correlations between structure, topographic arrangement
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 266~398-412 (1987)
Correlations Between Structure, Topographic Arrangement, and Spectral
Sensitivity of Sound-Sensitive Interneurons in Crickets
GORDON ATKINS AND GERALD S. POLLACK Department of Biology, McGill University, Montreal, Quebec, Canada H3A 1B1
ABSTRACT The morphology of nine prothoracic, sound-activated, interganglionic
interneurons in Teleogryllus oceanicus is described. Only two of the neurons can, on anatomical grounds, receive input directly from auditory receptors. The morphology of many of the cells suggests that they may provide output to motor areas. The nine cells can be divided into two groups on the basis of their spectral sensitivity: high-frequency neurons and low-frequency neu- rons. Correlations were found between morphology and spectral sensitivity. High-frequency neurons have a ventromedial soma, dorsally positioned neu- ropile processes, and an axon in the lateral half of the promesothoracic connective. In contrast, low-frequency neurons have a dorsal and/or laterally positioned soma, neuropile processes in the ventral portion of the prothoracic ganglion, and an axon projecting in the medial half of the connective. These findings reveal the existence of a crude tonotopic organization of central neurons. In addition, they provide hints as to the type of output and the targets of these neurons.
Key words: auditory, insect, anatomy, tonotopy, orthoptera
In another paper (Atkins and Pollack, '871, we describe the response properties of nine sound-sensitive prothoracic interneurons in the cricket Teleogryllus oceanicus. These can be divided into two general classes on the basis of their spectral sensitivity: low-frequency cells and high-frequency cells. In the present paper we present a detailed description of their anatomy, with reference to internal ganglionic structures. One goal of this work is to gather anatomical information which, when compared with similar informa- tion for other neurons, can suggest or rule out connectivity patterns. A second goal is to extend our knowledge of the general anatomical organization of insect auditory systems. In particular, we examine the relationships between the morphology, spatial distribution, and physiological proper- ties of these interneurons.
Functionally relevant spatial distribution of auditory neurons is well known in vertebrates, where tonotopic or- ganization of receptors and central neurons has been thor- oughly studied (Lorente de N6, '33; Woolsey and Walzl, '42; von Bekesy, '60; Merzenich et al., '75). Among inverte- brates, tonotopy has been demonstrated in orthopteran in- sects for the somata, dendrites, and central terminals of receptor cells (Oldfield, '82, '84; Romer, '83, '85; Oldfield et
0 1987 ALAN R. LISS, INC.
al., '86). In the present paper, we show that the locations of the neuropile processes, axons, and somata of interneurons are correlated with spectral sensitivity, i.e., that central neurons of insects show a crude tonotopic organization.
MATERIALS AND METHODS Adult female T oceanicus crickets were prepared for in-
tracellular recording and staining according to the proce- dures described in an earlier paper (Atkins and Pollack, '87). Briefly, a cricket was immobilized ventral side up in a manner that allowed stimulation with sound, light, and wind, and the prothoracic ganglion and the promesothora- cic connectives were exposed.
Interneurons were impaled with glass micropipettes filled with 3% Lucifer Yellow (Aldrich) dissolved in 0.5 M LiCl and backfilled with 2 M LiCl (resistance 100-250 MQ). Dye was injected by applying -1 to -15 nA DC current for up to 20 minutes. To visualize mesothoracic projections, neu- rons were impaled in the promesothoracic connectives and
Accepted July 14, 1987.
SOUND-SENSITIVE INTERNEURONS IN THE CRICKET 399
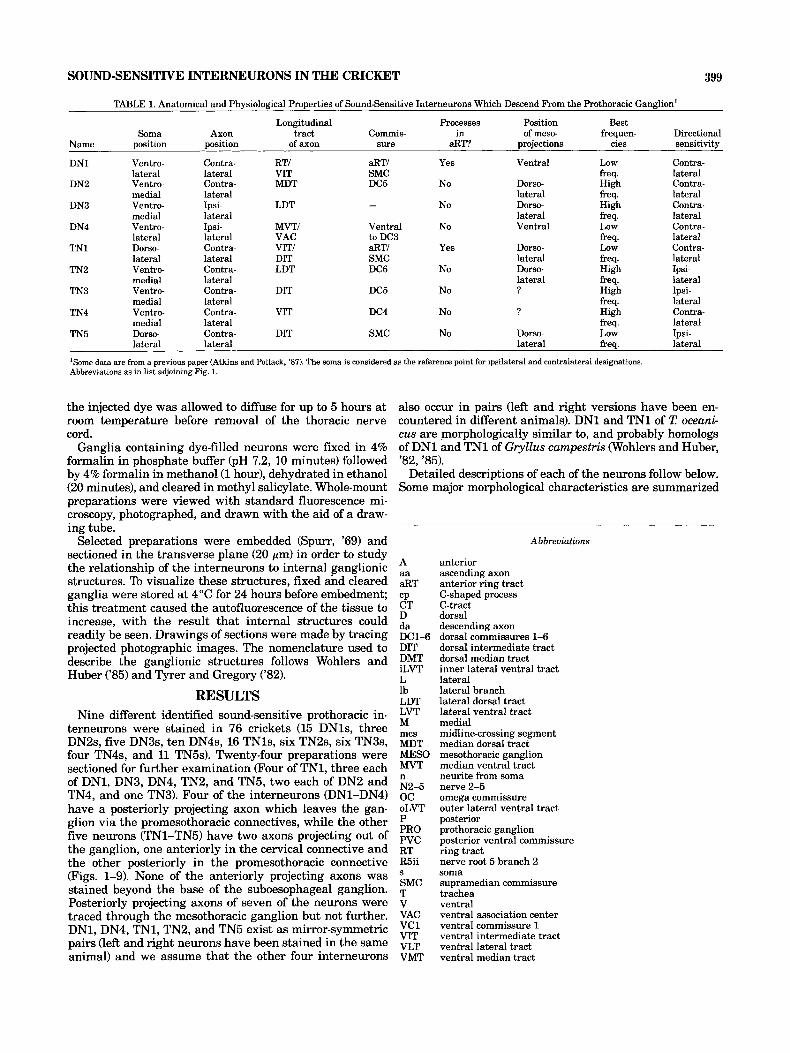
TABLE 1. Anatomical and Physiological Properties of Sound-Sensitive Interneurons Which Descend From the Prothoracic Ganglion'
Longitudinal Processes Position Best Soma Axon tract Commis- in of meso- frequen- Directional
Name oosition Dosition of axon sure aRT? oroiections cies sensitivitv
DN1 Ventro- Contra- RTI &TI Yes Ventral Low Contra-
DN2 Ventro- Contra- MDT DC5 No Dorso- High Contra-
DN3 Ventro- Ipsi- LDT - No Dorm- High Contra-
DN4 Ventro- Ipsi- MVTI Ventral No Ventral Low Contra-
TN1 Dorso- Contra- VITI &TI Yes Dorso- Low Contra-
TN2 Vent r o- Contra- LDT DC6 No Dorso- High Ipsi-
TN3 Ventro- Contra- DIT DC5 No ? High Ipsi-
TN4 Vent r o- Contra- VIT DC4 No ? High Contra-
TN5 Dorso- Contra- DIT SMC No Dorso- LOW Ipsi-
lateral lateral VIT SMC freq. lateral
medial lateral lateral freq. lateral
medial lateral lateral freq. lateral
lateral lateral VAC to DC3 freq. lateral
lateral lateral DIT SMC lateral freq. lateral
medial lateral lateral freq. lateral
medial lateral freq. lateral
medial lateral freq. lateral
lateral lateral lateral freq. lateral
'Some data are from a previous paper (Atkins and Pollack, '87). The soma is considered as the reference p i n t for ipsilateral and contralateral designations. Abbreviations as in list adjoining Fig. 1.
the injected dye was allowed to diffuse for up to 5 hours at room temperature before removal of the thoracic nerve cord.
Ganglia containing dye-filled neurons were fixed in 4% formalin in phosphate buffer (pH 7.2, 10 minutes) followed by 4% formalin in methanol (1 hour), dehydrated in ethanol (20 minutes), and cleared in methyl salicylate. Whole-mount preparations were viewed with standard fluorescence mi- croscopy, photographed, and drawn with the aid of a draw- ing tube.
