cold plasma interactions with plants: morphing and...
TRANSCRIPT

Bioelectrochemistry 118 (2017) 100–105
Contents lists available at ScienceDirect
Bioelectrochemistry
j ourna l homepage: www.e lsev ie r .com/ locate /b ioe lechem
Cold plasma interactions with plants: Morphing and movements ofVenus flytrap and Mimosa pudica induced by argon plasma jet
Alexander G. Volkov a,⁎, Kunning G. Xu b, Vladimir I. Kolobov c,d
a Department of Chemistry, Oakwood University, Huntsville, AL 35896, USAb Mechanical and Aerospace Engineering Department, The University of Alabama in Huntsville, Huntsville, AL 35899, USAc The Center for Space Plasma and Aeronomic Research, The University of Alabama in Huntsville, Huntsville, AL 35899, USAd CFD Research Corporation, Huntsville, AL 35806, USA
Abbreviations: RONS, reactive oxygen and nitrogenspecies.⁎ Corresponding author.
E-mail address: [email protected] (A.G. Volkov).
http://dx.doi.org/10.1016/j.bioelechem.2017.07.0111567-5394/© 2017 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 6 May 2017Received in revised form 22 July 2017Accepted 24 July 2017Available online 29 July 2017
Low temperature (cold) plasma finds an increasing number of applications in biology, medicine and agriculture.In this paper, we report a new effect of plasma induced morphing and movements of Venus flytrap andMimosapudica. We have experimentally observed plasma activation of sensitive plant movements and morphing struc-tures in these plants similar to stimulation of their mechanosensors in vivo. Application of an atmospheric pres-sure argon plasma jet to the inside or outside of a lobe, midrib, or cilia in Dionaea muscipula Ellis induces trapclosing. Treatment ofMimosa pudica by plasma inducesmovements of pinnules and petioles similar to the effectsof mechanical stimulation. We have conducted control experiments and simulations to illustrate that gas flowand UV radiation associated with plasma are not the primary reasons for the observed effects. Reactive oxygenand nitrogen species (RONS) produced by cold plasma in atmospheric air appear to be the primary reason of plas-ma-induced activation of phytoactuators in plants. Some of these RONS are known to be signaling molecules,which control plants' developmental processes. Understanding these mechanisms could promote plasma-based technology for plant developmental control and future use for plant protection from pathogens. Ourwork offers new insight into mechanisms which trigger plant morphing and movement.
© 2017 Elsevier B.V. All rights reserved.
Keywords:ElectrocultureMimosa pudicaPlasmaRONSVenus flytrap
1. Introduction
The universe is composed of plasma - the most abundant yet leastunderstood state of matter. On Earth, plasma is naturally produced inthe form of lightning and the aurora borealis. Since the beginning ofthe 20th century, the effects of aurora borealis on plants and treeshave been associated with ions generated near the Earth's surfaceby atmospheric electricity. The associated research was termed“Electroculture” [1,2].
In physics and chemistry, plasma denotes a quasi-neutral mixture ofelectrons, ions and neutral species formed during gas ionization by fastfree electrons. Due to the large difference of electron and atom mass,electron mean energy (temperature) may exceed gas temperature bytwo orders of magnitude when electricity passes through a gas. Lowtemperature (cold) plasma provides a non-equilibrium catalytic envi-ronment for a variety of chemical reactions at ambient gas temperatures[3]. In biology andmedicine, plasma has a very differentmeaning as the
species; ROS, reactive oxygen
water-like liquid component of blood. In the present paper, the termplasma refers to the fourth state of matter, i.e. a partially-ionizedquasi-neutral mixture of electrons, ions, and neutral species.
