comparative effect of viral and bacterial neuraminidase onthe
TRANSCRIPT

Clin. exp. Immunol. (1972) 10, 139-150.
COMPARATIVE EFFECT OF VIRAL AND BACTERIALNEURAMINIDASE ON THE COMPLEMENT
SENSITIVITY OF LYMPHOID CELLS
P. K. RAY AND R. L. SIMMONS
Department of Surgery, University of Minnesota,Minneapolis, Minnesota, U.S.A.
(Received 7 July 1971)
SUMMARY
C3H/HeJ mouse lymphoid cells were exposed to Vibrio cholerae neuraminidase(VCN), Clostridium perfringens neuraminidase (CPN), and influenza virus neura-minidase (IVN). Exposure of mouse lymphoid cells to VCN or CPN in non-cyto-toxic concentrations renders such cells susceptible to lysis by specific antibody andrabbit serum complement and by rabbit serum alone. Inactivation of complementeliminates the effect. Exposure of rabbit lymphoid cells to VCN or CPN rendersthem susceptible to lysis in the absence of antibody when autologous complement isactivated within the fluid phase by cobra venom factor. IVN does not sensitizelymphoid cells to complement in these systems.VCN and CPN are known to hydrolyse the 2-3', 2-6' and 2-8' glycosidic linkages
between sialic acid and mucopolysaccharides on the cell membrane. IVN hydro-lysed only the 2-3' and 2-8' glycosidic linkages. Thus, destruction of the 2-6'glycosidic linkage renders such cells highly susceptible to lysis by complement.Quantitation of sialic acid released from cells treated with VCN or IVN demon-strates that most of th esialic acid is bound to the cell surface by the 2-6' glycosidiclinkage.
INTRODUCTION
Previous reports have demonstrated that Vibrio cholerae neuraminidase (VCN) renderslymphoid cells highly susceptible to the cytolytic action of specific antiserum and comple-ment (C) (Ray, Gewurz & Simmons, 1970; Schlesinger & Amos, 1971). The increasedsensitivity can be only partially explained by unmasking of antibody-binding sites on thecell surface (Schlesinger & Amos, 1971; Kassulke, Stutman & Yunis, 1971) and is probablydue to an increased sensitivity of VCN-treated cells to C-mediated lysis (Ray et al., 1970;Ray et al., 1971). Bacterial neuraminidases lyse the 2-3', 2-6', and 2-8' glycosidic linkages
Correspondence: Dr R. L. Simmons, Box 185-Mayo, University of Minnesota, Minneapolis, Minnesota55455, U.S.A.
139

140 P. K. Ray and R. L. Simmons
between sialic acid and the cell membrane whereas viral neuraminidases lyse only the2-3' and 2-8' glycosidic linkages (Drzeniek, 1967; Drzeniek & Gaube, 1970). The presentexperiment was designed to compare the relative role of the various types of glycosidiclinkages between sialic acid and the cell membrane on the C sensitivity of lymphoid cells.
MATERIALS AND METHODSAnimalsAdult C3H/HeJ male mice obtained from the Jackson Laboratory, Bar Harbor, Maine,
were used for lymphoid cells. C57 BL/6J male mice were used as the source of anti-C3Hantiserum. New Zealand white rabbits were used as the source of complement (C) andlymphoid cells.
AntiserumAntisera to C3H/HeJ alloantigens were prepared in C57 BL/6J mice by grafting C57
BL/6J animals twice sequentially with C3H/HeJ skin. In addition 25 x 106 C3H mousespleen cells were injected i.p. into the C57 recipient weekly for 8-12 weeks (Kinne &Simmons, 1967). Bleeding was carried out 4 days after the last spleen cell injection. Theblood was allowed to clot and centrifuged at 2500 g for 20 min at 0C. The serum obtainedwas stored at - 20'C until used.
ComplementRabbit serum was usually used as the source of C. Blood was obtained from normal
adult rabbits by ear vein puncture, allowed to clot at room temperature (250C) for 30 minand then at 00C for 1 hr. The serum was obtained by centrifugation at 2500 g for 20 minat 0°C. The rabbit serum was repeatedly absorbed at 0°C with mouse liver powder (BBL,Division of Bioquist, Cockeysville, Maryland 21030, U.S.A.) (one part mouse liver powder:four parts serum) to absorb out lytic factors directed against mouse cells. The serum wasstored at - 200C till used. When necessary the serum C was inactivated by heat (56°C,30 min).Cobra venom factor was prepared (Shin, Gewurz & Snyderman, 1969) and used as an
activator of the late-acting C-cascade (G6tze & Muller-Eberhard, 1970; Shin et al., 1969).
