class 10. motor proteins that move along polymer tracks. kinesin. yildiz et al, kinesin walks...
TRANSCRIPT

Class 10. Motor proteins that move along polymer tracks. Kinesin. Yildiz et al, Kinesin walks hand-over-hand. Science303:676 (2004).
Goals:Learn about motor proteins that move along tracksPositioning the center of an object to better than lTotal internal reflection microscopy and detecting
fluorescence from single moleculesWaiting time distributionsMore general concept of Brownian motors
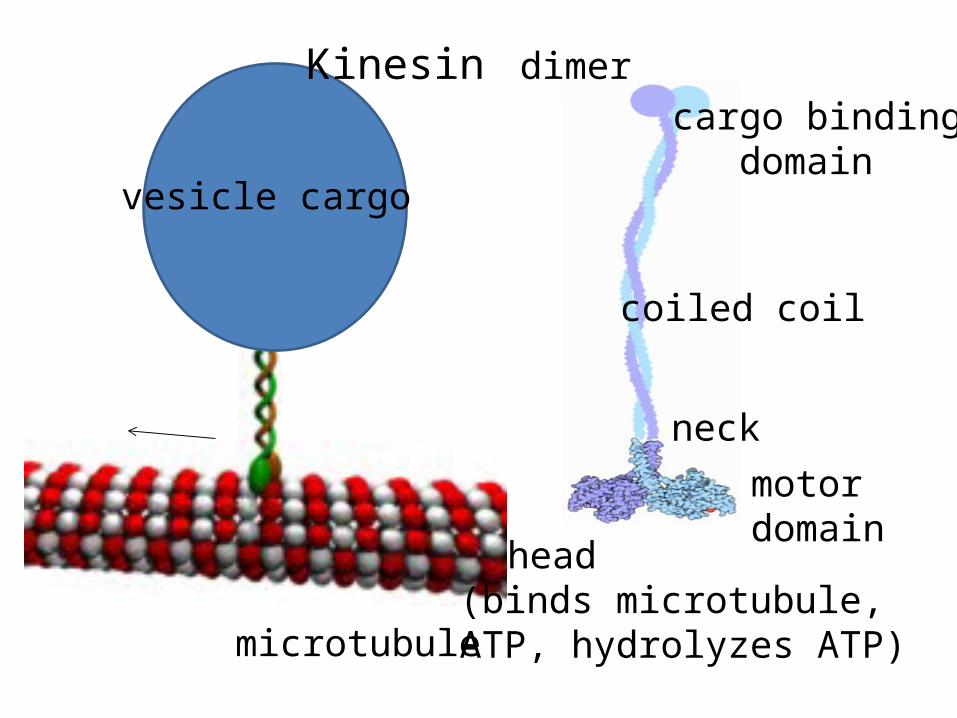
microtubule
vesicle cargo
Kinesin dimer
motor domain
neck
coiled coil
cargo binding domain
head (binds microtubule, ATP, hydrolyzes ATP)

Head groups go through cycles of binding b-tubulin and coming off
One head group stays bound when other releases so kinesinstays attached through many bind/release cycles
Head groups take turns binding sequential b-tubulins inprotofilament so kinesin “walks” bipedially
Cycles of binding and release coupled to cycles of bindingand hydrolyzing ATP, releasing ADP – details unclear
Interactions with tubulin, ATP, ADP etc change conformationof kinesin, which changes energy of binding other mol.
Details of walk - protein conformation at each chemo-mechanical step – are subjects of great interest
What makes the walk go in one direction?
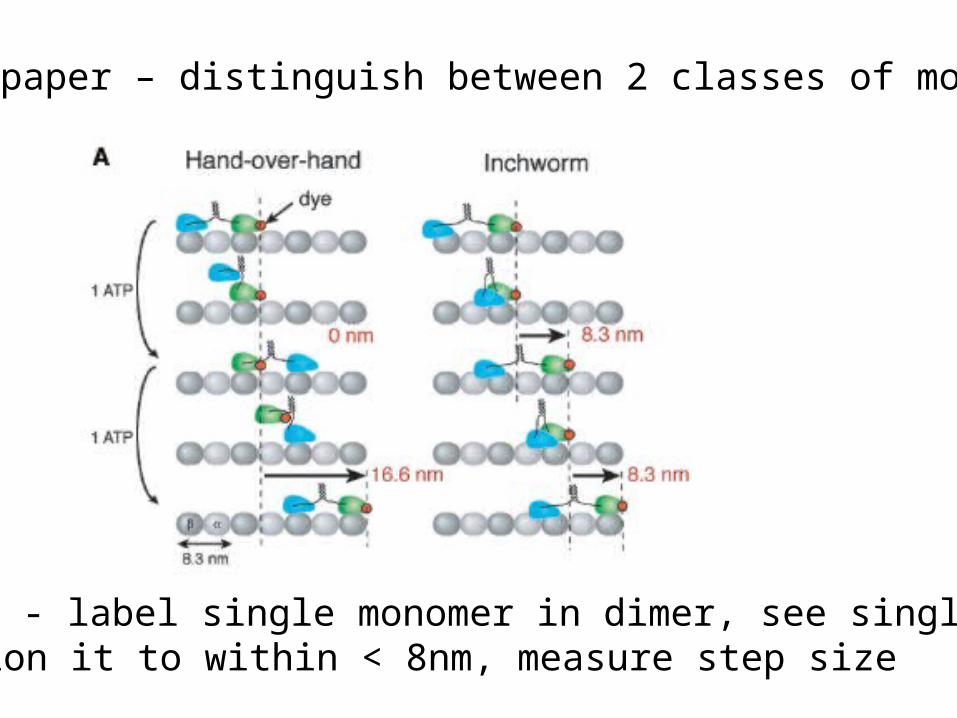
Goal of paper – distinguish between 2 classes of models
Strategy - label single monomer in dimer, see single label,position it to within < 8nm, measure step size

