crystallization and some properties of isocitrate lyase ... · to 1.8 x 1o-2 m (fig. 1). this range...
TRANSCRIPT

THE JOURNAL OF ILLOGICAL CHEMISTRY
Vol. 238, No. 5, May 1963
Franted in U.S.A.
Crystallization and Some Properties of Isocitrate Lyase from
Pseudomonas indigofera*
BRUCE A. MCFADDEX AND WILLIAM V. HowEst
From the Department of Chemistry, Washington State University, Pullman, Washington
(Received for publication, December 28, 1962)
lsocitrate lyase (n,-isocitrate glyosylate-lyase, EC 4.1 .3.1) has been purified from a variety of sources (l-5), although the enzyme has not. been crystallized. In most cases workers pre- sented evidence for a requirement for divalent cation and thiol (14). Smith and Gunsalus (I) reported an equilibrium constant of 34.6 at 27”, pH 7.6, for the condensation reaction catalyzed by a partially purified enzyme from Pseudomonas aeruginosa. In a recent review Olson has described several properties of partially purified isocitrate lyases, including inhibition by glyoxylstc and succinate (6). Very little work has been reported, however, on inhibition by other metabolites.
In the present articlc, data on the purification and crystalliza- tion of isocitrate lyasc from Pseudomonas indigofera are reported. Additionally, some properties of the enzyme are described, in- cluding its sensitivity to various plausible feedback inhibitors.
EXPERIMENTAL PROCEDURE
Growth and Preparation of Ihzymic Fraction-P. indigofera was grown in a basal inorganic salts medium (7) containing 0.3% sodium butyrate and 0.1 7; Difco yeast estract at 25”. This cul- turc was used in the late logarithmic phase as an inoculum at 3 fs (volume for volume) for 40 liters of an identical medium except that the yeast extract was omitted. Sterile air was forced through the culture at 25”. The cells were harvested in the late logarithmic phase by continuous centrifugation, washed once by suspension in 0.05 M Tris, pH 7.9, and suspended in 180 ml of an identical buffer except that 3 X 1O-3 M MgCl, and 10e3 M GSH’ were added. Typically about 75 g of cells, wet weight, were ob- tained. The cell suspension was ruptured in three equal aliquot portions for 6 minutes at full power in a Raytheon lo-kc sonic oscillator with tap water coolant. Undisrupted cells and debris were removed at 30,000 x g for 30 minutes at 4”. Further centrifugation of the supernatant at 144,000 X g for 2 hours at 4” afforded the “soluble” fraction. In prior studies it has been established that isocitrate lyase activity resides quantitatively in the “soluble” fraction from ethanol-grown P. indigofera (8).
Enzyme Pur$cation-,411 of the following purification proce- dures were conducted at 4’. To the ‘Lsoluble” fraction (about I20 ml) from 40 liters of culture, 40 g of (NHJz’SO, were added, bringing the volume to 160 ml and the solution to 46 Y0 satura-
* This investigation was supported in part by Research Grant RG-9039 from the National Institutes of Health, United States Public Health Service.
t Research Corporation Fellow, 1962. Present address, Depart- ment of Biology, Massachusetts Institute of Technology, Cam- bridge 39, Massachusetts.
