colitis-associated variant of tlr2 causes impaired mucosal ......colitis-associated variant of tlr2...
TRANSCRIPT

CB
D
*ME
BmitdtlaamdnalssmtuopsrmlRtcRihnut
Twctgtrs
GASTROENTEROLOGY 2009;137:209–220
olitis-Associated Variant of TLR2 Causes Impaired Mucosal Repairecause of TFF3 Deficiency
ANIEL K. PODOLSKY,*,‡ GUIDO GERKEN,§ ANNETTE EYKING,§ and ELKE CARIO§
Department of Internal Medicine, UT Southwestern Medical Center, Dallas, Texas; ‡Gastrointestinal Unit, Center for the Study of Inflammatory Bowel Disease,assachusetts General Hospital and Harvard Medical School, Boston, Massachusetts; and §Division of Gastroenterology and Hepatology, University Hospital of
ssen, and Medical School, University of Duisburg-Essen, Essen, Germany
irimmcmmveoc
fai(pebdmotelcaipdlnfplan
gTc
BA
SIC–
ALI
MEN
TARY
TRA
CT
ACKGROUND & AIMS: Goblet cells (GC) facilitateucosal protection and epithelial barrier repair, yet the
nnate immune mechanisms that selectively drive GC func-ions have not been defined. The aim of this study was toetermine whether Toll-like receptor (TLR) 2 and modula-ion of GC-derived trefoil factor (TFF) 3 are functionallyinked in the intestine. METHODS: GC modulation wasssessed using quantitative real-time polymerase chain re-ction analysis (qRT-PCR), Western blotting, and confocalicroscopy. Dextran sulfate sodium (DSS) colitis was in-
uced in wild-type, TFF3�/�, and TLR2�/� mice. Recombi-ant TLR2 ligand or TFF3 peptide were orally administeredfter DSS termination. Caco-2 cells overexpressing full-ength TLR2 or mutant TLR2-R753Q were tested for TFF3ynthesis and functional-related effects in a wounding as-ay. RESULTS: Data from in vitro (Ls174T) and ex vivo
odels of murine and human GC reveal that TLR2 activa-ion selectively induces synthesis of TFF3. In vivo studiessing TFF3�/� or TLR2�/� mice demonstrate the ability forral treatment with a TLR2 agonist to confer antiapoptoticrotection of the intestinal mucosa against inflammatorytress-induced damage through TFF3. Recombinant TFF3escues TLR2-deficient mice from increased morbidity and
ortality during acute colonic injury. Severe ulcerative co-itis (UC) has recently been found to be associated with the753Q polymorphism of the TLR2 gene. The relevance of
he observed functional effect of TLR2 in regulating GC isonfirmed by the finding that the UC-associated TLR2-753Q variant is functionally deficient in the ability to
nduce TFF3 synthesis, thus leading to impaired woundealing. CONCLUSIONS: These data demonstrate aovel function of TLR2 in intestinal GC that links prod-cts of commensal bacteria to innate immune protec-
ion of the host via TFF3.
he intestinal epithelium is covered by a protectivemucus layer that is in continuous intimate contact
ith myriad commensal bacteria. The mucus layer isomposed predominantly of mucin glycoproteins andrefoil factor (TFF) 3 that are synthesized and secreted byoblet cells (GC) throughout the small and large intes-ines. TFF3 plays a major role in wound healing andepair of the intestinal mucosa.1 TFF3�/� mice are highly
usceptible to chemical, hypoxia, or radiation stress-nduced colonic injuries and fail to mount an effectiveepair response.2– 4 TFF3 does not exhibit intrinsic activ-ty in regulating cell proliferation but promotes essential
igration during epithelial restitution.5 Commensalsay drive GC functions by modulating synthesis of mu-
us layer components, thus maintaining mucosal ho-eostasis in the intestine.6 Alterations of the intestinalucus composition may contribute to imbalanced acti-
ation of immune responses in inflammatory bowel dis-ases (IBD).7 However, the innate immune mechanismsf beneficial commensal-host interactions8 that specifi-ally affect GC dynamics have not been elucidated yet.
Toll-like receptor 2 (TLR2), one member of the TLRamily, recognizes conserved molecular patterns associ-ted with both gram-negative and gram-positive bacteria,ncluding lipopeptides, such as synthetic Pam3CysSK4PCSK).9 TLR2 has been shown to be functionally ex-ressed in 3 out of the 4 intestinal epithelial cell lineages:nterocytes,10 Paneth cells, and enteroendocrine cells11
ut not in GC so far. Recently, progress has been made inefining TLR2-dependent defense mechanisms that helpaintain functional tight junction (TJ) barrier integrity
f the intestinal epithelial layer. TLR2 directly enhancesransepithelial resistance via protein kinase C �/� of thenterocyte barrier in vitro. Treatment with the TLR2igand PCSK protects TJ-associated integrity and de-reases intestinal permeability, leading to significantmelioration of acute dextran sulfate sodium (DSS)-nduced colonic inflammation during the recoveryhase.12,13 Mice deficient in TLR2 exhibit delayed oriminished tissue repair responses.13,14 Absence of TLR2
eads to deficient antiapoptotic protection of the intesti-al mucosa against toxic stress-induced injury, which
urther compromises TJ-associated barrier integrity anderpetuates intestinal inflammation.13 However, the mo-
ecular and cellular mechanisms of TLR2-mediated anti-poptosis in mucosal inflammation of the intestine haveot yet been further defined.
Abbreviations used in this paper: DSS, dextran sulfate sodium; GC,oblet cell; IBD, inflammatory bowel diseases; PCSK, Pam3CysSK4;FF3, trefoil factor 3 (intestinal); TJ, tight junction; UC, ulcerativeolitis; WT, wild type.
© 2009 by the AGA Institute0016-5085/09/$36.00
doi:10.1053/j.gastro.2009.03.007

lGhacapemndTmcreTt
(cTCmhmt(tnfitob
Co
t(tFLs1c
APlCEi
tlaEtpss
f
t4
BA
SIC–
ALIM
ENTA
RY
TRA
CT
210 PODOLSKY ET AL GASTROENTEROLOGY Vol. 137, No. 1
In this study, we provide evidence of an essential mo-ecular link between innate immunity and host-protective
C function. We show that the benefit of commensal-ost interaction in the intestine is through TLR2-medi-ted induction of the GC-product TFF3, which criticallyonfers antiapoptotic protection of the intestinal mucosagainst inflammatory stress-induced damage. Of note,atients affected with ulcerative colitis (UC) can developxtensive colonic disease, a condition characterized byucosal inflammation and ulceration. This severe phe-
otype has recently been associated with innate immuneysfunction through the R753Q polymorphism of theLR2 gene,15 but the underlying pathophysiology re-ained so far unresolved. The relevance of our findings is
onfirmed by showing that the R753Q mutant of TLR2esulted in reduced TFF3 and impaired healing, thusstablishing the mechanistic link to disease pathogenesis.hese findings provide a new strategy for developing
herapeutic approaches in intestinal injuries.
Materials and MethodsReagents and AntibodiesSynthetic lipopeptide Pam3Cys-SKKKKx3HCl
PCSK; lot No. L08/02) was obtained from EMC Micro-ollections GmbH (Tübingen, Germany).12 RecombinantFF3 (rTFF3) peptide was kindly provided by The GIompany, Framingham, MA. Rabbit polyclonal andouse monoclonal antisera generated against rat TFF3
ave recently been described.16 Polyclonal antibody tourine MUC2 and monoclonal antibody to pancytokera-
in were purchased from Santa Cruz BiotechnologySanta Cruz, CA), and polyclonal antibodies to cleaved/otal caspases 7, 8, and 9 were obtained from Cell Sig-aling (Danvers, MA). ZO-1 polyclonal antibody was
rom Zymed-Invitrogen (Frankfurt, Germany). Horserad-sh peroxidase-conjugated anti-rabbit and anti-mouse an-ibodies were from Amersham (Munich, Germany). Allther reagents were obtained from Sigma-Aldrich (Ham-urg, Germany), unless otherwise specified.
CellsCaco-2, IEC-6, and Ls174T cells (American Type
ulture Collection) were cultured as previously described12
r as recommended by the manufacturer, respectively.
MiceTLR2�/� (Tlr2tm1Kir;�F10 [C57BL6/J]) with wild-
ype (WT) (TLR2�/�) controls (C57BL6/J) and TFF3�/�
Tff3tm1Dkpy;�F7 [129S2/SvPaf]) with WT (TFF3�/�) con-rols (129S2/SvPaf) have previously been described.2,17
urther details can be obtained online from The Jacksonaboratory (Bar Harbor, ME). Representative allele-pecific genotyping is provided in Supplementary Figure. Mice were housed under strict specific pathogen-free
onditions (Helicobacter species-, MNV-free) at the Central �nimal Facility, University Hospital of Essen, Germany.rotocols were in compliance with German law for use of
ive animals and approved by the Institutional Animalare and Use Committee at the University Hospital ofssen and the responsible district government. For more
nformation, please see Supplementary Materials.
3D-Human Intestinal Mucosa-Like CultureModel of BiopsiesTissue samples were obtained from healthy pa-
ients undergoing complete colonoscopy for regular co-on cancer screening examinations and/or polypectomyt the Endoscopy Unit (head: M. Rünzi, MD), Klinikenssen-Süd. Informed consent was obtained from all pa-
ients before the procedure, and the protocol was ap-roved by the Human Studies Committee, Kliniken Es-en-Süd, Essen, Germany. For processing of biopsypecimens, please see Supplementary Materials.
Organ Culture of Murine Small IntestineOrgan culture of murine small intestine was per-
ormed as previously described.13
Induction of Colitis and TreatmentPlease see Supplementary Materials.
Histologic and Morphometric AnalysisPlease see Supplementary Materials.
ImmunohistochemistryPlease see Supplementary Materials.
Confocal Immunofluorescence MicroscopyPlease see Supplementary Materials.
Analysis of Apoptosis in Colonic SpecimensPlease see Supplementary Materials.
Protein Analysis by Immunoblotting andCytokine ArrayPlease see Supplementary Materials.
RNA Extraction and Real-Time PolymeraseChain Reaction AnalysisPlease see Supplementary Materials.
