cryopreparation provides new insight into the effects of...
TRANSCRIPT

Journal of Structural Biology 130, 63–72 (2000)doi:10.1006/jsbi.2000.4230, available online at http://www.idealibrary.com on
Cryopreparation Provides New Insight into the Effects of Brefeldin Aon the Structure of the HepG2 Golgi Apparatus
Michael W. Hess,*,†,1 Martin Muller,‡ Paul L. Debbage,† Monika Vetterlein,†,§ and Margit Pavelka†,§
*Institute of Biotechnology, University of Helsinki, Helsinki, Finland; †Institute of Anatomy and Histology, University of Innsbruck,Innsbruck, Austria; ‡Laboratory for Electron Microscopy 1, Swiss Federal Institute of Technology/ETH–Zurich, Zurich, Switzerland;
and §Institute of Histology and Embryology II, University of Vienna, Vienna, Austria
Received October 9, 1999, and in revised form January 24, 2000
wbmssmcm
tsfifipfnbstrmctaiohspt
fl
dputBfiTest(BrwcGa(t1cstcleptacGn
mtf(
IFm
High-pressure freezing and freeze-substitutionere used to study Golgi ultrastructure and itsrefeldin A-induced transformations in HepG2 hu-an hepatoma cells. Cryoimmobilization arrested
ubcellular dynamics within milliseconds, thus con-iderably improving the temporal resolution inonitoring the very early effects of high brefeldin
oncentrations at the ultrastructural level (i.e., 20g/ml brefeldin applied for 35 s to 8 min). Moreover,
his approach ruled out possible cumulative and/orynergistic effects of the drug and fixatives. Severalndings differed from studies based on chemicalxation. In particular, Golgi breakdown did notroceed gradually but occurred in distinct steps. Weound a conspicuous lag between the absence ofonclathrin coats on Golgi membranes after 30 s ofrefeldin treatment and the disassembly of thetacks, which did not start until after 90 to 120 s. Athis time, domains at the trans and cis faces sepa-ated from the stacks, starting tubulation and frag-entation. After 3–5 min the Golgi apparatus was
ompletely replaced by loose meshworks of straightubules of different sizes and staining properties;lso frequent were bent tubules and vesicles form-ng glomerule-like structures. After 8 min all kindsf Golgi-derived structures had aggregated withinuge clusters. The morphologically highly distincttructures found after brefeldin treatment could inart be correlated with particular Golgi domains inhe control cells. r 2000 Academic Press
Key Words: cryofixation; drug effects; endosomes;reeze-substitution; high-pressure freezing; mamma-ian Golgi apparatus.
1 To whom correspondence should be addressed at EM Unit,nstitute of Biotechnology, University of Helsinki, P.O. Box 56,IN-00014 Helsinki, Finland. Fax: 1358-9-191 59498. E-mail:
63
INTRODUCTION
Brefeldin A (BFA) is a fungal metabolite, which,ue to its effects on the assembly of membrane coatroteins and on vesicle formation, has providednique insights into mechanisms of intracellularransport (see Klausner et al., 1992, for a review).FA blocks pre-Golgi and intra-Golgi vesicular traf-c by inhibiting the formation of transport vesicles.his is probably due to inhibition of nucleotidexchange on ADP-ribosylation factor (ARF), a neces-ary step preceding the assembly of coatomer duringhe formation of coat protein I (COP I)-coated vesiclesDonaldson et al., 1992; Helms and Rothman, 1992).lockage of vesicular traffic is accompanied by aapid tubulation of Golgi cisternae, trans-Golgi net-ork (TGN), and endosomal compartments (Lippin-
ott-Schwartz et al., 1991); as a consequence theolgi apparatus disappears and Golgi componentsre redistributed into the endoplasmic reticulumER; Lippincott-Schwartz et al., 1989, 1990). Explora-ion of Golgi dynamics in living cells (Sciaky et al.,997) has shown that BFA-induced tubulation oc-urs during a period of 5–10 min and is followed by audden and rapid disassembly of the Golgi appara-us (within 15–30 s), followed by enrichment of itsonstituents, including the membrane proteins andipids, in the ER. Many details of the BFA-inducedvents are perfectly known, but several importantoints require further investigation. In particularhey concern the very early events that take placefter BFA administration, as well as the processesonnected with the sudden disappearance of theolgi apparatus and the redistribution of its compo-ents.Electron microscopy has provided valuable infor-ation on BFA-induced ultrastructural changes of
he Golgi apparatus. The first transformations wereound to occur very early after BFA administrationPavelka and Ellinger, 1993), apparently coinciding
ith the removal of coat proteins from Golgi mem-1047-8477/00 $35.00Copyright r 2000 by Academic Press
All rights of reproduction in any form reserved.

