cholinergic signal transduction cascades in rat ... · teaching and research institutions in france...
TRANSCRIPT

HAL Id: hal-00900302https://hal.archives-ouvertes.fr/hal-00900302
Submitted on 1 Jan 1999
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Cholinergic signal transduction cascades in ratpinealocytes: functional and ontogenetic aspects
Christof Schomerus, Horst-Werner Korf
To cite this version:Christof Schomerus, Horst-Werner Korf. Cholinergic signal transduction cascades in rat pinealocytes:functional and ontogenetic aspects. Reproduction Nutrition Development, EDP Sciences, 1999, 39(3), pp.305-314. <hal-00900302>

Review article
Cholinergic signal transduction cascadesin rat pinealocytes: functional and ontogenetic aspects
Christof Schomerus Horst-Werner Korf
Dr. Senckenbergische Anatomie, Anatomisches Institut II,Johann Wolfgang Goethe-Universität Frankfurt, Theodor-Stern-Kai 7,
60590 Frankfurt/Main, Germany
(Received 21 January 1999; accepted 8 March 1999)
Abstract - In adult rat pinealocytes, acetylcholine activates nicotinic receptors whose stimulationcauses a depolarization of the cells, opening of voltage-gated cation channels of the L-type and sub-sequent increase in the intracellular calcium ion concentration. These events trigger a release of glu-tamate that, by its action on metabotropic glutamate type 3 receptors, activates an inhibitory cyclicAMP cascade and suppresses norepinephrine-induced melatonin biosynthesis. The nicotinic res-ponse is fully developed in the third postnatal week. Prior to this timepoint, rat pinealocytes possessfunctional muscarinic receptors whose activation causes a rise in the intracellular calcium ion concen-tration through a calcium release from thapsigargin-sensitive intracellular calcium stores and anopening of store-operated calcium channels. This cascade may influence tissue differentiation and matu-ration of the melatonin pathway. The demonstration of functional cholinoreceptors and the ontoge-netic switch from muscarinic to nicotinic signalling in rat pinealocytes supports the concept thatpineal functions in mammals are influenced by neuronal inputs other than the sympathetic innerva-tion which serves as the major regulatory system. © Inra/Elsevier, Paris
pinealocyte (rat) / nicotinic acetylcholine receptor / muscarinic acetylcholine receptor /norepinephrine / glutamate
Résumé ― Des cascades de signaux de transduction cholinergique dans les pinéalocytes durat : aspects fonctionnel et ontogénétique. Dans les pinéalocytes du rat adulte, l’acétylcholineactive des récepteurs nicotiniques dont la stimulation induit une dépolarisation cellulaire, l’ouverturedes canaux de cations voltage-dépendant du type L et l’augmentation subséquent de la concentrationd’ions calcium intracellulaires. Ces événements induisent la libération du glutamate qui, par sonaction sur des récepteurs métabotropiques de type glutamate 3, active une cascade d’inhibition del’AMP cyclique et réprime la biosynthèse de la mélatonine induite par la norépinéphrine. La réponsenicotinique est entièrement développée au cours de la troisième semaine postnatale. Avant ce moment,les pinéalocytes du rat possèdent des récepteurs fonctionnels muscariniques dont l’activation induitune hausse de la concentration intracellulaire d’ions calcium via la libération du calcium des réservesintracellulaires sensibles à la thapsigargine et l’ouverture des canaux de calcium actionnée par des
* Correspondence and reprintsE-mail: [email protected]

réserves. Cette cascade peut influencer la différentiation des tissus et la maturation des voies de syn-thèse de la mélatonine. La démonstration de l’existence de cholinorécepteurs fonctionnels et le chan-gement ontogénétique des signaux muscariniques en signaux nicotiniques dans les pinéalocytes durat soutiennent l’idée générale que les fonctions pinéales des mammifères sont influencées par des affé-rences différentes de l’innervation sympathique qui est le système principal de régulation. © Inra/
Elsevier, Paris
pinéalocyte de rat / récepteur d’acétylcholine nicotinique / récepteur de l’acétylcholine muscarinique /norépinéphrine / glutamate
1. INTRODUCTION
The mammalian pineal organ is animportant component of the photoneuroen-docrine system which rhythmically synthe-sizes and secretes melatonin during the nightin response to photoperiodic stimuli and sig-nals from the circadian clock in the hypotha-lamic suprachiasmatic nucleus. The mela-tonin rhythm generation depends on thesympathetic innervation of the pineal gland[12] which rhythmically releases nor-epinephrine (NE) at the onset of darkness[6]. By stimulating (X¡- and P,-adrenergicreceptors, NE causes increases in the intra-cellular concentrations of calcium ions and
