current neuropharmacology, 1-15 1 physiology and
TRANSCRIPT

Current Neuropharmacology, 2006, 4, 1-15 1
1570-159X/06 $50.00+.00 ©2006 Bentham Science Publishers Ltd.
Physiology and Pharmacology of the Vanilloid Receptor
Angel Messeguer1, Rosa Planells-Cases
2and Antonio Ferrer-Montiel
3,*
1Department of Biological Organic Chemistry, IIQAB-CSIC, J. Girona 23, 080034 Barcelona, Spain,
2Centro
Investigación Príncipe Felipe, FVIB, Av. Autopista Saler 16, 46013 Valencia, Spain, 3Instituto de Biología Molecular y
Celular, Universidad Miguel Hernández, Av. de la Universidad s/n, 03202 Elche (Alicante), Spain
Abstract: The identification and cloning of the vanilloid receptor 1 (TRPV1) represented a significant step for the
understanding of the molecular mechanisms underlying the transduction of noxious chemical and thermal stimuli by
peripheral nociceptors. TRPV1 is a non-selective cation channel gated by noxious heat, vanilloids and extracellular
protons. TRPV1 channel activity is remarkably potentiated by pro-inflammatory agents, a phenomenon that is thought to
underlie the peripheral sensitisation of nociceptors that leads to thermal hyperalgesia. Cumulative evidence is building a
strong case for the involvement of this receptor in the etiology of both peripheral and visceral inflammatory pain, such as
inflammatory bowel disease, bladder inflammation and cancer pain. The validation of TRPV1 receptor as a key
therapeutic target for pain management has thrust intensive drug discovery programs aimed at developing orally active
antagonists of the receptor protein. Nonetheless, the real challenge of these drug discovery platforms is to develop
antagonists that preserve the physiological activity of TRPV1 receptors while correcting over-active channels. This is a
condition to ensure normal pro-prioceptive and nociceptive responses that represent a safety mechanism to prevent tissue
injury. Recent and exciting advances in the function, dysfunction and modulation of this receptor will be the focus of this
review.
THE TRP RECEPTOR SUPERFAMILY
The Transient Receptor Potential (TRP) mammalian genesuperfamily consists of 28 different gene products encodingnon-selective cation channels that play a wide diversityof physiological functions [24,79]. These channels areconsidered molecular gateways in sensory systems, sinceseveral of these channels transduce chemical and physicalstimuli into neuronal activity, i.e. action potentials. Thus,TRP channels have emerged as major transducers ofchemically and physically evoked sensations.
Based upon their sequence homology, mammalian TRPchannels are divided into six subfamilies, namely TRPC,TRPV, TRPP, TRPM, TRPA and TRPML [24,79,82]. Anadditional family, TRPN, has been described in Drosophila.These proteins have a common topology of six trans-membrane segments (S1-S6) with a pore region between thefifth and sixth segment, and cytoplasmic N- and C-termini.TRPV and TRPC contain two-to-four ankyrin domains thatare thought to interact with the cytoskeleton, as well as withother cytosolic proteins [24]. TRP proteins are remarkablechannels because of the diversity of their activationmechanisms, cation selectivity and biological function. Somemembers are activated through the phospholipase C-inositoltrisphosphate pathway, although the specific mechanism ofactivation is still elusive [24,77]. Others, such as someTRPV members, are ionotropic receptors activated bychemical and physical stimuli [24,61]. The moleculardiversity of TRP proteins correlates with their wide numberof biological functions, ranging from fertility to vision, taste,olfaction, osmo/mechanosensation, and nociception [61,77,
*Address correspondence to this author at Instituto Biología Molecular yCelular, Ed. Torregaitan, Universidad Miguel Hernández, Av. de la
Universidad s/n, 03202 Elche, Alicante, Spain; Tel: +34 966658727; Fax:+34 966658758; E-mail: [email protected]
79]. Therefore, TRP channels constitute a family of sensoryreceptors.
THE TRPV1 CHANNEL
Among the TRP channel superfamily, TRPV1, TRPV2,TRPV3, TRPV4, TRPM8 and TRPA1 are thermoreceptors[80]. These receptors are designed to detect a wide range oftemperatures from hot (TRPV1 and TRPV2) and cold(TRPA1) noxious temperatures to innocuous thermal stimuli(TRPV3, TRPV4 and TRPM8) [24,80]. In addition, thesechannels are activated by chemical agents such as capsaicin(TRPV1), menthol (TRPM8), camphor (TRPV3) and mustard(TRPA1) [19,60,74,81,80,100]. Furthermore, TRPV1 is alsogated by protons and endocannabinoids [19,61]. Notably, thephysiological properties of the receptor have signalled it outas a key thermosensory transducer in the nervous system[19,80]. In support of this tenet, analysis of vanilloidreceptor null mice has substantiated the involvement of thischannel in pain sensation [18,30].
At the molecular level, TRPV1 is a calcium permeablenon-selective cation channel. The functional receptor is atetrameric membrane protein with four identical subunitsassembled around a central aqueous pore, although hetero-meric association with TRPV3 has been reported [98].Structurally, each TRPV1 subunit protein shows a membranedomain composed of six transmembrane segments (S1-S6),with an amphipathic region between the fifth and sixthsegment that forms the channel conductive pore. This regioncontains glutamic acids that are involved in the pH-dependent gating of the receptor [19]. The protein also has acytoplasmic N- and C-termini (Fig. 1). In the N-terminus,TRPV1 channels exhibit three ankyrin domains that mediateprotein-protein interactions with cytosolic proteins, andshow consensus sequences for protein kinases. The proteindisplays a cytosolic C-terminus domain containing phospho-inositide, and calmodulin binding (CAM) domains, as well

2 Current Neuropharmacology, 2006, Vol. 4, No. 1 Messeguer et al.
as phosphorylation sites [24]. In addition, the C-end has aTRP-like motif that functions as an association domain ofreceptor subunits [39].
TRPV1 is an integrator of noxious stimuli that gates inresponse to heat ( 42ºC), vanilloids, protons, and pro-algesic substances [19,61]. Temperature sensing seemsrelated to voltage dependent gating, as evidenced by the shiftin TRPV1 voltage-dependent activation in response tochanges in temperature [111]. Chemical activators of TRPV1channels can be grouped into those agents that directly gatethe channel, and those that allosterically modulate theprotein by post-translational modifications. Furthermore,agonists of the receptor act as gating modifiers reducing thetemperature threshold of the channel gating [111]. AlthoughTRPV1 is a temperature-gated channel with a temperaturethreshold of activation of 42ºC, it was recently shown thatTRPV1 may not be required for the detection of noxious heatin intact C-fiber afferents [117]. However, the channelbecomes a pivotal heat sensor under pathological/inflammatory states as concluded from genetic and pharma-cological TRPV1 knock-out studies [18,30,38]. The receptorhas two abundant single nucleotide polymorphisms thatproduce amino acid substitutions, one in codon 315 (Met
315
Ile) at the N-terminus domain, and the other at amino acid585 (Ile
585 Val) located in the fifth transmembrane segment.
Interestingly, gender, ethnicity and temperament seem tocontribute to individual variation in thermal and cold painsensitivity by interactions in part with these TRPV1 singlenucleotide polymorphisms [66].
A large body of evidence indicates that post-translationalmodification of TRPV1 increases the channel activity of thereceptor [11,32]. Indeed, TRPV1 has several consensussequences for protein kinases A (PKA), C (PKC), calmodulinkinase II (CAMKII) and the tyrosine specific kinase Src(Fig. 1). PKA phosphorylation potentiates TRPV1 responsesby reducing agonist-induced desensitisation. Desensitisationof the receptor depends on the presence of extracellular Ca
2+,
and it is characterised by a profound run-down of its channelactivity upon repetitive agonist stimulation. PKA exerts its
action through partial rescue of desensitised receptors bydirect phosphorylation of amino acids S116 and T370 at theN-terminus domain. Similarly, CaMKII-mediated phospho-rylation, along with calcineurin-induced dephosphorylationof the receptor, notably contributes to determine itsactivation and desensitisation state [63,78]. Likewise, incolonic dorsal root ganglion neurons, the channel activity ofTRPV1 is enhanced by tyrosine phosphorylation with thenonreceptor tyrosine kinase Src [58]. Tyrosine phospho-rylation of the protein may underlie in part to the IL-1 -induced potentiation of heat-activated currents in rat sensoryneurons [86].
PKC-dependent phosphorylation of TRPV1 can beprimarily induced either by extracellular ATP released frominjured cells, IL-1 , proteases, neurotrophins and bradykinin[86,102,110]. In vitro studies have identified S502, T704 andS800 as PKC phosphorylation sites (Fig. 1). Different PKCisoforms have been involved in TRPV1 sensitisation. Forinstance, phorbol esters primarily activate PKC [87],although they can also act as a direct ligands of TRPV1[10,83]. In contrast, stimulation of bradykinin B1 and B2receptors leads to PKC activation [20,102]. More recently,PKC , a kinase related to, but distinct of the PKC family hasbeen shown to phosphorylate and sensitise both hetero-logously expressed and native TRPV1 channels [115]. PKCphosphorylation of the vanilloid receptor notably enhancesits responsiveness by augmenting the channel openprobability [93]. Akin to vanilloids and pH, TRPV1 phospho-rylation by PKC decreases the receptor heat threshold, thusleading to activation of the channel at body temperature. Inaddition, PKC activation promotes the rapid recruitment ofvesicular receptors to the cell surface [83]. The occurrence ofboth events provides a molecular mechanism for the painexperienced in response to a warm stimulus, such asshowering sunburnt skin.
