cloning of a vacuolar h+-pyrophosphatase gene from the halophyte suaeda corniculata whose...
TRANSCRIPT

Journal of Integrative Plant Biology 2011, 53 (9): 731–742
Research Article
Cloning of a Vacuolar H+-pyrophosphatase Genefrom the Halophyte Suaeda corniculata whoseHeterologous Overexpression Improves Salt,Saline-alkali and Drought Tolerance in ArabidopsisLiang Liu1,2†, Ying Wang1†, Nan Wang1, Yuan-Yuan Dong1, Xiu-Duo Fan1,2, Xiu-Ming Liu1,Jing Yang1 and Hai-Yan Li1,2∗
1Ministry of Education Engineering Research Center of Bioreactor and Pharmaceutical Development, Jilin Agricultural University,Changchun 130118, China2College of Life Sciences, Jilin Agricultural University, Changchun 130118, China†These authors contributed equally to this work.∗Corresponding author
Tel: +86 431 8453 3427; Fax: +86 431 8453 3347; E-mail: [email protected] online on 14 July 2011 at www.jipb.net and www.wileyonlinelibrary.com/journal/jipbdoi: 10.1111/j.1744-7909.2011.01066.x
Abstract
Salt, saline-alkali conditions, and drought are major environmental factors limiting plant growth andproductivity. The vacuolar H+-translocating inorganic pyrophosphatase (V-H+-PPase) is an electrogenicproton pump that translocates protons into vacuoles in plant cells. Expression of V-H+-PPase increasesin plants under a number of abiotic stresses, and is thought to have an important role in adaptation toabiotic stress. In this work, we report the isolation and characterization of the gene, ScVP, encodinga vacuolar inorganic pyrophosphatase (V-H+-PPase) from the halophyte, Suaeda corniculata. Semi-quantitative reverse transcription-polymerase chain reaction analysis showed that ScVP was induced inroots, stems and leaves under treatment with salt, saline-alkali and drought. Compared with wild-type(WT) Arabidopsis, transgenic plants overexpressing ScVP accumulated more Na+ in leaves and roots,and showed increased tolerance to high salinity, saline-alkali and drought stresses. The germinationpercentage of transgenic Arabidopsis seeds was higher than that of WT seeds under the abiotic stresses.The root length of transgenic plants under salt stress was longer than that of WT plants. Furthermore, therate of water loss during drought stress was higher in WT than in transgenic plants. Collectively, theseresults indicate that ScVP plays an important role in plant tolerance to salt, saline-alkali and droughtstress.
Keywords: drought; H+-pyrophosphatase; salt; saline-alkali; Suaeda corniculata.
Liu L, Wang Y, Wang N, Dong YY, Fan XD, Liu XM, Yang J, Li HY (2011) Cloning of a vacuolar H+-pyrophosphatase gene from the halophyteSuaeda corniculata whose heterologous overexpression improves salt, saline-alkali and drought tolerance in Arabidopsis. J. Integr. Plant Biol. 53(9),731–742.
Introduction
Salinity is a major environmental factor limiting crop growthand productivity (Ashraf 1994). It is estimated that 20% of allcultivated land, and nearly half of irrigated land, is affected bysalinity (Rhoades and Loveday 1990). To cope with salt stress,
plants have developed multifarious adaptation mechanismsto tolerate high concentrations of salt in the environment(Hamada et al. 2001). The compartmentalization of Na+ in thevacuole, through vacuolar Na+/H+ antiporters, might reducethe deleterious effects of excess Na+ in the cytosol andmaintain osmotic balance by using Na+ accumulated in the
C© 2011 Institute of Botany, Chinese Academy of Sciences

732 Journal of Integrative Plant Biology Vol. 53 No. 9 2011
vacuole, thus enhancing water uptake and salt tolerance ofthe plant (Gaxiola et al. 2001). The Na+ compartmentalizationprocess is mediated by a vacuolar Na+/H+ antiporter that isdriven by the electrochemical gradient of protons across thetonoplast generated by vacuolar H+-pumps, H+-ATPase andH+-pyrophosphatase (H+-PPase) (Apse et al. 1999; Sze et al.1999; Li et al. 2004).
Vacuolar H+-pyrophosphatase (V-H+-PPase) is an electro-genic proton pump that catalyzes the hydrolysis of inorganicpyrophosphate (PPi) to energize proton transport from thecytoplasm into vacuoles (Maeshima 2000). Comparison of V-H+-PPases from different organisms has identified a numberof conserved motifs (Rea and Poole 1993; Baltscheffsky et al.1999; Maeshima 2000; Mimura et al. 2004), such as CS1,CS2 and CS3. The CS1 motif includes the catalytic sequenceDVGADLVGKVE, which is essential for substrate hydrolysis(Rea and Poole 1993; Takasu et al. 1997; Mimura et al.2004), thus, this region is exposed to the cytosolic side ofthe tonoplast (Tanaka et al. 1993). The CS2 motif is alsolocated in a hydrophilic loop, whereas CS3 is located in thecarboxy-terminal region and contains 12 charged residues. TheCS3 motif is exposed to the cytosol and plays a critical rolein catalytic function in combination with CS1 and CS2 (Reaet al. 1992). All characterized V-H+-PPases consist of a single75–82 kDa intrinsic membrane protein species comprising twostructurally and functionally distinct types: I and II. Type I V-H+-PPases are exemplified by the molecular prototype AVP1 fromArabidopsis (Sarafian et al. 1992), and type II V-H+-PPasesare exemplified by Arabidopsis AVP2 (Drozdowicz et al. 2000).
Transgenic plants overexpressing AVP1, a gene encodinga single subunit protein for V-H+-PPase, displayed enhancedsalt-stress tolerance (Gaxiola et al. 2001). Improved salt- anddrought-stress tolerance was also achieved in Arabidopsisthrough overexpression of the wheat H+-PPase TVP1 (Briniet al. 2007). Transgenic tomatoes expressing AVP1 were moreresistant to soil water deficit than controls because of their morerobust root systems (Perez-Alfocea et al. 1993; Torrecillaset al. 1995; Li et al. 2005; Park et al. 2005). These resultsindicate that enhanced vacuolar H+ pumping in the transgenicplants is an effective approach to improve crop salt and droughttolerance.
