contribution of postprandial lipemia to the dietary fat–mediated
TRANSCRIPT

Contribution of postprandial lipemia to the dietary fat–mediatedchanges in endogenous lipoprotein-cholesterol concentrations inhumans1–3
Byung-Hong Chung, BH Simon Cho, Ping Liang, Steve Doran, Laura Osterlund, Robert A Oster, Betty Darnell, andFrank Franklin
ABSTRACTBackground: Dietary fats alter LDL and HDL cholesterol whileserving as precursors of postprandial triacylglycerol-rich lipopro-teins (TRLs).Objective: We hypothesized that the saturated fatty acid (SFA)–mediated increase and the polyunsaturated fatty acid (PUFA)–me-diated decrease in endogenous lipoprotein cholesterol are promotedby postprandial TRLs.Design: We performed a 16-d crossover diet study to examine theeffect of PUFA-rich [ratio of PUFAs to SFAs (P:S) � 2.0] andSFA-rich (P:S � 0.25) diets on fasting and postprandial plasma lipidand lipoprotein-cholesterol concentrations in 16 normolipidemicsubjects.Results: Fasting plasma cholesterol decreased significantly after aPUFA-rich diet because of a decrease in LDL (�12.3%; P � 0.05)and HDL (�3.8%; NS), but did not change after an SFA-rich diet.The appearance of postprandial TRLs in plasma at 4 h was linked toa significant lowering of both LDL (�7.4%) and HDL (�4.8%) aftera PUFA-rich diet; no such effect was observed after the SFA-richdiet. At 7 h, LDL and HDL cholesterol returned to near fastingconcentrations without postprandial TRL accumulation after aPUFA-rich diet but with a significant postprandial TRL accumula-tion after an SFA-rich diet. Thus, the in vivo postprandial clearanceof cholesterol in LDL�HDL was greater after a PUFA-rich diet thanafter an SFA-rich diet. The appearance of postprandial TRLs inplasma increased the cholesteryl ester transfer protein–mediatedtransfer of cholesteryl ester from LDL�HDL to TRLs in vitro with-out a significant influence from dietary fat.Conclusion: Dietary fat–mediated alterations in the rate of hepaticremoval of postprandial TRLs, which carry cholesterol acceptedfrom LDL�HDL via cholesteryl ester transfer protein in vivo, maycontribute to the dietary fat–mediated change in endogenous li-poprotein cholesterol. Am J Clin Nutr 2004;80:1145–58.
KEY WORDS Chylomicrons, VLDL, LDL, HDL,lecithin:cholesterol acyltransferase, cholesteryl ester transfer pro-teins, postprandial lipemia, reverse cholesterol transport, polyunsat-urated fat, saturated fat
INTRODUCTION
It is well established that dietary fat composition affects theconcentration and distribution of cholesterol among lipoproteins
in fasting plasma and the risk of developing cardiovascular dis-ease (CVD) (1–3). A diet rich in polyunsaturated fatty acids(PUFA) lowers fasting plasma cholesterol by decreasing LDLand HDL cholesterol and thereby reduces CVD risk, whereas adiet rich in saturated fatty acids (SFA) has the opposite effect(1–3). There is a consensus that the SFA- and PUFA-mediatedchanges in fasting cholesterol concentrations are due to a changein the concentration and activities of hepatic LDL receptors (4,5). However, the SFA- and PUFA-mediated simultaneouschanges in LDL and HDL cholesterol cannot be fully explainedby changes in the concentration and activities of the LDL recep-tors alone because those changes may not affect and so do notexplain changes in HDL cholesterol. Most long-term studies ofdietary fat–induced changes in plasma cholesterol have focusedon endogenous lipoprotein-cholesterol concentrations in fastingplasma even though dietary fats are direct precursors of post-prandial TRL but do not directly contribute to the formation ofendogenous lipoproteins. Postprandial lipemia is due to an in-crease in both intestine-derived apolipoprotein (apo) B-48–con-taining chylomicrons and liver-derived apo B-100–containingVLDL (6, 7). SFA-induced postprandial lipemia is more pro-nounced and of a longer duration than is PUFA-induced post-prandial lipemia (8–11).
Recent studies indicate that plasma LDL-cholesterol concen-trations negatively correlate with the removal rate from plasma ofchylomicron-like emulsion particles (12) and that the pharma-cologic reduction of plasma LDL cholesterol is associated withan increase in the clearance of postprandial triacylglycerol-richlipoproteins (TRLs) or chylomicron-like emulsion particles inhumans (13–16). Conversely, an association between elevated
1 From the Atherosclerosis Research Unit (B-HC, PL, SD, and LO), theGeneral Clinical Research Center (RAS and BD), and the Departments ofMedicine (B-HC, PL, SD, and LO), Pediatrics (FF), and Nutrition (FF),Medical School, University of Alabama at Birmingham, and the Moore HeartResearch Foundation, University of Illinois, Champaign, IL (BHSC).
2 Supported by the US Public Health Service (grant NIH HL 60936) and bythe General Clinical Research Center, University of Alabama at Birmingham(grant RR0032).
3 Address reprint requests to B-H Chung, Department of Nutrition Sci-ence, Webb Building 248, University of Alabama, Birmingham, AL 35294.E-mail: [email protected].
Received September 18, 2003.Accepted for publication May 7, 2004.
1145Am J Clin Nutr 2004;80:1145–58. Printed in USA. © 2004 American Society for Clinical Nutrition
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

LDL-cholesterol concentrations and delayed chylomicron rem-nant clearance has been observed in the elderly (17). The abovein vivo observations suggest that LDL-cholesterol concentra-tions in plasma can be altered by the change in the clearance rateof postprandial TRLs. However, whether the PUFA- and SFA-mediated changes in LDL- and HDL-cholesterol concentrationsin fasting plasma are linked to alterations in the clearance rate ofpostprandial TRLs has not been fully explored. Many studieshave shown that the induction of postprandial lipemia is accom-panied by a transient increase in plasma lecithin:cholesterol acyl-transferase (LCAT) and cholesteryl ester transfer protein(CETP) activities (18–20), a net decrease in cholesteryl ester(CE) concentrations in both LDL and HDL as well as in plasmaas a whole (11, 21, 22-24) and a shift in the distribution of CEfrom LDL and HDL to chylomicron (25). In plasma, LCAT andCETP promote the esterification of unesterified cholesterol (UC)and the reciprocal exchange of CE and triacylglycerol betweenthe cholesterol-rich lipoproteins (LDL and HDL) and TRLs(VLDL�chylomicron) (20, 26). The possibility then exists, buthas not been fully explored, that the postprandial enhancement ofLCAT and CETP activities could possibly lower the concentra-tions of LDL and HDL cholesterol by transferring CE from LDLand HDL to postprandial TRLs and by subsequently clearingpostprandial TRLs.
We previously provided evidence that postprandial TRLs actas potent acceptors both of cholesterol from endogenous LDLand HDL and of that released from cell membranes via LCATand CETP activities in vitro (27). In another recent study (28), weshowed that the alcohol-mediated increase in postprandial TRLsfurther enhanced the postprandial reduction of LDL and HDLcholesterol in vivo and the acceptance of cholesterol by TRLs fromendogenous lipoproteins and cell membranes via LCAT and CETPin vitro. Given that postprandial TRLs are cleared from plasmamostly by hepatic uptake (29), our data suggest that postprandialTRLs can serve as vehicles for transporting cholesterol from endog-enous LDL and HDL and cell membranes to the liver via LCAT andCETP. Because dietary fat can alter the concentration and clearancerate of postprandial TRLs in vivo (8–11) and may even affect thepotencies of postprandial TRLs to serve as acceptors of LCAT andCETP reaction products, it could potentially alter the concentrationof endogenous lipoprotein cholesterol and the rate of reverse cho-lesterol transport in vivo.
To determine whether dietary fat alters endogenous lipopro-tein cholesterol and CVD risk by affecting the potency and rateof postprandial TRLs to carry cholesterol accepted from endog-enous LDL and HDL and from cell membranes via LCAT andCETP to the liver for its removal, we examined the acute andchronic effects of the consumption of a PUFA- or SFA-rich dietand meal on fasting plasma lipoprotein-cholesterol concentra-tions, on the extent of postprandial change in the levels anddistribution of cholesterol in lipoproteins in vivo, and on thepotency of TRLs in fasting and postprandial plasma to serve asacceptors of the LCAT and CETP reaction product in vitro.
SUBJECTS AND METHODS
Subjects
Healthy normolipidemic men and postmenopausal womenwere recruited. Interested volunteers underwent a screening ex-amination at the General Clinical Research Center (GCRC) of the
University of Alabama at Birmingham Medical Center. The ex-amination included a brief medical history and physical exami-nation as well as measurement of body weight, height, and fast-ing plasma lipid and lipoprotein-cholesterol concentrations.Subjects with a fasting triacylglycerol concentration above the75th percentile, plasma cholesterol above the 95th percentile, orHDL cholesterol below the 10th percentile for the respective agegroups were excluded from the study. Subjects taking medica-tion and subjects with unusual dietary habits (vegetarians, heavydrinkers, etc) were also excluded. Eight men (7 white and 1black) aged 33–49 y (35.3 � 4.5 y) and 8 postmenopausalwomen (5 white and 3 black) aged 45–62 y (51.9 � 6.6 y)participated in this study. The mean body mass indexes (inkg/m2) of men and women were 25.3 � 4.1 and 29.6 � 4.5,respectively. The research protocol for using human subjects inthis study was approved by the University of Alabama Institu-tional Review Board.
