.1.1. Biological material
The biological material was represented by: 7 races grown in 2009: AB, IBV, B1, RG 90, Ac 29/T, Ac/T , S8; 3 races grown in 2011: AB, B1, AC/T and 8 hybrids: S8 x Ac 29/T, Ac 29/T x S8, Ac x B1, B1 x Ac, Hesa1 x Svila 2, B1 x Svila 2, B1 x Hesa 2, Vratza 35 x Svila 2; For a more complete analysis 10 individuals were taken from each race and hybrid. 8.2. METHODS 8.2.1. DNA EXTRACTION
Individuals were taken from -20 ºC and DNA isolation was made from silk-secreting gland middle portion for each silkworm. Extraction was performed with Chelx 5%.
8.2.2. DNA quantification was determined using a NanoDrop ND-VIS spectrophotometer UV-2000;
8.2.3. PCR amplification PCR amplification was performed by repeating 35 cycles in a termocycler
(BioRad C 1000), nine SSR primers were tested, their sequences are taken from the literature, and for those that worked well (SAT 951, SAT 1423, TO1CTA07R, SAT 346, CA16G03R) forward forward primers were labeled and the products scored in a capillary electrophoresis. Also, to reduce the running costs of capillary electrophoresis, primers were labeled by different fluorochrome (D2, Cy5, Cy5.5) to run them in a multiplex mix. Fragment Analysis Protocol Genome Lab protocol was adapted so that they were prepared for their introduction into Beckman-Coulter equipment.
8.3. DATA ANALYSIS
Analysis of chromatograms was performed using CEQTM Fragments Analysis program distributed by Beckman Coulter.
GenAlEx Program 6.41, Chi-square test for evidence of genetic balance HW, Amova test (analysis of molecular variance coefficient), coefficient RST and Nei, StatSoft Inc. program, Statistics Data Analysis System software, version 10.
CHAPTER IX. RESULTS REGARDING THE POLYMORPHISM
OBTAINED WITH SSR PRIMERS
9.1. RESULTS OBTAINED WITH SSR PRIMERS The results on DNA purity, from 5% Chelex extraction, shows the advantage of
using the method of extraction, we can work faster and can be easily stored at -20°C. This extraction method was adapted in this doctoral thesis and is a first as a method of
44

extraction and purification of DNA of silkworm in Romania.
Table 5 Alleles obtained with SSR primers in silkworms
Primer SAT 951 SAT 1423 TO1CTA07R SAT 346 CA16G03R
No. of alleles 6 8 6 4 7
Alleles 97 159 244 131 22499 161 246 135 226105 165 248 139 236111 167 264 141 238113 169 266 244117 171 282 246
173 248 177
The number of alleles identified for each primer is between four and eight for loci SAT 346 and SAT 1423.
Regarding S8 race and S8xAC29/T hybrid, they are in Hardy-Weinberg balance for all 5 loci analyzed, being the only ones in genetic balance for all used loci.
The average number of analyzed individuals is 7.4 ± 0.30, the average alleles per locus is 2.40±0.11, effective number of alleles per locus is 1.83±0.08. The observed heterozygosity (H0), has lower values than expected heterozygosity, overall locuses presents a value of 0.290±0.031 while He which has a value of 0.374±0.026, which signifies that 29% of 180 analyzed individuals are heterozygotes.