Cell suspensionMouse or rabbit lymph nodes were excised after killing by cervical compression. The
nodes were teased apart in medium 199 (M 199) (Grand Island, New York) containing 0.10%bovine serum albumin (BSA) (Sigma Company). The cell suspensions were centrifugedat 250 g for 5 min at 40C and washed three times in M199 plus BSA. After the last wash,the cells were suspended in M199.
Labelling of cells with 51CrThe suspension of cells in M199 were mixed with 0-22 mCi of Na25'CrO4 solution
(Amersham/Searle) and incubated at 37°C for 45 min. After incubation the excess Na25 'CrO4solution was removed by repeated washing with large volumes of M199 and the cells werefinally suspended in 1 ml of M199 plus BSA. After labelling, the viability of the cells was

Viral and bacterial neuraminidase 141measured by their ability to exclude trypan blue. Only suspensions with greater than 9000viability were used. The cell suspension was diluted with M199 plus BSA to have a con-centration of 10 x 106 cells/ml.
Incubation of cells with neuraminidaseVibrio cholerae neuraminidase (VCN) (General Biochemicals, Chagrin Falls, Ohio)
contains 500 units/ml. A unit of activity is equivalent to 1 Pg of N-acetyl neuraminic acidreleased from a glycoprotein substrate in 15 min at 370C. Lee strain influenza virus neura-minidase (IVN) (General Biochemicals, Chagrin Falls, Ohio) contains 100 units/ml. Oneunit IVN causes release of 1 pmole of sialic acid per minute at 370C from bovine sub-maxillary mucin. Clostridium perfringens neuraminidase (CPN) (Sigma type VI) contains l1units/mg. One unit of CPN liberates 1 ymole of N-acetyl neuraminic acid per minute atpH 5*0 at 370C from N-acetyl neuramino-lactose. CPN is known to be impure and containstoxic contaminants (Kraemer, 1968). The enzyme solutions were diluted in M199 to thedesired concentrations prior to use. One ml of the diluted enzyme solution was mixed with1 ml (10 x 106 cells) cell suspension and incubated at 370C for 1 hour. All the tubes weremixed well every 15 min. After incubation, the excess enzyme was removed by repeatedwashing with M199 plus BSA. When appropriate the enzymes were inactivated by heatingto 100°C for 10 min (Codington, Sanford & Jeanloz, 1970).
Cytotoxicity assayThe method followed was that of Wigzell (1965). Serial dilutions of C57 anti-C3H serum
or rabbit serum (0 05 ml) were made with M199 plus BSA. To 0 05 ml of antiserum, 0 05 ml(5 x 105 cells) of enzyme treated cells were added and incubated at 37°C for 15 min.Controls with untreated cells and cells exposed to heat-inactivated enzyme were always run.After incubation, diluted rabbit serum C (0 05 ml) was added and again incubated for atotal of 1 hr at 37°C. At the end of incubation all the tubes were diluted with Ca"+ andMg' + deficient phosphate-buffered saline, pH 7-2. All the tubes were centrifuged at 800 gfor 5 min and 1 ml supernatant portions were taken out for counting in a gamma-radiationcounter (Gamma/guard, Tracer Lab).The spontaneous release of radioactivity was determined by counting 1 ml of supernatant
from one of the tubes containing only cells plus M199. The total cellular radioactivity wasdetermined by counting 1 ml of thoroughly mixed cell suspension from other tubes con-taining only cells in M199. Values for spontaneous release were subtracted from all othervalues. The results are expressed as a percentage of the total cellular radioactivity.
Several experiments were carried out in which CVF was used to activate the terminalcomponents of the C-cascade (C3-C9) within the fluid phase (Shin et al., 1969; Gotze &Muller-Eberhard, 1970). In these experiments it was possible to determine the complementsensitivity of cells in the absence of antibody. The three components of the incubationmixture, labelled rabbit cells (0 05 ml containing 5 x 105 cells), autologous rabbit serum(0 05 ml) and CVF (0-29 pg in 0 05 ml) were added to one another in varying sequences.Two of the components were mixed, incubated at 37°C for 30 min prior to the addition ofthe third component. After a second 30-min period of incubation at 37°C, the reaction wasstopped by the addition of 1 5 ml of calcium and magnesium deficient phosphate bufferedsaline (pH 7 2). After centrifugation the radioactivity in 1 ml supernatant was measured.

142 P. K. Ray and R. L. Simmons
Controls containing only cells plus M199, cells plus serum, or cells plus CVF were also runsimultaneously.