Diet and blood collection
A randomized crossover diet study was performed to deter-mine the chronic and acute effects of a normal PUFA-rich [ratioof PUFAs to SFAs (P:S) � 2.0] and SFA-rich (P:S � 0.25) dieton the concentrations of lipid and lipoprotein cholesterol in fast-ing and postprandial plasma. Subjects adopted each of the 2 dietsfor a period of 20 d, with a 3–4 wk return to their habitual adlibitum diet in between. Both diets provided 15% of energy fromproteins, 50% of energy from carbohydrates, and 35% of energyfrom fat and contained 175 mg cholesterol/1000 kcal. SFAs,monounsaturated fatty acids, and PUFAs provided 7.5%, 12%,and 15.5% of total energy in the PUFA-rich diet, respectively,and 18.8%, 11.5%, and 4.7% of total energy in the SFA-rich diet,respectively. All meals were prepared by the University of Ala-bama GCRC research kitchen and were provided daily to thesubjects. To examine the postprandial lipemic response, a fat-loading test was performed on day 16 of each diet by offering abreakfast that had the same P:S as did the chronic experimentaldiet. Another fat-loading study was performed on the last day(day 20) of each diet to examine the acute effect of changing ameal’s fat composition on the postprandial lipemic response. Inthe studies on day 20, the subjects consuming a chronic SFA-richdiet were given a PUFA-rich breakfast, whereas subjects con-suming a chronic PUFA-rich diet were given a SFA-rich break-fast.
To maximize the postprandial lipemic response, the challengemeal contained a slightly higher concentration of fat (40% ofenergy from fat) than did the background diet (35% of energyfrom fat). Briefly, on the evening before the fat-loading test (day16 and day 20), the subjects were admitted to the GCRC, wherea dinner meal was prepared for them. After they fasted overnightat the GCRC, the subjects were offered a challenge breakfastmeal. Because the subjects would not eat anything else for �7 h,the challenge meal consisted of whole foods that provided 50%of the total daily calories. Fasting and postprandial blood sampleswere collected from the subjects immediately before the meal (40mL) and 4 h (80 mL) and 7 h (40 mL) after the meal.
Treatment of blood and plasma samples
Blood samples were collected in tubes containing EDTA(0.1%) and immediately placed in an ice bath; plasma was sep-arated from red blood cells by centrifuging the blood at 2620 �
1146 CHUNG ET AL
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

g (3500 rpm) for 25 min at 4 °C in a low-speed centrifuge (Beck-man Co, Fullerton, CA). Plasma was kept in an ice bath untilanalyzed, and one aliquot of plasma was placed in a 37 °C waterbath (Forma Scientific Co, Division of Mallinckrodt, Marietta,OH) for 4–16 h to allow the lipoproteins in the plasma to interactwith the endogenous LCAT and CETP. A portion of each plasmasample was stored in several aliquots at �70 °C.
Measurement of plasma lipid, lipoprotein-cholesterol, andtriacylglycerol concentrations
Concentrations of plasma total cholesterol, UC, and triacyl-glycerol were measured by using enzymatic cholesterol, triacyl-glycerol, and UC assay reagents purchased from Sigma Co (StLouis), Boehringer Mannheim Co (Indianapolis), and Waco Di-agnostic Co (Richmond, VA). The distribution of cholesterol andtriacylglycerol in VLDL, LDL, and HDL density fractions ineach fresh fasting plasma sample obtained on day 1 and in fastingand 4-h and 7-h plasma samples obtained on day 16 and day 20was determined by the modified lipoprotein cholesterol auto-profiler method developed in this laboratory (30, 31). Thismethod involves 1) short (150 min) single-spin density-gradientultracentrifugal separation of the major lipoprotein fraction inplasma in a swing-out rotor (model SW 50.1; Beckman Co), 2)continuous-flow online mixing of effluent from density-gradienttubes with enzymatic cholesterol or triacylglycerol reagent, 3)online incubation and online measurement of absorbance produced,and 4) computerized calculation of lipoprotein-cholesterol and tri-acylglycerol concentrations (31). To determine the overall dietaryfat–mediated changes in the density spectrum of lipoprotein parti-cles as well as in their concentrations in fasting plasma, an aliquot(0.5 mL) of each frozen fasting plasma sample, collected from allstudy subjects (n � 16) on day 1 and day 16, was taken and pooled.The lipoprotein-cholesterol profiles of pooled plasma were thendetermined by the lipoprotein-cholesterol auto-profiler method. Weshowed previously that a single freezing and thawing of fastingplasma produces no appreciable change in its lipoprotein-cholesterol profile (30).
Measurement of TRL apo B-100 and apo B-48
A modified version of the sodium dodecyl sulfate (SDS) gelelectrophoresis method described by Kotite et al (32) was used toexamine the postprandial changes in concentrations of apo B-100and apo B-48 associated with TRLs. Briefly, 4 mL fasting andpostprandial plasma containing proteinase inhibitor mixtures(Roche Co, Indianapolis) were placed in the bottom of a Beck-man SW 50.1 ultracentrifuge tube containing 1 mL bufferedsaline (0.01 mol tris/L, 1.15 mol NaCl/L, ph 7.4) with the use ofan underlayering method. After the ultracentrifugation of plasmaat 269 971 � g (49 000 rpm) for 16 h at 4 °C, TRLs floated to thetop of the tubes (in a total volume of 0.8 mL) was quantitativelyremoved. Once the triacylglycerol and cholesterol concentrationof fasting VLDLs had been determined, a fasting VLDL solutioncontaining 10 �g triacylglycerol and 4-h and 7-h TRL solutionswith the same volume as the fasting VLDL solution were placedin a conical centrifuge tube containing transferrin as an internalstandard. The rotary vacuum evaporator rapidly dried the sam-ples, and the dried TRLs adhering to the tubes were exposedbriefly (5 min) to cold hexane:ethanol (3:1, by vol) mixtureswithout agitation of the tubes. After gentle aspiration of thesolvent mixtures, the partially delipidated TRLs that had adhered
to the walls of the tubes were then dissolved in an SDS-treatmentbuffer and loaded on to 4–20% SDS gradient gels (InvitrogenCo, Carlsbad, CA) to separate apo B-100, apo B-48, and otherapolipoproteins on TRLs. Apo B-100 prepared from purifiedLDL was used as a standard. After the gels were stained with gelcode blue stain (Pierce Co, Rockford, IL), the concentrations offasting and postprandial TRL apo B-100 and apo B-48 in the gelwere then determined based on an apo B-100 standard by usinga computer-driven UVP gel imaging system (Quest ScientificCo, Cumming, GA). Postprandial changes in the concentrationsof apo B-100 and apo B-48 associated with TRLs were examinedby determining the percentage of postprandial plasma TRL apoB-48 and apo B-100 and fasting VLDL apo B-48 relative tofasting plasma VLDL apo B-100 and by performing the statisti-cal analysis on the extent (percentage) of postprandial increasesin concentrations of apo B-48 and apo B-100 relative to those infasting plasma.
Measurement of the concentration and activity ofplasma CETP
The CETP concentrations in fasting and postprandial plasmawere determined by using a CETP enzyme-linked immunosor-bent assay kit purchased from Waco Diagnostic Co. The plasmaCETP activities were assayed by measuring the rate and extent oftransfer of [3H]CE from LDL�HDL to TRLs in fasting andpostprandial plasma in vitro. Briefly, [3H]CE-labeledLDL�HDL was prepared by using a TRL-free plasma fraction(density � 1.006 g/mL) as described by Thomas and Rudel (33).The distribution pattern of [3H]CE for LDL and HDL in theprepared samples mirrored that of LDL and HDL cholesterol.After isolation of [3H]CE-labeled LDL and HDL by one-stepultracentrifugation (269 971 � g, 150 min, 4 °C), equal (trace)amounts of [3H]CE-labeled LDL�HDL were added to freshfasting and postprandial plasma, and the mixtures were incu-bated for 3–9 h at 37 °C. After incubation, TRLs were separatedquantitatively from LDL and HDL by short (150 min), single-spin, density-gradient ultracentrifugation in a SW 50.1 swing-out rotor (31). The level of [3H]CE radioactivity transferred fromLDL�HDL into TRLs was measured by counting the radioac-tivity associated with the TRL fraction. The increases in choles-terol mass in TRLs after in vitro incubation of plasma for 16 h at37 °C was determined by analyzing the change in the lipoprotein-cholesterol profiles of plasma (31). To determine whether aCETP-mediated reciprocal transfer of CE and triacylglyceroloccurs between TRLs and the LDL�HDL fraction during post-prandial lipemia in vivo, we measured the changes in the fattyacyl chain composition of triacylglycerol associated with TRLsand the LDL�HDL fraction after quantitatively separating themfrom fresh and LCAT- and CETP-reacted fasting and postpran-dial plasma using the gas chromatographic method as previouslydescribed (34).
Statistical analysis
Quantitative variables were expressed as means � SDs. Sta-tistical tests were applied to compare the concentration of plasmalipids, lipoprotein cholesterol, and TRL apo B-100 and apo B-48in fasting plasma and in postprandial plasma obtained after sub-jects consumed an SFA- or PUFA-rich diet or a meal or after an
POSTPRANDIAL LIPEMIA AND FASTING PLASMA CHOLESTEROL 1147
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021
in vitro reaction of LCAT and CETP in plasma. These compar-isons were performed by using mixed-models repeated-measures analysis of variance, assuming an unstructured covari-ance matrix. Tukey’s multiple comparisons test was then used todetermine which specific pairs of means were significantly dif-ferent (P � 0.05). All statistical tests were two-sided and wereperformed at a 5% significance level (ie, � � 0.05). All statisticalanalyses were performed with the use of SAS software (version9.0; SAS Institute Inc, Cary, NC).