cyclic AMP and shapes the melatoninrhythm by regulating the arylalkylamine-N-acetyltransferase (AANAT), the keyenzyme of the melatonin biosynthesis [14],at transcriptional and post-transcriptionallevels. The transcriptional regulation of theAANAT involves activating and inhibitorytranscription factors, e.g. phosphorylatedCREB and ICER [17, 21, 25, 30, 32-34]; amajor post-transcriptional mechanism is thecyclic AMP-dependent rapid and reversiblecontrol of selective proteasomal proteoly-sis [9]. All these data have corroborated theessential role of the sympathetic innerva-tion of the pineal organ and its primary neu-rotransmitter, NE. In contrast, it is stillunclear whether and how non-sympatheticneuronal pathways (see [ 15, 16, 23]) or hor-mones are involved in the regulation ofmelatonin biosynthesis and pineal functionin mammals.
Out of the variety of putative neuronaland hormonal inputs to the mammalianpineal organ, the cholinergic system appearsof particular interest for the following rea-sons. 1) Several morphological investiga-tions point towards the existence of aparasympathetic innervation of the mam-malian pineal gland (for review, see [23])which may originate from the pterygopala-tine ganglion, employ acetylcholine (ACh)as the primary neurotransmitter and antag-onize the sympathetic effects. 2) A cholin-ergic innervation of the mammalian pinealorgan has been convincingly demonstratedby immunocytochemistry using antibodiesagainst the vesicular ACh transporter [26].3) Biochemical and immunocytochemicalresults suggest that rat pinealocytes containACh and that the ACh content increases ten-fold at night [37].
Possible effects of cholinergic agonistson melatonin production and release havebeen repeatedly investigated (for review,see [19]), but the data are equivocal withregard to the receptor types involved, theirlocation and the functional consequences oftheir activation. Transpineal in vivo micro-dialysis has shown that the infusion of thecholinergic agonists carbachol or oxotrem-orine into the pineal organ of adult ratsresulted in a marked decrease in melatoninrelease during the dark period by inhibitingthe NE release from sympathetic nervefibres [7]. Such data suggest the presenceof muscarinic acetylcholine receptors(mAChRs) in a presynaptic location, i.e. on

the sympathetic nerve endings. The exis-tence of nicotinic ACh receptors (nAChRs)in the rat pineal organ was inferred fromimmunocytochemical investigations [24]and binding studies with radiolabelled spe-cific ligands [31] and it has been suggestedthat nicotine has an inhibitory effect onpineal melatonin biosynthesis.
In the last 3 years, several investigationshave been performed in an attempt to clar-ify the cholinergic signal transduction mech-anisms, using the rat pineal organ as amodel. The results of these studies will bereviewed in the present contribution. More-over, new data will be presented on the onto-genetic development of cholinergic sig-nalling mechanisms in rat pinealocytes.
2. CHOLINERGIC SIGNALTRANSDUCTIONIN PINEALOCYTES OF ADULTRATS
Calcium imaging of isolated, immuno-cytochemically identified rat pinealocyteshas shown that ACh increases the intracel-lular calcium ion concentration ([Ca 2+] inmore than 90 % of the cells in a dose-depen-dent manner [29]. All ACh-sensitivepinealocytes also respond to NE with anincrease in [Ca2+];, but the two types ofresponses are quite different. The ACh-induced rise in [Ca2+]¡ is followed by a rapiddecrease to basal levels within a few minutesafter the onset of the stimulus. This decreaseis also seen under a constant exposure tothe ligand. The response is mediated by anicotinic receptor subtype (figure lA)because ACh and nicotine elicit virtuallyidentical effects, and both the ACh- and thenicotine-induced responses are blocked bythe specific nicotinic antagonist, d-tubocu-rarine. mAChRs do not play a role in thiscalcium response because pilocarpine, act-ing upon all presently known muscarinicreceptor subtypes, does not evoke a calciumresponse, and the muscarinic antagonistatropine does not block the ACh-induced
rise in [Ca2+]i. The response to ACh is totallyprevented when pinealocytes were kept incalcium-free saline, indicating that theresponse is based upon a calcium influx.