In recent years, cold plasma has found an increasing number of ap-plications in biology,medicine and agriculture, particularly for the treat-ment of crops, seeds, water and soil [4]. Experimental studies havedemonstrated plasma-assisted decontamination of seeds, enhancementof seed germination, growth enhancement of plants, and reduction ofmicrobial load in food [5,6]. The majority of the reported plasma treat-ment used low-pressure radio frequency systems, which require vacu-um equipment [7,8]. Recently, atmospheric-pressure cold plasmashave been produced using plasma jets, dielectric barrier discharges(DBD) and corona discharges. Such plasmas are actively being studiedfor applications inmedicine and biotechnology due to their ability to in-duce desirable biochemical responses in living organisms, plants andseeds [9–12]. Plasma treatment induced desirable changes of develop-mental and physiological processes in plants, improving seed resistanceto stress and diseases, modifying seed coat structures, increasing thepermeability of seed coats, and stimulating seed germination and seed-ling growth for several types of commercially significant food plants forhuman and animal consumption, such as wheat, barley, tomato, soy-bean, and Arabidopsis thaliana. The observed beneficial effects of plasma

Schematic 1. Plasma jet system.
101A.G. Volkov et al. / Bioelectrochemistry 118 (2017) 100–105
arise from the cocktail of reactive neutral species, charged species (elec-trons, ions), electric fields, and ultraviolet radiation produced in the dis-charge at ambient pressure and room temperature [13].
It is believed that reactive oxygen species (ROS) and reactive nitro-gen species (RNS) created by plasma are mainly responsible for its ben-eficial effects in biology, medicine, agriculture, and food treatment [14,15]. It is known that ROS participate in plant developmental processesby acting as signalingmolecules for cell proliferation anddifferentiation,programmed cell death, seed germination, gravitropism, root hairgrowth, senescence, and pollen tube development, etc. However, thespecific mechanisms responsible for the regulatory action of ROS at var-ious plant developmental stages remain unknown [16].While RONS arerequired for many important signaling reactions in plants, they are alsotoxic by-products of aerobicmetabolism. As cold plasma becomes an at-tractive method for protection from viruses, fungi and bacteria in biolo-gy, medicine, food processing, and agriculture, it may also have sideeffects due to undesirable interactions of RONSwith ion channels,mem-brane transport and bio-tissue.
Many plants can change shape and dimensions under external per-turbations. This morphing behaviour can involve micro, macro, struc-tural, and fluidic processes. The rapid closure of the carnivorous plantDionaea muscipula Ellis (Venus flytrap) has been widely investigated[17–22]. The Venus flytrap can be closed by mechanical stimulation oftrigger hairs [23] at the inner side of a lobe or by an electrical stimula-tion between a midrib (+) and a lobe (−) [24,25] causing the lobe tochange its shape from convex to concave.
Another plant with morphing behaviour is the Mimosa pudica.Volkov et al. have previously investigated the morphing structures inM. pudica induced by electrical or mechanical stimulation [26–29].
In this paper, we report the first observations of cold plasma activa-tion of the Venus flytrap andMimosa pudicamorphing structures usingan argon plasma jet.
2. Materials and methods
2.1. Plant material
Fifty bulbs of Dionaea muscipula (Venus flytrap) were purchased forthis experimental work from Fly-Trap Farm (Supply, North Carolina).The soil was treated with distilled water. The seeds of Mimosa pudicaL. were soaked in warm water (30 °C) for 48 h. After growing for twoweeks, the seedlings were transplanted into pots and placed inside aplant growing chamber. Two-three-month-old Mimosa pudica plantswere used for the experiments. All plants were grown in well drainedpots at 22 °C with a 12:12 h light: dark photoperiod. The plants werewatered every day. Humidity averaged 45–50%. Irradiance was 700μE/m2s. All experiments were performed on healthy adult specimens.
2.2. Chemicals
Hydrogen peroxide solution was purchased from Sigma-Aldrich(USA).