1 In all operations GSH was added to buffer shortly before use.
tion. After 5 minutes of stirring, the precipitate was removed by centrifugation and discarded. The supernatant solution was then brought to 64% saturation (a total of 56 g of (NHJzS04 per 169 ml), and the precipitate was collected as before. The 46 to 64oj, precipitate was dissolved in 50 ml of 0.01 ivf Tris, pH 7.9, containing 3 X lop3 M iL’IgC1, and lop3 M CSH (hereafter referred to as Tris-GSH-Mg++ buffer). The 46 to 64y0 fraction was then agitated for 45 minutes with an equal volume of calcium phosphate gel (9) containing 13 mg of solids per ml. The “gel” was removed by centrifugation and the supernatant was dialyzed overnight against 0.01 M Tris containing 3 X lo-” M MgClz and lop3 M cysteine. The dialyzed fraction was brought to 45% saturation by the addition of (NHJ2S04 solution, pH 7.9, sat- urated at 4’. The precipitate formed was collected by centrifu- Ration and discarded. The supernatant was then brought to 55% saturation by further addition of the alkaline (NH,),SO,. The precipitate was collected, dissolved in 20 ml of Tris-GSH- Mg++ buffer, and dialyzed overnight as before. The solution was then placed on a diethylaminoethyl cellulose (DEAE- cellulose) column, 1 X 25 cm, containing 2 g of DEAE-cellulose. The column had been previously washed with 150 ml of Tris- G.SH-Mg++ buffer. The enzyme fraction was washed in with about 2 ml of Tris-G-SH-Mgff buffer and an additional 10 ml of buffer were added. A gradient elution was accomplished by placing 200 ml of 0.015 M Tris, pH 7.9, containing 3 X lop3 M
MgCl, and lop3 M G-SH in the mixing chamber and 150 ml of 0.65 M Tris, pH 7.9, containing 3 X 1O-3 M CSH in the reservoir. Elution was conducted at a rate of 12 to 14 ml per hour, and 2%ml fractions were collected. The protein elution was fol- lowed by change in absorbancy at 280 rnp and enzyme assays were conducted. The three peak fractions were pooled and the two fractions on both sides of the peak were also pooled.
Crystallization of Isocitrate Lyase-.An aliquot of Ohe pooled peak fractions from the DEAE-cellulose column was diluted 1: 1 with Hz0 and was brought to 33% saturation by the addition of an (NH,),SO, solution, saturated at 4”, pH 7.9. The solution was placed in an ice bath under 4 mm of pressure Hg until the reduction in volume brought the saturation to 50 ‘$& During the 4-hour period microfine crystals separated from the solution. These crystals were collected by centrifugation, a portion was photomicrographed, and the remainder was dissolved in 0.05 M
Tris, pH 7.9, containing 3 X 1O-3 M hfgC&. The specific ac- tivity of isocitrate lyase in this solution was determined. The crystalline fraction was stored at -20” in the presence of 1O-3 M
GSH. Protein B&nation--Both GSH and cysteine were observed to
interfere with the Folin phenol (10) estimation of protein.
1737
by guest on May 19, 2020
http://ww
w.jbc.org/
Dow
nloaded from
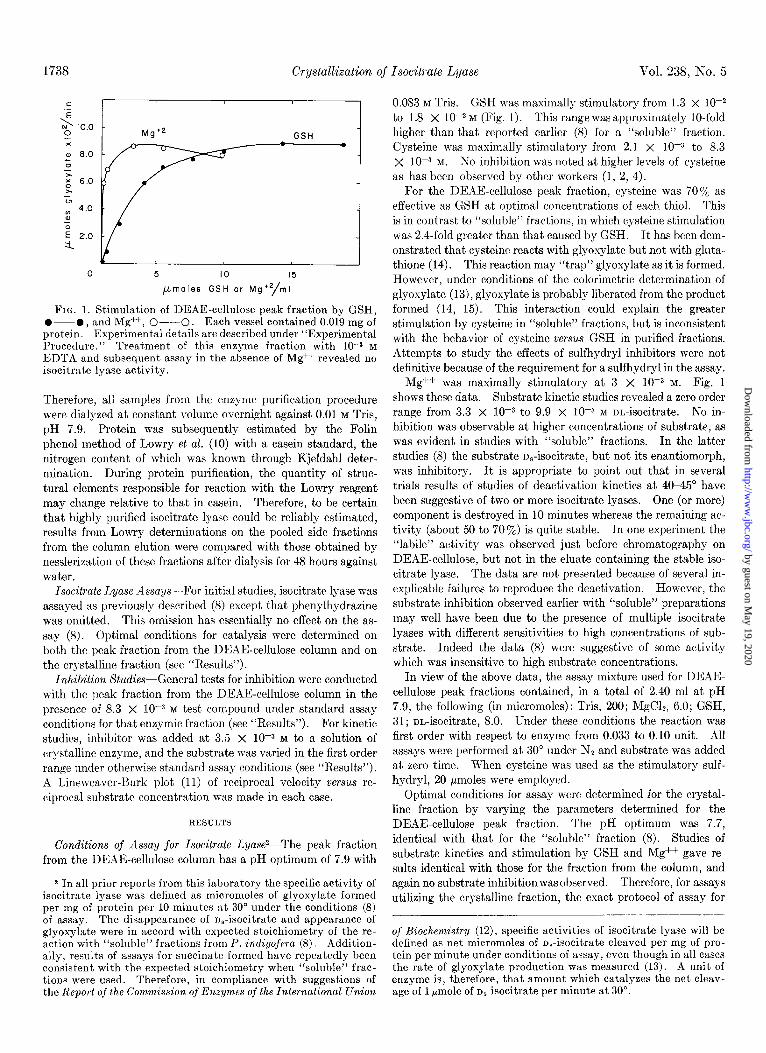
1738 Crystallization of Isocitrate Lyase Vol. 238, No. 5
Y I 0 5 IO 15
pmoles GSH or Mg”/ml
