collaborators: ulrich brose carol blanchette jennifer dunne sonia kefi neo martinez bruce menge
DESCRIPTION
Collaborators: Ulrich Brose Carol Blanchette Jennifer Dunne Sonia Kefi Neo Martinez Bruce Menge Sergio Navarrete Owen Petchey Philip Stark Rich Williams …. Predictability. Predict. Biodiversity. Biodiversity. Prediction. Changes in Focal Species. - PowerPoint PPT PresentationTRANSCRIPT


Collaborators:
Ulrich BroseCarol BlanchetteJennifer Dunne
Sonia KefiNeo MartinezBruce Menge
Sergio NavarreteOwen Petchey
Philip StarkRich Williams
…

Predictability

Predict
Biodiversity

Prediction
Biodiversity
Changes in Focal Species

….add bit about yosemite toad or mt yellow legged frog


How can we predict the consequences of species loss in complex ecosystems?
Little Rock Lake Food Web (Martinez 1991)

1 degree
How can we predict the consequences of species loss in complex ecosystems?
Little Rock Lake Food Web (Martinez 1991)

2 degrees
How can we predict the consequences of species loss in complex ecosystems?
Little Rock Lake Food Web (Martinez 1991)

3 degrees
Williams et al. PNAS 2002
How can we predict the consequences of species loss in complex ecosystems?
Little Rock Lake Food Web (Martinez 1991)

Complex

Complicated

some hope

metabolism

<1>
everything needs energy to stay alive

<2>
BIG things need more energy than small things

<2>
BIG things need more energy than small things( )3/4
allometric scaling of metabolism with body size

Feeding is Universal

Food Webs are the foundation of Ecological Networks

Body Size should predict the strength of interactions in food webs


Feeding is Universal

Universal ≠ The Only Thing

Ubiquitous≠ The Only Thing

non-metabolic interactions
R. Donovan

Question

can we explain with body size (metabolism)?
ALL interaction strengths

can we explain with body size (metabolism)?
ALL interaction strengths

can we explain with body size (metabolism)?
WHAT

can we explain with body size (metabolism)?
WHAT NOT

.

repeat it

can we explain with body size (metabolism)?
ALL interaction strengths

can we explain with body size (metabolism)?
WHAT

can we explain with body size (metabolism)?
WHAT NOT

abun
danc
e,in
tera
ction
str
engt
h,et
c.
?

abun
danc
e,in
tera
ction
str
engt
h,et
c.
feeding,body size,
metabolism,etc.

abun
danc
e,in
tera
ction
str
engt
h,et
c.
feeding,body size,
metabolism,etc.

can wedescribe a metabolic baseline of interactions
in complex networks?

can wedetrend metabolism
in complex networks?

Brose et al. 2005 EcologyBrose et al. 2006 EcologyPetchey et al. 2008 PNAS
Body Size also influences Food Web Structure

if each link obeys allometric rulesare those rules preserved at the network scale?

if each link obeys allometric ruleswill body size predict
the effect of species loss in the network?

does more complex = more complicated?

Approach

Simulation Results

Real World



Approach:
<1>
Simulate species dynamics in a wide variety of networks
stochastic variation instructural and dynamic
parameters

Approach:
<2>
all feeding links governed by (body size)¾

Approach:
<3>
delete each species and measure effects on all others

Approach:
<4>
Track variation for each simulation
interaction strengthsnetwork level structureneighborhood structure
species attributeslink attributes

Approach:
<5>
mine the variability for what best explains interaction strengths

The Model

The Modelscoupled

<1> Food Web Structure: Niche Model
(Williams and Martinez 2000)
<2> Predator-Prey Interactions: Bio-energetic Model
(Yodzis and Innes 1992, Brose et al. 2005, 2006 Eco Letts)
<3> Plant population dynamics: Plant-Nutrient Model
(Tilman 1982, Huisman and Weissing 1999)

consumersj
jijijjresourcesj
ijiiiii eFyBxFyBxBxB /'
Bioenergetic Predator-Prey Dynamics
Biomassi at time t
Biomass of each species (i) at time (t) is balance of1. gain from consuming prey species 2. loss to being consumed by other species 3. loss to metabolism

mass-specific metabolic rate
max metabolic-specific ingestion rate
Functional Response
assimilationefficiency
Bioenergetic Predator-Prey Dynamics
consumersj
jijijjresourcesj
ijiiiii eFyBxFyBxBxB /'
xi, yi scale with body size(body size correlated with web structure)
# Prey
Cons
umpti
on