EXPRESSION OF TRPV1 CHANNELS
In situ hybridisation, immunocytochemical analysis, anddrug binding assays have shown TRPV1 expression in 50%of dorsal root and trigeminal ganglion neurons, in dorsal
Fig. (1). Proposed model of the topology of TRPV1 subunits. Each subunit consists of six putative transmembrane segments and intracellular
N- and C- domains. Functional regulatory domains of the receptors and phosphorylation sites are indicated. The capsaicin binding site is
labelled withe the chili icon.

Physiology and Pharmacology of the Vanilloid Receptor Current Neuropharmacology, 2006, Vol. 4, No. 1 3
horn of spinal cord and caudal nucleus of spinal trigeminalcomplex [19,107]. The majority of TRPV1 positive neuronsalso colocalise with the nerve growth factor (NGF) receptortrkA , the lectin IB4, and the neuropeptides involved innociceptive transmission such as substance P (SP) andcalcitonin gene related peptide (CGRP) [19,61]. Vanilloidsensitive nociceptors are peptidergic, small diameter neuronsthat give rise to unmyelinated C fibers, although some Afibers are responsive to vanilloid derivatives [49]. Somaticand visceral primary afferents express TRPV1 at both thespinal and peripheral terminals. Furthermore, TRPV1expression has been reported in vagal afferents in jugular andnodose ganglion neurons [54,84].
In addition to a subset of nociceptors, TRPV1 is presentin neurons of the central nervous system and non-neuronalcells. For instance, TRPV1 mRNA or protein is widelyexpressed in brain regions such as the olfactory nuclei,cerebral cortex, dentate gyrus, central amygdala, striatum,centromedian and paraventricular thalamic nuclei, hypotha-lamus, substantia nigra, reticular formation, locus coeruleus,inferior olive and cerebellar cortex [see 84 and referencestherein]. The role of TRPV1 in the central nervous system isstill elusive, although it may mediate endovanilloid signallingpromoting the release of excitatory neurotransmitter such asL-glutamate, noradrenaline and dopamine [71,72].
In the skin, TRPV1 positive cells have been found in thedermis and epidermis [46], primarily in Meissner corpusclesand keratinocytes [56,88]. Moreover, this receptor is alsoexpressed in mast cells, where its activation could releasemast cell pro-algesic mediators that bind to histamine andproteinase-activated receptors on sensory terminals [99].More recently, functional TRPV1 channels were found in thehuman hair follicle [15]. Activation of this receptor incultured keratinocytes inhibited cellular proliferation, inducedapoptosis, up regulated known endogenous hair growthinhibitors, and down regulated hair growth promoters. Thus,these results suggest an important role of TRPV1 inepithelial growth disorders [15].
It is also worth mentioning that TRPV1 receptors appearto be highly expressed in visceral afferents [53], and arepresent in the gastrointestinal (GI) tract within the myentericand submucous plexus and some enteric intrinsic neurons[116]. Although capsaicin does not desensitise gastricepithelial cells, activation of TRPV1 expressing primaryafferents has been shown to thicken the protective barrier inthe stomach and duodenum [84], and sensitisation of thisreceptor plays a role in GI inflammation and function [41].In pathological conditions, TRPV1 has been implicated inthe hypermotility of the GI tract in abdominal painassociated to functional bowel disorders, and in theneurogenic component of pancreatitis [1,49].
TRPV1 expression is also prominent in other tissues. Inthe urinary bladder, functional channels are expressed in theuroepithelium in both the superficial and basal layers [4],where they are implicated in the development of themicturition reflex in both normal and pathological conditions[12]. In the lungs, capsaicin stimulates airway specific C-fibers and may play a role in the generation of non-productive cough [65]. Neurogenic inflammation of mucosal
nociceptors also seems to be a key factor in the pathogenesisof asthma [44]. Under normal conditions, these fibers do notexpress SP but begin to produce it after allergicinflammation and viral infection [17,51]. In the vascularsystem, TRPV1-expressing fibers accompany blood vesselsin all layers of the viscera. These receptors appear involvedin Bayliss myogenic constriction which leads tohypertension [97]. Furthermore, TRPV1 seems to contributeto myocardial protection by releasing CGRP and nitric oxidefrom capsaicin-sensitive afferents in the ischemic heart[101]. Another tissue that expresses TRPV1 channels is theinner ear, where the protein is present in inner and outer haircells, inner and outer pillar cells, Hensen cells and satellitecells [8,121]. These channels appear to be involved inhearing, as evidenced by the vanilloid-induced alteration ofcochlear sensitivity [104].
Taken together, all these findings illustrate that TRPV1channels are widely expressed in neuronal and non-neuronalcells of both endodermal and mesodermal origin, andsuggest that the receptor is involved in diverse physiologicalfunctions. These include thermosensory transduction, as wellas chemical signalling presumably mediated by endovanilloidcompounds. In addition, they hint that dysfunction of thechannel may underlie the etiology of pathological sensorytransduction such as that occurring in inflammation. Takentogether, all these observations underscore the notion thatTRPV1 is a widely expressed protein whose function may becritical for diverse physiological conditions.
TRPV1 A THERAPEUTIC TARGET FOR PAIN
MANAGEMENT
The pivotal role of TRPV1 in physiology has suggested acontribution of the channel to the mechanism of diversehuman diseases. In particular, cumulative evidence issubstantiating the tenet that nociceptor sensitisation byinflammatory agents is primarily achieved by TRPV1. Thisreceptor is the endpoint target of intracellular signallingpathways triggered by inflammatory mediators that lead topotentiation of its channel activity which, in turn, promotesthe hyperexcitability of nociceptors. Enhancement of TRPV1function by pro-algesic agents may be accomplished eitherby direct activation of the channel or by its post-translationalmodification mediated by intracellular metabolic cascades[24,80]. Direct activation of TRPV1 responses has beenreported for lipid mediators such as arachidonic acidmetabolites including anandamide, N-arachidonyl-dopamine(NADA), N-oleyldopamine and, 12-(hyperoxy)eicosatet-renoic acid (12-HPETE). These compounds act as weakagonists, but notably increase TRPV1-mediated [Ca
2+]i in
both heterologous expression systems, and in primarysensory neurons [22,50,52]. In addition, the acidosis thatdevelops in inflamed tissues is also a direct activator of theTRPV1 channel activity [19]. The potency and efficacy ofeach singular mediator is quite low but in inflammatoryconditions, several of these modulators are simultaneouslyreleased and act synergistically. Noteworthy, most of theseTRPV1 ligands act by reducing the heat threshold of channelactivation from 42ºC to body temperature ( 35ºC) [11].Therefore, direct gating of TRPV1 responses by inflammatoryagents acting as channel agonists notably increases the

4 Current Neuropharmacology, 2006, Vol. 4, No. 1 Messeguer et al.
excitability of nociceptors resulting in a hyperalgesiccondition. In addition, the TRPV1-mediated [Ca
2+]i rise, in
turn, triggers the release of pro-inflammatory agents atperipheral terminals, thus further increasing the excitabilityof the nociceptors. This feedback circuit notably contributesto enhance the hypersensitivity of inflamed tissue.
Activation of intracellular protein networks duringinflammation results in TRPV1 phosphorylation [11], releaseof tonically-inhibited receptors [23], and an increment of thesurface expression of functional channels [83], all beingmajor events underlying the nociceptor activation andsensitisation that leads to hyperalgesia. Indeed, TRPV1expression is upregulated in tissue samples from patientswith inflammatory bowel disease and Crohn´s disease [118],and also in patients with rectal hypersensitivity [21], as wellas those affected of vulvodynia [108]. Thus, TRPV1receptors are strong candidates for an important role in thesensitisation of primary afferents after injury or inflammation.The involvement of TRPV1 in heat hyperalgesia is under-scored by the reduced thermal hypersensitivity of micelacking TRPV1 [18,30]. Similarly, non-competitive anta-gonists of the TRPV1 channel notably attenuate the heathyperalgesia triggered by inflammation in vivo [38]. Theimportant contribution of TRPV1 receptor to the onset andmaintenance of neurogenic inflammation has validated it as atherapeutic target for inflammatory pain management.
In addition to the contribution of the vanilloid receptor asa target of the neurogenic inflammation underlying differentdiseases, TRPV1 is gaining interest for the treatment ofneuropathic, postoperative and chronic pain and, recently,for the therapy of epithelial disorders. Thus, for instance,topical capsaicin or resiniferotoxin have been used inpostherpetic neuralgia, diabetic neuropathy, postmastectomypain and arthritis [64,103]. Recently, TRPV1 has beenclearly validated as a key target for management of chronicpain in bone cancer [42]. As a result, the development ofspecific TRPV1 antagonists is a central focus of current drugdiscovery programs.
PHARMACOLOGY OF TRPV1 RECEPTORS
The TRPV1 receptor is a molecular entity with diversedrug binding sites. Structure-function studies have unmaskedthe molecular determinants of the capsaicin binding site inan intracellular domain of the receptor (Fig. 1) [40,59], andthose involved in the interaction with the non-competitiveantagonist ruthenium red at the extracellular vestibule of thepore domain [37]. The existence of these binding sites hasprompted the discovery of novel vanilloid-like agonists, aswell as competitive and non-competitive antagonists with thehope that they will be of clinical use to treat human TRPV1receptor dysfunction. Further summarized are the effortsundertaken to develop these three classes of TRPV1modulators.