FIG. 1. Stimulation of DEAE-cellulose peak fraction by GSH, O--O, and Mg++, O--O. Each vessel contained 0.019 mg of protein. Experimental details are described under “Experimental Procedure.” Treatment of this enzyme fraction with 10-s M EDTA and subsequent assay in the absence of Mg++ revealed no isocitrate lyase activity.
Therefore, all samples from the enzyme purification procedure were dialyzed at constant volume overnight against 0.01 M Tris, pH 7.9. Protein was subsequently estimated by the Folin phenol method of Lowry et al. (10) with a casein standard, the nitrogen content of which was known through Kjeldahl deter- mination. During protein purification, the quantity of struc- tural elements responsible for reaction with the Lowry reagent may change relative to that in casein. Therefore, to be certain that highly purified isocitrate lyase could be reliably estimated, results from Lowry determinations on the pooled side fractions from the column elution were compared with those obtained by nesslerization of these fractions after dialysis for 48 hours against water.
Isocitrate T,yase Assays-For initial studies, isocitrate lyase was assayed as previously described (8) except that phenylhydrazine was omitted. This omission has essentially no effect on the as- say (8). Optimal conditions for catalysis were determined on both the peak fraction from the DE4E-cellulose column and on the crystalline fraction (see “Results”).
Inhibition Studies-G.eneral tests for inhibition were conducted with the peak fraction from the DEAF-cellulose column in the presence of 8.3 X 10e3 M test compound under standard assay conditions for that enzymic fraction (see “Results”). For kinetic studies, inhibitor was added at 3.5 X lop3 M to a solution of crystalline enzyme, and the substrate was varied in the first order range under otherwise standard assay conditions (see “Results”). A Lineweaver-Rurk plot (11) of reciprocal velocity versus re- ciprocal substrate concentration was made in each case.
RESULTS
Conditions of Assay for Isocitrate Lyase’-The peak fraction from the DEAF-cellulose column has a pH optimum of 7.9 with
2 In all prior reports from this laboratory the specific activity of isocitrste lyase was defined as micromoles of glyoxylate formed per mg of protein per 10 minutes at 30” under the conditions (8) of assay. The disappearance of n,-isocitrate and appearance of glyoxylate were in accord with expected stoichiometry of the re- action with “soluble” fractions from P. indigofera (8). Addition- ally, results of assays for succinate formed have repeatedly been consistent with the expected stoichiometry when “soluble” frac- tions were used. Therefore, in compliance with suggestions of the Report of the Commission of Enzymes of the International Union
0.083 M Tris. G.SH was maximally stimulatory from 1.3 x lop2 to 1.8 X 1O-2 M (Fig. 1). This range was approximately lo-fold higher than that reported earlier (8) for a “soluble” fraction. Cysteine was maximally stimulatory from 2.1 X 10m3 to 8.3 X IO-3 M. So inhibition was noted at higher levels of cysteine as has been observed by other workers (1, 2, 4).