RESULTS
Chronic effect of a PUFA-rich and SFA-rich diet onchanges in plasma lipid and lipoprotein-cholesterolconcentrations and lipoprotein-cholesterol profiles offasting plasma
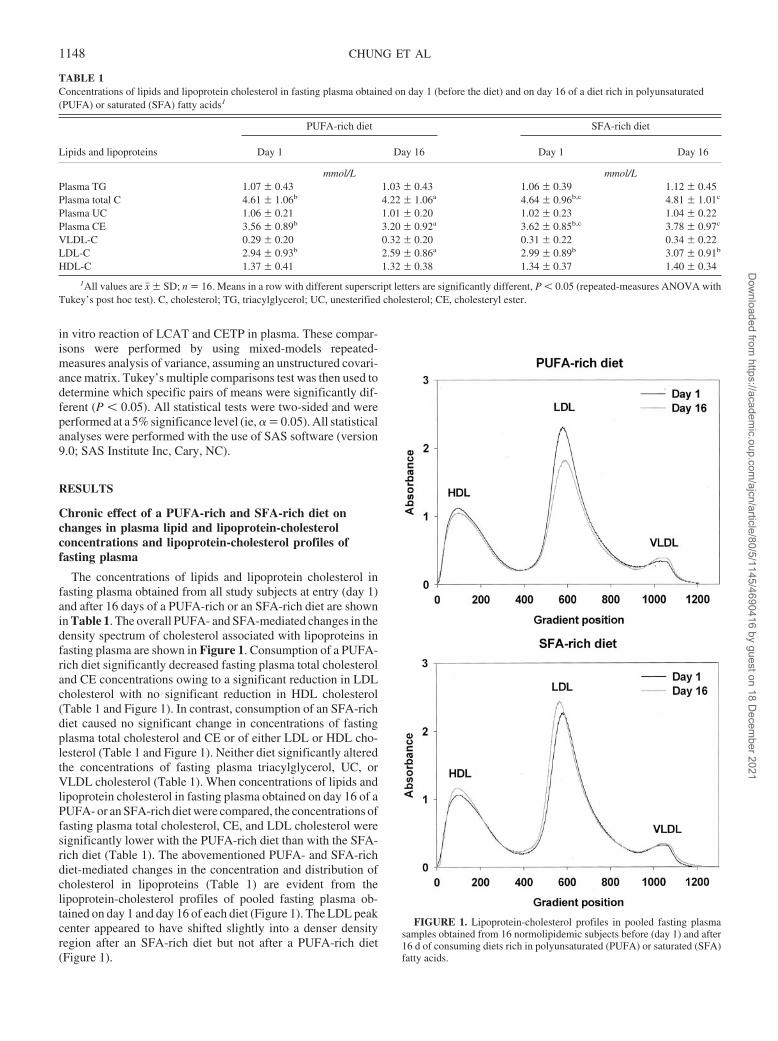
The concentrations of lipids and lipoprotein cholesterol infasting plasma obtained from all study subjects at entry (day 1)and after 16 days of a PUFA-rich or an SFA-rich diet are shownin Table 1. The overall PUFA- and SFA-mediated changes in thedensity spectrum of cholesterol associated with lipoproteins infasting plasma are shown in Figure 1. Consumption of a PUFA-rich diet significantly decreased fasting plasma total cholesteroland CE concentrations owing to a significant reduction in LDLcholesterol with no significant reduction in HDL cholesterol(Table 1 and Figure 1). In contrast, consumption of an SFA-richdiet caused no significant change in concentrations of fastingplasma total cholesterol and CE or of either LDL or HDL cho-lesterol (Table 1 and Figure 1). Neither diet significantly alteredthe concentrations of fasting plasma triacylglycerol, UC, orVLDL cholesterol (Table 1). When concentrations of lipids andlipoprotein cholesterol in fasting plasma obtained on day 16 of aPUFA- or an SFA-rich diet were compared, the concentrations offasting plasma total cholesterol, CE, and LDL cholesterol weresignificantly lower with the PUFA-rich diet than with the SFA-rich diet (Table 1). The abovementioned PUFA- and SFA-richdiet-mediated changes in the concentration and distribution ofcholesterol in lipoproteins (Table 1) are evident from thelipoprotein-cholesterol profiles of pooled fasting plasma ob-tained on day 1 and day 16 of each diet (Figure 1). The LDL peakcenter appeared to have shifted slightly into a denser densityregion after an SFA-rich diet but not after a PUFA-rich diet(Figure 1).
TABLE 1Concentrations of lipids and lipoprotein cholesterol in fasting plasma obtained on day 1 (before the diet) and on day 16 of a diet rich in polyunsaturated(PUFA) or saturated (SFA) fatty acids1
Lipids and lipoproteins
PUFA-rich diet SFA-rich diet
Day 1 Day 16 Day 1 Day 16
mmol/L mmol/LPlasma TG 1.07 � 0.43 1.03 � 0.43 1.06 � 0.39 1.12 � 0.45Plasma total C 4.61 � 1.06b 4.22 � 1.06a 4.64 � 0.96b,c 4.81 � 1.01c
Plasma UC 1.06 � 0.21 1.01 � 0.20 1.02 � 0.23 1.04 � 0.22Plasma CE 3.56 � 0.89b 3.20 � 0.92a 3.62 � 0.85b,c 3.78 � 0.97c
VLDL-C 0.29 � 0.20 0.32 � 0.20 0.31 � 0.22 0.34 � 0.22LDL-C 2.94 � 0.93b 2.59 � 0.86a 2.99 � 0.89b 3.07 � 0.91b
HDL-C 1.37 � 0.41 1.32 � 0.38 1.34 � 0.37 1.40 � 0.34
1All values are x� � SD; n � 16. Means in a row with different superscript letters are significantly different, P � 0.05 (repeated-measures ANOVA withTukey’s post hoc test). C, cholesterol; TG, triacylglycerol; UC, unesterified cholesterol; CE, cholesteryl ester.
FIGURE 1. Lipoprotein-cholesterol profiles in pooled fasting plasmasamples obtained from 16 normolipidemic subjects before (day 1) and after16 d of consuming diets rich in polyunsaturated (PUFA) or saturated (SFA)fatty acids.
1148 CHUNG ET AL
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

Chronic effect of PUFA- and SFA-rich diets on postprandialchanges in concentrations of plasma lipids, lipoprotein-cholesterol, TRL apo B-100 and apo B-48, and lipoprotein-cholesterol and triacylglycerol profiles
Postprandial change in plasma lipid and lipoprotein-cholesterolconcentrations, lipoprotein-triacylglycerol profiles, compositionand concentration of VLDL apo B, and lipoprotein-cholesterol pro-files of fasting plasma after 16 d of either a PUFA- or an SFA-richdiet are shown in Table 2, Figure 2, and Figure 3.
Postprandial changes in fasting plasma triacylglycerol andTRL apo B-100 and apo B-48 concentrations
With both diets, fasting plasma triacylglycerol concentrationsincreased significantly at 4 h (Table 2). At 7 h, the elevatedtriacylglycerol concentrations decreased to fasting concentrationwith the PUFA-rich diet but only decreased partially with theSFA-rich diet (Table 2). The postprandial changes in plasmalipoprotein-triacylglycerol profiles seen in a typical subject aftera PUFA-rich diet and an SFA-rich diet (Figure 2, A) indicate thatpostprandial increases and decreases in plasma triacylglycerolare primarily due to changes in concentrations of the triacylg-lycerols associated with the TRL (VLDL density) fraction. Sub-stantial accumulation of postprandial plasma triacylglycerol inTRLs 7 h after consumption of an SFA-rich but not after con-sumption of a PUFA-rich diet was evident from postprandialchanges in the triacylglycerol profiles (Figure 2, A). Separationand quantification of TRL apo B-100 and apo B-48 with SDSgradient gels showed that the postprandial increase in TRL-triacylglycerol seen at 4 h with both diets was due to a significantincrease in the concentration of both apo B-100–containingVLDLs and apo B-48–containing chylomicrons (Figure 2, Aand B). We detected a small variable amount of apo B-48 in mostfasting VLDLs obtained from our normolipidemic study subjects(Figure 2, B). The mean concentration of TRL apo B-48 in fasting
plasma after an SFA-rich diet (6.2% of fasting VDL apo B-100)was not different from that after a PUFA-rich diet (4.7% offasting VLDL apo B-100) (Figure 2, B).
A recent study (35) showed that normolipidemic subjects con-tained widely variable amounts of apo B-48–containing lipopro-tein particles in their fasting plasma. The significant postprandialincreases in TRL apo B-100 (158%) or apo B-48 (261%) 4 h afterconsumption of an SFA-rich diet were not significantly differentfrom those increases noted after a PUFA-rich diet [TRL apoB-100 (147%) and TRL apo B-48 (197%)] (Figure 2, B). At 7 h,TRL apo B-48 decreased to its fasting concentration with boththe PUFA-rich diet and the SFA-rich diet (Figure 2, B). With theSFA-rich diet, the concentration of apo B-100–containing TRLsat 7 h was still significantly greater than the fasting concentration(Figure 2, B). We determined further molar concentrations offasting VLDL apo B-100 and apo B-48 on the basis of the con-centration of fasting plasma VLDL triacylglycerol and the con-tent of apo B-100 and apo B-48 per 10 �g VLDL triacylglyceroland using a molecular weight of 555 486 for apo B-100 and amolecular weight of 260 416 for apo B-48. The molar concen-trations of fasting VLDL apo B-100 and apo B-48 were 61.5 �22.7 and 6.8 � 5.35 nmol/L with the PUFA-rich diet and 63.5 �24.9 nmol/L and 8.8 � 4.3 nmol/L with the SFA-rich diet, re-spectively. The concentrations of fasting VLDL apo B-100 andapo B-48 with the PUFA-rich diet were not significantly differentfrom those with the SFA-rich diet.