The components involved in the nico-tinic response have been identified using acombination of patch-clamp recordings andcalcium imaging [20]. These investigationshave shown that the average resting mem-brane voltage of isolated adult rat pinealo-cytes is -43 mV, and that the replacement ofextracellular NaCI by KCI completely depo-larizes the cells. This indicates that the rest-
ing membrane voltage is dominated by a K+ +
conductance. Single channel recordingsreveal the presence of a large conductance,Ca2+-activated, charybdotoxin-sensitive K+channel [4, 20]. The application of AChdepolarizes the pinealocytes by an averageof 16 mV. The depolarizing effect of ACh ismimicked by nicotine and prevented bytubocurarine. This depolarization is largelyabolished in the absence of extracellular Na+but is not significantly affected by extra-cellular Caz+ removal. Removal of extra-cellular Na+ also causes a large reductionin the ACh-induced rise in [Ca2+]i. Nifedip-ine suppresses the ACh-induced increase in
[Caz+]! by approximately 50 %. The find-ings indicate that ACh influences adult ratpinealocytes through stimulation of nAChRswhich induces a depolarization mainly bya (3Na+ influx through the receptor. Thedepolarization then activates voltage-gateL-type calcium channels (VCCs) which areresponsible for the nifedipine-sensitive por-tion of the [Ca2+]i increase.
The fact that nicotinic receptors andVCCs are present in the vast majority ofadult rat pinealocytes suggests their impor-tant role in the regulation of pinealmetabolism. One idea is that ACh may act
upon microvesicle-mediated glutamaterelease from pinealocytes which is elicitedby depolarization and activation of VCCs([39, 40]; figure 2A). Glutamate has beenshown to suppress the NE-induced mela-tonin production in the rat pineal organ kept

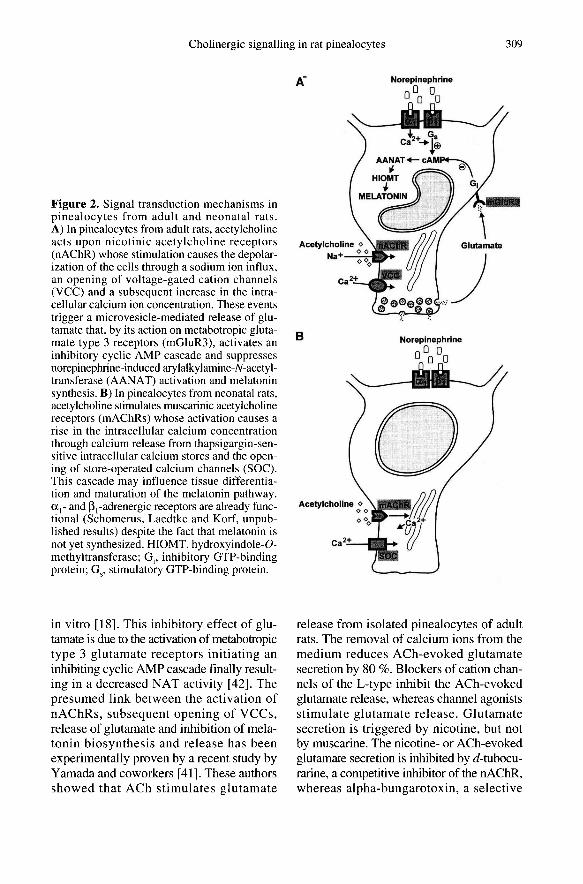
in vitro [18]. This inhibitory effect of glu-tamate is due to the activation of metabotropictype 3 glutamate receptors initiating aninhibiting cyclic AMP cascade finally result-ing in a decreased NAT activity [42]. Thepresumed link between the activation ofnAChRs, subsequent opening of VCCs,release of glutamate and inhibition of mela-tonin biosynthesis and release has beenexperimentally proven by a recent study byYamada and coworkers [41]. These authorsshowed that ACh stimulates glutamate
release from isolated pinealocytes of adultrats. The removal of calcium ions from themedium reduces ACh-evoked glutamatesecretion by 80 %. Blockers of cation chan-nels of the L-type inhibit the ACh-evokedglutamate release, whereas channel agonistsstimulate glutamate release. Glutamatesecretion is triggered by nicotine, but notby muscarine. The nicotine- or ACh-evokedglutamate secretion is inhibited by d-tubocu-rarine, a competitive inhibitor of the nAChR,whereas alpha-bungarotoxin, a selective

inhibitor of some subtypes of nAChR andthe muscarinic antagonists scopolamine andatropine are ineffective. These data showthat an alpha-bungarotoxin-insensitivenAChR is responsible for glutamate secre-tion from adult rat pinealocytes. Both nico-tine and ACh also strongly inhibit the NE-induced activation of AANAT and melatonin
biosynthesis. This inhibition is preventedby tubocurarine and a specific antagonist ofthe class II metabotropic-type glutamatereceptor.