For the DEAE-cellulose peak fraction, cysteine was 70% as effective as GSH at optimal concentrations of each thiol. This is in contrast to “soluble” fractions, in which cysteine stimulation was 2.4-fold greater than that caused by GSH. It has been dem- onstrated that cysteine reacts with glyoxylate but not with gluta- thione (I 4). This reaction may “trap” glyoxylate as it is formed. However, under conditions of the calorimetric determination of glyoxylate (13)) glyoxylate is probably liberated from the product formed (14, 15). This interaction could explain the greater stimulation by cysteine in “soluble” fractions, but is inconsistent with the behavior of cysteine versus GSH in purified fractions. Attempts to study the effects of sulfhydryl inhibitors were not definitive because of the requirement for a sulfhydryl in the assay.
ivIg++ was maximally stimulatory at 3 X low3 M. Fig. 1 shows these data. Substrate kinetic studies revealed a zero order range from 3.3 X lop3 to 9.9 X lop3 M m-isocitrate. No in- hibition was observable at higher concentrations of substrate, as was evident in studies with “soluble” fractions. In the latter studies (8) the substrate n,-isocitrate, but not its enantiomorph, was inhibitory. It is appropriate to point out that in several trials results of studies of deactivation kinetics at 40-45” have been suggestive of two or more isocitrate lyases. One (or more) component is destroyed in 10 minutes whereas the remaining ac- tivity (about 50 to 707,) is quite stable. In one experiment the “labile” activity was observed just before chromatography on DEAF-cellulose, but not in the eluate containing the stable iso- citrate lyase. The data are not presented because of several in- explicable failures to reproduce the deactivation. However, the substrate inhibition observed earlier with “soluble” preparations may well have been due to the presence of multiple isocitrate lyases with different sensitivities to high concentrations of sub- strate. Indeed the data (8) were suggestive of some activity which was insensitive to high substrate concentrations.
In view of the above data, the assay mixture used for DEAE- cellulose peak fractions contained, in a total of 2.40 ml at pH 7.9, the following (in micromoles): Tris, 200; MgC12, 6.0; G,SH, 31; on-isocitrate, 8.0. Under these conditions the reaction was first order with respect to enzyme from 0.033 to 0.10 unit. All assays were performed at 30” under Nz and substrate was added at zero time. When cysteine was used as the stimulatory sulf- hydryl, 20 pmoles were employed.
Optimal conditions for assay were determined for the crystal- line fraction by varying the parameters determined for the DEAE-cellulose peak fraction. The pH optimum was 7.7, identical with that for the “soluble” fraction (8). Studies of substrate kinetics and stimulation by GSH and Mg++ gave re- sults identical with those for the fraction from the column, and again no substrate inhibition was observed. Therefore, for assays utilizing the crystalline fraction, the exact protocol of assay for
of Biochemistry (12), specific activities of isocitrate lyase will be defined as net micromoles of o,-isocitrate cleaved per mg of pro- tein per minute under conditions of assay, even though in all cases the rate of glyoxylate production was measured (13). A unit of enzyme is, therefore, that amount which catalyzes the net cleav- age of 1 pmole of n,-isocitrate per minute at 30”.
by guest on May 19, 2020
http://ww
w.jbc.org/
Dow
nloaded from

May 1963 B. A. McFadden and W. V. Howes 1739
the DEAE-cellulose peak fraction was used except that assays were conducted at pH 7.7.
Protein Determination-Eecause GSH and cysteine both inter- fere with the Folin phenol protein estimation, it was necessary to dialyze the samples from the purification procedure before pro- tein estimation. This was accomplished in l-ml constant volume dialysis cells.
The pooled side fractions from the DEAE-cellulose column, after dialysis against water for 48 hours, contained 0.112 mg of nitrogen per ml by nesslerization. With the use of a factor of 6.38 for protein to nitrogen, a value of 0.71 mg of protein per ml was obtained. Protein estimation by the Folin phenol method resulted in a value of 0.70 mg of protein per ml. Therefore, it would seem that the Folin phenol method, with a casein standard, is an adequate estimate of protein in purified and dialyzed sam- ples of isocitrate lyase.