Suaeda corniculata, a member of the Chenopodiaceae, isnative to the saline-alkali soil (NaCl+NaHCO3) of NortheastChina and is one of the most important halophytes in China. Inthis study, we report the isolation of ScVP, the V-H+-PPasegene from S. corniculata, and demonstrate that transgenicArabidopsis plants overexpressing tonoplast H+-PPase (ScVP)are much more resistant to high concentrations of salt, saline-alkali stress and water deprivation than wild type (WT) plants.To our knowledge, this is the first study to report overexpressionof the H+-PPase enhanced resistance to saline-alkali stress intransgenic Arabidopsis.
Results
Cloning of ScVP cDNA and sequence analysis
The full-length cDNA of ScVP was obtained by reversetranscription-polymerase chain reaction (RT-PCR) and rapidamplification of cDNA ends (RACE). The ScVP cDNA con-tained an open reading frame (ORF) of 2 292 bp encodinga protein of 764 amino acids (GenBank accession num-ber: ADQ00196). Phylogenetic analysis (Figure 1A) showedthat ScVP formed a clade with the most closely relatedplant H+-PPase homologs. The highest identity was foundto be 97% similarity with the V-H+-PPase from Halostachyscaspica, a salt-tolerant grass. ScVP also showed high sim-ilarity to V-H+-PPases of Kalidium foliatum (96%), Suaedasalsa (94%), Chenopodium rubrum (89%), Beta vulgaris (89%),Chenopodium glaucum (88%) and Arabidopsis thaliana (87%).As shown in Figure 1B, the first conserved segment (CS1)transmembrane domain contains a putative catalytic motif (DV-GADLVGKVE) common to V-H+-PPases and was confirmed tobe exposed to the cytosol (Takasu et al. 1997). These resultsindicate that S. corniculata ScVP is closely related to V-H+-PPases of other Chenopodiaceae species.
Transcript analysis of ScVP in Suaeda corniculata
The transcript level of ScVP in seedlings of Suaeda corniculatawas assayed in the presence of 100 µmol/L abscisic acid(ABA), 250 mM NaCl, 200 mM NaCl and 100 mM NaHCO3,or 30% PEG8000 by semiquantitative RT-PCR. Salt, saline-alkali, drought stresses and ABA application induced transcriptaccumulation of ScVP (Figure 2A). The transcript level of ScVPwas induced for 0 h prior to salt and drought stresses, andreached a maximum after 24 h. Thereafter, the transcript leveldecreased only slightly after 48 h of treatment. In the saline-alkali stress treatment, transcripts of ScVP were detected at0 h and peaked at 12 h, and then decreased after 24 h ofsaline-alkali stress treatment. Interestingly, the transcript levelof ScVP also increased in response to exogenous ABA, whichfunctions as a signaling molecule and plays an important role instress tolerance (Figure 2A). In addition, we also examined theexpression patterns of ScVP in different tissues, including theroot, stem and leaf of S. corniculata seedlings under salt andsaline-alkali stress. The results showed that accumulation ofScVP transcript in root reached its maximum level within 12 hunder salt stress, but decreased at 24 h and reduced to almost abasal level at 48 h. In stem, the ScVP transcript level reached amaximum at 24 h, while experiencing a reduction at 48 h of salttreatment. In leaf, the ScVP transcript accumulation followedthe same course as in the stem (Figure 2B), which indicatedthat the expression pattern of ScVP in root under salt stress isdifferent from that in stem and leaf. The expression pattern of

Vacuolar H+-pyrophosphatase in Plant Stress Tolerance 733

734 Journal of Integrative Plant Biology Vol. 53 No. 9 2011
ScVP in root, stem and leaf in response to saline-alkali stress issimilar to that of salt treatment (Figure 2C). However, transcriptlevel was higher under saline-alkali stress than that under saltstress, suggesting that the gene may function differently indifferent stress conditions.
Overexpression of ScVP confers enhanced salt,drought, and saline-alkali tolerance in transgenicArabidopsis
To examine the function of ScVP, after transformation of Ara-bidopsis plants with 35S::ScVP and selection with the herbicideBasta, several transformants were produced. The expression ofScVP genes in all ScVP transgenic lines has been monitored byrelative quantitative RT-PCR. Figure 3 shows that ScVP trans-genic lines L8 and L45 show a slightly higher level of expressionof ScVP than the three other lines, thus they were used forsubsequent analysis. Overexpressing ScVP transgenic linesand WT plants were tested for salt, saline-alkali and droughttolerance, seedlings of both transgenic Arabidopsis lines andWT were grown in a Petri dish for 5 d, and then transferredto MS medium containing 100, 150, 200 mM NaCl, 7.5 mMNaHCO3 + 92.5 mM NaCl or 10.0 mM NaHCO3 + 90 mMNaCl. The results showed that the WT seedlings withered andthe leaves became chlorotic, whereas the transgenic seedlings
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 1. Phylogenetic tree and multiple alignments representing evolutionary relationships among ScVP and closely related
vacuolar H+-pyrophosphatases from other plant species.
(A) Phylogenetic analysis was based on typical vacuolar proton-pumping PPase from plants. The gene abbreviation and GenBank accession
numbers are as follows: Suaeda corniculata (ScVP; ADQ00196), Suaeda salsa (SsVP) (Guo et al. 2006), Chenopodium rubrum (CVP1;
AAM97920), Beta vulgaris (BVP1; AAA61610), B. vulgaris (BVP2; AAA61609), Hordeum vulgare (HVP; ACA63883), H. vulgare (HVP1;
BAB18681), H. vulgare (HVP2; BAA02717), H. brevisubulatum (HbVP1; AAP06752), Oryza sativa (OVP1; BAA08232), O. sativa (OVP2;
BAA08233), Vigna radiata (VVP2; BAA23649), Nicotiana tabacum (NtVP; CAA58701), N. tabacum (NtVP5; CAA54869), N. tabacum (NtVP9;
CAA58700), Lotus corniculatus (LcVP1; ABS01290), Kalidium foliatum (KfVP1; ABK91685), Arabidopsis thaliana (AVP1; NP_173021), A.