bRcttHaseabtltpl1sisafie3caenwTl
fptamsm
m(gw1iSq(1s3p
s2Hw
epcicsFTdssss
U
sscnpssacmascr(5mcmotcusSaebatwtfrnw
B
64 HESS ET AL.
ranes (Donaldson et al., 1990; Orci et al., 1991;obinson and Kreis, 1992). Electron microscopy alsoonfirmed subsequent tubulations and revealed dis-inct glomerular structures that persist in BFA-reated cells (De Lemos-Chiarandini et al., 1992;endricks et al., 1992; Hidalgo et al., 1992; Pavelkand Ellinger, 1993). However, chemical fixation is toolow to resolve properly the rapid transformations ofndomembrane compartments occurring immedi-tely after BFA administration and during redistri-ution of Golgi constituents into the ER. To visualizehese events, we have used a technique that immobi-izes subcellular constituents much more rapidlyhan does chemical fixation. We applied high-ressure freezing (Moor, 1987), which arrests subcel-ular dynamics in less than half a second (Sitte,996). This allowed precise timing of the drug expo-ure and the moment of fixation. This approachmproved the temporal resolution in depicting tran-ient structures (Knoll et al., 1987) but also ruled outny cumulative or synergistic effects of drugs andxatives. We were able to discriminate between veryarly BFA effects on Golgi membrane coats (i.e., after0 s) and the onset of dissociation of the stacks ofisternae after 90–120 s. It was possible to differenti-te between various kinds of ramifying tubulesmerging after 2–3 min from TGN and Golgi cister-ae, respectively. After 8 min, the tubular mesh-orks disappeared; instead, all kinds of Golgi- andGN-derived structures were found to be accumu-
ated in prominent intracellular clusters.
MATERIALS AND METHODS
Cell culture. HepG2 human hepatoblastoma cells (HB-8065rom the American Type Culture Collection, Manassas, VA) werelated onto carbon-coated sapphire disks (diameter 3 mm, 50 µmhick, from R. Brugger AG, Minusio Locarno, Switzerland) suit-ble for high-pressure freezing (Schwarb, 1990) and cultivated ininimal essential medium supplemented with 10% fetal calf
erum (both from Gibco, Grand Island, NY). Subconfluent cellonolayers were achieved after 3 days.Brefeldin A treatment and high-pressure freezing. The speci-ens were transferred for 0 s or 20 s to 7.5 min to culture medium
at 137°C) containing 20 µg/ml BFA (from Epicentre Technolo-ies, Madison, WI). Subsequently, BFA-treated and control samplesere cryoimmobilized by high-pressure freezing (HPF; Moor,987) according to Studer et al. (1989). Briefly, the specimens weremmersed for a few seconds in 1-hexadecene (from Fluka, Buchswitzerland), an inert paraffin oil essential for HPF, and subse-uently mounted into the sample holder of a HPF apparatusBAL-TEC HPM 010, Balzers, Liechtenstein). This procedure took5 to 20 s at maximum and was recorded separately for eachample; thus, the specimens were exposed to BFA for a total of 0 s,5 s, 50 s, 60 s, 75 s, 90 s, 120 s, 3 min, 4 min, 5 min, and 8 minrior to cryoimmobilization.Freeze-substitution and electron microscopy. The frozen
amples were freeze-substituted for at least 8 h at approximately90°C with acetone containing 1% (w/v) OsO4 (modified after Vanarrefeld and Crowell, 1964) followed by optional en bloc staining
ith 0.02% (w/v) uranyl acetate in acetone (30 min at 14°C) and imbedded in Epon. Sections (80–100 nm thick) were cut in thelane of the monolayer and collected on uncoated or formvar-oated copper grids. The sections were poststained by immersionn aqueous uranyl acetate (30 min at 125°C) followed by leaditrate (3 min at 125°C) and examined with an electron micro-cope at 60–100 kV (EM10A, Zeiss, Oberkochen, Germany; CM120,EI-Philips, Eindhoven, The Netherlands; JEM-1200 EX, Jeol,okyo, Japan). With respect to membrane staining no majorifferences were observed as to whether or not samples had beenubjected to en bloc staining. It was noteworthy, however, thatections thinner than approximately 80 nm or poststained on oneide only suffered from poor membrane staining of the Golgitacks, in contrast to the TGN and any other membranes.
RESULTS
ntreated HepG2 Control Samples
The Golgi apparatus of HepG2 cells consisted ofeveral clearly polarized stacks displaying discreteubsets of cisternae. The typically fenestrated cisisternae (Fig. 1), with their wide lumina of 50–60m, frequently showed irregular contours when cuterpendicularly. The membranes did not alwayshow a distinct contrast against the moderatelytained cisternal contents or the cytoplasm. Vesiclesnd tubules located in close vicinity to the cisisternae presumably are part of the pre-Golgi inter-ediate compartment (Fig. 1a; see also Ladinsky et
l., 1999). The medial cisternae (Fig. 1) showedtrongly stained membranes that enclosed lumina ofonstant width of 36 nm. Electron-dense, fuzzy mate-ial coated their luminal face. The trans cisternaeFig. 1) showed irregular outlines and wide lumina of0–60 nm. Fenestrations were common. The limitingembranes were clearly visible but lacked internal
oatings. The trans-most cisterna displayed at itsargins branching tubules with average diameters
f 40–80 nm, which sometimes appeared to peel offhe stack (Fig. 1); they regularly showed clathrin-oated buds and vesicles (Fig. 1b). These profiles aresually referred to as TGN. ER was often seenandwiched between the trans cisternae (Fig. 1).ometimes the space between the cisternae and thessociated ER was spanned at various points bylectron-dense material (possibly ‘‘intercisternalridges’’ according to Cluett and Brown (1992); seelso Senda and Yoshinaga-Hirabayashi (1998)). Mul-iple tubular and vesicular membrane profiles asell as buds at the rims of all Golgi cisternae (except
he trans-most) regularly displayed electron-dense,uzzy coats (Fig. 1); according to morphological crite-ia they are considered to be COP coats (see Ban-ykh and Balch, 1998). Endosomes and lysosomesere abundant.