Measurement of sialic acidSialic acid released by the action of neuraminidase was assayed by the method of Aminoff
(1961) with a little modification. The reagents used are (a) Periodate: 25 mm periodic acid in0 125 N-H2SO4 (pH 1 2); (b) Sodium arsenite: 2% solution of sodium arsenite in 0 5N-HCl. (c) Thiobarbituric acid; 01 M solution of 2-thiobarbituric acid (Sigma) in water,adjusted to pH 9 0 with NaOH. (d) Acid butanol: Butan-1-ol containing 500 (V/V) of 12N-HCl.Lymphoid cell suspensions from rabbits were made in Earle's balanced salt solution of
pH 7 0 without any phenol red or other indicator. 1 ml cell suspension containing 200 x 106cells were incubated with 1 ml enzyme solution at 370C for 1 hr. After incubation, the cellswere centrifuged (800 g, 5 min). To 2 ml of supernatant, 1 ml of periodate reagent was addedand incubated for 30 min at 37°C. The excess of periodate is then reduced with 0-8 ml ofsodium arsenite. After the colour of iodine has disappeared (30 sec-1 min) 8 ml of thio-barbituric acid reagent is added, mixed well in a covered test tube and heated at 100°C for10 min. The coloured solutions are then cooled in ice water and shaken with 5 ml of acidbutanol. The separation of the two phases is facilitated by centrifugation at 800 g for 5 minand the colours in the butanol layer are compared at 549 mju in a Beckman spectrophoto-meter, model DU. N-acetyl neuraminic acid (Sigma type III) was used as standard. Reagentcontrol using untreated cell supernate was made and adjusted to zero.
RESULTS
EFFECT OF BACTERIAL AND VIRAL NEURAMINIDASES ON THE LYSIS OF MOUSE
LYMPHOID CELLS
Radiochromated C3H/HeJ lymphoid cells were exposed to varying amounts of VCN,CPN, or IVN at 37°C for 1 hr. Cell lysis resulted in 51Cr release into the supernatantsolution. The results are shown in Table 1. CPN is toxic to mouse lymphoid cells at con-centrations greater than 25 ,g/ml/5 x 106 cells but neither IVN nor VCN destroy lymphoidcells at any concentration, up to 100 u/ml/5 x 106 cells.
COMPARATIVE ACTIVITY OF BACTERIAL AND VIRAL NEURAMINIDASES TO SENSITIZE
MOUSE LYMPHOID CELLS TO THE CYTOTOXIC ACTION OF ANTI-MOUSE SERUM AND
RABBIT SERUM COMPLEMENT
C3H/HeJ mouse lymphoid cells were incubated with VCN (25 u/ml/5 x 106 cells), CPN(25 ,g/ml/5 x 106 cells), and IVN (25 u/ml/5 x 106 cells) at 37°C for 1 hr. These cells werethen exposed to serial dilutions of C57 anti-C3H serum and rabbit serum C. The results areshown in Fig. 1. VCN and CPN increased the susceptibility of lymphoid cells to the cytolyticaction of specific antibodies and C. VCN appears to be most active while IVN was almostcompletely inactive in this regard. Heat inactivation of the C in rabbit serum prevented allcytotoxic effect. Cells incubated with heat inactivated VCN or CPN behaved as normaluntreated cells.
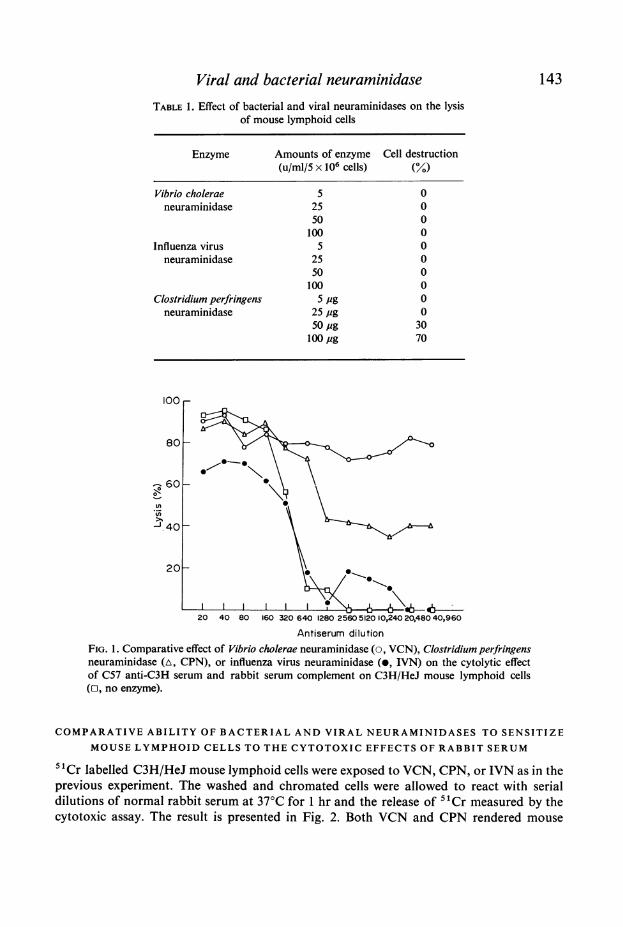
Viral and bacterial neuraminidaseTABLE 1. Effect of bacterial and viral neuraminidases on the lysis
of mouse lymphoid cells
Enzyme
143
Amounts of enzyme Cell destruction(u/ml/5 x 106 cells) (%)
Vibrio cholerae 5 0neuraminidase 25 0
50 0100 0
Influenza virus 5 0neuraminidase 25 0
50 0100 0
Clostridium perfringens 5 pg 0neuraminidase 25 pg 0
50pg 30100 jug 70
.0J40
20
20 40 80 160 320 640 1280 2560 5120 10,240 20,480 40,960Antiserum dilution
FIG. 1. Comparative effect of Vibrio cholerae neuraminidase (o, VCN), Clostridium perfringensneuraminidase (A, CPN), or influenza virus neuraminidase (0, IVN) on the cytolytic effectof C57 anti-C3H serum and rabbit serum complement on C3H/HeJ mouse lymphoid cells(r, no enzyme).