2.3. Experimental apparatus
The plasma source consisted of a nested pair of quartz tubes with acentral tungsten rod and an outer steel ring (Schematic 1). The rodand ring served as the powered and grounded electrodes respectively.The outer tube (3 mm ID, 6 mm OD) was open on both ends and oneend was held in the perpendicular leg of a plastic compression Teefitting. The inner tube (1 mm ID, 2 mm OD) had a closed end and wasinserted into the outer tube through a drilled hole in the Tee fittingand seal with epoxy. The 1 mm diameter tungsten rod was then placedinside the inner tube to rest against the seal end. The grounded ringelectrode was placed near the end of the outer tube. The source designprevented arcing that can occur if the powered rod were in direct
contact with plasma. Argon gas was flown at a rate 1.55 L/min betweenthe two tubes. The argon was ionized by the strong electric fields be-tween the electrodes, and formed a plasma jet approximately 2 cmlong measured from the exit of the outer tube. The plasma was drivenwith a high voltage pulsed DC circuit consisting of a Matsusada AU-10P60 10 kV DC power supply, a IXYS PVX-4110 pulsed generator,and a SRS DG-645 delay generator. The system was operated with8 kV pulse amplitude, 8 kHz pulse frequency, 500 ns pulse width, anda ~70 ns rise time. An electrical system for monitoring the voltage andcurrent waveform contained a PR-55 high voltage probe and a Pearson411 current monitor connected to a Tektronix MDO3024 oscilloscope.The voltage had a square pulse shape with a slight sloping during therising edge. The current trace had two distinct current pulses at the ris-ing and falling fronts of the voltage with damping oscillations after themain current peaks due to both displacement and conduction currents[30].
2.4. Video recording
Digital video recorders, Sony DCR-HC36 and HD AVCHD, were usedto monitor the Venus flytraps and to collect digital images, whichwere analysed frame by frame using Sony Vegas 13 software. All exper-imental results were reproduced at least 14 times using different plants.
2.5. Software
Simulation results were performed using commercial multi-physicssoftware CFD-ACE+. Software SigmaPlot 12 (Systat Software, Inc.) wasused for statistical analysis of experimental data.
3. Results
3.1. Venus flytrap
The Venus flytrap can be closed by placing the plume of the argonplasma jet near the internal or external sides of the lobes, midrib, orcilia at about 2 cmdistance from the plant surface (Fig. 1). However, ap-plication of the argonplasma jet to the lower leaf of theVenus flytrap for40 s does not close the trap. Pure gas flow, without plasma, has no clos-ing effects on the plant. All traps closed by the plasma were completelyopen again two or three days after the plasma treatment. Thus, coldplasma did not damage the plants. The same trap-closing experimentsby plasma exposure were reproduced a second time with the sameplants within a week.
