cipres: enabling tree of life projects
DESCRIPTION
CIPRES: Enabling Tree of Life Projects. Tandy Warnow The University of Texas at Austin. Reconstructing the “Tree” of Life. Handling large datasets: millions of species. The “Tree of Life” is not really a tree: reticulate evolution. Cyber Infrastructure for Phylogenetic Research - PowerPoint PPT PresentationTRANSCRIPT

CIPRES: Enabling Tree of Life Projects
Tandy Warnow
The University of Texas at Austin

Reconstructing the “Tree” of LifeHandling large datasets: Handling large datasets:
millions of speciesmillions of species
The “Tree of Life” is not The “Tree of Life” is not really a tree: really a tree:
reticulate evolutionreticulate evolution

Cyber Infrastructure for Phylogenetic Research
Purpose: to create a national infrastructure of hardware, open source software, database technology, etc., necessary to infer the Tree of Life. Group: 40 biologists, computer scientists, and mathematicians from 13 institutions.Funding: $11.6 M (large ITR grant from NSF).URL: http://www.phylo.org
CIPRes Members
University of New MexicoBernard Moret David Bader
UCSD/SDSCFran Berman Alex Borchers Phil Bourne John Huelsenbeck Terri LiebowitzMark Miller University of ConnecticutPaul O Lewis
University of PennsylvaniaJunhyong Kim Susan Davidson Sampath KannanVal Tannen
Texas A&MTiffani Williams
UT Austin Tandy Warnow David M. Hillis Warren Hunt Robert Jansen Randy Linder Lauren Meyers Daniel Miranker
University of ArizonaDavid R. Maddison
University of British ColumbiaWayne Maddison
North Carolina State UniversitySpencer Muse
American Museum of Natural HistoryWard C. Wheeler
NJITUsman Roshan
UC BerkeleySatish Rao Steve EvansRichard M Karp Brent MishlerElchanan MosselEugene W. MyersChristos M. PapadimitriouStuart J. Russell
Rice Luay Nakhleh
SUNY BuffaloWilliam Piel
Florida State UniversityDavid L. SwoffordMark Holder
Yale Michael DonoghuePaul Turner

CIPRES activity
• Databases - e.g. TreeBase II (Bill Piel and others)• Simulations of large-scale complex genome-scale
evolution (Junhyong Kim)• Outreach (Michael Donoghue and Brent Mishler)• Algorithms (Tandy Warnow)• Open source software (Wayne Maddison, Dave
Swofford, Mark Holder, and Bernard Moret)• Computer cluster at SDSC (Fran Berman and Mark
Miller) - available to ATOL projects and other groups with datasets above 1000 taxa

CIPRES research in algorithms
• Multiple sequence alignment
• Genomic alignment
• Heuristics for Maximum Parsimony and Maximum Likelihood
• Bayesian MCMC methods
• Supertree methods
• Whole genome phylogeny reconstruction
• Reticulate evolution detection and reconstruction
• Data mining on sets of trees, and compact representations of these sets