COMPARATIVE ABILITY OF BACTERIAL AND VIRAL NEURAMINIDASES TO SENSITIZEMOUSE LYMPHOID CELLS TO THE CYTOTOXIC EFFECTS OF RABBIT SERUM
5tCr labelled C3H/HeJ mouse lymphoid cells were exposed to VCN, CPN, or IVN as in theprevious experiment. The washed and chromated cells were allowed to react with serialdilutions of normal rabbit serum at 370C for 1 hr and the release of 5tCr measured by thecytotoxic assay. The result is presented in Fig. 2. Both VCN and CPN rendered mouse

144 P. K. Ray and R. L. Simmons
100_
80-
-60
4040
20
2 4 8 16 32Rabbit serum dilution
FIG. 2. Comparative effect of Vibrio cholerae neuraminidase (o, VCN), Clostridium perfringensneuraminidase (A, CPN), or influenza virus neuraminidase (0, IVN) on the cytolytic effect ofrabbit serum on C3H normal lymphoid cells (o, no enzyme).
lymphoid cells increasingly susceptible to the cytolytic action of normal rabbit serum;IVN had no effect. Heat inactivation of the rabbit serum prevented all cytotoxic effects.Cells incubated with heat inactivated VCN or CPN behaved as untreated cells.
COMPARATIVE ABILITY OF BACTERIAL AND VIRAL NEURAMINIDASE TOSENSITIZE RABBIT LYMPHOID CELLS TO THE FLUID PHASE ACTIVATION OF
AUTOLOGOUS RABBIT COMPLEMENT
Suspensions of rabbit lymphoid cells were exposed to VCN, CPN, or IVN. The washedlabelled rabbit cells were then mixed in various sequences with autologous rabbit serum andCVF. Cell lysis was then determined. The results of three separate experiments are shown inTable 2. Cells which had been exposed to VCN or CPN were lysed if CVF was added to amixture of rabbit cells and autologous serum. IVN treated cells were not lysed. All lyticeffects were prevented if (a) the enzyme had been inactivated by heat, (b) the CVF was addedto the autologous serum before the addition of cells, or (c) the autologous serum had beenheated to 560C for 30 min. Therefore activation of autologous serum C by CVF within thefluid phase permits the lysis only of CPN or VCN treated cells. In some experiments lysis(always less than 80%) could be seen by incubating VCN treated cells with the autologousserum without adding CVF, but this too was prevented by prior heating of the serum.
COMPARISON OF THE AMOUNT OF SIALIC ACID RELEASED FROM LYMPHOID
CELLS BY BACTERIAL OR VIRAL NEURAMINIDASE
Rabbit lymphoid cells were incubated with varying amounts of VCN, or IVN for 1 hr at37°C. The supernatant was assayed for sialic acid content by the thiobarbituric acid methodof Aminoff (1961). The results are presented in Fig. 3. There is a marked difference in the

Viral and bacterial neuraminidase 145TABLE 2. Comparative ability of bacterial and viral neuraminidases to render rabbit lymphoid cells sensitive
to the autologous serum complement activated by cobra venom factor
Enzyme treatment Amount of enzyme/ Complement activation Percent lysis ("Cr-release)of cells 5 x 106 cells/ml
Exp. 1* Exp. 2t Exp. 3t
(A) Cells incubated with rabbit serum followed by addition of CVF.None CVF 0 0 0Inactivated VCN 25 u CVF 0 0 0VCN 25 u CVF 34 42 54CPN 25 ug CVF 12 20 36IVN 25u CVF 0 0 0VCN 25u 5 6'5 8
(B) Cells incubated with heat inactivated rabbit serum followed by addition of CVF.None CVF 0 0 0Inactivated VCN 25 u CVF 0 0 0VCN 25u CVF 0 0 0CPN 25pg CVF 0 0 0IVN 25 u CVF 0 0 0VCN 25 u 0 0 0
(C) CVF incubated with rabbit serum followed by addition of cells.None CVF 0 0 0Inactived VCN 25 u CVF 0 0 0VCN 25u CVF 0 0 0CPN 254,g CVF 0 0 0IVN 25 u CVF 0 0 0VCN 25u 5-5 65 8
* 5 x 10' cells in 0 05 ml were incubated with 0 05 ml of autologous serum.t I x 105 cells in 0 05 ml were incubated with 0-05 ml of autologous serum.