Nutrient-DependentGrowth of Plants
Bioenergetic Predator-Prey Dynamics(Plants)
consumersj
jijijjiiiiii eFyBxBxBGrB /'
mass-specificgrowth rate metabolic loss loss to herbivores
ri, xj, y scale with body size

Nutrient-Dependent Growth of Plants
Growth determined by most limiting Nutrient
plantgrowth rate
Concentration of Nutrientsdetermined by
SupplyTurnover
Consumption
Half saturation conc. for uptake of that Nutrient
)(,)(22
2
11
1 tBNK
N
NK
NMINNG i
iii

Generate a food web (Niche Model)
Calculate trophic level for each species
Apply plant-nutrient model to plants, predator-prey model to rest.
Assign body sizes based on trophic level (mean pred: prey ratio = 10)
Run simulation with each species deleted individually to generate a complete removal matrix
Repeat for all species and for 600 Niche Model webs


R
T
Removed Species
Target Species

R
T
X+

R
T

R
T
X-
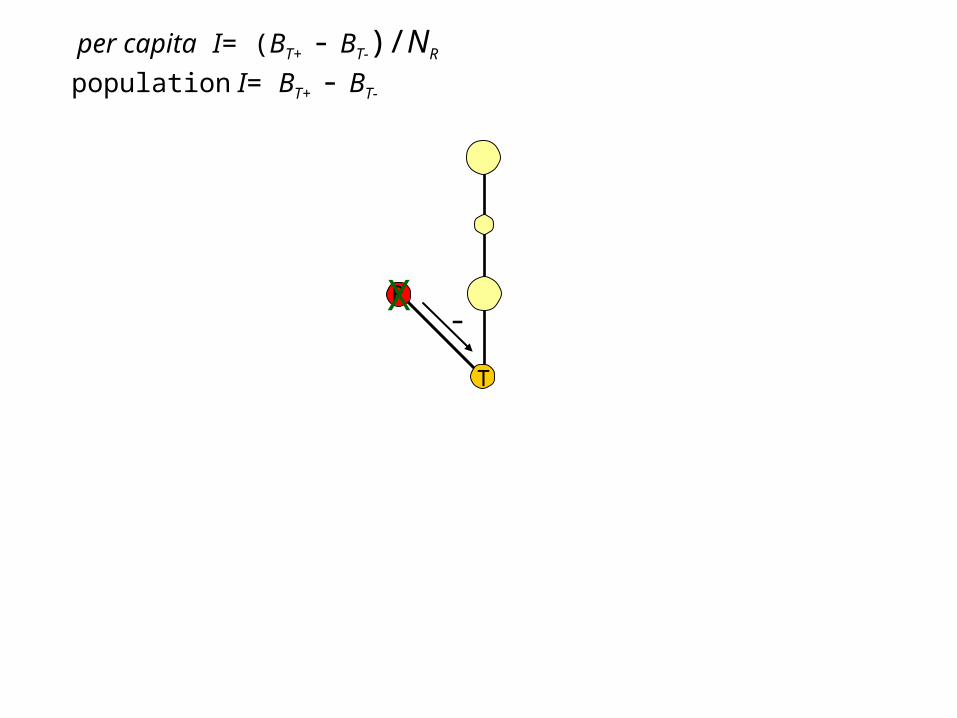
per capita I= (BT+ - BT-)/NR
population I= BT+ - BT-

R
T
X-
per capita I= (BT+ - BT-)/NR
population I= BT+ - BT-

R
T
X-
per capita I= (BT+ - BT-)/NR
population I= BT+ - BT-
R
T
X-
per capita I= (BT+ - BT-)/NR
population I= BT+ - BT-

R
T
X ?
T
per capita I= (BT+ - BT-)/NR
population I= BT+ - BT-

R
T
X 1° Consumers
2° Consumers
3° Consumers
1° Prey
2° Prey
3° Prey
?
T
per capita I= (BT+ - BT-)/NR
population I= BT+ - BT-

R
T
X 1° Consumers
2° Consumers
3° Consumers
1° Prey
2° Prey
3° Prey
?
T
per capita I= (BT+ - BT-)/NR
population I= BT+ - BT-

K
D S1
NK1
K = Keystone consumerNK = Non-Keystone consumer
D = Dominant basal speciesS = Subordinate basal species
R = ResourceR1 R2

