crl7smu1 e3 ligase complex driven h2b ubiquitination ... · chromatid cohesion so far. in this...
TRANSCRIPT

© 2018. Published by The Company of Biologists Ltd.
CRL7SMU1 E3 ligase complex driven H2B ubiquitination functions in sister
chromatid cohesion by regulating SMC1 expression
Varun Jayeshkumar Shah1, 2, & Subbareddy Maddika1*
1Laboratory of Cell Death & Cell Survival, Centre for DNA Fingerprinting and Diagnostics,
Hyderabad, INDIA
2Graduate studies, Manipal Academy of Higher Education, Manipal, INDIA
*To whom correspondence should be addressed
Dr. Subbareddy Maddika
Centre for DNA Fingerprinting and Diagnostics, Nampally, Hyderabad-500 001, INDIA
Tel: +91-40-24749353
Fax: +91-40-24749448
E-mail: [email protected]
Key words: E3 ligase, SMU1, H2B ubiquitination, RNF40, CUL7, DDB1, mitosis, chromatid
cohesion
Summary statement
We identified that SMU1, DDB1, CUL7 and RNF40 assemble an E3 ligase complex and
promotes H2B monoubiquitination to drive SMC1 expression, which is essential for sister
chromatid cohesion.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt
JCS Advance Online Article. Posted on 5 March 2018

Abstract
Cullin–RING E3 ligases (CRLs) control broad range of biological processes by ubiquitinating
numerous cellular substrates. However, role of CRL E3 ligases in chromatid cohesion is
unknown. In this study, we identified a new CRL type E3 ligase (designated as CRL7SMU1
complex) that has an essential role in maintenance of chromatid cohesion. We demonstrate that
SMU1, DDB1, CUL7 and RNF40 as integral components of this complex. SMU1 by acting as a
substrate recognition module, binds to H2B and mediates monoubiquitination at K120 site
through CRL7SMU1 E3 ligase complex. Depletion of CRL7SMU1 leads to loss of H2B
ubiquitination at SMC1a locus and thus subsequently compromised SMC1a expression in cells.
Knock down of CRL7SMU1 components or loss of H2B ubiquitination leads to defective sister
chromatid cohesion, which is rescued by restoration of SMC1a expression. Together, our results
unveil an important role of CRL7SMU1 E3 ligase in promoting H2B ubiquitination for
maintenance of sister chromatid cohesion during mitosis.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Introduction
Ubiquitination is an essential posttranslational modification that regulates wide array of cellular
processes in eukaryotes (Hershko and Ciechanover, 1998). Ubiquitin is covalently attached
through its C-terminal glycine to the ε-amino group of lysine or occasionally to the N-terminus
of the protein substrates. Proteins can be tagged with either single ubiquitin or with a
polyubiquitin chain. Linkage of ubiquitin with substrates occurs in a three-enzyme cascade
catalyzed by E1 (ubiquitin-activating enzyme), E2 (ubiquitin-conjugating enzyme), and E3
(ubiquitin ligase) enzymes. E3 ligases are the most heterogeneous class of enzymes, which bring
together the correct E2 with the right substrate and thus are critical for defining substrate
specificity during ubiquitination process (Berndsen and Wolberger, 2014; Hershko and
Ciechanover, 1998).
Depending on the domain architecture and on the mechanism of ubiquitin transfer to the
substrate, E3 ligases have been classified into three types: HECT E3s, RING E3s and RBR type
E3s. The HECT-type E3 ligases characterized by the presence of HECT (homologous to E6-AP
carboxyl terminus) domain possess intrinsic catalytic activity. HECT-E3s load ubiquitin on
themselves through formation of ubiquitin-thioester intermediate with the catalytic cysteine
residue within the HECT domain and then transfers the ubiquitin to the target protein (Berndsen
and Wolberger, 2014; Rotin and Kumar, 2009). HECT-E3s predominantly function as
monomeric enzymes, but existence of functional multimeric HECT E3s in the cell are recently
emerging (Maddika and Chen, 2009). For example, HECT-E3 EDD forms a multi-component E3
ligase with DDB1, DYRK2 and VPRBP as core components to regulate G2/M transition. RING-
class of E3s characterized by the presence of RING (Really Interesting New Gene) or U-box
domain are the most abundant type of ubiquitin ligases in the cell (Berndsen and Wolberger,
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

2014). RING-type E3s do not have intrinsic catalytic activity however, they function as scaffold
between E2 and the substrate and thereby mediate direct transfer of ubiquitin to the substrate.
RING E3s can function as monomers, homodimers, heterodimers and multi-protein complexes,
such as the cullin-RING ligases (CRLs) (Francis et al., 2013; Lydeard et al., 2013; Petroski and
Deshaies, 2005; Zimmerman et al., 2010). On the other hand, the RBR-type E3 ligases share
common features with both RING and HECT E3 ligase families. They recruit ubiquitin charged
E2 through the RING domain but, like HECT-E3s, catalyze the ubiquitin transfer through two-
step mechanism where first transfer occur on to E3 itself and later to the substrate (Spratt et al.,
2014).
CRLs [Cullin–RING E3 ubiquitin ligases] are a superfamily of RING E3s responsible for
as much as 20% of ubiquitin-dependent protein modification in cells (Bennett et al., 2010;
Petroski and Deshaies, 2005). Cullin based E3 ligases assemble into multimeric complexes to
enhance substrate diversity during ubiquitination. CRLs consist of two distinct modules: 1) a
substrate-targeting unit composed of a substrate-recognition protein and an adaptor protein that
links the module to the cullin, and 2) the RING component that is active in recruiting an
ubiquitin charged E2 enzyme(Zimmerman et al., 2010). CRLs ubiquitinate several cellular
substrates and thus control broad range of biological processes, including cell growth,
development, signal transduction, transcriptional control and tumor suppression (Harper and
Tan, 2012; Petroski and Deshaies, 2005). However, no CRL E3 ligase is implicated in sister
chromatid cohesion so far. In this study, we identified a new CRL type E3 ligase (CRL7SMU1
complex with SMU1-DDB1-CUL7 and RNF40 as core components) that has an essential role in
maintenance of chromatid cohesion. We demonstrated that CRL7SMU1 E3 ligase supports sister
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

chromatid cohesion through H2B ubiquitination at SMC1a locus and thereby controlling its
expression.
Results
Identification of CRL7SMU1 E3 ligase complex
Previous studies have demonstrated that DCAF1/VprBP, a protein with WD repeat region and
LisH domain acts as a substrate recognition component of HECT-type as well as RING-based E3
ubiquitin ligase complexes (Maddika and Chen, 2009; Nakagawa et al., 2013). To identify
proteins with a similar combination of LisH and WD repeat organization that may assemble E3
ligase complexes, we performed a global search for LisH domain (ID: PS50896) using UniProt
database. We retrieved 28 human proteins that contain LisH domain of which nine of them
exhibit a combination of LisH domain with WD repeats in their architecture (Figure 1A). To test
the possibility if these proteins assemble E3 ligase complexes, we isolated protein complexes
associated with SMU1, one of the listed proteins with such a domain organization. SMU1
(suppressor of mec-8 and unc-52) is a spliceosome accessory protein known for its role in DNA
replication, mitotic spindle assembly and maintenance of genomic stability (Ren et al., 2013;
Sugaya et al., 2006; Sugaya et al., 2005). Tandem affinity purification of SBP-Flag-S peptide
tagged (SFB)- SMU1 followed by mass spectrometric analysis revealed several known and
unknown interacting partners (Figure 1B). Interestingly, we found DDB1, CUL7 and RNF40 in
the list of SMU1 interacting proteins. CUL7 is a member of the Cullin family of proteins that
function as scaffold for E3 ubiquitin ligases. Like other cullins, CUL7 also assembles CRL by
associating with Skp1, Fbx29, and ROC1 (Dias et al., 2002). On the other hand DDB1 (Damaged
DNA-binding protein 1) along with DDB2 (damaged DNA binding protein 2) is known to
function in nucleotide excision repair. At the molecular level, DDB1 functions as an adaptor
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

protein for Cul4 ubiquitin E3 ligase complex and DYRK2-EDVP complex to regulate ubiquitin-
dependent degradation of various substrates like Cdt1, c-Jun, p21 and katanin p60 (Iovine et al.,
2011). RNF40 is a RING type of E3 ligase which heterodimerize with RNF20 to
monoubiquitinate histone H2B (Kim et al., 2009). As it is well known that CUL7 acts as a
scaffolding protein and DDB1 as adaptor for various E3 ligase complexes and further RNF40
being an E3 ligase, we thus propose a new CRL7 E3 ligase complex with SMU1 as substrate
recognition component.
By performing immunoprecipation using SMU1 antibody, we validated the endogenous
association of RNF40, DDB1 and CUL7 with SMU1 (Figure 1C). Importantly, DYRK2, a
scaffolding subunit of EDVP E3 ligase complex, do not interact with SMU1 suggesting that
CRL7SMU1 is a distinct complex. Further, exogenously expressed SMU1 efficiently interacted
with CUL7, RNF40 and DDB1 but not with DYRK2 (Figure S1A). Likewise, we found no
interaction of SMU1 with VPRBP (Figure 1D), which was known functional subunit of DYRK2-
EDVP complex as well as CUL4-DDB1 complex, again supporting our conclusion that
CRL7SMU1 is a discrete E3 ligase complex in cells. In fact, SMU1 is not part of any known
Cullin-RING based E3 ligase complexes since SMU1 neither interacts with SKP1 (Figure 1E)
nor ROC1 (Figure 1F), while ROC1 and SKP1 interacted positively. While DDB1 can associate
with CUL4A as well as CUL7 along with SMU1 and RNF40 (Figure S1B), we found no
interaction of CUL7 and RNF40 with CUL4A (Figure S1C & S1D), again supporting CRL7SMU1
as an independent complex. Previously, RNF40 was known to act as functional E3 ligase in
RNF40/RNF20 heterodimeric form. Surprisingly, however, we found that SMU1 does not
interact with RNF20 (Figure 1G), implying that SMU1-RNF40 E3 ligase complex is formed
independent of RNF20.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Furthermore, to investigate whether SMU1 forms a stable E3 ligase complex in vivo, we
analyzed these proteins in 293T cell extracts by using size-exclusion chromatography. SMU1,
DDB1, CUL7 and RNF40 were co-eluted in similar cell lysate fractions corresponding to a
molecular mass of approximately 440 kDa (Figure 1H), suggesting that CRL7SMU1 complex
proteins physically interact with each other and assemble a large complex in cells. WD repeats
are well established to assist in the assembly of E3 ligase complexes such as DDB1-CRLs. To
delineate the regions of SMU1 required for assembly of the E3 ligase complex, we made deletion
constructs of SMU1 that either lack WD repeat region or the N-terminal LisH domain-containing
region (Figure 1I). Interestingly, we found that that while WD repeat region is dispensable for
association with components of E3 ligase, LisH-domain region is required for assembly of E3
ligase complex (Figure 1J). Collectively, we demonstrate that SMU1 via its LisH domain region
assembles a novel E3 ligase complex by associating with DDB1, CUL7 and RNF40 in cells.
CRL7SMU1 complex regulates mono-ubiquitination of H2B at K120 site.
In search of substrate(s) of newly identified E3 ligase complex, we next examined the mass spec
data derived from SMU1 purification. We found Histone 2B (H2B) as one of the interacting
partners of SMU1. H2B along with H2A, H3 and H4 is a part of core nucleosome and is a well-
known substrate of RNF20/40 complex, which monoubiquitinates H2B at Lysine 120 site in
humans (K123 in yeast) (Kim et al., 2009; Wood et al., 2003). However, no substrate recognition
subunit for RNF40/RNF20 E3 ligase is reported yet. As we identified RNF40 as a core
component in the CRL7SMU1 complex, we proposed H2B as a probable substrate of this complex.
By performing the co-immunoprecipitation experiments using streptavidin pull down followed
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