thaliana (AVP2; AAF31163), A. thaliana (AVP3; AAA32754), Triticum aestivum (TVP1; AAP55210), Zygophyllum xanthoxylum (ZxVP1;
ABU92563), Thellungiella salsuginea (TsVP1; AAR08913), Vitis vinifera (VvVP; AAF69010), Medicago truncatula (MtVP1; ACI22377),
Leptochloa fusca (LfVP; ACT98610), Prunus persica (PpVP1; AAL11506), Prunus persica (PpVP2; AF367447), Populus trichocarpa
(PtVP; XP_002325187), Chenopodium glaucum (CgVP1; ABD98735), Ricinus communis (RcVP; XP_002530755), Zea mays (ZmVPP1;
NP_001105380), Pyrus communis (PcVP; BAC41250), Potamogeton distinctus (PdVP; BAF63470), Halostachys caspica (HcVP; ABO45933).
The protein sequences were aligned with CLUSTAL X and a phylogenetic tree constructed using MEGA 4.1. Bootstrap percentages are
shown at dendrogram branch points.
(B) Multiple alignment of the deduced amino acid sequence of ScVP with those of closely related vacuolar H+-pyrophosphatases from
other plant species. Alignments among eight H+-PPases of the segments containing highly conserved motifs are shown. The protein
abbreviations and GenBank accession numbers are as follows: Suaeda corniculata (ScVP; HQ412514), Suaeda salsa (SsVP), Chenopodium
rubrum (CVP1; AAM97920), Beta vulgaris (BVP1; AAA61610), Chenopodium glaucum (CgVP1; ABD98735), Halostachys caspica (HcVP;
ABO45933), Arabidopsis thaliana (AVP1; NP_173021), and Kalidium foliatum (KfVP1; ABK91685). The sequences were aligned with
ClustalW. A catalytic domain (indicated by the line above the alignment) has been proposed to be present in the first conserved segment
(CS1). Asterisks indicate identical amino acid residues, colons indicate amino acids with high similarity, periods indicate amino acids with
low similarity, and dashes indicate gaps. The line shows the score of the catalytic domain.
were slightly affected and could survive under 200 mM NaCland 10.0 mM NaHCO3 + 90 mM NaCl conditions (Figure 4A).After seedlings were transferred to MS medium containing 10%polyethylene glycol (PEG), controls showed reduced growthand chlorosis, whereas transgenic lines continued to grownormally (Figure 4A). In addition, all the seedlings grew normallyin NaCl up to 150 mM, while the transgenic seedlings formedlonger roots than the WT. However, we did not find a significantdifference in root length between transgenic plants and controlplants under saline-alkali and drought treatment (Figure 4B).
Effects of salt, saline-alkali and drought stress ongermination of Arabidopsis seeds
To evaluate the effect of ScVP on seed germination underabiotic stresses, the germination percentage of WT and trans-genic Arabidopsis seeds L8 and L45 were determined afterincubation for 2 weeks. In the controls, 99% of the seedshad germinated after 2 weeks. The germination percentageof transgenic seeds did not differ from that of WT seeds undernormal growth conditions. While when exposed to 100 mMNaCl, 10.0 mM NaHCO3 + 90 mM NaCl, and 10% PEG for10 d, the germination percentage of WT seeds decreased bymore than 4%, 86% and 72%, respectively, and the germinationpercentage of transgenic seeds was significantly higher than

Vacuolar H+-pyrophosphatase in Plant Stress Tolerance 735
Figure 2. Transcript patterns of ScVP in Suaeda corniculata
seedlings in response to different stresses.
(A) Transcript level of ScVP in the whole plant under salt (250 mM
NaCl), dehydration (30% polyethylene glycol [PEG]), abscisic acid
(ABA)(100 µM), and saline-alkali (200 mM NaCl and 100 mM
NaHCO3).
(B) Transcript level of ScVP in different organs under salt stress
(250 mM NaCl).
(C) Expression of ScVP in different organs under saline-alkali stress
(200 mM NaCl and 100 mM NaHCO3).
that of WT seeds under all abiotic stress conditions (Figure 5A).Thus, overexpression of ScVP in Arabidopsis enhanced thesalt, saline-alkali and drought stress tolerance of the transgenicplants at germination stage (Figure 5B).
Na+ accumulation in transgenic plants
To investigate whether expression of ScVP increases theconcentration and sequestration of Na+ into the vacuole, wemeasured the amounts of Na+ in the roots and leaves of ScVPtransgenic plants and WT under different NaCl and saline-alkaliconcentrations. The results showed that under salt and saline-alkali treatments, Na+ content significantly increased in theleaves and roots of both transgenic and WT plants when theconcentration of NaCl and saline-alkali in the medium increased
Figure 3. Analysis of ScVP expression in Arabidopsis trans-
genic lines by relative quantitative reverse transcription-
polymerase chain reaction (RT-PCR).
Specific PCR products of 456 bp were detected in five ScVP trans-
genic lines, respectively. A 375 bp Actin fragment was amplified by
RT-PCR as an internal control.
(Figure 6). However, transgenic plants accumulated more Na+
in their leaves and roots than WT plants under salt and saline-alkali treatments, which indicated that overexpression of ScVPin Arabidopsis increases the concentration and sequestrationof Na+ into the vacuole.
Relative water content in ScVP overexpressingArabidopsis and WT plants
After 9 d of water deprivation, WT plants showed growthreduction and chlorosis, whereas transgenic lines exhibitednormal growth. After 12 d of water-deficit stress, plants fromeach group were re-watered; the control plants died, whereasthe transgenic plants continued to grow normally. Leaf relativewater content (RWC) was evaluated during water deprivation.The RWC was reduced in both transgenic (L8 and L45) and WTplants, but the rates of decline were greater in the WT plants.After 7 d of water-deficit stress, a higher rate of water loss wasobserved in WT plants, in which the RWC decreased by morethan 50%, whereas the RWC of transgenic plants decreasedby only 20% (Figure 7). Thus, the transgenic plants showedan enhanced capacity for water uptake and increased droughttolerance during water-deficit stress.