The precise mechanism through whichACh inhibits NE-induced AANAT activa-tion remains to be elucidated. As mentionedabove AANAT can be regulated at the tran-scriptional and post-transcriptional level.The transcriptional regulation involves phos-phorylation of the activating transcriptionfactor which, however, is not affected byACh, nicotine or L-glutamate (figure 3A,B). These results suggest that the choliner-gic effects on AANAT activity are exertedat the post-transcriptional rather than thetranscriptional level.
3. ONTOGENETIC DEVELOPMENTOF CHOLINERGIC SIGNALTRANSDUCTIONIN RAT PINEALOCYTES
To study the ontogenetic developmentof cholinergic signal transduction mecha-nisms we have investigated pinealocytesisolated from newborn, 1-, 2-, 3-, 4-, 5, 7-,10-, 14- and 21-day-old rats. ACh at a con-centration of > 10! M induces a robust bi-
phasic rise in [Ca2+]¡ in approximately 90 %of neonatal pinealocytes (PO). A transientmaximum in [Ca2+]! is followed by a sus-tained elevation of [Ca2+]! which finallydrops to basal levels upon removal of thestimulus. Calibration of the semiquantita-tive ratio data revealed that, after ACh stim-ulation, [Ca2+]¡ increases from a basal levelof approximately 40 ± 10 nM to a maxi-mum value of approximately 400 ± 150 nM.The ACh-responsive cells also react to NE
with a rise in [Ca2+]¡. Like ACh, the mus-carinic agonists muscarine (non-selective)and oxotremorine-M (relatively M1-selec-tive) and the non-specific cholinergic agonistcarbachol elicit a calcium signal in 90 % ofthe cells at concentrations of > 10-6 M. Incontrast, nicotine (10! M) induces a weakcalcium signal in less than 10 % of cellsonly (figure IB). The muscarinic antago-nists atropine (non-selective) and piren-zepine (M1-selective) totally block the ACh-evoked calcium signal at a concentration of>_ 10! and> la-7M, respectively. In contrast,the nicotinic antagonist d-tubocurarine(10! M) only partially inhibits the ACh-evoked rise in [Ca 2,]i. In the second post-natal week, the percentage of cells whichclearly respond to 10! M nicotine with arise in [Ca2+]¡ increases to approximately10 %. These cells usually also react to AChand oxotremorine-M at concentrations of> 10-6 M. After the second postnatal week,the percentage of cells responding to mus-carinergic stimuli decreases from 90 % toapproximately 50 %. By the end of the thirdpostnatal week oxotremorine and muscarineat concentrations of up to 10! M are basi-
cally without effect in most pinealocytes. Incontrast, a robust calcium signal is elicited in90 % of the cells by ACh and nicotine whenapplied at 10-4 M. This type of pinealocyte isprevailing in adult rats (figure ]A).
These findings show that the cholinergicreceptor types in rat pinealocytes undergoa striking transformation from mAChRs inneonatal animals to nAChRs in adult rats.
They conform to binding studies showingthat the amount of muscarinic ACh bindingsites in the rat pineal organ declines in thecourse of ontogeny from relatively high lev-els early in development [27, 28] to low lev-els in adulthood [8, 19, 35]. ].