by immunoblotting with H2B antibody, we confirmed the specific interaction of H2B with
CUL7, DDB1, RNF40 and SMU1 (Figure 2A). To establish the architecture of the complex and
further test if SMU1 acts as substrate recognition component in the E3 ligase complex, we
performed knockdown experiments. Interaction between RNF40 and H2B is severely hampered
upon depletion of SMU1 where as DDB1 interaction with RNF40 is intact in these cells (Figure
2B). On the other hand, interaction between SMU1-H2B and SMU1-DDB1 was unaffected upon
knockdown of RNF40 (Figure 2C). However, depletion of DDB1 resulted in reduced binding of
SMU1 with CUL7 (Figure 2D). Also, depletion of CUL7 led to loss of RNF40-DDB1 interaction
with no effect on SMU1-DDB1 interaction (Figure 2E). This data suggest that H2B binds to
SMU1, which associates to CUL7 and further RNF40 through DDB1 (Figure 2F). To
substantiate the role of SMU1 as substrate recognition protein of the complex, we next checked
the direct interaction of recombinant bacterially expressed glutathione S-transferase (GST)-
tagged SMU1 (GST-SMU1) with Maltose binding protein (MBP) - tagged H2B (MBP-H2B).
Indeed, we found that SMU1 interacts with H2B directly under in vitro conditions (Figure 2G).
Together, these experiments suggest that SMU1 functions as a substrate recognition component
that links H2B with DDB1-CUL7-RNF40. Next, to examine the role of CRL7SMU1 complex in
histone H2B ubiquitination, we used specific si/shRNA to deplete complex proteins in HeLa
cells. It has been shown that knockdown of RNF20/40 abolishes H2B ubiquitination (Kim et al.,
2009). In agreement with previous studies we found knock down of RNF40 led to
downregulation of H2B monoubiquitination at K120 (Figure 2H). Interestingly, similar to
knockdown of RNF40, depletion of SMU1 (Figure 2I), DDB1 (Figure 2J) and CUL7 (Figure 2K)
also led to significant downregulation of H2B ubiquitination suggesting that these proteins
function together to ubiquitinate H2B K120 in vivo. Further, RNF40 mediated H2B
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

ubiquitination is known to stimulate H3K4 trimethylation via COMPASS complex (Kim et al.,
2009). Similar to RNF40 (Figure 2H), depletion of SMU1, DDB1 and CUL7 resulted in
reduction of H3K4 trimethylation (Figure 2I-2K). DDB1 depletion as well as RNF40 depletion
seems to affect H3K4 trimethylation levels more than H2B ubiquitination. Given that DDB1 is
part of multiple independent E3 ligase complexes, it might be possible that DDB1 regulates
H3K4me3 through additional mechanisms independent of H2B ubiquitination. Infact, it has been
reported that CUL4-DDB1 ubiquitin E3 ligase interacts with multiple WD repeat containing
proteins and regulate H3 methylations including H3 K4Me1, K4Me3, K9Me3 and K27Me3
(Higa et al., 2006). Likewise, RNF40 could also be forming other protein complexes to directly
regulate H3K4me3 independent of H2B ubiquitination. Nonetheless, our data suggest that
SMU1, DDB1, CUL7 and RNF40 are integral components of functional E3 ligase complex that
regulate histone modifications.
Intact CRL7SMU1 complex is required for mitotic progression
Next, we sought to understand the functional role of CRL7SMU1 complex in cells. We observed
that cells depleted with SMU1 (Figure S2A) proliferated at a significantly slower rate than
control cells (Figure S2B). Furthermore, we found that depletion of SMU1 led to the
accumulation of 4N cells as well as polyploid (>4N) cells (Figure 3A). This increase in the 4N
population is due to arrest in the mitotic phase, as time-lapse imaging analysis confirmed that
cells lacking SMU1 spent several hours in mitosis while control siRNA cells took 60 minutes on
an average to complete mitosis (Figure 3B & 3C). Likewise, we found that knock down of all the
individual components of CRL7SMU1 complex led to accumulation of phospho-H3 positive cells
(Figure 3D & 3E), and significant accumulation of morphologically round cells in culture
(Figure S2C), typical of possibly mitotically arrested cells. Since we found that CRL7SMU1
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

complex proteins are required for normal progression of mitosis, we next tested if loss of SMU1
leads to any mitotic defects. We observed various chromosomal and spindle defects upon SMU1
knockdown (Figure 4A). Loss of SMU1 resulted in significant increase in cells displaying
lagging chromosomes, anaphase/nuclear bridges and multipolar spindles (Figure 4B). Similarly,
we also found that depletion of CUL7, DDB1 or RNF40 individually in cells led to severe
mitotic defects (Figure 4C & 4D). Since loss of CRL7SMU1 complex proteins resulted in
numerous mitotic defects, we next tested if loss of H2B monoubiquitination at K120
phenocopies loss of E3 ligase complex from cells. Expression of H2B K120R, but not wild type
H2B, resulted in accumulation of cells with multiple mitotic defects (Figure 4E, 4F & 4G), thus
suggesting that H2B ubiquitination at K120 position is critical for normal mitotic progression
and further prevention of genomic instability.
CRL7SMU1 complex drives SMC1a gene expression
It is well known that H2B ubiquitination is enriched at sites of active gene transcription and
modulates transcription elongation. Since we found that CRL7SMU1 complex promotes H2B
ubiquitination and H3K4 trimethylation (sites of active gene transcription), we hypothesized that
this complex may control transcription of specific set of genes required for mitosis. Previously, a
microarray analysis upon SMU1 depletion had revealed alterations in the expression of various
mitotic genes (Papasaikas et al., 2015). We tested if SMU1 is enriched on these set of mitotic
genes by using chromatin immunoprecipitation assay (ChIP). SMU1 was significantly enriched
on different mitotic genes such as SMC1a, ANAPC12, ANAPC5, Aurora A and CDCA2 (Figure
5A). Consequently, we tested if depletion of CRL7SMU1 components affects H2B ubiquitination
at these loci. Although, depletion (Figure S3) of individual components of the E3 ligase complex
reduced H2Bub levels at distinct loci, we found SMC1a to be common locus where H2B
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

ubiquitination is hampered with loss of CRL7SMU1 complex (Figure 5B). As H2B ubiquitination
is directly associated with gene transcription, we further tested whether this E3 ligase complex
regulate transcript levels of mitotic genes. Interestingly, again we observed that expression levels
of only SMC1a were reduced upon depletion of CUL7, DDB1, RNF40 and SMU1 (Figure 5C).
Consequently, we also found significant reduction in SMC1a protein levels upon depletion of
individual components of CRL7SMU1 complex (Figure 5D). Notably, expression of H2B K120R
mutant also reduced the gene expression of SMC1a (Figure 5E) consistent with our hypothesis
that H2B ubiquitination is required for expression of this critical mitotic gene.
CRL7SMU1 complex is essential for sister chromatid cohesion
SMC1a (Structural maintenance of chromosomes protein 1A) is a central component of cohesin
complex, which is required for the cohesion of sister chromatids and is essential for accurate
chromosome segregation at the onset of mitosis (Brooker and Berkowitz, 2014; Sumara et al.,
2000). To determine if CRL7SMU1 complex is required for sister chromatid cohesion we prepared
metaphase spreads on HeLa cells depleted with individual components of E3 ligase complex.
Control cells display normal “X-shaped” mitotic chromosomes with sister chromatids tightly
linked at the centromere and chromosome arms separated. However, strikingly, knockdown of
CUL7, DDB1, RNF40 or SMU1 led to parallel chromatids and single chromatids in isolation in a
large fraction of mitotic cells (Figure 6A & 6B) suggesting a premature loss of sister chromatid
cohesion in these cells. We next measured the distance between metaphase sister kinetochores in
CRL7SMU1 complex -depleted cells using CENP-A immunostaining. Sister chromatid
interkinetochore distances were significantly increased in aligned metaphases of SMU1, CUL7,
DDB1 and RNF40 depleted cells compared with control metaphases (Figure 6C & 6D), further
supporting a critical role of CRL7SMU1 complex in establishing cohesion between sister
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