0 ~~~~~~~~~~~~~~-O30
*0
0
E 20
I-0
, 5 25 50 100
Enzyme units /200x o06 cells
FIG. 3. Sialic acid liberated by Vibrio cholerae neuraminidase (o, VCN-treated) or influenzavirus neuraminidase (0, IVN-treated) from rabbit lymphoid cells at varying concentrations ofenzyme.
K

146 P. K. Ray and R. L. Simmons
amount of sialic acid liberated by bacterial and viral neuraminidases. VCN releases moresialic acid at the relatively low concentrations used in the previous experiment (25 u/5 x 106cells) than are released even by very high concentrations of IVN. CPN at concentrations of25 ug/200 x 106 cells releases amounts of sialic acid comparable to that released by VCN(30 nanomoles/109 cells). Since CPN at doses higher than 25 jg is toxic towards cells, thehigher concentrations were not used in this experiment.
DISCUSSION
Sialic acid residues are major constituents on the surface of plasma membranes (Benedetti &Emmelot, 1967) and make the major contribution to the negative charge of the cell surface(Ambrose, 1966). A number of functional alterations of cells may be induced by removal ofsialic acid residues from the surface membrane by VCN (Sanford, 1967; Currie, Doorninck& Bagshawe, 1968; Currie & Bagshawe, 1968; Currie & Bagshawe, 1967). These includeenhanced phagocytosis (Weiss, Mayhew & Ulrich, 1966; Lee, 1968), inhibition of viral(Baylor, 1964) or mycoplasma (Gesner & Thomas, 1966) induced haemagglutination,inhibition of cell aggregation (Kemp, 1968), increased cell deformability (Weiss, 1965), alter-ations in patterns of cellular migration (Woodruff & Gesner, 1969), and interference withamino acid transport across treated cell membranes (Brown & Michael, 1969). In addition, anumber of investigators have demonstrated that removal of sialic acid from cell mem-branes increases the immunogenicity of malignant (Sanford, 1967; Currie & Bagshawe,1968; Bagshawe & Currie, 1967; Lindenmann & Klein, 1967, Simmons et al., 1971c) andnormal cells (Simmons, Rios & Ray, 1970; Simmons, Rios & Ray, 1971a; Simmons et al.,1971b; Lundgren et al., 1971).Previous studies have shown that lymphoid cells are rendered extremely susceptible to
the cytolytic action of alloantibody and C by VCN (Ray et al., 1970; 1971). Absorptionexperiments, however, failed to demonstrate any increase in alloantibody-binding capacityby VCN treated cells (Ray & Simmons, 1971) even though certain other antigens could be'unmasked' by VCN (Schlesinger & Amos, 1971; Kassulke et al., 1971). The increasedcytolytic effect of antibody and C appeared to be primarily due to an increased susceptibilityof such cells to C (Ray et al., 1971). Activation of the terminal C-cascade in autologousserum by CVF (Shin et al., 1969; Gttze & Mifller-Eberhard, 1970) in the total absence ofantibody, was shown to be capable of lysing VCN treated cells but not cells exposed toheat-inactivated VCN. The findings were, in addition, shown to be the result of the enzy-matic action of VCN, since the effect was inhibited by sialic acid (Ray et al., 1970), aspecific feedback inhibitor of neuraminidase.The present studies demonstrate that VCN shares the property of increasing the sensitivity
of lymphoid cells to the lytic action of serum C with another bacterial neuraminidase,CPN. IVN appears to be totally inactive in this regard. The bacterial and viral neuramini-dases, despite their similar nomenclature and their similar ability to lyse the 0-glycosidiclinkage between sialic acid residues and carbohydrates, are serologically different enzymes(Madoff, Eyler & Weinstein, 1960; Ada, Lind & Laver, 1963). In addition they have differentsubstrate specificities. VCN can apparently split all three of the identified types of alpha-keto glycosidic linkages (2-3', 2-6' and 2-8') between the neuraminic acids and carbo-hydrates. Viral neuraminidases can lyse only the 2-3' and 2-8' linkages (Drzeniek, 1967;Drzeniek & Gaube, 1970; Schneir & Rafelson, 1966). The present results demonstrate that

Viral and bacterial neuraminidase 147
breaking the 2-3' and/or 2-8' linkage between the terminal sialic acid residues and othermucopolysaccharides on the cell surface has no discernible effect on the sensitivity of thecell to C. However, breaking the 2-3', 2-6' and 2-8' glycosidic linkages markedly increasesthe sensitivity of the cells to C.