The developmental switch of the
cholinoceptor type of rat pinealocytes is par-alleled by a change in the signalling cas-cade distal to the receptors. As mentionedabove, the nicotinic calcium response ofadult rat pinealocytes involves the induc-

tion of a depolarizing Na+ influx and the
subsequent activation of VCCs leading toa transient rise in [Ca2+]¡ which is followed
by a gradual drop to basal levels in the pres-ence of the stimulus [20]. The muscariniccalcium response in neonatal pinealocytesconsists of a primary phase mainly associ-ated with the mobilization of Ca2+ from
thapsigargin-sensitive intracellular storesand a secondary phase associated with Ca2+entry into the cell (figure 2B). Interestingly,these thapsigargin-sensitive calcium storesalso contribute to the calcium signal elicitedby stimulation of <Xl-adrenergic receptors.When neonatal pinealocytes are kept inCa2+-free saline and pretreated with NE,they exhibit a transient increase in [Ca 2+1,due to the release of calcium from intracel-lular compartments, but do not respond tosubsequent stimulation with ACh in cal-cium-free saline. Vice versa, the depletion ofthese stores by ACh application preventsthe response to NE. The calcium channels
mediating the influx of Ca2+ in the secondaryphase of the muscarinic response have notyet been identified and it remains to beestablished whether the calcium entry mech-anism in the secondary phase of the mus-carinic calcium signal is identical to thatmediating the calcium influx in response toNE stimulation.
VCCs are apparently not yet developed inneonatal pinealocytes since treatment withdepolarizing concentrations of KCI (figureIB) or the agonist Bay K 8644 is withoutan effect on [Ca2+]¡. Accordingly, specificL-type channel antagonists (nifedipine, ver-apamil) do not affect the ACh-evoked, mus-carinic calcium signalling.
The transformation of the cholinergic sig-nalling system during the development ofrat pinealocytes strikingly changes the spa-tial and temporal patterns of the ACh-evoked calcium response. Since these pat-terns are considered important determinantswhich enhance the flexibility of Ca2+ to reg-ulate diverse cellular processes [2, 5, 10,22], the biphasic response evoked via

mAChRs in neonatal pinealocytes is suitedto promote the activation of Ca2+-sensitiveevents different from those activated vianAChRs in adult pinealocytes. A directimpact of muscarinic signalling on the reg-ulation of melatonin production can beexcluded since melatonin is not yet synthe-sized in the first postnatal week [13]. Morelikely, activation of mAChRs may play aregulatory role in the development of thepineal gland, in general, and in the matura-tion of the melatonin-generating system, inparticular. The activation of mAChRs hasbeen shown to promote cellular prolifera-tion in neuronal and non-neuronal cells [1,11]. In the retina, signalling via mAChRshas been proposed to be related to morpho-genesis [43] and proliferation [38]. Inter-estingly, retinal cells also appear to expressdifferent types of cholinergic receptors dur-ing development which, it is proposed, havediverse and temporally regulated roles intheir differentiation [38]. Similar differen-tiation processes may be controlled viamAChRs in the developing pineal organwhich shares the diencephalic origin as wellas photoreceptive and phototransducingproperties with the retina [17]. The mus-carinic response disappears concomitantlywith the completion of mitosis [36]. Thisraises the possibility that the stimulation ofmAChRs may also promote cell division inthe pineal gland. Alternatively, muscariniccholinergic signalling may be related to theexpression of phototransduction moleculesmany of which are present at high levels inneonatal rat pineal organs and at low levelsin adult rat pineal organs [3]. ] .
Pinealocytes lose the capacity to respondto muscarinic stimuli by the third postnatalweek when the rhythmic melatonin synthe-sis is fully developed. The loss of the ’mus-carinic phenotype’ may be caused by adecrease in receptor number, decreasedaffinity between receptor and ligand, and/oractions distal from the mAChRs. Concomi-
tantly, pinealocytes gain responsiveness toACh via nAChRs. The factors responsiblefor the differential maturation of cholinergic
calcium signalling mechanisms remainunclear. The switch is obviously not causedby cell death of those pinealocytes whichare endowed with a muscarinic signallingcascade since the decrease in sensitivity toACh and muscarinic agonists is paralleledby an increase in responsiveness to nicotinein one and the same cell. Interestingly, thefunctional maturation of VCCs precedes thedevelopment of a calcium response medi-ated by nAChRs. This raises the interestinghypothesis of whether the development ofVCCs may induce the maturation of the
’proximal’ nicotinic receptor.