Plasmid Constructs and Cell TransfectionPlease see Supplementary Materials.
Restitution (Migration) in an in Vitro Modelof WoundingPlease see Supplementary Materials.
Statistical AnalysisDifferences between means were calculated using
he 2-tailed, unpaired t test (GraphPad Prism, version.03; GraphPad Software, San Diego, CA). P values of
.05 were considered as significant. All data are ex-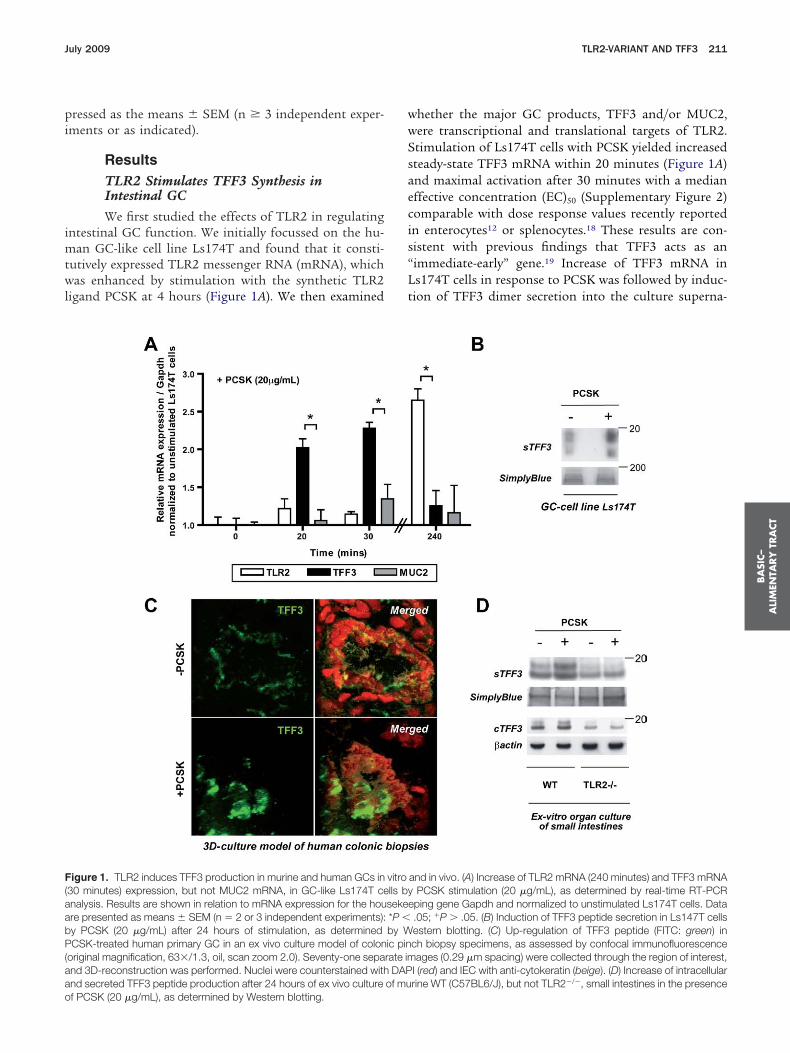
pi
imtwl
wwSsaecis“Lt
F(aabP(aao
BA
SIC–
ALI
MEN
TARY
TRA
CT
July 2009 TLR2-VARIANT AND TFF3 211
ressed as the means � SEM (n � 3 independent exper-ments or as indicated).
ResultsTLR2 Stimulates TFF3 Synthesis inIntestinal GCWe first studied the effects of TLR2 in regulating
ntestinal GC function. We initially focussed on the hu-an GC-like cell line Ls174T and found that it consti-
utively expressed TLR2 messenger RNA (mRNA), whichas enhanced by stimulation with the synthetic TLR2
igand PCSK at 4 hours (Figure 1A). We then examined
igure 1. TLR2 induces TFF3 production in murine and human GCs in30 minutes) expression, but not MUC2 mRNA, in GC-like Ls174T cenalysis. Results are shown in relation to mRNA expression for the houre presented as means � SEM (n � 2 or 3 independent experiments):y PCSK (20 �g/mL) after 24 hours of stimulation, as determinedCSK-treated human primary GC in an ex vivo culture model of colon
original magnification, 63�/1.3, oil, scan zoom 2.0). Seventy-one sepand 3D-reconstruction was performed. Nuclei were counterstained withnd secreted TFF3 peptide production after 24 hours of ex vivo culture
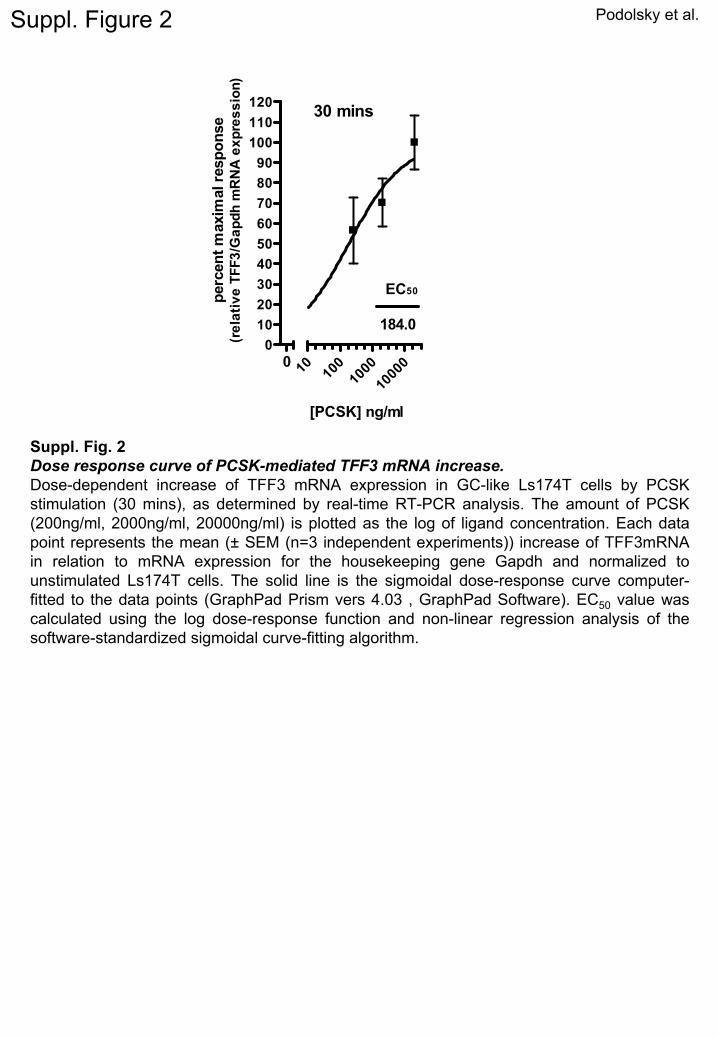
f PCSK (20 �g/mL), as determined by Western blotting.hether the major GC products, TFF3 and/or MUC2,ere transcriptional and translational targets of TLR2.timulation of Ls174T cells with PCSK yielded increasedteady-state TFF3 mRNA within 20 minutes (Figure 1A)nd maximal activation after 30 minutes with a medianffective concentration (EC)50 (Supplementary Figure 2)omparable with dose response values recently reportedn enterocytes12 or splenocytes.18 These results are con-istent with previous findings that TFF3 acts as animmediate-early” gene.19 Increase of TFF3 mRNA ins174T cells in response to PCSK was followed by induc-ion of TFF3 dimer secretion into the culture superna-
and in vivo. (A) Increase of TLR2 mRNA (240 minutes) and TFF3 mRNAPCSK stimulation (20 �g/mL), as determined by real-time RT-PCRping gene Gapdh and normalized to unstimulated Ls174T cells. Data.05; �P � .05. (B) Induction of TFF3 peptide secretion in Ls147T cellsestern blotting. (C) Up-regulation of TFF3 peptide (FITC: green) inch biopsy specimens, as assessed by confocal immunofluorescenceages (0.29 �m spacing) were collected through the region of interest,
I (red) and IEC with anti-cytokeratin (beige). (D) Increase of intracellularrine WT (C57BL6/J), but not TLR2�/�, small intestines in the presence
vitrolls bysekee*P �by Wic pinrate im
DAPof mu

ts
cipPippclswspmtees
mtiai
sTdalpphfleDostddpF
cech
BA
SIC–
ALIM
ENTA
RY
TRA
CT
212 PODOLSKY ET AL GASTROENTEROLOGY Vol. 137, No. 1
ant (Figure 1B), with similar kinetics as previously de-cribed for other stimuli of the TFF3 peptide.20
To validate the results obtained from the immortalizedell line in primary human GC, we established a novelntestinal mucosa-like culture model of human colonicinch biopsy specimens in the presence or absence ofCSK. As assessed by confocal immunofluorescence us-
ng 3-dimensional (3D) reconstruction, TFF3 peptide ex-ression was markedly enhanced in PCSK-treated humanrimary GC but attenuated in untreated (Figure 1C). Toonfirm the functional dependence between TLR2 signal-ing and PCSK-induced TFF3 synthesis, we culturedmall intestines from WT and TLR2�/� mice with orithout PCSK ex vivo and assessed levels of cellular and
ecreted TFF3 peptide. As shown in Figure 1D, murinerimary small intestinal WT-GC produced substantiallyore TFF3 after PCSK stimulation compared with un-
reated GC. In contrast, small intestinal TLR2�/� GCxpressed significantly less TFF3, which was not influ-nced by PCSK, suggesting that PCSK-induced TFF3ynthesis directly requires TLR2.