TRPV1 AGONISTS
Agonists are molecules that directly gate the channel by amechanism that involves the reduction of the heat thresholdof activation. Prolonged exposure of the receptor to theagonist in the presence of Ca
2+ induces channel closure by
desensitisation and tachyphylaxia [103], by a mechanism
that involves phosphorylation of a key residue at the C-terminus of the protein [11]. Agonist-induced desensitisationis thought to underlie the analgesic activity of capsaicin.However, capsaicin analgesia may also arise from selectivedeletion of TRPV1-expressing nociceptors due to the Ca
2+
overload that may induce the ligand [64]. Thus, high-affinityagonists that promote receptor tachyphylaxia and/or noci-ceptor ablation could be used as efficacious pain relievers.
The identification of several natural products that act asTRPV1 ligands prompted the use of these molecules aspharmacological tools to study this receptor even before itsmolecular identity was known. In addition, these studiesopened different medicinal chemistry programs based onstructure-activity relationships (SAR) directed to improvethe therapeutic profile of the initial hits.
Vanilloids are the best known kind of TRPV1 agonists.Capsaicin (1, Fig. 2) is the representative example of thisfamily. This molecule bears the 4-hydroxy-3-methoxybenzylmoiety characteristic for homovanillin derivatives, an amidelinker and a lipophilic region. The homovanillyl motif andthe amide linker contain polar groups capable of forminghydrogen bonds. The SAR studies performed on capsaicinanaloges suggest that these polar moieties are essential forexciting sensory neurons. In contrast, the hydrophobicregion, with an optimal chain length of 8-15 carbon atoms,would interact with a hydrophobic region in the receptor.Unlike other ligand-gated channels that produce fast synaptictransmission, vanilloids show a slow activation kinetics, inpart because their binding site is located in the intracellularportion of the receptor (Fig. 1) [31,40,59,62]. Indeed, acytosolic region spanning the third transmembrane domainin TRPV1 appears to be essential for capsaicin binding,presumably through hydrophobic interactions with thevanilloid carbon chain [40,59]. The structural determinantsinvolved in capsaicin binding are being unravelled andmolecular models for the vanilloid site have been proposedwhich, upon refinement, may facilitate the design of agonistswith higher therapeutic index [40].
There are other known classes of naturally occurringvanilloids, such as Resiniferatoxin (RTX, 2), a compoundisolated from Euphorbia resinifera, that exhibits a TRPV1agonistic activity much more potently than capsaicin (IC50=10 pM). As shown in Fig. 2 , RTX is a rather complexmolecule and the pharmacophoric groups have not beenclearly defined yet. SAR studies suggest that the homo-vanillic moiety of RTX, the C3-keto group and the orthoesterphenyl moiety in ring C are essential structural elements foreliciting its extremely high potency desensitising TRPV1channel activity. As a consequence, RTX is being developedas a sensory neuron desensitising agent for the treatment ofurinary urge incontinence and pain associated with diabeticneuropathy [105], two human pathologies mediated byTRPV1 dysfunction. In spite of these results, the limitedavailability of RTX from natural sources and its difficultchemical synthesis hamper its therapeutic use.
The efficacy of both capsaicin and RTX for treatingoveractive bladder and neuropathic pain has stimulated theefforts for identifying orally active TRPV1 agonists. A majorshortcoming for the therapeutic use of capsaicin and related
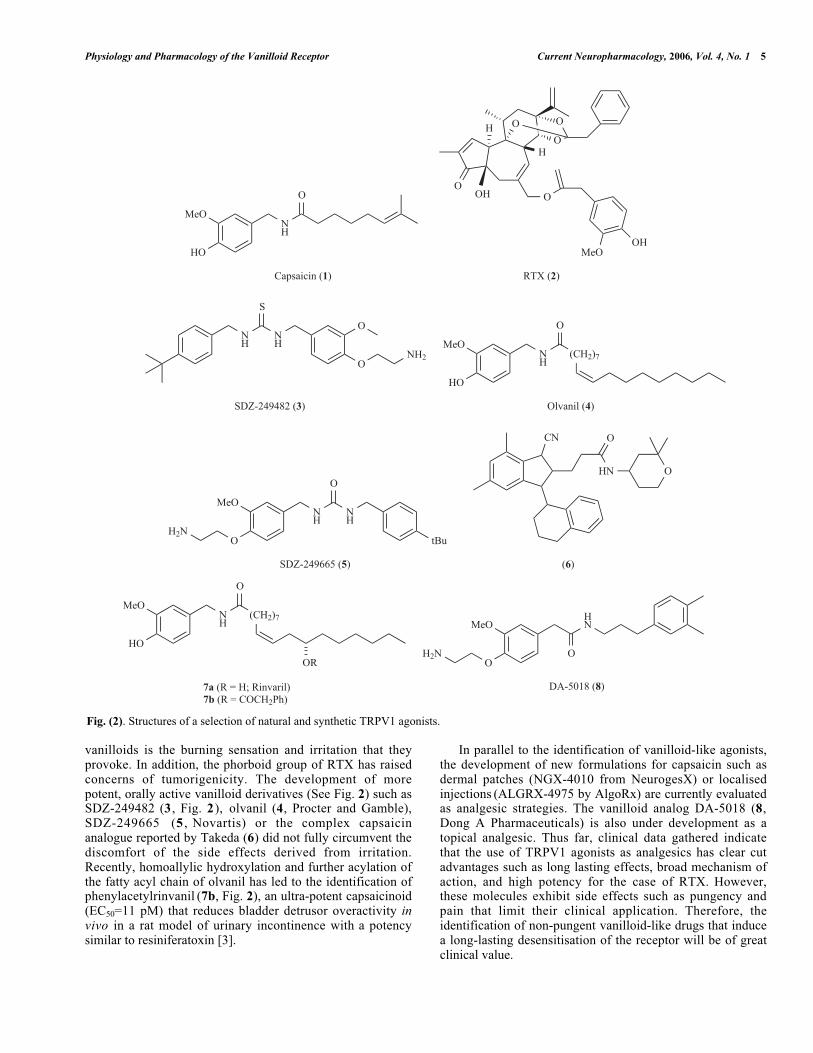
Physiology and Pharmacology of the Vanilloid Receptor Current Neuropharmacology, 2006, Vol. 4, No. 1 5
vanilloids is the burning sensation and irritation that theyprovoke. In addition, the phorboid group of RTX has raisedconcerns of tumorigenicity. The development of morepotent, orally active vanilloid derivatives (See Fig. 2) such asSDZ-249482 (3 , Fig. 2), olvanil (4, Procter and Gamble),SDZ-249665 (5 , Novartis) or the complex capsaicinanalogue reported by Takeda (6) did not fully circumvent thediscomfort of the side effects derived from irritation.Recently, homoallylic hydroxylation and further acylation ofthe fatty acyl chain of olvanil has led to the identification ofphenylacetylrinvanil (7b, Fig. 2), an ultra-potent capsaicinoid(EC50=11 pM) that reduces bladder detrusor overactivity invivo in a rat model of urinary incontinence with a potencysimilar to resiniferatoxin [3].
In parallel to the identification of vanilloid-like agonists,the development of new formulations for capsaicin such asdermal patches (NGX-4010 from NeurogesX) or localisedinjections (ALGRX-4975 by AlgoRx) are currently evaluatedas analgesic strategies. The vanilloid analog DA-5018 (8,Dong A Pharmaceuticals) is also under development as atopical analgesic. Thus far, clinical data gathered indicatethat the use of TRPV1 agonists as analgesics has clear cutadvantages such as long lasting effects, broad mechanism ofaction, and high potency for the case of RTX. However,these molecules exhibit side effects such as pungency andpain that limit their clinical application. Therefore, theidentification of non-pungent vanilloid-like drugs that inducea long-lasting desensitisation of the receptor will be of greatclinical value.
Fig. (2). Structures of a selection of natural and synthetic TRPV1 agonists.
NH
O
MeO
HO
NH
(CH2)7
O
MeO
HO
OOH
H O
O
O
O
H
OHMeO
NH
NH
O
MeO
OH2N
tBu
CN
HN O
O
HNMeO
OH2N O
DA-5018 (8)
NH
NH
S
O
ONH2
NH
(CH2)7
O
MeO
HO
OR
Capsaicin (1)
Olvanil (4)
SDZ-249665 (5)
RTX (2)
SDZ-249482 (3)
(6)
7a (R = H; Rinvaril)
7b (R = COCH2Ph)

6 Current Neuropharmacology, 2006, Vol. 4, No. 1 Messeguer et al.
There is good evidence on the occurrence of endogenousvanilloid receptor agonists, coined with the term endo-vanilloids, and that these agents modulate the sensitivity ofTRPV1 channels to thermal stimuli [109]. For instance, ithas been described that a polyunsaturated compound derivedfrom arachidonic acid, anandamide (9, Fig. 3) activates bothnative and recombinant TRPV1 receptors [122]. Thestructure of the molecule does not give evidence that it is avanilloid analog. This fact caused some confusion about theterm “vanilloid” depending on whether it is used chemicallyor pharmacologically. Moreover, anandamide is alsoconsidered as an endocannabinoid, since it activatescannabinoid (CB) receptors CB1 and CB2 at concentrationslower than those required to gate TRPV1 [122]. Similarly,several eicosanoids, particularly those derived from theenzymatic action of 5-lipoxygenase or 12-lipoxygenase, arecapable of activating rat TRPV1. In particular, 12-HPETE(10 , Fig. 3) and leukotriene B4 (11 , LTB4, Fig. 3) haveexhibited the most potent agonistic activity [50]. It isbelieved that eicosanoids function as intracellular vanilloidsof TRPV1 receptors in the cells where they are synthesised.Recently, NADA (12, Fig. 3) has been identified as a brainendovanilloid [52]. NADA is an endogenous anandamideanalog several times more potent than anandamide onTRPV1, although it still activates CB1 receptors.
