comparative physiological studies of the yeast - journal of
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Mar. 1971, p. 690-700Copyright © 1971 American Society for Microbiology
Comparative Physiological Studies of the Yeastand Mycelial Forms of Histoplasma capsulatum:
Uptake and Incorporation of L-LeucineRISHAB K. GUPTA' AND DEXTER H. HOWARD
Department of Medical Microbiology and Immunology, School of Medicine, University of California, LosAngeles, California 90024
Received for publication 16 November 1970
L-Leucine entered the cells of both morphological forms of Histoplasma capsu-
latum by a permease-like system at low external concentrations of substrate. How-ever, at levels greater than 5 x 10-5 M L-leucine, the amino acid entered the cellsboth through a simple diffusion-like process and the permease-like system. The rateof the amino acid diffusion into yeast and mycelial forms appeared to be the same,whereas the initial rate of accumulation through the permease-like system was 5 to10 times faster in the mycelial phase than it was in the yeast phase. The Michaelisconstants were 2.2 x 10-5 M in yeast phase and 2 x 10-5 M in mycelial phase cells.Transport of L-leucine at an external concentration ol ll0) M showed all of the char-acteristics of a system of active transport, which was dependent on temperature andpH. Displacement or removal of the a-amino group, or modification of the a-car-boxyl group abolished amino acid uptake. The process was competitively inhibitedby neutral aliphatic side-chain amino acids (inhibition constants ranged from 1.5 x10-5 to 6.2 x 10-5 M). Neutral aromatic side-chain amino acids and the D-isomersof leucine and valine did not inhibit L-leucine uptake. These data were interpreted tomean that the L-leucine transport system is stereospecific and is highly specific forneutral aliphatic side-chain amino acids. Incorporation of L-leucine into macromo-lecules occurred at almost the same rate in both morphological forms of the fungus.The mycelial phase but not the yeast phase showed a slight initial lag in incorpora-tion. In both morphological forms the intracellular pool of L-leucine was of limitedcapacity, and the total uptake of the amino acid was a function of intracellular poolsize. The initial rate of L-leucine uptake was independent of the level of intracellularpool. Both morphological forms deaminated and degraded only a minor fraction ofthe accumulated leucine.
The transport system(s) involved in the uptakeof amino acids by the yeast and mycelial forms ofHistoplasma capsulatum (13, 23, 25) is the sub-ject of this report. The purpose of our researchwas (i) to compare amino acid transport and in-corporation in the two morphologic forms of a
dimorphic zoopathogic fungus, and (ii) to extendobservations on transport mechanisms in fila-mentous fungi in general (9, 27, 28). In addition,changes in amino acid uptake and incorporationwhich occur during the transition of the yeastphase to the mycelial phase of H. capsulatum are
reported (5, 17, 19).
MATERIALS AND METHODSOrganism. H. capsulatum strain 505 (Fungus Collec-
tion, Department of Medical Microbiology and Immu-nology, University of California, School of Medicine,
Present address: Department of Microbiology, School of Medicine,Yale University, New Haven, Conn. 06510.
690
Los Angeles) was used. The organism was maintainedin the yeast phase on blood-glucose-cysteine agar slants(7) incubated at 37 C for 3 days and stored in the refrig-erator. Subcultures were made at monthly intervals.
Cultivation of the yeast phase. Cells were grown in
Salvin's medium [10 g of casein hydrolysate, 3 g of
yeast extract, 3 g of glucose, 2.5 g of NaCl, 2.5 g ot'KCI, 4 g of sodium phosphate (dibasic), and 0.5 g of
cysteine hydrochloride per liter of distilled water, pH 7(reference 26)] at 37 C on a gyratory shaker (NewBrunswick Scientific Co., New Brunswick, N.J.) for 72hr. Cells were harvested by centrifugation at 1,100 x gfor 10 min at room temperature, washed once, and sus-
pended with a salt solution of low ionic strength [KCI,0.75 g; CaCl2 (anhydrous), 0.27 g; and MgCla.6H2O,0.5 g per liter of distilled water (reference 29)]. Salvin'smedium (50 ml) in a 250-ml Erlenmeyer flask was inoc-ulated with washed cells (105/ml), and the flasks were
incubated under conditions mentioned above. After 48hr of incubation, 5 ml of the culture was transterred toanother 45 ml of fresh Salvin's medium, and this cul-ture was incubated for an additional 48 hr. This was
Vol. 105, No. 3Printed in U.S.A.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.

L-LEUCINE IN H. CAPSULATUM
done so that the cells, at the time of harvest, would beof the same approximate physiological age for uptakestudies.
Cultivation of mycelial phase. Five-milliliter culturesof yeast phase cells were inoculated into 45 ml of Sal-vin's medium and incubated at 37 C on a shaker for 24hr. The flasks were transferred to room temperature (23to 24 C) and shaken for an additional 60 to 72 hr.About 85% of the yeast cells germinated under theseconditions. The germinated cells were used for uptakeand incorporation studies of the mycelial phase of H.capsulatum.
Preparation of cells for uptake studies. Cultures were
used only when the viability was 95% or above as deter-mined by the eosin-Y exclusion technique (12). Cellswere harvested, washed three times with 0.02 M sodium-potassium phosphate buffer at pH 7, and resuspendedin the buffer. The suspensions were diluted to a celldensity of 2 mg (dry weight) per ml and stored in an icebath until used.
Measurement of L-leucine uptake and incorporation.Throughout these studies, uniformly labeled "C-L-leu-cine, purchased from New England Nuclear Corp.,Boston, Mass., was used. Because of high specific ac-tivity (316 mCi/mmole) of "C-L-leucine, it was dilutedwith unlabeled L-leucine (Calbiochem, Los Angeles,Calif.) for uptake studies.