Next, we determined whether in vivo TLR2 activationay modulate GC function in the intestine. Oral PCSK
reatment of healthy mice induced GC proliferation andncreased GC size in small intestines (data not shown)nd colons via TLR2 (Supplementary Table 1A), correlat-ng with enhanced TFF3 peptide production (data not
hown). To investigate whether TLR2 may also induceFF3 under conditions of inflammatory stress-inducedamage, we used DSS-colitis, a well-established model ofcute intestinal mucosal injury. Indeed, PCSK treatmented to rapid colonic GC regeneration (Figure 2, left; Sup-lementary Table 1B), as evidenced by enhanced TFF3eptide production (Figure 2, right) during the post-DSSealing phase, which correlated with inhibition of in-ammatory destruction of the intestinal mucosa.13 How-ver, expression levels of TFF3 mRNA were elevated in allSS mice during the recovery phase (day 12), regardless
f PCSK administration (Supplementary Figure 3A), pos-ibly reflecting TLR2-mediated differences in mRNA/pro-ein stability or posttranscriptional regulation of TFF3uring GC regeneration. In contrast, TLR2 stimulationid not induce MUC2 gene transcription or protein ex-ression in vitro and in vivo (Figure 1A, Supplementaryigure 3A and B).
Antiapoptotic Efficacy of Treatment WithTLR2 Ligand in Colitis Depends on TFF3InductionInduction of antiapoptosis has been reported to
ontribute to TLR2-induced accelerated mucosal recov-ry after acute inflammatory injury,13 yet the underlyingellular mechanism has not been resolved so far. TFF3as been shown to block apoptosis, thus accelerat-
Figure 2. Oral treatment withTLR2 agonist induces GC re-generation and increases TFF3peptide expression in acute DSScolitis. DSS-colitis was inducedin WT mice (C57BL6/J) that re-ceived 2.7% DSS for 6 days, fol-lowed by oral treatment with 150�g/mL PCSK or water for 6 sub-sequent days. Mice were killedon day 12. Representative his-tology of distal DSS-colon withor without PCSK treatment isshown: left: alcian blue (AB)/pe-riodic acid-schiff (PAS); right:TFF3 (CY5: white) immunofluo-rescence assessed by confocallaser microscopy (original mag-nification, 40�/1.3, oil, scan
zoom 0.7).
icwltvDiacewdam(hsw[wmtwTTmettZctFin
bitm(fcalpCiTtTp
wi4eWaTBnc
skhTcppcacd
mtpmWovd1TscCTt
pTptdctsiD
BA
SIC–
ALI
MEN
TARY
TRA
CT
July 2009 TLR2-VARIANT AND TFF3 213
ng wound healing and repair of the intestinal mu-osa.2– 4,16,21,22 Given these properties, we investigatedhether TLR2-mediated therapeutic effects in acute co-
onic inflammation may depend functionally on induc-ion of antiapoptotic TFF3 in GC. Consistent with pre-ious findings,2 TFF3�/� mice were highly susceptible toSS-colitis with an overall mortality rate of 40% (vs 0%
n the DSS-TFF3�/� groups). Whereas PCSK treatmentmeliorated all colitis-associated signs in TFF3�/� mice,olitis in TFF3�/� mice was not influenced during thearly phase by the TLR2 ligand (Figure 3A–C). Treatmentith PCSK abolished mucosal apoptosis in DSS-TFF3�/�
uring the acute phase of colitis by day 8. However,bsence of TFF3 led to complete inhibition of TLR2-ediated antiapoptosis in acute mucosal inflammation
Figure 3A), resulting in significantly delayed mucosalealing of inflammatory stress-induced injury, as as-essed by any of the several parameters (including bodyeight, histology [Figure 3B and C], and colon length
Supplementary Figure 4A]). Increased cellular apoptosisas found throughout the lamina propria and sub-ucosa as well as surface intestinal epithelium, leading
o recruitment of prominent leukocyte infiltrationsith transmural involvement, which persisted in DSS-FF3�/� up to day 12, irrespective of PCSK therapy.hese findings indicate that TLR2 critically suppressesucosal apoptosis in acute colitis via TFF3 in GC. How-
ver, compared with untreated DSS-TFF3�/�, PCSK-reated DSS-TFF3�/� mice showed significantly less rec-al bleeding and demonstrated improved restoration ofO-1-associated barrier integrity in differentiated entero-ytes during the recovery phase— even in close proximityo inflammatory infiltrates (Figure 3D; Supplementaryigure 4B and C), suggesting that TLR2 stabilizes colitis-
nduced, TJ-associated barrier disassembly in the intesti-al epithelial cell layer independent of TFF3.
TLR2�/� Mice Exhibit a Selective Defect inTFF3 During Intestinal GC MaturationTo substantiate further the direct molecular link
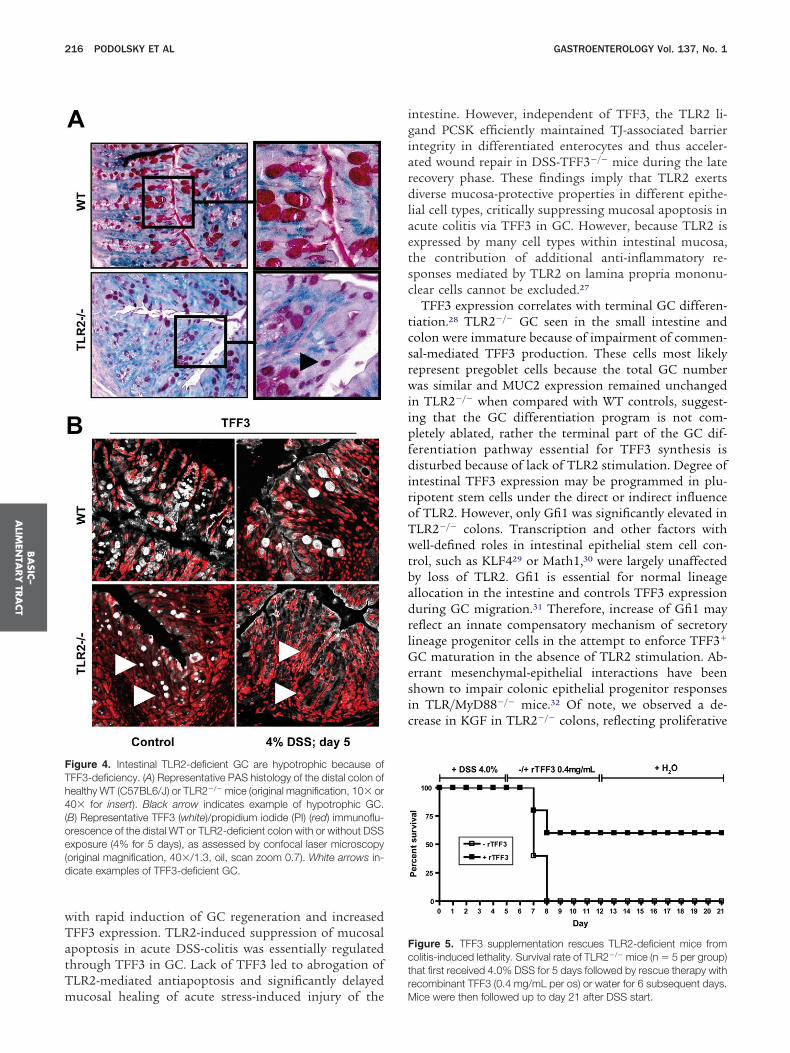
etween TLR2 and TFF3 in the intestine, we next exam-ned GC morphology and related product expression inhe absence of TLR2. Intestinal GC in healthy TLR2�/�
ice were hypotrophic along the upper half of the cryptFigure 4A), apparently lacking the ability to transformrom pre-GC to mature GC during migration from mid-rypt to villus surface. Quantative morphometry revealedlmost a one-third decrease in GC size in TLR2�/� co-ons, but no reduction in total GC number, when com-ared with TLR2�/� colons (Supplementary Table 1A).olonic TFF3 mRNA levels were decreased by almost 50%
n healthy TLR2�/� mice (Supplementary Figure 5A), andFF3 peptide expression was significantly diminished in
he upper portions of the crypt epithelium of healthyLR2�/� mice (Figure 4B). Furthermore, during the acute
hase of DSS-induced inflammation, expression of TFF3 pas almost completely lost on mRNA and peptide levelsn the absence, but not in the presence, of TLR2 (FigureB; Supplementary Figure 5A). In contrast, MUC2 mRNAxpression in TLR2�/� mice was comparable with control
T and MUC2 protein was abundantly expressed in GClong the crypt-villus axis in both healthy and inflamedLR2�/� and WT colons (Supplementary Figure 5A and). There were no microscopic signs of inflammation ando evidence of infection, essentially excluding secondaryauses of GC hypoplasia and TFF3 deficiency.
We considered whether perturbation of stem cell tran-criptional factors or associated stromal mediatorsnown to be involved in terminal GC differentiation mayave contributed to the observed GC phenotype inLR2�/�. We observed only a statistically significant in-rease in Gfi-1 mRNA levels in healthy TLR2�/� com-ared with WT colons, whereas the mRNA levels of otherrogenitor/postmeitotic GC regulators remained un-hanged (Supplementary Table 2). However, baseline ker-tinocyte growth factor (KGF) was decreased, potentiallyontributing secondarily to aberrant GC-specific lineageifferentiation in the absence of TLR2.