4. CONCLUSIONS
Cholinergic signal transduction cascadesoperate in rat pinealocytes at all postnatalstages. In adult pinealocytes ACh acts uponnAChRs whose stimulation causes depo-larization of the cells, opening of VCCs anda subsequent increase in the intracellularcalcium ion concentration [20, 29]. Theseevents trigger the release of glutamate that,by its action on metabotropic glutamate type3 receptors, activates an inhibitory cyclicAMP cascade and suppresses NE-inducedNAT activation and melatonin synthesis[41, 42]. In adult rats, mAChRs are absentfrom the pinealocyte membrane, but are pre-sent on sympathetic nerve fibres. Activa-tion of these receptors blocks NE releasefrom sympathetic nerve terminals, thus lead-ing to a decreased melatonin output [7].Thus, ACh appears to employ two differ-ent pathways to inhibit NE-induced mela-tonin synthesis and release in adult rats. Thenicotinic response of rat pinealocytes is fullydeveloped in the third postnatal week. Priorto this timepoint, rat pinealocytes expressfunctional mAChRs whose activation causesa rise in the intracellular calcium ion con-centration through a calcium release fromthapsigargin-sensitive intracellular calciumstores and the opening of store-operated cal-cium channels. The functional significanceof the muscarinic signal transduction cas-

cade in pinealocytes during early postnataldevelopment remains to be clarified. It maybe assumed that this cascade influences tis-sue differentiation and maturation of themelatonin pathway. The demonstration offunctional cholinoreceptors and the onto-genetic switch from muscarinic to nicotinicsignalling in rat pinealocytes supports theconcept that pineal functions in mammalsare modulated by a variety of inputs thatmay fine-tune the signals from the majorregulatory system, the sympathetic inner-vation.
ACKNOWLEDGEMENTS
This investigation has been supported by theDeutsche Forschungsgemeinschaft (SFB 269,Teilprojekt B2). The authors are grateful to MrsE. Laedke for expert technical assistance andMrs I. Szasz for excellent graphical work.
REFERENCES
[1] ] Ashkenazi A., Ramachandran J., Capon D.J.,Acetylcholine analogue stimulates DNA syn-thesis in brain-derived cells via specific mus-carinic receptor subtypes, Nature 340 (1989)146-150.
[2] Berridge M.J., The AM and FM of calcium sig-nalling, Nature 386 (1997) 759-760.
[3] Blackshaw S., Snyder S.H., Developmentalexpression pattern of phototransduction com-ponents in mammalian pineal implies a light-sensing function, J. Neurosci. 17 (1997)8074-8082.
[4] Cena V., Halperin J.L, Yeandle S., Klein D.C.,Norepinephrine stimulates potassium efflux frompinealocytes: evidence for involvement of bio-chemical ’AND’ gate operated by calcium andadenosine 3’,5’-monophosphate, Endocrinol-ogy 128 (1991) 559-569.
[5] Clapham D.E., Calcium signalling, Cell 80(1995) 259-268.
[6] Drijfhout W.J., van der Linde A.G., Kooi S.E.,Grol C.J., Westerink B.H.C., Norepinephrinerelease in the rat pineal gland: the input fromthe biological clock measured by in vivo micro-dialysis, J. Neurochem. 66 (1996) 748-755.
[7] Drijfhout W.J., Grol C.J., Westerink B.H.C.,Parasympathetic inhibition of pineal indolemetabolism by prejunctional modulation of nora-drenergic release, Eur. J. Pharmacol. 308 (1996)117-124.
[8] Finocchiaro L.M.F., Scheucher A., Finkielman S.,Nahmod V.E., Pirola C.J., Muscarinic effectson the hydroxy- and methoxyindole pathway inthe rat pineal gland, J. Endocrinol. 123 (1989)205-211. 1 .
[9] Gastel J.A., Roseboom P.H., Rinaldi P.A.,Weller J.L., Klein D.C., Melatonin production:proteosomal proteolysis in serotonin N-acetyl-transferase regulation, Science 279 (1998)1358-1360.
