crocodile remains from the burdigalian (lower miocene) of...
TRANSCRIPT

309GEODIVERSITAS • 2004 • 26 (2) © Publications Scientifiques du Muséum national d’Histoire naturelle, Paris. www.geodiversitas.com
Crocodile remains from the Burdigalian (lower Miocene) of Gebel Zelten (Libya)
Eduardo LLINÁS AGRASARDepartment of Earth Sciences, University of Bristol,
Wills Memorial Building, Queen’s Road BS8 1RJ, Bristol (England)[email protected]
Llinás Agrasar E. 2004. — Crocodile remains from the Burdigalian (lower Miocene) of GebelZelten (Libya). Geodiversitas 26 (2) : 309-321.
ABSTRACTThe Gebel Zelten locality in Cirenaica, North Central Libya (Burdigalian,lower Miocene) has yielded numerous fossil remains of crocodilians. Althoughfragmentary in nature, several forms can be distinguished, among them thelongirostrines Euthecodon arambourgi Ginsburg & Buffetaut, 1978 andTomistoma lusitanica Antunes, 1967 and the brevirostrine Rimasuchus lloydiFourtau, 1918. There are also other remains present in the collections fromthis locality that cannot be assigned with confidence to any known croco-dilian species due to their fragmentary condition. Nevertheless, these remainsindicate the presence of at least one additional taxon, a previously unidenti-fied brevirostrine form differing from Rimasuchus lloydi in the morphology ofits lower jaw. The crocodilian fauna from this locality shows similarities withthat from the Burdigalian of Wadi Moghara (Egypt), pointing to the exis-tence of a crocodilian assemblage in North Africa during the early Miocene,showing both forms endemic to the African continent (Euthecodon arambourgi,Rimasuchus lloydi) and forms that were also present at that time all around theMediterranean basin and Asia (Tomistominae and Gavialoidea). The pres-ence of crocodilians in North Africa at that time also indicates the existence ofa tropical, warm and humid climate, before the onset of more arid conditionsin the region that led to the disappearance of the crocodilians and thedevelopment and expansion of the Sahara desert.
KEY WORDSReptilia,
Crocodylia, Crocodylidae, Tomistomina,
Burdigalian, Gebel Zelten,
Libya, North Africa,
climate.

INTRODUCTION
The Gebel Zelten locality of Sirte Basin(Cirenaica, Libya) (Fig. 1) consists of outcrops ofthe Marada Formation. This formation shows aseries of facies ranging from continental to fluvi-atile sediments in the south to marine (littoraland offshore) deposits in the north, which indi-cate the existence of large rivers flowing north-ward into the Tethyan basin (Savage &Hamilton 1973). This locality has yield abun-dant vertebrate remains, especially those of a richmammalian fauna including diverse groups suchas sirenians, proboscideans, hyracoids, perisso-dactyls, artiodactyls, creodonts and carnivorans(Savage & Hamilton 1973). Besides the mam-mals, this locality has also yielded other verte-brates such as fish, birds, chelonians andcrocodilians, including both long-snouted taxaand taxa of a more generalized cranial morpho-logy. They are represented mainly by fragmentarycranial elements, although there are also many
isolated postcranial bones, especially vertebrae,which cannot be assigned to any of the forms rep-resented by cranial material. Previous accounts ofthe crocodilians present at this locality reportedthe existence of at least four different taxa:Euthecodon arambourgi Ginsburg & Buffetaut,1978 (Ginsburg & Buffetaut 1978; Tchernov1986), Rimasuchus lloydi Fourtau, 1918(Tchernov 1976, 1986; Storrs 2003), Tomistomadowsoni Fourtau, 1918 (Savage & Hamilton1973; Tchernov 1986), and an undetermined“gavialid” (Ginsburg & Buffetaut 1978;Buffetaut 1982).In the present paper, additional material from thesame locality is reported and described. Fossilsare housed at the Department of Earth Sciences,University of Bristol (BRSUG) and the Muséumnational d’Histoire naturelle, Paris (MNHN).The evidence indicates the existence of a tomis-tomine (Tomistoma lusitanica) together with apreviously undescribed brevirostrine form. Allthese elements point to the existence of a diverse
Llinás Agrasar E.
310 GEODIVERSITAS • 2004 • 26 (2)
RÉSUMÉLes restes de crocodiles provenant du Burdigalien (Miocène inférieur) de GebelZelten (Libye).Le gisement de Gebel Zelten, en Cyrénaïque, Libye nord-centrale(Burdigalien, Miocène inférieur) a produit de nombreux restes de crocodi-liens. Même fragmentaires, quelques formes peuvent être distinguées, parmicelles-ci les longirostrines Euthecodon arambourgi Ginsburg & Buffetaut,1978 et Tomistoma lusitanica Antunes, 1967 et le brevirostrine Rimasuchuslloydi Fourtau, 1918. Il y a aussi d’autres restes dans les collections provenantde ce gisement qui ne peuvent être identifiées avec confiance parce qu’ils sonttrès fragmentaires. Néanmoins, ces restes indiquent la présence d’au moins unautre taxon, un brevirostrine pas encore identifié qui diffère de Rimasuchuslloydi dans la morphologie de sa mâchoire. La faune de crocodiliens provenantde ce gisement montre les ressemblances avec celle du Burdigalien du WadiMoghara (Egypte), indiquant l’existence d’un assemblage de crocodiliens enl’Afrique du Nord durant le Miocène inférieur et montrant les deux formesendémiques d’Afrique (Euthecodon arambourgi, Rimasuchus lloydi) et lesformes qui étaient présentes aussi à l’époque autour du bassin méditerranéenet de l’Asie (Tomistominae et Gavialoidea). La présence de crocodiliens enAfrique du Nord à cette époque indique également l’existence d’un climattropical, chaud et humide, antérieur à des conditions plus arides dans larégion qui ont mené à la disparition des crocodiliens et au dévelopement et àl’augmentation du désert du Sahara.