Unless described as otherwise, L-leucine uptake was
measured under the following standard conditions. Thecell suspension was placed in a 50-ml Erlenmeyer flaskand treated with a solution of 14C-L-leucine; thus, thefinal mixture contained 1.0 mg (dry weight) of cells and10 nmole (approximately 10,000 counts/min) of 14C-L-leucine per ml of 0.02 M phosphate buffer at pH 7. Themixtures were incubated at 37 C on a gyratory waterbath shaker (New Brunswick Scientific Co., NewBrunswick, N.J.). Samples (1.0 ml) were withdrawn andfiltered through membrane (0.45-mm pore size) filters(Millipore Corp., Bedford, Mass.). Portions (0.5 ml) ofthe filtrate were used to measure the "4C-activity. Thedecrease in radioactivity at various time intervals com-
pared to the whole suspension represented the uptake ofL-leucine.To measure incorporation of 14C activity, duplicate
1.0-ml samples were withdrawn at appropriate timeintervals. One sample was treated as mentioned above toassess total uptake. The other sample was mixed with1.0 ml of precooled (0 to 2 C) 10% trichloroacetic acid.After 30 min, exactly 1.0-ml portions of this mixturewere filtered, and a 0.5-ml portion was used for radio-active measurements. The membrane filter which re-tained the cells was washed with 15 ml of 5% cold tri-chloroacetic acid. The washed filter was solubilized in 10ml of the scintillation liquid and tested for radioactivity(cold trichloroacetic acid-insoluble fraction). Radioac-tivity associated with the cold trichloroacetic acid-sol-uble fraction was computed by subtracting the 14C ac-
tivity of the filtrate collected without acid treatmentfrom the 14C activity of the filtrate collected after coldacid treatment.
All radioactive measurements were made in a Nu-clear-Chicago liquid scintillation counter (Nuclear-Chi-cago Corp., Des Plaines, 111.) which had approximately70% efficiency for 14C in a scintillation liquid of the fol-lowing composition: 7 g of 2,5-diphenyloxazol, 0.05 g
of 1,4-bis-2-(5-phenyloxazolyl)-benzene, and 70 g ofnaphthalene in 1.0 liter of p-dioxane (4). Samples werecounted for a minimum of 10 min, or for a time ade-quate to give at least 2,000 counts per min per sample.
Chromatographic analysis of accumulated L-leucine.Five-milliliter yeast phase cell suspensions, exposed to10-5 M 14C-L-leucine for 15 min, were collected on amembrane filter, washed twice with 15 ml of cold (4 C)distilled water, and suspended in 1.0 ml of distilledwater. The suspension was heated in a boiling-waterbath for 10 min and filtered again through a membranefilter. The filtrate was collected and used to analyze thechemical nature of accumulated 14C activity by silica gelthin-layer chromatography. Similar procedures wereused to extract mycelial phase cells exposed to 10-' M4C-L-leucine for 5 min.Extracts (20 ,liters) were placed on a silica gel
chromagram sheet 6061 (Eastman Kodak Co., Roches-ter, N.Y.). "C-L-leucine was used as standard control.The silica gel sheet was developed in a methanol-n-bu-tanol-benzene- water (2:1:1) solvent system. An auto-radiogram of the chromatogram was prepared by ex-posing a no-screen X-ray film (Eastman Kodak Co.,Rochester, N.Y.) to the silica gel sheet for I week.Most of the radioactivity in the extract was associatedwith the spot corresponding to the authentic L-leucine.
Isolation and identification of a-ketoisocaproic acid.a-Ketoisocaproate was precipitated as its 2,4-dinitro-phenyl hydrazone derivative. Two milliliters of the testmaterial was treated with I ml of 0.2% 2,4-dinitro-phenyl hydrazine dissolved in 2 N HCI. The mixturewas allowed to stand for 12 hr in the dark at 4 C. Au-thentic a-ketoisocaproate (Calbiochem, Los Angeles,Calif.) was used as carrier at a concentration of 0.5% toobtain a detectable amount of the precipitate. The pre-cipitate generated during the reaction was collected bycentrifugation at 2,000 x g for 20 min, washed threetimes with 5-ml portions of cold distilled water, andrecrystallized from absolute ethanol. Uniformly labeled"4C-L-leucine was treated similarly as a control. Thepurified precipitate of the a-keto acid hydrazone wasdissolved in I ml of ethanol and tested for 14C activityin a scintillation counter. The 14C activity associatedwith the precipitate was determined by thin-layer chro-matography on silica gel plates (Eastman Kodak Co.,Rochester, N.Y.) and autoradiography as described byDancis et al. (6).
RESULTSTime course of L-leucine incorporation. Figures
I and 2 represent the results of one of severalexperiments on the uptake and incorporation ofL-leucine by the yeast and mycelial phases of H.capsulatum. Yeast phase cells accumulated theamino acid as a linear function of time until theexogenous supply became limiting. The accumu-lated amino acid was incorporated into the coldtrichloroacetic acid-insoluble (acid-insoluble)fraction without any lag (Fig. IA). The absenceof a lag was noted even when samples were takenat 1.0-min instead of 5-min intervals (data notshown). After 40 min, the rate of incorporationinto the acid-insoluble fraction slowed down to
691VOL. 105, 1971
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.
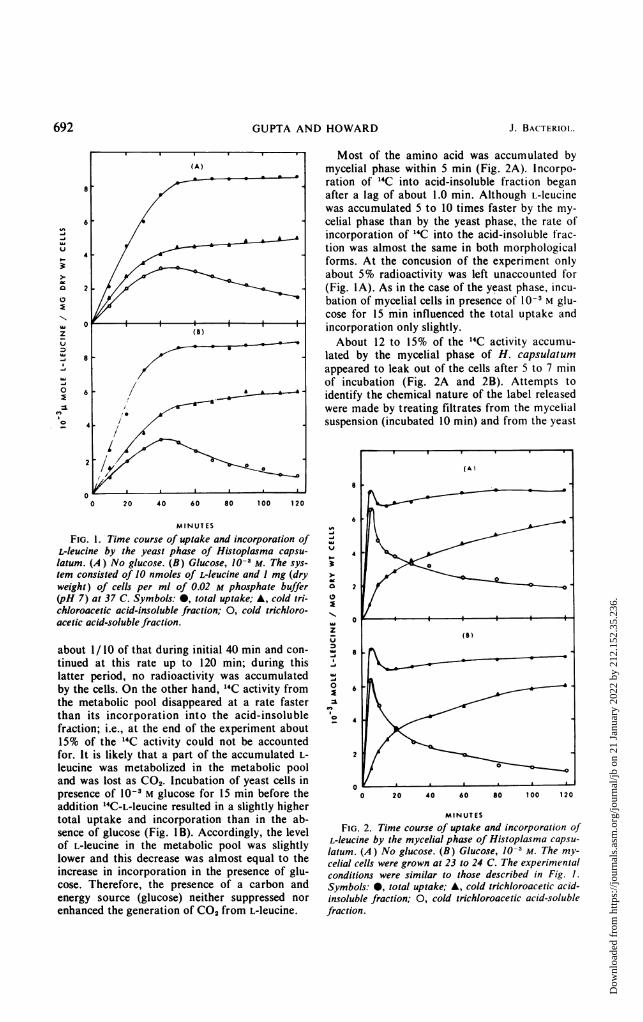
692 GUPTA AND HOWARD J. BACTERIOI.
Most of the amino acid was accumulated by(A) mycelial phase within 5 min (Fig. 2A). Incorpo-
ration of 14C into acid-insoluble fraction beganafter a lag of about 1.0 min. Although L-leucinewas accumulated 5 to 10 times faster by the my-
6 . C celial phase than by the yeast phase, the rate of-J /incorporation of 14C into the acid-insoluble frac-v
4tion was almost the same in both morphologicalforms. At the concusion of the experiment onlyabout 5% radioactivity was left unaccounted for
2-//.....(Fig. IA). As in the case of the yeast phase, incu-bation of mycelial cells in presence of 10-3 M glu-
.....cose for 15 min influenced the total uptake andzu 0 _ § § § I § § incorporation only slightly.