Selected preparations were embedded (Spurr, '69) and sectioned in the transverse plane (20 pm) in order to study the relationship of the interneurons to internal ganglionic structures. To visualize these structures, fixed and cleared ganglia were stored at 4°C for 24 hours before embedment; this treatment caused the autofluorescence of the tissue to increase, with the result that internal structures could readily be seen. Drawings of sections were made by tracing projected photographic images. The nomenclature used to describe the ganglionic structures follows Wohlers and Huber ('85) and Tyrer and Gregory ('82).
RESULTS Nine different identified sound-sensitive prothoracic in-
terneurons were stained in 76 crickets (15 DNls, three DNZs, five DN3s, ten DN4s, 16 TNls, six TNZs, six TN3s, four TN4s, and 11 TN5s). Twenty-four preparations were sectioned for further examination (Four of TN1, three each of DN1, DN3, DN4, TN2, and TN5, two each of DN2 and TN4, and one TN3). Four of the interneurons (DN1-DN4) have a posteriorly projecting axon which leaves the gan- glion via the promesothoracic connectives, while the other five neurons (TNl-TN5) have two axons projecting out of the ganglion, one anteriorly in the cervical connective and the other posteriorly in the promesothoracic connective (Figs. 1-9). None of the anteriorly projecting axons was stained beyond the base of the suboesophageal ganglion. Posteriorly projecting axons of seven of the neurons were traced through the mesothoracic ganglion but not further. DN1, DN4, TN1, TN2, and TN5 exist as mirror-symmetric pairs (left and right neurons have been stained in the same animal) and we assume that the other four interneurons
also occur in pairs (left and right versions have been en- countered in different animals). DN1 and TN1 of l! oceuni- cus are ,morphologically similar to, and probably homologs of DN1 and TN1 of Gryllus cumpestris (Wohlers and Huber, '82, '85).
Detailed descriptions of each of the neurons follow below. Some major morphological characteristics are summarized
Abbreviations
A aa aRT CP CT D da DC1-6 DIT DMT iLVT L lb LDT LVT M mcs MDT MESO MVT n N2-5 oc oLVT P PRO PVC RT R5ii
SMC T V VAC vc1 VIT VLT VMT
8
anterior ascending axon anterior ring tract C-shaped process C-tract dorsal descending axon dorsal commissures 1-6 dorsal intermediate tract dorsal median tract inner lateral ventral tract lateral lateral branch lateral dorsal tract lateral ventral tract medial midlinecrossing segment median dorsal tract mesothoracic ganglion median ventral tract neurite from soma nerve 2-5 omega commissure outer lateral ventral tract posterior prothoracic ganglion posterior ventral commissure ring tract nerve root 5 branch 2 soma supramedian commissure trachea ventral ventral association center ventral commissure 1 ventral intermediate tract ventral lateral tract ventral median tract
A
B
C
00
I J’ MESO A
L-
H
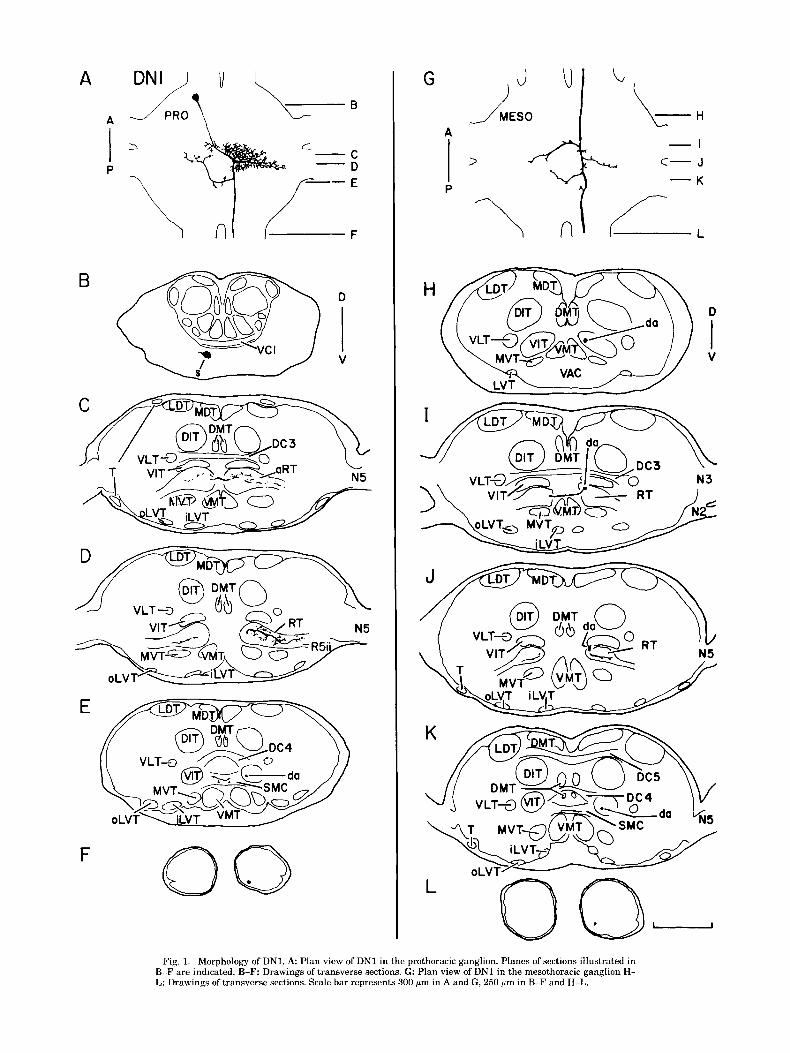
Fig. 1. Morphology of DNI. A. Plan view of DNI in the prothoracic ganglion. Planes of sections illustrated in B-F are indicated. R-F: Drawings of transverse sections. G: Plan view of DN1 in the mesothoracic ganglion H- I,: Drawings of transverse sections. Scale bar represents 300 pm in A and G, 250 pm in B-F and H-L.

SOUND-SENSITIVE INTERNEURONS IN THE CRICKET
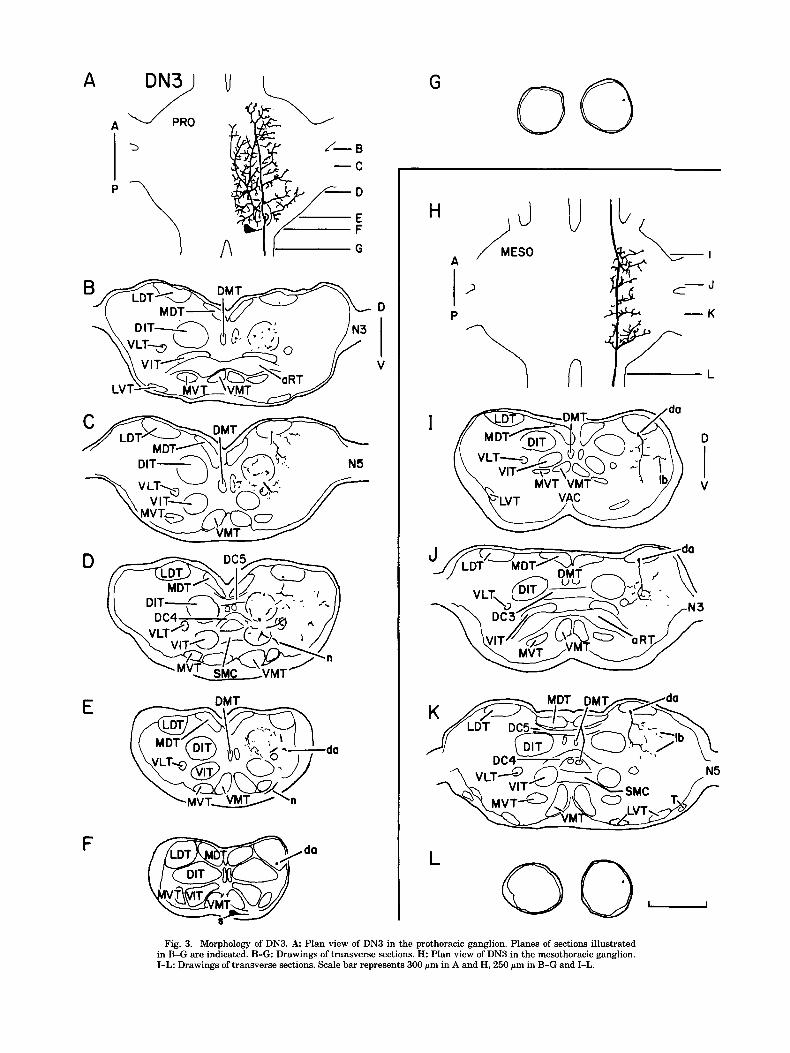
DN3 in Table 1, which also includes physiological characteristics that are described more thoroughly elsewhere (Atkins and The soma of DN3 is located medially on the ventral sur- Pollack, '87). face of the ganglion near the origin of the promesothoracic
connective (Fig. 3A,F). A neurite extends laterally from the soma (Fig. 3E,F), bends dorsomedially, traverses through DN1