S1
S2
K
D S1
NK1
+
Keystone Present
R1 R2
Consumption
Resource competition
Indirect Facilitation

S1
S2
K
D S1
NK1
+
Keystone Present
R1 R2Increased Resources
Consumption
Resource competition
Indirect Facilitation

S1
S2
K
D S1
NK1
Sn
Other Competitors
+
Keystone Present
R1 R2
Consumption
Resource competition
Indirect Facilitation

S1
K
D S1
NK1
Secondary Consumers
+
R1 R2
NK2n
Consumption
Resource competition
Indirect Facilitation

S1
S2
K
D S1
NK1
NK3nTertiary Consumers
+
Secondary Consumers
R1 R2
NK2n
Consumption
Resource competition
Indirect Facilitation

S1
S2
K
D S1
NK1
NK2n
NK3nTertiary Consumers
+
Secondary Consumers
R1 R2
and so on… NK4n

S1
K
D S1
+
S2
Bottom up
Top down
Horizontal

add noise

track the consequences of that noise

add noise:
<1>
Web Structuresize, connectance, architecture

add noise:
<2>
Animal Attributes metabolic and max consumption rate,
pred-prey body size ratiofunctional response type
predator interference

add noise:
<3>
Plant Attributesgrowth rate
half saturation concentrations

track:
90 predictors to explain
variation in the strengths of
254,032 interactions among
12,116 species in
600 webs

track:
<1>Global network structure
<2>Species attributes of R and T
<3>Local network structure around each R and T
<4>Attributes of the interaction

prey predatorpredator prey
+
-
R
T
R T
attributes of the interaction+-

shortest path = 2 degrees
2 degree paths: +, +, -prey predator
predator prey
+
-
R
T
R T+-
attributes of the interaction

shortest path = 2 degrees
2 degree paths: +, +, -3 degree paths: +, +, +, -prey predator
predator prey
+
-
R
T
R T+-
attributes of the interaction

shortest path = 2 degrees
2 degree paths: +, +, -3 degree paths: +, +, +, -4 degree paths: -
prey predatorpredator prey
+
-
R
T
R T+-
sign shortest path = +1sign next shortest path = +2un-weighted sum (shortest + next shortest) = +3weighted sum (shortest + (next shortest / 2)) = +2
attributes of the interaction



-2
0
2
4
6
8
Log
(Bod
y M
ass)
1 2 3 4 5 6 7 8
Trophic Level
Body Size and Food Web Structure

R2 = 0.90Slope = 0.74
Lo
g (
per
ca
pit
a co
nsu
mp
tio
n)
-12
-9
-6
-3
0
3
6
9
PC
Lin
IS)
-4 -2 0 2 4 6 8 10log (SR mass)
Log (R body mass)
Each Feeding Interaction Scales with (Body Size)3/4
R
T

R2 = 0.90Slope = 0.74
Lo
g (
per
ca
pit
a co
nsu
mp
tio
n)
Per Capita Linear Interaction Strength
-12
-9
-6
-3
0
3
6
9
PC
Lin
IS)
-4 -2 0 2 4 6 8 10log (SR mass)
Log (R body mass)
R
T
= Per Capita Removal Interaction Strength?

-12
-9
-6
-3
0
3
6
9
log
(|D
iff P
C|)
-4 -2 0 2 4 6 8 10log (SR mass)
Lo
g |
per
ca
pit
a I|
R2 = 0.32Slope = 0.74
Log (R body mass)
Per Capita Removal Interaction Strength
R
T

-12
-9
-6
-3
0
3
6
9
log
(|D
iff P
C|)
-4 -2 0 2 4 6 8 10log (SR mass)
Lo
g |
per
ca
pit
a I|
R2 = 0.14Slope = 1.3
Log (R body mass)
Per Capita Removal Interaction Strength
R
T

-12
-9
-6
-3
0
3
6
9
log
(|D
iff P
C|)
-4 -2 0 2 4 6 8 10log (SR mass)
Log (R body mass)
Lo
g |
per
ca
pit
a I|
R2 = 0.45Slope = 1.4
Per Capita Removal Interaction Strength
R
T