Postprandial changes in cholesterol concentrations ofplasma and lipoproteins and the lipoprotein-cholesterolprofile
With both diets, fasting plasma UC concentrations increasedsignificantly at 4 h without a significant change in plasma totalcholesterol, which resulted in a net decrease in the plasma CEconcentration (Table 2). The postprandial decrease in plasma CE
TABLE 2Plasma lipid and lipoprotein-cholesterol concentrations in fasting, 4 h, and 7 h plasma samples after the consumption of a diet rich in eitherpolyunsaturated (PUFA) or saturated (SFA) fatty acids for 16 d (chronic effect) and on day 20 after the consumption of a breakfast meal from the oppositediet (acute effect)1
Lipids and lipoproteins
PUFA-rich diet SFA-rich diet
Fasting 4 h 7 h Fasting 4 h 7 h
Chronic effect of diets (day 16) mmol/L mmol/LPlasma TG 1.03 � 0.43a 1.64 � 0.71b,c 1.10 � 0.45a 1.12 � 0.45a 2.02 � 0.89c 1.49 � 0.52b
Plasma total C 4.22 � 1.06a 4.15 � 1.08a 4.16 � 0.99a 4.81 � 1.01b 4.85 � 1.03b 4.85 � 0.95b
Plasma UC 1.01 � 0.20a,b 1.11 � 0.28c,d,e 1.03 � 0.22a,b 1.04 � 0.22a,c 1.16 � 0.26b,c 1.10 � 0.29a,b,c
Plasma CE 3.21 � 0.92c 3.04 � 0.89a,b 3.13 � 0.85b,c 3.77 � 0.97c 3.69 � 0.98d 3.75 � 0.88d
TRL-C 0.32 � 0.20a 0.48 � 0.25c 0.34 � 0.19a,b 0.34 � 0.22a 0.54 � 0.35b 0.45 � 0.22b,c
LDL-C 2.59 � 0.86a 2.42 � 0.87b,c 2.52 � 0.82a,c 3.07 � 0.91d 2.95 � 0.81d 3.00 � 0.81d
HDL-C 1.32 � 0.38b,d 1.25 � 0.38a,c,e 1.30 � 0.37b,f 1.40 � 0.34b,d 1.36 � 0.35b,c 1.39 � 0.36a,d,f
Acute effects of a meal (day 20)2
Plasma TG 1.00 � 0.38a 1.57 � 0.53b,c,d 1.27 � 0.49e,f 1.11 � 0.42a,e 1.88 � 0.77c 1.48 � 0.50d,f
Plasma total C 4.23 � 1.04a 4.27 � 1.06a 4.29 � 1.00a 4.80 � 0.98b 4.77 � 0.93b 4.78 � 0.97b
Plasma UC 1.01 � 0.23a,b 1.10 � 0.25c,d 1.05 � 0.23a,b 1.04 � 0.21a,c,e 1.15 � 0.26b,d 1.10 � 0.28b,d,e
Plasma CE 3.22 � 0.90a 3.17 � 0.90a 3.22 � 0.89a 3.76 � 0.91b 3.62 � 0.89b 3.67 � 0.84b
TRL-C 0.31 � 0.19a 0.47 � 0.23b,c 0.40 � 0.22b,d 0.35 � 0.19a 0.52 � 0.26b 0.44 � 0.22c,d
LDL-C 2.60 � 0.91a,b 2.56 � 0.98a 2.59 � 1.01a,b 3.04 � 0.90c 2.90 � 0.79b,c 2.93 � 0.82c
HDL-C 1.31 � 0.37a,b 1.26 � 0.36a 1.29 � 0.36a,b 1.40 � 0.32b 1.37 � 0.34a,b 1.40 � 0.32b
1 All values are x� � SD; n � 16. Means in a row with different superscript letters are significantly different, P � 0.05 (repeated-measures ANOVA withTukey’s post hoc test). C, cholesterol; TG, triacylglycerol; UC, unesterified cholesterol; CE, cholesteryl ester; TRL, triacylglycerol-rich lipoproteins (VLDLin fasting plasma or VLDL and chylomicrons in postprandial plasma).
POSTPRANDIAL LIPEMIA AND FASTING PLASMA CHOLESTEROL 1149
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

concentrations at 4 h was significant with the PUFA-rich diet butnot with the SFA-rich diet (Table 2). The increased concentra-tions of plasma UC with both diets at 4 h and decreased concen-trations of plasma CE at 4 h with the PUFA-rich diet returned tonear fasting concentrations at 7 h (Table 2). The chronic effectsof a diet rich in PUFAs or SFAs on the postprandial changes inthe concentrations of cholesterol in VLDL-, LDL-, and HDL-density fractions were further examined by determining thelipoprotein-cholesterol profiles in fasting and postprandialplasma. On the basis of postprandial changes in the cholesterolprofile of fasting plasma obtained from a subject with a briskpostprandial lipemic response (Figure 3, A and B), induction ofpostprandial lipemia after both diets caused an increase in cho-lesterol concentrations in TRLs and a concomitant decrease incholesterol concentrations in LDL and HDL in 4 h plasma.
Overall, the postprandial increase in TRL cholesterol at 4 hwas statistically significant with both diets, but the postprandialdecrease in concentrations of cholesterol in LDL and HDL at 4 hwas statistically significant only with the PUFA-rich diet (Table2). The postprandial increase in cholesterol in TRLs with a con-comitant postprandial decrease in the concentration of LDL andHDL cholesterol and the lack of significant changes in plasmatotal cholesterol suggests a shift in the distribution of cholesterolfrom LDL and HDL to TRLs. We observed that the postprandialdecrease in LDL and HDL cholesterol occurs with little or nopostprandial increase in TRL cholesterol in subjects with a highHDL-cholesterol concentration (data not shown). Becauseplasma UC increased significantly at 4 h (Table 2), the postpran-dial decreases in cholesterol in LDL and HDL were observed tobe due to decreases in their CE concentration (data not shown).
FIGURE 2. A: Lipoprotein triacylglycerol profiles (top) and sodium dodecyl sulfate gradient gel electropherograms showing apolipoprotein (apo) B-100and apo B-48 concentrations in triacylglycerol-rich lipoproteins (TRLs; bottom) in fasting, 4 h, and 7 h plasma samples obtained from a subject after 16 d ofconsuming a diet rich in polyunsaturated (PUFA) or saturated (SFA) fatty acids. B: Mean (�SD) percentage of postprandial TRL apo B-100 and fasting andpostprandial TRL apo B-48 relative to fasting VLDL apo B-100 in 8 subjects after 16 d of consuming a PUFA-rich or SFA-rich diet. Bars within each panelwith different letters are significantly different, P � 0.05 (repeated-measures ANOVA with Tukey’s post hoc test).
1150 CHUNG ET AL
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

The postprandial changes in lipoprotein-cholesterol concentra-tions (Table 2) indicate further that the mean postprandial de-crease in cholesterol concentrations of the LDL�HDL fraction(0.24 mmol/L) exceeded the mean postprandial increase in cho-lesterol concentrations of TRLs (0.17 mmol/L) after a PUFA-rich diet; in contrast, the mean postprandial increase in TRLcholesterol (0.21 mmol/L) exceeded the mean postprandial de-crease in cholesterol of the LDL�HDL fraction (0.16 mmol/L)after an SFA-rich diet (Table 2). Because the extent of the post-prandial increase in TRL-cholesterol concentration at 4 h withboth diets was about equal, the PUFA-mediated net decrease andthe SFA-mediated net increase in plasma cholesterol were attrib-uted primarily to the differences in the extent of the postprandialdecrease in LDL and HDL cholesterol. At 7 h, the loweredplasma concentrations of LDL and HDL cholesterol at 4 h in-creased back to near fasting concentrations with both the PUFA-and SFA-rich diets, but the significant accumulation of postpran-dial TRLs in the plasma after the SFA-rich diet was not observedafter the PUFA-rich diet (Table 2 and Figure 3, A and B).
Acute effects of substituting a meal of the alternate dieton day 20 of the experimental diet on the postprandialchanges in plasma lipid and lipoprotein-cholesterolconcentrations and lipoprotein-cholesterol profiles
The concentrations of lipids and lipoprotein cholesterol infasting plasma obtained on day 20 of the experimental diet werenot significantly different from those obtained on day 16 (Table2; acute and chronic effects). The substitution of a PUFA-richmeal for an SFA-rich meal or vice versa did not significantly alterthe extent of postprandial increases in plasma triacylglycerol andUC concentrations in 4 h plasma (Table 2; acute effects). Theextent of postprandial plasma triacylglycerol accumulated in 7 hplasma significantly increased after the substitution of an SFA-rich meal for a PUFA-rich meal in the subjects who consumed thePUFA-rich diet but was not altered after the reverse substitution(Table 2; acute effects). The effect of substituting an SFA-rich
meal for a PUFA-rich meal or vice versa on the postprandialchanges in lipoprotein-cholesterol concentration was deter-mined further by examining the alteration in postprandialchanges in lipoprotein-cholesterol profiles (Figure 3). It is evi-dent that in a subject with a brisk postprandial lipemic response,the substitution of an SFA-rich meal for a PUFA-rich meal insubjects consuming a chronic PUFA-rich diet diminished thepostprandial lipemia-mediated reduction of LDL and HDL cho-lesterol at 4 h and increased the accumulation of postprandialTRLs at 7 h (Figure 3, A and C). It was also evident that thereverse substitution in subjects consuming a chronic SFA-richdiet only minimally altered postprandial lipoprotein-cholesterolprofiles (Figure 3, B and D). In the subjects who consumed thechronic PUFA-rich diet, the postprandial lipemia produced aftersubstitution of an SFA-rich meal for a PUFA-rich meal wasassociated with no significant reduction in plasma CE and LDLand HDL cholesterol at 4 h and a significant accumulation ofpostprandial TRL cholesterol at 7 h (Table 2; acute effects). Thisfinding is in contrast with a significant reduction in plasma CEand LDL and HDL cholesterol at 4 h and no significant accumu-lation of postprandial TRL cholesterol 7 h after a PUFA-richmeal (Table 2; chronic effects). No statistically significant dif-ference in postprandial changes in plasma lipid and lipoprotein-cholesterol concentrations were noted in subjects consuming achronic SFA-rich diet when a PUFA-rich meal was acutely given(Table 2; chronic and acute effects).
Comparison of the chronic and acute effects of PUFA-rich and SFA-rich diets on the extent of postprandialchanges in plasma lipid and lipoprotein-cholesterolconcentrations
The effects of acute and chronic SFA- and PUFA-mediatedpostprandial lipemia on the percentage change in plasma lipidsand lipoprotein cholesterol are shown in Table 3. The extent ofthe postprandial increase in plasma triacylglycerol and TRL cho-lesterol 4 h after a chronic PUFA-rich diet did not differ from that
FIGURE 3. Lipoprotein-cholesterol profiles in fasting, 4 h, and 7 h plasma samples obtained from a normolipidemic subject after 16 d of consuming a chronicdiet and a single meal rich in polyunsaturated (PUFA; A) or saturated (SFA; B) fatty acids and after 20 d of consuming a chronic diet rich in PUFAs and anacute meal rich in SFAs (C) or a chronic diet rich in SFAs and an acute meal rich in PUFAs (D). A brisk postprandial lipemic response was observed.