chromatids. Interestingly, expression of H2B K120R mutant, but not wild type H2B, also
resulted in defective chromatid cohesion (Figure 6E & 6F) and enhanced interkinetochore
distance (Figure 6G & 6H), thus suggesting that H2B ubiquitination by CRL7SMU1 complex is
critical for maintenance of sister chromatid cohesion.
Based on these observations, we hypothesized that expression of H2B-Ub fusion protein,
which mimics H2B K120 ubiquitination (Zhang et al., 2013), could rescue the cohesion defects
caused due to depletion of CRL7SMU1 E3 ligase from cells. To our surprise, over-expression of
H2B-Ub fusion protein could not rescue cells from cohesion defects (Figure S4A & S4B) caused
due to E3 ligase loss. This data prompted us to reason that dynamic H2B ubiquitination and
deubiquitination might be necessary for maintenance and later dissolution of sister chromatid
cohesion during appropriate phases of mitosis. Therefore, constitutive loss or presence of H2B
monoubiquitination might be deleterious for normal chromatid cohesion. This is in agreement
with previous data that loss of USP44, a deubiquitinase for H2B, (mimicking constitutive H2B
ubiquitination) also led to various mitotic abnormalities (Zhang et al., 2012). In support of the
dynamic nature of H2B ubiquitination during chromatid cohesion, we found that transient
expression of H2B-Ub fusion protein alone is sufficient to induce sister chromatid cohesion
defects (Figure S4C & S4D), increased interkinetochore distance (Figure S4E) as well as mitotic
defects in cells (Figure S4F & S4G). Nonetheless, we next tested if restoration of SMC1a in cells
through plasmid-based expression would rescue the chromatid cohesion defects caused due to E3
ligase loss. Indeed, exogenous expression of SMC1a (Figure 6A) significantly prevented
defective chromatid cohesion in cells resulted due to depletion of E3 ligase components (Figure
6B). Also, SMC1a expression in CUL7, DDB1, RNF40 or SMU1 depleted cells rescued the
interkinetochore distance that was enhanced due to loss of E3 ligase complex (Figure 6C). In
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

conclusion, we identified that SMU1, DDB1, CUL7 and RNF40 assemble a CRL type E3 ligase
complex and promotes monoubiquitination of H2B to drive the expression of SMC1a, which is
essential for maintenance of sister chromatid cohesion during mitosis.
Discussion
H2B ubiquitination is one of the critical histone modifications associated with gene expression
(Cole et al., 2015; Xie et al., 2017). In humans, RNF20/40 is considered as a major E3 ligase for
H2B monoubiquitination (Kim et al., 2009). Although, H2Bub1 is known to be globally
associated with transcribed genes and its levels correlate positively with the gene expression
(Minsky et al., 2008), knockdown of RNF20 affects transcription of only a subset of genes
(Shema et al., 2008). This possibly suggests the presence of multiple E3 ligase complexes,
independent of RNF20, to regulate H2B monoubiquitination in the cell. In this study, we
provided multiple lines of evidence supporting an essential role of CRL7SMU1 E3 ligase complex
in mediating H2B monoubiquitination. Though H2B ubiquitination has been implicated in wide
range of cellular processes such as transcription initiation and elongation, DNA damage
response, replication, stem cell differentiation, RNA processing and export, its role in chromatid
cohesion and mitosis is unexplored. We clearly have shown that our newly identified E3 ligase
complex promotes H2B ubiquitination at SMC1a to maintain sister chromatid cohesion during
mitosis. Moreover, ectopic expression of the H2Bub1 mutant (H2BK120R) caused an alteration
of the transcription of SMC1a and a significant increase in mitotic defects including cohesion
loss, which is perhaps direct evidence that H2B ubiquitination is critical for mitotic progression.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

E3 ligases play an essential role in the final step of ubiquitination process to catalyze the
transfer of ubiquitin to appropriate substrates. E3 ligases, in particular cullin based class, often
assemble into multimeric complexes to enhance substrate diversity and specificity during the
process of ubiquitination (Lydeard et al., 2013; Zimmerman et al., 2010). Several interaction
domains such as F-Box, SOCS-box, β-domains, WD40 repeats, ankyrin, Kelch, WW and RLD
motifs were known to assist in the assembly of multi-component E3 ligases. In this study, by
using SMU1 as an example, we demonstrated that LisH domain proteins participate in the
organization of a functional E3 ligase complex. LisH (LIS1 homology) is a highly conserved
domain found in eukaryotic proteins that is proposed to mediate protein-protein interactions.
Although proteins harboring LisH domain are known to participate in processes such as
microtubule dynamics, chromosome segregation and are implicated in pathogenesis (Emes and
Ponting, 2001), studies on molecular function of LisH domain are limited. There are certain
examples to suggest that LisH domain participates in organization of multimeric complexes to
regulate protein stability. For instance, in S. cerevisiae, LisH domain proteins such as GID1,
GID7 and GID8 along with GID2, GID4, GID5, and GID9 assemble a multimeric glucose
induced degradation deficient (GID) complex. The GID complex mediates polyubiquitination of
fructose-1,6-bisphosphatase (FBPase) via E3 ubiquitin ligase activity of GID2, that contains a
RING domain (Menssen et al., 2012). However, the role of LisH domain containing proteins in
the assembly of an E3 ligase in humans is unknown. Although earlier, we and others have shown
that DCAF1/VprBP that contains LisH domain acts as a substrate recognition protein in RING
E3s as well as HECT type of E3 ligase complexes (Maddika and Chen, 2009; Nakagawa et al.,
2013), whether LisH domain is required for the assembly of E3 ligases is unexplored. In another
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

example, WDR26 in complex with Axin1 controls β-catenin levels and negatively regulate Wnt
signaling (Goto et al., 2016). Although, LisH domain of WDR26 is critical for β-catenin
degradation, whether it participates in the organization of E3 ligase in this case is not studied.
Thus our current study provides clues in corroborating the critical role of LisH domain proteins
in organizing E3 ligase complexes.
SMC1 is an essential component of Cohesin, a multi-protein complex (made up of four
subunits Smc1, Smc3, an α-kleisin subunit – Mcd1/Scc1 (mitosis)/Rec8 (meiosis), and
Irr1/Scc3), which is conserved from yeast to humans. Cohesin has a well-documented role in
chromatid cohesion where it provides stable but reversible connections between sister
chromatids during both mitosis and meiosis (Nasmyth and Haering, 2009). However, recent
findings indicate that in higher eukaryotes sister chromatid cohesion may not be the only major
function of cohesin, but may also regulate other processes including transcriptional regulation,
DNA repair, chromosome condensation and morphogenesis (Dorsett, 2011). Indeed, elegant
studies in yeast indicate that severe reduction in the level of chromatin bound cohesin does not
entirely affect its function of holding the sister chromatids together, but drastically affects the
non-canonical functions (Dorsett, 2011; Heidinger-Pauli et al., 2010; Mehta et al., 2013). Thus,
although our studies have clearly demonstrated defective sister chromatid cohesion due to
CRL7SMU1 complex loss is dependent on SMC1, it still remains to be determined whether these
effects are mediated directly through SMC1 localization at centromeres or indirectly through
regulation of unknown gene expression.
In addition to H2B ubiquitination, CRL7SMU1 complex may also directly regulate
ubiquitination of mitotic proteins. In support of this speculation, we found proteins such as
Adenomatous polyposis coli (APC) in the list of SMU1 associated proteins. Further studies are
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

required to test if APC acts as a substrate of this E3 ligase complex. Lastly, our work revealed
that a LisH domain protein SMU1 associate with RNF40 to form a RING type CRL E3 ligase.
Interestingly, the list of SMU1 associated proteins also contains HECT type E3 ligases, EDD and
HUWE1. In fact, proteins such as VPRBP and DDB1 were shown previously to associate with
RING as well as HECT E3s (Maddika and Chen, 2009; Nakagawa et al., 2013). Thus, in future
studies it would be interesting to test if SMU1 may participate in the assembly of both RING and
HECT-type of E3 ligases to regulate different cellular processes by controlling distinct set of
substrates.
Materials and methods
Plasmids
Full length SMU1, DDB1, CUL7, RNF40, RNF20, SMC1a, Roc1, VprBP, Rab7, H2B wild type
and K120R mutant were cloned into SFB triple tagged (S protein/Flag/Streptavidin binding
protein) destination vectors using Gateway cloning system (Invitrogen). H2B and SMU1 were
cloned into GST and MBP destination vectors using the same system. SMU1 and DYRK2 were
also cloned into Myc destination vectors and SKP1 was cloned into HA destination vector using
gateway cloning method. The point mutations for H2B was generated by PCR- based site –
directed mutagenesis and cloned into SFB and GST-tagged destination vectors. RNF40 plasmid
was a kind gift from Dr. Steven Johnsen (University Medical Center Göttingen, Göttingen).
Myc-tagged RNF20 was kindly provided by Dr. Jae Bum Kim (Seoul National University, South
Korea). Flag H2B-Ub fusion construct was a kind gift from Dr. Hengbin Wang (UBA,
Alabama). cMyc-SMC1a plasmid was a gift from Michael Kastan (Addgene plasmid #
32363)(Kim et al., 2002), myc3-CUL7 was a gift from Yue Xiong (Addgene plasmid #
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