Another related approach to modulate TRPV1 signallingis the identification of the so-called hybrid cannabinoids/TRPV1 ligands. Since TRPV1 colocalises with CB1receptors in brain, and both proteins have overlapping ligandrecognition properties, it is possible to conceive compoundscapable of activating both receptor types simultaneouslyeven by different mechanisms. Arvanil (N-[3’-methoxy-4’-
hydroxybenzyl]arachidonamide, 13, Fig. 3) is one of theseagents. Arvanil has affinity for CB1 receptors and activatesTRPV1 receptors more potently than capsaicin or anandamide[75]. This compound is an antiproliferative agent for humanbreast cancer cells sensitive to both CB1 and TRPV1receptor antagonists, as well as a spinal analgesic and, arelaxant of mouse vas deferens. Similarly, arachidonylethanolamine, an endogenous cannabinoid that also acts onthe TRPV1 channel, induced strong apoptosis of uterinecancer cells that aberrantly express the vanilloid receptor[24]. Accordingly, it is of great interest for the futuredevelopment of improved hybrid CB1/TRPV1 agonists.
TRPV1 ANTAGONISTS
Intense efforts have been carried out to design TRPV1competitive and non-competitive antagonists [91,106]. Thecompetitive antagonists bind to the agonist binding site, andlock the channel in the closed, nonconductive state. Incontrast, non-competitive antagonists interact with additionalbinding sites on the receptor structure preventing receptoropening by the agonist or blocking its aqueous pore [91].Non-competitive antagonists acting as open channel blockersare therapeutically attractive because they preferentiallyrecognise the population of over-activated TRPV1 channelswith a marginal recognition of physiologically workingreceptors. Thus, this kind of drugs target pathologically-activated receptors, which can reduce the potential unwantedside effects.
TRPV1 COMPETITIVE ANTAGONISTS
The first competitive TRPV1 antagonist, capsazepine(14, Fig. 4), was identified by Sandoz (now Novartis) and
Fig. (3). Structures of TRPV1 endogenous ligands and hybrid canabinoid-TRPV1 modulators.
NH
OH
O
OH
O
HO
O
NH
OOH
OH
OH
OH
OOH
HO
OCH3
NH
O
Anandamide (9) 12-HPETE (10)
NADA (12)LTB4 (11)
Arvanil (13)
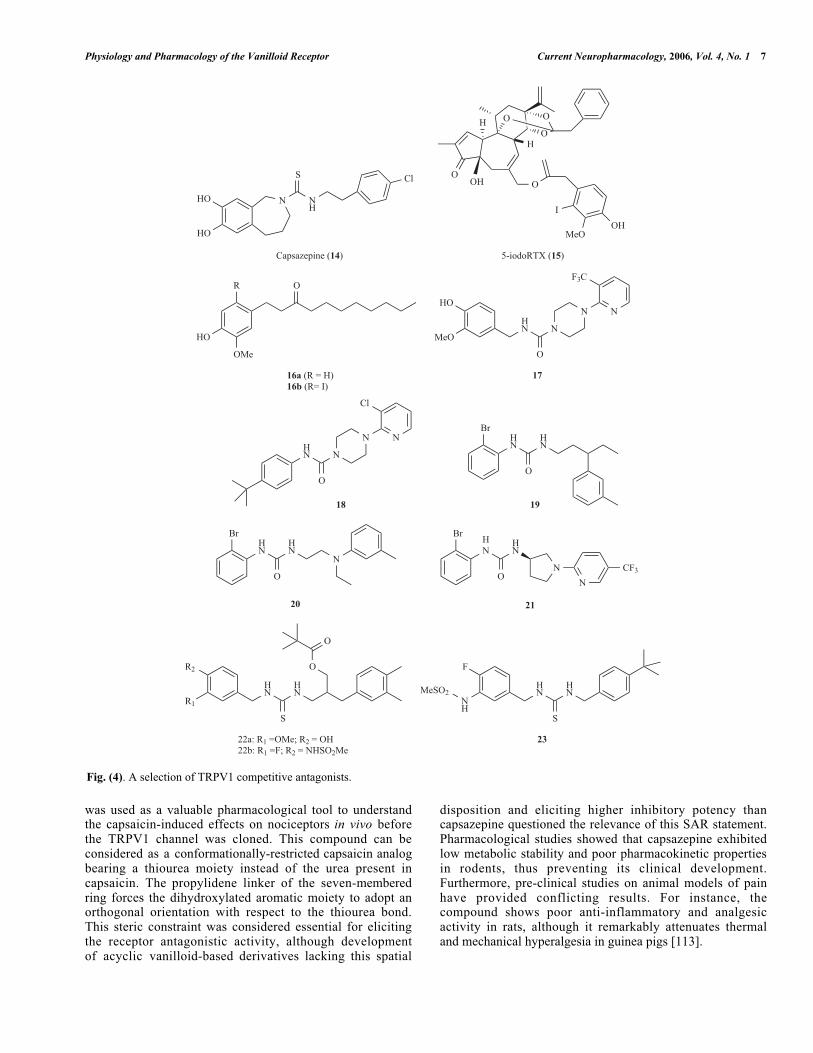
Physiology and Pharmacology of the Vanilloid Receptor Current Neuropharmacology, 2006, Vol. 4, No. 1 7
was used as a valuable pharmacological tool to understandthe capsaicin-induced effects on nociceptors in vivo beforethe TRPV1 channel was cloned. This compound can beconsidered as a conformationally-restricted capsaicin analogbearing a thiourea moiety instead of the urea present incapsaicin. The propylidene linker of the seven-memberedring forces the dihydroxylated aromatic moiety to adopt anorthogonal orientation with respect to the thiourea bond.This steric constraint was considered essential for elicitingthe receptor antagonistic activity, although developmentof acyclic vanilloid-based derivatives lacking this spatial
disposition and eliciting higher inhibitory potency thancapsazepine questioned the relevance of this SAR statement.Pharmacological studies showed that capsazepine exhibitedlow metabolic stability and poor pharmacokinetic propertiesin rodents, thus preventing its clinical development.Furthermore, pre-clinical studies on animal models of painhave provided conflicting results. For instance, thecompound shows poor anti-inflammatory and analgesicactivity in rats, although it remarkably attenuates thermaland mechanical hyperalgesia in guinea pigs [113].
Fig. (4). A selection of TRPV1 competitive antagonists.
HO
HO
N NH
ClS OOH
HO
O
O
O
H
OHMeO
I
R
HO
O
OMe
HN N
N NHO
MeO
O
F3C
HN
HN
R2
R1
S
O
O
HN N
N N
Cl
O
HN
HN
Br
O
HN
HN
Br
O
N
HN
HN
F
NH
S
MeSO2
BrH
NHN
ON
N
CF3
Capsazepine (14) 5-iodoRTX (15)
16a (R = H)
16b (R= I)
17
22a: R1 =OMe; R2 = OH
22b: R1 =F; R2 = NHSO2Me
18 19
20
23
21

8 Current Neuropharmacology, 2006, Vol. 4, No. 1 Messeguer et al.
The discovery that the introduction of an iodine atom onthe vanillyl moiety of RTX agonist modulates itspharmacological activity depending on the position of thehalogen ended up with the identification of 5-iodoRTX (15,Fig. 4) as a potent antagonist of TRPV1 (IC50 = 3.9 nM)[73,112]. This compound produced analgesic activity in vivoand it is currently in clinical studies. Similar to RTX, a majordrawback of compound 15 is its complex chemical structureand, the cost of its synthesis from RTX itself. In addition, 5-iodoRTX exhibits low oral bioavailability probably due to itspoor pharmacokinetic properties. Nevertheless, the discoveryof compound 15 stimulated the SAR studies on the halo-genation of the vanillyl moiety present in simple capsaicinderivatives. For instance, taking nonivamide as a model(16a, Fig. 4), the substitution at C-6 afforded more potentinhibitors than at C-5 and the inhibitory potency correlatedwith the size of the halogen (I > Br > Cl) [2]. The mostpotent derivative identified was that bearing the iodine atomat C-6 (16b). Similarly, iodination of phenylacetylrinvanil(7b) produced a potent TRPV1 antagonist (IC50 = 0.8 nM)[3]. These results showed that halogenation could be anappropriate strategy for reversing the activity of vanilloids.This observation, together with the aim of synthesisingsimpler derivatives of RTX that preserve the inhibitorypotency of the parent compound, would constitute anattractive field for developing orally active, competitiveTRPV1 antagonists with improved pharmaceutical profilesfor clinical use.
A related family of capsaicin analogues was developedby Neurogen through the formal amidation of the homo-vanillylamino moiety with different 4-( -pyridyl)piperidine-1-acyl residues. The urea 17 (Fig. 4) is a representativeexample of this family [6]. The presence of an amino moietyat the hydrophobic region of the capsacinoid appeared to becrucial for obtaining a potent inhibitor that was, at the sametime, a highly improved drug lead in terms of hydrophilicity.This observation was also important to recognise that the C-region of capsaicin should be considered as a domain thatplays a key role in the interaction with the receptor. Anotherpiperazinyl urea (BCTC, 18) was reported by Purdue-
Pharma to be more selective than capsazepine against a widevariety of enzymes and channels. This compound exhibitedin vivo oral activity in animal models of inflammatory andneuropathic pain [92]. Less conformationally-restrictedanalogs of BCTC such as compounds 19 and 20, along withthe recently reported pyrrolidine urea 21 (Fig. 4), also elicit agood antagonistic TRPV1 potency maintaining the receptorselectivity. The ongoing pre-clinical characterization of thesecompounds will uncover whether they have a highertherapeutic index than BCTC [94]. It is worth noting thatmost of these molecules were identified from the screeningof chemical combinatorial libraries, thus showing theusefulness of this strategy in drug discovery programs.