Fig. 1 illustrates the dynamics of trap closure induced by applicationof the plasma jet to an internal side of a lobe (Fig. 1a, and curve 1 on Fig.
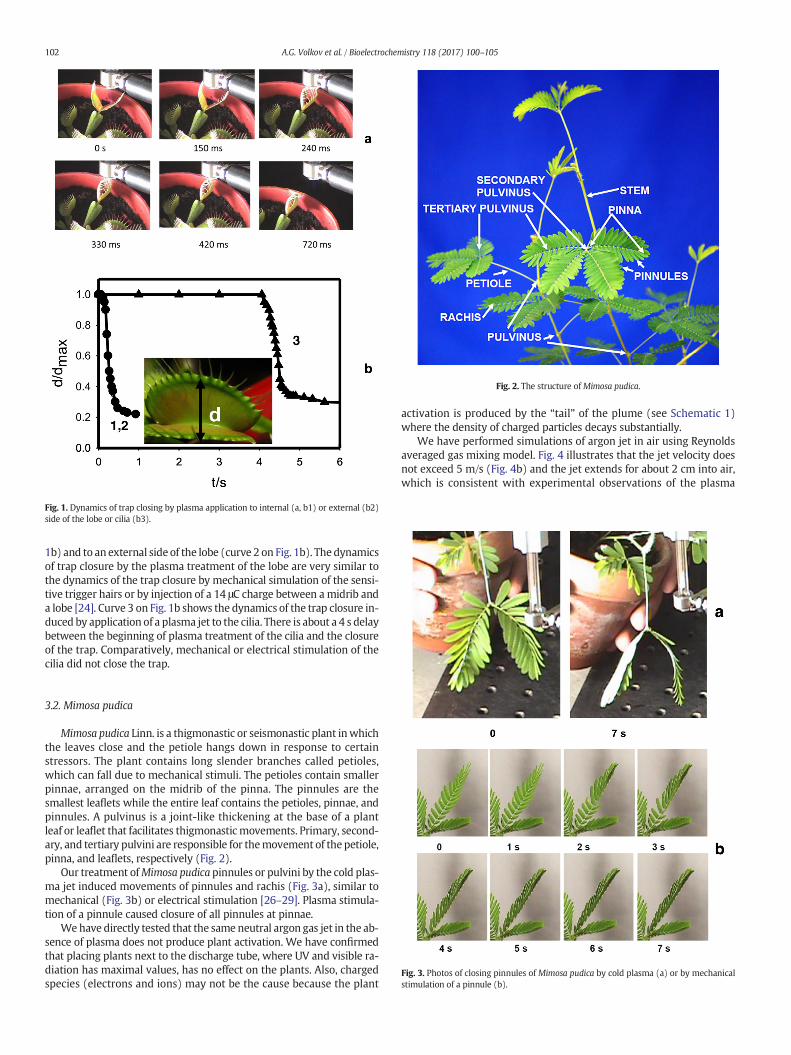
Fig. 1. Dynamics of trap closing by plasma application to internal (a, b1) or external (b2)side of the lobe or cilia (b3).
Fig. 2. The structure ofMimosa pudica.
102 A.G. Volkov et al. / Bioelectrochemistry 118 (2017) 100–105
1b) and to an external side of the lobe (curve 2 on Fig. 1b). The dynamicsof trap closure by the plasma treatment of the lobe are very similar tothe dynamics of the trap closure by mechanical simulation of the sensi-tive trigger hairs or by injection of a 14 μC charge between a midrib anda lobe [24]. Curve 3 on Fig. 1b shows the dynamics of the trap closure in-duced by application of a plasma jet to the cilia. There is about a 4 s delaybetween the beginning of plasma treatment of the cilia and the closureof the trap. Comparatively, mechanical or electrical stimulation of thecilia did not close the trap.
Fig. 3. Photos of closing pinnules of Mimosa pudica by cold plasma (a) or by mechanicalstimulation of a pinnule (b).
3.2. Mimosa pudica
Mimosa pudica Linn. is a thigmonastic or seismonastic plant inwhichthe leaves close and the petiole hangs down in response to certainstressors. The plant contains long slender branches called petioles,which can fall due to mechanical stimuli. The petioles contain smallerpinnae, arranged on the midrib of the pinna. The pinnules are thesmallest leaflets while the entire leaf contains the petioles, pinnae, andpinnules. A pulvinus is a joint-like thickening at the base of a plantleaf or leaflet that facilitates thigmonasticmovements. Primary, second-ary, and tertiary pulvini are responsible for themovement of the petiole,pinna, and leaflets, respectively (Fig. 2).
Our treatment ofMimosa pudica pinnules or pulvini by the cold plas-ma jet induced movements of pinnules and rachis (Fig. 3a), similar tomechanical (Fig. 3b) or electrical stimulation [26–29]. Plasma stimula-tion of a pinnule caused closure of all pinnules at pinnae.
We have directly tested that the same neutral argon gas jet in the ab-sence of plasma does not produce plant activation. We have confirmedthat placing plants next to the discharge tube, where UV and visible ra-diation has maximal values, has no effect on the plants. Also, chargedspecies (electrons and ions) may not be the cause because the plant
activation is produced by the “tail” of the plume (see Schematic 1)where the density of charged particles decays substantially.
We have performed simulations of argon jet in air using Reynoldsaveraged gas mixing model. Fig. 4 illustrates that the jet velocity doesnot exceed 5 m/s (Fig. 4b) and the jet extends for about 2 cm into air,which is consistent with experimental observations of the plasma

Fig. 4. Photo of argon plasma jet (a), calculated mass fraction of argon and gas flowvelocity (b) for the experimental conditions.