refeldin A—35 s
At this first time point no changes were observed
n the architecture of the Golgi stacks or of the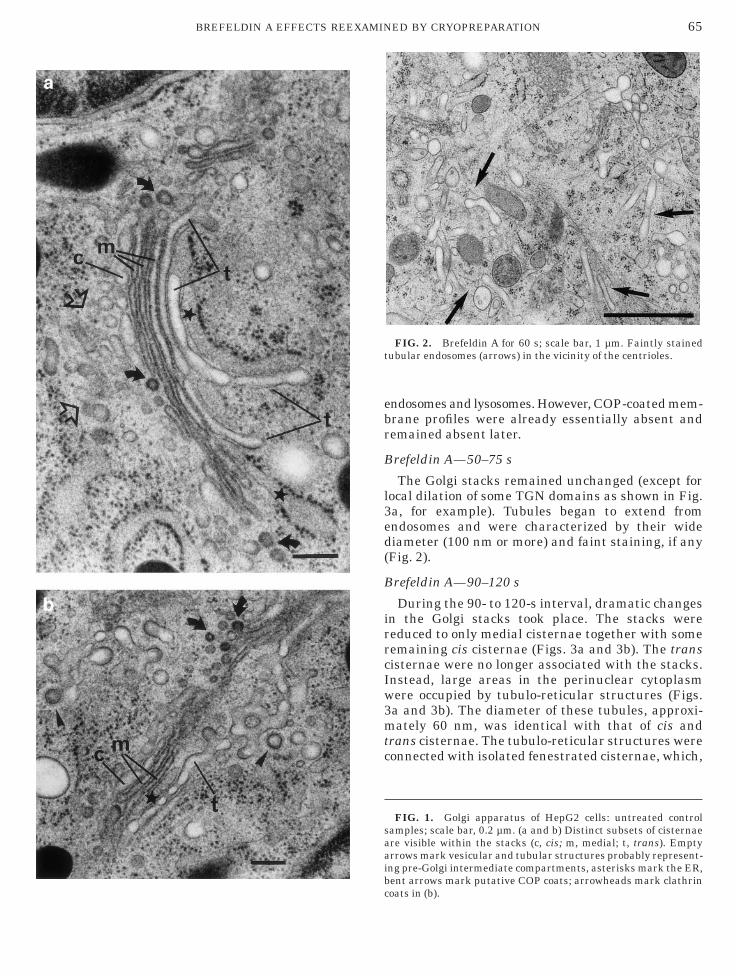
ebr
B
l3ed(
B
irrcIw3mtc
saaib
t
65BREFELDIN A EFFECTS REEXAMINED BY CRYOPREPARATION
c
ndosomes and lysosomes. However, COP-coated mem-rane profiles were already essentially absent andemained absent later.
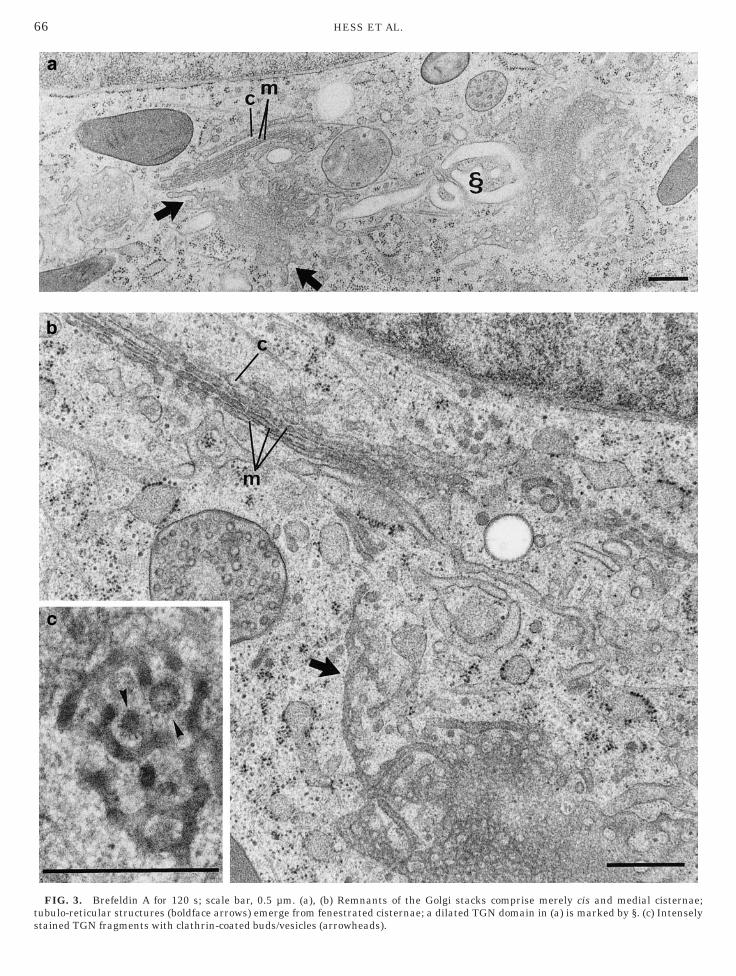
refeldin A—50–75 s
The Golgi stacks remained unchanged (except forocal dilation of some TGN domains as shown in Fig.a, for example). Tubules began to extend fromndosomes and were characterized by their wideiameter (100 nm or more) and faint staining, if anyFig. 2).