20695)(Andrews et al., 2006) and GFP-H2B was a gift from Geoff Wahl (Addgene plasmid #
11680)(Kanda et al., 1998).
Antibodies
SMU1 (Abgent # AT3965a) (1:1000), RNF40 (SIGMA # R9029) (1:2000), CUL7 (SIGMA #
C1743) (1:2000), DDB1 (Bethyl # A300-462A) (1:5000), SMC1a (Abcam #ab133643) (1:1000),
H2B (Millipore # 07-371) (1:5000), Ubiquityl-Histone H2B (Lys120) (CST # 5546) (1:1000),
Cyclin A (BD # 611269) (1:1000), CDT1 (Bethyl # A300-786A) (1:1000), pH3 (S10) (Cell
signaling technologies #9701L) (WB- 1:1000, IF- 1:200), Cul4a (Bethyl # A300-739A) (1:5000),
RNF20 (abcam # ab32629) (1:1000), Myc (Santa Cruz # 9E10) (1:1000), Flag (Sigma, F3165)
(1:10000), HA (Bethyl, A190-108A) (1:1000), Actin (Sigma, A5441) (1:10000), and Alpha
tubulin (Sigma, T6074) (WB, 1:5000, IF- 1:200) antibodies were used in this study. HRP/ FITC
conjugated anti-Mouse and anti-Rabbit secondary antibodies were obtained from Jackson
Immunologicals.
Cell lines and transfection
HEK 293T, HeLa and BOSC23 cell lines were used in this work. All cell lines were purchased
from American Type Culture Collection, which were tested and authenticated by the cell bank
using their standard STR (short tandem repeats) based techniques. Cells were also continuously
monitored by microscopy to maintain their original morphology and also tested for mycoplasma
contamination by using DAPI staining. HEK 293T or HeLa were transfected with various
plasmids using PEI (Polysciences) according to the manufacturer’s protocol. Briefly, the plasmid
was mixed with PEI (1 mg/ml) in 1:3 ratio in serum free RPMI medium. Then, DNA-PEI
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

mixture was incubated at room temperature (RT) for 20 min and the complexes were added to
cells to allow the transfection of plasmid.
Tandem affinity purification
SMU1-associated proteins were isolated by using tandem affinity purification as described
before (Maddika and Chen, 2009). Briefly, 293T cells expressing SFB-triple tagged SMU1 were
lysed with NETN lysis buffer (20mM Tris-HCl at pH 8.0, 100mM NaCl, 1mM EDTA, 0.5%
Nonidet P-40) containing protease inhibitors (phenylmethylsulfonyl fluoride (PMSF), Pepstatin
A and aprotinin) on ice for 20 min. The cell lysates were added on to streptavidin–sepharose
beads (Amersham Biosciences) and incubated for 1.5 h at 4°C. Then beads were washed thrice
with lysis buffer and the associated proteins were eluted using 2 mg/ml biotin (Sigma) for 1 h at
4°C. The eluates from the first step of purification were then incubated with S-protein–agarose
beads (Novagen) for 1 h at 4°C. After clearing the unbound proteins by washing, the proteins
associated with S-protein–agarose beads were eluted by boiling in SDS-loading buffer for 10
min at 95 °C. Eluted protein lysate was loaded on SDS-PAGE. The associated proteins were
identified by in gel trypsin digestion followed by LC-MS/MS analysis at Taplin biological mass
spectrometry facility (Harvard University).
Immunoprecipitation and Western blotting
For immunoprecipitation assays, cells were lysed with NETN buffer. The whole-cell lysates
obtained by centrifugation were incubated with 2 µg of specified antibody bound to protein G–
sepharose beads or with streptavidin–sepharose beads (GE) for 1.5 h at 4 °C. The
immunocomplexes were then washed with NETN buffer three times and applied to SDS–PAGE.
Western blotting was carried out by following standard protocols. Proteins were separated by
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

denaturing SDS–PAGE and then transferred on to polyvinylidene difluoride (PVDF) membrane.
The membranes were blocked in 5% non-fat dried milk in Tris-buffered saline (TBS) and then
incubated overnight with the primary antibodies at 4 °C. Then, the blots were incubated with the
corresponding secondary antibodies conjugated with HRP at room temperature for 1 h.
Visualization was carried out by enhanced chemiluminescence detection (Thermo Fisher
Scientific).
Histone extraction
Histones were isolated using acid extraction protocol. Briefly, cells were lysed under hypotonic
conditions and the intact nuclei were collected. Isolated nuclei were acid extracted using 0.4N
H2SO4. Histones were precipitated using 33% TCA and residual TCA was removed by ice-cold
acetone. The pellets were air-dried and milli Q water was added to dissolve histones. Histone
fractions were subjected to SDS-gel followed by western blotting with antibodies of interest.
GST pulldown assays
Bacterially expressed GST and GST-SMU1 bound to glutathione-Sepharose beads were
incubated with eluted MBP-H2B for 1 hr, at 4°C. Beads were washed and proteins were eluted
by boiling in 2X SDS Laemmli buffer and then separated by SDS-PAGE; the interactions were
analysed by western blotting.
RNA interference and lentiviral infection
Control siRNA and pre validated SMU1 siRNA were purchased from Dharmacon (Catalogue no:
J-021129-10). TRC Lentiviral SMU1 shRNA (RHS4533-EG55234), DDB1 shRNA (RHS4533-
EG1642), CUL7 shRNA (RHS4533-EG9820) and RNF40 shRNA (RHS4533-EG9810) were
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

purchased from Dharmacon. Transfection was performed twice, 24 h apart, with 200 nM siRNA
using Oligofectamine reagent in accordance with the manufacturer’s protocol (Invitrogen).
shRNAs were transfected transiently using PEI (Invitrogen) in BOSC23 packaging cells along
with packaging vectors. Forty-eight hours post transfection, the viral medium was collected and
added to the target cells along with polybrene (8 mg/ml). 48 or 72 hours post infection, cells
were collected and processed for various assays and immunoblotting was performed with the
specific antibodies to check the efficiency of knockdown.
Immunofluorescence staining
Cells grown on coverslips were fixed with 3% paraformaldehyde solution in PBS containing
50mM sucrose at room temperature (RT) for 15 min. After permeabilization with 0.5% Triton X-
100 buffer containing 20 mM HEPES at pH 7.4, 50 mM NaCl, 3 mM MgCl2 and 300 mM
sucrose at RT for 5 min, cells were incubated with a 1% BSA for blocking at RT for 30 min.
After washing with PBS, cells were incubated with primary antibodies for 2–3 hrs at RT
followed by three times washes with 1X PBS 5min each. Then, cells were incubated with FITC
or Rhodamine-conjugated secondary antibodies at RT for 60 min followed by three times wash
with 1X PBS 5min each. Nuclei were counterstained with DAPI. After a final wash with PBS,
coverslips were mounted with glycerine containing paraphenylenediamine. Images were taken
using Zeiss confocal microscope (LSM Meta 510 or 700).
Jo
urna
l of C
ell S
cien
ce •
Acc
epte
d m
anus
crip
t

Cell cycle analysis
HeLa cells transfected with the desired expression vectors and si/sh RNA were harvested,
washed with phosphate-buffered saline and fixed with ice-cold 70% ethanol for at least 1 h. Cells
were washed thrice in PBS and treated for 30 min at 37 °C with RNase A (5 μg/ml) and
propidium iodide (25 μg/ml), then analysed using BD accuri C6 flow cytometer.
Cell proliferation assay
HeLa cells were transfected with Control and SMU1 siRNA. 48 hr post transfection 1x10^5 cells
were seeded in five different plates and cell counting was done using trypan blue dye exclusion
method.
Gel filtration
Sephacryl 300 column from GE healthcare was used to separate the protein complexes in the
range of 10 kDa –1500kDa. Column was calibrated using four gel filtration markers
Thioglobulin (669 kDa), Ferritin (440 kDa), Conalbumin (75 kDa) and Ovalbumin (44 kDa).
After calibration, lysates (0.8-1 ml, Concentration 1-2 mg/ml) were injected and allowed to pass
through column. 1 ml fractions were collected using a Bio-Rad 2110 fraction collector within the
molecular weight range of interest and all 1 ml fractions were concentrated separately using
protein concentration columns to final volume of 80 µl. All the fractions were subjected to SDS-
gel followed by western blotting with antibodies of interest.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Quantitative RT-PCR
Total RNA was isolated using Trizol reagent (Invitrogen) as per manufacturer's instructions.
Two microgram of total RNA was reverse transcribed in the presence of anchored oligo dT using
Superscript-III (Invitrogen) as per manufacturer's protocol. Quantitative PCR (qPCR) was then
initiated using the SYBR Pre mix Ex Taq (Tli RNaseH plus) kit (Clontech Laboratories) in 7500
real time PCR systems (Applied Biosystems) as per manufacturer's protocol. The threshold cycle
(Ct) values for particular genes were normalized to GAPDH for each sample. Sequences for
Primers used for qRT-PCR analysis were included in supplementary information.
Chromatin Immunoprecipitation (ChIP) assay
4ug Mouse IgG (bethyl), 4ug SMU1 (Abgent # AT3965a) or 4 µl of H2Bub1 (CST # 5546)
antibodies were used for ChIP assay. Briefly, HeLa cells were crosslinked using 1%
formaldehyde solution. Cross-linking was allowed to proceed for 10 min at room temperature
and was then stopped by the addition of glycine to a final concentration of 0.125 M. Cells were
washed twice with ice-cold 1X PBS and cells were lysed using cell lysis buffer (5mM PIPES
pH-8, 85mM KCL, 0.5% NP-40 and protease inhibitors) followed by nuclei lysis buffer (50mM
Tris pH-8.1, 10mM EDTA, 1% SDS and protease inhibitors). Isolated chromatin was sonicated
using Diagenode Bioruptor at medium power and checked for fragment size. After
microcentrifugation, the supernatant was diluted with 1X IP dilution buffer and preclearing was
performed using Protein G beads. Then equal amount of antibody was added and incubated on a
rotating platform for 12 to 16 h at 4°C. Next day, protein G beads were added to chromatin.
Beads were washed two times with IP Dialysis buffer (2mM EDTA and 50mM Tris pH-8) and
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