Labeling method – mutate to eliminate solvent exposed cysreplace E215 or T324 or S43 with cyscovalently couple fluorescent dye to mutant kinesinat dye:dimer ratio 0.4; most dimers get <1 dye
dye-labeled sites (in diff.molecules)
ATP binding site
What would happen if both monomers got labeled?

Experimental set-up
Adhere sea urchin axonemes (microtubules)to glass slide
Add dye-labeled kinesins
Preliminary expts – dye-labeled kinesins bind in absenceof ATP but don’t move; move when ATP added

How to see fluorescence from single dye molecule
Total Internal Reflection (TIR) Microscopy
Limiting illumination to evanescentfield markedly decreases background Sako and Uyemura, Cell Struct
and Function 27:357 (2002)

How do you know you are looking at single dye molecules?
Fluorescent molecules “bleach” (photo-chemicalmodification -> loss of fluorescence)
Single step loss of intensity to 0 => single molecule
~ 400 photons/sec for20 sec, then nothing
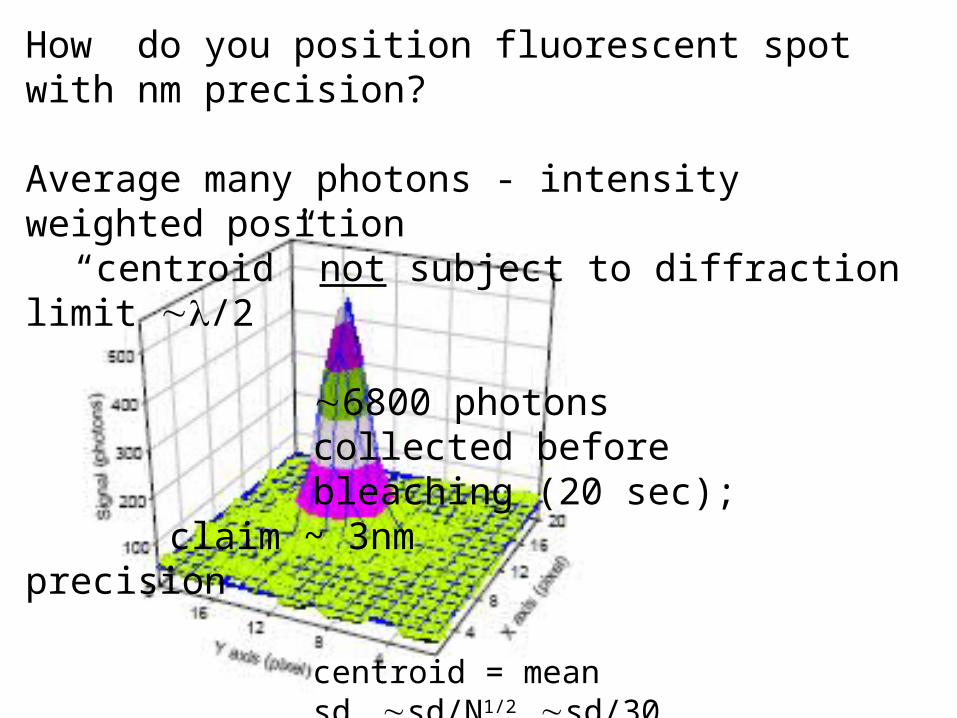
How do you position fluorescent spot with nm precision?
Average many photons - intensity weighted position“centroid” not subject to diffraction limit ~l/2
~6800 photonscollected beforebleaching (20 sec); claim ~ 3nm precision
centroid = meansdm ~sd/N1/2 ~sd/30
if sd ~1 pixel ~100nm sdm~0.03 pixel
~3nm