-6
-3
0
3
6
Res
idua
ls
log
(abs
PC
)
-15 -12 -9 -6 -3 0log (TS biom)
-12
-9
-6
-3
0
3
6
9
log
(|D
iff P
C|)
-4 -2 0 2 4 6 8 10log (SR mass)
Log (R body mass)
Lo
g |
per
ca
pit
a I|
Per Capita Interaction Strength
Low R BiomassHigh R Biomass
Res
idu
als
Log (T biomass)
Per Capita Removal Interaction Strength
R
T

Predicted by:Log (T biomass) +Log (R biomass) +Log (R body mass)
Lo
g |
per
ca
pit
a I|
Per Capita Interaction Strength
-12
-9
-6
-3
0
3
6
9
log
(|D
iff P
C|)
-10 -8 -6 -4 -2 0 2 4 6 8Pred Formula log (|Diff PC|)
R2 = 0.88
R
T

population I
(population interaction strength)

population I
(total effect on T of removing R)

Classification and Regression Trees (CART)on log transformed |Interaction Strengths|
best predictors of absolute magnitude of log(population I)
T biomassR biomass
(Degrees Separated)
of the 90 variables tracked
R2 = 0.65

-12
-9
-6
-3
0
log
(|D
iff T
E|)
-12 -9 -6 -3 0log (TS biom)
Lo
g |
po
pu
lati
on
I|
Log (T biomass)

-12
-9
-6
-3
0
log
(|D
iff T
E|)
-12 -9 -6 -3 0log (TS biom)
Low R BiomassHigh R Biomass
Log (T biomass)
Lo
g |
po
pu
lati
on
I|
R2 = 0.65

Sign (strong interactions)
0.00
0.25
0.50
0.75
1.00
-1 0 1
-1
1
≤ -1 ≥ 1
Weighted Sum Path Signs
Pro
po
rtio
n O
bse
rved
0.00
0.25
0.50
0.75
1.00
-1 0 1
-1
1
Sign (weak interactions)
≤ -1 ≥ 1
Weighted Sum Path Signs
positive
negative

-18
-15
-12
-9
-6
-3
0
log
(abs
TE
)
-15 -12 -9 -6 -3 0log (TS biom)
-6
-3
0
3
6
Res
idua
ls
log
(abs
PC
)-15 -12 -9 -6 -3 0
log (TS biom)
-15
-12
-9
-6
-3
0
log
(abs
TE
)
-4 -2 0 2 4 6 8 10log (SR mass)
-12
-8
-4
0
4
8
12
log
(abs
PC
)
-4 -2 0 2 4 6 8 10log (SR mass)
Lo
g (
|per
cap
ita
I|)L
og
(|p
op
ula
tio
n I|
)
Res
idu
als
fro
m (
a)
Log (T biomass)Log (R Body Mass)
Lo
g (
|po
pu
lati
on
I|)
(a)
2. strongest per capita I:large bodied, low biomass R
effects on high biomass TR2 = 0.88
3. strongest population I:high biomass R
effects on high biomass TR2 = 0.65
Summary:
1. 3/4 scaling disappears in complex networks
Low R BiomassHigh R Biomass
-2
0
2
4
6
-4 -2 0 2 4 6 8 10log (SR mass)
Lo
g (
per
cap
ita
linea
r I)

Strong Strong per capita per capita effectseffects

Strong population effects

How can it be so simple?

Is it circular?
-15
-12
-9
-6
-3
0
log
(|D
iff T
E|)
-15 -12 -9 -6 -3 0
log (TS biom)Log (T biomass)
Lo
g (
|po
pu
lati
on
I|)
predicting: (BT+ - BT-)using: BT+
Log (BT+)
BT+ = T biomass (R present)BT- = T biomass (R removed)

Is it circular?
-15
-12
-9
-6
-3
0
TE
shu
ff|)
-15 -12 -9 -6 -3 0log (TS biom)
-15
-12
-9
-6
-3
0
log
(|D
iff T
E|)
-15 -12 -9 -6 -3 0
log (TS biom)Log (T biomass)
Lo
g (
|po
pu
lati
on
I|)
predicting: (BT+ - BT-) 2° extinction of T
Log (T biomass)
reshuffled interactions
using: BT+
Log (BT+) Log (BT+)
R2 = 0.59 R2 = 0.19

-6
-4
-2
0
2
Diff
TE
1 2 3 4 5 6 7degrees_separated
po
pu
lati
on
I
Degrees Separated
Chains of interactions tend to dampen with distance

Species Richness
10 15 20 25 30
R2
0.75
0.80
0.85
0.90
0.95
Species Richness
10 15 20 25 30
R2
0.45
0.50
0.55
0.60
0.65
0.70
Prop
ortio
n of
Var
iatio
n Ex
plai
ned
R2 = 0.88
Number of Species
R2 = 0.73
More Complex is More Simple
per capita I population I



What about the real world?