Software distributions
The first distribution (in the next months) will focus on Rec-I-DCM3(PAUP*): fast heuristic searches for maximum parsimony on large datasets for PAUP* users
All software will be open source
Community contributions to software will be enabled
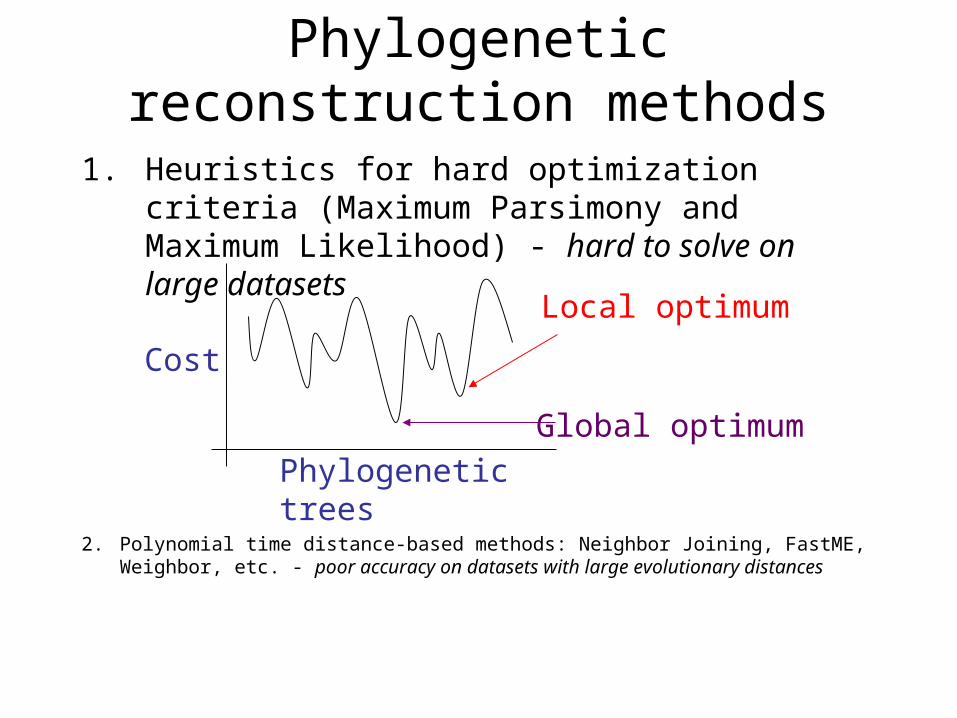
1. Heuristics for hard optimization criteria (Maximum Parsimony and Maximum Likelihood) - hard to solve on large datasets
Phylogenetic reconstruction methods
Phylogenetic trees
Cost
Global optimum
Local optimum
2. Polynomial time distance-based methods: Neighbor Joining, FastME, Weighbor, etc. - poor accuracy on datasets with large evolutionary distances

DCMs: Divide-and-conquer for improving phylogeny reconstruction

“Boosting” phylogeny reconstruction methods
• DCMs “boost” the performance of phylogeny reconstruction methods.
DCMBase method M DCM-M

DCMs (Disk-Covering Methods)
• DCMs for polynomial time methods improve topological accuracy (empirical observation), and have provable theoretical guarantees under Markov models of evolution
• DCMs for hard optimization problems reduce running time needed to achieve good levels of accuracy (empirically observation)

DCM1-boosting distance-based methods[Nakhleh et al. ISMB 2001]
•DCM1-boosting makes distance-based methods more accurate
•Theoretical guarantees that DCM1-NJ converges to the true tree from polynomial length sequences
NJ
DCM1-NJ
0 400 800 16001200No. Taxa
0
0.2
0.4
0.6
0.8
Err
or R
ate

Major challenge: MP and ML
• Maximum Parsimony (MP) and Maximum Likelihood (ML) remain the methods of choice for most systematists
• The main challenge here is to make it possible to obtain good solutions to MP or ML in reasonable time periods on large datasets

Solving NP-hard problems exactly is … unlikely
• Number of (unrooted) binary trees on n leaves is (2n-5)!!
• If each tree on 1000 taxa could be analyzed in 0.001 seconds, we would find the best tree in
2890 millennia
#leaves #trees
4 3
5 15
6 105
7 945
8 10395
9 135135
10 2027025
20 2.2 x 1020
100 4.5 x 10190
1000 2.7 x 102900

How good an MP analysis do we need?
• Our research shows that we need to get within 0.01% of optimal (or better even, on large datasets) to return reasonable estimates of the true tree’s “topology”

Problems with current techniques for MP
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2
0 4 8 12 16 20 24
Hours
Average MP score above
optimal, shown as a percentage of
the optimal
Shown here is the performance of a heuristic maximum parsimony analysis on a real dataset of almost 14,000 sequences. (“Optimal” here means best score to date, using any method for any amount of time.) Acceptable error is below 0.01%.
Performance of TNT with time

Observations
• The best MP heuristics cannot get acceptably good solutions within 24 hours on most of these large datasets.
• Datasets of these sizes may need months (or years) of further analysis to reach reasonable solutions.
• Apparent convergence can be misleading.