Snapshots from movie of walking kinesin dye. Pixels 86nm
Amazing that what seems like fairly coarse binning does not significantly impair precision
Thompson et al, Biophys J 82:2775 (2002)Valentine and Rana, IEEE Trans Nucl Sci 43: 2501 (1996)

Step trajectories for individual kinesin moleculeswith dye in different positions
step wait times ~1sec => ~1000 photons/step

Step size distribution for 124 steps from 22 moleculesof E215C, 12 steps from 3 molecules of T324C, and 7 steps from 1 molecule of S43C/T324C heterodimer
mean + sd = 17.3 + 3.3 nmrules out inch worm model (8.3 nm steps)fits hand-over-hand model (2 x 8.3 nm)

Distribution of waiting times between steps
Hand-over- hand model predicts 2 random events – What are they?
Initial dip not consistent with single random eventreasonable fit to P(t) = t k2 exp(-kt) (2 random events)
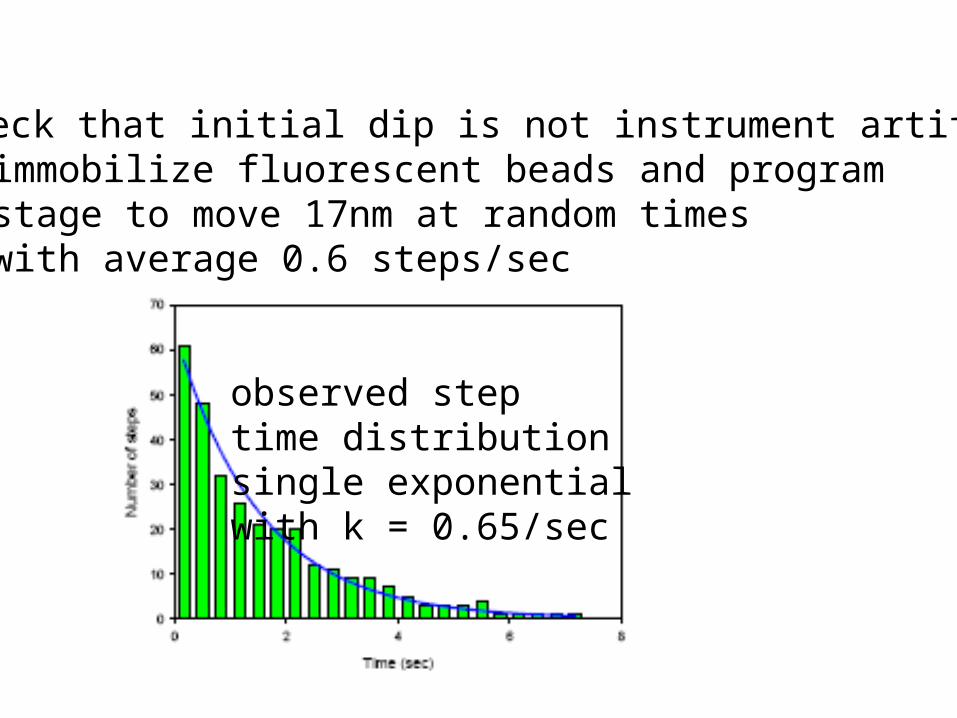
Check that initial dip is not instrument artifact:immobilize fluorescent beads and programstage to move 17nm at random timeswith average 0.6 steps/sec
observed steptime distributionsingle exponentialwith k = 0.65/sec
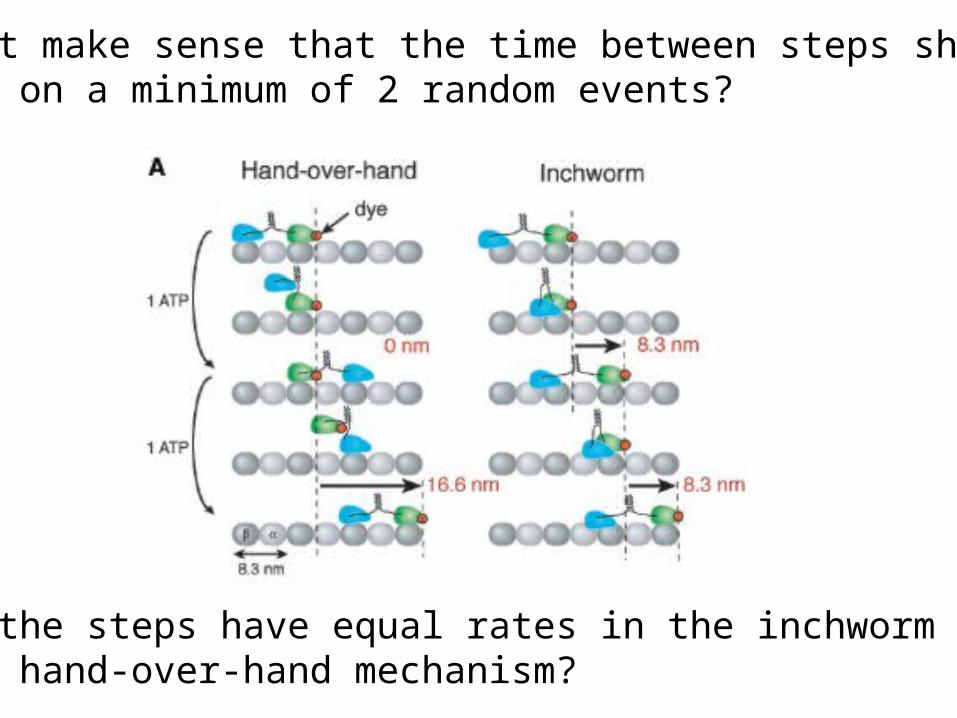
Does it make sense that the time between steps shoulddepend on a minimum of 2 random events?
Would the steps have equal rates in the inchworm model?In the hand-over-hand mechanism?