Predictions:
<1> Purely metabolic interactions
should be well predicted by simple attributes of R and T.

Predictions:
<2>Deviations from simple metabolic predictions
should point to strong non-metabolic influences.

Goal:
De-trend the "metabolic baseline" of complex systems to gain insight into other important ecological processes.

SuccessfullyPredict

FailPredictably

Berlow 1999 Nature 398:330

R
T
Whelks
MusselsBarnacles
Space

R
T
Whelks
MusselsBarnacles
Space
Field Experiment Conditions

R
T
Whelks
MusselsBarnacles
Space

R
T
Whelks
MusselsBarnacles
Space
+-
+- -
- -
Metabolic

R
T
Whelks
MusselsBarnacles
Space
+-
+- -
- -
Metabolic+
Non-Metabolic

R
T
WhelksMussels
Barnacles
+-
Metabolic+
Non-Metabolic
R
T
R
T+-
+- -
T+-
+- -
R
T
R
T
-
T
-Metabolic
Experimental Design
WhelksExcluded
Low WhelkBiomass
High WhelkBiomass
Natural Variationin Mussels
and Barnacles
4 blocks x 3 start datesx 1-3 yrs

-18
-15
-12
-9
-6
-3
0
log
(abs
TE
)
-15 -12 -9 -6 -3 0log (TS biom)
-6
-3
0
3
6R
esid
uals
log
(abs
PC
)
-15 -12 -9 -6 -3 0log (TS biom)
Lo
g (
|per
ca
pit
a I|
)
Log (T biomass)
Lo
g (
|po
pu
lati
on
I|)
Simulation ResultsLow R BiomassHigh R Biomass

Log (Mussel Biomass)
-1 0 1 2 3
Lo
g (|p
op
ulatio
n
I|)
-3
-2
-1
0
1
-1 0 1 2 3
Lo
g (
| per
cap
ita
I|)
-4
-3
-2
-1
0
Log (Mussel biomass)
Low Whelk BiomassHigh Whelk Biomass
Central TendencyPredicted by Simulations
predicted
Lo
g (
|per
ca
pit
a I|
)L
og
(|p
op
ula
tio
n I
|)

-1 0 1 2 3
Lo
g (
| per
cap
ita
I|)
-4
-3
-2
-1
0
Log (Mussel Biomass)
-1 0 1 2 3
Lo
g (
|po
pu
lati
on
I|)
-3
-2
-1
0
1
R
T
-Metabolic predicted
Log (Mussel biomass)
Lo
g (
|per
ca
pit
a I|
)L
og
(|p
op
ula
tio
n I
|)
Low Whelk BiomassHigh Whelk Biomass

-1 0 1 2 3
Lo
g (
| per
cap
ita
I|)
-4
-3
-2
-1
0
-1 0 1 2 3
Log (Mussel Biomass)
-1 0 1 2 3
Lo
g (
|po
pu
lati
on
I|)
-3
-2
-1
0
1
Log (Mussel Biomass)
-1 0 1 2 3
R
T
R
T+
+- --
R2 = 0.49
R2 = 0.43
Metabolic predicted
observed
Log (Mussel biomass)
Lo
g (
|per
ca
pit
a I|
)L
og
(|p
op
ula
tio
n I
|)
Low Whelk BiomassHigh Whelk Biomass
Low Whelk BiomassHigh Whelk Biomass

-1 0 1 2 3
Lo
g (
| per
cap
ita
I|)
-4
-3
-2
-1
0
-1 0 1 2 3
Log (Mussel Biomass)
-1 0 1 2 3
Lo
g (
|po
pu
lati
on
I|)
-3
-2
-1
0
1
Log (Mussel Biomass)
-1 0 1 2 3
R
T
R
T+
+- --
R2 = 0.49
R2 = 0.43
Metabolic Metabolic+
Non-Metabolic
Log (Mussel biomass) Log (Mussel biomass)
predicted
observed
Lo
g (
|per
ca
pit
a I|
)L
og
(|p
op
ula
tio
n I
|)

Summary<1>
¾ power law signal disappears andnew simple patterns emerge in a network context.
<2>magnitude of per capita and population I
explained by 2-3 simple species attributes (of 90)
<3>effects dampen with distancemore complex = more simple
<4>predictable fit and lack-of-fit in field experiment

Conclusions
<1>metabolic "webbiness" of life not necessarily a big source of uncertainty.