MOTS CLÉSReptilia,
Crocodylia, Crocodylidae, Tomistomina,
Burdigalien, Gebel Zelten,
Libye, Afrique du Nord,
climat.

DESCRIPTION
Snout (Fig. 2)The snout is represented by three fragments:MNHN.LBE.300, representing the anterior partof the snout; MNHN.LBE.301, representing itsposterior part; and MNHN.LBE.302, whichincludes only a small fragment of the anteriorportion of the snout. The premaxillae are almostcompletely preserved, lacking only their anterior-most portion, including the anterior border ofthe external naris. This is heart-shaped and its lat-eral borders converge backward. The posterodor-sal processes of the premaxillae reach backward tothe level of the posterior border of the third max-illary alveolus. The premaxillae are not expandedon its anterior part and there is not notch behindthe first maxillary alveolus, unlike the conditionseen in gavialoids. The foramen incisivum is notpreserved. Only the last three pairs of premaxil-lary alveoli are preserved. Of this, the anterior-most is the largest, the following one is slightlysmaller and the posteriormost one is the smallest,being clearly smaller than the two other. Thepremaxillary alveoli are well apart from eachother, being the lateral margin of the premaxillaeindented.
Miocene crocodiles from Libya
311GEODIVERSITAS • 2004 • 26 (2)
*
AFRICA
Libya*
FIG. 1. — Map of Africa showing location of the Gebel Zeltenlocality (*) in Northern Libya.
crocodilian fauna in a tropical climatic regimeduring the Burdigalian of North Africa.
MATERIALS
MATERIAL HOUSED IN BRSUGCrocodylidae indet. 1: BRSUG 27360: partialskull table accompanied by fragmentary occipitaland quadrate regions and braincase.
MATERIALS HOUSED IN MNHNTomistoma lusitanica: MNHN.LBE.300: anteriorportion of snout; MNHN.LBE.301: posteriorportion of snout; MNHN.LBE.302: fragment ofanterior part of snout.Crocodylidae indet. 1: MNHN.LBE.311: com-plete skull table accompanied by almost completeoccipital region and braincase and fragmentaryright quadrate region.Crocodylidae indet. 2: MNHN.LBE.306: almostcomplete skull table (lacking part of the rightpostorbital and squamosal) accompanied by theright quadrate region and a fragmentary occipitalregion lacking the basioccipital; MNHN.LBE.307:anterior part of right dentary; MNHN.LBE.308:posterior part of a right lower jaw. All these spec-imens were found in association, possibly comingfrom a single individual.
SYSTEMATICS
CROCODYLIFORMES Clark in Benton & Clark, 1988
MESOEUCROCODYLIA Whetstone & Whybrow, 1983
NEOSUCHIA Clark in Benton & Clark, 1988EUSUCHIA Huxley, 1875
CROCODYLIA Gmelin, 1789Family CROCODYLIDAE Cuvier, 1807
Subfamily TOMISTOMINAE Kälin, 1955Genus Tomistoma Müller, 1846
Tomistoma lusitanica Antunes, 1961MATERIAL EXAMINED. — MNHN.LBE.300,MNHN.LBE.301 and MNHN.LBE.302.