refeldin A—90–120 s
During the 90- to 120-s interval, dramatic changesn the Golgi stacks took place. The stacks wereeduced to only medial cisternae together with someemaining cis cisternae (Figs. 3a and 3b). The transisternae were no longer associated with the stacks.nstead, large areas in the perinuclear cytoplasmere occupied by tubulo-reticular structures (Figs.a and 3b). The diameter of these tubules, approxi-ately 60 nm, was identical with that of cis and
rans cisternae. The tubulo-reticular structures wereonnected with isolated fenestrated cisternae, which,
FIG. 1. Golgi apparatus of HepG2 cells: untreated controlamples; scale bar, 0.2 µm. (a and b) Distinct subsets of cisternaere visible within the stacks (c, cis; m, medial; t, trans). Emptyrrows mark vesicular and tubular structures probably represent-ng pre-Golgi intermediate compartments, asterisks mark the ER,ent arrows mark putative COP coats; arrowheads mark clathrin
FIG. 2. Brefeldin A for 60 s; scale bar, 1 µm. Faintly stainedubular endosomes (arrows) in the vicinity of the centrioles.
oats in (b).
ts
66 HESS ET AL.
FIG. 3. Brefeldin A for 120 s; scale bar, 0.5 µm. (a), (b) Remnants of the Golgi stacks comprise merely cis and medial cisternae;ubulo-reticular structures (boldface arrows) emerge from fenestrated cisternae; a dilated TGN domain in (a) is marked by §. (c) Intenselytained TGN fragments with clathrin-coated buds/vesicles (arrowheads).

itntstduw
wc
B
stss
a
67BREFELDIN A EFFECTS REEXAMINED BY CRYOPREPARATION
nterestingly, were sectioned predominantly tangen-ially in contrast to perpendicularly cut stack rem-ants in their vicinity (Figs. 3a and 3b). Nearby,here could be observed interconnected tubules ofmaller diameter, approximately 40 nm, and sincehese bore clathrin-coated buds they evidently wereerived from the TGN (Fig. 3c). In contrast tontreated TGN elements, however, their contentsere electron-dense (Fig. 3c) and their membranes
FIG. 4. Brefeldin A for 3 min; scale bar, 0.5 µm. The Golgi appa
nd branching 60-nm tubules; double arrowheads mark intensely stainedere frequently remarkably distinct. Endosomesontinued to extend tubules.
refeldin A—3–5 min
Three minutes after brefeldin application the Golgitacks had disrupted completely. Rearrangements ofhe disrupted elements resulted in glomerule-liketructures (Figs. 4 and 5a), consisting of moderatelytained, bent 50- to 60-nm tubules together with
s replaced by glomerule-like structures (marked by empty arrows)
ratus i TGN remnants.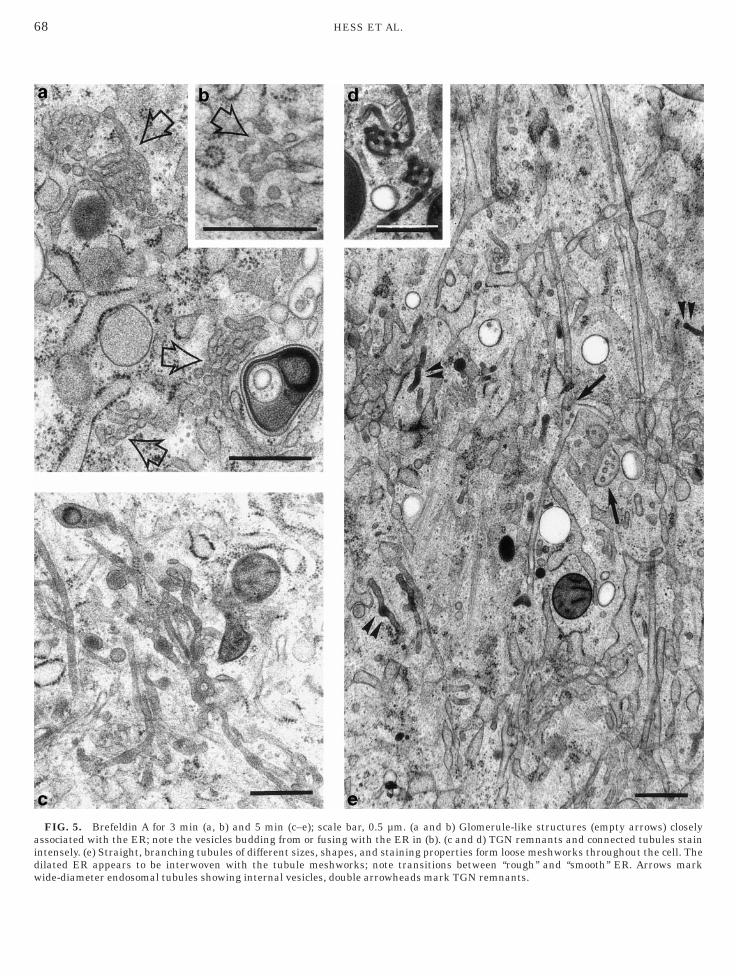
aidw
68 HESS ET AL.
FIG. 5. Brefeldin A for 3 min (a, b) and 5 min (c–e); scale bar, 0.5 µm. (a and b) Glomerule-like structures (empty arrows) closelyssociated with the ER; note the vesicles budding from or fusing with the ER in (b). (c and d) TGN remnants and connected tubules stainntensely. (e) Straight, branching tubules of different sizes, shapes, and staining properties form loose meshworks throughout the cell. Theilated ER appears to be interwoven with the tubule meshworks; note transitions between ‘‘rough’’ and ‘‘smooth’’ ER. Arrows mark
ide-diameter endosomal tubules showing internal vesicles, double arrowheads mark TGN remnants.