Structurally connected to capsaicin related compounds,thiourea 22a (Fig. 4) elicited a potent agonist activity (Ki <10 nM). Interestingly, the presence of fluorine at C-3’ andthe methanesulfonamido moiety at C-4’, replacing themethoxy and hydroxy groups, respectively, affordedcompound 22b, a molecule exhibiting an antagonistic TRPV1activity [68]. Further modification of these derivatives, nowat the lipophilic region, led AmorePacific to obtainoptimized compound 23, bearing a tert-butyl group at thearomatic ring [114].
The continuous efforts to improve the pharmacologicalproperties of TRPV1 antagonists have resulted in thedevelopment of novel templates (See Fig. 5). In this regard,several non-symmetric ureas have been reported by Janssen[25], Abbott [69] and Merck [16], where the 5-aminoisoquinoline moiety appears as a common feature (cf.24-26). Alternatively, Bayer has reported the 4-aminoindole27 [119] and the hydroxylated tetrahydronaphthalene 28
[119] urea derivatives. All these compounds show aninhibitory potency 10 nM. Taken together, these resultsemphasise the relevance for antagonistic activity of afunctionalised lipophilic moiety, preferentially with a basicgroup, for interacting with the capsaicin binding site onTRPV1.
The replacement of the urea moiety by other isostericgroups has also been explored. Euroceltique reported
Fig. (5). TRPV1 antagonists based on novel templates.
N
HN NH
R
O
R1
R1 = H, F
R
24-26NH
HN NH
PhO
HN
HO
NH
Cl
CF3
O
27 28
NH
N N
NtBu
Cl
N
CN
29
NH
O
OMe
Cl
N NH
O
Ph
30 31
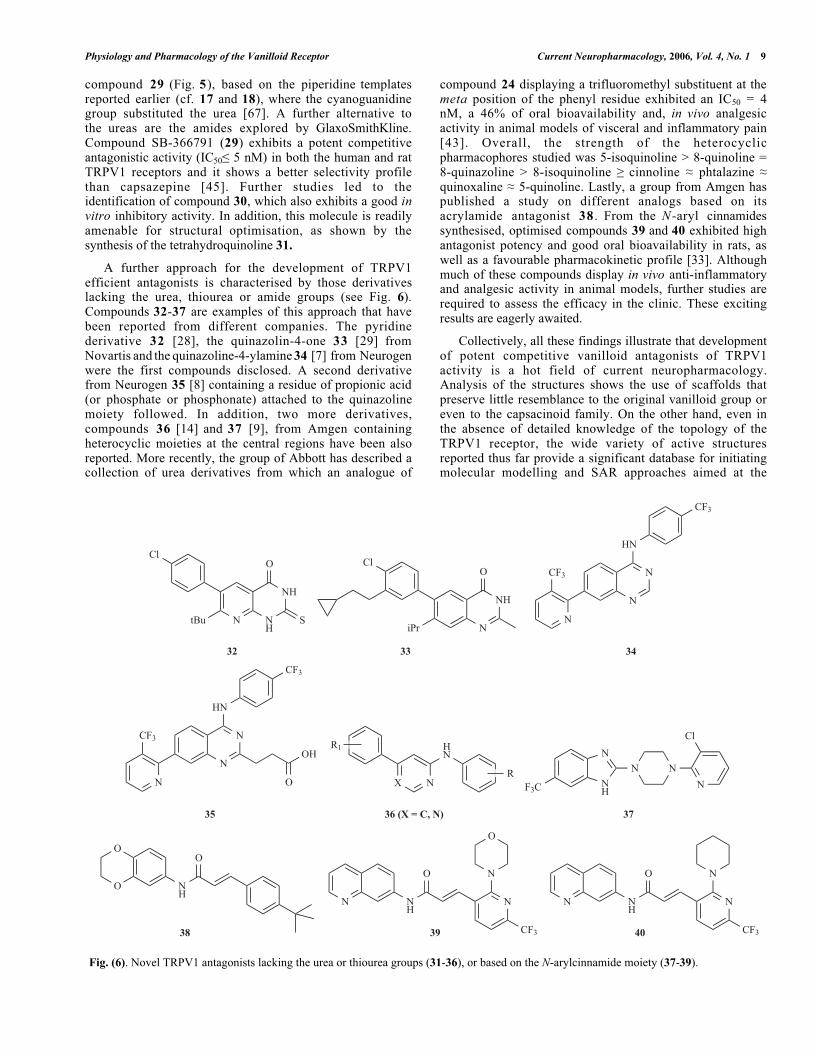
Physiology and Pharmacology of the Vanilloid Receptor Current Neuropharmacology, 2006, Vol. 4, No. 1 9
compound 29 (Fig. 5), based on the piperidine templatesreported earlier (cf. 17 and 18), where the cyanoguanidinegroup substituted the urea [67]. A further alternative tothe ureas are the amides explored by GlaxoSmithKline.Compound SB-366791 (29) exhibits a potent competitiveantagonistic activity (IC50 5 nM) in both the human and ratTRPV1 receptors and it shows a better selectivity profilethan capsazepine [45]. Further studies led to theidentification of compound 30, which also exhibits a good invitro inhibitory activity. In addition, this molecule is readilyamenable for structural optimisation, as shown by thesynthesis of the tetrahydroquinoline 31.
A further approach for the development of TRPV1efficient antagonists is characterised by those derivativeslacking the urea, thiourea or amide groups (see Fig. 6).Compounds 32-37 are examples of this approach that havebeen reported from different companies. The pyridinederivative 32 [28], the quinazolin-4-one 33 [29] fromNovartis and the quinazoline-4-ylamine 34 [7] from Neurogenwere the first compounds disclosed. A second derivativefrom Neurogen 35 [8] containing a residue of propionic acid(or phosphate or phosphonate) attached to the quinazolinemoiety followed. In addition, two more derivatives,compounds 36 [14] and 37 [9], from Amgen containingheterocyclic moieties at the central regions have been alsoreported. More recently, the group of Abbott has described acollection of urea derivatives from which an analogue of
compound 24 displaying a trifluoromethyl substituent at themeta position of the phenyl residue exhibited an IC50 = 4nM, a 46% of oral bioavailability and, in vivo analgesicactivity in animal models of visceral and inflammatory pain[43]. Overall, the strength of the heterocyclicpharmacophores studied was 5-isoquinoline > 8-quinoline =8-quinazoline > 8-isoquinoline cinnoline phtalazine quinoxaline 5-quinoline. Lastly, a group from Amgen haspublished a study on different analogs based on itsacrylamide antagonist 38 . From the N-aryl cinnamidessynthesised, optimised compounds 39 and 40 exhibited highantagonist potency and good oral bioavailability in rats, aswell as a favourable pharmacokinetic profile [33]. Althoughmuch of these compounds display in vivo anti-inflammatoryand analgesic activity in animal models, further studies arerequired to assess the efficacy in the clinic. These excitingresults are eagerly awaited.
Collectively, all these findings illustrate that developmentof potent competitive vanilloid antagonists of TRPV1activity is a hot field of current neuropharmacology.Analysis of the structures shows the use of scaffolds thatpreserve little resemblance to the original vanilloid group oreven to the capsacinoid family. On the other hand, even inthe absence of detailed knowledge of the topology of theTRPV1 receptor, the wide variety of active structuresreported thus far provide a significant database for initiatingmolecular modelling and SAR approaches aimed at the
Fig. (6). Novel TRPV1 antagonists lacking the urea or thiourea groups (31-36), or based on the N-arylcinnamide moiety (37-39).
N NH
NH
tBu
O
S
Cl
N
NH
iPr
OCl
N
N
HN
CF3
N
CF3
N
N
HN
CF3
N
CF3
OH
O X N
HN
R1
RNH
N
N N
N
Cl
F3C
32 33 34
35 36 (X = C, N) 37
O
O
NH
O
38
N NH
N
NO
CF3
O
N NH
N
NO
CF339 40
10 Current Neuropharmacology, 2006, Vol. 4, No. 1 Messeguer et al.
identification of the pharmacophoric features required fororally active, potent antagonistic activity. These studies arealso necessary to clarify whether the above structural varietyis due to the interaction with sames of different drug bindingsites on the receptor.