Enzyme Purification-The purification procedure and results are summarized in Table I. The relatively high specific activity of the “soluble” fraction compared with earlier studies (16) may be due, in part, to protein estimation on a dialyzed sample. In earlier studies “soluble” fractions were not prepared in the pres- ence of a thiol. Possibly low molecular weight polypeptides or other compounds, or both, interfering with the Folin phenol method were removed in the present studies and thus gave a lower protein value per unit volume. It is also possible that the inclusion of GSH in the sonication procedure stabilized the en- zyme. This latter aspect is now under investigation.
Calcium phosphate gel treatment, when tested on aliquots, generally indicated an enzyme purification in the supernatant of approximately 2-fold. When a gross treatment was used and samples for protein estimation were dialyzed, there was no change in specific activity, and a loss of 37 % of total protein was observed. However, the substantial increase in the Azso:Apc,~ ratio made this step valuable in the over-all purification procedure. Attempts to fractionate the supernatant from the calcium phosphate gel by pH adjustment resulted in totaldeactiva- tion of isocitrate lyase below pH 5.0. Little protein was pre-
TABLE I
PuriJcation of isocitrate lyase
Purification step /
Protein
COWXII tration
1. Oscillation and centrifu- gation at 144,000 X g..
2. Precipitate from 46 to 64% saturation with (NH,) zSOn.
3. Supernatant from calciun phosphate gel adsorp- tion..........
4. Precipitate from satura- tion with 45 to 55% (NH,) 2 SOa, pH 7.9.
5. DEAE-cellulose column elution, peak fractions..
6. Crystallization..
w/ml
7.2 1150 2.7 3100 0.84
5.2 260 6.3 1650 0.92
1.8
3.7
1.09 0.15
pt-cific acti- vity*
165 6.3 1050 1.21
74
9.2 1.3
8.2 610 1.50
20 185 46 60
1.72
Units
* Specific activities were determined by protocol described in “Experimental Procedure” for the DEAE-cellulose peak frac- tion.
tlsocitrate lyase A units/m, -i ‘;;-
15 .z
0.5 0.4
22 25 30 35 40
5
3
tube number FIG. 2. Gradient elution of isocitrate lyase from DEAE-cellu-
lose column. Assays for isocitrate Iyase, O--O, are described in “Experimental Procedure,” as are details of elution procedure. Protein absorbancy (A 280), O--O, was determined in l-cm cu- vettes at 280 mp.
cipitated above pH 5.0. Therefore, these attempts were discon- tinued. The saturated solution of the (NHJ2S04 used in these studies before calcium phosphate gel treatment had a pH of ap- proximately 5.5. To avoid possible acid deactivation of the enzyme, the salt solution used after gel treatment was adjusted to pH 7.9. Essentially no protein was precipitated below 40% saturation.
As can be seen in Fig. 2 and Table I, elution from the DEAE- cellulose column resulted in a 2.5.fold purification with the en- zyme peak being eluted slightly after the protein peak. The pooled peak fractions (8.4 ml) will be referred to hereafter as the peak fraction. GSH was added to the peak fraction at 10-S M,
and the fraction was stored at -20”. The enzyme was stable under these conditions for several weeks. The omission of GSH from the purification procedure resulted in a substantial loss of activity in the peak fraction. The protective action of a sulf- hydryl during purification has also been observed by Smith and Gunsalus (1).
Fig. 3 is a photomicrograph of the crystals of isocitrate lyase. Table I indicates a 2-fold purification upon crystallization. It is then of interest that ultracentrifugation at 260,000 X g of the peak fraction resulted in a single peak in the Tris-GSH-Mg++ buffer or in a medium of about 5-fold greater ionic strength, 0.10 M Tris, pH 7.7. Fig. 4 is a schlieren pattern in the Tris-GSH- Mg++ buffer. Apparently, contaminating protein in the peak column fraction sediments much like isocitrate lyase. However, the possibility that crystallization increases the catalytic activity and, therefore, that the peak fraction is homogeneous has not been excluded.
inhibition Studies-I3y use of the assay protocol described for the peak fraction, several compounds were tested at 8.3 X 1OF M as inhibitors of isocitrate lyase in the peak fraction. Table II summarizes these results. Compounds known to interfere with the calorimetric determination of glyosylate (13) were not tested.