_ . . . . About 12 to 15% of the 14C activity accumu-8 lated by the mycelial phase of H. capsulatum
appeared to leak out of the cells after 5 to 7 min.J /of incubation (Fig. 2A and 2B). Attempts too 6 / . _ _ _ identify the chemical nature of the label releasedZL / were made by treating filtrates from the mycelialO 4 t Z / 1 suspension (incubated 10 min) and from the yeast
,,/2~~~~~~~~0(A~/~~~~~~~~~~~~~~~~~~~~
00 20 40 60 80 100 120
6MINUTES
FIG. 1. Time course of uptake and incorporation of tVL-leucine by the yeast phase of Histoplasma capsu- 4
latum. (A) No glucose. (B) Glucose, l0- M. The sys- 3tem consisted of 10 nmoles Of L-leucine and I mg (dry >.weight) of cells per ml of 0.02 M phosphate buffer 2(pH 7) at 37 C. Symbols: 0, total uptake; A, cold tri- £chloroacetic acid-insoluble fraction; 0, cold trichloro- N.
acetic acid-soluble fraction. 0zZ (B)
about 1/10 of that during initial 40 min and con- ' 8tinued at this rate up to 120 min; during this ,!.latter period, no radioactivity was accumulated ,by the cells. On the other hand, 14C activity from 0 6the metabolic pool disappeared at a rate faster \than its incorporation into the acid-insoluble O 4fraction; i.e., at the end of the experiment about15% of the 14C activity could not be accountedfor. It is likely that a part of the accumulated L- 2leucine was metabolized in the metabolic pooland was lost as CO2. Incubation of yeast cells in ____._. _. _. _.__presence of 10-3 M glucose for 15 min before the 0 20 40 60 80 100 120addition "4C-L-leucine resulted in a slightly highertotal uptake and incorporation than in the ab- MINUTESsence of glucose (Fig. IB). Accordingly, the level FIG. 2. Time course of uptake and incorporation ofof L-leucine in the metabolic pool was slightly L-leucine by the mycelial phase of Histoplasma capsu-lofer aeunethis decrease was almost equal tothe latum. (A) No glucose. (B) Glucose, 10-3 M. The my-ower and this decrease was almost equal to the celial cells were grown at 23 to 24 C. The experimentalincrease in incorporation in the presence of glu- conditions were similar to those described in Fig. 1.cose. Therefore, the presence of a carbon and Symbols: 0, total uptake; A, cold trichloroacetic acid-energy source (glucose) neither suppressed nor insoluble fraction; 0, cold trichloroacetic acid-solubleenhanced the generation of CO2 from L-leucine. fraction.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.

L-LEUCINE IN H. CAPSULATUM
suspension (incubated 30 min) with 2,4-dinitro-phenyl hydrazine in the presence of authentic a-ketoisocaproic acid. The precipitates were chro-matographed on a silica gel thin-layer platewhich was exposed to X-ray film for 5 weeks.The spots from the chromatogram were scrapedoff and tested for radioactivity. The starting ma-terial, 14C-L-leucine, contained no radioactive a-ketoisocaproate, whereas the precipitates fromthe filtrates of yeast and mycelial suspensionswere radioactive. Under identical conditions a-ketoisocaproate from yeast phase suspension fil-trate had 30 counts/min, whereas that from my-celial phase suspension filtrate had 130counts/min, about a fourfold difference. There-fore, a part of the L-leucine accumulated by themycelial phase cells was deaminated to a-keto-isocaproate which was released into the medium.Yeast phase cells were much less active in deami-nating L-leucine.
Kinetics of L-leucine uptake. The initial ratesof L-leucine uptake by yeast and mycelial formsat external concentrations of the amino acid areshown in Fig. 3 and 4. Neither of the two mor-
1-.
I _
ZI
zD
LLJ x
L_ X
I -
< oU
- I
z
5
4
3
2
1
0
phological forms of H. capsulatum showed acomplete saturation phenomenon. At low ex-ternal concentrations of L-leucine, the rate ofuptake of the amino acid by yeast phase in-creased rapidly with increasing external concen-tration. This rapid increase started to diminishwhen the external concentration of L-leucine wasapproximately 5 x 10-' M; beyond this level, anyincrease in the external concentration resulted ina slow but linear increase in the initial rate of L-leucine uptake (Fig. 3). The saturation curve ob-tained with the mycelial phase was similar to thatwith the yeast phase except that the initial rate ofL-leucine uptake was always higher at any pointcompared to yeast phase (Fig. 4).The biphasic nature of the saturation curves in
both morphological forms indicated that theamino acid uptake was a complex process, andwas due to at least two distinct transport systems.At low external concentrations a permease systemsaturable at higher concentrations of L-leucinewas operative; however, at higher external con-centrations (greater than 5 x 10-5 M), the aminoacid probably entered the cells through simple
0 1 0 20 30 40 50
10 M L-LEUCINEFIG. 3. Effect of external substrate concentration on the initial rate of L-leucine uptake by the yeast phase of
Histoplasma capsulatum. Initial rate of L-leucine uptake per minute per milligram (dry weight) of cells was com-puted from observations made at 5-, 10-, and 15-min intervals. The reaction mixture (37 C) contained 1.0 mg (dryweight) ofyeast cells per ml suspended in 0.02 M phosphate buffer (pH 7) and 10-5 M 14C- L-leucine (approximately10,000 counts per min per ml). Double-reciprocal plot of the data. V = micromoles of L-leucine per minute permilligram (dry weight) ofcells.
VOL. lOS, 1971 693
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.

GUPTA AND HOWARD J. BACTERIOL.