vdpaBbwfawoeoeqnacrasvcaadtaocssmno
B
bc1bsCµlrpst
iwt
TbattmctwoCtretfkfa1
plvc
69BREFELDIN A EFFECTS REEXAMINED BY CRYOPREPARATION
esicles (Fig. 5a); some of these vesicles were bud-ing off from, or fusing with, closely associated ERrofiles (Fig. 5b). The glomerule-like structures hadn average diameter of 0.6 µm and about 3 min afterFA application they appeared in the Golgi region,ut by 5 min they were dispersed throughout thehole cell. A variety of straight tubule types now also
ormed loose meshworks throughout the whole cell,nd their characteristics indicated that they tooere derived from the Golgi stacks (Figs. 4 and 5e);ne tubule type was approximately 60 nm in diam-ter and stained moderately (Figs. 4 and 5e). An-ther type was approximately 40 nm in diameter andlectron-dense (Figs. 4 and 5e); this type was fre-uently continuous with conspicuously stained cister-al fragments (Figs. 5c and 5d), which were presum-bly remnants of the TGN (see also Fig. 3c), thoughlathrin-coated buds and vesicles were only veryarely observed. These loose meshworks of tubulesnastomosing with one another (Fig. 5e) and repre-enting remnants of the Golgi stacks were interwo-en with the dilated ER (Fig. 5e) but never becameontinuous with it. During this interval, 3–5 minfter BFA application, the wide-diameter tubularnd/or pleiomorphic compartments (100–200 nm),erived from endosomes, assumed their final posi-ions mainly in the cell periphery and concentratedround the centrioles. Their lumina were unstainedr faintly stained (Figs. 5e and 6) and occasionallyontained internal vesicles, distinguishing them fromimilarly shaped compartments derived from lyso-omes, which contained readily stained fine granularaterial (Fig. 6) and sometimes membrane rem-ants. Tubules were often paralled by microtubulesr even connected to them via bridge-like structures.
refeldin A—8 min
At this time point ramifying straight tubulesecame less frequent. By contrast, there emergedlusters comprising bent tubules measuring 60 or00 nm in diameter, together with structures resem-ling fenestrated cisternae. These moderately stainedtructures (Fig. 7a) were interwoven with the ER.lose to the clusters, which reached diameters of 2m or more, there regularly were located glomerule-
ike structures (Fig. 7a) and sometimes also TGNemnants (Fig. 7b). Interestingly, the clusters alsoersisted during longer incubation times (data nothown). The patterns of endosomal and lysosomalubules remained unchanged.
DISCUSSION
The present results show that high-pressure freez-ng and freeze-substitution preserve particularlyell the delicate ultrastructure of the Golgi appara-
us in HepG2, a widely used mammalian cell model.
he subtle differences thus revealed between mem-ranes (and also luminal contents) of the cis, medial,nd trans cisternae highlighted morphological dis-inctions between the Golgi subcompartments. Fur-hermore we provided detailed ultrastructural infor-ation on the very early effects of high BFA
oncentrations: a variety of morphologically distinctubular and cisternal endomembrane compartmentsere depicted with an accuracy that cannot bebtained with conventional specimen preparation.ryoimmobilization with subsequent stabilization of
he frozen samples by means of freeze-substitutionuled out possible cumulative and/or synergisticffects of the drug and fixatives (e.g., osmotic effects);he morphological patterns observed should there-ore reliably reflect genuine drug effects. To ournowledge, similar investigations have been per-ormed so far only with plants and fungi (Bourettnd Howard, 1996; Domozych, 1999; Driourich et al.,993, 1997; Ueda et al., 1996; Zhang et al., 1993).
FIG. 6. Brefeldin A for 5 min; scale bar, 1 µm. Tubular and/orleiomorphic wide-diameter compartments in the cell periphery:ysosomes (double arrows) with clearly stained granular contentsersus faintly to moderately stained endosomes (single arrows)haracterized by internal vesicles.