The soma of DN1 is situated anteriorly in the prothoracic VIT, and connects to a process occupying the DIT (Fig. ganglion near the origin of the cervical connectives (Fig. 3D,E). Two heavily branched processed extend anteriorly 1A,B). A neurite projects posteromedially, extends a few from their point of origin on the axon (Fig. 3A). The more processes into the soma-ipsilateral portion of the anterior ventral and medial of these projects into the DIT and also ring tract (aRT) (Fig. 1C), crosses the ganglionic midline extends branches to the DMT, the VIT, and the ventral (Fig. lC), and gives rise to a laterally directed neuropile lateral tract (VLT) (Fig. 3B-D). The other major process segment that projects through the aRT into the region projects anteriorly into the LDT (Fig. 3A-D) and also pro- occupied by nerve 5 root 2 (R5ii) (Fig. ID). A posteriorly duces branches which extend ventrally to the DIT and directed axon arises from the neuropile segment, exits the laterally toward the periphery of the ganglion (Fig. 3C,D). aRT, and enters the ventral intermediate tract WIT) (Fig. The descending axon originates from the fusion of the two ID-E). A medially projecting branch originates from the major anteriorly projecting processes and enters the LDT axon, crosses the ganglionic midline in the supramedian (Fig. 3F). The axon exits the ganglion in the dorsolateral commissure (SMC) (Fig. lE), and projects a few processes quadrant of the promesothoracic connective (Fig. 3G). anteriorly into the aRT (Fig. 1D). The axon leaves the In the mesothoracic ganglion, the axon enters the LDT prothoracic ganglion in the ventromedial quadrant of the and produces several ventrally projecting and laterally pro- promesothoracic connective (Fig. 1F). jecting branches along its length (Fig. 3H). These branches
The branching pattern of DN1 in the mesothoracic gan- are primarily confined to the dorsal half of the hemigan- glion strongly resembles that in the prothoracic ganglion glion and do not appear to enter any of the major longitu- (Fig. lA,G). Initially, the descending axon enters the VIT dinal tracts (Fig. 31-K). The axon projects from the ganglion (Fig. 1H). Further posteriorly, the axon leaves that tract in the ventromedial quadrant of the mesometathoracic con- and enters the ring tract (RT), where it extends branches nective (Fig. 3L).
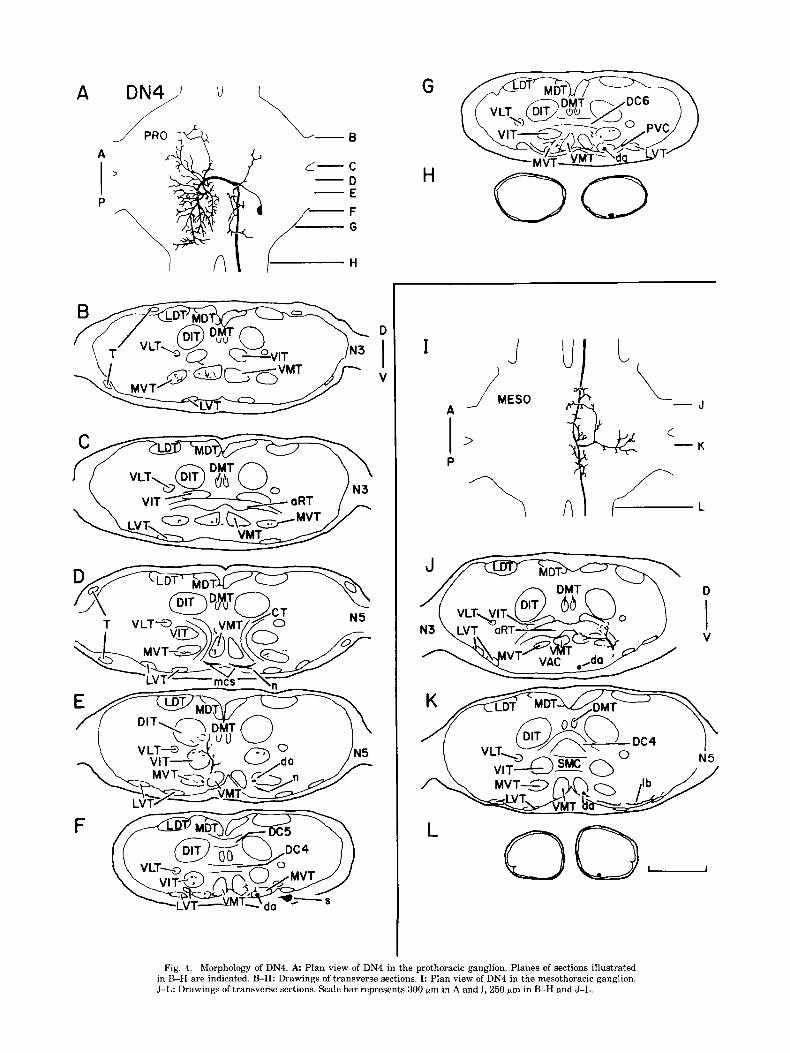
DN4 into the RT on both sides of the ganglion (Fig. 11,J). The axon leaves the RT and rejoins the VIT, from which yet another branch crosses the ganglionic midline in the SMC The soma of DN4 is found posteriorly on the ventral (Fig. 1K). The axon exits the ganglion and occupies a ven- surface of the ganglion (Fig. 4A,F). A transverse segment tromedial position in the mesometathoracic connective crosses the ganglionic midline at a point ventral to the (Fig. 1L). ventral median tracts (VMTs), at the level of the C-tracts
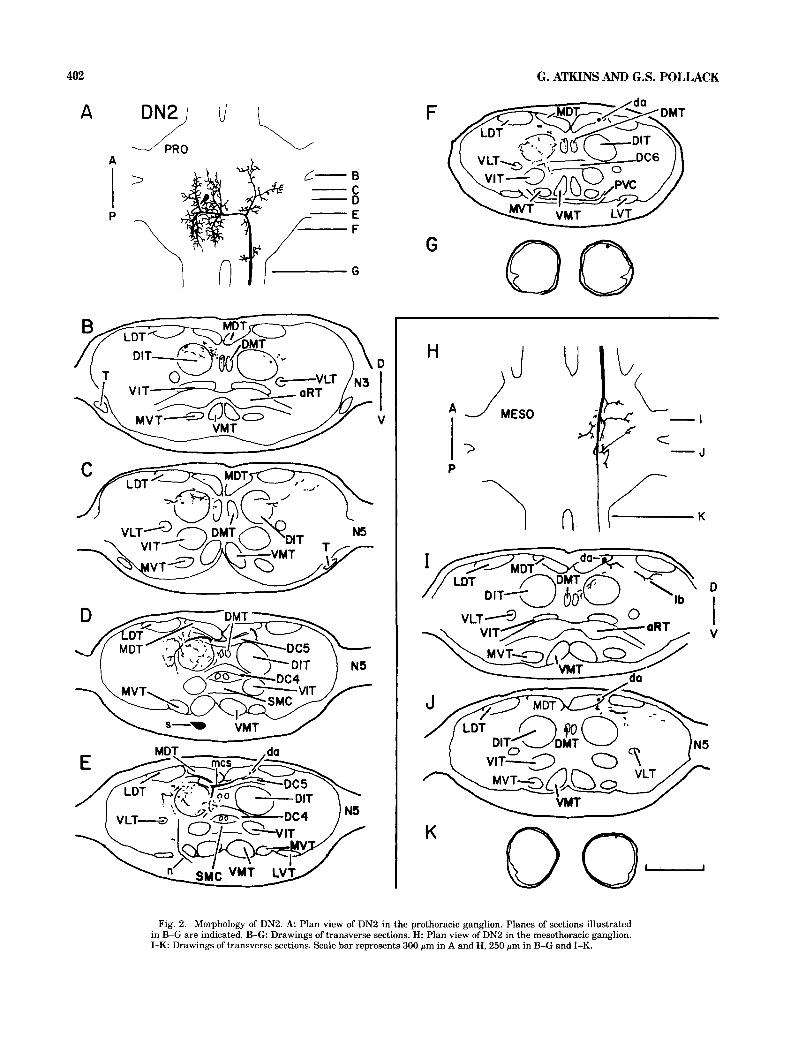
(CTs), and gives rise to several branches (Fig. 4A,D). Ante- riorly projecting branches extend into the MVT and the DN2
The soma of DN2 is positioned ventro-medially in the VMT (Fig. 4B,C), posteriorly projecting branches project prothoracic ganglion (Fig. 2A,D). A neurite extends lat- into and between the VIT, the MVT, and the VMT (Fig. erally from the soma, passes between the medial ventral 4F,G), while a large dorsally projecting branch extends tract (MVT) and the lateral ventral tract (LVT), and projects processes into the VIT and the DIT (Fig. 4E). A neurite posteriorly toward the dorsal intermediate tract (DIT) (Fig. extends anteromedially from the soma and connects to the 2E). The neurite connects to a dorsal segment that crosses neuropile segment near the ganglionic midline (Fig. 4A,D- the ganglionic midline through dorsal commissure 5 (DC5) F). A descending axon originates from the soma-ipsilateral (Fig. 2D,E). Four processes, two anteriorly projecting and side