0 20 40 60 80 100
10CM L-LEUCINEFIG. 4. Effect of external substrate concentration on the initial rate of L-leucine uptake by mycelial phase of
Histoplasma capsulatum. The experimental conditions were similar to those described for Fig. 3 except that theuptake rate was computed from the observations made at 1-, 2.5-, and 5-min intervals. Double-reciprocal plot ofthe data. V = micromoles of L-leucine per minute per milligram (dry weight) of cells.
diffusion in addition to the permease-like satu-rable system.
Also shown in Fig. 3 and 4 are the uptake dataplotted by the method of Lineweaver-Burk (18).At higher concentrations the curves bent towardthe origin in both cases. The Michaelis-Mentenconstants (Km) at low external concentrations cal-culated for the amino acid transporting system ofyeast and mycelial forms of H. capsulatum were2.2 x 10-5 M and 2 x 10-5 M, respectively. TheKm values for the transport system operative athigher external amino acid concentration was the-oretically infinite, because the second phase of thedouble-reciprocal plot passed almost through theorigin, indicating that L-leucine was entering thecell by simple diffusion. If a process other thansimple diffusion were operative, a second butfinite value of the Km should have been observed(3, 8, 10, 30).
It was of interest to note (Fig. 3 and 4) that in-creases in the initial rate of uptake at higher ex-
ternal concentration of the amino acid were paral-lel, indicating that the amino acid was diffusinginto the yeast and mycelial cells at the same rate.On the other hand, the Vinax of the permease-liketransport system of the mycelial form was ap-proximately fivefold greater (6.3 x 10-3 ,moles/mg, dry weight) than that of the yeast form(1.66 x 10-3,umoles/mg, dry weight).
The results shown in Fig. 5 establish thatamino acid uptake was pH dependent and thatthe optimal level for uptake was pH 7. In bothmorphological forms the initial rate of L-leucineuptake increased linearly from pH 3 to 7, de-creased above pH 7, and was more sensitive topH below 7 than to pH above 7. Transport of L-leucine by yeast phase cells decreased more rap-
idly than that of mycelial phase cells when thepH of the system was lowered from 7 to 3. Theuptake rate in both morphological forms was af-fected to a similar extent in the range of pH from7 to 10.
694
121
101D 0u I
Z (
Im x-J
zI -
O X0
- I
z
_ :2
Zi _1-
8
6
4
2
0
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.

L-LEUCINE IN H. CAPSULATUM
The initial rate of L-leucine uptake increasedwith increasing temperature from 0 to 40 C; therewas, however, a sharp decrease in the rate of up-take when the temperature was raised from 40 to45 C. This decrease was more pronounced in theyeast phase (about sixfold) than it was in themycelial phase (about twofold). Since the de-creased rate of uptake was not restored when thecells exposed to 45 C for 30 min were used tomeasure the uptake rate at 37 C, the decrease inuptake rate was probably due to the thermaldeath of the cells at 45 C. Both morphologicalforms of H. capsulatum showed maximum initialrate of L-leucine uptake at 40 C. The temperaturecoefficient (Q,0) values calculated between 20 and30 C were approximately 2.1 and 2.5 for yeastand mycelial phases, respectively. Mycelial phasetransported L-leucine about 15-fold faster thanyeast phase between 0 and 40 C at an externalsubstrate concentration of 10-5 M.
Effect of metabolic inhibitors on L-leucine up-take. Sodium azide (10-3 M), which interfereswith the electron transport system of cells, inhib-ited uptake about 30% in both morphologicalforms. 2,4-Dinitrophenol (10-3 M), which uncou-ples oxidative phosphorylation, reduced uptake23 and 33% in yeast and mycelial phases, respec-tively. Arsenate, which competes for phosphate inenergy-yielding reactions, inhibited uptake about20% in both morphologic forms. That these met-abolic inhibitors did not exert a more pronouncedeffect on uptake was undoubtedly due to the factthat, when the permease-like transport system
2.0
uiidP.--
t .;0
UAz -19 0:2ti x2, -1t z
:2LU x1-- .jI 0-i :2P I=LZ o
cl,
was partly or completely inactivated, the aminoacid could still enter the cells by diffusion.
Effect of other amino acids and structurally re-lated compounds on L-leucine uptake. Arginine,lysine, and histidine (basic amino acids), aspar-tate (acidic amino acid), and tryptophan, phenyl-alanine, and tyrosine (neutral amino acids withan aromatic side chain) did not inhibit leucineuptake. Indeed, lysine and phenylalanine stimu-lated leucine uptake 70 to 75% above controlvalues by yeast phase but not mycelial phasecells.
Cysteine, asparagine, and glutamine did not oronly slightly inhibited the uptake of leucine byeither morphological form of the fungus; how-ever, other neutral amino acids with aliphatic sidechains inhibited leucine uptake by both forms ofthe fungus. The preceding data clearly suggestthat the permease-like system specifically boundonly neutral amino acids with an aliphatic sidechain, and that removal of the negative chargefrom the side chain of the amino acids did notenable them to inhibit leucine uptake. Further-more, since the initial rate of L-leucine uptakewas not inhibited by the D-isomers of leucine orvaline, the permease-like system was stereospe-cific.Among neutral amino acids, alanine, serine,
methionine, valine, and isoleucine were thestrongest inhibitors of L-leucine uptake by bothmorphological forms of H. capsulatum. The con-clusion that the aliphatic side chain of L-leucinewas required for binding to the transport site was
3.0
2.5
x
Z 2.0
x
X I.0
I
pH
3 5 7 9
pH
FIG. 5. Influence ofpH on L-leucine uptake by the yeast and mycelial phases of Histoplasma capsulatum at 37
C. Note: ordinate scale of mycelial phase is 20-fold larger than that ofyeast phase. Buffer systems used in this ex-
periment were: pH 3 and 4, phthalate (0.02 M); pH 5 to 7, phosphate (0.02 M); pH 8 and 9, tris(hydroxymethyl)
ethanolamine (0.02 M); pH 10, borate (0.02 M). Ingredients at the concentrations used in these buffer systems had
no effect on L-leucine uptake under standard conditions of Fig. 3. The initial rate of uptake of the amino acid was
computed from observations made at 5-, 10-, and IS-min intervals for the yeast phase, andfrom observations made
at 1-, 2.5-, and 5-min intervals for the mycelial phase.
MYCEUAL PRASE
695VOL. 105, 197 1
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.