TFF3 Supplementation RescuesTLR2–Deficient Mice From TissueInjury-Induced LethalityWe and others have recently demonstrated that
ice deficient in TLR2 exhibit severe morbidity and mor-ality during the acute phase of DSS-colitis.13,14 We hy-othesized that increased tissue injury-induced lethalityay be caused by lack of TFF3 in the absence of TLR2.e therefore assessed the effect of oral supplementation
f recombinant TFF3 (rTFF3 [0.4 mg/mL]4) on the sur-ival rate of TLR2�/� mice after DSS exposure (4%; 5ays). As shown in Figure 5, the overall mortality rate was00% in DSS-TLR2�/� control mice vs only 40% in DSS-LR2�/� mice that received rTFF3, implying that TFF3
upplementation can rescue TLR2�/� mice from earlyolitis-induced wasting disease and subsequent death.onsistent with previous findings,13 the mortality rate ofLR2�/� controls that did not receive rTFF3 was less
han 10% (data not shown).
rTFF3 Ameliorates Colitis in TLR2-DeficientMice by Suppressing Mucosal ApoptosisWe hypothesized that the selective lack in TFF3
roduction because of impaired GC maturation inLR2�/� is an important contributing factor for im-aired healing of the intestinal mucosa after inflamma-ory stress-induced damage. To investigate further theisease course during mucosal restitution, we lowered theoncentration of DSS (2.0% for 6 days) to reduce itsoxicity in TLR2�/� mice. DSS-alone TLR2�/� mice stillhowed marked colitis-associated signs, whereas admin-stration of rTFF3 significantly ameliorated all signs of
SS-induced colonic inflammation (Figure 6A–C; Sup-
lementary Figure 6A–C). Mucosal cells in DSS-TLR2�/�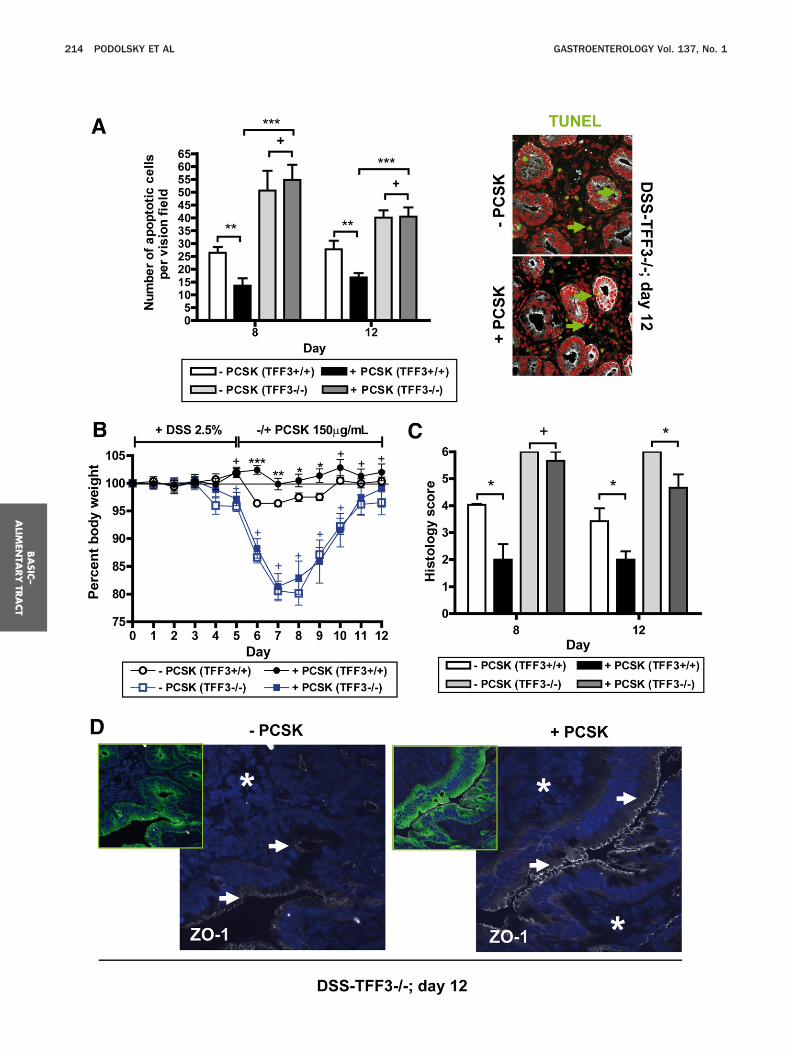
BA
SIC–
ALIM
ENTA
RY
TRA
CT
214 PODOLSKY ET AL GASTROENTEROLOGY Vol. 137, No. 1

micEilflRiTc
ltpelptTTacs(bemcspmpmtwctpsm
ClTrwR
dmtiiwtcTnt
sttMsscilektaT
tsdibmdm
4F(w(i(Eoac
BA
SIC–
ALI
MEN
TARY
TRA
CT
July 2009 TLR2-VARIANT AND TFF3 215
ice underwent mitochondrial-dependent apoptosis vianitiation of the apoptosome-catalyzed caspase-9¡7-cas-ade (intrinsic cell death pathway) (Figure 6C and D).xcessive apoptosis in DSS-TLR2�/� mice coincided with
ncreased chemokine expression, peak migration, and de-ayed clearance of leukocytes at the site of mucosal in-ammation (Supplementary Table 3; Figure 6B and C).emarkably, TFF3 supplementation completely abol-
shed ongoing caspase-dependent apoptosis in DSS-LR2�/� mice and consequently reduced apoptosis-asso-iated inflammatory recruitment of leukocytes.
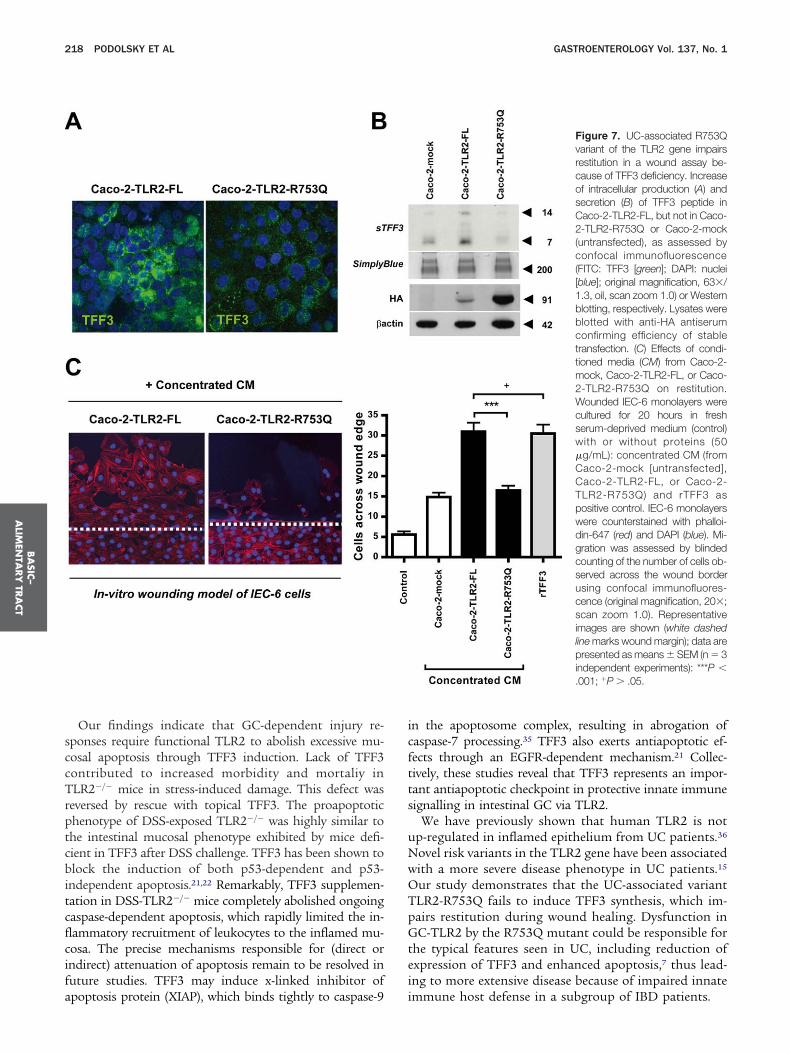
UC-Associated R753Q Variant of the TLR2Gene Impairs Wound Healing Because ofTFF3 DeficiencyUp to 40% of UC patients develop extensive co-
onic disease, called pancolitis,7 which still represents aherapeutic challenge for the clinician. This more severehenotype has recently been associated with the het-rozygous TLR2-R753Q polymorphism,15 but the under-ying pathogenesis remained so far unresolved. TFF3romotes essential migration during epithelial restitu-ion.5 We therefore hypothesized that this UC-associatedLR2 variant delays wound repair by failing to induceFF3 synthesis, thus leading to disease exacerbation. Toddress this, we stably transfected the intestinal epithelialell line Caco-2 (which inducibly express TFF323) withelf-active24 HA-tagged plasmids of TLR2 full-lengthTLR2-FL) or TLR2 mutant (TLR2-R753Q) and assessedasal production of TFF3 and its functional effects onpithelial cell migration in an in vitro wound healingodel of IEC-6 monolayers. Overexpression of Caco-2
ells with TLR2-FL led to significant induction of cellularynthesis (Figure 7A) and constitutive secretion of TFF3eptide into the culture supernatant (Figure 7B), whicharkedly promoted restitution (Figure 7C). Other factors
resent in the conditioned media of Caco-2-TLR2-FLay have secondarily contributed to increased IEC-6 res-
itution, but the degree of migration was comparableith that observed of rTFF35 using the same protein
oncentration. In contrast, Caco-2 cells overexpressinghe TLR2-R753Q mutant did not lead to intracellularroduction (Figure 7A) or secretion of TFF3 into theupernatants (Figure 7B), which impaired epithelial cell
igration (Figure 7C), comparable with untransfected
™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™igure 3. Therapeutic efficacy of TLR2 agonist in acute inflammatory
129S2/SvPaf) (n � 10 per group) received 2.5% DSS for 5 days followeere killed on days 8 or 12. Data are presented as means � SEM. (�P
FITC: green) assay of colonic tissues using confocal immunofluorescenn TFF3�/� mice. Cells were counterstained with propidium iodide (PI) fororiginal magnification 40�/1.3, scan zoom 0.7). Representative immvolution of (B) body weight changes and (C) histology scores during Df ZO-1 (CY5: white)-associated IEC barrier integrity after PCSK treatmenalysis (original magnification, 40�/1.3 oil, scan zoom 1.0). Arrow ind
ounterstained with DAPI for nuclei (blue) and anti-pancytokeratin (FITC: greaco-2 cells (mock). Although stable protein expressionevels of HA-tagged full-length were lower than mutantLR2, transfection efficiency was sufficient to initiate
obust TFF3 production in Caco-2-TLR2-FL cells, whichas neither evident in Caco-2-mock nor in Caco-2-TLR2-753Q cells.
DiscussionCommensals serve as an important stimulus for
iverse innate immune functions that protect intestinalucosal homeostasis. TLRs are involved in host recogni-
ion and response to commensals and play a key role innnate immunity of the gastrointestinal tract. Here, wedentify a previously unappreciated TLR2-regulated path-ay necessary for induction of TFF3 synthesis during
erminal GC maturation, thus critically balancing mu-osal homeostasis against proinflammatory apoptosis.hese studies provide a molecular link between the in-ate immune system and commensal-mediated modula-ion of GC-derived TFF3 in the intestine.