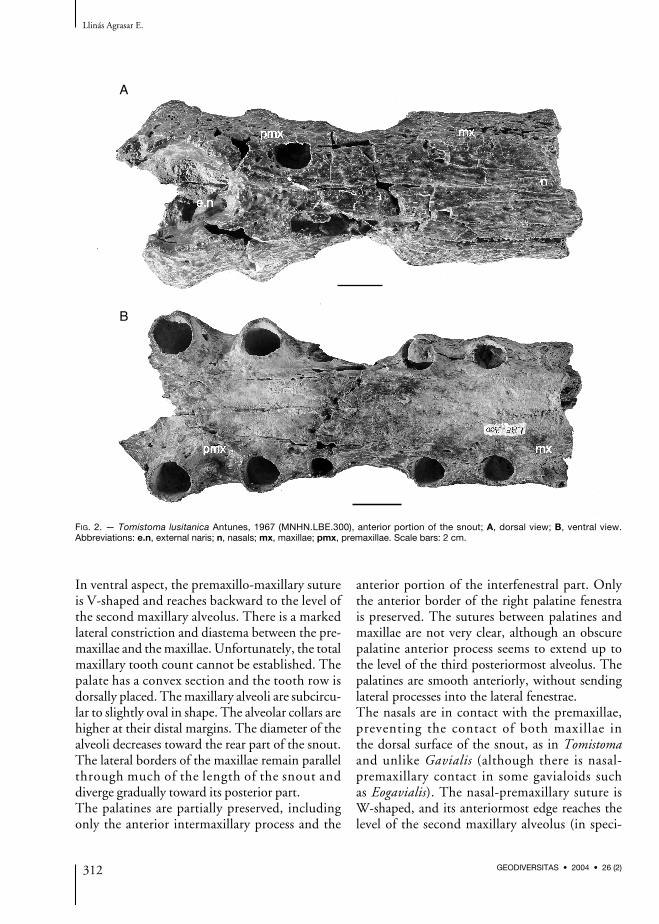
In ventral aspect, the premaxillo-maxillary sutureis V-shaped and reaches backward to the level ofthe second maxillary alveolus. There is a markedlateral constriction and diastema between the pre-maxillae and the maxillae. Unfortunately, the totalmaxillary tooth count cannot be established. Thepalate has a convex section and the tooth row isdorsally placed. The maxillary alveoli are subcircu-lar to slightly oval in shape. The alveolar collars arehigher at their distal margins. The diameter of thealveoli decreases toward the rear part of the snout.The lateral borders of the maxillae remain parallelthrough much of the length of the snout anddiverge gradually toward its posterior part.The palatines are partially preserved, includingonly the anterior intermaxillary process and the
anterior portion of the interfenestral part. Onlythe anterior border of the right palatine fenestrais preserved. The sutures between palatines andmaxillae are not very clear, although an obscurepalatine anterior process seems to extend up tothe level of the third posteriormost alveolus. Thepalatines are smooth anteriorly, without sendinglateral processes into the lateral fenestrae.The nasals are in contact with the premaxillae,preventing the contact of both maxillae inthe dorsal surface of the snout, as in Tomistomaand unlike Gavialis (although there is nasal-premaxillary contact in some gavialoids suchas Eogavialis). The nasal-premaxillary suture isW-shaped, and its anteriormost edge reaches thelevel of the second maxillary alveolus (in speci-
Llinás Agrasar E.
312 GEODIVERSITAS • 2004 • 26 (2)
A
B
FIG. 2. — Tomistoma lusitanica Antunes, 1967 (MNHN.LBE.300), anterior portion of the snout; A, dorsal view; B, ventral view.Abbreviations: e.n, external naris; n, nasals; mx, maxillae; pmx, premaxillae. Scale bars: 2 cm.

men MNHM.LBE.302, it reaches the level of thefirst one). The nasals are isolated from the poste-rior border of the external naris by the premaxil-lae. The nasals remain moderately narrow andparallel through much of the length of the snoutbefore becoming gradually broader and divergenttoward the rear part. The dorsal surface of thesnout is slightly sculptured by numerous smallshallow pits.There are certain features (nasal-premaxillarycontact, enlarged anterior maxillary alveoli,absence of a neat expansion of the premaxillaeforming a notch behind the first premaxillaryalveolus) that are reminiscent of a tomistominecrocodylid. The material is very similar to that ofT. dowsoni Fourtau, 1918 from the Burdigalianof Wadi Moghara, Egypt, and seems to confirmprevious reports of this form at Gebel Zelten(Savage & Hamilton 1973; Tchernov 1986).However, Brochu (Brochu & Gingerich 2000)synonymized all the early Miocene forms of theMediterranean basin previously referred to asTomistoma (including T. dowsoni) under thename T. lusitanica Antunes, 1961. Therefore,I refer the present material to T. lusitanicaAntunes, 1961.
Family CROCODYLIDAE Cuvier, 1807
CROCODYLIDAE indet. 1
MATERIAL EXAMINED. — BRSUG 27360 andMNHN.LBE.311.
DESCRIPTION
Skull table (Fig. 3)The skull table is rectangular in shape, wider thanlong. Its surface is transversely concave and itsornamentation consists of small irregular pits.The posterior margin of the skull table is almostlinear and horizontal. Its lateral margins are par-allel to subparallel, their anterior (postorbital)margins deflect medially toward the orbits. Thebars formed by the postorbitals and squamosalsare narrow. The plate between the supratemporalfossae is so narrow that the ornamentation on its
surface is formed by a low longitudinal ridge.This ridge runs backward to the posterior marginof the skull table over the dorsal surface of theparietal and supraoccipital. The dermal bones ofthe skull table do not overhang the borders of thesupratemporal fossae.The supratemporal fenestrae are wide and subrec-tangular in shape. The orbito-temporal foramen,situated in the posterior wall of the supratempo-ral fenestra, is large and is bordered by thesquamosal, quadrate and parietal.The postorbital constitutes about half the lengthof the lateral border of the supratemporal fenes-tra, and extends posteriorly about half the lengthof the lateral borders of the skull table. Its antero-lateral corner is rounded and overhangs the baseof the postorbital bar. The postorbital bar is par-tially preserved. There is a large nutrient foramenon its base.The squamosal is also large. Its dorsal surface hasa depression on its posterolateral corner separat-ing it from the large posterolateral squamosalprojection. On its lateral aspect, the squamosalforms the dorsal wall of the otic meatus. It has ashallow longitudinal groove for the attachment ofthe external ear valve musculature whose bordersremain parallel before flaring anteriorly towardthe postorbital. In lateral view, the postorbital-squamosal suture is oriented ventrally and it doesnot pass medially ventral to the skull table. Thesquamosal-quadrate suture extends dorsally alongthe posterior margin of the external auditorymeatus.The parietal is posteriorly wide. It narrows at theinterfenestral space and its anteriormost portionhas a short length. Only the posterior part of thefrontal is preserved. It is excluded from the ante-rior border of the supratemporal fossa by theparietal and the postorbitals. The fronto-parietalsuture is concavo-convex, with the convex sidetoward the parietal. The dorsal surface of the pos-terior part of the frontal is concave, especially inthe interorbital plate, which is wider than theinterfenestral plate. Only the posterior and pos-teromedial borders of the orbits are preserved.They were larger than the supratemporal fenes-trae and directed upward and slightly outward.