According to linking value for all analyzed races and hybrids in this study, the result was that they all present a homozygotes excess. 9.2. Alleles distribution
Figure 9. Alleles distribution
45
9.3. ANALYSIS OF GENETIC VARIATION AND DIFFERENTIATION AMONG
RACES AND HYBRIDS
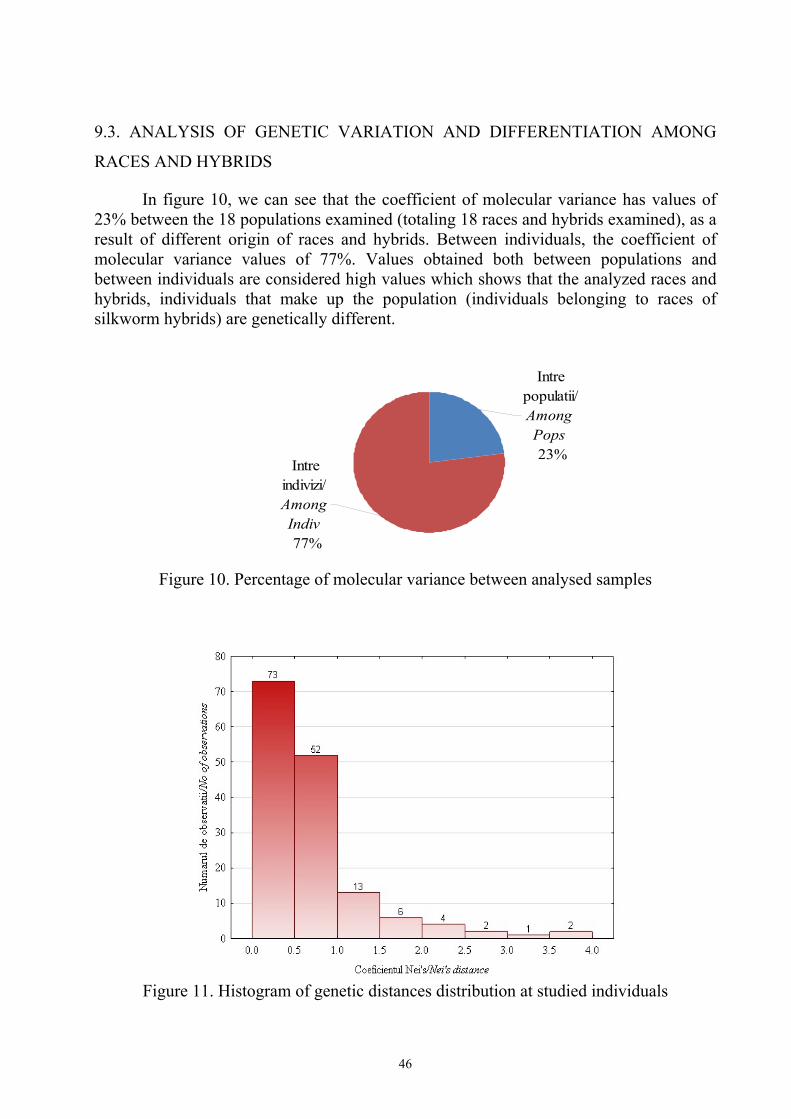
In figure 10, we can see that the coefficient of molecular variance has values of 23% between the 18 populations examined (totaling 18 races and hybrids examined), as a result of different origin of races and hybrids. Between individuals, the coefficient of molecular variance values of 77%. Values obtained both between populations and between individuals are considered high values which shows that the analyzed races and hybrids, individuals that make up the population (individuals belonging to races of silkworm hybrids) are genetically different.
Intre indivizi/ Among Indiv 77%
Intre populatii/Among Pops 23%
Figure 10. Percentage of molecular variance between analysed samples
Figure 11. Histogram of genetic distances distribution at studied individuals
46

In figure 11 genetic distances are visible for the 153 samples analyzed, the average distance of them being 0.51.
Large genetic distances are found to race IBV, this confirms that race IBV is different from the other races, because it is a race polyvoltine by origins, this is not considered an impediment because over the years it has been shown that this race is a stable and homogeneous, as illustrated in SSR analysis. 9.4. PRELIMINARY CONCLUSIONS
1. Five primers were used that mark I annotated (SAT 951, SAT 1423, TO1CTA07R, SAT 346, CA16G03R), each primer was used for amplification of individual samples until they are placed in Beckman, was therefore developed a multiplex system to reduce the costs of running the electrophoresis capillary.
2. The coefficient of variance identified significant values between races and hybrids analyzed (23%) and between individuals analyzed (77%).
CHAPTER X. GENERAL CONCLUSIONS
1. This study is the first of its kind in Romania, the importance of being given
the need for molecular characterization of races and hybrids of species of silkworm Bombyx mori L., which are part of the genetic sericulture native of Romania.
2. In this study we used two molecular techniques, specifically for the analysis
proposed two molecular markers: RAPD-markers used in screening tests and SSR-markers used for molecular characterization of races and hybrids of silkworm Bombyx mori in Romania.
3. As a result of experimental raceing performed in this study, for each
investigated parameter the following hierarchy of the races and hybrids was determined:
• Hatching biological parameters: race AC/T; hybrids B1 X Hesa2; • Hatching percentage: race IBV; hybrids Hesa1 x Svila2; • Larval lenght: race AC29/T; hybrids B1X AC; • Larval weight: race AC/T; hybrids B1 X Hesa2; • Weight of silkglands: race B1; hybrids B1 X Svila 2; • ent: hybrids B1 X Svila 2; Weight of raw cocoons and incartam• Weight of dry cocoon- race RG90; • Weight of dry incartament: rasa AC29/T; • Length of silk filament: hybrids: AC29/T X S8.