of the neuropile segment (Fig. 4A,D) and produces two posteriorly projecting, arise from the soma-ipsilateral anteriorly projecting processes that extend into the MVT portion of the dorsal segment (Fig. 2A). These processes, (Fig. 4C,D) and posteriorly projecting processes that project which are in the DIT, produce numerous branches that into and between the MVT, the VMT, and the VIT (Fig. invade the DIT, the dorsal medial tract (DMT), and regions 4F,G), The axon leaves the ganglion and assumes an ex- between the DIT and the medial dorsal tract (MDT) (Fig. tremely ventral position in the promesothoracic connective 2B-F). The axon of DN2 arises from the soma-contralateral (Fig. 4H). side of the dorsal segment (Fig. 2A), bends posteriorly, and The axon of DN4 enters the mesothoracic ganglion and enters the MDT (Fig. 2E,F). A large anteriorly projecting courses posteriorly through the ventral region of the ven- branch arises from the axon at the point where it bends tral association center WAC) (Fig. 45). At the posterior posteriorly (Fig. 2A) and extends processes dorsolaterally border of the VAC the axon enters the VMT (Fig. 4K). It (Fig. 2B,C). The descending axon increases in diameter and produces several small branches along its length as well as produces a few small dorsal processes before entering the one large, laterally projecting process (Fig. 41), which ex- promesothoracic connective, where it maintains a dorsal tends to a position ventral to the MVT and then bifurcates, and slightly lateral position (Fig. 2G). one branch extending further laterally to the LVTs (Fig.
The mesothoracic projection of DN2 (Fig. H-K) is re- 4K), and the other branch projecting anterodorsally into stricted to the dorsal half of the hemiganglion. As the axon the RT and the VIT (Fig. 45). The axon projects out of the passes through the ganglion in the MDT, its diameter de- ganglion through the mesometathoracic connective, where creases steadily (Fig. 2H-K). The axon supports several it assumes a ventromedial position (Fig. 4L).
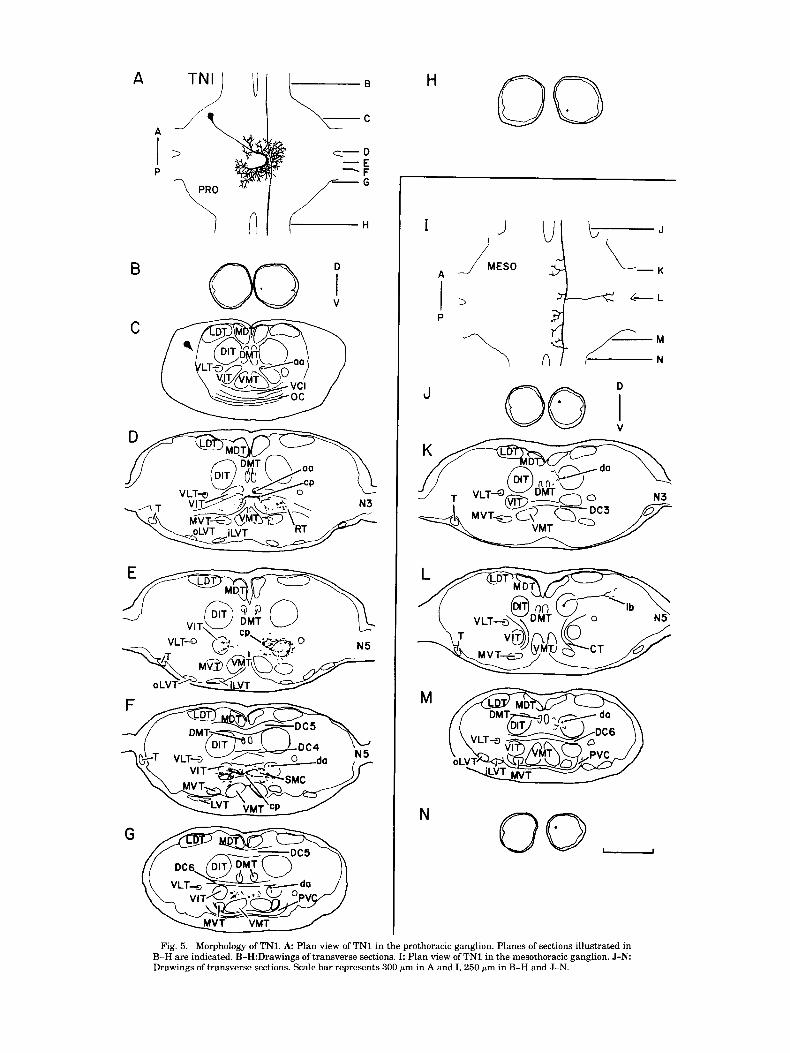
TN1 branches that project laterally past the lateral dorsal tract (LDT) and ventromedially to the DIT (Fig. 21,J). The axon leaves the ganglion via the mesometathoracic connective, The dorsal, lateral soma of TN1 is connected to a large maintaining a dorsal position (Fig. 2K). C-shaped neuropile segment by a thin, branchless neurite
401
G. ATKINS AND G.S. POLLACK 402
A F
Fig. 2. Morphology of DN2. A Plan view of DN2 in the prothoracic ganglion. Planes of sections illustrated in B-G are indicated. B-G Drawings of transverse sections. H: Plan view of DN2 in the mesothoracic ganglion. I-K: Drawings of transverse sections. Scale bar represents 300 pm in A and H, 250 pm in B-G and I-K.
A DN3) u I
I A II G
F da
G 00 H U
/ MESO A
P
I D
I V
Fig. 3. Morphology of DN3. A Plan view of DN3 in the prothoracic ganglion. Planes of sections illustrated in B-G are indicated. B-G Drawings of transverse sections. H: Plan view of DN3 in the mesothoracic ganglion. I-L: Drawings of transverse sections. Scale bar represents 300 p m in A and H, 250 pm in B-G and ILL.
A
A
I > /
P
c- C D E F
- -
/-. H I
F
D
V
H
I
P
1\
Fig. 4. Morphology of DN4. A Plan view of DN4 in the prothoracic ganglion. Planes of sections illustrated in B-H are indicated. B-H: Drawings of transverse sections. I: Plan view of DN4 in the mesothoracic ganglion. J-L: Drawings of transverse sections. Scale bar represents 300 pm in A and I, 250 pm in B-H and J-L.
D
I B
V
00 H
M
Fig. 5. Morphology of TNI. A Plan view of TN1 in the prothoracic ganglion. Planes of sections illustrated in B-H are indicated. B-H:Drawings of transverse sections. I: Plan view of TN1 in the mesothoracic ganglion. J -N Drawings of transverse sections. Scale bar represents 300 pm in A and I, 250 pm in B-H and J-N.