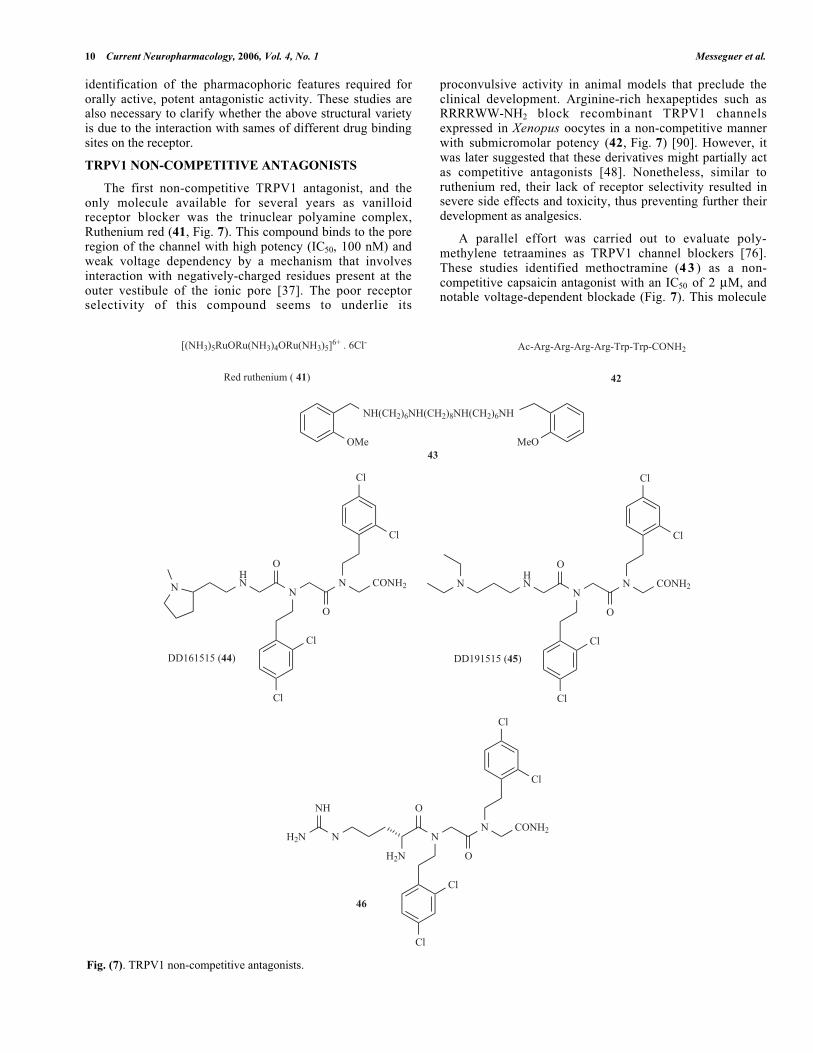
TRPV1 NON-COMPETITIVE ANTAGONISTS
The first non-competitive TRPV1 antagonist, and theonly molecule available for several years as vanilloidreceptor blocker was the trinuclear polyamine complex,Ruthenium red (41, Fig. 7). This compound binds to the poreregion of the channel with high potency (IC50, 100 nM) andweak voltage dependency by a mechanism that involvesinteraction with negatively-charged residues present at theouter vestibule of the ionic pore [37]. The poor receptorselectivity of this compound seems to underlie its
proconvulsive activity in animal models that preclude theclinical development. Arginine-rich hexapeptides such asRRRRWW-NH2 block recombinant TRPV1 channelsexpressed in Xenopus oocytes in a non-competitive mannerwith submicromolar potency (42, Fig. 7) [90]. However, itwas later suggested that these derivatives might partially actas competitive antagonists [48]. Nonetheless, similar toruthenium red, their lack of receptor selectivity resulted insevere side effects and toxicity, thus preventing further theirdevelopment as analgesics.
A parallel effort was carried out to evaluate poly-methylene tetraamines as TRPV1 channel blockers [76].These studies identified methoctramine (4 3 ) as a non-competitive capsaicin antagonist with an IC50 of 2 M, andnotable voltage-dependent blockade (Fig. 7). This molecule
Fig. (7). TRPV1 non-competitive antagonists.
HN
NN CONH2N
O
O
Cl
Cl
Cl
Cl
HN
NN CONH2
O
O
Cl
Cl
Cl
Cl
N
Ac-Arg-Arg-Arg-Arg-Trp-Trp-CONH2
Red ruthenium ( 41)
DD161515 (44) DD191515 (45)
42
NN CONH2
O
O
Cl
Cl
Cl
Cl
46
OMe
NH(CH2)6NH(CH2)8NH(CH2)6NH
MeO
43
H2N
NH2N
NH
[(NH3)5RuORu(NH3)4ORu(NH3)5]6+ . 6Cl-

Physiology and Pharmacology of the Vanilloid Receptor Current Neuropharmacology, 2006, Vol. 4, No. 1 11
antagonised native TRPV1 receptors in dorsal root ganglionneurons activated by either capsaicin or protons. Therefore,the inhibitory activity of methoctramine analogs hasprovided a new pharmacophoric scaffold for non-competitiveantagonists of TRPV1. Thus far, the lack of receptorselectivity has restrained the use of these compounds in vivo.
The first small organic molecules identified as non-competitive TRPV1 emerged from the screening of apeptidomimetic combinatorial library. N-Alkylglycines (alsoknown as peptoids), constitute a class of compounds ofsynthetic origin that exhibit interesting biological propertiesderived from their structural features, i.e, the absence of CO-NH bonds and then of the hydrogen bond interactions thatthey induce, an improved stability against proteases and ahigh degree of conformational freedom. In addition, thestructural simplicity of peptoids makes these peptidomimeticcompounds amenable to structural manipulation, thusfacilitating the optimisation of identified hits for improvingtheir drug-like properties [47]. This is important if it isconsidered that the high conformational mobility of thesemolecules can generate selectivity problems due to undesiredinteractions with non-target receptors.
The screening of the library of trimers of N-alkylglycinesled to the identification of two compounds referred to asDD161515 (44) and DD191515 (45) that preferentially blockTRPV1 channel activity with micromolar potency andmoderate voltage-dependency (Fig 7) [38]. These peptoidsappear to recognise a receptor site different from that ofcapsaicin. Intraperitoneal administration of both trialkyl-glycines into mice significantly attenuated pain andneurogenic inflammation induced by injection of capsaicininto the hindpaws and, reduced thermal hyperlalgesia due tomustard oil-evoked tissue irritation. In contrast, thesepeptoids did not affect capsaicin triggered-mechanical hyper-sensitivity [38]. These findings suggested that TRPV1 activetrialkylglycines may be developed into analgesics to treatinflammatory pain. However, the in vivo doses required foranalgesic and anti-inflammatory activity were too high ( 25mg/kg), thus preventing their development into useful drugs.Therefore, the design of more potent, non-competitiveantagonists of TRPV1 is a yet unmet goal of drug discoveryplatforms. Recent progress in this arena, along withmolecular modelling of the TRPV1 pore domain [35], isproviding this kind of compounds. For instance, bycombining the chemical features of arginine-rich peptidesand peptoid DD161515, a potent non-competitive antagonist(H-Arg-15-15C, 46) of TRPV1 was discovered (Fig. 7) [34].Compound H-Arg-15-15C blocked capsaicin-inducedTRPV1 activity with 10-fold higher potency than the originalpeptoid DD161515, without compromising the receptorselectivity. Noteworthy, at variance with the parental peptoids,compound H-Arg-15-15C potently abrogated pH-activatedTRPV1 responses. As for DD161515, the blockademechanism was non-competitive acting as a channel blocker.Thus, introduction of a strong positive charge in thepharmacophore provided by peptoid DD161515 results in asignificant increase in the binding efficacy of the molecule tothe receptor. Noteworthy, compound 46 notably attenuatedthe inflammatory discomfort exerted by intraplantar injectionof capsaicin. The i.p. dose required to produce analgesia was
10-fold lower than that of peptoid DD161515. In addition,the compound showed activity on the Phase II of theformalin model, although at doses relatively high, suggestinga modest activity of the antagonist in neuropathic painmodels [96]. Indeed, H-Arg-1515C did not mitigatemechanical allodynia in the partially ligated sciatic nervemodel of neuropathic pain. In contrast, 46 completelyreversed the complete Freund Adjuvant (CFA)-inducedthermal hyperalgesia at therapeutic doses (5 mg/kg, i.p.).Collectively, these findings show that compound H-Arg-15-15C is a potent non-competitive antagonist of the receptorthat exhibits notable anti-inflammatory and analgesic activityin pre-clinical models of acute and chronic pain. However,compound 46 displays limited oral and i.p. bioavailability invivo, presumably because of its lack of compliance with theLipinski rule of 5 [70]. Thus, chemical modification on theH-Arg-15-15C structure will be necessary to increase itstherapeutic potential. All these results demonstrate thatTRPV1 antagonists that act on a site different from thevanilloid binding site are also promising candidates foranalgesic/anti-inflammatory drug development.
THE TRPV1 RECEPTOR COMPLEX AS A THERA-
PEUTIC TARGET
Similar to other TRP channels, TRPV1 is arranged inmajor molecular complexes establishing high ordersignalling networks that notably determine the response toexternal stimuli. These protein networks are in turn the targetof intracellular signaling pathways that modulate theircomposition, structure and function. Thus, an emergingnotion is that ion channels are not isolated entities in cellmembranes but pivotal components of these macromolecularassemblies. These complexes are composed of a plethora ofdiverse polypeptides including scaffolding proteins,receptors, enzymes and cytoskeletal proteins. The molecularcomponents of the TRPV1 complex identified thus farinclude the high affinity neurotrophic receptor (TrkA),phospholipase C (PLC), and calmodulin [85,95]. Thecomposition of these assemblies appears important for theregulation of the receptor activity. Activation of trkA andPLC, along with translocation of PKC, decrease the tonicinhibition that phosphatidylinositol bisphosphate exerts onTRPV1, thus promoting channel activity [23]. Similarly,activation of PKA and CAMKII gives rise to a decrease inCa
2+-induced receptor desensitisation and tachyphylaxia.
Cumulative evidence indicates that modulation of receptorcomplexes underlie the TRPV1 potentiation by inflammatorymediators.