It thus appears that the 2-6' glycosidic linkage between the terminal sialic acid residuesand the cell surface carbohydrates is crucial to the observed increase in C sensitivity. Whatis not clear, however, is whether breaking the 2-6' linkage is important by itself, or onlyimportant when the 2-3' and/or 2-8' linkages are also broken. Furthermore it is not clearwhether the results are a function of the possibility that the 2-6' linkage predominates onthe lymphoid cell surface. If this were so, merely removing a given quantity of sialic acidwith its associated electro-negative charge from the cell might, in itself, sensitize the cell toC regardless of the linkage involved.
Quantitative studies of the amount of sialic acid released from lymphoid cells by VCNand IVN were, therefore, carried out. The preliminary results reported here demonstratethat the 2-6' glycosidic linkage between sialic acid and mucopolysaccharides is the pre-dominant linkage by which sialic acid is bound to the lymphoid cell surface. The 2-3' and2-8' linkages appear to be relatively uncommon on the cell surfaces as evidenced by therelatively small amount of sialic acid released by IVN. Since an IVN unit will release moresialic acid from a standardized substrate than will a VCN unit, and since increasing theconcentration of IVN did not lead to further increases in sialic acid release, it would appearthat the accessible sialic acid residues are bound primarily by the 2-6' linkage.
This data alone, however, does not permit one to answer the question whether it is theabsolute number of sialic acid residues removed that renders the cell susceptible to C me-diated lysis, or whether it is the specific rupture of the 2-6' linkage which does so. Lowconcentrations ofVCN release an amount of sialic acid equivalent to that released by IVN.Our previous studies have shown that very dilute concentrations of VCN are capable ofsensitizing the cells to C mediated lysis. These facts give support to the idea that it is not thequantitative amount of sialic acid released from the cell surface that is important but ratherthe number of 2-6' linkages ruptured. If this conclusion is correct, then it is probably not thechange in surface negative charge which influences C sensitivity, but rather the change in thestearic configurations of the cell surface induced by rupturing the 2-6' linkage.Whatever the case, it appears that sialic acid bound by the 2-6' linkage has an inhibitory
effect on the action of C on the cell membrane. It may be significant in this regard that theesterase activity (Cl esterase) associated with the first component of human C can beblocked by a protein inhibitor in normal human serum (Ratnoff & Lepow, 1957) which hassince been identified as an alpha2-neuramino glycoprotein (Pensky & Schwick, 1969),which is, in turn, a substrate for neuraminidase. Furthermore, an inhibitory role of mem-brane sialic acid in the interaction between red blood cells andChas been previously suggested(Yachnin, Laforet & Gardner, 1961; Arquilla Hamashige & Miller, 1964; Dalmasso &Muller-Eberhard, 1964). There is little information, however, whether breaking the 2-6'linkage acts to (a) unmask specific C binding sites, (b) change the avidity of such sites for C,(c) sensitize the C binding sites so that less C needs to be bound to effect lysis, (d) causenonspecific damage to the cell membrane so that less C is required to effect lysis, or (e)merely changes the cell charge so that the highly negatively charged C molecules are morereadily attracted to the relatively positively charged cell surface. The findings of Dalmasso& Muller-Eberhard (1964) that there is increased binding of C in the presence of polyeth-

148 P. K. Ray and R. L. Simmons
ylene glycol by human red blood cells which had been treated with VCN would suggestthat either C binding sites were unmasked, or their avidity changed by VCN treatment.The difference in activity of bacterial and viral neuraminidases may play some role in the
pathogenesis of bacterial and viral infection. Bacterial neuraminidases could, theoretically,sensitize host cells to endogenous C. Preformed antibody interaction with bacterial endo-toxins, which are known to activate the C-cascade may then contribute to host cell death(Gewurz, 1971). It appears that influenza virus neuraminidase must act by a differentmechanism since rupturing of the 2-3' and 2-8' linkages on some cells at least, does notrender the cells sensitive to C. It is still possible, however, that the effect ofC sensitivity of thecells depends not on the specific linkage, but merely on removing a large amount of sialicacid from the cell surface itself. If this were true, cells which bound sialic acid via the 2-3'and 2-8' linkages would be susceptible to influenza virus neuraminidases and those cellswhich contained primarily the 2-6' linkage would be susceptible to bacterial neuraminidase.Such surface chemical linkages may well play a role in the determination of the target organof a given infecting organism. For example, the respiratory epithelium which appears tobe a target organ for influenza virus may bear an increased number of 2-3' or 2-8' linkagesbetween the sialic acid and carbohydrate on the cell surface whereas the gut epitheliumattacked by Vibric cholerae organism may possess primarily the 2-6' linkages.