The accuracy of our morphological data encour-

acldaidscdtmnBtpltbam
aotmssOtas‘Tgfpfapct
c ants (d
70 HESS ET AL.
ges us to correlate some of the endomembraneompartments in BFA-treated HepG2 with particu-ar Golgi domains in the control cells. Electron-ense, clathrin-decorated cisternal fragments andpproximately 40-nm-wide tubules, occurring fromncubation periods of 90 s onward, are most likelyerived from the TGN/trans-most cisterna. Theytained intensively in drug-treated cells but not inontrol cells, indicating an accumulation of proteinsue to arrest in intracellular transport. The persis-ence of clathrin coats on transforming Golgi do-ains for as long as 90–120 s of BFA treatment hasot been reported previously (see, e.g., Wong androdsky, 1992) and visualization of these fragile
ubules is probably due to the improved samplereservation achieved with cryopreparation. Iso-ated fenestrated cisternae with tubulo-reticular ex-ensions were transient structures occurring mainlyetween 90 s and approximately 3 min. They presum-bly represent modified trans cisternae and to a
FIG. 7. Brefeldin A for 8 min; scale bar, 0.5 µm. Mixed clusisternae; glomerule-like structures (open arrows) and TGN remn
inor extent cis cisternae that have shifted off the m
xis of the stack. This is indicated by the abundancef tangentially cut profiles of such isolated struc-ures in the vicinity of perpendicularly cut stackededial cisternae; such patterns were almost never
een in identically oriented and thus comparableections through the Golgi region of control cells.ne is tempted to speculate that this breakdown of
he stacks’ 3-dimensional organization might bettributable to alterations of the Golgi membranekeleton (see, e.g., Beck et al., 1994, 1997) and/or of‘intercisternal bridges’’ (Cluett and Brown, 1992).he 50- to 60-nm tubules and vesicles forminglomerule-like structures after 3 min may be derivedrom the pre-Golgi intermediate compartment andossibly from the cis-most cisterna as well, as judgedrom their structural characteristics, i.e., size, shape,nd staining properties. Assuming that these inter-retations concerning TGN, pre-Golgi intermediateompartments, and cis-most cisterna are correct,hen the majority of the moderately stained, anasto-
onsisting of different kinds of tubules and putative fenestratedouble arrowheads) are regularly associated as well.
ters c
osing 60-nm tubules should represent transforma-

tiimtGmLpGnf
nntctcApfGeqfotrfww
himictcsHem
ctlfa(IsamB
vddPasurccttdrm(tfmmd1
odatdGbatri3fm(w
KsHa
B
B
B
71BREFELDIN A EFFECTS REEXAMINED BY CRYOPREPARATION
ions of all other Golgi domains. It remains to benvestigated whether there exist possibly short-livedntermediates between medial cisternae and anasto-
osing tubules similar to the tubulo-reticular struc-ures. The prominent clusters made up of all kinds ofolgi-derived structures that emerged after 8 minay correspond to ‘‘residual Golgi elements’’ (Deemos-Chiarandini et al., 1992); it has been pro-osed that such persistent structures are involved inolgi recovery (De Lemos-Chiarandini et al., 1992), aotion also supported by our own preliminary datarom recovery experiments.
Taken together, it is clear that further work iseeded to complement our morphological data,amely to verify the proposed relationships betweenhe endomembrane compartments in BFA-treatedells and Golgi domains in the controls. However,his involves the development of tools that areompatible with the technique of freeze-substitution.s to cytochemistry, for example, one is limited toostembedding labeling. This approach is still farrom being well established for the localization ofolgi markers in animal cells. Low-temperaturembedding with Lowicryl resins is in many casesuite suitable for labeling; such samples, however,ail to display the very subtle ultrastructural detailsf the Golgi apparatus (see Monaghan et al., 1998)hat can be seen in our preparations. Likewise,eplacing OsO4 with uranyl acetate or tannic acidollowed by ambient-temperature embedding in LR-hite, Unicryl, or Epon proved unsatisfactory asell (data not shown).Another aspect that merits further investigation is
ow BFA-induced redistribution of Golgi moleculesnto the ER is brought about. We did not find any
embrane continuities between the ER and BFA-nduced compartments, apart from the vesicles asso-iated with the glomerule-like structures. Accordingo recent studies on BFA effects carried out in livingells, multiple fusion events might not be a prerequi-ite for Golgi–ER redistribution (Sciaky et al., 1997).owever, ultrastructural localization of marker mol-
cules in cryoprocessed specimens should yield aore differentiated insight into this process as well.Finally, it seems worth mentioning that the use of
ryoimmobilization improved the temporal resolu-ion in monitoring drug effects at the ultrastructuralevel. The timing of nonclathrin coat disappearancerom the Golgi membranes in our cryosamples (i.e.,fter 30 s) matched well the existing data on b-COPDonaldson et al., 1990; Robinson and Kreis, 1992).t is amazing, however, to note that the Golgi stacksubsequently withstood major structural changes fornother 60–90 s—as did clathrin-coated TGN do-ains—despite the high concentration of 20 µg/ml
FA employed. Once transformations became clearlyisible, breakdown of the Golgi stacks proceeded inistinct steps. These features differ from previousata obtained from chemically fixed samples (e.g.,avelka and Ellinger, 1993). On the contrary, theyre consistent with the kinetics of ‘‘explosive dissas-embly/Golgi blinkout’’ recorded in living cells bysing green fluorescent protein–Golgi protein chime-as (Sciaky et al., 1997). Together, our finding of aonspicuous lag between the disappearance of COPoats and the first signs of Golgi stack transforma-ions should be taken into account when consideringhe hypotheses on the causalities of BFA action. Theata support the idea that Golgi breakdown does notesult merely from the perturbance of coatomeretabolism. Thus, other Golgi-associated proteins
e.g., spectrin, ankyrin: Beck et al., 1994, 1997) andheir possible interactions with a NAD1-dependentactor(s) (Mironov et al., 1997; Spanfo et al., 1999)ight also play key roles in this process—as in Golgiaintenance in general (for further references and
iscussion, see Mironov et al., 1997; Sciaky et al.,997; Spanfo et al., 1999).