GUPTA AND HOWARD
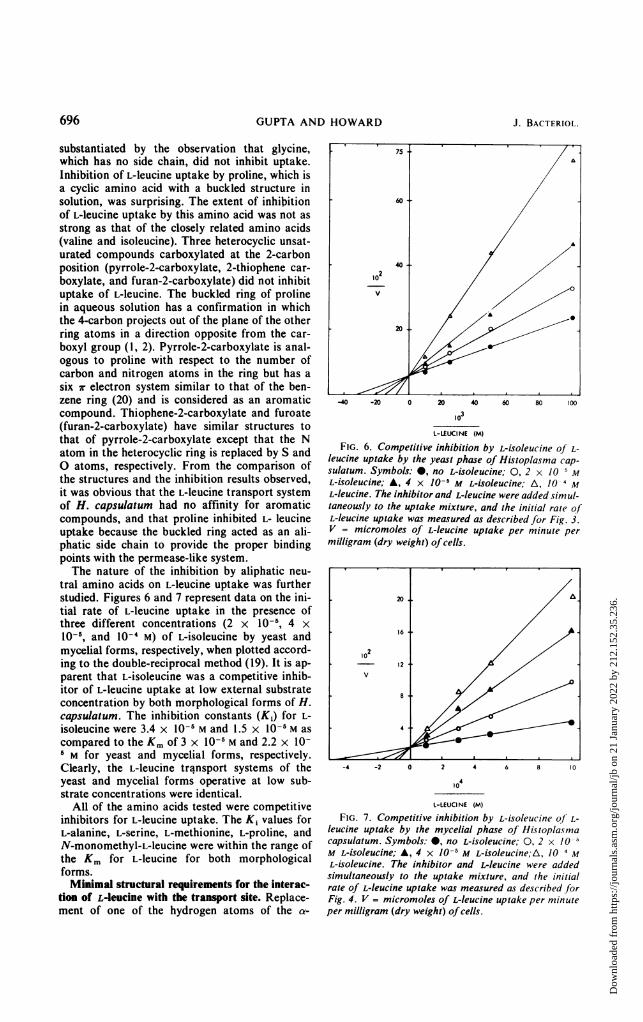
substantiated by the observation that glycine,which has no side chain, did not inhibit uptake.Inhibition of L-leucine uptake by proline, which isa cyclic amino acid with a buckled structure insolution, was surprising. The extent of inhibitionof L-leucine uptake by this amino acid was not asstrong as that of the closely related amino acids(valine and isoleucine). Three heterocyclic unsat-urated compounds carboxylated at the 2-carbonposition (pyrrole-2-carboxylate, 2-thiophene car-boxylate, and furan-2-carboxylate) did not inhibituptake of L-leucine. The buckled ring of prolinein aqueous solution has a confirmation in whichthe 4-carbon projects out of the plane of the otherring atoms in a direction opposite from the car-boxyl group (1, 2). Pyrrole-2-carboxylate is anal-ogous to proline with respect to the number ofcarbon and nitrogen atoms in the ring but has asix wr electron system similar to that of the ben-zene ring (20) and is considered as an aromaticcompound. Thiophene-2-carboxylate and furoate(furan-2-carboxylate) have similar structures tothat of pyrrole-2-carboxylate except that the Natom in the heterocyclic ring is replaced by S andO atoms, respectively. From the comparison ofthe structures and the inhibition results observed,it was obvious that the L-leucine transport systemof H. capsulatum had no affinity for aromaticcompounds, and that proline inhibited L- leucineuptake because the buckled ring acted as an ali-phatic side chain to provide the proper bindingpoints with the permease-like system.The nature of the inhibition by aliphatic neu-
tral amino acids on L-leucine uptake was furtherstudied. Figures 6 and 7 represent data on the ini-tial rate of L-leucine uptake in the presence ofthree different concentrations (2 x 10-5, 4 x10-6, and 10-4 M) of L-isoleucine by yeast andmycelial forms, respectively, when plotted accord-ing to the double-reciprocal method (19). It is ap-parent that L-isoleucine was a competitive inhib-itor of L-leucine uptake at low external substrateconcentration by both morphological forms of H.capsulatum. The inhibition constants (K1) for L-isoleucine were 3.4 x 10-5 M and 1.5 x 10-5 M ascompared to the Km of 3 x 10-5 M and 2.2 x 10-6 M for yeast and mycelial forms, respectively.Clearly, the L-leucine transport systems of theyeast and mycelial forms operative at low sub-strate concentrations were identical.
All of the amino acids tested were competitiveinhibitors for L-leucine uptake. The K1 values forL-alanine, L-serine, L-methionine, L-proline, andN-monomethyl-L-leucine were within the range ofthe Km for L-leucine for both morphologicalforms.Minimal structural requirements for the interac-
tion of L-leucine with the transport site. Replace-ment of one of the hydrogen atoms of the a-
-40 -20 0 20 40 60 so 100
lo'L-LEUCINE (M)
FIG. 6. Competitive inhibition by L-isoleucine of L-leucine uptake by the yeast phase of Histoplasma cap-sulatum. Symbols: 0, no L-isoleucine; 0, 2 x 10 5 ML-isoleucine; A, 4 x 10-5 M L-isoleucine; A, 10 4 ML-leucine. The inhibitor and L-leucine were added simul-taneously to the uptake mixture, and the initial rate ofL-leucine uptake was measured as described for Fig. 3.V = micromoles of L-leucine uptake per minute permilligram (dry weight) ofcells.
to4L-LEUCINE (M)
FIG. 7. Competitive inhibition by L-isoleucine of L-leucine uptake by the mycelial phase of Histoplasmacapsulatum. Symbols: 0, no L-isoleucine; 0, 2 x 10 5
M L-isoleucine; A, 4 x 10-5 M L-isoleucine;A, /0 4 ML-isoleucine. The inhibitor and L-leucine were addedsimultaneously to the uptake mixture, and the initialrate of L-leucine uptake was measured as described forFig. 4. V = micromoles of L-leucine uptake per minuteper milligram (dry weight) ofcells.
696 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.