47

4. It is obvious that there are major differences at the genetic level between analyzed races and hybrids, which shows that there are populations with strong genetic profile. Restricted interpopulation genetic flow generated the cleavage of a stabile assembly of coadaptated genes.
5. Regarding S8 race and S8XAC29/T hybrid, they are in Hardy-Weinberg balance for all 5 locuses that we have analyzed.
6. The observed heterozygosity (H0) has lower values than expected heterozygosity overall locuses presents a value of 0.290±0.031 while He which has a value of 0.374±0.026, which signifies that 29% of 180 analyzed individuals are heterozygotes.
7. According to linking value for all analyzed races and hybrids in this study, the result was that they all present a homozygotes in excess.
ORIGINAL ISSUES
1. Degree of novelty of this thesis is that races and hybrids of silkworm Bombyx mori have been fully characterized both phenotypically by standard methods and by analyzing primary molecular RAPD and microsatellite analysis of the complex.
2. DNA extraction protocol by Chelex 5% and proteinase K used, is a first in
the use of silkworm in Romania.
3. Because of polymorphism loci highlighted by the study were able to
identify races and hybrids susceptible to inraceing depression for the next few years, they still showing an excess of homozygosity now.
RECOMMANDATIONS AND PERSPECTIVES
1. It is recommended further research regarding the study of SSR
polymorphism analysis especially to all races and hybrids of the native gene stock to increase the amount of information about them and to identify parental races for heterozygous hybrids.
2. In order to achieve a better conservation of races and hybrids of silkworms
that are part of the genetic stock of Romania, it is necessary to establish accurate genotyping of them. It is good to be determined on two levels: phenotypic and genotypic.
48

REFERENCES 1. Akkaya, M. S., Shoemaker, R. C., Specht, J.E., Bhagwat, A. A., and P. B.,
Cregan, 1995, Integration of simple sequence repeats DNA markers into a soybean linkage map. Crop. Sci. 35, 1439-1445.
2. Aufauvre-Brown, A., J. Cohen, and D.W. Holden, 1992, Use of Randomly Amplified Polymorphic DNA Markers to Distinguish Isolates of Aspergillus fumigatus, J. Clin. Microbiol., 30, 299l -2993.
3. Ballinger-Crabtree, M. E., W. C. V. Black, and B. R. Miller (1992). Use of Genetic Polymorphisms Detected by the Random Amplified Polymorphic DNA Polymerase DNA Polymerase Chain Reaction (RAPD-PCR) for Differentiation and ldentification of Aedes Aegpti Subspecies and Population, Amer. J. Trop. Med & Hyg., 47, 893-901.
4. Bruford, M.W. and R.K. Wyne, 1993, Microsatellites and their application to population genetic studies. Curr. Opin. Genet. Dev. 3, 939-943.
5. Bura, M., 1992, Curs de tehnologia producţiei sericicole. Universitatea de Ştiinţe Agricole a Banatului Timişoara.
6. Chatterjee, S.N. and T. Tanushree, 2004, Molecular Profiling of Silkworm Biodiversity in India: An Overview. Russian Journal of Genetics, Vol. 40, No. 12, 2004, pp. 1339-1347. From Genetika, Vol. 40, No. 12, 1618-1627.
7. Crooijman, R.P.M.A., P.A.M. Vanoers, J.A. Strijk, J.J. Vanderpoel, M.A. M., Groenen, 1996, Preliminary linkage map of the chicken (Gallus domesticus) genome based on microsatellite markers- 77 new markers mapped. Poult. Sci. 75: 746-754.
8. Dib, C., Sabine Fauré, Cécile Fizames, Delphine Samson, Nathalie Drouot, A. Vignal, P. Millasseau, Sophie Marc, J. Kazan, E. Seboun, M. Lathrop, G. Gyapay, J. Morissette and J. Weissenbach, 1996, A comprehensive genetic map of the humain genome based on 5264 microsatellites, Nature 380:152-154.
9. Dietrich, W.F., J. Miller, R. Steen, M.A. Merchant, D. Damronboles, Z. Husain, R. Dredge, M.J. Daly, K.A. Ingalls, T.J. O’Connor, C.A. Evans, M. M., DeAngelis, D.M. Levinson, L. Kruglyak, N. Goodman, N.G. Copeland, N.A. Jenkins, T.L. Hawkins, L.Stein, D.C. Page, and E.S. Lander, 1996, A comprehensive genetic map of the mouse genome, Nature, 380:149-152.