406
A
B
G. ATKINS AND G.S. POLLACK
I
A
P
L
M n
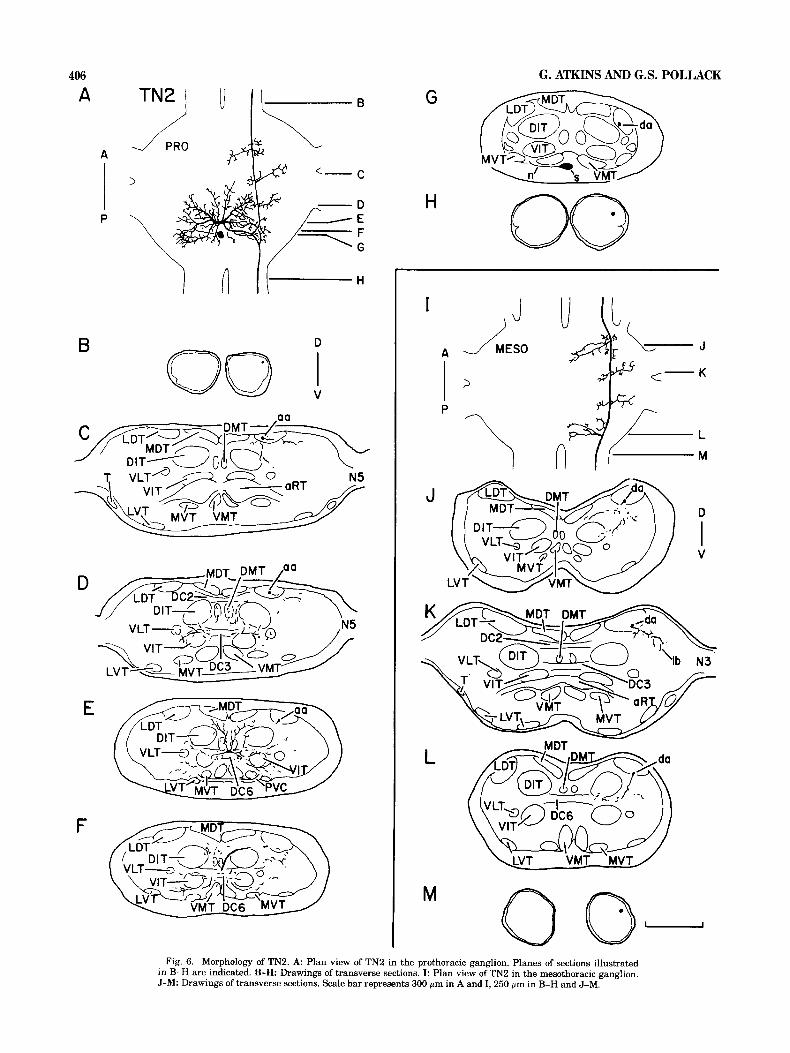
Fig. 6. Morphology of TN2. A Plan view of TN2 in the prothoracic ganglion. Planes of sections illustrated in B-H are indicated. B-H Drawings of transverse sections. I: Plan view of TN2 in the mesothoracic ganglion. J-M: Drawings of transverse sections. Scale bar represents 300 p m in A and I, 250 pm in B-H and J-M.
SOUND-SENSITIVE INTERNEURONS IN THE CRICKET 407
A
P
B
B
F
D
I Q V
G
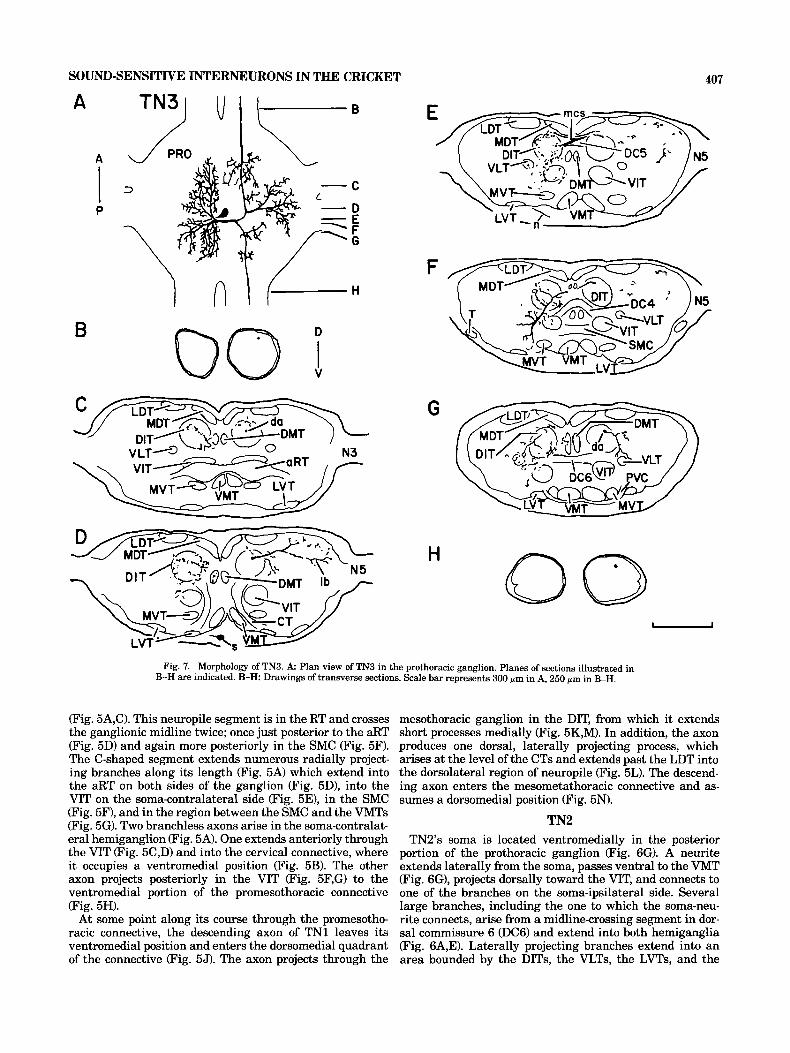
H 0 Fig. 7. Morphology of TN3. A Plan view of TN3 in the prothoracic ganglion. Planes of sections illustrated in
B-H are indicated. B-H: Drawings of transverse sections. Scale bar represents 300 pm in A, 250 sm in B-H.
(Fig. 5A,C). This neuropile segment is in the RT and crosses the ganglionic midline twice; once just posterior to the aRT (Fig. 5D) and again more posteriorly in the SMC (Fig. 5F). The C-shaped segment extends numerous radially project- ing branches along its length (Fig. 5A) which extend into the aRT on both sides of the ganglion (Fig. 5D), into the VIT on the soma-contralateral side (Fig. 5E), in the SMC (Fig. 5F), and in the region between the SMC and the VMTs (Fig. 5G). Two branchless axons arise in the soma-contralat- era1 hemiganglion (Fig. 5A). One extends anteriorly through the VIT (Fig. 5C,D) and into the cervical connective, where it occupies a ventromedial position (Fig. 5B). The other axon projects posteriorly in the VIT (Fig. 5F,G) to the ventromedial portion of the promesothoracic connective (Fig. 5H).
At some point along its course through the promesotho- racic connective, the descending axon of TN1 leaves its ventromedial position and enters the dorsomedial quadrant of the connective (Fig. 54. The axon projects through the
mesothoracic ganglion in the DIT, from which it extends short processes medially (Fig. 5K,M). In addition, the axon produces one dorsal, laterally projecting process, which arises at the level of the CTs and extends past the LDT into the dorsolateral region of neuropile (Fig. 5L). The descend- ing axon enters the mesometathoracic connective and as- sumes a dorsomedial position (Fig. 5N).