103A.G. Volkov et al. / Bioelectrochemistry 118 (2017) 100–105
plume emanating from the tube (Fig. 4a). Although the plasma plumeappears steady (Fig. 4a), it consists of guided ionization waves travelingalong the argon jet channel at speeds orders of magnitude faster thanthe gas flow velocity. Detailed simulations of gas breakdown, plasmaformation, propagation of the ionization waves, and electron-drivenplasma chemistry in the argon-air mixing layer are beyond the scopeof this paper. In particular plasma chemistry is rather complex includinga multitude of species and dozens of reactions [31]. Hence simulationsof plasma jets have been performed so far only with simplified chemis-tries [32,33] or with global plasma models [34].
The most likely species responsible for the observed effects of coldplasma on plants are ROS, including peroxides (H2O2). Hence, we havetested whether hydrogen peroxide can indeed induce closing of theVenus flytrap in the absence of plasma. Two 10 μL drops of H2O2 wereplaced on the midrib of the trap. After a few seconds, about 80% oftrapswere closed. Higher concentration of H2O2 producedmore closingtraps. For 0.88 M about 75% of 20 traps were closed, for 8.82 M about85% of 20 trapswere closed. Drops of distilledwater insteadof hydrogenperoxide did not close the trap. Traps activated by H2O2 not only closedbut also converted into constricted traps without further stimulation.These experiments confirmed that hydrogen peroxide (typical ROS inbiosystems) can activate morphing of the Venus flytrap similar to thatinduced by cold plasma.
4. Discussion
Why did cold plasma cause such effects on the plants? Which plas-ma components are responsible for the observed plant activation?
Our simulations of gas flow using commercial CFD software showedthat gas velocity of the argon jet does not exceed 5 m/s (Fig. 4b). Such alow flow speed cannot affect our plants. Indeed, we have directly testedthat the argon jet in the absence of plasma does not produce plantactivation.
To evaluate effects of plasma radiation, we have placed plants nextto the discharge tube, where visible and UV radiation both have maxi-mal values. No effect on the plants has been observed.
To evaluate effects of electric fields and ions, we have changed thedistance between theplant surface and the jet exit. It is known that plas-ma jets in He or Ar gas exist in the form of an ionization wave (plasmabullet) traveling with velocities in the range of 104–106 m/s, which areorders of magnitude larger than the gas flow velocities [31]. The maxi-mal densities of electrons and ions inside an argon plasma bullet mea-sured under similar conditions have maximal values of 1018 m−3 andelectron temperatures do not exceed 4.5 eV [32]. The densities ofcharged species decay rapidly outside the bullet due to volume
recombination. Thus, “plasma bullets”, i.e. guided ionization wavespropagating along the plume deliver net zero charge to the plants butcould induce electric field splashes associated with propagation of ioni-zation fronts. Since the effects are induced by the tails of the plume anddo not change significantly with decreasing distance to the plume, webelieve that electric fields are not themain cause of the observed effects.Thus, we believe that long-living active neutral species produced bycold plasma are responsible for the observed effects. The typical activespecies produced by a cold plasma jet include O, 1O2, O2
−, O3, OH,H2O2, NO, NO2, N2O5, and N2O. Most of these RONS are known to inter-act with livingmatter and have been studied by the free radical biologycommunity [33]. The actual composition of the RONS cocktail dependson the plasma gas mixture, the discharge type, and the operatingconditions.
RONS are involved in signal transduction and cell death [4,6,15]. Thespecific biological response of a plant to RONS depends on the chemicalidentity of the RONS, the intensity of the signal, sites of production, theplant developmental stage, previous stresses encountered, and interac-tions with other signaling molecules such as nitric oxide, lipid messen-gers, and plant hormones. The most important ROS radicals includesuperoxide, hydroxyl, hydroperoxyl, hydrogen peroxide, alkoxy,peroxy, and singlet oxygen. These species are cytotoxic to plants andcan lead to cell death [34]. For example, the reaction of nitric oxidewith the superoxide radical produces a short-lived oxidantperoxynitrile, which is a potent inducer of cell death [35].