Discussion
Vacuolar H+ pumps play a key role in the maintenance ofthe H+ electrochemical gradient across the vacuolar mem-brane in plant stress responses. In this study, a novel geneencoding a V-H+-PPase, ScVP, was isolated from halophyteS. corniculata. We examined the expression patterns of ScVPin response to various environmental stresses. Expression ofScVP in S. corniculata seedlings was significantly inducedby high salinity, saline-alkali, drought and ABA application(Figure 2). This pattern of expression is similar to that ofTsVP in Thellungiella halophila (Lv et al. 2008). In addition,comparing gene expression under salt and saline-alkali stress,expression was higher in all the tissues under saline-alkali

736 Journal of Integrative Plant Biology Vol. 53 No. 9 2011
Figure 4. Root growth assay to determine stress sensitivity. Seedlings were photographed 8 d after transplantation, when the
transgenic stressed seedlings (L8 and L45) showed a higher survival percentage compared with the wild type (WT) (controls).
(A) Arabidopsis seedlings grown in standard culture medium for one week (the first two true leaves had already emerged) before transferal
to medium containing either 0 mM, 100 mM, 200 mM NaCl, 7.5 mM NaHCO3 + 92.5 mM NaCl or 10.0 mM NaHCO3 + 90 mM NaCl and
10% PEG8000.
(B) Effect of ScVP expression on salt tolerance in transgenic Arabidopsis and WT seedlings. Data shown are root length in response to 0 mM,
100 mM, 150 mM or 200 mM NaCl. Data were obtained from three independent experiments and are means ± SE. ∗P < 0.05, ∗∗P < 0.01.
ScVP, transgenic plants overexpressing ScVP; WT, non-transgenic controls.
stress than that of salt treatment, suggesting that the gene mayfunction differently under salt and saline-alkali stress. Furtherstudies are now in progress in our laboratory to investigate thisrelationship.
To avoid the toxic effects of salt, plants have developedmechanisms to sequester Na+ into vacuoles (Amtmann andSanders 1999). V-H+-PPase is an electrogenic proton pump
that translocates protons across the tonoplast into vacuoles,thereby supporting strong ion differences and membrane po-larization (Felle 2005), and alleviating acidification of the cytosol(Stitt 1998; Maeshima 2000). H+-PPase can enhance theaccumulation of Na+ in vacuoles. Accumulation of Na+ invacuoles instead of in the cytoplasm can avoid the toxic effectsof excessive Na+ in plant cells (Gaxiola et al. 2001; Ohta
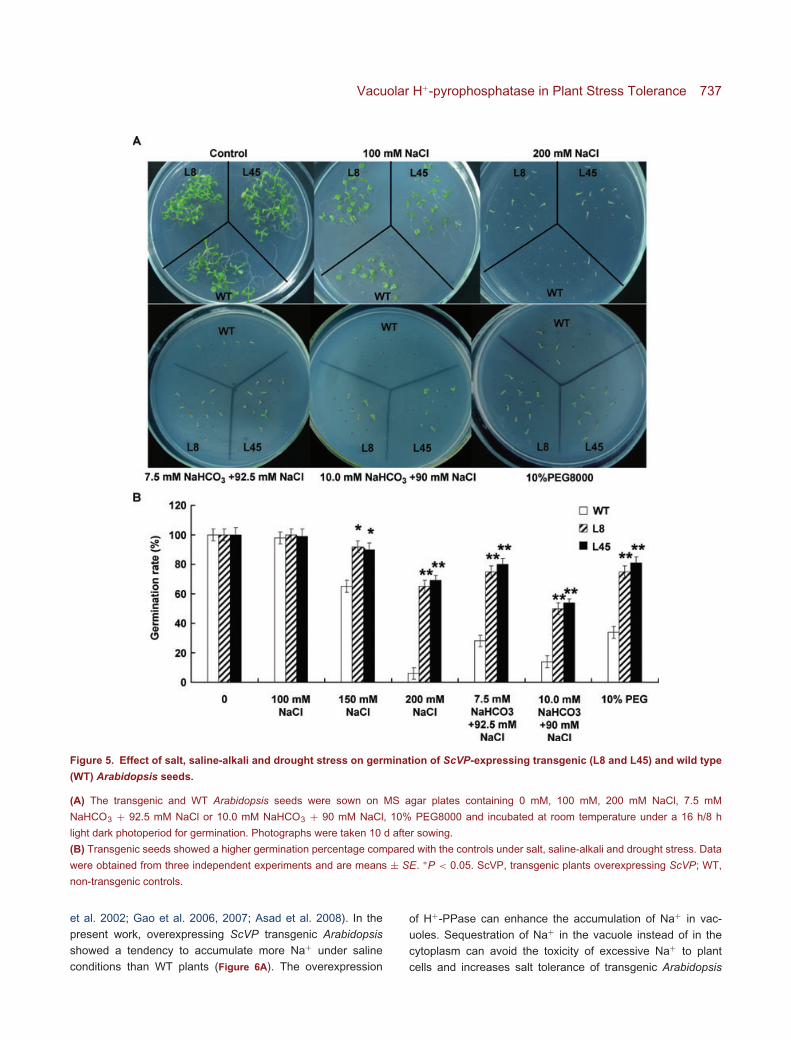
Vacuolar H+-pyrophosphatase in Plant Stress Tolerance 737
Figure 5. Effect of salt, saline-alkali and drought stress on germination of ScVP-expressing transgenic (L8 and L45) and wild type
(WT) Arabidopsis seeds.
(A) The transgenic and WT Arabidopsis seeds were sown on MS agar plates containing 0 mM, 100 mM, 200 mM NaCl, 7.5 mM
NaHCO3 + 92.5 mM NaCl or 10.0 mM NaHCO3 + 90 mM NaCl, 10% PEG8000 and incubated at room temperature under a 16 h/8 h
light dark photoperiod for germination. Photographs were taken 10 d after sowing.