three times with IP wash buffer (100mM Tris pH-8, 250mM LiCl, 1% NP-40 and 1% Na
deoxycholate) for five minutes at RT. Then beads were quickly washed with 1X TE and bound
DNA fragments were eluted using elution buffer (50mM NaHCO3 and 1% SDS) at 65°C. Input
and elution products were kept for decrosslinking at 65°C O/N. Then, samples were treated with
RNase A and proteinase K for 3 hours at 37°C and DNA was extracted using PCI treatment.
Quantitative PCR (qPCR) was then initiated using the SYBR Pre mix Ex Taq (Tli RNaseH plus)
kit (Clontech Laboratories) in 7500 real time PCR systems (Applied Biosystems) as per
manufacturer's protocol. Sequences for primers used for ChIP analysis were included in
supplementary information.
Metaphase spreads
Metaphase chromosome spreads were performed as described before(Maddika et al., 2009).
Briefly, cells treated with colcemid for 4 hours were collected, washed with phosphate buffered
saline (PBS) and treated with 75 mM KCl at room temperature for 30 min. The treated cells were
then fixed in fresh methanol:acetic acid (3:1) solution and dropped onto glass slides. Cells were
allowed to air dry and were stained with giemsa (5%) and visualized under the microscope.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Author contributions
SM conceptualized and managed the project. SM and VJS designed the experiments, analysed
the data and wrote the manuscript. VJS performed all the experiments.
Acknowledgements
This work was supported in part by a grant from Council of Scientific & Industrial Research,
India (to S.M; 37(6371)/17) and CDFD core funds. S.M is a senior fellow of Wellcome
Trust/DBT India Alliance and a recipient of Department of Biotechnology’s Sr. IYBA award.
V.J.S acknowledge the fellowship support from Department of Biotechnology (DBT), India. We
thank CDFD core facility as well as Dr. Rohit Joshi’s lab for their assistance in confocal
imaging. We thank Nanci Rani for technical assistance, Shaffiqu T S for providing assistance in
gel filtration experiment, Sawant Suresh, Zaffer Ullah Zargar and Amit M Karole for their
critical inputs during gene expression and ChIP experiments, Dr, Ashwin Dalal and Dr. Usha
Dutta for their inputs during mitotic spread experiments. We also thank all members of LCDCS
for their suggestions and critical inputs at various stages of the project.
Competing financial interests
The authors declare no competing financial interests.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

References
Andrews, P., He, Y. J. and Xiong, Y. (2006). Cytoplasmic localized ubiquitin ligase cullin 7 binds to p53 and promotes cell growth by antagonizing p53 function. Oncogene 25, 4534-48. Bennett, E. J., Rush, J., Gygi, S. P. and Harper, J. W. (2010). Dynamics of cullin-RING ubiquitin ligase network revealed by systematic quantitative proteomics. Cell 143, 951-65. Berndsen, C. E. and Wolberger, C. (2014). New insights into ubiquitin E3 ligase mechanism. Nat Struct Mol Biol 21, 301-7. Brooker, A. S. and Berkowitz, K. M. (2014). The roles of cohesins in mitosis, meiosis, and human health and disease. Methods Mol Biol 1170, 229-66. Cole, A. J., Clifton-Bligh, R. and Marsh, D. J. (2015). Histone H2B monoubiquitination: roles to play in human malignancy. Endocr Relat Cancer 22, T19-33. Dias, D. C., Dolios, G., Wang, R. and Pan, Z. Q. (2002). CUL7: A DOC domain-containing cullin selectively binds Skp1.Fbx29 to form an SCF-like complex. Proc Natl Acad Sci U S A 99, 16601-6. Dorsett, D. (2011). Cohesin: genomic insights into controlling gene transcription and development. Curr Opin Genet Dev 21, 199-206. Emes, R. D. and Ponting, C. P. (2001). A new sequence motif linking lissencephaly, Treacher Collins and oral-facial-digital type 1 syndromes, microtubule dynamics and cell migration. Hum Mol Genet 10, 2813-20. Francis, O., Han, F. and Adams, J. C. (2013). Molecular phylogeny of a RING E3 ubiquitin ligase, conserved in eukaryotic cells and dominated by homologous components, the muskelin/RanBPM/CTLH complex. PLoS One 8, e75217. Goto, T., Matsuzawa, J., Iemura, S., Natsume, T. and Shibuya, H. (2016). WDR26 is a new partner of Axin1 in the canonical Wnt signaling pathway. FEBS Lett 590, 1291-303. Harper, J. W. and Tan, M. K. (2012). Understanding cullin-RING E3 biology through proteomics-based substrate identification. Mol Cell Proteomics 11, 1541-50. Heidinger-Pauli, J. M., Mert, O., Davenport, C., Guacci, V. and Koshland, D. (2010). Systematic reduction of cohesin differentially affects chromosome segregation, condensation, and DNA repair. Curr Biol 20, 957-63. Hershko, A. and Ciechanover, A. (1998). The ubiquitin system. Annu Rev Biochem 67, 425-79. Higa, L. A., Wu, M., Ye, T., Kobayashi, R., Sun, H. and Zhang, H. (2006). CUL4-DDB1 ubiquitin ligase interacts with multiple WD40-repeat proteins and regulates histone methylation. Nat Cell Biol 8, 1277-83. Iovine, B., Iannella, M. L. and Bevilacqua, M. A. (2011). Damage-specific DNA binding protein 1 (DDB1): a protein with a wide range of functions. Int J Biochem Cell Biol 43, 1664-7. Kanda, T., Sullivan, K. F. and Wahl, G. M. (1998). Histone-GFP fusion protein enables sensitive analysis of chromosome dynamics in living mammalian cells. Curr Biol 8, 377-85. Kim, J., Guermah, M., McGinty, R. K., Lee, J. S., Tang, Z., Milne, T. A., Shilatifard, A., Muir, T. W. and Roeder, R. G. (2009). RAD6-Mediated transcription-coupled H2B ubiquitylation directly stimulates H3K4 methylation in human cells. Cell 137, 459-71. Kim, S. T., Xu, B. and Kastan, M. B. (2002). Involvement of the cohesin protein, Smc1, in Atm-dependent and independent responses to DNA damage. Genes Dev 16, 560-70. Lydeard, J. R., Schulman, B. A. and Harper, J. W. (2013). Building and remodelling Cullin-RING E3 ubiquitin ligases. EMBO Rep 14, 1050-61.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Maddika, S. and Chen, J. (2009). Protein kinase DYRK2 is a scaffold that facilitates assembly of an E3 ligase. Nat Cell Biol 11, 409-19. Maddika, S., Sy, S. M. and Chen, J. (2009). Functional interaction between Chfr and Kif22 controls genomic stability. J Biol Chem 284, 12998-3003. Mehta, G. D., Kumar, R., Srivastava, S. and Ghosh, S. K. (2013). Cohesin: functions beyond sister chromatid cohesion. FEBS Lett 587, 2299-312. Menssen, R., Schweiggert, J., Schreiner, J., Kusevic, D., Reuther, J., Braun, B. and Wolf, D. H. (2012). Exploring the topology of the Gid complex, the E3 ubiquitin ligase involved in catabolite-induced degradation of gluconeogenic enzymes. J Biol Chem 287, 25602-14. Minsky, N., Shema, E., Field, Y., Schuster, M., Segal, E. and Oren, M. (2008). Monoubiquitinated H2B is associated with the transcribed region of highly expressed genes in human cells. Nat Cell Biol 10, 483-8. Nakagawa, T., Mondal, K. and Swanson, P. C. (2013). VprBP (DCAF1): a promiscuous substrate recognition subunit that incorporates into both RING-family CRL4 and HECT-family EDD/UBR5 E3 ubiquitin ligases. BMC Mol Biol 14, 22. Nasmyth, K. and Haering, C. H. (2009). Cohesin: its roles and mechanisms. Annu Rev Genet 43, 525-58. Papasaikas, P., Tejedor, J. R., Vigevani, L. and Valcarcel, J. (2015). Functional splicing network reveals extensive regulatory potential of the core spliceosomal machinery. Mol Cell 57, 7-22. Petroski, M. D. and Deshaies, R. J. (2005). Function and regulation of cullin-RING ubiquitin ligases. Nat Rev Mol Cell Biol 6, 9-20. Ren, L., Liu, Y., Guo, L., Wang, H., Ma, L., Zeng, M., Shao, X., Yang, C., Tang, Y., Wang, L. et al. (2013). Loss of Smu1 function de-represses DNA replication and over-activates ATR-dependent replication checkpoint. Biochem Biophys Res Commun 436, 192-8. Rotin, D. and Kumar, S. (2009). Physiological functions of the HECT family of ubiquitin ligases. Nat Rev Mol Cell Biol 10, 398-409. Shema, E., Tirosh, I., Aylon, Y., Huang, J., Ye, C., Moskovits, N., Raver-Shapira, N., Minsky, N., Pirngruber, J., Tarcic, G. et al. (2008). The histone H2B-specific ubiquitin ligase RNF20/hBRE1 acts as a putative tumor suppressor through selective regulation of gene expression. Genes Dev 22, 2664-76. Spratt, D. E., Walden, H. and Shaw, G. S. (2014). RBR E3 ubiquitin ligases: new structures, new insights, new questions. Biochem J 458, 421-37. Sugaya, K., Hongo, E., Ishihara, Y. and Tsuji, H. (2006). The conserved role of Smu1 in splicing is characterized in its mammalian temperature-sensitive mutant. J Cell Sci 119, 4944-51. Sugaya, K., Hongo, E. and Tsuji, H. (2005). A temperature-sensitive mutation in the WD repeat-containing protein Smu1 is related to maintenance of chromosome integrity. Exp Cell Res 306, 242-51. Sumara, I., Vorlaufer, E., Gieffers, C., Peters, B. H. and Peters, J. M. (2000). Characterization of vertebrate cohesin complexes and their regulation in prophase. J Cell Biol 151, 749-62. Wood, A., Krogan, N. J., Dover, J., Schneider, J., Heidt, J., Boateng, M. A., Dean, K., Golshani, A., Zhang, Y., Greenblatt, J. F. et al. (2003). Bre1, an E3 ubiquitin ligase required for recruitment and substrate selection of Rad6 at a promoter. Mol Cell 11, 267-74. Xie, W., Nagarajan, S., Baumgart, S. J., Kosinsky, R. L., Najafova, Z., Kari, V., Hennion, M., Indenbirken, D., Bonn, S., Grundhoff, A. et al. (2017). RNF40 regulates gene expression in an epigenetic context-dependent manner. Genome Biol 18, 32. Zhang, Y., Foreman, O., Wigle, D. A., Kosari, F., Vasmatzis, G., Salisbury, J. L., van Deursen, J. and Galardy, P. J. (2012). USP44 regulates centrosome positioning to prevent aneuploidy and suppress tumorigenesis. J Clin Invest 122, 4362-74.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Zhang, Z., Jones, A., Joo, H. Y., Zhou, D., Cao, Y., Chen, S., Erdjument-Bromage, H., Renfrow, M., He, H., Tempst, P. et al. (2013). USP49 deubiquitinates histone H2B and regulates cotranscriptional pre-mRNA splicing. Genes Dev 27, 1581-95. Zimmerman, E. S., Schulman, B. A. and Zheng, N. (2010). Structural assembly of cullin-RING ubiquitin ligase complexes. Curr Opin Struct Biol 20, 714-21.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Figures
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Figure 1: SMU1 assembles CRL type E3 ligase complex by interacting with DDB1, CUL7
and RNF40. (A) Graphical representation of proteins containing LisH domain and WD repeats.
(B) Partial list of SMU1-associated proteins identified by biochemical purification followed by
mass spectrometric analysis were listed together with the number of peptides for each
protein. (C) Immunoprecipitation (IP) with control IgG or anti-SMU1 antibody was performed
with extracts prepared from 293T cells. The presence of RNF40, DDB1, CUL7 and DYRK2 in
these immunoprecipitates was evaluated by immunoblotting with their respective antibodies. (D)
SFB-VPRBP along with either Myc-SMU1 or Myc-DYRK2 was expressed in cells and the
interaction of these proteins was detected by immunoblotting with the indicated antibodies after
pulling down the complexes with streptavidin sepharose. (E) HA tagged-SKP1 along with either
SFB-SMU1 or SFB-ROC1 and (F) SFB-ROC1 along with Myc-SMU1 or HA-SKP1 were
expressed in cells and the their interaction was detected as described in figure 1D. (G) HeLa
cells expressing Myc-RNF20 were lysed and immunoprecipitation was carried out using either
IgG or Myc antibody. The presence of SMU1 and RNF40 was detected in these
immunoprecipitates by immunoblotting using specific antibodies. (H) 293T cell extracts was
analysed by size-exclusion chromatography using Sephacryl 300 column. Proteins eluted from
the different fractions were immunoblotted with the indicated antibodies. (I) Graphical
representation of domain architecture of SMU1 and its deletion mutants. (J) SFB tagged SMU1
FL and deletion mutants were transfected in HeLa cells. 24hr Post transfection, cells were lysed
and pulldown was carried out using SBP beads. The presence of DDB1, CUL7 and RNF40 in
these precipitates were evaluated by immunoblotting with their respective antibodies.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Figure 2: CRL7SMU1 complex regulates the mono-ubiquitination of H2B at K120 site. (A)
SFB tagged CUL7, DDB1, RNF40, SMU1, Rab7 or empty vector (EV) were transfected and the
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