Our data from in vitro and ex vivo studies indicate thattimulation of TLR2 with PCSK promptly induces sus-ained TFF3 synthesis in murine and human GC. TFF3ranscriptional activation occurs through Ras/MEK/
APK and PI3K/Akt pathways,19,25 which are bothhared by TLR2 signaling.10,13 Downstream, TFF3 tran-cription is regulated through induction of the gobletell–specific transcription factor GCSI-BP.26 Future stud-es will need to determine whether TLR2-induced signal-ing modules may recruit GCSI-BP or other regulatorylements that may distinctly drive the expression of thisey GC product. TLR2 did not modulate MUC2 generanscription in vitro and in vivo, suggesting a selectivend direct GC-regulatory effect on TFF3 synthesis byLR2 in the intestine.Furthermore, our findings demonstrate that TLR2 ac-
ivation distinctly modulates GC function in vivo. Aignificant increase in TFF3-producing GC via TLR2 wasetected in PCSK-treated intestine. The specific and anti-
nflammatory effects of PCSK via TLR2 have previouslyeen shown,13 yet the full array of cell-specific protectiveechanisms of TLR2 remains to be resolved. We now
emonstrate that PCSK-mediated inhibition of inflam-atory destruction of the intestinal mucosa correlated
™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™is dependent on antiapoptotic TFF3. (A–D) TFF3�/� or TFF3�/� mice
treatment with PCSK 150 �g/mL or water for 7 subsequent days. Mice) vs (�PCSK): *P � .05; **P � .01; ***P � .001; �P � .05. (A) TUNELday 12 after DSS exposure with or without subsequent PCSK therapy
ei (rhodamine: red) and anti-pancytokeratin (CY5: white) specific for IECuorescent images are shown; green arrows indicate apoptotic cells.litis and subsequent treatment with or without PCSK. (D) PreservationSS-TFF3�/� on day 12, as assessed by confocal immunofluorescenceZO-1 staining and star subepithelial infiltrate, respectively. Cells were
™™™injuryd byCSK
ce onnuclunoflSS cont in Dicates
en), as presented in inserts.
wTatTm
igiardlaetsc
tcsrwiipfdiroTwtbadrlGesic
FTh4(oe(d
Fctr
BA
SIC–
ALIM
ENTA
RY
TRA
CT
216 PODOLSKY ET AL GASTROENTEROLOGY Vol. 137, No. 1
ith rapid induction of GC regeneration and increasedFF3 expression. TLR2-induced suppression of mucosalpoptosis in acute DSS-colitis was essentially regulatedhrough TFF3 in GC. Lack of TFF3 led to abrogation ofLR2-mediated antiapoptosis and significantly delayed
igure 4. Intestinal TLR2-deficient GC are hypotrophic because ofFF3-deficiency. (A) Representative PAS histology of the distal colon ofealthy WT (C57BL6/J) or TLR2�/� mice (original magnification, 10� or0� for insert). Black arrow indicates example of hypotrophic GC.
B) Representative TFF3 (white)/propidium iodide (PI) (red) immunoflu-rescence of the distal WT or TLR2-deficient colon with or without DSSxposure (4% for 5 days), as assessed by confocal laser microscopyoriginal magnification, 40�/1.3, oil, scan zoom 0.7). White arrows in-icate examples of TFF3-deficient GC.
ucosal healing of acute stress-induced injury of the M
ntestine. However, independent of TFF3, the TLR2 li-and PCSK efficiently maintained TJ-associated barrierntegrity in differentiated enterocytes and thus acceler-ted wound repair in DSS-TFF3�/� mice during the lateecovery phase. These findings imply that TLR2 exertsiverse mucosa-protective properties in different epithe-
ial cell types, critically suppressing mucosal apoptosis incute colitis via TFF3 in GC. However, because TLR2 isxpressed by many cell types within intestinal mucosa,he contribution of additional anti-inflammatory re-ponses mediated by TLR2 on lamina propria mononu-lear cells cannot be excluded.27
TFF3 expression correlates with terminal GC differen-iation.28 TLR2�/� GC seen in the small intestine andolon were immature because of impairment of commen-al-mediated TFF3 production. These cells most likelyepresent pregoblet cells because the total GC numberas similar and MUC2 expression remained unchanged
n TLR2�/� when compared with WT controls, suggest-ng that the GC differentiation program is not com-letely ablated, rather the terminal part of the GC dif-erentiation pathway essential for TFF3 synthesis isisturbed because of lack of TLR2 stimulation. Degree of
ntestinal TFF3 expression may be programmed in plu-ipotent stem cells under the direct or indirect influencef TLR2. However, only Gfi1 was significantly elevated inLR2�/� colons. Transcription and other factors withell-defined roles in intestinal epithelial stem cell con-
rol, such as KLF429 or Math1,30 were largely unaffectedy loss of TLR2. Gfi1 is essential for normal lineagellocation in the intestine and controls TFF3 expressionuring GC migration.31 Therefore, increase of Gfi1 mayeflect an innate compensatory mechanism of secretoryineage progenitor cells in the attempt to enforce TFF3�
C maturation in the absence of TLR2 stimulation. Ab-rrant mesenchymal-epithelial interactions have beenhown to impair colonic epithelial progenitor responsesn TLR/MyD88�/� mice.32 Of note, we observed a de-rease in KGF in TLR2�/� colons, reflecting proliferative
igure 5. TFF3 supplementation rescues TLR2-deficient mice fromolitis-induced lethality. Survival rate of TLR2�/� mice (n � 5 per group)hat first received 4.0% DSS for 5 days followed by rescue therapy withecombinant TFF3 (0.4 mg/mL per os) or water for 6 subsequent days.
ice were then followed up to day 21 after DSS start.
aefimwT
daStt
Frs�
cswa1w
BA
SIC–
ALI
MEN
TARY
TRA
CT
July 2009 TLR2-VARIANT AND TFF3 217
lteration in the pericryptal mesenchyme. KGF is anndogenous paracrine effector synthesized by stromalbroblasts in the colonic stem cell niche.33 KGF regulatesouse TFF3 transcription through the GCSI element,hich is essential for goblet cell–specific expression of
igure 6. Administration of rTFF3 ameliorates acute DSS colitis in TLRecruitment. TLR2�/� mice (n � 8 per group) received 2.0% DSS forubsequent days. Mice were killed on days 8 or 12. Data are presentedP � .05. (A) Evolution of body weight during DSS colitis and subsequeolon with or without rTFF3 treatment on day 12 (H&E staining on frozentar indicates ulcer. (C) TUNEL (FITC: green) assay of distal colonic tissuith or without subsequent rTFF3 therapy in TLR2�/� mice. Cells wernti-pancytokeratin (CY5: white) specific for IEC (original magnification, 42) are shown; arrows indicate apoptotic cells. (D) Assessment of cleavith or without rTFF3 treatment on day 12 by Western blotting.
FF3.26 Reduction of homeostatic KGF in the TLR2- t
eficient intestinal mucosa may have contributed second-rily to the defect in GC differentiation. The IL-6/Gp130/TAT-pathway has also been linked to TFF3 produc-ion,34 but we were also not able to detect any modula-ion of baseline interleukin-6 in healthy TLR2�/� intes-
cient mice by abolishing mucosal apoptosis and associated leukocyteys followed by treatment with rTFF3 (0.4 mg/mL p.o.) or water for 6eans � SEM. (�rTFF3) vs (�rTFF3): *P � .05; **P � .01; ***P � .001;pplementation of rTFF3. (B) Representative histology of the distal DSSons [original magnification, 2.5�]). Arrows indicate leukocyte infiltrates;ing confocal immunofluorescence on days 8 or 12 after DSS exposurenterstained with propidium iodide (PI) for nuclei (rhodamine: red) and.3, oil, scan zoom 0.7). Representative immunofluorescent images (dayf caspases 9, 8, and 7 in whole distal colonic tissues of DSS-TLR2�/�
2-defi6 daas mnt susecties use cou0�/1age o
ines.
sccTrptcbitcflcifa
icftts
uNwOTpGtei
BA
SIC–
ALIM
ENTA
RY
TRA
CT
218 PODOLSKY ET AL GASTROENTEROLOGY Vol. 137, No. 1
Our findings indicate that GC-dependent injury re-ponses require functional TLR2 to abolish excessive mu-osal apoptosis through TFF3 induction. Lack of TFF3ontributed to increased morbidity and mortaliy inLR2�/� mice in stress-induced damage. This defect was
eversed by rescue with topical TFF3. The proapoptotichenotype of DSS-exposed TLR2�/� was highly similar tohe intestinal mucosal phenotype exhibited by mice defi-ient in TFF3 after DSS challenge. TFF3 has been shown tolock the induction of both p53-dependent and p53-
ndependent apoptosis.21,22 Remarkably, TFF3 supplemen-ation in DSS-TLR2�/� mice completely abolished ongoingaspase-dependent apoptosis, which rapidly limited the in-ammatory recruitment of leukocytes to the inflamed mu-osa. The precise mechanisms responsible for (direct orndirect) attenuation of apoptosis remain to be resolved inuture studies. TFF3 may induce x-linked inhibitor of
poptosis protein (XIAP), which binds tightly to caspase-9 in the apoptosome complex, resulting in abrogation ofaspase-7 processing.35 TFF3 also exerts antiapoptotic ef-ects through an EGFR-dependent mechanism.21 Collec-ively, these studies reveal that TFF3 represents an impor-ant antiapoptotic checkpoint in protective innate immuneignalling in intestinal GC via TLR2.