CONCLUSION
Cryotechniques represent a considerable method-logical improvement in unraveling the complexynamics of the Golgi apparatus under physiologicalnd experimentally altered conditions with highemporal and spatial resolution. Summarizing ourata on BFA-induced disassembly of the HepG2olgi apparatus, three major, distinct phases coulde recognized, following the immediate disappear-nce of COP coats after 30 s: (i) after 90–120 s of BFAreatment domains at the trans and cis faces sepa-ated from the remaining stack and started tubulat-ng, and this process also included the TGN; (ii) after–5 min the Golgi apparatus had completely trans-ormed into glomerule-like structures, loose tubuleeshworks, and fragments of trans-Golgi/TGN; and
iii) after 8 min all kinds of Golgi-derived structuresere found to aggregate within huge clusters.
The authors thank Eeva-Liisa Eskelinen, Eija Jokitalo, Esauismanen, Jaakko Saraste, and Paul Walther for helpful discus-
ions and for critically reading the manuscript and thank Rudolfaring, Mervi Lindman, and Tuire Koro for excellent technicalssistance.
REFERENCES
annykh, S. I., and Balch, W. E. (1998) Selective transport ofcargo between the endoplasmic reticulum and Golgi apparatus,Histochem. Cell Biol. 109, 463–475.
eck, K. A., Buchanan, J. A., Malhotra, V., and Nelson, W. J.(1994) Golgi spectrin: Identification of an erythroid beta-spectrin homolog associated with the Golgi complex, J. CellBiol. 127, 707–723.
eck, K. A., Buchanan, J. A., and Nelson, W. J. (1997) Golgi
membrane skeleton: Identification, localization and oligomeriza-
B
C
D
D
D
D
D
D
H
H
H
K
K
L
L
L
L
M
M
M
O
P
R
S
S
S
S
S
S
U
V
W
Z
72 HESS ET AL.
tion of a 195 kDa ankyrin isoform associated with the Golgicomplex, J. Cell Sci. 110, 1239–1249.
ourett, T. M., and Howard, R. J. (1996) Brefeldin A-inducedstructural changes in the endomembrane system of a filamen-tous fungus, Magnaporthe grisea, Protoplasma 190, 151–163.
luett, E. B., and Brown, W. J. (1992) Adhesion of Golgi cisternaeby proteinaceous interactions: Intercisternal bridges as puta-tive adhesive structures, J. Cell Sci. 103, 773–784.e Lemos-Chiarandini, C., Ivessa, N. E., Black, V. H., Tsao, Y. S.,Gumper, I., and Kreibich, G. (1992) A Golgi-related structureremains after the brefeldin A-induced formation of an ER–Golgihybrid compartment, Eur. J. Cell Biol. 58, 187–201.omozych, D. S. (1999) Perturbation of the secretory network inClosterium acerosum by Na1-selective ionophores, Protoplasma206, 41–56.onaldson, J. G., Finazzi, D., and Klausner, R. D. (1992) BrefeldinA inhibits Golgi membrane-catalysed exchange of guaninenucleotide onto ARF protein, Nature 360, 350–352.onaldson, J. G., Lippincott-Schwartz, J., Bloom, G. S., Kreis,T. E., and Klausner, R. D. (1990) Dissociation of a 110kDperipheral membrane protein from the Golgi apparatus is anearly event in brefeldin A action, J. Cell Biol. 111, 2295–2306.riouich, A., Jauneau, A., and Staehelin, L. A. (1997) 7-Dehydro-brefeldin A, a naturally occurring brefeldin A derivative, inhib-its secretion and causes a cis-to-trans breakdown of Golgi stacksin plant cells, Plant Physiol. 113, 487–492.riouich, A., Zhang, G. F., and Staehelin, L. A. (1993) Effect ofbrefeldin A on the structure of the Golgi apparatus and on thesynthesis and secretion of proteins and polysaccharides insycamore maple (Acer pseudoplatanus) suspension-culturedcells, Plant Physiol. 101, 1363–1373.elms, J. B., and Rothman, J. E. (1992) Inhibition by brefeldin Aof a Golgi membrane enzyme that catalyses exchange of gua-nine nucleotide bound to ARF, Nature 360, 352–354.endricks, L. C., McClanahan, S. L., McCaffery, M., Palade, G. E.,and Farquhar, M. G. (1992) Golgi proteins persist in thetubulovesicular remnants found in Brefeldin A-treated pancre-atic acinar cells, Eur. J. Cell Biol. 58, 202–213.idalgo, J., Garcia-Navarro, R., Gracia-Navarro, F., Perez-Vilar,J., and Velasco, A. (1992) Presence of Golgi remnant membranesin the cytoplasm of Brefeldin A-treated cells, Eur. J. Cell Biol.58, 214–227.lausner, R. D., Donaldson, J. G., and Lippincott-Schwartz, J.(1992) Brefeldin A: Insights into the control of membrane trafficand organelle structure, J. Cell Biol. 116, 1071–1080.noll, G., Verkleij, A. J., and Plattner, H. (1987) Cryofixation ofdynamic processes in cells and organelles, in Steinbrecht, R. A.,and Zierold, K. (Eds.), Cryotechniques in Biological ElectronMicroscopy, pp. 258–271, Springer-Verlag, Berlin/Heidelberg.