[10] Ginty D.D., Calcium regulation of gene expres-sion: Isn’t that spatial?, Neuron 18 (1997)I 83-186.
[11] Gutkind J.S., Novotny E.A., Brann M.R., Rob-bins K.C., Muscarinic acetylcholine receptorsubtypes as agonist-dependent oncogenes, Proc.Natl. Acad. Sci. USA 88 (1991) 4703-4707.
[ 12] Klein D.C., Photoneural regulation of the mam-malian pineal gland, in: Evered D., Clark S.(Eds), Photoperiodism, Melatonin and the PinealGland, Ciba Foundation Symposium 117, Pit-man, London, 1985, pp. 38-56.
[13] Klein D.C., Namboodiri M.A.A., Auerbach D.A.,The melatonin rhythm generating system: devel-opmental aspects, Life Sci. 28 (1981) 1975-1986.
[14] Klein D.C., Coon S.L., Roseboom P.H.,Weller J.L., Bernard M., Gastel J.A., Zatz M.,luvone P.M., Rodriguez I.R., Begay V., Falcon J.,Cahill G.M., Cassone V.M., Baler R., The mela-tonin rhythm-generating enzyme: molecular reg-ulation of serotonin N-acetyltransferase in thepineal gland, Rec. Progr. Hormone Res. 52(1997) 307-358.
[IS] Korf H.W., Innervation of the pineal gland, in:Burnstock G., Unsicker K. (Ed.), Series on theAutonomous Nervous System, Vol. 10, Auto-nomic-Endocrine Interactions, Harwood, Ams-terdam, 1996, pp. 129-180.
[16] Korf H.W., Moller M., The innervation of themammalian pineal gland with special referenceto central pinealopetal projections, Pineal Res.Rev. 2 ( 1984) 41-86.
[17] Korf H.W., Schomerus C., Stehle J.H., Thepineal organ, its hormone melatonin, and thephotoneuroendocrine system, Adv. Anat.Embryol. Cell Biol. 146, Springer-Verlag Berlin,Heidelberg, 1998.
[ l8] Kus L., Handa R.J., McNulty J.A., Glutamateinhibition of the adrenergic-stimulated produc-tion of melatonin in rat pineal gland in vitro,J. Neurochem. 62 (1994) 2241-2245.
[ 19J Laitinen J.T., Laitinen K.S.M., Kokkola T.,Cholinergic signalling in the rat pineal gland,Cell. Mol. Neurobiol. 15 (1995) 177-192.
[20] Letz B., Schomerus C., Maronde E., Korf H.W.,Korbmacher C., Stimulation of a nicotinic AChreceptor causes depolarization and activation ofL-type Ca2’ channels in rat pinealocytes, J. Phys-iol. 499 (1997) 329-340.

[21] Maronde E., Schomerus C., Stehle J.H.,Korf H.W., Control of CREB phosphorylationand its role for induction of melatonin synthesisin rat pinealocytes, Biol. Cell 89 (1997)505-511.
[22] Meldolesi J., Oscillation, activation, expression,Nature 392 (1998) 863-865.
[23] Moller M., Fine structure of the pinealopetalinnervation of the mammalian pineal gland,Microsc. Res. Tech. 21 (1992) 188-204.
[24] Reuss S., Schrbder H., Maelicke A., Nicotiniccholinoceptors in the rat pineal gland as ana-lyzed by western blot, light- and electronmicroscopy, Brain Res. 573 (1992) 114-1 18.
[25] Roseboom P.H., Klein, D.C., Norepinephrinestimulation of pineal cyclic AMP response ele-ment-binding protein phosphorylation: involve-ment of a (3-adrenergic/cyclic AMP mechanism,Mol. Pharmacol. 47 (1995) 439!49.
[26] SchAfer M.K.H., Eiden L.E., Weihe E., Cholin-ergic neurons and terminal fields revealed byimmunohistochemistry for the vesicular acetyl-choline transporter. I. Central nervous system,Neuroscience 84 (1998) 331-359.
[27] Schlumpf M., Bruinink A., Lichtensteiger W.,Cort6s R., Palacios J.M., Pazos A., Beta adren-ergic binding sites in fetal rat central nervoussystem and pineal gland: their relation to otherreceptor sites, Dev. Pharmacol. Ther. 10 (1987)422-435.