Miocene crocodiles from Libya
313GEODIVERSITAS • 2004 • 26 (2)

Quadrate region (Figs 4; 6)The articular surface of the quadrate is badlyeroded but it seems to have been divided in twohemicondyles (medial and lateral). The quadrateforms the floor and much of the anterior andposterior walls of the external auditory meatus,where it contacts the squamosal. The posteriormargin of the otic aperture is invaginate. There isa foramen preoticum just in front of the externalotic aperture. The distal end of the quadratojugalis also preserved. It abuts the lateral border of thequadrate and reaches backward just to the level ofthe lateral articular hemicondyle. Its lateral sur-face seems to have been sparsely sculptured. The quadratojugal did not extend dorsally tocontact the squamosal and the quadrate thatformed the dorsocaudal border of the infratem-
poral fenestra. The quadrate contacted the pos-torbital at the medial aspect of the base of thepostorbital bar. The foramen aereum cannot beobserved; however, a shallow groove is visible onits usual position on the dorsomedial surface ofthe quadrate. On its ventral surface, the quadratepresents a longitudinal crest running for aboutthree fourths of its length and reaching its distalend, near which it thickens. The quadrate abutsthe lateral wall of the braincase and shows a sharpridge near the basisphenoid.
Occipital region (Fig. 5)The occipital surface is slightly inclined back-ward, but not to the degree seen in gavialids.Dorsal to the foramen magnum, its posterior sur-face is vertical. The exoccipital borders of the
Llinás Agrasar E.
314 GEODIVERSITAS • 2004 • 26 (2)
po
f
q
exoc
boc
p
soc
stf
sqp
sq
pop
FIG. 3. — Crocodylidae indet. 1 (MNHN.LBE.311), skull table in dorsal view. Abbreviations: boc, basioccipital condyle; exoc, exoccipital; f, frontal; p, parietal; po, postorbital; pop, paraoccipital process; q, quadrate; soc, supraoccipital; sq, squamosal;sqp, squamosal projection; stf, supratemporal fenestra. Scale bar: 2 cm.

foramen magnum and the basioccipital are visiblein dorsal view. The supraoccipital is large and subtriangular inshape, being far wider than high. Its dorsal bor-der is clearly concave. The supraoccipital extendsdownward to about two thirds the distance to thedorsal border of the foramen magnum, fromwhich it is excluded by the exoccipitals. Thesupraoccipital has a low median vertical crestaccompanied by a shallow depression on eachside of it. Toward both of its dorsolateral ends,the supraoccipital shows the two large tuberosi-ties of the ventromedial borders of the postempo-ral fenestrae. The postemporal fenestrae arebordered by the squamosals, the supraoccipital,the exoccipitals and the parietal bone. In dorsalview, the supraoccipital is triangular in outline,
being approximately as long as wide, and it occu-pies the central quarter of the posterior margin ofthe skull table.The squamosals also participate in the occiput.The squamosals form the posterolateral projec-tions over the paraoccipital processes of the exoc-cipitals, although these projections do not reachtheir distal extremities. The occipital view of thesquamosal extends along its contact with theexoccipital to the ventrolateral corner of thesupratemporal fenestra. The exoccipitals have long paraoccipital processesthat are devoid of sculpture on their surface andextend for a long distance lateral to the entrance ofthe cranio-quadrate passage. The foramen mag-num is elliptical in shape, wider than tall. The ex-occipitals form the dorsal and lateral borders of
Miocene crocodiles from Libya
315GEODIVERSITAS • 2004 • 26 (2)
f
pob
q
boc
ls
bo
bs
m.e.f
po
stf
FIG. 4. — Crocodylidae indet. 1 (MNHN.LBE.311), skull table and braincase in ventral view. Abbreviations: bo, basioccipital;boc, basioccipital condyle; bs, basisphenoid; f, frontal; ls, laterosphenoid; m.e.f, median eustachian foramen; po, postorbital;pob, postorbital bar; q, quadrate; stf, supratemporal fenestra. Scale bar: 2 cm.