A yeast-two hybrid screen of a rat brain library using theN-terminus of TRPV1 as a bait identified two synapticvesicle proteins, Snapin and Synaptotagmin IX, as interactingpartners of TRPV1 [83]. Noteworthy, these proteins bind toSNARE proteins and participate in neuronal exocytosis[36,55], suggesting that surface delivery of TRPV1 channelsis a highly regulated, Ca
2+-dependent exocytotic process.
Indeed, the interaction of both vesicular proteins withTRPV1 appears temporal and seems not involved in theformation of the molecular complexes at the cell surface.However, these interactions are pivotal for the traffickingand surface expression of TRPV1 channels. In support of

12 Current Neuropharmacology, 2006, Vol. 4, No. 1 Messeguer et al.
this tenet, PKC-induced TRPV1 trafficking to the plasmamembrane was blocked by botulinum neurotoxins, thusindicating that PKC- sensitisation of TRPV1 receptors is dueat least in part to the regulated exocytosis of channels locatedin a reserve pool of cytosolic vesicles [83]. A recentobservation that botulinum neurotoxin attenuates heathyperalgesia substantiates this hypothesis [27], althoughadditional in vivo experiments are needed to determine theprecise role of receptor exocytosis to the onset and main-tenance of neurogenic inflammation. Therefore, regulation ofTRPV1 surface density in nociceptor peripheral terminalsappears to be an important mechanism for both thedevelopment and preservation of inflammatory hyperalgesia.CFA-induced inflammation triggers an increased translationand trafficking of TRPV1 channels to the peripheralterminals [57]. These findings imply that modulation ofTRPV1 trafficking may be a therapeutic approach for painmanagement. Accordingly, botulinomimetic compoundssuch as small peptides that inhibit SNARE-dependentexocytosis could be developed as anti-inflammatory andanalgesic compounds [13].
CONCLUSION
The recognition of the important role the TRPV1 in theetiology of neurogenic inflammation and pain transductionhas thrust the development of down regulators of its channelactivity as analgesic drugs. Several of these compounds areinitiating clinical studies in humans and their results areawaited. However, a note of caution must be sound becauseof the pivotal and diverse physiological functions played bythe TRPV1 channel. Its widespread distribution in differenttissues should be taken into account since undiscriminatepharmacological block of the receptor may lead to severeside effects that may complicate the use of antagonists ofTRPV1 activity. Thus, the challenge of molecularneuropharmacology would be to develop receptor-selectivedrugs that preserve the physiological activity of TRPV1while down regulating the function of overactive receptors.This kind of compound would not compromise the normalsensory signaling that is essential for survival.
ACKNOWLEDGMENTS
We thank all members of our group and colleagues ofcollaboration groups for their fundamental contribution tothe results herein presented. We are indebted to the Financialsupport from the MEC, FIS, GVA, and Fundació La Caixa.
ABBREVIATIONS
TRPV1 = transient receptor potential vanilloid subunit 1
PKA = protein kinase A
PKC = protein kinase C
NGF = nerve growth factor
RTX = resiniferatoxin
SP = substance P
CGRP = calcitonin gene related peptide
NADA = N-arachidonyl-dopamine
SAR = structure-activity relationships
CFA = complete Freund adjuvant
CAMK = calmodulin kinase
PLC = phospholipase C
GI = gastrointestinal
TrkA = high affinity neurotrophic receptor.
REFERENCES
[1] Akiba,Y., Furukawa, O., Guth, P.H., Engel, E., Nastaskin, I. andKaunitz, J.D. (2001) Sensory pathways and cyclooxygenase
regulate mucus gel thickness in rat duodenum. Am. J. Physiol.Gastrointest. Liver Physiol., 280, G470-474.
[2] Appendino, G., Harrison, S., De Petrocellis, L., Daddario, N.,Bianchi, F., Schiano, F., Moriello, A., Trevisani, M., Benvenuti, F.,
Geppetti, P. and Di Marzo, V. (2003) Halogenation of a capsaicinanalogue leads to novel vanilloid TRPV1 receptor antagonists. Br.
J. Pharmacol., 139, 1417-1424.[3] Appendino, G., De Petrocellis, L., Trevisani, M., Minassi, A.,
Daddario, N., Moriello, A.S., Gazzieri, D., Ligresti, A., Campi, B.,Fontana, G., Pinna, C., Geppetti, P. and Di Marzo, V. (2005)
Development of the first ultra-potent "capsaicinoid" agonist attransient receptor potential vanilloid type 1 (TRPV1) channels and
its therapeutic potential. J. Pharmacol. Exp. Ther., 312, 561-570.[4] Avelino, A., Cruz, C., Nagy, I. and Cruz, F. (2002) Vanilloid
receptor 1 expression in the rat urinary tract. Neuroscience, 109,787-798.
[5] Bakthavatchalam, R., Hutchison, A., Desimone, R.W., Hodgetts,K.J., Krause, J.E. and White, G.G. (2002) Capsaicin receptor
ligands. WO 0208221-A.[6] Bakthavatchatam, R., Blum, C.L., Brielmann, H.L., Caldwell,
T.M., Delombaert, S., Hodgetis, K.J., Yoon, T. and Zheng, X.(2003) Substituted quinazolin-4-ylamine analogues as modulators
of capsaicin. WO 03/062209.[7] Bakthavatachalam, R., Blum, C.A., Brielmann, H., Caldwell, T.M.,
De Lombaert, S., Hodgetts, K.J. and Zhen, X. (2004) Carboxylic
acid, phosphate or phosphonate substituted quinazolin-4-ylamine
analogues as capsaicin receptor modulators. WO 04/055004.[8] Balaban, C.D., Zhou, J. and Li, H.S., (2003) Type I vanilloid
receptor expression by mammalian inner ear glanglion cells. HearRes., 175, 165-170.
[9] Balan, C., Bo, Y., Dominguez, C., Christopher, H., Gore, U.K.,Ma, V.V., Norman, M.H., Ognyanov, V.Y., Quian, Y.-X., Wang,
X., Xi, N. and Xu, S. (2004) Benzimidazole derivatives and their
use as vanilloid receptor ligands. WO 04/035549.
[10] Bhave, G., Hu, H.J., Glauner, K.S., Zhu, W., Wang, H., Brasier,D.J., Oxford, G.S. and Gereau, R.W. 4th. (2003) Protein kinase C
phosphorylation sensitizes but does not activate the capsaicinreceptor transient receptor potential vanilloid 1 (TRPV1). Proc.
Natl. Acad. Sci. USA, 100, 12480-12485.[11] Bhave, G. and Gereau, R.W. 4th (2004) Posttranslational
mechanisms of peripheral sensitization. Int. J. Neurobiol., 61, 88-106.
[12] Birder, L.A., Nakamura, Y., Kiss, S., Nealen, M.L., Barrick, S.,Kanai, A.J., Wang, E., Ruiz, G., De Groat, W.C., Apodaca, G.,
Watkins, S. and Caterina, M.J. (2002) Altered urinary bladderfunction in mice lacking the vanilloid receptor TRPV1. Nat.
Neurosci., 5, 856-860.[13] Blanes-Mira, C., Merino, J.M., Valera, E., Fernández-Ballester, G.,
Gutiérrez, L.M., Viniegra, S., Pérez-Payá, E. and Ferrer-Montiel,A. (2004) Small peptides patterned after the N-terminus domain of
SNAP25 inhibit SNARE complex assembly and regulatedexocytosis. J. Neurochemistry, 88, 124-135.
[14] Bo, Y.Y., Chakrabarti, P.P., Chen, N., Doherty, E.M., Fotsch, C.H.,Han, N., Kelly, M.G., Liu, Q., Norman, M.H., Wang, X., Zhu, J.
and Ornyanov, V.I. (2003) Vanilloid receptor ligands and their usein treatments. WO 03/049702.
[15] Bodó, E., Biró, T., Telek, A., Czifra, G., Griger, Z., Tóth, B.I.,Mescalchin, A., Ito, T., Bettermann, A., Kovács, L. and Paus, R.
(2005) Involvement of vanilloid receptor -1 (VR1/TRPV1)

Physiology and Pharmacology of the Vanilloid Receptor Current Neuropharmacology, 2006, Vol. 4, No. 1 13
signalling in human hair growth control. Am. J. Pathol., 166, 985-998.
[16] Brown, R.E., Doughty, V.A., Hollingworth, G.J., Brian, J.A.,Lindon, M.J., Moyes, C.R. and Rogers, L. (2003) Heteroaromatic
urea derivatives as VR-1 receptor modulators for treating pain. WO03/080578.
[17] Carr, M.J., Hunter, D.D., Jacoby, D.B. and Undem, B.J. (2002)Expression of tachykinins in nociceptive vagal afferent neurons
during respiratory viral infection in guinea pigs. Am. J. Respir.,165, 1071-1075.
[18] Caterina, M.J., Leffler, A., Malmberg, A.B., Martin, W.J., Trafton,J., Petersen-Zeitz, K.R., Koltzenburg, M., Basbaum, A.I. and
Julius, D. (2000) Impaired nociception and pain sensation in micelacking the capsaicin receptor. Science, 288, 306-313.
[19] Caterina, M.J. and Julius, D. (2001) The vanilloid receptor: Amolecular gateway to the pain pathway. Annu. Rev. Neurosci., 24,
487-517.[20] Cesare, P. and McNaughton, P. (1997) Peripheral pain
mechanisms. Curr. Opin. Neurobiol., 7, 457-482.[21] Chan, C.L., Facer, P., Davis, J.B., Smith, G.A., Egerton, J.,
Bountra, C., Williams, N.S. and Anand, P. (2003) Sensory fibresexpressing capsaicin receptor TRPV1 in patients with rectal
hypersensitivity and faecal urgency. Lancet, 361, 385-391.[22] Chu, C.J., Huang, S.M., De Petrocellis, L., Bisogno, T., Ewing,
S.A., Miller, J.A., Zipkin, R.E., Daddario, N., Appendino, G., DiMarzo, V. and Walker, J.M. (2003) N-oleoyldopamine, a novel
endogenous capsaicin-like lipid that produces hyperalgesia. J. Biol.Chem., 278, 13633-13639.