[28] Schlumpf M., Palacios J.M., Cort6s R., Licht-ensteiger W., Regional development of mus-carinic cholinergic binding sites in the prenatalrat brain, Neuroscience 45 (1991) 347-357.
[29] Schomerus C., Laedtke E., Korf H.W., Calciumresponses of isolated, immunocytochemicallyidentified rat pinealocytes to noradrenergic,cholinergic and vasopressinergic stimulations,Neurochem. Int. 27 (1995) 163-175.
[30] Schomerus C., Maronde E., Laedtke E.,Korf H.W., Vasoactive intestinal peptide (VIP)and pituitary adenylate cyclase-activatingpolypeptide (PACAP) induce phosphorylation ofthe transcriptional factor CREB in subpopula-tions of rat pinealocytes: immunocytochemicaland immunochemical evidence, Cell Tissue Res.286 (1996) 305-313.
[31] Stankov B., Cimino M., Marini P., Lucini V.,Fraschini F., Clementi F., Identification andfunctional significance of nicotinic cholinergicreceptors in the rat pineal gland, Neurosci. Lett.156 (1993) 131-134.
[32] Stehle J.H., Pineal gene expression: dawn in adark matter, J. Pineal Res. 18 (1995) 179-190.
[33] Stehle J.H., Foulkes N.S., Molina C.A., Simon-neaux V., Pevet P., Sassone-Corsi P., Adrener-gic signals direct rhythmic expression of tran-scriptional repressor CREM in the pineal gland,Nature 356 (1993) 314-320.
[34] Tamotsu S., Schomerus C., Stehle J.H., Rose-boom P.H., Korf H.W., Norepinephrine-inducedphosphorylation of the transcription factorCREB in isolated rat pinealocytes: an immuno-cytochemical study, Cell Tissue Res. 282 (1995)219-226.
[35] Taylor R.L., Albuquerque M.L.C., Burt D.R.,Muscarinic receptors in pineal, Life Sci. 26(1980)2195-2200.
[36] Vollrath L., The pineal organ, in: Oksche A.,Vollrath L. (Eds.), Handbuch der Mikroskopis-chen Anatomie des Menschen, vol. VI/7,Springer, Berlin, 198L. 1 .
[37] Wessler L, Reinheimer T., Bittinger F., Kirk-patrick C.J., Schenda J., Vollrath L., Day-nightrhythm of acetylcholine in the rat pineal gland,Neurosci. Lett. 224 (1997) 173-176.
[38] Wong R.O.L., Cholinergic regulation of [Ca2+J;during cell division and differentiation in themammalian retina, J. Neurosci. 15 (1995)2696-2705.
[39] Yamada H., Yamamoto A., Takahashi M.,Michibata H., Kumon H., Moriyama Y., TheL-type Ca2’ channel is involved in microvesicle-mediated glutamate exocytosis from rat pinealo-cytes, J. Pineal Res. 21 (1996) 165-174.
[40] Yamada H., Yamamoto A., Yodozawa S.,Kozaki S., Takahashi M, Morita M., Michibata H.,Furuichi T., Mikoshiba K., Moriyama Y.,Microvesicle-mediated exocytosis of glutamateis a novel paracrine-like chemical transductionmechanism and inhibits melatonin secretion inrat pinealocytes, J. Pineal Res. 21 (1996)175-191. 1 .
[41 ] Yamada H., Ogura A., Koizumi S., Yamaguchi A.,Moriyama Y., Acetylcholine triggers L-gluta-mate exocytosis via nicotinic receptors andinhibits melatonin synthesis in rat pinealocytes,J. Neurosci. 18 (1998) 4946-4952.
[42] Yamada H., Yastushiro S., Ishio S., Hayashi M.,Nishi T., Yamamoto A., Futai M., Yamaguchi A.,Moriyama Y., Metabotropic glutamate recep-tors negatively regulate melatonin synthesis inrat pinealocytes, J. Neurosci. 18 (1998)2056-2062.
[43] Yamashita M., Fukuda Y., Incurvation of earlyembryonic neural retina by acetylcholine throughmuscarinic receptors, Neurosci. Lett. 163 (1993)215-218.