ACKNOWLEDGMENTS
This work was supported by Grant No. 5 Rol CA 11605-02 from the United States PublicHealth Service.Dr R. L. Simmons is a John and Mary Markle Scholar in Academic Medicine.
REFERENCES
ADA, G.L., LIND, P.E. & LAVER, W.G. (1963) An immunological study of avain, viral and bacterial neura-
minidases based on specific inhibition of enzyme by antibody. J. gen. Microbiol., 32, 225.AMBROSE, E.J. (1966) Electrophoretic behavior of cells. Prog. Biophys. molec. BiC. 16, 241.AMINOFF, D. (1961) Methods for the quantitative estimation of N-acetyl neuraminic acid and their application
to hydrolysates of sialomucoids. Biochem. J. 81, 384.ARQUILLA, E.R., HAMASHIGE, S. & MILLER, A. (1964) Inhibition of immune hemolysis by glycoprotein
antigens. Fed. Proc. 23, 506.BAGSHAWE, K.D. & CURRIE, G.A. (1967) Immunogenicity of L 1210 murine leukaemia cells after treatment
with neuraminidase. Nature (Lond.), 218, 1254.BAYLOR, M.E. (1964) The reaction of receptor glycoprotein with influenza virus and neuraminidase: An
electron microscopic study. Trans. N. Y. Acad. Sci. 26, 1103.BENEDETTI, E.L. & EMMELOT, P. (1967) Studies on plasma membranes. IV. The ultrastructural localization
and content of sialic acid in plasma membranes isolated from rat liver and hepatoma. J. Cell Sci. 2, 499.BROWN, D.M. & MICHAEL, A.F. (1969) Effect of neuraminidase on the accumulation of alpha-amino-iso-
butyric acid in Hela cells. Proc. Soc. exp. Biol. (N. Y.), 131, 568.CODINGTON, J.F., SANFORD, B.H. & JEANLOZ, R.W. (1970) Glycoprotein coat of the TA3 cell. I. Removal of
carbohydrates and protein material from viable cells. J. nat. Cancer Inst. 45, 637.CURRIE, G.A. & BAGSHAWE, K.D. (1967) The masking of antigens on trophoblast and cancer cells. Lancet, i,
708.CURRIE, G.A. & BAGSHAWE, K.D. (1968) The role of sialic acid in antigenic expression: Further studies of the
Landschutz ascites tumour. Brit. J. Cancer, 22, 843.CURRIE, G.A., VAN DOORNINCK, W. & BAGSHAWE, K.D. (1968) Effect of neuraminidase on the immuno-
genicity of early mouse trophoblast. Nature (Lond.), 219, 191.

Viral and bacterial neuraminidase 149CURRIE, G.A. & BAGSHAWE, K.D. (1969) Tumour specific immunogenicity of methylcholanthrene-induced
sarcoma cells after incubation in neuraminidase. Brit. J. Cancer, 23, 141.DALMASSO, A.P. & MOLLER-EBERHARD, H.J. (1964) Interaction of autologous complement with red cells in
the absence of antibody. Proc. Soc. exp. Biol. (N. Y.) 117, 643.DRZENIEK, R. (1967) Differences in splitting capacity of virus and Vibrio cholerae neuraminidases on sialic
acid type substrates. Biochem. biophys. Res. Commun. 26, 631.DRZENIEK, R. & GAUBE, A. (1970) Differences in substrate specificity of myxovirus neuraminidases.
Biochem. biophys. Res. Commun. 38, 651.GESNER, B. & THOMAS, L. (1966) Sialic acid binding sites: Role in hemagglutination by Mycoplasmagallisepti-
cum. Science, 151, 590.GEWURZ, H. (1971) Interaction between endotoxic lipopolysaccharides (LPS) and complement (C) system:
Solubilization of C-consuming substances during brief absorptions at 00. Proc. Soc. exp. Biol. (N. Y.)136, 561.
GOTZE, 0. & MULLER-EBERHARD, H.J. (1970) Lysis of erythrocytes by complement in absence of antibody.J. exp. Med. 132, 898.
KASSULKE, J.T., STUTMAN, O., YUNIS, E.J. (1971) Blood-group isoantigens in leukemic cells: Reversibility ofisoantigenic changes by neuraminidase. J. nat. Cancer Inst. (In press).
KEMP, R.B. (1968) Effect of the removal of cell surface sialic acids on cell aggregation in vitro. Nature(Lond.), 218, 1255.