POSTPRANDIAL LIPEMIA AND FASTING PLASMA CHOLESTEROL 1151
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

after a chronic SFA-rich diet and was not altered by the substi-tution of a PUFA-rich for an SFA-rich meal or vice versa (Table3). The percentage of postprandial plasma triacylglycerol andTRLs accumulated 7 h after a chronic PUFA-rich diet was sig-nificantly lower than that after a chronic SFA-rich diet. Theextent of accumulation of postprandial plasma triacylglyceroland TRL cholesterol at 7 h increased significantly after the sub-stitution of an SFA-rich meal for a PUFA-rich meal (Table 3) butwas not altered by the substitution of a PUFA-rich meal for anSFA-rich meal (Table 3). Postprandial lipemia at 4 and 7 h tended tocause a net decrease and a net increase in plasma cholesterol after achronic PUFA-rich diet and SFA-rich diet, respectively; however,this difference was not statistically significant (Table 3).
The substitution of an SFA-rich meal for a PUFA-rich mealcaused a statistically significant net increase in cholesterol inboth 4 h and 7 h plasma, whereas the reverse substitution causedno significant change (Table 3). The postprandial lipemia-mediated percentage reduction in LDL cholesterol at 4 h tendedto be greater after a chronic PUFA-rich diet than after a chronicSFA-rich diet (Table 3). The substitution of an SFA-rich meal fora PUFA-rich meal caused a statistically significant lowering ofthe postprandial decrease in LDL cholesterol (Table 3). Thereverse substitution, however, did not significantly alter the ex-tent of the postprandial lipemia-mediated reduction in LDL cho-lesterol (Table 3). The percentage change in HDL cholesterol at4 and 7 h was not significantly altered by either a chronic diet oran acute meal. The above data indicated that an acute SFA-richmeal can blunt the chronic effect of a PUFA-rich diet-mediatedchange in postprandial plasma lipids and lipoprotein-cholesterolconcentration; however, an acute PUFA-rich meal does not havethe same blunting effects on chronic SFA-mediated changes.
Effect of in vitro reactions of endogenous LCAT andCETP in fasting and postprandial plasma or blood on thechanges in the cholesterol content of TRLs
To determine whether the postprandial lipemia-mediated de-crease in LDL and HDL cholesterol and the concomitant increasein TRL cholesterol (Table 2 and Figure 3) were due to a transfer
of CE from LDL and HDL to TRLs via the postprandial lipemia-mediated enhancement of LCAT and CETP activities in vivo(18–20), we sought to determine in vitro whether these endog-enous LCAT and CETP reactions increased the cholesterol con-tent of TRLs or decreased the content of the LDL�HDL fractionin fasting and postprandial plasma (Figure 4, left panel). In vitroincubation at 37 °C of fasting and postprandial plasma obtainedafter both types of diets produced a net increase in the cholesterolcontent of TRLs via a concomitant decrease in the cholesterolcontent of the LDL�HDL fraction (Figure 4, left panel A). Afterincubation, the amount of cholesterol accepted by TRLs from theLDL�HDL fraction was significantly (�2 times) more in 4 hplasma than in fasting plasma after both types of diets (Figure 4,left panels A and B). The amount of cholesterol accepted byTRLs from the LDL�HDL fraction in 7 h plasma was lower thanthat at 4 h because of in vivo clearance of postprandial TRLs(Figure 4, left panels A and B).
When the extent (percentage) of increase in the cholesterolcontent of TRLs in fasting and postprandial plasma after incu-bation was compared, the percentage increases in the cholesterolcontent of TRLs in 4 h plasma with both diets was not signifi-cantly different from that in fasting plasma or in 7 h plasma(Figure 4, left panel C). The percentage increase in the choles-terol content of TRLs in fasting and 4 h and 7 h plasma obtainedafter a PUFA-rich diet did not differ significantly from that (infasting and 4 h and 7 h plasma) obtained after an SFA-rich diet(Figure 4, left panel C). The above data indicate that dietary fatcomposition has no significant influence on the ability of TRLsin fasting and postprandial plasma to accept cholesterol from theLDL�HDL fraction during the in vitro reaction of LCAT andCETP. We reported previously that TRLs in fasting and post-prandial plasma can serve as potent acceptors of cholesterolreleased from red blood cell membranes via LCAT and CETP(27, 28). We found in this study that the level of cholesterolaccepted from red blood cell membranes by TRLs in fastingplasma via LCAT and CETP increased with the appearance ofpostprandial TRLs and then decreased with the clearance ofpostprandial TRLs from plasma in vivo with both diets (Figure 4,
TABLE 3Comparison of changes in the percentage of plasma lipid and lipoprotein-cholesterol concentrations after a diet rich in polyunsaturated (PUFA) orsaturated (SFA) fatty acids (chronic effect) or a single meal (acute effect)1
Lipids and lipoprotein C
SFA-rich diet PUFA-rich diet
SFA-rich meal PUFA-rich meal PUFA-rich meal SFA-rich meal
% of fasting value % of fasting valueChanges at 4 h (4 h vs fasting)
Plasma TG 82.7 � 48.8 70.4 � 50.2 53.5 � 25.8 55.4 � 34.8Plasma C 0.6 � 3.2a,b �1.0 � 3.7a,b �2.3 � 4.4a 0.7 � 5.2b
TRL-C 61.6 � 34.2 48.5 � 28.6 50.9 � 27.2 64.2 � 40.4LDL-C �4.2 � 5.0a,b �4.9 � 6.4a,b �7.7 � 5.7a �3.4 � 6.6b
HDL-C �2.9 � 5.9 �2.8 � 4.0 �4.8 � 5.0 �3.2 � 6.7Changes at 7 h (7 h vs fasting)
Plasma TG 45.8 � 48.8b 41.3 � 46.2b 7.4 � 16.1a 29.2 � 33.2b
Plasma C 0.9 � 3.0a,b �0.9 � 3.7a,b �1.5 � 4.9a 0.8 � 3.7b
TRL-C 34.8 � 33.2b 28.3 � 47.0a,b 5.6 � 16.2a 36.5 � 39.6b
LDL-C �2.2 � 5.9 �4.0 � 6.7 �3.1 � 6.2 �3.6 � 6.4HDL-C 1.4 � 6.6 0.9 � 6.9 �0.9 � 6.9 �0.1 � 6.8
1 All values are x� � SD; n � 16. Means in a row with different superscript letters are significantly different, P � 0.05 (repeated-measures ANOVA withTukey’s post hoc test). TG, triacylglycerol; C, cholesterol; TRL, triacylglycerol-rich lipoproteins (VLDL in fasting plasma or VLDL and chylomicrons inpostprandial plasma).
1152 CHUNG ET AL
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

right panel A). After incubation with red blood cells, the per-centage increase in the cholesterol content of TRLs in fastingplasma, 4 h plasma, and 7 h plasma obtained after an SFA-richdiet did not differ significantly from that in TRLs obtained aftera PUFA-rich diet (Figure 4, right panel C). The above data indi-cate that the potencies to accept cholesterol released from redblood cells by TRL particles in fasting and postprandial plasmawere not affected by dietary fat composition.
Effects of postprandial lipemia on plasma CETPconcentrations and activity
We further examined whether the postprandial decrease incholesterol in the LDL�HDL fraction and the concomitant in-crease in TRL cholesterol were due to a postprandial lipemia-mediated increase in plasma CETP concentration and activity
(Figure 5). As indicated in Figure 5 (panel A), the CETP con-centration in fasting plasma was not affected by either the con-sumption of a PUFA-rich diet for 16 d or by the induction ofpostprandial lipemia. Similarly, no significant change in theplasma CETP concentration was observed when subjects con-sumed the SFA-rich diet for 16 d and when postprandial lipemiawas subsequently induced (data not shown). We examined CETPactivity by incubating fresh fasting and postprandial plasma withan equal amount of [3H]CE-labeled LDL and HDL and thenmeasured the amount of [3H]CE that appeared in TRLs. Thisstudy showed that the rate and extent of transfer of [3H]CE fromthe LDL�HDL fraction to TRLs were much greater in 4 h plasmathan in fasting plasma (Figure 5, B). The decrease in TRL con-centrations in 7 h plasma via the clearance of postprandial TRLsin vivo largely eliminated the earlier increase in CETP activity in
FIGURE 4. Effect of incubating fresh (4 °C) fasting, 4 h, and 7 h plasma samples, which were obtained after the consumption of a diet rich in polyunsaturated(PUFA) or saturated (SFA) fatty acids for 16 d at 37 °C (for 16 h) in the absence (left panel) or presence (right panel) of red blood cells (RBCs) on the changein the lipoprotein-cholesterol profile of a normolipidemic subject (A) and on the increase in mean (�SD) concentrations (B) and percentages (C) oftriacylglycerol-rich lipoprotein [TRL: VLDL � chylomicrons (CM)] cholesterol in all subjects (n � 16). Bars within each panel with different letters aresignificantly different, P � 0.05 (repeated-measures ANOVA with Tukey’s post hoc test).
POSTPRANDIAL LIPEMIA AND FASTING PLASMA CHOLESTEROL 1153
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

4 h plasma (Figure 5, B). Further study showed that the transferof [3H]CE from the LDL�HDL fraction to TRLs in 4 h plasmawas markedly reduced by the centrifugal removal of chylomi-crons from 4 h plasma but was increased by the enrichment of 4 hplasma with autologous chylomicron before its incubation (Fig-ure 5, C). Because postprandial lipemia did not change CETPconcentrations (Figure 5, A) and because CETP is responsible forthe intraplasma transfer of CE (20), it is possible that postprandiallipemia increases the transfer of CE from LDL or HDL to TRLsin vivo by increasing the concentration of chylomicrons thatserve as acceptors of the CETP reaction. We observed also thatthe appearance of postprandial TRLs in 4 h plasma significantlyincreased the partitioning of [3H]CE formed from [3H]UC inplasma by LCAT into TRLs with a concomitant significant de-crease in its partitioning into the LDL�HDL fraction (data notshown). The above observations suggest that the CETP-mediated transfer of endogenous CE in LDL and HDL and inLCAT-produced CE to TRLs may be increased by the appear-ance of postprandial TRLs in plasma.