Fig. 5 shows the proposedmechanism of closure of the Venus flytrapbymechanical stimulation of trigger hairs (E) or by cold plasma (F). It isknown that the lobes of the Venus flytrap move because of the waterflux across biological membranes driven by osmosis [19]. The waterflow occurs as passive diffusion across the lipid bilayer [36] facilitatedby aquaporins, which may play a pivotal role in osmoregulation inplants. The cold plasma treatment of lobes, midrib or cilia induces thetrap closure without triggering the mechanosensitive channels. In pre-vious experiments, the trap was closed by direct electrostimulationwith positive electrodes in a midrib and negative electrodes in a lobe[21–25]. Since cold plasma can close traps by stimulation of any partof the traps, no direct electrostimulation by plasma is involved. RONScan activate ion channels [37,38] and ion transport in membranes[38], which can induce osmotic water flow through aquaporins andplantmorphing [21]. Thus, RONS can be the primary reason of the trap'sclosing by cold plasma.
Plasma treatment ofMimosa pudica induces plantmovement similarto stimulation of mechanosensors by the mechanism shown in Fig. 6. Itis well known that reactive oxygen species [38] or electrical signaltransduction to pulvini [26–29] can activate cation channels in plants[38]. The activation of voltage-gated ion channels induces redistributionof ions between extensor and flexor layers of the pulvinus [26–29]. Thedifferential volume changes of the flexor and extensor sites are causedby the transport of ions accompanied by osmotic transport of water.Ion fluxes activate fast water transport through aquaporins betweenthe lower half and the upper half of the pulvinus.
5. Conclusions
We have observed for the first time that argon plasma jet inducesmorphing of Venus flytrap andMimosa pudica. We have evaluated pos-sible contributions of different plasma components on the plant activa-tion and concluded that plasma-generated ROS should be responsiblefor the observed effect. Direct experiments have confirmed that dropsof H2O2 on the midrib (common ROS in biosystems) can induce closingof the Venus flytrap similar to that induced by cold plasma.
The present work offers new insight into bioelectrochemical mecha-nisms of plant activation and movement. Cold plasma treatment oflobes, midrib or cilia induces the trap closure without triggering themechanosensitive channels.

Fig. 5.Mechanisms of the Venus flytrap activation.Morphing structures in the Venus flytrap induced by stimulation ofmechanophytosensors (left) and by direct stimulation of the trap bycold plasma (right): open trap (A); mechanosensitive trigger hair (B); closed trap (C); locked and constricted trap (D).
104 A.G. Volkov et al. / Bioelectrochemistry 118 (2017) 100–105
Further studies of the underlying mechanisms could promote plas-ma technology for plant developmental control and future use forplant protection from pathogens, as well as from biotic and abioticstresses on crop plants. Understanding plasma control of plantmorphing could help in designing adaptive structures and bioinspiredintelligent materials.
Disclosure of potential conflicts of interest
The authors declare no competing financial interests.
Fig. 6.Morphing structures inMimosa pudica induced by stimulation of mechanosensors (A) o(B).
Acknowledgements
This workwas supported in part by the Plasma Science Center oper-ating under the US Department of Energy Office of Fusion Energy Sci-ence Contract DE-SC0001939 and by the National Institutes of Healthunder grant number 1R25GM106994-01.
Author contributions
AGV, KGX and VIK designed and performed experiments. All authorsrecorded and evaluated plants responses to cold plasma activation. All
r by plasma stimulation of a pinnule or a pulvinus without stimulation of mechanosensors

105A.G. Volkov et al. / Bioelectrochemistry 118 (2017) 100–105
authors wrote the manuscript. All authors read and approved themanuscript.
References
[1] K. Lemström, Electricity in Agriculture and Horticulture, Electrician Publications,London, 1904.
[2] L.E. Murr, Plant growth responses in a stimulated electric field environment, Nature200 (1963) 490–491.