(B) Transgenic seeds showed a higher germination percentage compared with the controls under salt, saline-alkali and drought stress. Data
were obtained from three independent experiments and are means ± SE. ∗P < 0.05. ScVP, transgenic plants overexpressing ScVP; WT,
non-transgenic controls.
et al. 2002; Gao et al. 2006, 2007; Asad et al. 2008). In thepresent work, overexpressing ScVP transgenic Arabidopsisshowed a tendency to accumulate more Na+ under salineconditions than WT plants (Figure 6A). The overexpression
of H+-PPase can enhance the accumulation of Na+ in vac-uoles. Sequestration of Na+ in the vacuole instead of in thecytoplasm can avoid the toxicity of excessive Na+ to plantcells and increases salt tolerance of transgenic Arabidopsis

738 Journal of Integrative Plant Biology Vol. 53 No. 9 2011
Figure 6. Na+ content in the leaf and root of transgenic and wild type (WT) Arabidopsis seedlings grown under different NaCl and
saline-alkali concentrations.
(A) Na+ content was measured after 15 d of salt treatment.
(B) Na+ content was measured after 15 d of saline-alkali treatment. Data were obtained from three independent experiments and are
means ± SD (n = 6). ∗P < 0.05, ∗∗P < 0.01. ScVP, transgenic plants overexpressing ScVP; WT, non-transgenic controls.
plants (Figure 4A, 5A). Moreover, transgenic Arabidopsis plantsincrease drought tolerance (Figure 4A). The RWC in leaves oftransgenic plants decrease less than WT plants (Figure 7). Thismay be a consequence of ScVP overexpression that resultsin an increase in the concentration of solutes in the cell (Guoet al. 2006). The increase in the concentration of solutes mustlead to an increase in the uptake of water and the retention ofwater under drought conditions.
In spite of the considerable volume of research on NaClstress, one very important aspect has been neglected: saline-alkali stress, which is more complex than salt stress alone,and exerts the same stress factors as salt stress but with theaddition of high pH stress. In the present work, in order tocharacterize the saline-alkali tolerance function of ScVP, twotreatments of salinity, salt (100 mM NaCl, pH 5.8) and saline-alkali (10.0 mM NaCl + 90 mM NaHCO3, pH 8.5), which containthe same Na+ concentration but different pH, were performed tothe overexpressing ScVP transgenic Arabidopsis. The resultsshowed that although salt stress had some inhibitory effecton germination of the WT seeds, many WT seeds remainedviable under 100 mM NaCl salt stress, and there was no
inhibition of transgenic seed germination (Figure 5A). However,WT seedlings withered and died under 10.0 mM NaCl + 90 mMNaHCO3 aline-alkali stress, while the transgenic seeds sown ina saline-alkali environment (10.0 mM NaHCO3+ 90 mM NaCl)remained viable (Figure 5A). This suggests that high pH maylead to lack of protons, the destruction or inhibition of trans-membrane electrochemical-potential gradients in root cells,and the loss of normal physiological root functions such as Na+
absorption (Yang et al. 2006; Li et al. 2008; Zhang et al. 2008).Whereas in this work, the Arabidopsis plants overexpressingScVP showed a tendency to accumulate more Na+ than WTplants under saline-alkali conditions (Figure 6B). The increasedaccumulation of sodium is likely to be a consequence of theactivity of the vacuolar secondary transports, carrying protonsinto the cytoplasm and leading to cytosolic acidification inthe plant cell; then protons accumulated in the cytosol maybe exported across the plasma membrane by proton pumps.Arabidopsis plants overexpressing ScVP are more tolerant ofhigh external pH due to extrusion of protons to the extracellularspace. Therefore, our results indicated that the saline-alkalitolerance in transgenic Arabidopsis may be accompanied by

Vacuolar H+-pyrophosphatase in Plant Stress Tolerance 739
Figure 7. Relative water content of wild type (WT) and ScVP-
expressing transgenic Arabidopsis plants (L8 and L45) grown
under water-deficit stress.
Values are means ± SD (n = 4 for each plant type). ScVP, trans-
genic plants overexpressing ScVP; WT, non-transgenic controls.
changing the vacuolar pH, which is brought about by increasingthe activities of V-PPase. In addition, Maeshima (2001) havereported that plant V-PPase maintain the acidic pH of vacuoles.However, the mechanisms underlying this phenomenon requirefurther investigation.
The present finding that the vacuolar H+-PPase gene from S.corniculata conferred salt, saline-alkali and drought tolerance inArabidopsis suggests that it may be worthwhile to elucidate thecontribution of these proteins to stress tolerance in crop plant.The overexpression of ScVP gene in economically importantcrop plants might be a strategy for engineering cultivars ofagriculturally important plants that are adapted to importantabiotic stresses, such as saline and saline-alkali soils.
Materials and Methods
Plant material and treatments
Seeds of Suaeda corniculata were collected from a saline-alkalisoil area located in Baicheng, northeast China. The seeds weresterilized and germinated on MS medium supplemented with3% (w/v) sucrose and 0.7% (w/v) agar in a growth chamberfor 8–10 d at 22 ◦C. Seedlings were grown in pots filledwith a 5:4:1 (v/v/v) mixture of peat/forest soil, vermiculite andperlite in a greenhouse at 24/16 ◦C (day/night) with a 16 hlight/8 h dark photoperiod and 60% relative humidity. Six-week-old plants of S. corniculata were treated with half-strength MSsalt solution containing either 100 µmol/L ABA, 250 mM NaCl(salt stress treatment), 200 mM NaCl and 100 mM NaHCO3
(saline-alkali stress treatment), or 30% PEG8000 (w/v; droughtstress treatment). The roots and leaves of these plants werecollected at 0, 6, 12, 24, and 48 h after initiation of stresstreatment. Plant material was placed directly into liquid nitrogenand stored at –80 ◦C until use.