interaction of H2B was detected by immunoblotting with specific antibody after streptavidin
sepharose pull down. (B) Hela cells were transduced with either control or SMU1 specific
shRNA followed by over expression of SFB-RNF40. 72 hours post transduction, pulldown was
performed with streptavidin sepharose beads and the interaction of DDB1 and H2B with RNF40
was evaluated by immunoblotting with their respective antibodies. (C) SFB tagged SMU1 was
overexpressed in cells transduced with either control or RNF40 shRNA. The interaction of
SMU1 with H2B and DDB1 was detected through immunoblotting using specific antibodies
after immunoprecipitation. (D) Cells were transduced with either control or DDB1 shRNA and
(E) Control or CUL7 shRNA containing viral particles. Pulldown followed by detection of
different indicated proteins in precipitates were done as described in figure 2B. (F) Model shows
the assembly of CRL7SMU1 complex in association with its substrate H2B. (G) GST pulldown
assay was performed with immobilized control GST or GST–SMU1 fusion proteins on
glutathione beads, followed by incubation with bacterially purified MBP-H2B. The interaction of
SMU1 with H2B was assessed by immunoblotting with MBP antibody. Expression of the GST,
recombinant GST-SMU1 and MBP-H2B was shown by coomassie staining. (H) HeLa cells were
transduced using either control or RNF40 shRNA. Post 72 hours, cells were collected and lysed
to isolate soluble and histone fractions. Lysates were subjected to SDS-PAGE followed by
immunoblotting using the indicated antibodies. (I) Cells were transfected/transduced with either
control or SMU1 siRNA, (J) or DDB1 shRNA, (K) or CUL7 shRNA. Soluble and acid extracted
histone fractions were subjected to SDS-PAGE followed by immunoblotting using indicated
antibodies. The data presented here is representation of three independent experiments.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Figure 3: Intact CRL7SMU1 complex is required for mitotic progression. (A) HeLa cells
transfected with either control siRNA or SMU1 specific siRNAs were stained with Propidium
Iodide and cell cycle analysis was performed by flow cytometry. (B) HeLa cells were transfected
with indicated siRNAs. The transition of cells through mitosis was analyzed by live cell time-
lapse microscopy after synchronizing cells using double thymidine block. (C) Time taken by
each cell from mitotic entry to division was calculated and the data was plotted for control and
SMU1 depleted cells (n=15). (D) HeLa cells were transduced with control shRNA, Cul7 shRNA,
DDB1 shRNA, RNF40 shRNA and SMU1 shRNA separately. 72 hrs post transduction, cells
were probed with pH3 antibody to measure mitotic index. Nuclei were counterstained with
DAPI. (E) Quantification of mitotic index results is shown.
Jo
urna
l of C
ell S
cien
ce •
Acc
epte
d m
anus
crip
t

Figure 4: Depletion of CRL7SMU1 complex proteins induce mitotic defects. (A) HeLa cells
were transfected with either control or SMU1 siRNAs. Cells were stained with alpha- tubulin
antibody to check the spindle defects (multipolar spindles) and nuclei were counterstained with
DAPI to check for chromosomal defects (lagging chromosomes and anaphase bridges) (scale
bar, 10μm). (B) Quantification of results from figure 4A is shown (n=50 cells each). (C) HeLa
cells were transduced with control shRNA, CUL7 shRNA, DDB1 shRNA or RNF40 shRNA.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Cells were stained with alpha- tubulin antibody to check the spindle defects (multipolar spindles)
and nuclei were counterstained with DAPI to check for chromosomal defects (lagging
chromosomes and anaphase bridges) (Scale bar, 5μm). (D) Quantification of results from figure
4C is shown, (n=40 cells each). (E) Cells were transfected with H2B Wild type (WT) and K120R
mutant. (F) Various mitotic abnormalities in cells expressing H2B Wild type (WT) and K120R
mutant were checked using immunofluorescence after staining with alpha- tubulin antibody and
DAPI (Scale bar, 5μm). (G) Quantification of results from figure 4F is shown (n=50 cells each).
Statistical analysis: error bars- standard deviation, ***P<0.001 **P<0.01; *P<0.05, Student’s t-
test.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Figure 5: CRL7SMU1 complex is necessary for driving SMC1 gene expression. (A)
Exponentially growing HeLa cells were subjected to ChIP analysis using either SMU1 or IgG
antibody. SMU1 enrichment at various loci is shown. The data shown is derived from three
independent experiments. (B) Cells expressing control shRNA, CUL7 shRNA, DDB1 shRNA,
RNF40 shRNA and SMU1 shRNA were subjected to ChIP analysis using H2Bub antibody. Fold
change of H2Bub enrichment at indicated loci with respect to control shRNA was shown. The
data shown is derived from three independent experiments. (C) Total RNA was extracted from
HeLa cells transfected with control or SMU1 siRNA or CUL7 shRNA or DDB1 shRNA and
RNF40 shRNA, and expression levels of various genes measured by qRT-PCR from three
independent experiments was shown. (D) HeLa cells transduced with control, CUL7 shRNA,
DDB1 shRNA, RNF40 shRNA or SMU1 shRNA and levels of SMC1a protein was measured by
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

immunoblotting with specific antibody. (E) HeLa cells were transfected with H2B wild type and
K120R mutant. Relative expression of indicated genes measured by using qRT-PCR from three
independent experiments was plotted. Statistical analysis: error bars- standard deviation,
***P<0.001; **P<0.01; *P<0.05, Student’s t-test.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt
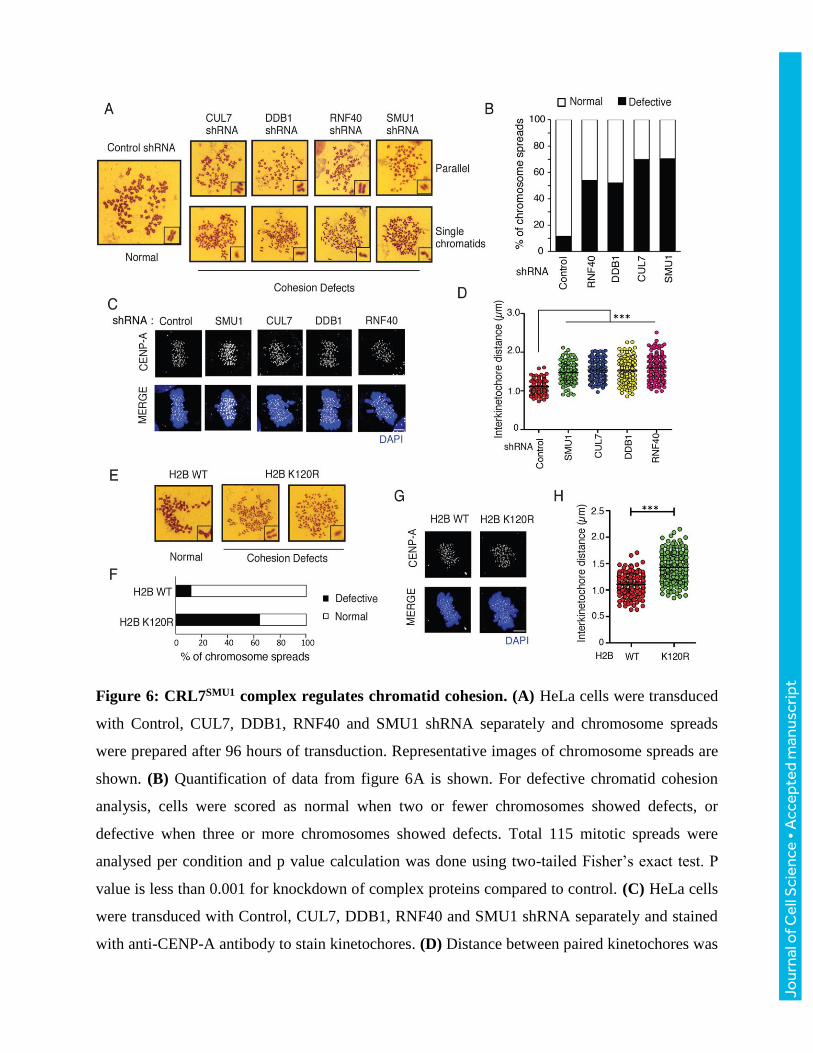
Figure 6: CRL7SMU1 complex regulates chromatid cohesion. (A) HeLa cells were transduced
with Control, CUL7, DDB1, RNF40 and SMU1 shRNA separately and chromosome spreads
were prepared after 96 hours of transduction. Representative images of chromosome spreads are
shown. (B) Quantification of data from figure 6A is shown. For defective chromatid cohesion
analysis, cells were scored as normal when two or fewer chromosomes showed defects, or
defective when three or more chromosomes showed defects. Total 115 mitotic spreads were
analysed per condition and p value calculation was done using two-tailed Fisher’s exact test. P
value is less than 0.001 for knockdown of complex proteins compared to control. (C) HeLa cells
were transduced with Control, CUL7, DDB1, RNF40 and SMU1 shRNA separately and stained
with anti-CENP-A antibody to stain kinetochores. (D) Distance between paired kinetochores was
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