[3] Z. Machala, K. Hensen, Y. Akishev (Eds.), Plasma for Bio-decontamination, Medicineand Food Security, Springer, Berlin, 2012.
[4] N.N. Misra, O. Schlüter, P.J. Cullen (Eds.), Cold Plasma in Food and Agriculture,Elsevier, Amsterdam, 2016.
[5] C.M. Edelblute, M.A. Malik, L.C. Heller, Surface-dependent inactivation of model mi-croorganisms with shielded sliding plasma discharges and applied air flow,Bioelectrochemistry 103 (2015) 22–27.
[6] F. Van Breusegem, J.F. Dat, Reactive oxygen species in plant cell death, Plant Physiol.141 (2006) 384–390.
[7] L.K. Randeniya, G.J.J.B. de Groot, Non-thermal plasma treatment of agricultural seedsfor stimulation of germination, removal of surface contamination and other bene-fits: a review, Plasma Process. Polym. 12 (2015) 608–623.
[8] S.A. Mir, M.A. Shah, M.M. Mir, Understanding the role of plasma technology in foodindustry, Food Bioprocess Technol. (2016).
[9] R. Zhou, X. Zhang, J. Zhuang, S. Yang, K. Bazaka, K. Ostrikov, Effects of atmospheric-pressure N2, He, air, and O2 microplasmas on mung bean seed germination andseedling growth, Sci Rep 6 (2016) 32603, http://dx.doi.org/10.1038/srep32603.
[10] B. Surowsky, O. Schluter, D. Knorr, Interactions of non-thermal atmospheric pres-sure plasma with solid and liquid food systems: a review, Food Eng. Rev.(2014)http://dx.doi.org/10.1007/s12393-014-9088-5.
[11] E. Dolrzalova, P. Lukes, Membrane damage and active but nonculturable state in liq-uid cultures of Escherichia coli treated with an atmospheric pressure plasma jet,Bioelectrochemistry 103 (2015) 7–14.
[12] R. Bonascik, G. Burchhardi, K. Zocher, S. Hammerschmidt, Comparison of pulsed co-rona plasma and pulsed electric fields for the decomposition of water containingLegionella pneumophila as model organism, Bioelectrochemistry 112 (2016) 83–90.
[13] X. Lu, G.V. Naidis, M. Laroussi, S. Reuter, D.B. Graves, K. Ostrikov, Reactive species innon-equilibrium atmospheric pressure plasmas: generation, transport, and biologi-cal effects, Phys. Rep. 630 (2016) 1–84.
[14] D.B. Graves, Low temperature plasma biomedicine: a tutorial review, Phys. Plasmas21 (2014), 080901. (12 pp.).
[15] S. Gilroy, M. Białasek, N. Suzuki, M. Górecka, A.R. Devireddy, S. Karpinski, R. Mittler,ROS, calcium, and electric signals: key mediators of rapid systemic signaling inplants, Plant Physiol. 171 (2016) 1606–1615.
[16] R. Singh, P. Parihar, R.K. Mishra, D.K. Tripathi, V.P. Singh, D.K. Chauhan, S.M. Prasad,Reactive Oxygen Species (ROS): beneficial companions of plants' developmentalprocesses, Front. Plant Sci. 7 (2016)http://dx.doi.org/10.3389/fpls.2016.01299.
[17] C. Darwin, Insectivorous Plants, Murray, London, 1875.[18] F.E. Lloyd, The Carnivorous Plants, Chronica Botanica, Waltham, 1942.[19] V.S. Markin, A.G. Volkov, E. Jovanov, Active movements in plants. Mechanism of trap
closure by Dionaea muscipula Ellis, Plant Signal. Behav. 3 (2008) 778–783.
[20] M. Escalante-Perez, S. Scherzer, K.A.S. Al-Rasheid, C. Döttinger, E. Neher, R. Hedrich,Mechano-stimulation triggers turgor changes associated with trap closure in theDarwin plant Dionaea muscipula, Mol. Plant 7 (2014) 744–746.