Isolation of ScVP from Suaeda corniculata by RT-PCRand 5′ and 3′ RACE
To isolate the H+-PPase gene from S. corniculata, degener-ate primers were designed, based on the sequence of theconserved CS1 domain and the 3′ end region of all knowndicot H+-PPase genes. The primer sequences were as fol-lows: forward primer 5′-YGGTCTTGGAGGRTCWTCCA-3′ andreverse primer 5′-AAGRGCRTCRGTTCTCTCSC-3′, where Rdenotes A or G, Y denotes C or T, S denotes G or C, andW denotes A or T. Total RNA was extracted from leaves ofS. corniculata plants treated with 250 mM NaCl for 2 h usingthe Trizol reagent (Invitrogen, Carlsbad, CA, USA) and theRNA extract was treated with RNase-free DNase according tothe manufacturer’s protocol (Invitrogen). Treated RNA (1 µg)was used for first-strand synthesis and PCR according to themanufacturer’s protocol (Invitrogen). The reverse-transcriptionproducts were amplified by PCR under the following thermalcycle conditions: 94 ◦C for 8 min, followed by 35 cycles of94 ◦C for 30 s, 55 ◦C for 30 s, and 72 ◦C for 60 s. The PCRproducts were cloned into the pMD18-T vector (Takara) andsequenced. Specific primers were then designed for RACEaccording to the sequence information of the partial cDNAfragment to obtain the full-length sequence of the gene. For5′-RACE, two antisense gene-specific primers were designed(GSP1, 5′-ATGGGTGYWGYTCTTCTTCC-3′; and GSP2, 5′-AAGGGCATCAGTTCTCTCAC-3′). Primers for 3′-RACE were:GSP1, 5′-GTATTGCTGTGGCTGCTCTT-3′; and GSP2, 5′-GGTATTGCTGAAATGGCTGG-′3. The RACE reactions wereperformed according to the manufacturer’s protocol (Invitro-gen). The RACE products were cloned and sequenced. Afull-length cDNA sequence was obtained by combining the5′-RACE fragment, partial cDNA fragment, and the 3′-RACEfragment. The full-length cDNA was amplified by PCR usingthe primers ScVPF (5′-ATGAGTGGCATTCTTCTTC-3′) andScVPR (5′-TTAGAAGATCTTCAAGAGCA-3′) under the follow-ing thermal cycle conditions: 94 ◦C for 8 min, followed by 35cycles of 94 ◦C for 40 s, 58 ◦C for 40 s, and 72 ◦C for 90 s.The resulting PCR products were cloned into the pMD18-Tvector and independently amplified clones were sequenced.The sequences of the amplified fragments were assembled toprovide the full-length ScVP cDNA. Homology analysis of thededuced protein ScVP was performed and a phylogenetic anal-ysis involving the Neighbor-Joining method was undertakenusing CLUSTAL W and MEGA 4.1 software.
Relative quantitative RT-PCR
Total RNA (1 µg) was reverse-transcribed in a total vol-ume of 10 µL with 1 µL oligo (dT) 18 primer (100 pM),1.0 µl dNTPs (10 mM), 2 µL 5× M-MLV buffer, 0.25 µLRNase inhibitor (40 U/µL), and 0.5 µL reverse-transcriptase

740 Journal of Integrative Plant Biology Vol. 53 No. 9 2011
M-MLV (200 U/µL). The relative expression levels of ScVPwere analyzed with semi-quantitative RT-PCR. The PCR wasperformed in a total volume of 25 µL with 30 cycles. Thefollowing gene-specific primers were used: ScVP-P1 (5′-CTGCTGGAAACACTACCGCT-3′) and ScVP-P2 (5′-CATTGTCCCAAGCACCTCCT-3′); actin-P1 (5′-TACCACATCCAAGGAAGGCA-3′) and actin-P2 (5′-ACCCAAAGTCCAACTACGAG-3′)were used to amplify the Actin gene as an internal control.
Construction of plant expression vectors andArabidopsis transformation
The complete ORF of ScVP was amplified with the primersScVP-P3 (5′-AAAAAGCAGGCTATGAGTGGCATTCTTCTTC-3′) and ScVP-P4 (5′-AgAAAGCTGGGTTTAGAAGATCTTCAAGAGCA-3′) under the following thermal cycle conditions: 94 ◦Cfor 8 min, followed by 35 cycles of 94 ◦C for 40 s, 58 ◦C for40 s, and 72 ◦C for 80 s. The PCR products were purifiedand cloned into the pDONR221 entry vector via the BP re-action (Gateway Technology, Invitrogen). For ScVP cDNAs,pENTRY clones from individual bacterial colonies were usedsubsequently in LR reactions (Invitrogen) with the expressionvector pCB35sR1R2 to generate pCB35s-ScVP (Sambrooket al. 1989) containing the BAR gene conferring resistanceto the herbicide Basta as a selectable marker. Arabidopsisthaliana (ecotype Columbia) was transformed with Agrobac-terium tumefaciens strain EHA105 (Voelker et al. 1987) car-rying the above recombinant plasmids. Transgenic plants forScVP were selected by spraying the herbicide Basta (40 mg/L)on the seedlings three times at 2 or 3 d intervals. The positiveplants were transplanted into soil and grown in a greenhouseat 24/16 ◦C (day/night) with a 16 h light/8 h dark photope-riod and 50% relative humidity. Plants were subsequentlyselected for two further generations to identify transgenicplants homozygous for the transgene. The relative expressionlevels of ScVP were analyzed with semiquantitative RT-PCR.The identity of these plantlets was confirmed by PCR usingthe primer pairs ScVP-P5 (5′-GGGCTACCTCCTTCCTTTAT-3′) and ScVP-P6 (5′-TACCTCCACCAACTCTACCA-3′). actin-P3 (5′-CGATGAAGCTCAATCCAAACG-3′) and actin-P4 (5′-TCACGACCAGCAAGATCAAG-3′) were used to amplify theArabidopsis Actin gene fragment as an internal control.