Effects of postprandial lipemia and in vitro reactions ofLCAT and CETP on the changes in the fatty acyl chaincomposition of triacylglycerol associated with TRLs orthe LDL�HDL fractions
CETP in plasma catalyzes the reciprocal transfer of CE andtriacylglycerol between TRLs and the LDL�HDL fractions(20). To determine whether the CETP-mediated transfer of tri-acylglycerol from postprandial TRLs to the LDL�HDL fractionoccurs in vivo, we examined the effect of postprandial lipemia onthe appearance of dietary fat–derived triacylglycerol on post-prandial TRLs and the LDL�HDL fraction by analyzing thepostprandial change in the fatty acyl chain composition (P:S) oftriacylglycerol associated with TRLs and the LDL�HDL frac-tion.
We previously observed that the P:S of triacylglycerol asso-ciated with TRLs in fasting plasma obtained from subjects ontheir habitual diets increased significantly after the intake of ameal rich in PUFAs (34). Thus, we obtained fresh fasting andpostprandial plasma from the subjects while they were consum-ing their habitual diets (day 1) before and after they consumed aPUFA-rich meal (P:S � 2.0) or an SFA-rich meal (P:S � 0.25).The mean P:S ratios of triacylglycerol in the VLDL andLDL�HDL fractions were not significantly different (0.82 and0.88, respectively) in fasting plasma, which indicated that triac-ylglycerol in VLDL equilibrated with triacylglycerol in theLDL�HDL fraction (Table 4). When a PUFA-rich meal (P:S �2.0) was fed acutely, the mean P:S of chylomicron triacylglycerolin 4 h plasma was 1.71 (Table 4). The P:S of triacylglycerol inVLDL isolated from chylomicron-free 4 h plasma obtained aftera PUFA-rich meal was significantly higher than that of VLDL infasting plasma (Table 4). The mean P:S of chylomicron triacyl-glycerol in 4 h plasma was observed to be 0.47 when an SFA-richmeal (P:S � 0.25) was fed acutely to the same subjects on theirhabitual diet (data not shown). The above observations suggestthat the triacylglycerol composition of chylomicron and VLDLin postprandial plasma reflects the fat composition of an acutelyingested meal.
The triacylglycerol P:S of the LDL�HDL fraction in 4 hplasma was also significantly higher than that in fasting plasma,whereas the triacylglycerol P:S of the LDL�HDL fraction washigher in 7 h plasma than in 4 h plasma and was not significantly
FIGURE 5. Cholesteryl ester transfer protein (CETP) concentrations(A) and activities (B and C) in fasting plasma obtained from normolipidemicsubjects before (day 1) and in fasting, 4 h, and 7 h plasma samples obtainedafter 16 d of consuming a diet rich in polyunsaturated fatty acids (PUFAs). A:Mean (�SD) CETP concentrations in fasting plasma before (day 1) and infasting, 4 h, and 7 h plasma obtained after 16 d of the consumption of aPUFA-rich diet (n � 12) as measured by an enzyme-linked immunosorbentassay. There were no significant differences between fasting concentrationson day 1 and day 16 or between 4 h and 7 h plasma on day 16. B: Mean (�SD)CETP activities (in triplicate) in fasting, 4 h, and 7 h plasma and in plasma freeof triacylglycerol-rich lipoproteins (TRLs) (density � 1.006 g/mL) as as-sayed by measuring the rate and extent of transfer of [3H]cholesteryl ester(CE) from the LDL�HDL fraction to TRLs. C: Mean (�SD) CETP activitiesafter the centrifugal removal of chylomicrons (CMs) from 4 h plasma andafter the addition of isolated CMs to 4 h plasma, as assayed by measuring therate and extent of transfer of CE from the LDL�HDL fraction to TRLs. TG,triacylglycerol. PP, postprandial; cpm, counts per minute.
1154 CHUNG ET AL
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

different from the triacylglycerol P:S of TRLs (Table 4). Afterincubation of fasting and postprandial plasma, the triacylglycerolconcentration of the LDL�HDL fraction increased significantly(2.2- to 2.7-fold) because of the CETP-mediated transfer of tri-acylglycerol from TRLs to the LDL�HDL fraction; however,this incubation minimally altered the P:S of triacylglycerol as-sociated with the LDL�HDL fraction (Table 4). The above dataindicate that, at 4 h, a near equilibrium existed between triacyl-glycerol in TRLs and the LDL�HDL fraction, which suggestedthat the CETP-mediated reciprocal transfer of triacylglyceroland CE between the postprandial TRLs and LDL�HDL frac-tions occurred in vivo. The occurrence of a significant postpran-dial change in the triacylglycerol P:S on the LDL�HDL fractionwithout a significant change in its concentration at 4 h furthersuggested that triacylglycerol accepted by LDL and HDL frompostprandial TRLs are rapidly hydrolyzed by hepatic lipase invivo.
DISCUSSION
We examined the effects of the appearance and clearance ofpostprandial TRLs serving as acceptors of LCAT and CETPreaction products on the postprandial changes in endogenouslipoprotein-cholesterol concentrations in normolipidemic sub-jects who have followed PUFA- or SFA-rich diets for 20 d. Ourstudy showed that both diets affected not only the cholesterolconcentrations of lipoproteins in fasting plasma but also thepostprandial changes in the distribution of cholesterol amonglipoproteins in plasma. LDL and HDL cholesterol in fastingplasma decreased after a PUFA-rich diet but increased after aSFA-rich diet (Table 1), which further confirmed the knowncholesterol-lowering effect of a PUFA-rich diet and thecholesterol-raising effect of an SFA-rich diet in humans (1–3).
Many previous studies conducted in human volunteersshowed that subjects who consumed an SFA-rich diet had higherconcentrations of postprandial TRLs and their remnants and thatthese postprandial TRL remnants cleared more slowly than insubjects who consumed a PUFA-rich diet (8–11). Consistentwith reports from other investigators (8–11), we observed in thecurrent study that at 7 h, postprandial TRL cholesterol, apoB-100, apo B-48, and plasma triacylglycerol accumulated to agreater degree after an SFA-rich diet than after a PUFA-rich diet(Table 2 and Figures 2 and 3). The SFA-rich background dietdelayed the clearance of apo B-100–containing TRLs more than
it did the clearance of apo B-48–containing TRLs and was foundto have a more potent influence on the clearance of postprandialTRLs than did a single meal (Table 2, Figures 2 and 3), whichconfirmed the previous findings of Bergeron and Havel (11). Theincreased accumulation of postprandial TRLs in plasma 7 h afteran SFA-rich diet or meal could possibly have been due to thedelayed hepatic clearance of the lipolytic remnants of postpran-dial TRLs or to the delayed appearance of postprandial TRLs inplasma because SFA is less well absorbed than is PUFA (36, 37).
Our study showed that the induction of postprandial lipemiaafter both types of diets resulted in a significant increase in TRLcholesterol and triacylglycerol and a concomitant transient de-crease in LDL and HDL cholesterol (Table 2 and Figure 3).Although the abovementioned postprandial lipemia-mediatedchanges in lipoprotein-cholesterol concentrations have been re-ported by many other investigators (11, 21-25), the mechanismresponsible for these changes has not been fully explored in otherstudies. The simultaneous postprandial increase in concentra-tions of TRL-associated triacylglycerol and cholesterol in 4 hplasma (Figures 2 and 3) could be due to increased secretions ofpostprandial TRLs from the intestines or liver induced by in-gested triacylglycerol and cholesterol. However, many studies(38, 39) showed that dietary cholesterol (deuterated), when pro-vided as a single fat-containing meal, does not simultaneouslyappear with triacylglycerol but is secreted in intestinally derivedchylomicrons during several subsequent phases in healthy hu-mans. The maximal dietary (deuterated) cholesterol appearing inpostprandial chylomicron at the postprandial lipemic peak wasonly 1% of ingested cholesterol (38), which indicated that thebulk of cholesterol associated with chylomicrons at the postpran-dial lipemic peak was derived from endogenous sources. Be-cause the postprandial increase in TRL cholesterol occurred con-comitant with a decrease in LDL and HDL cholesterol (Table 2and Figure 3), postprandial enhancement in the CETP-mediatedtransfer of CE from LDL and HDL to TRLs in vivo could havebeen the cause. The CETP-mediated transfer of cholesterol fromendogenous lipoproteins to TRLs in fasting plasma in vitro in-creased with the appearance of postprandial TRLs in plasma at4 h without a change in plasma CETP mass (Figure 4). Thisobservation suggests that the postprandial enhancement of CETPactivity may be due to the appearance of postprandial TRLs,which serve as acceptors of a CETP reaction. Furthermore, thepostprandial lipemia-mediated decrease in cholesterol in theLDL�HDL fraction was associated with an increase in the P:S of
TABLE 4Ratio of polyunsaturated to saturated fatty acids (P:S) and concentrations of triacylglycerol (TG) associated with TG-rich lipoproteins (TRLs) and theLDL�HDL fraction isolated from control and incubated fasting and postprandial plasma1
Plasma fraction
Fasting plasma 4 h plasma 7 h plasma
Control Incubated Control Incubated Control Incubated
P:S of TGTRL 0.82 � 0.18a — 1.17 � 0.14b — 1.29 � 0.14c —CM — — 1.71 � 0.17 — — —LDL�HDL 0.88 � 0.13a 0.90 � 0.18a 1.07 � 0.08b 1.11 � 0.14b 1.20 � 0.18e 1.24 � 0.14c
TG concentration (mmol/L)LDL�HDL 0.32 � 0.07a 0.71 � 0.22b 0.32 � 0.08a 0.89 � 0.27c 0.33 � 0.08a 0.76 � 0.26b
1 All values are x� � SD; n � 7. Means in a row with different superscript letters are significantly different, P � 0.05 (repeated-measures ANOVA withTukey’s post hoc test). Fasting and postprandial plasma samples were obtained after the habitual (home) diet on day 1 followed by consumption of the PUFA-richmeal. CM, chylomicrons; TRL indicates the VLDL in fasting plasma or VLDL and CM in postprandial plasma.