For several of the compounds tested (Table II) and for glyox- ylate, inhibition constants were determined with the crystalline enzyme preparation. Fig. 5 is a plot of the data.
As can be noted, the slopes representing data obtained for
by guest on May 19, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Crystallization of Isocitrate Lyase
FIG. 3. Photomicrograph showing the crystalline habit of isocitrate lyase. The distance between calibration marks is 0.1 mm.
FIG. 4. Schlieren pattern of DEAE-cellulose peak fraction in the Tris-GSH-Mg++ buffer. A Spinco model E analytical ultra- centrifuge was employed. The bar angle was 30” and temperature was 23 f 1”. Sedimentation is shown from Zest to right and ex- posures were taken at 1.5, 9.5, 17.5, 33.5, and 55.5 minutes after 59,780 r.p.m. had been reached.
glyosylate, a-ketoglutarate, and L-malate are quite similar to tor from the y intercept, which equals (l/V,,,) (1 + (1)/K& that of the control. The data are thus suggestive of uncompeti- where V,,, is the maximal velocity and (I) is inhibitor molarity. tive inhibition (18) for these inhibitors. Even the extrapolated The values of KEsr at 30” were: for glyoxylate, 5.3 X 10W3 M; plot for succinate inhibition intersects the --z axis at some dis- for cy-ketoglutarate, 6.8 X 10V3 M; for L-malate, 5.9 X low3 M; tance from the extrapolated control plot. Kssr, the dissociation for succinate, 1.4 X 10m3 M. The Michaelis constant, K,, was constant for ESI = I&S’ + I, has been evaluated for each inhibi- 8.2 x lo-4 M.
by guest on May 19, 2020
http://ww
w.jbc.org/
Dow
nloaded from

May 1963 B. A. McFadden and W. V. Howes 1741
TABLE II
Effect of various compounds on isocitrate lyase-
catalyzed reaction*
Compoundt Percentage of control rate
None (control). ........................ Acetate ......... ....................... 1,.Alanine ..... ........................ 1,.AspartateI. .......................... Butyrate ................................ Citrate. ................... ............ Fumarate ............. .............. L-Glutamate. .........................
Glycine. ............................. Glycolate ............................... a-Ketoglutarate. ...................... I,-Malate .............................. Maleate. ...... ...................... Malonate ... .................... Succinate ...........................
(100) 88
>90 >90 >90
88 70
>90 80 0
01 33
0 30 18
* Assays were conducted with a DEAE-cellulose peak fraction exactly as described for that fraction in “Experimental Pro- cedure,” except that the total volume of the reaction mixture was 2.35 ml.
t Compounds were used at 8.5 X 10-a M. $ Added as the DL form at 1.7 X low2 M.
7.0
6.0
5.0
t
b -
-7,
4.0
3.0
2.0
1.0
0
/
tsuccinate
/
IlY( lxylate
nalate
tetoglutarate
inhibitor
0.5 1.0 1.5 2.0
& x 10-z
FIG. 5. Lineweaver-Burk plot of kinetic data obtained with crystalline isocitrate lyase in the absence of inhibitor, O---O, and in the presence of the following inhibitors, each at 3.5 X 1OW M: glyoxylate, O--O; a-ketoglutarate, A; L-malate, A.; and succinate, 0. For glyoxylate inhibition, succinate was deter- mined by the method of Rodgers (17). Substrate concentration is in terms of mole liter-1 and velocity in terms of mole liter-1 mine1. Other details are described in “Experimental Procedure.”