L-LEUCINE IN H. CAPSULATUM
amino group of L-leucine with a hydrophobicmethyl group (as in the case of N-monomethyl-L-leucine) did not affect the uptake of the com-pound. Under the experimental conditions of thepresent investigation the test compound, N-mono-methyl-L-leucine, possessed a positivecharge on the secondary amino (imino) group.The presence of a methyl group on one side of thea-amino group and the rest of the molecules onthe other may not have allowed the functional (a-amino) group to come into close enough prox-imity to interact directly with the transport site.Therefore, the positive charge present on a-amino group was important, and this functionalgroup participated by an electrostatic interactionwith the transport site. Complete elimination ofthe a-amino group (as in a-ketoisocaproic acid)completely abolished L-leucine uptake. L-Leucineamide, a compound which is analogous to L-leu-cine except that the a-carboxyl group is blockedby converting it to an amide group, did not inter-fere with L-leucine uptake, indicating the a-car-boxyl group as an absolute requirement for L-leucine binding. a-Aminoisobutyric acid de-pressed L-leucine uptake about 50% in both mor-phological forms of the fungus. Thus, the a-hy-drogen of L-leucine molecule was not an absoluterequirement of affinity for the transport site be-cause it could be replaced with a methyl groupwithout affecting the transport of the amino acid.Neither f,-alanine nor y-aminobutyric acid,
lIg I
-IlLUIu
0cL
zLU
-J
-J
-J
I,)0
8
6
4
2
0
which represent a displacement of the aminogroup of leucine from a-position to ,3- and y-po-sition, respectively, inhibited L-leucine uptake.The inability of these compounds to decrease therate of the amino acid uptake meant that theamino group had to be in the a-position of themolecule to have an affinity for the transport site.Dependence of the rate of L-leucine uptake on
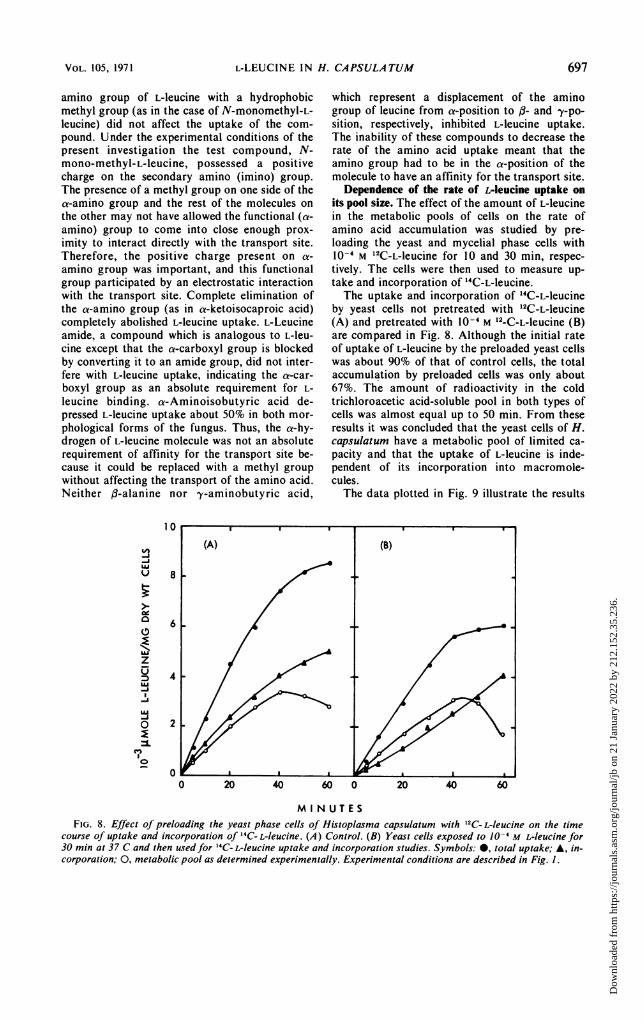
its pool size. The effect of the amount of L-leucinein the metabolic pools of cells on the rate ofamino acid accumulation was studied by pre-loading the yeast and mycelial phase cells with10-4 M 12C-L-leucine for 10 and 30 min, respec-tively. The cells were then used to measure up-take and incorporation of 14C-L-leucine.The uptake and incorporation of 14C-L-leucine
by yeast cells not pretreated with 12C-L-leucine(A) and pretreated with 10-4 M 12-C-L-leucine (B)are compared in Fig. 8. Although the initial rateof uptake of L-leucine by the preloaded yeast cellswas about 90% of that of control cells, the totalaccumulation by preloaded cells was only about67%. The amount of radioactivity in the coldtrichloroacetic acid-soluble pool in both types ofcells was almost equal up to 50 min. From theseresults it was concluded that the yeast cells of H.capsulatum have a metabolic pool of limited ca-pacity and that the uptake of L-leucine is inde-pendent of its incorporation into macromole-cules.The data plotted in Fig. 9 illustrate the results
0 20 40 60 0 20 40 60
MI NUTESFIG. 8. Effect of preloading the yeast phase cells of Histoplasma capsulatum with 12C- L-leucine on the time
course of uptake and incorporation of "C- L-leucine. (A) Control. (B) Yeast cells exposed to 10-4 M L-leucine for30 min at 37 C and then used for 14C- L-leucine uptake and incorporation studies. Symbols: 0, total uptake; A, in-corporation; 0, metabolic pool as determined experimentally. Experimental conditions are described in Fig. 1.
697VOL. 105, 1971
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.

GUPTA AND HOWARD
20 40 60
MINUTES
FIG. 9. Effect ofpreloading the mycelial phase cellsof Histoplasma capsulatum with 12C- L-leucine on thetime course of uptake and incorporation of 4C- L-leu-cine. (A) Control. (B) Mycelial cells were exposed to10-4 M L-leucine for 10 min at 37 C and then used foruptake and incorporation studies. Symbols: 0, totaluptake: A, incorporation; 0, metabolic pool as deter-mined experimentally. Experimental conditions are de-scribed in Fig. 1.
of similar experiments conducted with mycelialcells. The time course of L-leucine uptake in-creased from 5 to 7 min (non-preloaded cells) to10 to 12 min in preloaded cells. But, unlike yeastphase cells, the total accumulation within 10 minof incubation was about 86% that of untreatedcells. The total incorporation of the radioactivityinto acid-insoluble fraction was only 70% of thatof untreated cells.Uptake and incorporation of L-Ieucine during
transition of yeast phase to mycelial phase. Sincethe yeast phase of H. capsulatum can easily beconverted to the mycelial phase, and since thetwo morphological forms showed a significantdifference in the initial rate of L-leucine uptake,it was of interest to determine whether the in-crease in the rate of the amino acid uptake oc-
curred at a particular time during the morpholog-ical transition or whether it was a slow contin-uous process accompanying the germination ofblastospores. As the yeast cells started to germi-nate the rate of L-leucine uptake increased at a
faster rate than its incorporation. The increasedrate of L-leucine uptake resulted in a larger poolof the amino acid in the mycelial phase cells thanin the yeast phase cells. These differences becamemore obvious when total uptake, cold acid-sol-uble and the acid-insoluble fractions calculatedfor the first minute of incubation were plottedagainst the time at which the cells were harvested(Fig. 10). Microscopic examination of wetmounts of the cells revealed that at 0 hr all of thecells were in the yeast form. By 18 hr of incuba-tion most of the cells (about 60%) had short pre-
mordia of germ tubes. After 26 hr of incubationmore than 70% of the yeast cells had short germtubes, and by 40 hr the cells were completely inmycelial form.