KINNE, D.W. & SIMMONS, R.L. (1967) Passive serologic transfer of allograft immunity in immunologicallysuppressed mice. I. Immunosuppression by heterologous antilymphocyte serum. J. Immunol. 98,251.
KRAEMER, P.M. (1968) Cytotoxic, hemolytic and phospholipase contaminants of commercial neuraminidases.Biochim. biophys. Acta. (Amst.) 167, 205.
LEE, A. (1968) Effect of neuraminidase on the phagocytosis of heterologous red cells by mouse peritonealmacrophages. Proc. Soc. exp. Biol. (N. Y.), 128, 891.
LINDENMANN, J. & KLEIN, P.A. (1967) Recent Results in Cancer Research, 9. Immunologic aspects of viraloncolysis, p. 66, Springer-Verlag, New York.
LUNDGREN, G., JEITZ, L., LUNDIN, L. & SIMMONS, R.L. (1971) Increased stimulation by neuraminidasetreated cells in mixed lymphocyte cultures. Fed. Proc. 30, 395.
MADOFF, M.A., EYLER, E.H. & WEINSTEIN, L. (1960) Serologic studies of the neuraminidases of Vibriocholerae, Diplococcus pneumoneae and influenza virus (PR-8). J. Immunol. 85, 603.
PENSKY, J. & SCHWICK, H.G. (1969) Human serum inhibitor of C'1 esterase: Identity with alpha 2-neura-minoglycoprotein. Science, 163, 698.
RATNOFF, O.D. & LEPOW, I.H. (1957) Some properties of an esterase derived from preparations of the firstcomponent of complement. J. exp. Med. 106, 327.
RAY, P.K., GEWURZ, H. & SIMMONS, R.L. (1970) The mechanism of increased sensitivity of neuraminidasetreated cells to antibody induced cytolysis. Fed. Proc. 29, 573.
RAY, P.K., GEWURZ, H. & SIMMONS, R.L. (1971) Complement sensitivity of neuraminidase treated lymphoidcells. Transplantation, 12, 327.
RAY, P.K., & SIMMONS, R.L. (1971) Failure of neuraminidase to unmask allogeneic antigens on cell suraces.Proc. Soc. exp. Biol. (N. Y.) (In press).
SANFORD, B.H. (1967) An alteration in tumor histocompatibility induced by neuraminidase. Transplanation,5, 1273.
SCHLESINGER, M. & AMos, D.B. (1971) The effect of neuraminidase on the serological properties of murinelymphoid cells. Transplant. Proc. 111, 895.
SCHNEIR, M.L. & RAFELSON, M.E. (1966) Isolation and characterization of two structural isomers of N-acetyl neuraminyllactose from bovine colostrum. Biochim. Biophys. Acta. (Amst.) 130, 1.
SHIN, H.S., GEWURZ, H. & SYNDERMAN, R. (1969) Reactions of a cobra venom factor with guinea pigcomplement and generation of an activity chemotactic for polymorphonuclear leukocytes. Proc. Soc.exp. Biol. (N. Y.) 131, 203,
SIMMONS, R.L., RIos, A. & RAY, P.K. (1970) Mechanism of neuraminidase induced antigen 'unmasking'Surg. Forum, 21, 265.
SIMMONS, R.L., RIos, A. & RAY, P.K. (1971a) Immunogenicity and antigenicity of lymphoid cells treated withneuraminidase. Nature New Biol. 231, 179.

150 P. K. Ray and R. L. Simmons
SIMMONS, R.L., LIPSCHULTZ, M.L., Rios, A. & RAY, P.K. (1971b) Failure of neuraminidase to unmaskhistocompatibility antigens on trophoblasts. Nature New Biol. 231, 111.
SIMMONS, R.L., Rios, A., LUNDGREN, G., RAY, P.K. MCKHANN, C.F. & HAYWOOD, G. (1971c) Immuno-specific regression of methylcholanthrene fibrosarcoma using neuraminidase, Surgery, 70, 38.
WEISS, L., (1965) Studies on cell deformability. I. Effect of surface charge. J. Cell Biol. 26, 735.WEISS, L., MAYHEW, E. & ULRICH, K. (1966) The effect of neuraminidase on the phagocytic process in
human monocytes. Lab. Invest. 15, 1304.WIGZELL, H. (1965) Quantitative titrations of mouse H-2 antibodies using Cr-" labelled target cells. Trans-
plantation, 3, 380.WOODRUFF, J.J. & GESNER, B.M. (1969) The effect of neuraminidase on the fate of transfused lymphocytes.
J. exp. Med. 129, 551.YACHNIN, S., LAFORET, M.T. & GARDNER, F.H. (1961) PH dependent hemolytic systems. I. Their relationship
to paroxysmal nocturnal hemoglobinuria. Blood, 17, 83.