that foramen. The ventral processes of the exoc-cipitals are short and cover only the dorsalmostpart of the lateral borders of the exoccipital. Thehypoglossal foramen is small and is placed dorso-medially to the foramen vagus. It is oval-shapedwith acute ends and very deep. The foramencaroticum posterious is also oval-shaped and deepand it is situated immediately ventral to theformer and over the suture with the quadrate.The basioccipital forms the basioccipital condyleand the ventral border of the foramen magnum.The basioccipital condyle is subcircular in outline.The basioccipital shelf is tall and does not form theprominent tuberosities seen in gavialoids. It pres-ents a prominent vertical median crest. The lateraleustachian canals opened dorsolaterally to the me-dial one. They are placed at the level of the lowestpoint of the ventral process of each exoccipital.The median eustachian canal opened in the ven-tral aspect of the basisphenoid, just anterior to thesuture with the basioccipital.
Braincase (Figs 4; 6)Its walls are formed by the parietal, the lat-erosphenoids, the quadrates, the exoccipitals, theprootics, the basisphenoid and the basioccipital.The foramen ovale is wide and its dorsal and pos-terior borders extend posterodorsally as a shallowrecess over the posterior margin of the laterosphe-noid and the anterior margin of the quadrate.The laterosphenoids are relatively well preserved.They form the anterior border of the foraminaovale. Their capitate processes are orientedanteroposteriorly toward the middline and theydo not seem to have been very long. The lateralwing-like process of the laterosphenoid contactsthe base of the postorbital bar medially and itsanterior border is slightly constricted.The basisphenoid is incompletely preserved, lack-ing the basisphenoid rostrum. Only a very smallpart of the pterygoids is preserved, covering thelateral surfaces of the basisphenoid. The caudalpart of the basisphenoid placed between the
Llinás Agrasar E.
316 GEODIVERSITAS • 2004 • 26 (2)
f.m
exoc
soc
ptf
boc
pop
cqp
bo
sqp
sqp
FIG. 5. — Crocodylidae indet. 1 (MNHN.LBE.311), occipital region. Abbreviations: bo, basioccipital; boc, basioccipital condyle;cqp, cranio-quadrate passage; exoc, exoccipital; f.m, foramen magnum; p, parietal; pop, paraoccipital process; ptf, postemporalfenestra; soc, supraoccipital; sq, squamosal; sqp, squamosal projection. Scale bar: 2 cm.

posterior border of the pterygoids and the anteri-or border of the basioccipital is thick and notreduced to a thin lamina, but it is not extensivelyexposed on the braincase wall anterior to themedian eustachian foramen. The basisphenoid isnot broadly exposed ventral to the basioccipital.The quadrate-pterygoid suture is linear from thebasisphenoid to the foramen ovale and there isnot significant ventral quadrate process on thelateral wall of the braincase.
CROCODYLIDAE indet. 2
MATERIAL EXAMINED. — MNHN.LBE.306,MNHN.LBE.307 and MNHN.LBE.308.
DESCRIPTION
Skull table and associated regions (Fig. 7A)The skull table is almost as wide as long. Its later-al borders converge anteriorly. The surface of the
skull table is nearly flat and sculptured by numer-ous wide but shallow pits that are ovate in shape.The supratemporal fenestrae are small and sub-oval. Their lateral margins are clearly longer thanthe medial ones. The interfenestral plate and thelateral borders of the skull table are broad. Thesquamosals form about two thirds of the lengthof the lateral borders of the skull table, the post-orbitals making up the other third. The antero-lateral margin of the postorbital is broadlyrounded. The frontal is excluded from the anteri-or border of the supratemporal fenestrae by theparietal and the postorbitals. The fronto-parietalsuture is concavo-convex, with the convexity fac-ing toward the parietal. The frontal is slightlydepressed with respect to the postorbitals.The posterior margin of the skull table is slightlyconcave. The parietal and the supraoccipitalmake up its central half and the squamosals makeup the other. The posterior part of the parietal is
Miocene crocodiles from Libya
317GEODIVERSITAS • 2004 • 26 (2)
e.a.m
pob
Is
f.o
bo
boc
bs
f
posq
q
FIG. 6. — Crocodylidae indet. 1 (MNHN.LBE.311), skull table and braincase in right lateral view. Abbreviations: bo, basioccipital;boc, basioccipital condyle; bs, basisphenoid; e.a.m, external auditory meatus; f, frontal; f.o, foramen ovale; ls, laterosphenoid;po, postorbital; pob, postorbital bar; q, quadrate; sq, squamosal. Scale bar: 2 cm.