Conclusions
<2>“module” approaches may work best
when embedded in complexity

Conclusions
<3>metabolic "null model" may describe
a universal baseline of species interactions in a complex network.

Conclusions
<4>"de-trend" metabolism in ecological networks
to better understand non-metabolic interactions and processes

"I would not give a fig for simplicity on this side of complexity, but I'd give my life for the simplicity on the other side of complexity"
Oliver Wendell Holmes, Jr.

Acknowledgements
Alexander von Humboldt Foundation




-14
-12
-10
-8
-6
-4
-2
0
2
4L
og
(P
op
ula
tio
n D
ensi
ty)
-6 -4 -2 0 2 4 6 8 10 12
Log (Body Mass)
R2 = 0.96 slope = -1.05High Biomass
R2 = 0.36 slope = -1.17Low Biomass
R2 = 0.59slope = -1.4All Points
0.05
0.10
0.15
Pro
babi
lity
-16 -14 -12 -10 -8 -6 -4 -2 0 1
0.05
0.10
0.15
-16 -14 -12 -10 -8 -6 -4 -2 0 1
PositiveEffects
NegativeEffects
Pro
bab
ility
0.10
0.05
0.15
0.10
0.05
0.15
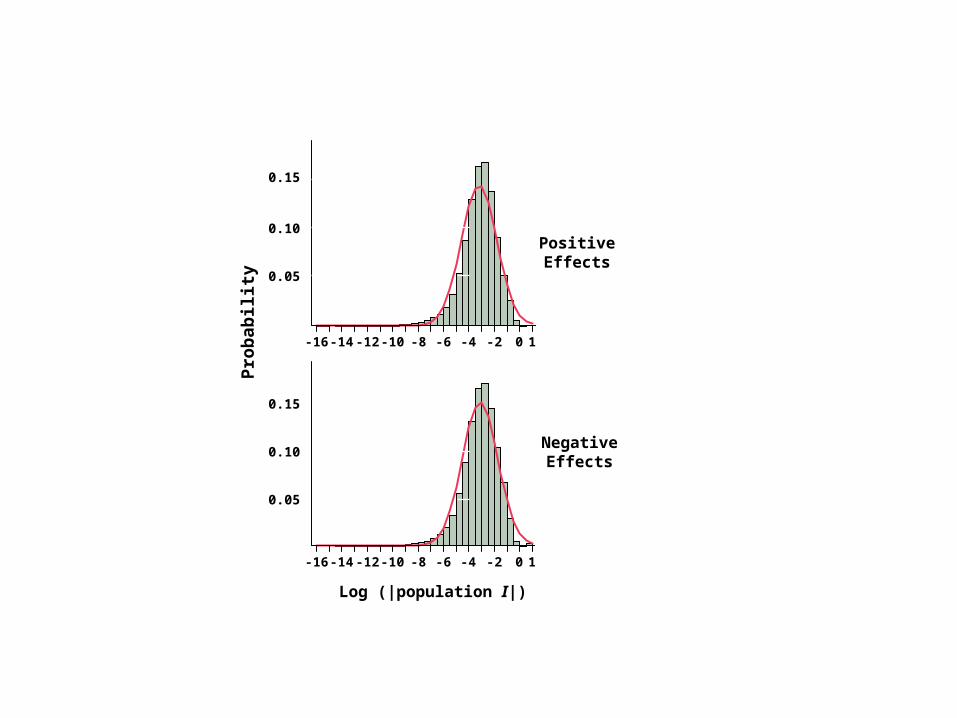
Log (|population I|)

n = 5 random subsamplesof 10,000 interactions
trophic
leve
l
1dgr p
rey
biom
log (b
ody m
ass)
#1 d
gr pre
d
% o
f E
xpla
ined
Var
iati
on
0
10
20
30
40
50
R presentR removed
(a)+
+
-
-

Degrees Separated1 2 3 4 5 6 7
Ma
x L
og
(|p
er c
apit
a I|
)
2
4
6
8
10
Ma
x L
og
(|p
op
ula
tio
n I
|)
-1.6
-1.2
-0.8
-0.4
Chains of interactions tend to dampen with distance