measured at individual Z planes and plotted (n= 150). (E) Chromosome spreads were prepared
from HeLa cells transfected with either H2B wild type or H2B K120R mutant. Representative
images of chromosome spreads were shown. (F) Quantification of data from figure 6E was
shown (n= 100) p < 0.001. (G) Cells expressing either H2B WT or K120R mutant were stained
with anti-CENPA antibody and (H) the distance between paired kinetochores was measured.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Figure 7: Exogenous SMC1a expression partially rescues cohesion defects caused due to
loss of CRL7SMU1 complex. (A) HeLa cells were transduced with control shRNA, CUL7
shRNA, DDB1 shRNA, RNF40 shRNA and SMU1 shRNA alone or together with SMC1a
plasmid and SMC1a levels were analysed by immunoblotting. (B) Cells expressing control
shRNA, Cul7 shRNA, DDB1 shRNA, RNF40 shRNA and SMU1 shRNA alone or together with
SMC1a were analysed for chromatid cohesion (n= 100, P value is less than 0.001 for knockdown
of complex proteins compared to control and less than 0.01 between shRNA alone and along
with SMC1a (rescue) (C) and inter kinetochore distance (n= 190). Statistical analysis: error bars-
standard deviation, ***P<0.001; **P<0.01, Student’s t-test.
Jour
nal o
f Cel
l Sci
ence
• A
ccep
ted
man
uscr
ipt

Supplementary Data
Supplementary table 1: List of primers used in this study for ChIP experiments were shown.
Gene name Primer sequence Location chr12:121756213-121756365, strand= reverse
ANAPC5 For: GGAGATGTGGTGGTGGATCT ANAPC5 Rev: ACCGGTACTCCTTGGAGAGG SMC1A For: CGGTGAACCTCTGAAATGGT chrX: 53449441-53449547,
strand= forward SMC1A Rev: TGGGGTTCCTGAAACTGATT MUS81 For: TCTCCAAACCCTCTCTGCTC chr11:65632546-65632715,
strand= forward MUS81 Rev: AAACACTTCTCGCACCGACT CDCA2 For: CCTGAAACCAAGGAGTCTGC chr8:25317784-25317933,
strand= forward CDCA2 Rev: TCTTCCCAGTTCCCAAAATG ANAPC12 For: GACCCGTAAGAAACAGAAGGAA chr9:116029543-116029694, ANAPC12 Rev: TGAACGATTATTGGGCTTGG strand= reverse AURKA For: CAGGCCAATCGGCTTTCTA chr20:54967248-54967345,
strand= reverse AURKA Rev: TTCCAAGAGCTCAGCCGTTA
J. Cell Sci. 131: doi:10.1242/jcs.213868: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion

Gene name Primer sequence
DDB1 For: TGCCTACAAGCCCATGGAAG DDB1 Rev: TGGCAGCGCTATCCTTTTGA RNF40 For: AAGAGGGCTCAGGAGGACAT RNF40 Rev: CATCTGCGTAGACCTCCACC DDB1 For: ATCATTGACCCTGAGTGCCG DDB1 Rev: CCTGAGGGTCCTGGTAGACA CUL7 For: TACCAGGAGGGGTCCTCAAG CUL7 Rev: TTCTCCAAGTTCTGGCCGTC SMU1 For: TGGCACTGTAAAGATCTGGAA SMU1 Rev: CTGTTGCACACCACAAAGTG SMC1a_For: TACCTCAGTTCTCGGGCGTA SMC1a_Rev: TGACTTACCAGAGCCATTGGG MUS81_For: GGCCCAGGAGACCAATCCT MUS81_Rev: TTCAGCCGGAACTTCTGCTC AURKA_For: GTCAAGTCCCCTGTCGGTTC AURKA_Rev: GCAATGGAGTGAGACCCTCT ANAPC5_For: CGGTCACTATCAACAGGCAGA ANAPC5_Rev: GCGAGGTACGGTAACCCAAA ANAPC12_For: AGGCCGGCTTGAAAAGAGATTA ANAPC12_Rev: GGGCCAAACCCAGTGAACTA CDCA2_For: CCGAAGACTGGGTTCAGGTTA CDCA2_Rev: TGCCAAGCGATGAGGAACTT
Supplementary table 2: List of primers used in this study for qRT-PCR analysis were
shown.
J. Cell Sci. 131: doi:10.1242/jcs.213868: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion
C
Inpu
t
SFB
- EV
SFB
- CU
L7
Flag (CUL7)
DDB1
RNF40
SMU1
CUL4a
IP :
SBP
(CUL
7)
SFB
- EV
SFB
- CU
L7
DDB1
RNF40
SMU1
CUL4a
CUL7
DDB1
SMU1
CUL4a
SFB
- EV
SFB
- RN
F40
IP :
SBP
(RNF
40)
Flag (RNF40)
DDB1
SMU1
CUL4a
CUL7
SFB
- EV
SFB
- RN
F40
Inpu
t
SMU1
CUL4a
CUL7
RNF40
IP :
SBP
(DDB
1)
SFB
- EV
SFB
- DD
B1
Flag (DDB1)
SMU1
CUL4a
CUL7
RNF40
SFB
- EV
SFB
- DD
B1
Inpu
t
A BS
BP
IgG
Flag (SMU1)
Myc (CUL7)
SFB
-SM
U1
Myc
-DY
RK
2
Flag (SMU1)
SFB
-SM
U1
Myc
-CU
L7
Myc (DYRK2)
IP
Inpu
t
Flag (SMU1)
SFB
-SM
U1
Fla
g-D
DB
1 S
FB-S
MU
1 M
yc-R
NF4
0
Myc (RNF40)
Flag (SMU1)
Flag (DDB1)
D
Figure S1
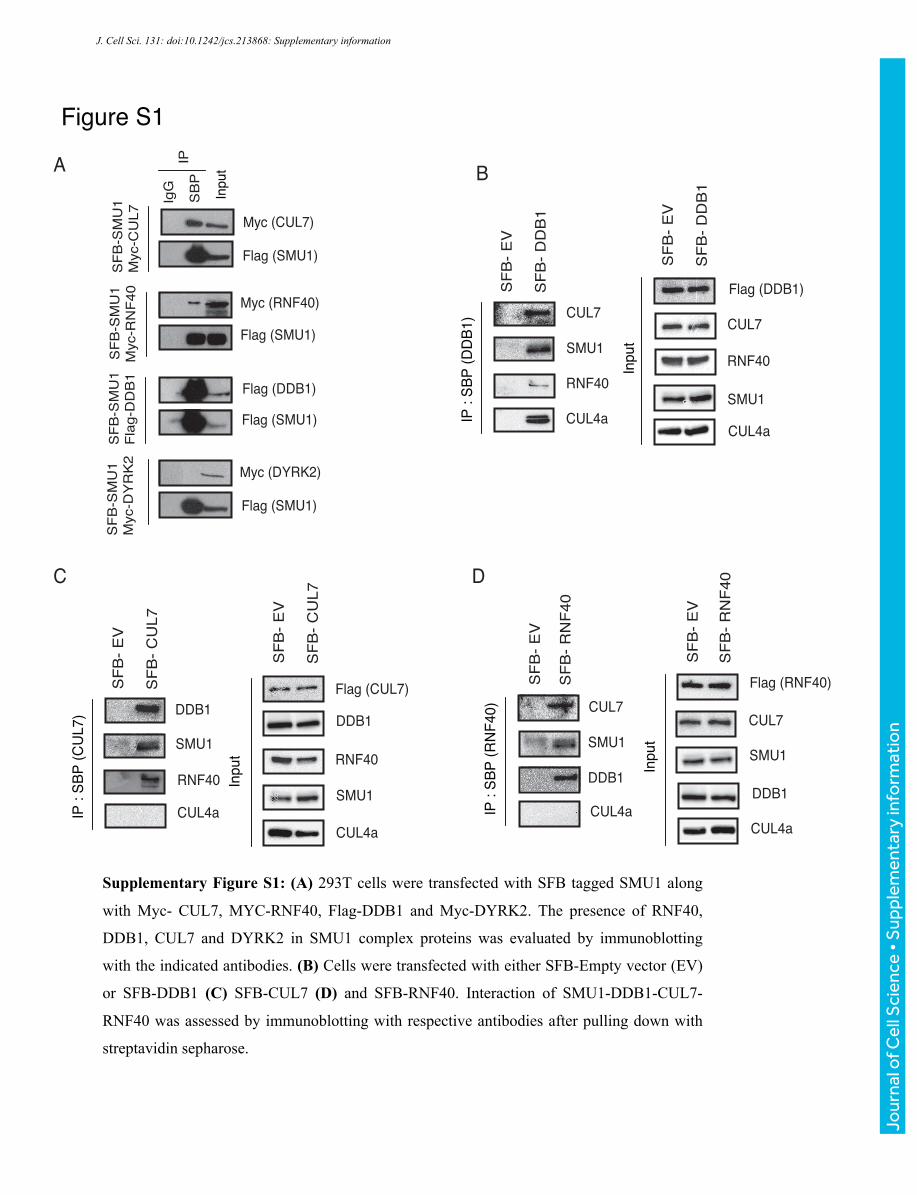
Supplementary Figure S1: (A) 293T cells were transfected with SFB tagged SMU1 along
with Myc- CUL7, MYC-RNF40, Flag-DDB1 and Myc-DYRK2. The presence of RNF40,
DDB1, CUL7 and DYRK2 in SMU1 complex proteins was evaluated by immunoblotting
with the indicated antibodies. (B) Cells were transfected with either SFB-Empty vector (EV)
or SFB-DDB1 (C) SFB-CUL7 (D) and SFB-RNF40. Interaction of SMU1-DDB1-CUL7-
RNF40 was assessed by immunoblotting with respective antibodies after pulling down with
streptavidin sepharose.
J. Cell Sci. 131: doi:10.1242/jcs.213868: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion

Figure S2A B
Day 1 2 3 4 5
20
15
10
5
0
Control siRNA SMU1 siRNA
Prol
ifera
tion
(105
)
Control shRNA SMU1 shRNA
DDB1 shRNA CUL7 shRNA
RNF40 shRNA AN
Ris lortnoC
AN
Ris 1U
MS
SMU1
Actin
C
Supplementary Figure S2: (A) HeLa cells transfected with control or SMU1 siRNA. Cell
lysates were checked for knockdown of SMU1 using SMU1 specific antibody and actin was
used as loading control. (B) HeLa cells were transfected with control and SMU1 siRNA
separately. 48 hr post transduction 1*10^5 cells were seeded in five different plates and rate
of cell proliferation was measured by cell counting after staining with trypan blue dye. (C)
Cells were transduced with control shRNA, RNF40 shRNA, SMU1 shRNA, DDB1 shRNA
or CUL7 shRNA. 72 hours post transduction, mitotic cells (rounded off cells) were imaged in
phase contrast microscope and representative images were shown.
J. Cell Sci. 131: doi:10.1242/jcs.213868: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion
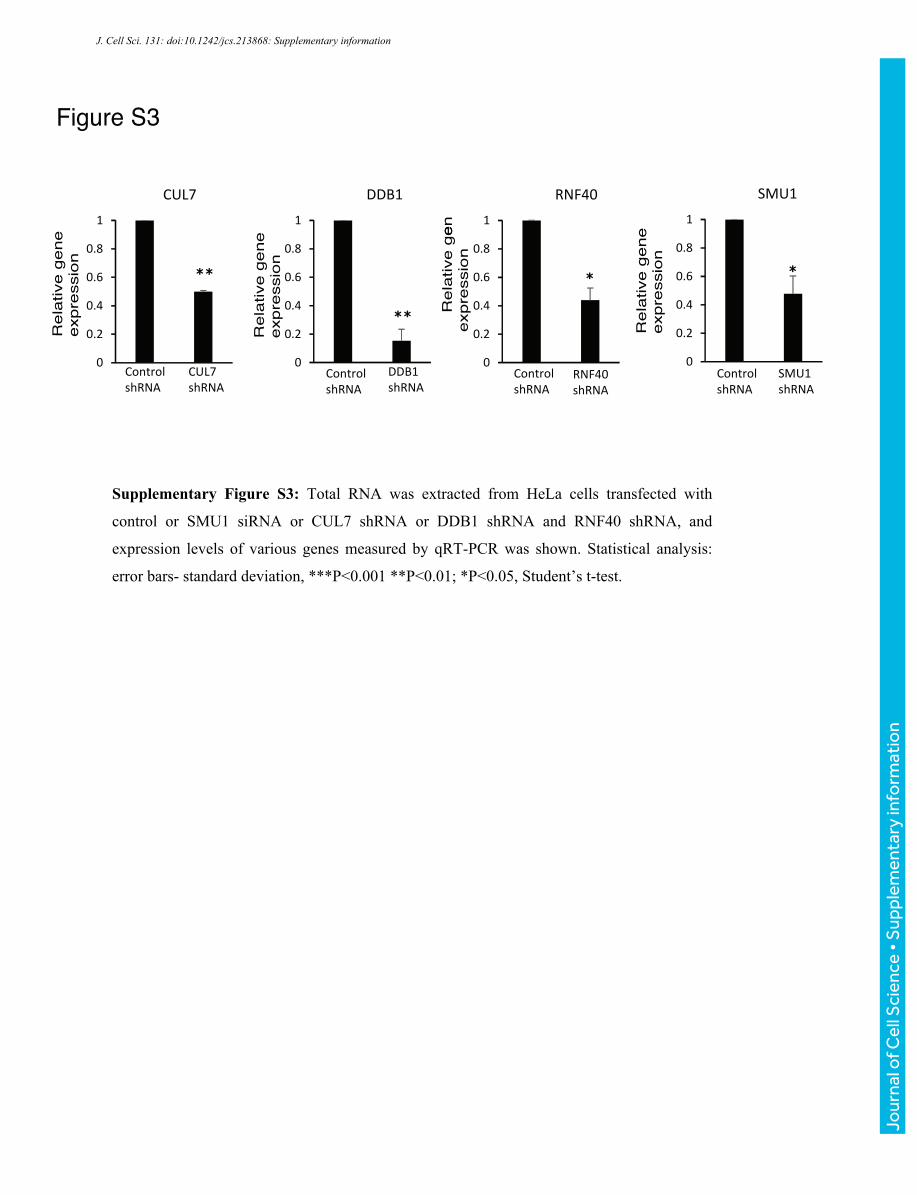
Figure S3
0
0.2
0.4
0.6
0.8
1
Control shRNA
CUL7 shRNA
CUL7
0
0.2
0.4
0.6
0.8
1
DDB1 shRNA
DDB1
eneg evitaleR
noisserpxe
eneg evitaleR
noisserpxe
eneg evitaleR
noisserpxe
eneg evitaleR
noisserpxe
**
**
0
0.2
0.4
0.6
0.8
1
RNF40 shRNA
RNF40
0
0.2
0.4
0.6
0.8
1
SMU1 shRNA
SMU1
* *
Control shRNA
Control shRNA
Control shRNA
Supplementary Figure S3: Total RNA was extracted from HeLa cells transfected with
control or SMU1 siRNA or CUL7 shRNA or DDB1 shRNA and RNF40 shRNA, and
expression levels of various genes measured by qRT-PCR was shown. Statistical analysis:
error bars- standard deviation, ***P<0.001 **P<0.01; *P<0.05, Student’s t-test.
J. Cell Sci. 131: doi:10.1242/jcs.213868: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion

Figure S4
Cohesion DefectsNormal
H2B - Ub fusion
H2B WT
% of chromosome spreads
0 20 40 60 80 100
H2B - Ub fusionH2B WT
NormalDefective
C
A B
Cohesion Defects
Normal
Control shRNA
+ H2B ub fusion
D
0
20
40
60
80
100
shRNA : Control CUL7 DDB1 RNF40 SMU1 H2B Ub – – + – + – + – +Fusion:
Defective Normal
% o
f Ch
rom
osom
e s
prea
ds
*** *** **
Multipolar spindles
Anaphase Bridges
Lagging chromosomes
0
10
20
30
% o
f cel
ls
H2B WT H2B-Ub fusion
F
H2B WT
H2B ub fusion
Lagging chromosomes
Anaphase bridges
Multipolar spindles
DAPI
E
Inte
rkin
etoc
hore
dist
ance
(µm
)
H2B WT Ub fusion
2.0
1.0
0
3.0 ***
G
CUL7 shRNA
DDB1 shRNA
RNF40shRNA
SMU1 shRNA
J. Cell Sci. 131: doi:10.1242/jcs.213868: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion

Supplementary Figure S4: (A) Cells expressing control shRNA, CUL7 shRNA, DDB1
shRNA, RNF40 shRNA and SMU1 shRNA alone or together with H2B-Ub fusion were
analysed for chromatid cohesion (n= 75). (B) Percentage of chromosome spreads showing
normal and defective cohesion was plotted. P value is less than 0.001 for knockdown of
complex proteins compared to control and non-significant between shRNA alone and along
with H2B-Ub fusion (C) Chromosome spreads were prepared form HeLa cells transfected
with either H2B wild type or H2B-Ub fusion. Representative images of chromosome spreads are shown. (D) Quantification of data from figure S3C is shown. (n= 90), p < 0.001. (E) The
distance between paired kinetochores was measured for cells transfected with either H2B WT
or H2B-Ub fusion. (F) Cells were transfected with H2B Wild type (WT) or H2B-Ub fusion
protein. Mitotic defects were detected using immunofluorescence after staining with DAPI
(Scale bar, 10µm). (G) Quantification of the data from figure S5F is shown (n=25 cells each).
Statistical analysis: error bars- standard deviation, ***P<0.001 **P<0.01; *P<0.05, Student’s
t-test. (Scale bar, 5µm).
J. Cell Sci. 131: doi:10.1242/jcs.213868: Supplementary information
Jour
nal o
f Cel
l Sci
ence
• S
uppl
emen
tary
info
rmat
ion