DISCUSSION
P. indigofera grown on ethanol or butyrate is an excellent source of isocitrate lyase. In general, specific activities of 0.7
to 1.2 can be expected for “soluble” fractions prepared in the presence of GSH. When samples for protein estimation were dialyzed, these values increased by approximately 28%. The specific activity of 2.7 for the “soluble” fraction used in the puri- fication procedure reported in Table I was greater than expected. Unusually high specific activities also have been observed occa- sionally in “soluble” fractions prepared from cells grown on ethanol. An explanation for this behavior is not obvious. The addition of 0.05% yeast extract to the growth medium is known to have essentially no effect on the specific activity of isocitrate lyase but does stimulate growth (19).
Several compounds were tested as possible inhibitors of iso- citrate lyase in the peak fraction from the K)EhE-cellulose col- umn. Acetate, L-alanine, L-aspartate, butyrate, citrate, L-glu- tamate, and glycine had little or no cffcct at 8.3 X lop3 M. It is
of some interest that glycine is without effect, since catalysis by isocitrate lyase has been proposed as a source of glyoxylatc, from which glycine could be derived (20). Thus glycine may not regulate its own biosynthesis by feedback inhibition (21) of iso- citrate lyase.
Kornberg and Krebs (22) first postulated that the glyosylate cycle could be used to replenish tricarbosylic acid cycle intermc- diates required for cellular syntheses during microbial growth on 2 carbon substrates. Hence inhibition of isocitrate lyase by fumarate, ar-ketoglutarate, malatc, and succinate is of some in- terest. Feedback inhibition by any of these metabolites may afford P. indigofera the capacity for fine adjustment of the ratio of oxidation by the tricarboxylic acid cycle and synthesis by way of the glyoxylate cycle. However, it is well to remember that interpretations of studies in vitro should be extrapolated to phenomena in vivo with reservation. An illustration of this is the fact that endogenous concentrations of succinate in P. indigo- fera do not vary markedly even when succinate is added to the medium, although succinate utilization occurs (19). Therefore, pronounced control of isocitratc lyase by succinate as a feedback inhibitor seems unlikely.
Inhibition by malonate and by glycolate is not too surprising since these compounds may be regarded as structural analogues of known inhibitors such as Cq dicarbosylic acids and glyoxylate, respectively. Malonate inhibits the isocitrate lyase-catalyzed incorporation of Cl4 from C%uccinate plus glyoxylate into iso- citrate (3). The striking inhibition by maleate is interesting because binding of the inhibitory Cd dicarboxylic acids in a “cis conformation” would be unexpected. Knowledge of the mecha- nism of inhibition by maleate must await studies of the mode of inhibition and of the nature of catalysis by isocitrate lyase.
Z1lthough kinetics was studied with a single inhibitor concen-
tration only, inhibition by glyoxylate, ar-ketoglutaratc, and L-
malate resembles uncompetitive inhibition. This is suggestive of binding which is dependent upon prior association of enzyme
with isocitrate (18). Each of these three inhibitors may well function as a feedback inhibitor in viva. Hence it is of interest that Gerhart and Pardee (23) have recently present,ed evidence
that a feedback inhibitor, cytidine triphosphate, of aspartic transcarbamylase is bound at a site which is completely (or at least partially) different from that for substrate. Perhaps a simi-
lar situation obtains during feedback control of isocitrate lyase, although a precise description of inhibitor-enzyme interaction must await further experimentation.
by guest on May 19, 2020
http://ww
w.jbc.org/
Dow
nloaded from

1742 Crystallization of Isocitrate Lyase Vol. 238, No. 5
SUMMARY 3. CARPENTER. W. D.. AND BEEVERS, H., Plant Physiol., 34, 403
4
5.
6.