POSTPRANDIAL LIPEMIA AND FASTING PLASMA CHOLESTEROL 1155
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

triacylglycerol in the LDL�HDL fraction that mirrors that of thefatty acids in the ingested meal (Table 4). This postprandialincrease in the P:S of triacylglycerol in the LDL�HDL fractionwas likely a consequence of the CETP-mediated reciprocal ex-change of CE and triacylglycerol between the TRL andLDL�HDL fractions in vivo, because dietary triacylglycerol isnot directly incorporated into LDL and HDL during a short post-prandial lipemic period. Studies that examined the fate of [3H]CEon HDL in humans and in rabbits, with active CETP in plasma,showed that [3H]CE on HDL, injected intravenously, appeared inboth VLDL and LDL (40, 41). In rabbits, the peak activity of[3H]CE in injected HDL was seen in VLDL and LDL within30–60 min and 60–120 min, respectively (41). These findingsindicated that the CETP-mediated transfer of CE from HDL toTRLs in vivo is a rapid process. Under normal conditions, the rateof the CETP-mediated transfer of CE from HDL to TRLs in vivowas higher than the catabolic rate of endogenous lipoproteinsand, thus, the pools of CE among lipoproteins in the fasting stateapproach equilibrium in vivo (42). However, it is probable thatthe CETP reaction during the postprandial lipemic state, whichlasts several hours, would be sufficient to alter the equilibrium ofCE between TRLs and endogenous lipoproteins in vivo becausepostprandial TRLs serving as additional acceptors of the CETPreaction clear at a faster rate than that of the CETP reaction orendogenous lipoprotein catabolism.
Oliviera et al (43) reported that, in chyluric patients, 58% and18% of CE in lymph chylomicrons derived from HDL and LDL,respectively. This finding further supports the hypothesis that theCETP-mediated CE transfer from LDL and HDL to postprandialchylomicrons occurs in humans in vivo. Thus, the CETP-mediated transfer of CE from LDL and HDL to postprandialTRLs and the clearance of postprandial TRLs via hepatic uptakein vivo could lead to the net decrease in concentrations of CE inplasma and cholesterol in both LDL and HDL observed in thisstudy (Table 2 and Figure 3). In a comparison of the effects of aPUFA- and an SFA-rich diet on postprandial changes in plasmalipid and lipoprotein-cholesterol concentrations (Figure 3, Ta-bles 2 and 3), we observed that the SFA- and PUFA-mediatedinduction of postprandial lipemia produced qualitatively similar,but significantly different, quantitative postprandial changes inplasma lipid and lipoprotein-cholesterol concentrations. Thesequantitative differences include 1) the net postprandial decreasein plasma total cholesterol 4 h after a PUFA-rich diet comparedwith the net increase 4 h after an SFA-rich diet; 2) the significantpostprandial reduction in the concentrations of plasma CE, LDLcholesterol, and HDL cholesterol 4 h after a PUFA-rich dietcompared with a nonsignificant reduction after an SFA-rich diet,and 3) the near complete clearance of postprandial TRLs 7 h aftera PUFA-rich diet compared with the significant accumulation ofpostprandial TRLs after an SFA-rich diet (Table 2 and Figure 3).In earlier studies performed in young men, the extent of post-prandial reductions in concentrations of cholesterol in LDL andHDL (�mg/dL) observed with a PUFA-rich diet and meal wereshown to be similar to those observed with an SFA-rich diet andmeal (11, 24). Because LDL- and HDL-cholesterol concentra-tions in fasting plasma were significantly greater after an SFA-rich diet than after a PUFA-rich diet (Table 1), the postprandiallipemia-mediated percentage reduction in LDL cholesterol 4 hafter the PUFA-rich diet tended to be greater than that after achronic SFA-rich diet; the change was significantly minimizedby the substitution of an SFA-rich meal for a PUFA-rich meal
(Table 3). The mechanisms responsible for the above differencesin the extent of postprandial changes in the percentage of endog-enous lipoprotein cholesterol and plasma CE in vivo are notentirely clear but could be the result of different clearance ratesfor postprandial TRLs. Because the ability of TRLs to serve asacceptors of cholesterol from LDL and HDL after the in vitroreaction of LCAT and CETP was not affected by the type ofdietary fat (Figure 4, left panel), the rapid clearance of postpran-dial TRLs carrying CE accepted from LDL and HDL via LCATand CETP in vivo could cause a significant postprandial reduc-tion in the concentration of plasma CE and LDL and HDL cho-lesterol and a net decrease in plasma cholesterol at 4 h with thePUFA-rich diet. Because humans are mostly in the postprandiallipemic state during the day because of the consumption of mealsat regular intervals, the CETP-mediated transfer of CE from LDLand HDL to postprandial TRLs and the subsequent rapid clear-ance of postprandial TRLs during the consumption of a PUFA-rich diet would lead to the reduction of both LDL and HDLcholesterol in fasting plasma. For the diet rich in SFA, the lack ofa significant postprandial lipemia-mediated reduction in plasmaCE and LDL and HDL cholesterol and significant accumulationof postprandial TRLs at 7 h (Table 2) could have led to anincrease in cholesterol in plasma LDL, HDL, and total choles-terol. Fielding and Fielding (44) reported that the generation ofCE by LCAT, in situ, inhibited LCAT activity, provided that theproduct of LCAT activity remained in the HDL. It is possible thatthe accumulation of cholesterol accepted from LDL and HDL bypostprandial TRLs in plasma, due to an SFA-mediated delayedclearance, could limit the CETP-mediated transfer of CE fromLDL and HDL to TRLs via a feedback inhibition mechanism.Thus, rapid removal of postprandial TRLs carrying cholesterolaccepted from LDL and HDL via CETP could be of potentialimportance in lowering endogenous lipoprotein-cholesterol con-centrations. It is possible that the extent of flux of cholesterol inLDL and HDL via postprandial TRLs may be substantially lowerwith an SFA-rich than with a PUFA-rich diet because of a de-layed hepatic removal of postprandial TRLs with an SFA-richdiet. Our current study also showed that postprandial TRLs ap-pearing in plasma can serve as a potent acceptor of cholesterolreleased from cell membranes via LCAT and CETP and thatdietary fat composition had no significant influence on the po-tencies of postprandial TRLs to accept cholesterol released fromred blood cells via LCAT and CETP (Figure 4, right panel).Because postprandial TRLs will be cleared from circulatingplasma primarily via hepatic uptake (29), the LCAT- and CETP-mediated transfer of cholesterol from cell membranes to post-prandial TRLs may lead to the removal of that cholesterol by theliver. It is possible that the transport of cholesterol on cell mem-branes to the liver via postprandial TRLs are enhanced with aPUFA-rich diet because of the rapid clearance of postprandialTRLs and are impaired with an SFA-rich diet because of thedelayed clearance of postprandial TRLs. Many case control stud-ies have shown that the delayed clearance of postprandial TRLsis more prevalent in CVD patients than in control subjects (45–47). Thus, the SFA-mediated increase or PUFA-mediated de-crease in the risk of CVD (2) could be mediated by differences intheir respective clearance rates of postprandial TRLs carryingcholesterol accepted from cell membranes via LCAT and CETP.In summary, data obtained from our current and previous studies(27, 28) suggest that postprandial lipemia in humans is associatedwith an active transfer of CE from endogenous lipoproteins or
1156 CHUNG ET AL
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

cell membranes to postprandial TRLs via LCAT and CETP. Ourcurrent study also indicates that the clearance rate of postprandialTRLs is influenced by dietary fat composition. With a chronicPUFA-rich diet, the rapid clearance of postprandial TRLs, whichcarry cholesterol accepted from LDL and HDL via CETP andLCAT, may lead to a decrease in endogenous lipoprotein cho-lesterol in fasting plasma. Conversely, with a chronic SFA-richdiet, the delayed clearance of postprandial TRLs, which carrycholesterol accepted from cell membranes, may lead to an in-crease in both LDL and HDL cholesterol. Thus, the clearance rateof postprandial TRLs may play an important role in regulatingfasting plasma cholesterol concentrations. Because postprandialTRLs act as potent ultimate acceptors of cholesterol releasedfrom cell membranes via LCAT and CETP, dietary fat–mediatedalterations in the clearance rate of postprandial TRL carryingcholesterol accepted from cell membranes could affect the risk ofCVD by altering the rate at which cholesterol is transported to theliver for excretion, a process often referred to as reverse choles-terol transport.
B-HC and FF developed the study concept. LO recruited and characterizedthe study subjects and monitored the subjects’ adherence to the test diet. BDformulated and prepared the test meals and diets. SD and PL measured theplasma lipids and lipoproteins. BHSC analyzed the fatty acyl chain compo-sition of dietary fat and lipoprotein triacylglycerols. RAO performed thestatistical analysis. B-HC drafted the manuscript. None of the authors had anyfinancial interest in this work or with the sponsors of this work.
REFERENCES1. Howell HW, McNamara DJ, Tosca MA, Smith BT, Gaines JA. Plasma
lipid and lipoprotein responses to dietary fat and cholesterol: a meta-analysis. Am J Clin Nutr 1997;65:1747–64.
2. Caggiula AW, Mustard AA. Effects of dietary fat and fatty acids oncoronary artery disease risk and total lipoprotein-cholesterol concentra-tions: epidemiological studies. Am J Clin Nutr 1997;65(suppl):1597S–610S.