10. Field, D., and C. Willis, 1996, Long polymorphic microsatellites in simple microorganisms. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 263:209, 215.
11. Goldsmith, M.R., T. Shimada and A. Hiroaki, 2005, The genetics and genomics of the silkworm, Bombyx mori, Annual Review of Entomology, Proquest Agricultura Journals,71.
12. Gotelli, D., C. Sillero-Zubiri, G.D. Applebaum, M.S. Roy, D.J. Girman, J. Garcia-Moreno, E.A. Ostrander, and R.K. Wayne, 1994, Molecular genetics of the most endangered canid: The Ethiopian wolf Canis simensis., Mol. Ecol. 3, 301-312.
13. Hubell, S., 2001, Shrinking the Cat: Genetic Engineering Before We Knew about Genes. Boston: Houghton Mifflin, 175.
14. Jeffreys, A.J., A.J. Allen, E. Hagelberg, and A. Sonnberg, 1991, Identification of the skeletal remains of Josef Mengele by DNA analysis. Nature, 352, 427-429.
49

15. Jung, D.S., I.J. Rhe, S.M. Lee, 1989, Classification and selection of the breeding materials in the silkworm, Bombyx mori, by multivariate analysis. Classification of the silkworm genetic stoks by principal component analysis and cuuster analysis. Miryang National Junin. Coll. Of Agriculture and Sericiculture, Miryana, Korean Journal Of Sericicultural Science (Korea Republic), 31:2, 102-112.
16. Kazan, K., J. M. Manners, and D.F. Cameron, 1993, Genetic Relationships and Variation in the Stylosanthes guianensis complex assessed by Random Amplified Polymorphic DNA, Genome, 36, 43-49.
17. Kurin, R., 2002, The Silk Road: Conecting Cultures, Creating Trust” Talk Story, Fall, Smith-sonian Center for Folklife and Cultural Heritage 21, 1-11.
18. Lehmann, P.F., D. Lin, and B.A. Lasker, 1992, Genotypic Identification of Characterization of Species and Strains within the Genus Candida by Using Random Amplified Polymorphic DNA, J. Clin. Microbiol., 30, 3249-3254.
19. Morin, P.A., J.J. Moore, R. Chakraborty, L. Jin, J. Goodall, and D.S. Woodruff, 1994, Kin selection, social structure, gene, flow and evolution of chimpanzees, Science, 265, 1193-1201.
20. Nagaraja, G.M. and J. Nagaraju, 1995, Genome fingerprinting of the silkworm, Bombyx mori, using randim arbitrary primers, Electrophoresis, 16, 1633-1638.
21. Nicese, F.P., J.I. Hormaza, and G.H. Mcgranahan, 1998, Molecular characterization and genetic relatedness among walnut (Juglans regia L.) genotypes based on RAPD markers, Euphytica 101, 199–206.
22. Ostander, E.A., J.F. Sprague, and J.Jr. Rine, 1993, Identification and characterization of dinucleotide repeat (CA)n markers for genetic mapping in dog, Genomics 16, 207-213.
23. Paşca, I., L.Al. Mărghitaş, D. Dezmirean, Laura Laslo, Georgeta Diniţă, O. Maghear, Dana Pusta, R. Morar, A. Cîmpean, R. Oroian, Claudia Bagita, 2008b, Technological features of parental breeds dry cocoon and hybrid combinations on mulberry silkworm (Bombyx mori L.) – SERISTECH, The Proceedings of the First Internatiolnal Conference „Sericulture – From Tradition to Modern Biotechnology”, U.S.A.M.V. Cluj-Napoca, Editura AcademicPres, 17 – 18 aprilie, ISBN 978-973-744-109-6, 119 – 132.
24. Prasad, D.M., M. Muthulakshmi, M. Madhu, Sunil Archak, K. Mita and J. Nagaraju, 2005a, Survey and Analysis of Microsatellites in the Silkworm, Bombyx mori: Frequency, Distribution, Mutations, Marker Potential and Their Conservation in Heterologous Species, Genetics 169, 197–214.
25. Suzuki, Y., L. Gage, and D.D. Brown, 1972, The genes for silk fibroin in Bombyx mori, J., Mol. Biol., 70, 637–649.
26. Williams, J.G. K., A.R. Kubelik, K.J. Livak, J.A. Rafalski, and S.V. Tingey, 1990, DNA Polymorphism Amplified by Arbitrary Primers Are Useful Genetic Markers, Nucl. Acids Res.,18, 653l -6535.
50