Our objective: speed up the best MP heuristics
Time
MP scoreof best trees
Performance of hill-climbing heuristic
Desired Performance
Fake study

DCM3 decomposition
Input: Set S of sequences, and guide-tree T
1. Compute short subtree graph G(S,T), based upon T
2. Find clique separator in the graph G(S,T) and form subproblems
DCM3 decompositions (1) can be obtained in O(n) time(2) yield small subproblems(3) can be used iteratively(4) can be applied recursively

Iterative-DCM3
T
T’
Base methodDCM3

New DCMs
• DCM31. Compute subproblems using DCM3 decomposition
2. Apply base method to each subproblem to yield subtrees
3. Merge subtrees using the Strict Consensus Merger technique
4. Randomly refine to make it binary
• Recursive-DCM3• Iterative DCM3
1. Compute a DCM3 tree
2. Perform local search and go to step 1
• Recursive-Iterative DCM3

Rec-I-DCM3 significantly improves performance
Comparison of TNT to Rec-I-DCM3(TNT) on one large dataset
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2
0 4 8 12 16 20 24
Hours
Average MP score above
optimal, shown as a percentage of
the optimal
Current best techniques
DCM boosted version of best techniques

Datasets
• 1322 lsu rRNA of all organisms• 2000 Eukaryotic rRNA• 2594 rbcL DNA• 4583 Actinobacteria 16s rRNA • 6590 ssu rRNA of all Eukaryotes• 7180 three-domain rRNA• 7322 Firmicutes bacteria 16s rRNA• 8506 three-domain+2org rRNA• 11361 ssu rRNA of all Bacteria• 13921 Proteobacteria 16s rRNA
Obtained from various researchers and online databases

Rec-I-DCM3(TNT) vs. TNT(Comparison of scores at 24 hours)
Base method is the default TNT technique, the current best method for MP. Rec-I-DCM3 significantly improves upon the unboosted TNT by returning trees which are at most 0.01% above optimal on most datasets.
00.010.020.030.040.050.060.070.080.090.1
Average MP score above
optimal at 24 hours, shown as a
percentage of the optimal
1 2 3 4 5 6 7 8 9 10
Dataset#
TNT Rec-I-DCM3

Observations
• Rec-I-DCM3 improves upon the best performing heuristics for MP.
• The improvement increases with the difficulty of the dataset.

DCMs• DCM for NJ and other distance methods produces
absolute fast converging (afc) methods• DCMs for MP heuristics • DCMs for use with the GRAPPA software for whole
genome phylogenetic analysis; these have been shown to let GRAPPA scale from its maximum of about 15-20 genomes to 1000 genomes.
• Current projects: DCM development for maximum likelihood and multiple sequence alignment.

Part II: Whole-Genome Phylogenetics
A
B
C
D
E
F
X
Y
ZW
A
B
C
D
E
F
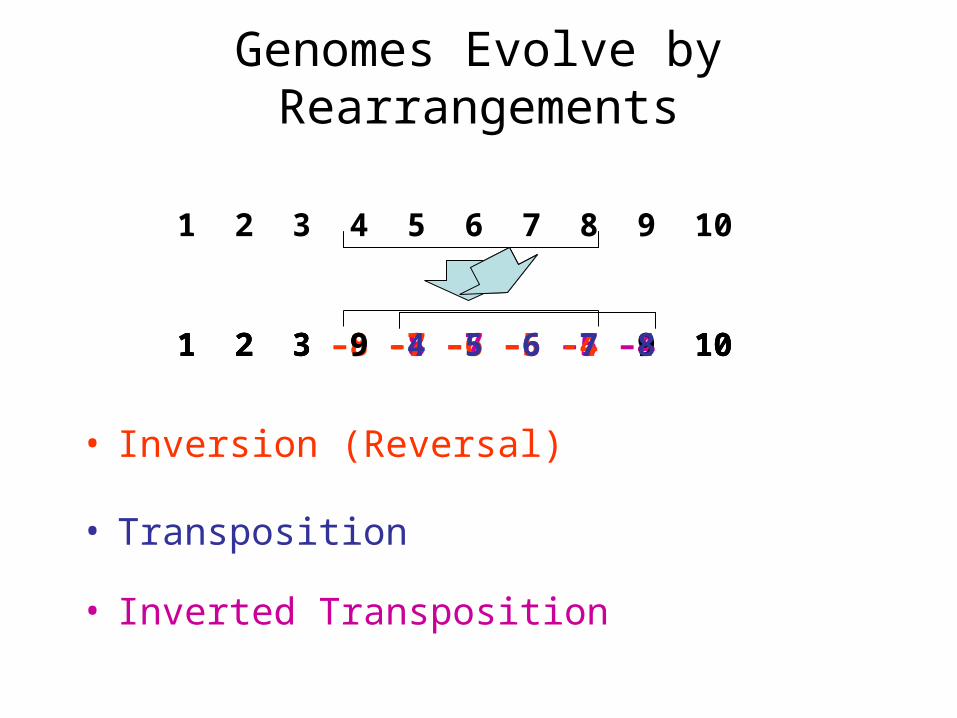
Genomes Evolve by Rearrangements
• Inverted Transposition
1 2 3 9 -8 –7 –6 –5 –4 10
1 2 3 4 5 6 7 8 9 10
• Inversion (Reversal)
1 2 3 –8 –7 –6 –5 -4 9 10
• Transposition
1 2 3 9 4 5 6 7 8 10