3. Schaefer EJ. Lipoproteins, nutrition and heart disease. Am J Clin Nutr2002;75:191–212.
4. Wollet LA, Spady DK, Dietschy JM. Saturated and unsaturated fattyacids independently regulate low density lipoprotein receptor activityand production rate. J Lipid Res 1992;33:77–88.
5. Dietschy JM. Theoretical consideration of what regulates low densitylipoproteins and high density lipoprotein-cholesterol. Am J Clin Nutr1997;65(suppl):1581S–9S.
6. Cohn JS, Johnson EJ, Millar JS, et al. Contribution of apo B-48 and apoB-100 triglyceride-rich lipoproteins (TRL) to post-prandial increases inthe plasma concentration of TRL triglycerides and retinyl esters. J LipidRes 1993;34:2033–40.
7. Schneeman BA, Kotite L, Todd KM, Havel RJ. Relationships betweenresponse of triglyceride-rich lipoproteins in blood containing apoli-poprotein B-48 and 100 to a fat-containing meal in normolipidemichumans. Proc Natl Acad Sci U S A 1993;90:2069–73.
8. Weintraub MS, Zechner R, Brown A, Eisenberg S, Breslow J. Dietarypolyunsaturated fats of �-6 and �-3 series reduce post-prandial lipopro-tein concentrations. J Clin Invest 1988;82:1884–93.
9. Harris WS, Connor WE, Alam L, Illingworth DR. Reduction of post-prandial triglyceridemia in humans by dietary n�3 fatty acids. J LipidRes 1988;29:1451–60.
10. Demacker PN, Reijnen IGM, Katan MB, Stuyt PMJ, Stalenhoef AFH.Increased removal of remnants of triglyceride-rich lipoproteins on a dietrich in polyunsaturated fatty acids. Eur J Clin Invest 1991;21:197–203.
11. Bergeron N, Havel RJ. Influence of diets-rich in saturated fat andomega-6 polyunsaturated fatty acids on the post-prandial response ofapolipoproteins B-48, B-100, E and lipids in triglyceride-rich lipopro-teins. Arterioscler Thromb Vasc Biol 1995;15:2111–21.
12. Sposito A, Santos RD, Hueb W, et al. LDL concentration is correlatedwith the removal from plasma of a chylomicron-like emulsion in subjectswith coronary artery disease. Atherosclerosis 2002;161:447–53.
13. Reitsma JB, Castro CC, de Bruin TW, Erkelens DW. Relationship be-tween improved post-prandial lipemia and low density lipoprotein me-tabolism during treatment with tetrahydrolipstatin, a pancreatic lipaseinhibitor. Metabolism 1994;43:293–8.
14. Boquist S, Karpe F, Danell-Tverud K, Hamsten A. Effect of atorvastatinon post-prandial plasma lipoproteins in post-infarction patients withcombined hyperlipidemia. Atherosclerosis 2002;162:163–70.
15. Sposito AC, Santos RD, Amancio RF, Ramires JAF, Chapman MJ,Maranhao RC. Atorvastatin enhances the plasma clearance ofchylomicron-like emulsion in subjects with atherogenic dyslipidemia:relevance to the in vivo metabolism of triglyceride-rich lipoproteins.Atherosclerosis 2003;166:311–21.
16. Parhofer KG, Laubach E, Barrett PHR. Effect of atorvastatin on post-prandial lipoprotein metabolism in hypertriglyceridemic patients. JLipid Res 2003;44:1192–98.
17. Schaefer EJ, Lichtenstein AH, Lamon-Fava S, McNamara JR, OrdovaJM. Lipoproteins, nutrition, aging and atherosclerosis. Am J Clin Nutr1995;61(suppl):726S–40S.
18. Rose HG, Juliano J. Regulation of plasma lecithin cholesterol acyltrans-ferase in man. Activation during alimentary lipemia. J Lab Clin Med1977;89:524–32.
19. Castro GR, Fielding CJ. Effect of post-prandial lipemia on plasma cho-lesterol metabolism. J Clin Invest 1985;75:874–82.
20. Tall AR. Plasma cholesterylester transfer protein. J Lipid Res 1993;34:1255–74.
21. Dubois C, Armand A, Azais-Braesco V, et al. Effect of a moderateamount of emulsified dietary fat on post-prandial lipemia and lipopro-teins in normolipidemic adults. Am J Clin Nutr 1994;60:374–82.
22. Lassel TS, Guerin M, Auboiron S, Guy-Grand B, Chapman MJ. Evi-dence for a cholesterylester donor activity of LDL particles during ali-mentary lipemia in normolipidemic subjects. Atherosclerosis.1999;147:41–8.
23. Fielding PE, Jackson EJ, Fielding CJ. Chronic dietary fat and cholesterolinhibit the normal post-prandial stimulation of plasma cholesterol me-tabolism. J Lipid Res 1990;30:1211–7.
24. Fielding CJ, Havel RJ, Tod KM, et al. Effect of dietary cholesterol andfat saturation on plasma lipoproteins in an ethnically diverse populationin healthy young men. J Clin Invest 1995;85:611–8.
25. Dullaart RPF, Groener JEM, Wijk HV, Sluiter WJ, Erkelens DW. Ali-mentary lipemia-induced redistribution of cholesterylester between li-poproteins: Studies in normolipidemic, combined hyperlidemic and hy-percholesterolemic men. Arteriosclerosis 1989;9:614–22.
26. Glomset JA, Norum KR. The metabolic role of lecithin:cholesterol acyl-transferase: perspective from pathology. Adv Lipid Res 1973;11:1–65.
27. Chung BH, Franklin F, Cho SBH, Segrest JP, Hart K, Darnell B. Poten-cies of lipoproteins in fasting and post-prandial plasma to accept addi-tional cholesterol molecules released from cell membranes. ArteriosclerThromb Vasc Biol 1998;18:1217–30.
28. Chung BH, Doran S, Liang P, et al. Alcohol-mediated enhancement ofpost-prandial lipemia: a contributing factor to an increase in plasma HDLand a decrease in risk of cardiovascular disease. Am J Clin Nutr 2003;78:391–9.
29. Cooper AD. Hepatic uptake of chylomicron remnants. J Lipid Res 1997;38:2173–92.
30. Chung BH, Segrest JP, Pfau J, Cone JT, Geer JC, Duncan LA. Highresolution plasma lipoprotein cholesterol profiles by a rapid high volumeautomated method. J Lipid Res 1981;22:1003–14.
31. Chung BH, Segrest JP, Ray MJ, et al. Single vertical spin density gra-dient ultracentrifugation. Methods Enzymol 1986;128:181–209.
32. Kotite L, Bergeron N, Havel RJ. Quantification of apolipoproteinsB-100, B-48 and E in human triglyceride-rich lipoproteins. J Lipid Res1995;36:890–4.
33. Thomas MS, Rudel LL. 3H-cholesteryl ester labeling and transfer amonghuman and nonhuman primate plasma lipoproteins. Anal Biochem 1983;130:215–22.
34. Chung BH, Hennig H, Cho SBH, Darnell B. Effect of the fat compositionof a single meal on the composition and cytotoxic potencies of lipoly-tically released free fatty acids in post-prandial plasma. Atherosclerosis1998;141:321–32.
35. Sakai N, Uchida Y, Ohashi K, et al. Measurement of fasting serum apoB-48 levels in normolipidemic and hypertriglyceridemic subjects byELISA. J Lipid Res 2003;44:1256–62.
36. Bracco U. Effect of triglyceride structure on fat absorption. Am J ClinNutr 1994;60(suppl):1002S–9S.
POSTPRANDIAL LIPEMIA AND FASTING PLASMA CHOLESTEROL 1157
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021

37. Emken EA. Metabolism of dietary stearic acid relative to other fattyacids in human subjects. Am J Clin Nutr 1994;60(suppl):1023S–8S.
38. Dubois C, Armand M, Ferezou J, et al. Post-prandial appearance ofdietary deuterated cholesterol in the chylomicron fraction and wholeplasma in healthy subjects. Am J Clin Nutr 1996;64:47–52.
39. Beaumier-Gallon G, Dubois C, Senft M, et al. Dietary cholesterol issecreted in intestinally derived chylomicrons during several subse-quent post-prandial phases in healthy humans. Am J Clin Nutr 2001;73:870 –7.
40. Nestel PJ, Reardon M, Billington T. In vivo transfer of cholesterylestersfrom high lipoproteins to very low density lipoproteins in humans Bio-chim Biophys Acta 1979;573:403–7.
41. Lally JI, Barter PJ. The in vivo metabolism of esterified cholesterol in theplasma high density lipoproteins in rabbits. J Lab Clin Med 1979;93:570–82.
42. Barter PJ, Brewer HB Jr, Chapman MJ, Hennekens CH, Rader DJ, Tall AR.
Cholesterylester transfer protein: a novel target for raising HDL and inhib-iting atherosclerosis. Arterioscler Thromb Vasc Biol 2003;23:160–7.
43. Oliviera HCF, Nilausen K, Meinertz H, Quintao ECR. Cholesterylesterin lymph chylomicrons: contribution from high density lipoproteins inplasma into intestinal lymph. J Lipid Res 1993;34:1729–36.
44. Fielding CJ, Fielding PE. Cholesterol transport between cells and bodyfluids. Med Clin North Am 1982;66:368–73.
45. Groot PHE, Van Stipout WAHJ, Kraus XH, et al. Post-prandial lipopro-tein metabolism in normolipidemic men with and without coronary heartdisease. Arterioscler Thromb 1991;11:653–62.
46. Patsch JR, Misenbock G, Hofpfenweiser T, et al. Relation of triglyceridemetabolism and coronary artery metabolism in normolipidemic menwith and without coronary artery disease: studies in post-prandial state.Arterioscler Thromb 1992;12:1336–45.
47. Weintraub MS, Grosskopf I, Rassin T, et al. Clearance of chylomicronremnants in normolipidemic patients with coronary artery disease: casecontrol study over three years. BMJ 1996;312:936–9.
1158 CHUNG ET AL
Dow
nloaded from https://academ
ic.oup.com/ajcn/article/80/5/1145/4690416 by guest on 18 D
ecember 2021