[21] A.G. Volkov, T. Adesina, V.S. Markin, E. Jovanov, Kinetics and mechanism of Dionaeamuscipula trap closing, Plant Physiol. 146 (2008) 694–702.
[22] A.G. Volkov, V. Forde-Tuckett, M.I. Volkova, V.S. Markin, Morphing structures of theDionaea muscipula Ellis during the trap opening and closing, Plant Signal. Behav. 9(2014), e27793-1-7.
[23] A.G. Volkov, K.J. Coopwood, V.S. Markin, Inhibition of the Dionaea muscipula Ellistrap closure by ion and water channels blockers and uncouplers, Plant Sci. 175(2008) 642–649.
[24] A.G. Volkov, T. Adesina, E. Jovanov, Charge induced closing ofDionaeamuscipula Ellistrap, Bioelectrochemistry 74 (2008) 16–21.
[25] A.G. Volkov, T. Adesina, E. Jovanov, Closing of Venus flytrap by electrical stimulationof motor cells, Plant Signal. Behav. 2 (2007) 139–144.
[26] A.G. Volkov, J.C. Foster, V.S. Markin, Signal transduction in Mimosa pudica: biologi-cally closed electrical circuits, Plant Cell Environ. 33 (2010) 816–827.
[27] Volkov, J.C. Foster, K.D. Baker, V.S. Markin, Mechanical and electrical anisotropy inMimosa pudica pulvini, Plant Signal. Behav. 5 (2010) 1211–1221.
[28] A.G. Volkov, L. O'Neal, M.I. Volkova, V.S. Markin, Morphing structures and signaltransduction in Mimosa pudica L. induced by localized thermal stress, J. Plant Phys-iol. 170 (2013) 1317–1327.
[29] A.G. Volkov, J.C. Foster, T.A. Ashby, R.K. Walker, J.A. Johnson, V.S. Markin, Mimosapudica: electrical and mechanical stimulation of plant movements, Plant Cell Envi-ron. 33 (2010) 163–173.
[30] M. Laroussi, X. Lu, V. Kolobov, R. Arslanbekov, Power consideration in the pulsed di-electric barrier discharge at atmospheric pressure, J. Appl. Phys. 96 (2004)3028–3030.
[31] X. Lu, G.V. Naidis, M. Laroussi, K. Ostrikov, Guided ionization waves: theory and ex-periments, Phys. Rep. 540 (2014) 123–166.
[32] S. Hubner, S. Hofmann, E.M. van Veldhuizen, P.J. Bruggeman, Electron densities andenergies of a guided argon streamer in argon and air environments, Plasma SourcesSci. Technol. 22 (2013), 065011. (8 pp.).
[33] B. Halliwell, J.M.C. Gutteridge, Free Radicals in Biology and Medicine, 4th ed.Clarendon Press, New York, 2007.
[34] T. Karuppanapandian, J.C. Moon, C. Kim, K. Manoharan, W. Kim, Reactive oxygenspecies in plants: their generation, signal transduction, and scavengingmechanisms,Aust. J. Crop Sci. 5 (2011) 709–725.
[35] C. Szabo, C.H. Ischiropoulos, R. Radi, Peroxynitrile: biochemistry, pathology and de-velopment of therapeutics, Nat. Rev. Drug Discov. 6 (2007) 662–680.
[36] A.G. Volkov, S. Paula, D.W. Deamer, Twomechanisms of permeation of small neutralmolecules and hydrated ions across phospholipid bilayers, Bioelectrochemistry 42(1997) 153–160.
[37] S. Shabala, J. Bose, Application of non-invasive microelectrode fluxmeasurements inplant stress physiology, in: A.G. Volkov (Ed.), Plant Electrophysiology. Methods andCell Electrophysiology, Springer, Berlin 2012, pp. 91–126.
[38] V. Demidchik, Reactive oxygen species, oxidative stress and plant ion channels, in:V. Demidchik, F. Maathuis (Eds.), Ion Channels and Plant Stress Responses, Spring-er-Verlag, Berlin 2010, pp. 207–232.