If alternate steps in hand-over-hand model werenot identical due to alternately overwinding/underwinding the dimer coiled coil, alternatewaiting time distributions could be different. How could they look for this in their data?
See if waiting time distribution fits twostochastic events with different waiting times: p(t) = [k1k2/(k2-k1)](e-k1t - e-k2t)
Compare waiting time distributions of odd # vseven # steps: podd(t) = k1e-k1t , peven(t) = k2e-k2t

Variant hand-over-hand models: monomer detaches,pauses at relaxed position waiting for some event,e.g. ATP binding, then moves to next binding site
predicts alternate steps oflength x, 16.6 – x
they don’t see intermediate step sizes
alternate steps have samesize distribution to within 2 nm
x
16.6 - x

What would you expect for step size and waiting time distribution if both monomers were labeled?
step size: centroid “middle” of dimerh-over-h: 8nm stepsinchworm: 4nm steps
waiting time: p(t) = ke-kt , since you see movement when each head steps (k1 and k2 if alt. steps differ)
What would you predict for step sizes if the monomers were labeled with different color (red, green) dyes?
hand-over-hand: red, green alternate 16nminchworm: each step 8nm, alternately overlapping

Brownian motor – concept underlying biomolecular motorsand other nanoscale devices for directed motion
simply turning on and off an asymmetric potential can -> uphill motion
doesn’t violate 2nd law because takes work to turn on and off potential
http://monet.physik.unibas.ch/~elmer/bm/ - Br. motor simulation
http://www.physik.uni-augsburg.de/theo1/hanggi/Papers/309.pdf
Asymmetric potential along “track”
object that binds
and diffuses when released

Applied to kinesin motor:
ATP/ADP-induced conformational changes turn on and off different interactions along length of tubulin, equivalent to changing potential energy landscape
Kinesin heads might usually go forward but sometimes go backward between attachments steps
ATP binding/hydrolysis effect not as simple as“ATP binding or hydrolysis powers forward movement”

Conclusions
Beautiful paper showing that kinesin arms move in ~17nmsteps, consistent with hand-over-hand model
Uses single-molecule tracking – can’t see how results could have been obtained with bulk measurements
Similar methods applicable to study other motors that move along tracks – e.g. myosin on actin (in fact, authors’ previous paper was “myosin walks hand-over-hand”)
Biomolecular motors suggest general principles for other nano-motors that must work in Brownian environment: alternately change binding interactions with polymer

More than half-way through course – basic ideas so far:
Knowledge of structure of DNA and other biopolymers-> ways to engineer them -> novel nanostructures
elastic properties of biopolymers – WLC -> ways to manipulate and measure single-molecule interactions
Brownian motion in liquids at nanoscale -> diffusion, viscous drag, Brownian ratchet motors
Few more examples – rotary motors, stretching proteins
Think about what’s missing in understanding molecular motors and how to make new nanomachines using principles learned from biological systems