Salt, saline-alkali and drought stress treatment intransgenic Arabidopsis
For the salt and saline-alkali tolerance assay, seeds of WTand two lines (L5 and L18) of transgenic T3 plants from ScVP-transgenic Arabidopsis were germinated on MS agar medium(Murashige and Skoog 1962) under a 16 h/8 h light/darkphotoperiod at 22 ◦C and 50% relative humidity. After 10 d,young seedlings were transplanted to fresh MS agar medium
supplemented with one of four salt solutions (0 mM NaCl,100 mM NaCl, 150 mM NaCl, and 200 mM NaCl) and oneof two saline-alkali solutions (7.5 mM NaHCO3 + 92.5 mMNaCl, pH 8.0; and 10.0 mM NaHCO3 + 90 mM NaCl, pH 8.5)or onto plain MS medium as a control. The seedlings weremaintained at 22 ◦C, and their growth was monitored for 15 d.Root length of the seedlings was measured 15 d after initiationof the stress treatments. The experiments were repeated threetimes.
For the drought-stress assay, 10-d-old WT and transgenicArabidopsis plants were transplanted to fresh MS mediumsupplemented with 10% PEG8000 and grown under a 16 h/8 hlight/dark photoperiod at 25 ◦C and 50% relative humidity. Theexperiment was repeated three times. Photographs were takenof 20-d-old plants.
For germination experiments, transgenic and WT Arabidop-sis seeds were used for stress-tolerance assays. Seeds fromWT and transgenic plants were placed on MS agar platescontaining either 0 mM NaCl, 100 mM NaCl, 150 mM NaCl,200 mM NaCl, 7.5 mM NaHCO3 + 92.5 mM NaCl, 10.0 mMNaHCO3 + 90 mM NaCl, or 10% PEG8000 and germinatedat room temperature under a 16 h/8 h light/dark photoperiod.Photographs were taken of 10-d-old plants.
Measurement of Na+ content
The amounts of Na+ in Arabidopsis leaves and roots weremeasured. All leaves lacking midribs were used to measure theNa+ concentration. The roots were rinsed in deionized waterfor 10 s and washed with cold LiNO3 solution isotonic with MSmedium containing different concentrations of NaCl and saline-alkali according to the method described by Flowers et al.(1986). The leaves and roots were dried for 48 h at 70 ◦C andthe dry weight was measured. The dried leaves and roots wereextracted with 1 M HNO3 as described by Storey (1995). Thesupernatants were analyzed by atomic absorption to quantifyNa+ content.
Measurement of relative water content
The relative water content (RWC) of drought-stressed trans-genic and WT Arabidopsis plants was measured (Gaxiola et al.2001). The drought tolerance of WT and transgenic plantswas tested by growing the plants for 5 weeks under a fullywatered regime and then no further addition of water. Leavesfrom the rosette stage of development were excised and theirfresh weights were recorded immediately. After floating leavesin deionized water at 4 ◦C overnight, their rehydrated weightwas determined. Finally, they were dried in an oven at 70 ◦Covernight and weighed. Each measurement was repeatedthree times. The RWC was calculated using the formula:

Vacuolar H+-pyrophosphatase in Plant Stress Tolerance 741
RWC = (fresh weight–dry weight)/ (rehydrated weight–dryweight).
Acknowledgements
This work was supported by grants from the Program for NewCentury Excellent Talents in University (Grant No. NCET-08-0693), the Special Program for Research of Transgenic Plants(Grant No. 2008ZX08010-002), the National Natural ScienceFoundation of China (Grant Nos. 30400300 and 30971804),the Science and Technology Development Project of JilinProvince and Changchun City, China (Grant Nos. 20080252and 2009024), the Scientific Research Foundation for ReturnedOverseas Chinese Scholars, State Education Ministry, China,and the State Key Laboratory of Crop Biology at ShandongAgricultural University, China (Grant No. 2010KF02).
Received 16 Feb 2011 Accepted 16 Jun 2011
References
Amtmann A, Sanders D (1999) Mechanisms of Na+ uptake by plant
cells. Adv. Bot. Res. 29, 76–112.
Apse MP, Aharon GS, Sneddon WA, Blumwald E (1999) Salt toler-
ance conferred by overexpression of a vacuolar Na+/H+ antiport in
Arabidopsis. Science 285, 1256–1258.
Asad S, Mukhtar Z, Nazir F, Hashmi JA, Mansoor S, Zafar Y, Arshad
M (2008) Silicon carbide whisker-mediated embryogenic callus
transformation of cotton (Gossypium hirsutum L.) and regeneration
of salt tolerant plants. Mol. Biotechnol. 40, 161–169.
Ashraf M (1994) Breeding for salinity tolerance in plants. Crit. Rev.
Plant Sci. 13, 17–42.
Baltscheffsky M, Schultz A, Baltscheffsky H (1999) H+-PPases: A
tightly membrane-bound family. FEBS Lett. 457, 527–533.
Brini F, Hanin M, Mezghani I, Berkowitz GA, Masmoudi K (2007)
Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-
pyrophosphatase TVP1 improve salt and drought stress tolerance
in Arabidopsis thaliana plants. J. Exp. Bot. 58, 301–308.
Drozdowicz YM, Kissinger JC, Rea PA (2000) AVP2, a sequence-
divergent, K+-insensitive H+-translocating inorganic pyrophos-
phatase from Arabidopsis. Plant Physiol. 123, 353–362.
Felle HH (2005) pH regulation in anoxic plants. Ann. Bot. 96, 519–532.
Flowers TJ, Flowers SA, Greenway H (1986) Effects of sodium
chloride on tobacco plants. Plant Cell Environ. 9, 645–651.
Gao F, Gao Q, Duan XG, Yue GD, Yang AF, Zhang JR (2006)
Cloning of an H+-PPase gene from Thellungiella halophila and its
heterologous expression to improve tobacco salt tolerance. J. Exp.
Bot. 57, 3259–3270.
Gao JP, Chao DY, Lin HX (2007) Understanding abiotic stress tol-
erance mechanisms: Recent studies on stress response in rice.
J. Integr. Plant Biol. 49, 742–750.
Gaxiola RA, Li JS, Undurraga S, Dang LM, Allen GJ, Alper SL,
Fink GR (2001) Drought- and salt-tolerant plants result from over-
expression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. U.S.A. 98,
11444–11449.