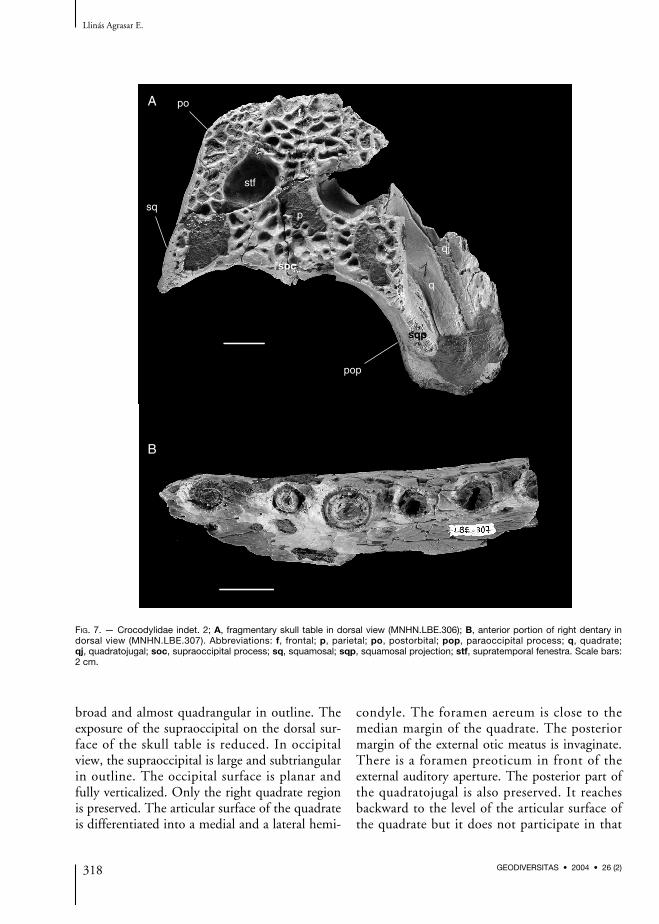
broad and almost quadrangular in outline. Theexposure of the supraoccipital on the dorsal sur-face of the skull table is reduced. In occipitalview, the supraoccipital is large and subtriangularin outline. The occipital surface is planar andfully verticalized. Only the right quadrate regionis preserved. The articular surface of the quadrateis differentiated into a medial and a lateral hemi-
condyle. The foramen aereum is close to themedian margin of the quadrate. The posteriormargin of the external otic meatus is invaginate.There is a foramen preoticum in front of theexternal auditory aperture. The posterior part ofthe quadratojugal is also preserved. It reachesbackward to the level of the articular surface ofthe quadrate but it does not participate in that
Llinás Agrasar E.
318 GEODIVERSITAS • 2004 • 26 (2)
fpo
sq
stf
p
soc
pop
stf
p
soc
pop
sqp
q
qj
A
B
po
sq
f
FIG. 7. — Crocodylidae indet. 2; A, fragmentary skull table in dorsal view (MNHN.LBE.306); B, anterior portion of right dentary indorsal view (MNHN.LBE.307). Abbreviations: f, frontal; p, parietal; po, postorbital; pop, paraoccipital process; q, quadrate;qj, quadratojugal; soc, supraoccipital process; sq, squamosal; sqp, squamosal projection; stf, supratemporal fenestra. Scale bars:2 cm.

surface. Its lateral surface is ornamented withlarge and shallow pits, although the ornamenta-tion has somewhat been eroded.
Lower jaw (Fig. 7B)Only the right dentary and splenial are fragmen-tary preserved. The mandibular ramus is robust.The total dentary tooth count cannot be deter-mined with certainty. The dentary symphysis isshort and its rear part is damaged, so its exactposterior extent cannot be determined, althoughit probably extended backward to the level of thefourth or fifth dentary tooth. It is unclearwhether the splenial participated in the sym-physis, although this seems unlikely. Althoughshort, the dentary symphysis is clearly more elong-ated than that of R. lloydi or C. niloticus. Theventral border of the symphysis is nearly flat andhorizontal, resembling that of R. lloydi. The ante-rior fragment of the lower jaw preserves six alve-oli (second-seventh), all of which are subcircularin outline. The second alveolus was apart fromthe first one, which is not preserved. It is alsoclearly apart from the third, this being the great-est interalveolar distance. From the third alveolusbackward, all the alveoli are clearly apart fromeach other and regularly spaced, including thosefrom the fourth to the seventh. This is in sharpcontrast with the condition observed in R. lloydior C. niloticus, in which these alveoli are closetogether (in some individuals of C. niloticus thesealveoli are slightly apart and there are pits for thereception of the first maxillary teeth slightly labi-ally to them; there are no such pits in the presentspecimen).The posterior fragment of the lower jaw preservesseven alveoli, most probably those from the ninthto the 15th. All are apart from each other but thedistance is not too great in any case. They are cir-cular to subcircular in outline and there are nogreat differences in size among them. The sple-nial forms the medial border of the last threealveoli, unlike in R. lloydi or C. niloticus. The lat-eral border of the lower jaw is strongly festooned.The fourth dentary alveolus occupies the highestpoint in the first convexity, whereas the 11thoccupies that of the second. The ventrolateral
surface of the dentary is sculptured with longitu-dinal grooves and small pits.