TN2 TN2's soma is located ventromedially in the posterior
portion of the prothoracic ganglion (Fig. 6G). A neurite extends laterally from the soma, passes ventral to the VMT (Fig. 6G), projects dorsally toward the VIT, and connects to one of the branches on the soma-ipsilateral side. Several large branches, including the one to which the soma-neu- rite connects, arise from a midline-crossing segment in dor- sal commissure 6 (DC6) and extend into both hemiganglia (Fig. 6A,E). Laterally projecting branches extend into an area bounded by the DITs, the VETS, the LVTs, and the
408
A B
00 E B
C
G. ATKINS AND G.S. POLLACK
F
G
H
n s
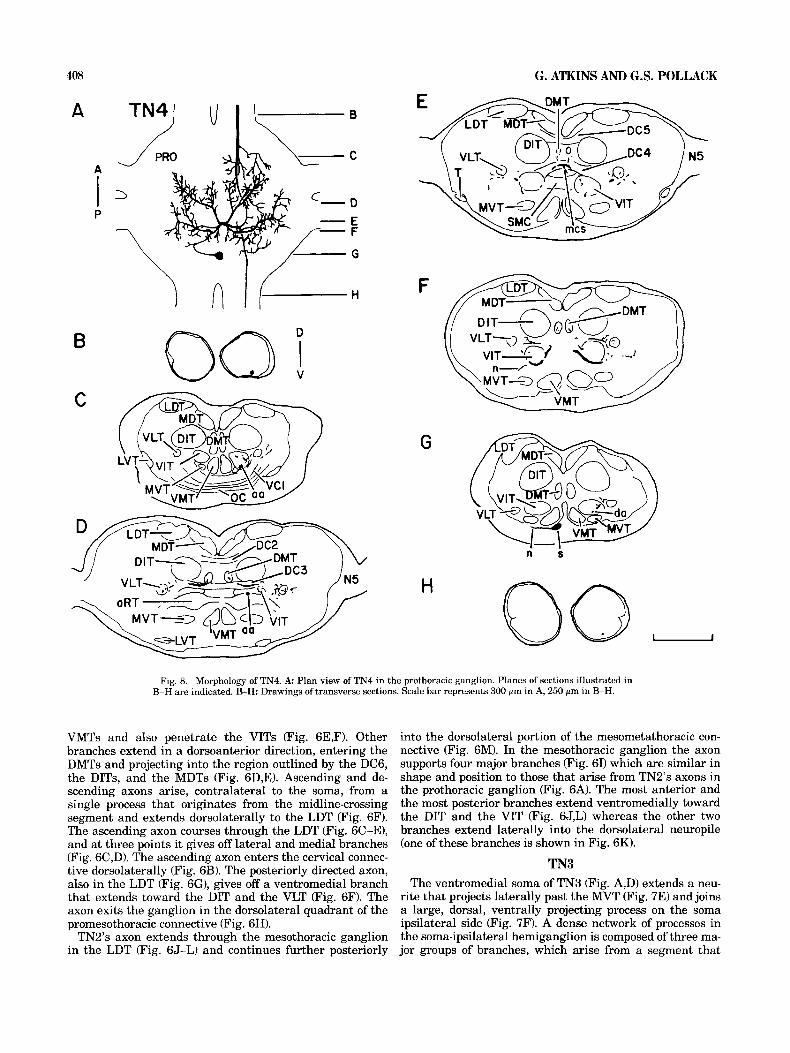
Fig. 8. Morphology of TN4. A: Plan view of TN4 in the prothoracic ganglion. Planes of sections illustrated in B-H are indicated. B-H: Drawings of transverse sections. Scale bar represents 300 pm in A, 250 pm in B-H.
VMTs and also penetrate the VITs (Fig. 6E,F). Other branches extend in a dorsoanterior direction, entering the DMTs and projecting into the region outlined by the DC6, the DITs, and the MDTs (Fig. 6D,E). Ascending and de- scending axons arise, contralateral to the soma, from a single process that originates from the midline-crossing segment and extends dorsolaterally to the LDT (Fig. 6F). The ascending axon courses through the LDT (Fig. 6C-E), and at three points it gives off lateral and medial branches (Fig. 6C,D). The ascending axon enters the cervical connec- tive dorsolaterally (Fig. 6B). The posteriorly directed axon, also in the LDT (Fig. 6G), gives off a ventromedial branch that extends toward the DIT and the VLT (Fig. 6F). The axon exits the ganglion in the dorsolateral quadrant of the promesothoracic connective (Fig. 6H).
TN2's axon extends through the mesothoracic ganglion in the LDT (Fig. 6J-L) and continues further posteriorly
into the dorsolateral portion of the mesometathoracic con- nective (Fig. 6M). In the mesothoracic ganglion the axon supports four major branches (Fig. 61) which are similar in shape and position to those that arise from TN2's axons in the prothoracic ganglion (Fig. 6A). The most anterior and the most posterior branches extend ventromedially toward the DIT and the VIT (Fig. 6J,L) whereas the other two branches extend laterally into the dorsolateral neuropile (one of these branches is shown in Fig. 6K).
TN3 The ventromedial soma of TN3 (Fig. A,D) extends a neu-
rite that projects laterally past the MVT (Fig. 7E) and joins a large, dorsal, ventrally projecting process on the soma ipsilateral side (Fig. 7F). A dense network of processes in the soma-ipsilateral hemiganglion is composed of three ma- jor groups of branches, which arise from a segment that
A
oE B
N
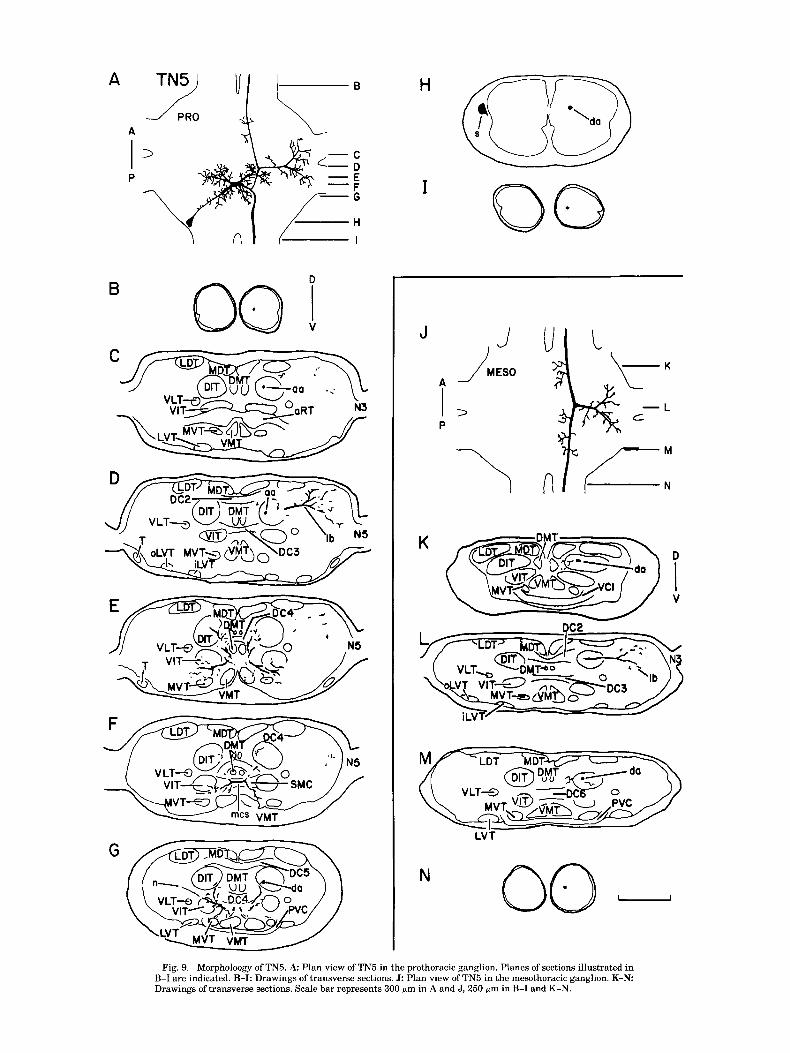
Fig. 9. Morpholoogy of TN5. A: Plan view of TN5 in the prothoracic ganglion. Planes of sections illustrated in B-I are indicated. B-I: Drawings of transverse sections. J: Plan view of TN5 in the mesothoracic ganglion. K-N Drawings of transverse sections. Scale bar represents 300 p m in A and J, 250 pm in B-I and K-N.

410 G. ATKINS AND G.S. POLLACK
spans the ganglionic midline in the DC5 (Fig. 7E). One set of branches extends anteriorly in the DIT and projects pro- cesses in the DIT, the VIT, and the DMT (Fig. 7C,D). An- other projects ventrally through the DIT toward the MVT, while extending branches into the DIT, the VLT, and the VIT (Fig. 7F). The remaining branches course posteriorly into the DIT and extend processes into the DIT, the DMT, the VLT, and the VIT (Fig. 7G). The midline-crossing seg- ment bifurcates on the soma-contralateral side and forms two axons (Fig. 7A). The anteriorly projecting one travels in the DIT (FIG. 7C,D) and enters the dorsolateral quadrant of the cervical connective (Fig. 7B). Several small, dorsally projecting branches originate from the ascending axon (Fig. 7C), as does one large, laterally projecting branch, which extends past the LDT to the periphery of the ganglion (Fig. 7D). The posteriorly projecting axon enters the DIT and produces small branches that project ventrolaterally to the VLT (Fig. 7G). The axon lies in the dorsolateral quadrant of the promesothoracic connective (Fig. 7H). We have not stained the mesothoracic projection of TN3.