Genome Rearrangement Has A Huge State Space
• DNA sequences : 4 states per site• Signed circular genomes with n genes:
states, 1 site
• Circular genomes (1 site)
– with 37 genes: states
– with 120 genes: states
)!1(2 1 −− nn
521056.2 ×2321070.3 ×

Why use gene orders?
• “Rare genomic changes”: huge state space and relative infrequency of events (compared to site substitutions) could make the inference of deep evolution easier, or more accurate.
• Our research shows this is true, but accurate analysis of gene order data is computationally very intensive!

Maximum Parsimony on Rearranged Genomes (MPRG)
• The leaves are rearranged genomes.• Find the tree that minimizes the total number of rearrangement events (NP-hard)
A
B
C
D
3 6
2
3
4
A
B
C
D
EF
Total length= 18

“Solving” the inversion phylogeny
Phylogenetic trees
MP score
Global optimum
Local optimum
• Usual issue of getting stuck in local optima, since the optimization problems are NP-hard
• Additional problem: finding the best trees is enormously hard, since even the “point estimation” problem is hard (worse than estimating branch lengths in ML).

Benchmark gene order dataset: Campanulaceae
• 12 genomes + 1 outgroup (Tobacco), 105 gene segments• NP-hard optimization problems: breakpoint and inversion phylogenies
(techniques score every tree)Joint work with Bob Jansen, Linda Raubeson, Jijun Tang, and Li-San Wang
1997: BPAnalysis (Blanchette and Sankoff): 200 years (est.)
2000: Using GRAPPA v1.1 on the 512-processor Los Lobos Supercluster machine: 2 minutes (200,000-fold speedup per processor)
2003: Using latest version of GRAPPA: 2 minutes on a single processor (1-billion-fold speedup per processor)

GRAPPA (Genome Rearrangement Analysis under Parsimony and other
Phylogenetic Algorithms)http://www.cs.unm.edu/~moret/GRAPPA/• Heuristics for NP-hard optimization problems• Fast polynomial time distance-based methods• Contributors: U. New Mexico, U. Texas at
Austin, Universitá di Bologna, Italy• Freely available in source code at this site.• Project leader: Bernard Moret (UNM)

Limitations and ongoing research
• Current methods are mostly limited to single chromosomes with equal gene content (or very small amounts of deletions and duplications).
• We have made some progress on developing a reliable distance-based method for chromosomes with unequal gene content (tests on real and simulated data show high accuracy)
• Handling the multiple chromosome case is harder

Acknowledgements
• NSF
• The David and Lucile Packard Foundation
• The Program in Evolutionary Dynamics at Harvard
• The Institute for Cellular and Molecular Biology at UT-Austin
See http://www.phylo.org and http://www.cs.utexas.edu/~tandy for more info