DISCUSSION
Despite its fragmentary nature, the crocodilianfauna at Gebel Zelten seems to have been diverse.Other crocodilian faunas from the Burdigalian ofNorth Africa, such as that at Wadi Mogahara(Egypt), were also diverse, with at least four dif-ferent taxa, including both longirostrine and bre-virostrine forms (Fourtau 1918).Euthecodon arambourgi Ginsburg & Buffetaut,1978 is the oldest species of its genus, datingfrom the Burdigalian of Libya. It may be repre-sented by fragmentary remains of the same agefrom other North African localities such as WadiMoghara in Egypt (Fourtau 1918). It is replacedin the Pliocene by E. nitriae Fourtau, 1920, pres-ent at Wadi Natrum (Fourtau 1920; Tchernov1986) (this form could be also present in thePliocene of Sahabi, Libya; however the materialreferred from this locality is too fragmentary topermit an identification at species level; Hecht1987). Between both forms there is a consider-able time gap spanning the middle and lateMiocene, although the genus is represented byfragmentary jaw material from some localitiesscattered across North Africa, such as Beglia(Tunisia) (Pickford 2000; Llinás Agrasar 2003).In East Africa, the genus is already present in theearly Miocene of Kenya, although the fragmen-tary nature of its remains renders an identifica-tion at specific level impossible (Tchernov & VanCouvering 1978; Buffetaut 1979). In the Plio-Pleistocene it is represented by the speciesE. brumpti Joleaud, 1920 (Joleaud 1920;Tchernov 1976, 1986; Storrs 2003) whichbecomes extinct in the middle Pleistocene(Tchernov 1986; Storrs 2003).Rimasuchus lloydi Fourtau, 1918 was first foundat Wadi Moghara, Egypt and described under thename Crocodylus lloydi Fourtau, 1918. It is alsopresent in Gebel Zelten (Savage & Hamilton1973; Tchernov 1976, 1986; Storrs 2003) andat a slightly later date in the early Miocene of
Miocene crocodiles from Libya
319GEODIVERSITAS • 2004 • 26 (2)

Namibia and South Africa (Pickford 1996). Thename C. lloydi was also applied to materials col-lected in younger beds (Plio-Pleistocene) in EastAfrica (Tchernov 1976, 1986; Pickford 1994;Leakey et al. 1996), but some authors regardedthe tangibility of some of these identifications asquestionable (Brochu 1997, 2000). However,they are not so different from the early Mioceneform, the only differences being their larger sizeand greater robustness, an even more brevi-rostrine snout (and a more bulldog-like aspect),an external naris with lateral borders convergingmore clearly backward and a wider and more lat-erally open premaxillo-maxillary notch. All thesedifferences could well be the result of intraspe-cific variability due to the broad geographic range(and presumably habitat diversity) of this species.Recent cladistic analysis (Brochu 1997, 2000;Brochu & Gingerich 2000) show that this formdoes not belong to the genus Crocodylus and inhis review of this form, Storrs (2003) erected anew genus, Rimasuchus Storrs, 2003 to refer to it.Besides Rimasuchus lloydi, there seems to beanother brevirostrine crocodile at Gebel Zelten(MNHN.LBE.306, 307 and 308; see above).This seems to differ from the former in the mor-phology of its lower jaw, having a more elongatedsymphyseal region, alveoli fourth to seventhclearly set apart from each other and a splenialthat forms the medial border of the last threealveoli. However, due to its fragmentary condi-tion, very little can be said about its affinities.Tomistomine remains are known from earlyMiocene deposits all around the Mediterranean.Although they were given many names depend-ing on their place of origin, all these forms seemto have been the same animal and they were syn-onymized by Brochu (Brochu & Gingerich2000) under the name Tomistoma lusitanicaAntunes, 1961. In North Africa, they are foundat Wadi Moghara, Egypt (Fourtau 1918), andGebel Zelten (Savage & Hamilton 1973). Theonly other record from a putative African tomis-tomine from a later date is that of Tomistomacoppensi Pickford, 1994 from the late Miocene-early Pliocene of Uganda and Zaire (Pickford1994).
The presence of this crocodilian fauna points tothe existence of tropical climatic conditions inthe Early Miocene of North Africa (Markwick1998; Pickford 2000) before the onset of morearid climatic conditions and the early stages ofthe development of the Sahara desert (Pickford2000).
Acknowledgements I wish to thank Michael J. Benton (BRSUG),Éric Buffetaut (CNRS, Paris), Sandra Chapman(The Natural History Museum, London),Phillipe Taquet, Nathalie Bardet, Xavier Pereda-Suberbiola, Stéphane Jouve, France de Lapparentde Broin, Bernard Battail and Martin Pickford(MNHN) for all the help and assistance providedfor this study, valuable comments and correc-tions, and for granting me both access to speci-mens under their care and their precious time.I also thank the two reviewers, ChristopherA. Brochu (University of Iowa) and AngelaD. Buscalioni (Universidad Autónoma deMadrid), whose valuable comments and criti-cisms helped to improve the quality of this paper.I also wish to thank Simon Powell (PhotographicUnit, BRSUG) for his help and assistance withphotography and Denis Serrette (MNHN), whotook the pictures of the specimens housed atMNHN. Funding was provided by a grant fromthe Bob Savage Memorial Fund (BRSUG) to theauthor.
REFERENCES
BENTON M. J. & CLARK J. M. 1988. — Archosaurphylogeny and the relationships of the Crocodylia,in BENTON M. J. (ed.), The Phylogeny andClassification of the Tetrapods. Volume 1:Amphibians, Reptiles and Birds . SystematicAssociation Special Volume No. 35a. ClarendonPress, Oxford: 295-338.