TN4 TN4's soma is located ventromedially in the posterior
region of the prothoracic ganglion (Fig. 8A,G). It extends a neurite that travels laterally past the VMT and the MVT (Fig. 8G) and then dorsally to join one of the major processes on the soma-ipsilateral side (Fig. 8A). A short segment crosses the midline in the dorsal commissure 4 (DC4) (Fig. 8E) and gives rise to a posterolateral and anterolateral branch in each hemiganglion (Fig. 8A). The two anterolat- eral branches pass through the lateral portions of the dorsal commissure 3 (DC3) and project processes into and around the VLTs and the DMTs (Fig. 8D). The two posterolateral branches extend ventrally around the VITs and send pro- cesses to the VITs, the VLTs, and the DMTs (Fig. 8F). The two axons, which arise on the soma contralateral side of the midline-crossing segment (Fig. 8A), travel in the VIT (Fig. 8C,D,G). The anteriorly directed axon gives off a few dorsally projecting and laterally projecting processes that extend toward the DIT and VIT at the level of the omega commissure (OC) (Fig. 8C). The axon continues into the ventrolateral quadrant of the cervical connective (Fig. 8s). The posteriorly projecting axon gives off a few small branches that extend to the VLT (Fig. 8G) before entering, ventrolaterally, the promesothoracic connective (Fig. 8H). The mesothoracic projection of TN4 has not been examined.
TN5 TN5's soma is found dorsally in the prothoracic ganglion,
lateral to the origin of the promesothoracic connectives (Fig. 9A,H). A neurite extends anteromedially and supports several short processes along its length which project to the VIT, the MVT, and the VMT (Fig. 9A,G). A short segment crosses the ganglion midline in the SMC (Fig. 9F) and supports several large processes as well as the neurite that extends from the soma (Fig. 9A). In the soma-contralateral hemiganglion, the midline-crossing segment gives rise to a posterolateral branch that sends processes toward the VIT, the VMT, and the MVT (Fig. 9G). The middle-crossing seg- ment also sends a branch laterally in each hemiganglion to the VITs and the MVTs. These two branches extend large processes dorsally to the DITs and the DMTs at the level of the DC4 (Fig. 9E). The two soma-contralateral axons, which course through the DIT, arise from a process that extends from the midline-crossing segment to the DIT (Fig. 9G). The
ascending axon supports a large, heavily branched process that extends dorsolaterally past the LDT (Fig. 9D). As the axon projects further anteriorly it enters the cervical con- nective and maintains a medial position (Fig. 9B). The descending axon does not produce any branches in the ganglion, and exits in the ventromedial quadrant of the promesothoracic connective (Fig. 91).
In the mesothoracic ganglion the axon enters the DIT (Fig. 9K-M) and gives off several short processes along its length, which penetrate the DIT and extend medially to- ward the DMT (Fig. 9K,M). In addition, the axon supports a large branch that terminates in the dorsolateral portion of the ganglion (Fig. 9L) and resembles, in shape and posi- tion, the dorsolateral branch observed in the prothoracic ganglion (Fig. 9J,L). The axon leaves the ganglion in the mesometathoracic connective (Fig. 9N).
DISCUSSION Sources of input
Auditory receptor cells terminate in the ventral portion of the aRT of the prothoracic ganglion (Wohlers and Huber, '85). Wohlers and Huber ('85) described the morphology of six auditory interneurons in G. cumpestris (ONl,ON2, AN1, AN2, DN1, and TN1) and found that each of these has processes in the aRT. Accordingly, each of these interneu- rons is potentially directly postsynaptic to receptor termi- nals. Of the nine interneurons described in the present paper, only DN1 and TN1 have processes that penetrate the aRT (Figs. 1C,D, 5D); thus these two cells are the only ones described here whose morphology is consistent with their being first-order interneurons. Intracellular record- ings show that the remaining cells do receive synaptic input in the prothoracic ganglion (Atkins and Pollack, '87); however, this must be from other interneurons.
Correlations between spectral sensitivity and morphology
The neurons described in this paper fall into two classes on the basis of spectral sensitivity; high-frequency neurons and low-frequency neurons (Atkins and Pollack, '87) (Table 1). Several correlations exist between neuronal structure and spectral sensitivity.
Neuropile processes. The neuropile processes of low-fre- quency and high-frequency cells are found in different re- gions of the prothoracic ganglion. Processes of high- frequency cells (DN2, DN3, TN2, TN3, and TN4) are lo- cated primarily in the dorsal regions of the ganglion (Fig. 10A). Conversely, processes of low-frequency neurons (DN1, DN4, TN1, and TN5) are primarily ventral (Fig. 10B). Con- sistent with this, high-frequency neurons cross the gangli- onic midline in dorsal commissures, while low-frequency cells do so more ventrally (Table 1). One possible interpre- tation of this spatial distinction between low- and high- frequency cells is that they receive input in different re- gions of the ganglion. Many of the processes of these cells in the prothorax are smooth in texture and may be sites of input (Peters et al., '86). The major exception to the dorso- ventral distribution of processes is the prominent dorsolat- eral projection of TN5, a low-frequency cell. This branch, which projects to a region of the ganglion known to contain dendrites of motor neurons (Kein, '80; Honegger et al., '84; Laurent and Richard, '861, has numerous varicosities and may be an output process (Peters et al., '86). (It must be noted, however, that light-microscope observations alone

SOUND-SENSITIVE INTERNEURONS IN THE CRICKET 411
LDT MDT DMl
, I MVT VMT
LDT MDT DMT
DN2
DN3
TN2
TN3
TN4
DN1
DN4
TN1
TN5
MVT VMT
Fig. 10. Topographic relationships of neuropile segments of low-fre- quency and high-frequency cells. Drawings showing the extent and position of the neurons in the transverse plane of the prothoracic ganglion (dorsal side uppermost) for (A) high-frequency neurons and (B) low-frequency neu- rons. Outlined areas do not include the soma on the branchless portion of the primary neurite.
cannot reliably distinguish between input and output sites; Watson and Burrows, '83; Peters et al., '86.) Our results suggest, therefore, that low-frequency cells and high-fre- quency cells receive input in different regions of the gan- glion; i.e., that their input processes show a crude tonotopic organization.
Low and high frequencies are involved in different behav- iors; the former are used for intraspecific signals that me- diate pair formation and courtship and elicit positive phonotactic responses, while the later are produced by pred- ators and elicit negative phonotactic responses Woiseff et al., '78). The present report shows that this behavioral dichotomy has a morphological correlate.
D
v 0 High frequency neurons 0 Low frequency neurons
Fig. 11. Schematic diagram showing the position of the descending axons of the interneurons in the right promesothoracic connective. Not drawn to scale.
Romer ('85) showed that the anatomy of three identified auditory neurons of tettigoniids was related to their spec- tral sensitivity. All of these cells are thought to be directly postsynaptic to auditory receptors. Receptors project to the central nervous system in a tonotopically organized man- ner (Oldfield, '83; Romer, '83, '%), and these first-order interneurons have dendritic branches in those regions of the receptor projection that encode the frequencies to which the interneurons respond (Romer, '85). The dorsoventral distribution of processes described in the present paper differs from the tettigoniid example because the interneu- rons described here are, with two possible exceptions, not directly postsynaptic to receptors. Their morphology, there- fore, is not constrained by the receptor projection. Our find- ings show, for the first time in insects, that tonotopy exists at levels of neural circuitry removed from the receptor projection.