DISCUSSIONThe evidence obtained in this investigation
clearly demonstrates that leucine uptake by thetwo morphological forms (yeast and mycelium)of H. capsulatum occurred at different rates.Contrary to expectation, the mycelial phasewhich has less surface area than the yeast phaseon an equivalent dry-weight basis accumulatedthe amino acid about 5 to 10 times faster than theyeast phase. This difference in the rate of L-leu-cine uptake could be attributed to one or more ofthe following factors: (i) a higher rate of proteinsynthesis in mycelial phase than in yeast phasecells, (ii) a mycelial phase which was metaboli-cally more active than the yeast phase, (iii) a dif-ference in the amino acid transport system ofyeast and mycelial phase cells, and/or (iv) theexistence of more transport sites per unit area ofcell membrane in mycelial phase cells than inyeast phase cells. The first possibility was ruledout on the basis of the results illustrated by Fig. I
2.4
2.0
160
°1.2
z
X 0.8
0
=. 0.410
0 1A(AI .AOI*v ICv J v 14v
HOURS OF INCUB ATION AT 24CFIG. 10. Initial rate of uptake and incorporation of
L-leucine during transition from yeast phase to mycelialphase of Histoplasma capsulatum. The initial rateswere computed from the observations made at 5- and10-min intervals. Symbols: *, total uptake; A, incorpo-ration; 0, metabolic pool.
698 J. BACTERIOL.
tn (A) (B)
LI.
O 6
z0
4I0
o 2CI~
0 20 40 60 0
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.

L-LEUCINE IN H. CAPSULATUM
and 2, in which the rates of 14C-L-leucine incorpo-ration of yeast and mycelial forms are almostequal as opposed to the rates of total uptake. Thesecond possibility was eliminated on the basis ofrespirometric studies in our laboratory whichrevealed that 02 consumption of mycelial phasecells was not greater than yeast phase cells (C.M. Yen and D. H. Howard, unpublished data).Furthermore, Nickerson and Edwards (21) re-ported that oxygen consumption by the yeastcells of Blastomyces dermatitidis was five- to six-fold higher than by mycelial cells. The third pos-sibility was not supported by the kinetics of theamino acid uptake. Therefore, the fourth factor,i.e., more transport sites per unit area of themembrane, seems to be the most likely possibilityfor an increased rate of L-leucine uptake by my-celial phase.The complex nature of the L-leucine uptake
process in both morphological forms of H. capsu-latum is indicated by the break in saturationcurves at high amino acid concentrations. It wasreported that nonlinear, double-reciprocal plotsare characteristic of many amino acid transportsystems in Pseudomonas aeruginosa (14), andsuch nonlinearity was discovered in other bacteriaand yeasts (3, 8, 10, 11, 30). In general, thesereports point out that a particular amino acid istransported by two different carrier sites: one hasa high affinity (Kmn) and is completely specific forthe amino acid, and the other is nonspecific witha low affinity (Kin). Thus, the biphasic nature ofthe double-reciprocal plots (Fig. 3, 4) of L-leucineuptake by the two morphological forms of H.capsulatum indicated that L-leucine is transportedby two different processes, and since the curve,after it breaks, almost passes through the origin,the second system would be unsaturable withinphysiological range of the amino acid concentra-tion.The properties of the transport system func-
tioning at low levels of L-leucine (5 x 10-5 M orless) in the two morphological forms of H. capsu-latum are those of an active transport mediatedby an enzyme-like carrier (24). At these externalsubstrate concentrations, the system was pH andtemperature dependent. The process was sensitiveto common metabolic inhibitors, required energy(22), and was specified for aliphatic neutralamino acids. The K1 values for these amino acidswere almost identical to the range of Km valuesfor L-leucine, indicating that the transport systemhas an equal affinity for these amino acids. Theelectro-negative nature of the aromatic ring ofthe aromatic neutral amino acids probably en-abled them to compete for L-leucine. The reasonwhy lysine and phenylalanine stimulated L-leu-cine uptake was not investigated.
The inability of basic and acidic amino acids tocompete with L-leucine for the transport site wasprobably because the side chains of these aminoacids possess a net positive and negative charge,respectively, which interfered with binding of themolecule to the transport system. That an ali-phatic side chain determined the affinity of theamino acid was further substantiated by thefinding that glycine did not inhibit L-leucine up-take. Asparagine and glutamine were onlyslightly inhibitory in mycelial phase cells but notat all in yeast phase cells. Inhibition of L-leucineuptake by a cyclic imino acid (proline) can beexplained on the basis of its structural confirma-tion in solution which has been studied byAbraham and McLauchlan (1, 2). The buckledring of proline seems to provide all the pointsnecessary for interaction with the transport site.The minimal structural requirements for a
molecule to bind to the transport site under dis-cussion were an a-amino group, an a-carboxylgroup, and an uncharged aliphatic side chain.The a-amino group could be converted to a sec-ondary a-amino (imino) group. These propertieswere the same for the transport system func-tioning in both yeast and mycelial forms of H.capsulatum.
Requirements for a-amino, a-carboxyl groupsand an uncharged aliphatic side chain for anamino acid to be transported by the permease-like system strongly suggest a three-point attach-ment to the transport site. Such a three-point at-tachment system for tryptophan was described byWiley and Matchett (28) for N. crassa.Kay and Gronlund (15) demonstrated that in P.
aeruginosa the size of the intracellular prolinepool and its formation depend on the capacity ofthe cells to transport the amino acid and theirability to utilize the accumulated amino acid forprotein synthesis. On the contrary, in yeast phaseof H. capsulatum it appeared that the metabolicpool was of limited capacity and that the initialrate of L-leucine uptake was independent of theintracellular pool size, but that the total uptakeof the amino acid was a function of intracellularpool size. A slightly slower rate of incorporationof "4C-L-leucine was observed when the cells werepreloaded with 12C-L-leucine. However, whetherthe intracellular L-leucine pool consisted of twocomponents (an expandable pool and an internalpool) as described by Kempner and Cowie (16) inCandida utilis is not firmly established by ourdata. The slower rate of incorporation of radioac-tivity by L-leucine-preloaded yeast cells was be-cause of dilution of the radioactivity in the in-ternal pool by the previously accumulated aminoacid (Fig. 8). The mycelial phase cells of H. cap-sulatum appeared to have an intracellular pool
VOL. lOS, 1971 699
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.