We have previously shown that human TLR2 is notp-regulated in inflamed epithelium from UC patients.36
ovel risk variants in the TLR2 gene have been associatedith a more severe disease phenotype in UC patients.15
ur study demonstrates that the UC-associated variantLR2-R753Q fails to induce TFF3 synthesis, which im-airs restitution during wound healing. Dysfunction inC-TLR2 by the R753Q mutant could be responsible for
he typical features seen in UC, including reduction ofxpression of TFF3 and enhanced apoptosis,7 thus lead-ng to more extensive disease because of impaired innate
Figure 7. UC-associated R753Qvariant of the TLR2 gene impairsrestitution in a wound assay be-cause of TFF3 deficiency. Increaseof intracellular production (A) andsecretion (B) of TFF3 peptide inCaco-2-TLR2-FL, but not in Caco-2-TLR2-R753Q or Caco-2-mock(untransfected), as assessed byconfocal immunofluorescence(FITC: TFF3 [green]; DAPI: nuclei[blue]; original magnification, 63�/1.3, oil, scan zoom 1.0) or Westernblotting, respectively. Lysates wereblotted with anti-HA antiserumconfirming efficiency of stabletransfection. (C) Effects of condi-tioned media (CM) from Caco-2-mock, Caco-2-TLR2-FL, or Caco-2-TLR2-R753Q on restitution.Wounded IEC-6 monolayers werecultured for 20 hours in freshserum-deprived medium (control)with or without proteins (50�g/mL): concentrated CM (fromCaco-2-mock [untransfected],Caco-2-TLR2-FL, or Caco-2-TLR2-R753Q) and rTFF3 aspositive control. IEC-6 monolayerswere counterstained with phalloi-din-647 (red) and DAPI (blue). Mi-gration was assessed by blindedcounting of the number of cells ob-served across the wound borderusing confocal immunofluores-cence (original magnification, 20�;scan zoom 1.0). Representativeimages are shown (white dashedline marks wound margin); data arepresented as means � SEM (n � 3independent experiments): ***P �.001; �P � .05.
mmune host defense in a subgroup of IBD patients.

sdiocTatrpfTnpcgsotTatmIicTb
aG1
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
BA
SIC–
ALI
MEN
TARY
TRA
CT
July 2009 TLR2-VARIANT AND TFF3 219
In summary, these studies provide first evidence that apecific TLR, namely TLR2, acts to control terminal GCifferentiation by selectively regulating TFF3 expression
n the intestine, thus conferring antiapoptotic protectionf the intestinal mucosa. They suggest that TLR2 defi-iency results in an innate immune defect of GC-derivedFF3, contributing to exacerbation of mucosal apoptosisnd associated leukocyte influx during acute inflamma-ory stress-induced damage of the intestine, which can beeversed by supplementation with recombinant TFF3eptide. They also demonstrate that cell type–specificunctional differences in mucosa-protective effects viaLR2 exist within the intestinal epithelial lineages. Fi-ally, the more severe disease phenotype seen in UCatients with the TLR2-mutant haplotype is pathogeneti-ally linked to intestinal TFF3 reduction. As recognitionrows for TLRs to play a major role in IBD pathogenesis,7
ignificant efforts have begun to find a cellular approachf therapeutic TLR manipulation in the gastrointestinalract.37 Our results suggest that specifically targetingLR2 in intestinal GC could help in the design of andjuvant therapeutic means by enhancing cell survivalhrough TFF3 induction, thus protecting the inflamed
ucosa during acute gastrointestinal injuries, such asBD, but potentially also other causes of damage, includ-ng, eg, radiation- or chemotherapy-induced mucositis. Inonclusion, these data demonstrate a novel function ofLR2 in intestinal GC that links products of commensalacteria to innate immune protection of the host.
Supplementary Data
Note: To access the supplementary materialccompanying this article, visit the online version ofastroenterology at www.gastrojournal.org, and at doi:0.1053/j.gastro.2009.03.007.
References
1. Taupin D, Podolsky DK. Trefoil factors: initiators of mucosalhealing. Nat Rev Mol Cell Biol 2003;4:721–732.
2. Mashimo H, Wu DC, Podolsky DK, et al. Impaired defense ofintestinal mucosa in mice lacking intestinal trefoil factor. Science1996;274:262–265.
3. Furuta GT, Turner JR, Taylor CT, et al. Hypoxia-inducible factor1-dependent induction of intestinal trefoil factor protects barrierfunction during hypoxia. J Exp Med 2001;193:1027–1034.
4. Beck PL, Wong JF, Li Y, et al. Chemotherapy- and radiotherapy-induced intestinal damage is regulated by intestinal trefoil factor.Gastroenterology 2004;126:796–808.
5. Dignass A, Lynch-Devaney K, Kindon H, et al. Trefoil peptidespromote epithelial migration through a transforming growth factor�-independent pathway. J Clin Invest 1994;94:376–383.
6. Hooper LV, Wong MH, Thelin A, et al. Molecular analysis ofcommensal host-microbial relationships in the intestine. Science2001;291:881–884.
7. Xavier RJ, Podolsky DK. Unravelling the pathogenesis of inflam-
matory bowel disease. Nature 2007;448:427–434.8. Cario E. Innate immune signalling at intestinal mucosal surfaces:a fine line between host protection and destruction. Curr OpinGastroenterol 2008;24:725–732.
9. Jin MS, Kim SE, Heo JY, et al. Crystal structure of the TLR1-TLR2heterodimer induced by binding of a tri-acylated lipopeptide. Cell2007;130:1071–1082.
0. Cario E, Rosenberg IM, Brandwein SL, et al. Lipopolysaccharideactivates distinct signaling pathways in intestinal epithelial celllines expressing Toll-like receptors. J Immunol 2000;164:966–972.
1. Palazzo M, Balsari A, Rossini A, et al. Activation of enteroendo-crine cells via TLRs induces hormone, chemokine, and defensinsecretion. J Immunol 2007;178:4296–4303.
2. Cario E, Gerken G, Podolsky DK. Toll-like receptor 2 enhancesZO-1-associated intestinal epithelial barrier integrity via proteinkinase C. Gastroenterology 2004;127:224–238.
3. Cario E, Gerken G, Podolsky DK. Toll-like receptor 2 controlsmucosal inflammation by regulating epithelial barrier function.Gastroenterology 2007;132:1359–1374.
4. Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, et al. Recogni-tion of commensal microflora by toll-like receptors is required forintestinal homeostasis. Cell 2004;118:229–241.
5. Pierik M, Joossens S, Van Steen K, et al. Toll-like receptor-1, -2,and -6 polymorphisms influence disease extension in inflamma-tory bowel diseases. Inflamm Bowel Dis 2006;12:1–8.
6. Itoh H, Beck PL, Inoue N, et al. A paradoxical reduction insusceptibility to colonic injury upon targeted transgenic ablationof goblet cells. J Clin Invest 1999;104:1539–1547.
7. Wooten RM, Ma Y, Yoder RA, et al. Toll-like receptor 2 is requiredfor innate, but not acquired, host defense to Borrelia burgdorferi.J Immunol 2002;168:348–355.
8. Buwitt-Beckmann U, Heine H, Wiesmuller KH, et al. TLR1- andTLR6-independent recognition of bacterial lipopeptides. J BiolChem 2006;281:9049–9057.
9. Taupin D, Wu DC, Jeon WK, et al. The trefoil gene family arecoordinately expressed immediate-early genes: EGF receptor-and MAP kinase-dependent interregulation. J Clin Invest 1999;103:R31–R38.
0. Paulsen FP, Woon CW, Varoga D, et al. Intestinal trefoil factor/TFF3 promotes re-epithelialization of corneal wounds. J BiolChem 2008;283:13418–13427.
1. Taupin DR, Kinoshita K, Podolsky DK. Intestinal trefoil factorconfers colonic epithelial resistance to apoptosis. Proc Natl AcadSci U S A 2000;97:799–804.
2. Kinoshita K, Taupin DR, Itoh H, et al. Distinct pathways of cellmigration and antiapoptotic response to epithelial injury: struc-ture-function analysis of human intestinal trefoil factor. Mol CellBiol 2000;20:4680–4690.
3. Gustafsson AC, Kupershmidt I, Edlundh-Rose E, et al. Globalgene expression analysis in time series following N-acetyl L-cysteine induced epithelial differentiation of human normal andcancer cells in vitro. BMC Cancer 2005;5:75.
4. Chang YJ, Wu MS, Lin JT, et al. Induction of cyclooxygenase-2overexpression in human gastric epithelial cells by Helicobacterpylori involves TLR2/TLR9 and c-Src-dependent nuclear factor-�Bactivation. Mol Pharmacol 2004;66:1465–1477.
5. Durual S, Blanchard C, Estienne M, et al. Expression of humanTFF3 in relation to growth of HT-29 cell subpopulations: in-volvement of PI3-K but not STAT6. Differentiation 2005;73:36–44.
6. Iwakiri D, Podolsky DK. A silencer inhibitor confers specific ex-pression of intestinal trefoil factor in gobletlike cell lines. Am JPhysiol Gastrointest Liver Physiol 2001;280:G1114–G1123.
7. Cario E. Barrier-protective function of intestinal epithelial Toll-like
receptor 2. Mucosal Immunol 2008;(Suppl 1):S62–S66.
2
2
3
3
3
3
3
3
3
3
R
GIe
A
okDkE
C
ed
F
I
BA
SIC–
ALIM
ENTA
RY
TRA
CT
220 PODOLSKY ET AL GASTROENTEROLOGY Vol. 137, No. 1
8. Iwakiri D, Podolsky DK. Keratinocyte growth factor promotesgoblet cell differentiation through regulation of goblet cell si-lencer inhibitor. Gastroenterology 2001;120:1372–1380.
9. Katz JP, Perreault N, Goldstein BG, et al. The zinc-finger transcrip-tion factor Klf4 is required for terminal differentiation of gobletcells in the colon. Development 2002;129:2619–2628.
0. Yang Q, Bermingham NA, Finegold MJ, et al. Requirement ofMath1 for secretory cell lineage commitment in the mouse intes-tine. Science 2001;294:2155–2158.
1. Shroyer NF, Wallis D, Venken KJ, et al. Gfi1 functions down-stream of Math1 to control intestinal secretory cell subtypeallocation and differentiation. Genes Dev 2005;19:2412–2417.
2. Pull SL, Doherty JM, Mills JC, et al. Activated macrophages are anadaptive element of the colonic epithelial progenitor niche nec-essary for regenerative responses to injury. Proc Natl Acad SciU S A 2005;102:99–104.
3. Housley RM, Morris CF, Boyle W, et al. Keratinocyte growth factorinduces proliferation of hepatocytes and epithelial cells throughoutthe rat gastrointestinal tract. J Clin Invest 1994;94:1764–1777.
4. Tebbutt NC, Giraud AS, Inglese M, et al. Reciprocal regulation ofgastrointestinal homeostasis by SHP2 and STAT-mediated trefoilgene activation in gp130 mutant mice. Nat Med 2002;8:1089–1097.