Axon position. The posteriorly projecting axons of low- frequency neurons travel in the medial portion of the con- nective, whereas those of high-frequency neurons are lo- cated more laterally (Fig. ll). Preliminary data from 13 other interneurons (five low-frequency cells, eight high- frequency cells) are consistent with this (Atkins, unpub- lished results). It seems unlikely that this distinction be- tween the two classes of cells carries any functional significance. It may, rather, be a consequence of the gangli- onic regions within which the axons arise. Axons originat- ing in dorsal regions of the ganglion (those of high-frequency cells) enter dorsal tracts which, upon entering the intergan- glionic connective, assume dorsal and lateral positions; ax- ons that originate ventrally (those of low-frequency cells) join ventral tracts, which come to lie ventrally and medially in the connective (see, e.g., Fig. 3F).
Once the axons enter the mesothoracic ganglion, the spa- tial separation of high- and low-frequency neurons is less clear. All three high-frequency cells whose mesothoracic projections have been stained (DN2, DN3, TN2) have dor- solateral branches, which bear numerous varicosities, and may be outputs (Figs. 2H-J, 3H-K, 61-L). Two of the low- frequency cells (TN1 and TN5) have dorsal branches simi- lar to the high-frequency cells (Figs. 5I,L, 9J,L). The other two (DN1, DN4) branch more ventrally in the mesothoracic ganglion (Figs. 1G-L, 41-L).
Inhibitory-type morphology. In the locust, Pearson and Robertson ('87) identified three anatomical features which, when occurring together, are diagnostic of inhibitory inter- neurons: a ventromedial cell body, a laterally bowed neu- rite, and a soma-contralateral interganglionic axon. These features are shared by 12 identified inhibitory interneurons in locust and by none of 19 identified excitatory interneu- rons. The inhibitory-type morphology is shown by four of the five high-frequency cells described here (DN2, TN2, TN3, TN4) (Fig. 12). The fifth, DN3, has a ventromedial soma and laterally bowed neurite, but its axon is ipsilateral to the soma (Figs. 3, 12). In contrast, none of the low- frequency cells have either a ventromedial soma or a lat- erally bowed neurite, though three do have soma-contralat- era1 axons (Fig. 12). If the correlation noted by Pearson and Robertson ('87) in locusts is also valid for crickets, then at least four of the five high-frequency cells are likely to be inhibitory. Although all identified interneurons that share these three anatomical features are inhibitory, the converse is not true; inhibitory neurons exist which lack one or more of these traits (Pearson and Robertson, '87). Therefore, the

412 G. ATKINS AND G.S. POLLACK
Low frequency High frequency neurons neurons
I= Soma-ipsilateral axon
Fig. 12. Summary diagram showing the positions of the somata and the paths of neurites in the prothoracic ganglion. Cells labelled I have soma- ipsilateral axons; the others have soma-contralateral axons.
fact that low-frequency cells do not express all three fea- tures is not a strong predictor that these cells are excitatory.
Possible targets DN2, DN3, TN2, TN3, and TN5 have branches in the
dorsolateral region of the prothoracic ganglion (Figs. 2C- D, 3C, 6D, 7D, 9D), which also contains the processes of neurons that control muscles of the neck and prothoracic legs (Kein, '80; Honneger et al., '84; Laurent and Richard, '86). DN2, DN3, TN1, TN2, and TN5 (and possibly others whose processes we have not traced past the promesothor- acic connectives) branch in the homologous region of the mesothoracic ganglion (21, 31-K, 5L, 6K, 9L), where neu- rons associated with the wings and mesothoracic legs are found (Elepfandt, '80; Furukawa et al., '83; Robertson, '87; Wang and Robertson, '87). The seven cells that have been stained as far as the mesothoracic ganglion all continue further posteriorly and may send branches to motor areas of lower ganglia as well. It is thus possible (though we have no direct evidence for this) that the cells described here play a role in motor responses to sound.
LITERATURE CITED Atkins, G., and G.S. Pollack (1987) Response properties of prothoracic,
interganglionic, sound-activated interneurons in the cricket, Teleogryl- lus oceanicus. J. Comp. Physiol. 161r681-693.
Elepfandt, A. (1980) Morphology and output coupling of wing muscle motor- neurons in the field cricket (Gryllidae, Orthoptera). Zool. Jahrb. Physiol.
84:26-45. Furukawa, N., K. Tomioka, and T. Yamaguchi (1983) Functional anatomy
of the musculature and innervation of the neck and thorax in the cricket Gryllus bimaculatus. Zool. Mag. 92:371-385.
Honegger, H.W., J.S. Altman, J. Kien, R. Muller-Tautz, and E. Pollerberg (1984) A comparative study of neck muscle motor neurons in a cricket and a locust. J. Comp. Neurol. 230:517-535.
Kien, J. (1980) Morphology of locust neck muscle motorneurons and some of their inputs. J. Comp. Physiol. 140t321-336.
Laurent, G., and D. Richard (1986) The organization and role during loco- motion of the proximal musculature of the cricket foreleg. 1. Anatomy and innervation. J. Exp. Biol. 123255-283.
Lorente de NO, R. (1937) The sensory endings in the cochlea. Trans. Am. Otol. SOC. 27:86-90.
Merzenich, M.M., P.L. Knight, and G.L. Roth (1975) Representation of cochlea within primary auditory cortex in the cat. J. Neurophysiol. 38:231-249.
Moiseff, A., G.S. Pollack, and R.R. Hoy (1978) Steering responses of flying crickets to sound and ultrasound Mate attraction and predator avoid- ance. Roc. Natl. Acad. Sci. USA 75:4052-4056.
Oldfield, B.P. (1982) Tonotopic organization of auditory receptors in Tetti- goniidae (Orthoptera: Ensifera). J. Comp. Physiol. 147t461-469.
Oldfield, B.P. (1983) Central projections of primary auditory fibers in Tetti- goniidae (Orthoptera: Ensifera). J. Comp. Physiol. 151:389-395.
Oldfield, B.P. (1984) Physiology of auditory receptors in two species of Tettigoniidae (Orthoptera: Ensifera). Alternative tonotopic organizations of the auditory organ. J. Comp. Physiol. 155t689-696.
Oldfield, B.P., H.U. Kleindienst, and F. Huber (1986) Physiology and ton- topic organization of auditory receptors in the cricket Gryllus bimacu- latus DeGeer. J. Comp. Physiol. I59:457-464.
Pearson, K.G., and R.M. Robertson (1987) Structure predicts synaptic func- tion of two classes of interneurons in the thoracic ganglia of Locusta migratoria. Cell Tissue Res. 250:105-114.
Peters, B.H., H. Romer, and V. Marquart (1986) Spatial segregation of synaptic inputs and outputs in a locust auditory interneurone. J. Comp. Neurol. 254:34-50.
Robertson, R.M. (1987) Interneurons in the flight system of the cricket Teleogryllus oceanicus. J. Comp. Physiol. 160:431-445.
Romer, H. (1983) Tonotopic organization of the auditory neuropile in the bushcricket Tettigonia uiridissima Nature 306:60-62.
Romer, H. (1985) Anatomical representation of frequency and intensity in the auditory system of orthoptera. In K. Kalmring and N. Elsner (eds): Acoustic and Vibration Communication in Insects. Berlin: Paul Parey, pp. 25-32.
Spurr, A.R. (1969) A low-viscosity epoxy resin embedding medium for elec- tron microscopy. J. Ultrastruct. Res. 2631-43.
Tyrer, N.M., and G.E. Gregory (1982) A guide to the neuroanatomy of locust suboesophageal and thoracic ganglia. Philos. Trans. R. SOC. Lond. [Biol.] 297:91-123.
von Bekesy, G. (1960) Experiments in Hearing. New York: McGraw-Hill. Wang, S., and R.M. Robertson (1987) Wing muscle motorneurons and flight
motor patterns of a cricket. Bull. Can. Soc. Zool. 18t44. Watson, A.D.H., and M. Burrows (1983) The morphology, ultrastructure,
and distribution of synapses on an intersegmental interneurone of the locust. J. Comp. Neurol. 214:154-169.
Wohlers, D.W., and F. Huber (1982) Processing of sound signals by six types of neurons in the prothoracic ganglion of the cricket, Gryllus campestris L. J. Comp. Physiol. 146161-173.
Wohlers, D.W., and F. Huber (1985) Topographical organization of the audi- tory pathway within the prothoracic ganglion of the cricket Gryllus campestris L. Cell Tissue Res. 239r555-565.
Woolsey, C.N., and E.M. Walzl(1942) Topical projections of nerve fibers from local regions of the cochlea to the cerebral cortex of the cat. Bull. Johns Hopkins Hosp. 71:315-344.