The enzyme isocitrate lyase has been crystallized from Pseudo-
monas indigofera. The pH optimum is 7.7 and a thiol such as glutathione or cysteine is required for catalysis. Additionally a metal ion such as Mg++ is required. Highly purified enzyme is inhibited by 8.3 x 10m3 M fumarate, glycolate, oc-ketoglutarate, maleate, L-malate, malonate, and succinate. Acetate, n-alanine, L-aspartate, butyrate, citrate, L-glutamate, and glycine, each at the above concentration, have little or no effect. Inhibition of crystalline enzyme by glyoxylate, ol-ketoglutarate, or n-malate appears to be uncompetitive, with KEsI values (for ESI = ES + I) being 5.3 X lop3 M, 6.8 X 10-S M, and 5.9 X 10-s M, respectively, at 30”. KEsr for succinate is 1.4 X 10e3 M and the Michaelis constant, K,, is 8.2 x low4 M at 30”. The data thus suggest binding of plausible feedback inhibitors such as glyoxylate, ar-ketoglutarate, malate, and succinate at sites which are different from that for substrate, although the possibility of a single inhibitor binding site has not been excluded.
(1959).
L4cknowZedgments-Performance by Mr. R. M. Sylvester of the sedimentation studies is gratefully acknowledged, as are the Nessler determinations conducted by Mrs. M. E. Vogel, Depart- ment of Agricultural Chemistry. Thanks are also due Dr. D. H. Freeman for preparation of photomicrographs of isocitrate lyase crystals.
REFERENCES
1. SMITH, R. A., AND GUNSALUS, I. C., J. Biol. Chem., 229, 305 (1957).
19.
20. 21. 22. 23.
SMITH, R. A., AND GUNSALUS, I. C., Nature, 176,774 (1955). UMBARGER, H. E., Science, 123, 848 (1956). KORNBERG, H. L., AND KREBS, H. A., Nature, 179, 988 (1957). GERHART, J. C., AND PARDEE, A. B., J. Biol. Chem., 237, 891
2. OLSON, J. A., J. Biol. Chem., 234, 5 (1959). (1962).
7.
8.
9.
10.
11.
12.
13.
14. 15.
16.
17. 18.
MCCURDY, H. E., AND CANTINO, E. C., Plant Physiol., 36, 463 (1960).
KREMMETER, A. F., SCHLIEP, H. J., AND BUKATSCH, F., 2. Naturforsch., Pt. b., 16b, 678 (1961).
OLSON, J. A., in P. D. BOYER, H. LARDY, AND K. MYRBXCK (Editors), The enzymes, Vol. 5, Ed. 2, Academic Press, Inc., New York, 1961, p. 387.
MCFADDEN, B. A., AND HOWES, W. V., J. Bacterial., 81, 858 (1961).
MCFADDEN, B. A., AND H~WES, W. V., Biochim. et Biophys. Acta, 50, 179 (1961).
KEILIN, D., AND HARTREE, E. F., Proc. Roy. Sot. (London), Ser. B, 124, 397 (1937-1938).
LOWRY. 0. H.. ROSEBROUGH. N. J.. FARR. A. L., AND RANDALL, R. J.: J. B&l. Chem., 193,‘265 (1951).
LINEWEAVER, H., AND BURK, D., J. Am. Chem. Sot., 56, 658 (1934).
Report of the commission on enzymes of the international union of biochemistry, Pergamon Press, New York, 1961, p. 8.
MCFADDEN, B. A., AND HOWES, W. V., Anal. Biochem., 1,240 (1960)
GADAL, P., AND SERIS, J., Compt. rend., 264, 1341 (1962). RAO, N. A. N., AND RAMAKRISHNAN, T., Biochim. et Biophys.
Acta, 58, 262 (1962). MCFADDEN, B. A., AND HOWES, W. V., J. Bacterial., 84, 72
(1962). RODGERS, K., Biochem. J., 80,240 (1961). ALBERTY, R. A., in F. F. NORD (Editor), Advances in enzymol-
ogy, Vol. 17, Interscience Publishers, Inc., New York, 1956, p. 1.
HOWES, W. V., AND MCFADDEN, B. A., J. Bacterial., 84, 1216 (1962).
by guest on May 19, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Bruce A. McFadden and William V. Howesindigofera
PseudomonasCrystallization and Some Properties of Isocitrate Lyase from
1963, 238:1737-1742.J. Biol. Chem.
http://www.jbc.org/content/238/5/1737.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/238/5/1737.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 19, 2020
http://ww
w.jbc.org/
Dow
nloaded from