BROCHU C. A. 1997. — Morphology, fossils, diver-gence timing, and the phylogenetic relationships ofGavialis. Systematic Biology 46: 479-522.
BROCHU C. A. 2000. — Phylogenetic relationshipsand divergence timing of Crocodylus based on mor-phology and the fossil record. Copeia 2000:657-673.
Llinás Agrasar E.
320 GEODIVERSITAS • 2004 • 26 (2)

BROCHU C. A. & GINGERICH P. D. 2000. — Newtomistomine crocodylian from the Middle Eocene(Bartonian) of Wadi Hitan, Fayum province,Egypt. Contributions from the Museum of Paleon-tology, The University of Michigan, Ann Arbor 30:251-268.
BUFFETAUT É. 1979. — Présence du crocodilienEuthecodon dans le Miocène inférieur d’Ombo(golfe de Kavirondo, Kenya). Bulletin de la Sociétégéologique de France 21: 321-322.
BUFFETAUT É. 1982. — Systématique, origine et évo-lution des Gavialidae sud-américains. Geobiosmémoire spécial 6: 127-140.
FOURTAU R. 1918. — Contribution à l’étude desvertébrés miocènes de l’Egypte. Geological Survey ofEgypt 22-26: 1-121.
FOURTAU R. 1920. — Contribution à l’étude desvertébrés miocènes de l’Egypte. Geological Survey ofEgypt 22-26:1-121.
GINSBURG L. & BUFFETAUT É. 1978. — Euthecodonarambourgi n. sp., et l’évolution du genreEuthecodon, crocodilien du Néogène d’Afrique.Géologie méditerranéenne 5: 291-302.
HECHT M. K. 1987. — Fossil snakes and crocodiliansfrom the Sahabi Formation of Libya, in BOAZ N. T.(ed.), Neogene Paleontology and Geology of Sahabi.Alan R. Liss, Inc., New York: 101-106.
JOLEAUD L. 1920. — Sur la présence d’un gavialidé dugenre Tomistoma dans le Pliocène d’eau douce del’Éthiopie. Comptes Rendus de l’Académie desSciences, Paris 170: 816-818.
LEAKEY M. G., FEIBEL C. S., BERNOR R. L., HARRIS J.M., CERLING T. E., STEWARD K. M., STORRS G.W., WALKER A., WERDELIN L. & WINKLER A. J.1996. — Lothagam: a record of faunal change inthe late Miocene of East Africa. Journal of VertebratePaleontology 16: 556-570.
LLINÁS AGRASAR E. 2003. — New fossil crocodiliansfrom the middle/upper Miocene of Tunisia. Annalesde Paléontologie 89: 103-110.
MARKWICK P. 1998. — Fossil crocodilians as indica-tors of Late Cretaceous and Cenozoic climates:implications for using palaeontological data inreconstructing palaeoclimate. PalaeogeographyPalaeoclimatology Palaeoecology 137: 207-271.
PICKFORD M. 1994. — Late Cenozoic Crocodiles(Reptilia: Crocodylidae) from the Western Rift,Uganda, in PICKFORD M. & SENUT B. (eds),Geology and Palaeobiology of the Albertine Rift Valley,Uganda-Zaire, Volume 2: Palaeobiology. CIFFEGOccasional Publications, Orléans: 137-155.
PICKFORD M. 1996. — Fossil crocodiles (Crocodyluslloydi) from the lower and middle Miocene ofSouthern Africa. Annales de Paléontologie 82:235-250.
PICKFORD M. 2000. — Crocodiles from the BegliaFormation, middle/late Miocene Boundary,Tunisia, and their significance for Saharan palaeo-climatology. Annales de Paléontologie 86: 59-67.
SAVAGE R. J. G. & HAMILTON W. R. 1973. —Introduction to the Miocene mammal faunas ofGebel Zelten, Libya. Bulletin of the British Museum(Natural History), Geology 22: 515-527.
STORRS G. W. 2003. — Late Miocene-early Pliocenecrocodilian fauna of Lothagam, Southwest Turkanabasin, Kenya, in LEAKEY M. G. & HARRIS J. M.(eds), Lothagam: the Dawn of Humanity in EasternAfrica. Columbia University Press, New York:137-159.
TCHERNOV E. 1976. — Crocodilians from the LateCenozoic of the Rudolf Basin, in COPPENS Y.,HOWELL F. C., ISAAC G. L. & LEAKEY R. E. F.(eds), Earliest Man and Environments in the LakeRudolf Basin. University of Chicago Press, Chicago:370-378.
TCHERNOV E. 1986. — Evolution of the crocodiles inEast and North Africa. Cahiers de Paléontologie, 65 p.
TCHERNOV E. & VAN COUVERING J. 1978. — Newcrocodiles from the early Miocene of Kenya.Palaeontology 21: 857-867.
Submitted on 12 March 2003;accepted on 27 October 2003.
Miocene crocodiles from Libya
321GEODIVERSITAS • 2004 • 26 (2)