Guo SL, Yin HB, Zhang X, Zhao FY, Li PH, Chen SH, Zhao YX, Zhang
H (2006) Molecular cloning and characterization of a vacuolar H+-
pyrophos-phatase gene, SsVP, from the halophyte Suaeda salsa
and its overexpression increases salt and drought tolerance of
Arabidopsis. Plant Mol. Biol. 60, 41–50.
Hamada A, Shono M, Xia T, Ohta M, Hayashi Y, Tanaka A,
Hayakawa T (2001) Isolation and characterization of a Na+/H+
antiporter gene from the halophyte Atriplex gmelini. Plant Mol. Biol.
46, 35–42.
Li B, Wei AY, Song CX, Li N, Zhang JR (2008) Heterologous
expression of the TsVP gene improves the drought resistance of
maize. Plant Biotechnol. J. 6, 146–159.
Li HP, Chen M, Wang BS (2004) Effect of K+ nutrition on growth and
activity of leaf tonoplast V-H+-ATPase and V-H+-PPase of Suaeda
salsa under NaCl stress. J. Integr. Plant Biol. 4, 433–440.
Li J, Yang H, Peer WA, Richter G, Blakeslee J (2005) Arabidopsis
H+-PPase AVP1 regulates auxin-mediated organ development.
Science 310, 121–125.
Lv SL, Zhang KW, Gao Q, Lian LJ, Song Y, Zhang J (2008) Overex-
pression of an H+-PPase gene from Thellungiella halophila in cotton
enhances salt tolerance and improves growth and photosynthetic
performance. Plant Cell Physiol. 49, 1150–1164.
Maeshima M (2000) Vacuolar H+-pyrphos-phatase. Biochem. Biophys.
Acta. 1465, 37–41.
Maeshima M (2001) Tonoplast transporters: Organization and function.
Annu. Rev. Plant Physiol. Plant Mol. Biol. 52, 469–497.
Mimura H, Nakanishi Y, Hirono M, Maeshima M (2004) Membrane
topology of the H+-pyrophosphatase of Streptomyces coelicolor
determined by cysteine-scanning mutagenesis. J. Biol. Chem. 279,
35106–35112.
Murashige T, Skoog F (1962) A revised medium for rapid growth
and bioassays with tobacco cultures. Physiol. Plant 15, 473–
497.
Ohta M, Hayashi Y, Nakashima A, Hamada A, Tanaka A, Naka-
mura T, Hayakawa T (2002) Introduction of a Na+/H+ antiporter
gene from Atriplex gmelini confers salt tolerance in rice. FEBS Lett.
532, 279–282.
Park S, Li J, Pittman JK, Berkowitz GA, Yang H, Undurraga S,
Morris J, Hirschi KD, Gaxiola RA (2005) Up-regulation of an H+-
pyrophosphatase (H+-PPase) as a strategy to engineer drought
resistant crop plants. Proc. Natl. Acad. Sci. U.S.A. 102, 18830–
18835.
Perez-Alfocea F, Estan MT, Caro M, Guerrier G (1993) Osmotic
adjustment in Lycopercon esculentum and L. pennelli under NaCl
and polyethylene glycol 6000 iso-osmotic stress stresses. Physiol.
Plant 87, 493–498.
Rea P, Kim Y, Sarafian V, Poole R, Davies J, Sanders D (1992)
Vacuolar H+-translocating pyrophosphatase: A new category of ion
translocase. TIBS 17, 348–352.

742 Journal of Integrative Plant Biology Vol. 53 No. 9 2011
Rea PA, Poole RJ (1993) Vacuolar H+-translocating pyrophosphatase.
Annu. Rev. Plant Physiol. Mol. Biol. 44, 157–180.
Rhoades JD, Loveday J (1990) Salinity in irrigated agriculture. In
Steward BA, Nielsen DR eds. American Society of Civil Engineers,
Irrigation of Agricultural Crops (Monograph 30). American Society
of Agronomists. pp. 1089–1142.
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning: A
Laboratory Manual, 2nd edition. New York Cold Spring Harbor.
Sarafian V, Kim Y, Poole RJ, Rea PA (1992) Molecular cloning and
sequence of cDNA encoding the pyrophosphate-energized vacuolar
membrane proton pump of Arabidopsis thaliana. Proc. Natl. Acad.
Sci. U.S.A. 89, 1775–1779.
Stitt M (1998) Pyrophosphate as an energy donor in the cytosol of plant
cells: An enigmatic alternative to ATP. Bot. Acta. 111, 167–175.
Storey R (1995) Salt tolerance, ion relations and the effects of root
medium on the response of citrus to salinity. Aust. J. Plant Physiol.
22, 101–114.
Sze H, Li X, Palmgren MG (1999) Energization of plant membranes
by H+-pumping ATPases: Regulation and biosynthesis. Plant Cell
11, 677–689.
Takasu A, Nakanishi Y, Yamauchi T, Maeshima M (1997) Analysis
of the substrate binding site and carboxyl terminal region of vac-
uolar H+-pyrophosphatase of mung bean with peptide antibodies.
J. Biochem. 122, 883–889.
Tanaka Y, Chiba K, Maeda M, Maeshima M (1993) Molecular cloning
of cDNA for vacuolar membrane proton-translocating inorganic
pyrophosphatase in Hordeum vulgare. Biochem. Biophys. Res.
Commun. 190, 1110–1114.
Torrecillas A, Guillaume C, Alarcon JJ, Ruiz-Sanchez MC (1995)
Water relations of two tomato species under water stress and
recovery. Plant Sci. 105, 169–176.
Voelker T, Sturm A, Chrispeels MJ (1987) Differences in expression
between two seed lectin alleles obtained from normal and lectin-
deficient beans are maintained in transgenic tobacco. EMBO J. 6,
3571–3577.
Yang CW, Jianaer A, Shi D C, Zhang Y, Yang YJ (2006) Ef-
fects of complex salt and alkali conditions on the germination
of seeds of Puccinellia tenuilor . Acta Prataculturae Sin. 15,
45–51.
Zhang DP, Cao BH, Jia B, Tang Q (2008) Germination and physio-
logical response of Albizia julibrissin seeds under alkali-salt stress.
Scientia Silvae Sinicae. 44, 157–161.
(Co-Editor: Zhizhong Gong)