GUPTA AND HOWARD
which was about two times larger for L-leucinethan that of yeast phase cells.
ACKNOWLEDGMENT
This work was supported by Public Health Service grantAl 07461-03 from the National Institute of Allergy and Infec-tious Diseases.
LITERATURE CITED
1. Abraham, R. J., and K. A. McLaughlan. 1962. The protonresonance and confirmations of the prolines. 1. Theproton resonance spectra of trans hydroxy-L-proline andcis (allo) hydroxy-L-proline. Mol. Phys. 5:195-203.
2. Abraham, R. J., and K. A. McLauchlan. 1962. The protonresonance spectra and conformations of the prolines.Part II. The conformations of trans hydroxy-L-prolineand cis (allo) hydroxy-L-proline in solution. Mol. Phys.5:513-523.
3. Ames, G. F. 1964. Uptake of amino acids by Salmonellatyphimurium. Arch. Biochem. Biophys. 106:1-18.
4. Bray, G. A. 1960. A simple efficient liquid scintillator forcounting aqueous solutions in a liquid scintillationcounter. Anal. Biochem. 1:219-285.
5. Brown, 0. R., and S. Reda. 1967. Enzyme and permea-bility changes during morphogenesis of Nocardia coral-lina. J. Gen. Microbiol. 47:169-194.
6. Dancis, J., J. Hutzler, and M. Levitz. 1963. Thin-layerchromatography and spectrophotometry of a-ketoacidhydrazones. Biochim. Biophys. Acta 78:85-100.
7. Emmons, C. W., C. H. Binford, and J. P. Utz. 1963. Med-ical mycology. Lea & Febiger, Philadelphia.
8. Gale, E. F., and J. P. Folkes. 1967. The effect of lipids onthe accumulation of certain amino acids by Staphylo-coccus aureus. Biochim. Biophys. Acta 144:461466.
9. Gilbert, B. E., and D. H. Howard. 1970. Uptake of cystineby the yeast phase of Histoplasma capsulatum. Infec.Immun. 2:139-144.
10. Grenson, M. 1966. Multiplicity of the amino acid per-meases in Saccharomyces cerevisiae. 11. Evidence for a
specific lysine-transporting system. Biochim. Biophys.Actc 127:339-346.
11. Halpern, Y. S., and A. Evan-Shoshan. 1967. Properties ofthe glutamate transport system in Escherichia coli. J.Bacteriol. 93:1009-1016.
12. Hanks, J. H., and J. H. Wallace. 1958. Determination ofcell viability. Proc. Soc. Exp. Biol. Med. 98:188-192.
13. Howard, D. H. 1962. The morphogenesis of the parasiticforms of dimorphic fungi. Mycopathol. Mycol. Appl. 10:303-3 16.
14. Kay, W. W., and A. F. Gronlund. 1969. Proline transport
by Pseudomonas aeruginosa. Biochim. Biophys. Acta193:444-455.
15. Kay, W. W., and A. F. Gronlund. 1969. Amino acid poolformation in Pseudomonas aeruginosa. J. Bacteriol. 97:282-291.
16. Kempner, E. S., and D. B. Cowie. 1960. Metabolic poolsand the utilization of amino acid analogues for protein
synthesis. Biochim. Biophys. Acta 42:401 -408.17. Krulwich, T. A., and J. C. Ensign. 1969. Alteration of glu-
cose metabolism of Arthrobacter crystallopoietes bycompounds which induce sphere to rod morphogenesis. J.Bacteriol. 97:526-534.
18. Lineweaver, H., and D. Burk. 1934. The determination ofenzyme dissociation constants. J. Amer. Chem. Soc. 56:658-666.
19. Lubin, M. 1962. Some properties and potential uses of bac-terial mutants defective in amino acid transport, p. 610611. In J. T. Holden (ed.), Amino acid pools. ElsevierPublishing Co., New York.
20. Morrison, R. T., and R. N. Boyd. 1962. Organic chemistry,p. 837-839. Allyn and Bacon, Inc., Boston, Mass.
21. Nickerson, W. J., and G. A. Edwards. 1949. Studies on thephysiological bases of morphogenesis in fungi. 1. Therespiratory metabolism of dimorphic pathogenic fungi. J.Gen. Physiol. 33:41-55.
22. Palvasova, E., and F. M. Harold. 1969. Energy coupling in
the transport of ,-galactosides by Escherichia coli: effectof proton conductors. J. Bacteriol. 98:198 -204.
23. Pine, L. 1962. Nutritional determinants of fungus mor-
phology. In G. Dalldorf (ed.), Fungus and fungus dis-eases. Charles C Thomas, Publisher, Springfield, 111.
24. Rickenberg, H. V., G. N. Cohen, G. Buttin, and J. Monod.1956. La galactoside-permease d'Escherichia coli. Ann.Inst. Pasteur 91:829-857.
25. Romano, A. H. 1966. Dimorphism, p. 181-207. In G. C.Ainsworth and A. S. Sussman (ed.), The fungi, vol. 2.Academic Press Inc., New York.
26. Salvin, S. B. 1950. Growth of yeast-like phase of Histo-plasma capsulatum in a fluid medium. J. Bacteriol. 59:312-313.
27. Slayman, C. W., and E. L. Tatum. 1965. Potassium trans-port in Neurospora. 111. Isolation of a transport mutant.Biochim. Biophys. Acta 109:184-193.
28. Wiley, W. R., and W. H. Matchett. 1966. Tryptophantransport in Neurospora crassa. 1. Specificity and ki-netics. J. Bacteriol. 92:1698-1705.
29. Williamson, D. H., and A. W. Scopes. 1960. The behaviorof nucleic acids in synchronously dividing cultures ofSaccharomyces cerevisiae. Exp. Cell Res. 20:338 -349.
30. Yabu, K. 1970. Amino acid transport in Mycobacteriumsmegmatis. J. Bacteriol. 102:6-13.
700 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Janu
ary
2022
by
212.
152.
35.2
36.