5. Twiddy D, Cohen GM, Macfarlane M, et al. Caspase-7 is directlyactivated by the approximately 700-kDa apoptosome complexand is released as a stable XIAP-caspase-7 approximately 200-kDa complex. J Biol Chem 2006;281:3876–3888.
6. Cario E, Podolsky DK. Differential alteration in intestinal epithe-lial cell expression of toll-like receptor 3 (TLR3) and TLR4 ininflammatory bowel disease. Infect Immun 2000;68:7010–
7017. (7. Kanzler H, Barrat FJ, Hessel EM, et al. Therapeutic targeting ofinnate immunity with Toll-like receptor agonists and antagonists.Nat Med 2007;13:552–559.
Received January 13, 2009. Accepted March 10, 2009.
eprint requestsAddress requests for reprints to: Elke Cario, MD, Division of
astroenterology and Hepatology, University Hospital of Essen,nstitutsgruppe I, Virchowstr. 171; D-45147 Essen, Germany. e-mail:[email protected]; fax: (49) 201 723 4527.
cknowledgmentsThe authors thank The GI Company (Framingham, MA) for the gift
f recombinant TFF3, Dr Michael Rünzi (Kliniken Essen-Süd) forindly providing human colonic biopsy specimens, Kathryn L.evaney (MGH-GI Unit) for breeding and genotyping the TFF3-nockout mice, and Yvonne Schwafertz (University of Hospitalssen) for technical assistance.
onflicts of interestThe authors disclose the following: D.K.P. is a founder and holds
quity in The GI Company, which has licensed rights for commercialevelopment of TFF3. The remaining authors disclose no conflicts.
undingSupported by CCFA grant SRA-1790, DFG grant Ca226/4-2,
FORES program (to E.C.), and NIH grants DK60049 and DK43351
to D.K.P.).
Podolsky et al.
Supplemental figures and tables
to
Colitis-associated variant of TLR2 causesimpaired mucosal repair due to TFF3 deficiency
Podolsky
DK, Gerken
G, Eyking
A, and Cario E

Podolsky et al.Suppl. Figure
1
A
B
674-
A
674-
B
674-
C
674-
D
674-
E
661-
G
661-
H
661-
I
661-
J
661-
K
H2O
TLR2-/- WT
499bp [WT]334bp [MUT]
500300
WT TFF3-/-
500300 400bp [WT]
183bp [MUT]
687-
A
687-
B
687-
C
687-
D
687-
E
686-
K
686-
N
686-
M
686-
J
686-
Q
H2O
Suppl. Fig. 1Genotyping of TLR2-/-
and TFF3-/-
mice by PCR.Allele-specific genotyping by PCR of mice deficient in TLR2
(A)
or TFF3 (B)
genes. Tail-
extracted genomic DNA was amplified as described in Supplemental Materials and Methods. Representative results (n=5 mice per group) are shown.
Podolsky et al.Suppl. Figure
2
00
10
20
30
40
50
60
70
80
90
100
110
120
10 100
1000
1000
0
[PCSK] ng/ml
EC50
184.0
30 mins
per
cen
t m
axim
al r
esp
on
se(r
ela
tiv
e T
FF
3/G
ap
dh
mR
NA
ex
pre
ss
ion
)
Suppl. Fig. 2Dose response curve of PCSK-mediated TFF3 mRNA increase.Dose-dependent increase of TFF3 mRNA expression in GC-like Ls174T cells by PCSK stimulation (30 mins), as determined by real-time RT-PCR analysis. The amount of PCSK (200ng/ml, 2000ng/ml, 20000ng/ml) is plotted as the log of ligand
concentration. Each data point represents the mean (±
SEM (n=3 independent experiments)) increase of TFF3mRNA
in relation to mRNA expression for the housekeeping gene Gapdh
and normalized to unstimulated
Ls174T cells. The solid line is the sigmoidal
dose-response curve computer-
fitted to the data points (GraphPad
Prism vers
4.03
, GraphPad
Software). EC50
value was calculated using the log dose-response function and non-linear regression analysis of the software-standardized sigmoidal
curve-fitting algorithm.

Podolsky et al.Suppl. Figure
3
A B
+ P
CS
K-
PC
SK
DS
S-W
T; d
ay
12
MUC2
contro
l
- PCSK (D
SS)
+ PCSK (D
SS)
contro
l
- PCSK (D
SS)
+ PCSK (D
SS)
1
2
3
4
5
+
TFF3 MUC2
++
**re
lati
ve
mR
NA
ex
pre
ss
ion
/ G
ap
dh
no
rma
lize
d t
o h
ea
lth
y W
T c
olo
ns
Suppl. Fig. 3The GC-specific product MUC2 is not regulated by TLR2.(A)
Increase of TFF3 mRNA, but not MUC2 mRNA, in distal DSS-colon with or without PCSK treatment during the recovery phase of colitis (day 12), as determined by real-time RT-PCR analysis. Results are shown in relation to mRNA expression for the housekeeping gene Gapdh
and normalized to control H2O-colon (set
as 1 arbitrary
unit). Data are presented as means ±
SEM (n=5-6 mice per group): **, p<0.01; +, p>0.05.
(B)
Representative MUC2 (white) -
immunofluorescence
of distal DSS-colon with or without PCSK treatment on day 12, as assessed by confocal
laser microscopy (40x/1.3, oil, scan zoom 0.7).

Podolsky et al.
0 1 2 3 4 5 6 7 8 9 10 11 120
1
2
3
4
- PCSK (TFF3-/-) + PCSK (TFF3-/-)+ PCSK (TFF3+/+)- PCSK (TFF3+/+)
+ DSS 2.5% -/+ PCSK 150µg/ml
*****+
** +
+ ***++
** *****
+
day
blo
od
sco
re
A B
-P
CS
K+
PC
SK
TF
F3-
/-
10x40x
10x
40x
C
8 120.0
2.5
5.0
7.5
10.0
- PCSK (TFF3+/+) + PCSK (TFF3+/+)
- PCSK (TFF3-/-) + PCSK (TFF3-/-)
+
* +*
day
colo
n l
eng
th (
cm)
Suppl. Figure
4
Suppl. Fig. 4Additional parameters for assessment of the therapeutic effects of TLR2 ligand
therapy in TFF3-deficient mice.TFF3+/+ or TFF3-/-
mice [129S2/SvPaf] received 2.5% DSS for 5 days followed by treatment with PCSK 150µg/ml or water for 7 subsequent days. Mice were sacrificed on days 8 or 12. Assessment of total colon length (A). Evolution of blood scores (B)
during DSS colitis and subsequent treatment with or without PCSK. Data are presented as
means ±
SEM. (+PCSK) vs. (-
PCSK): *p<0.05; **p<0.01; ***p<0.001; +p>0.05. Representative histology (C)
on day 12 of DSS-
TFF3-/-
colon with or without PCSK treatment (hematoxylin-eosin staining; original magnification: x10 or x40 as indicated). White arrows indicate immature epithelial cells over a large wound defect, black arrows indicate improved epithelization
over damaged mucosa.
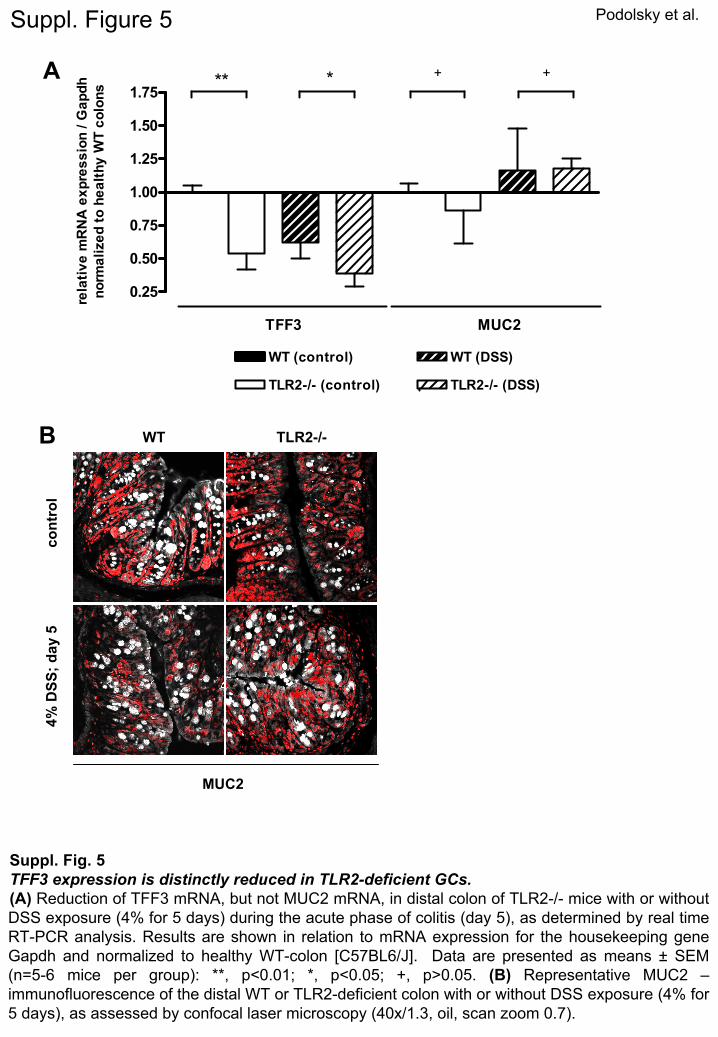
Podolsky et al.
A
B
con
tro
l4
% D
SS
; d
ay5
MUC2
WT TLR2-/-
0.25
0.50
0.75
1.00
1.25
1.50
1.75
WT (control)
TLR2-/- (control)
WT (DSS)
TLR2-/- (DSS)
TFF3 MUC2
** * + +re
lati
ve
mR
NA
ex
pre
ss
ion
/ G
ap
dh
no
rma
lize
d t
o h
ea
lth
y W
T c
olo
ns
Suppl. Figure
5
Suppl. Fig. 5TFF3 expression is distinctly reduced in TLR2-deficient GCs. (A)
Reduction of TFF3 mRNA, but not MUC2 mRNA, in distal colon of TLR2-/-
mice with or without DSS exposure (4% for 5 days) during the acute phase of colitis (day 5), as determined by real time RT-PCR analysis. Results are shown in relation to mRNA expression for the housekeeping gene Gapdh
and normalized to healthy WT-colon [C57BL6/J]. Data are presented as means ±
SEM (n=5-6 mice per group): **, p<0.01; *, p<0.05; +, p>0.05. (B)
Representative MUC2 –
immunofluorescence
of the distal WT or TLR2-deficient colon with or without DSS exposure (4% for 5 days), as assessed by confocal
laser microscopy (40x/1.3, oil, scan zoom 0.7).