adinsky, M. S., Mastronarde, D. N., McIntosh, J. R., Howell,K. E., and Staehelin, L. A. (1999) Golgi structure in threedimensions: Functional insights from the normal rat kidneycell, J. Cell Biol. 144, 1135–1149.
ippincott-Schwartz, J., Yuan, L. C., Bonifacio, J. S., and Klaus-ner, R. D. (1989) Rapid redistribution of Golgi proteins into theER in cells treated with brefeldin A: Evidence for membranecycling from Golgi to ER, Cell 56, 801–813.
ippincott-Schwartz, J., Donaldson, J. G., Schweizer, A., Berger,E. G., Hauri, H-P., Yuan, L. C., and Klausner, R. D. (1990)Microtubule-dependent retrograde transport of proteins intothe ER in the presence of brefeldin A suggests an ER recyclingpathway, Cell 60, 821–836.
ippincott-Schwartz, J., Yuan, L., Tipper, C., Amherdt, M., Orci,L., and Klausner, R. D. (1991) Brefeldin A’s effects on endo-
somes, lysosomes, and TGN suggests a general mechanism forregulating organelle structure and membrane traffic, Cell 67,601–616.ironov, A., Colanzi, A., Silletta, M. G., Fiucci, G., Flati, S.,Fusella, A., Polishchuk, R., Mironov, A., Ditullio, G., Weigert, R.,Malhotra, V., Corda, D., Dematteis, M. A., and Luini, A. (1997)Role of NAD1 and ADP-ribosylation in the maintenance of theGolgi structure, J. Cell Biol. 139, 1109–1118.onaghan, P., Perusinghe, N., and Muller, M. (1998) High pres-sure freezing for immunocytochemistry, J. Microsc. 192, 248–258.oor, H. (1987) Theory and practice of high pressure freezing, inSteinbrecht, R. A., and Zierold, K. (Eds.), Cryotechniques inBiological Electron Microscopy, pp. 175–191, Springer-Verlag,Berlin/Heidelberg.rci, L., Tagaya, M., Amherdt, M., Perrelet, A., Donaldson, J. G.,Lippincott-Schwartz, J., Klausner, R. D., and Rothman, J. E.(1991) Brefeldin A, a drug that blocks secretion, prevents theassembly of non-clathrin-coated buds on Golgi cisternae, Cell64, 1183–1195.
avelka, M., and Ellinger, A. (1993) Early and late transforma-tions occurring at organelles of the Golgi area under theinfluence of brefeldinA:An ultrastructural and lectin cytochemi-cal study, J. Histochem. Cytochem. 41, 1031–1042.
obinson, M. S., and Kreis, T. E. (1992) Recruitment of coatproteins onto Golgi membranes in intact and permeabilizedcells: Effects of brefeldin A and G protein activators, Cell 69,129–138.
chwarb, P. (1990) Morphologische Grundlagen zur Zell–ZellInteraktion bei adulten Herzmuskelzellen in Kultur. Ph.D.Thesis, Swiss Federal Institute of Technology, Zurich. [Diss.ETH-Zurich, 9195]
ciaky, N., Presley, J., Smith, C., Zaal, K. J., Cole, N., Moreira,J. E., Terasaki, M., Siggia, E., and Lippincott-Schwartz, J.(1997) Golgi tubule traffic and the effects of brefeldin A visual-ized in living cells, J. Cell Biol. 139, 1137–1155.
enda, T., and Yoshinaga-Hirabayashi, T. (1998) Intermembranebridges within membrane organelles revealed by quick-freezedeep-etch electron microscopy, Anat. Rec. 25, 339–345.
itte, H. (1996) Advanced instrumentation and methodologyrelated to cryoultramicrotomy: A review, Scanning Microsc.Suppl. 10, 387–466.
panfo, S., Silletta, M. G., Colanzi, A., Alberti, S., Fiucci, G.,Valente, C., Fusella, A., Salmona, M., Mironov, A., Luini, A., andCorda, D. (1999) Molecular cloning and functional characteriza-tion of brefeldin A–ADP-ribosylated substrate, J. Biol. Chem.274, 17705–17710.
tuder, D., Michel, M., and Muller, M. (1989) High-pressurefreezing comes of age, Scanning Microsc. Suppl. 3, 253–269.eda, T., Tsukaya, H., Anai, T., Hirata, A., Hasegawa, K., andUchimiya, H. (1996) Brefeldin A induces the accumulation ofunusual membrane structures in elongating pollen tubes ofNicotiana tabacum L., J. Plant Physiol. 149, 683–689.
an Harreveld, A., and Crowell, J. (1964) Electron microscopyafter rapid freezing on a metal surface and substitution fixa-tion, Anat. Rec. 149, 381–386.ong, D. H., and Brodsky, F. M. (1992) 100-kD proteins of Golgi-and trans-Golgi network-associated coated vesicles have re-lated but distinct membrane binding properties, J. Cell Biol.117, 1171–1179.
hang, G. F., Driouich, A., and Staehelin, L. A. (1993) Effect ofmonensin on plant Golgi: Re-examination of the monensin-induced changes in cisternal architecture and functional activi-ties of the Golgi apparatus of sycamore suspension-cultured
cells, J. Cell Sci. 104, 819–831.