Podolsky et al.
8 120
1
2
3
4
5
6
- rTFF3 (TLR2-/-)
+ rTFF3 (TLR2-/-)
*
+
day
his
tolo
gy
sco
re
8 120.0
2.5
5.0
7.5
10.0
- rTFF3 (TLR2-/-)
+ rTFF3 (TLR2-/-)
****
day
colo
n l
eng
th (
cm)
A B
0 1 2 3 4 5 6 7 8 9 10 11 120
1
2
3
4
+ DSS 2.0% -/+ rTFF3 0.4mg/ml
*** ******
****
+
+
- rTFF3 (TLR2-/-) + rTFF3 (TLR2-/-)
day
blo
od
sco
re
C
Suppl. Figure
6
Suppl. Fig. 6Additional parameters for assessment of the effects of recombinant TFF3
supplementation in TLR2-deficient mice.TLR2-/-
mice received 2.0% DSS for 6 days followed by treatment with rTFF3 (0.4mg/ml p.o.) or water for 6 subsequent days. Mice were sacrificed on day 12. Evolution of
rectal bleeding (A)
during DSS colitis and subsequent supplementation of rTFF3. Assessment of (B)
total colon length and (C)
histology score after DSS exposure with or without rTFF3 on day
12.
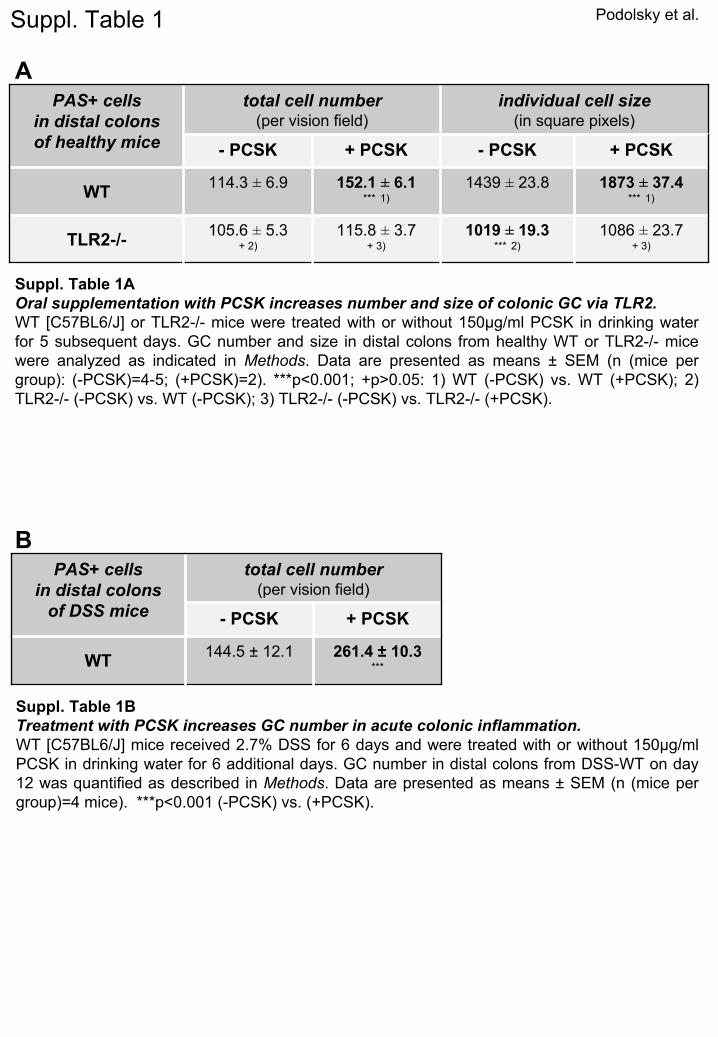
Podolsky et al.
PAS+ cells in distal colons of healthy mice
total cell number (per vision field)
individual cell size (in square pixels)
- PCSK + PCSK - PCSK + PCSK
WT114.3 ±
6.9 152.1 ±
6.1***
1)1439 ±
23.8 1873 ±
37.4***
1)
TLR2-/-105.6 ±
5.3 + 2)
115.8 ±
3.7+ 3)
1019 ±
19.3 ***
2)1086 ±
23.7 + 3)
Suppl. Table 1
A
PAS+ cells in distal colons
of DSS mice
total cell number (per vision field)
- PCSK + PCSK
WT144.5 ±
12.1 261.4 ±
10.3***
B
Suppl. Table 1BTreatment with PCSK increases GC number in acute colonic inflammation.WT [C57BL6/J] mice received 2.7% DSS for 6 days and were treated
with or without 150µg/ml PCSK in drinking water for 6 additional days. GC number in distal colons from DSS-WT on day 12 was quantified as described in Methods. Data are presented as means ±
SEM (n (mice per group)=4 mice). ***p<0.001 (-PCSK) vs. (+PCSK).
Suppl. Table 1AOral supplementation with PCSK increases number and size of colonic GC via TLR2.WT [C57BL6/J] or TLR2-/-
mice were treated with or without 150µg/ml PCSK in drinking water for 5 subsequent days. GC number and size in distal colons from healthy WT or TLR2-/-
mice were analyzed as indicated in Methods. Data are presented as means ±
SEM (n (mice per group): (-PCSK)=4-5; (+PCSK)=2). ***p<0.001; +p>0.05: 1) WT (-PCSK) vs. WT (+PCSK); 2) TLR2-/-
(-PCSK) vs. WT (-PCSK); 3) TLR2-/-
(-PCSK) vs. TLR2-/-
(+PCSK).

Podolsky et al.
gene mean
ratio
#
Gfi1 1.59**
Hes5 1.00+
Klf4 0.93+
Math1 0.95+
Notch1 0.84+
IL-6 1.24+
KGF 0.63*
# mean
ratio
(TLR2-/-
/ WT) of normalized
colonic
expression
levels
Suppl. Table 2
Suppl. Table 2Gene expression changed in healthy TLR2-/-
colons.Results are shown as ratios of mRNA expression levels of healthy
distal colons between TLR2-/-
and WT mice [C57BL6/J], normalized for the housekeeping gene Gapdh.
Values are presented as average fold-change,
as determined by real-time RT-PCR analysis (n=5-6 mice per group): * p<0.05; ** p<0.01; + p>0.05.
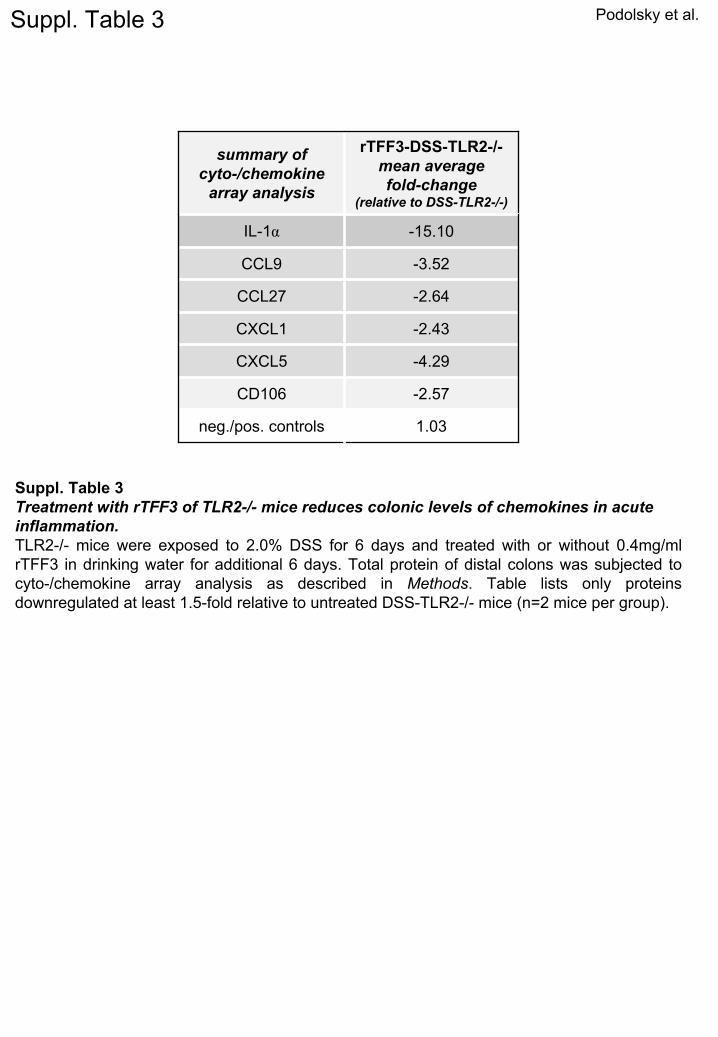
Podolsky et al.
summary ofcyto-/chemokine
array analysis
rTFF3-DSS-TLR2-/-mean
averagefold-change
(relative to DSS-TLR2-/-)
IL-1α -15.10
CCL9 -3.52
CCL27 -2.64
CXCL1 -2.43
CXCL5 -4.29
CD106 -2.57
neg./pos. controls 1.03
Suppl. Table 3
Suppl. Table 3Treatment with rTFF3 of TLR2-/-
mice reduces colonic levels of chemokines
in acute inflammation.TLR2-/-
mice were exposed to 2.0% DSS for 6 days and treated with or without 0.4mg/ml rTFF3 in drinking water for additional 6 days. Total protein of distal colons was subjected to cyto-/chemokine
array analysis as described in Methods. Table lists only proteins downregulated
at least 1.5-fold relative to untreated DSS-TLR2-/-
mice (n=2 mice per group).