[23] Chuang, H.H., Prescott, E.D., Kong, H., Shields, S., Jordt, S.E.,Basbaum, A.I., Chao, M.V. and Julius, D. (2001) Bradykinin and
nerve growth factor release the capsaicin receptor fromPtdIns(4,5)P2-mediated inhibition. Nature, 411, 957-962.
[24] Clapham, D.E. (2003) TRP channels as cellular sensors. Nature,426, 517-524.
[25] Codd, L., Dax, S., Jetter, M., Mcdonell, M., Mcnally, J. andYoungman, M. (2003) Aminotetralin-derived urea modulators of
vanilloid vr1 receptor. WO 03/097586.[26] Contassot, E., Tenan, M., Schnüriger, V., Pelte, M.-F. and Dietrich,
P.-Y. (2004) Arachidonyl ethanolamide induces apoptosis ofuterine cervix cancer cells via aberrantly expressed vanilloid
receptor-1. Gynecol. Oncol., 93, 182-188.[27] Cui, M., Khanijou, S., Rubino, J. and Aoki, K. (2004)
Subcutaneous administration of botulinum Toxin A reducesformalin-induced pain. Pain, 107, 125-133.
[28] Culshaw, A.J., Gull, P., Hallet, A., Kim, H.-Y., Seiler, M.P.,Zimmermann, K., Liu, Y. and Prashad, M. (2002) Pyridine
derivatives. WO 02/076946.[29] Culshaw, A.J., Dziadulewicz, E.D., Hallet, A. and Hart, T.W.
(2004) Quinazolinone derivatives useful as anti-hyperalgesic
agents. WO 04/033435.
[30] Davis, J.B., Gray, J., Gunthorpe, M.J., Hatcher, J.P., Davey, P.T.,Overend, P., Harries, M.H., Latcham, J., Clapham, C., Atkinson,
K., Hughes, S.A., Rance, K., Grau, E., Harper, A.J., Pugh, P.L.,Rogers, D.C., Bingham, S., Randall, A. and Sheardown, S.A.
(2000) Vanilloid receptor-1 is essential for inflammatory thermalhyperalgesia. Nature, 405, 183-187.
[31] De Petrocellis, L., Bisogno, T., Maccarrone, M., Davis, J.B.,Finazzi-Agro, A. and Di Marzo, V. (2001) The activity of
anandamide at vanilloid VR1 receptors requires facilitatedtransport across the cell membrane and in limited by intracellular
metabolism. J. Biol. Chem., 276, 12856-12863.[32] De Petrocellis, L., Harrison, S., Bisogno, T., Tognetto, M., Brandi,
I., Smith, G.D., Creminon, C., Davis, J.B., Geppeti, P. and DiMarzo, V. (2001) The vanilloid receptor (VR1)-mediated effects of
anandamide are potently enhanced by the cAMP-dependent proteinkinase. J. Neurochem., 77, 1660-1663.
[33] Doherty, E.M., Fotsch, C., Bo, Y., Chakrabarti, P.P., Chen, N.,Gavva, N., Han, N., Kelly, M.G., Kincaid, J., Klionsky, L., Liu, Q.,
Ognyanov, V.I., Tamir, R., Wang, X., Zhu, J., Norman, M.H., andTreanor, J.J. (2005) Discovery of potent, orally available vanilloid
receptor-1 antagonists. Structure-activity relationship of N-arylcinnamides. J. Med. Chem., 48, 71-90.
[34] Ferrer-Montiel, A., Fernández-Carvajal, A., García-Martínez, C.,Belmonte-Martínez, C., Van Den Nest, W. and Carreño, C. (2003)
Compounds which can block the responses to chemical substances
or thermal stimuli or mediators of inflammation of nociceptors,producing method thereof and composition containing them. WO
03/097670.[35] Ferrer-Montiel, A., Garcia-Martinez, C., Morenilla-Palao, C.,
Garcia-Sanz, N., Fernandez-Carvajal, A., Fernandez-Ballester, G.and Planells-Cases, R. (2004) Molecular architecture of the
vanilloid receptor. Insights for drug design. Eur. J. Biochem., 271,1820-1826.
[36] Fukuda, M. (2002) Vesicle-associated membrane protein-2/synaptobrevin binding to synaptotagmin I promotes O-
glycosylation of synaptotagmin I. J. Biol. Chem., 277, 30351-30358.
[37] García-Martínez, C., Morenilla-Palao, C., Planells-Cases, R.,Merino, J.M. and Ferrer-Montiel, A.V. (2000) Identification of an
aspartic residue in the P-loop of the vanilloid receptor thatmodulates pore properties. J. Biol. Chem., 275, 32552-32558.
[38] García-Martínez, C., Humet, M., Planells-Cases, R., Gomis, A.,Caprini, M., Viana, F., De La Pena, E., Sanchez-Baeza, F.,
Carbonell, T., De Felipe, C., Perez-Paya, E., Belmonte, C.,Messeguer, A. and Ferrer-Montiel, A. (2002) Attenuation of
thermal nociception and hyperalgesia by VR1 blockers. Proc. Natl.Acad. Sci. USA, 99, 2374-2379.
[39] Garcia-Sanz, N., Ferández-Carvajal, A., Morenilla-Palao, C.,Planells-Cases, R., Fajardo-Sanchez, E., Fernández-Ballester, G.
and Ferrer-Montiel, A. (2004) Identification of tetramerizationdomain in the C-terminus of the vanilloid receptor. J. Neurosci.,
24, 5306-5314.[40] Gavva, N.R., Klionsky, L., Qu, Y., Shi, L., Tamir, R., Edenson, S.,
Zhang, T.J., Viswanadhan, V.N., Toth, A., Pearce, L.V., Vanderah,T.W., Porreca, F., Blumberg, P.M., Lile, J., Sun, Y., Wild, K.,
Louis, J.C. and Treanor, J.J. (2004) Molecular determinants ofvanilloid receptor sensitivity in TRPV1. J. Biol. Chem., 2 7 9,
20283-20295.[41] Geppetti, P. and Trevisani, M. (2004) Activation and sensitisation
of the vanilloid receptor, role in gastrointestinal inflammation andfunction. Br. J. Pharmacol., 141,1313-1320.
[42] Ghilardi, J.R., Rohrich, H., Lindsay, T.H., Sevcik, M.A., Schwei,M.J., Kubota, K., Halvorson, K.G., Poblete, J., Chaplan, S.R.,
Dubin, A.E., Carruthers, N.I., Swanson, D., Kuskowski, M., Flores,C.M., Julius, D. and Mantyh, P.W. (2005) Selective blockade of the
capsaicin receptor TRPV1 attenuates bone cancer pain. J .Neurosci., 25, 3126-3131.
[43] Gomtsyan, A., Bayburt, E.K., Schmidt, R.G., Zeng, G.Z., Perner,R.J., Didomenico, S., Koenig, J.R., Turner, S., Jinkerson, T.,
Drizin, I., Hannick, S.M., Macri, B.S., McDonald, H.A., Honore,P., Wismer, C.T., Marsh, K.C., Wetter, J., Stewart, K.D., Oie, T.,
Jarvis, M.F., Surowy, C.S., Faltynek, C.R., and Lee, C.-H. (2005)Novel transient receptor potential vanilloid 1 receptor antagonists
for the treatment of pain, Structure-activity relationships for ureaswith quinoline, isoquinoline, quinazoline, phthalazine, quinoxaline
and cinnoline moieties. J. Med. Chem., 48, 744-752.[44] Groneberg, D.A., Quarcoo, D., Frossard, N. and Fischer, A. (2004)
Neurogenic mechanisms in bronchial inflammatory diseases.Allergy, 59, 1139-1152.
[45] Gunthorpe, M.J., Rami, H.K., Jeman, J.C., Smart, D., Gill, C.H.,Soffin, E.M., Hannan, S.L., Lappin, S.C., Egerton, J., Smith, G.D.,
Worby, A., Howett, L., Owen, D., Nasir, S., Davies, C.H.,Thompson, M., Wyman, P.A., Randall, A.D. and Davis, J.B. (2004)
Identification and characterization of SB-366791, a potent and aselective vanilloid receptor (VR1/TRPV1) antagonist.
Neuropharmacol., 46,133-149.[46] Guo, A., Vluchanova, L., Wang, J., Li, X. and Elde, R. (1999)
Immunocytochemical localization of the vanilloid receptor I (VR1),relationship to neuropeptides, the P2X3 purinoreceptor and IB4
binding sites. Eur. J. Neurosci., 11, 946-958.[47] Hann, M.M., Leach, A.R. and Harper, G. (2001) Molecular
complexity and its impact on the probability of finding leads fordrug discovery. J. Chem. Inf. Comput. Sci., 41, 856-864.
[48] Himmel, H.M., Kiss, T., Borvendeg, S.J., Gillen, C. and Ille, P.(2002) The arginine rich hexapeptide R4W2 is a stereoselective
antagonist of the vanilloid receptor 1, A Ca2+imaging study in adultrat dorsal root ganglion neurons. J. Pharmacol. Exp. Ther., 301,
981-986.

14 Current Neuropharmacology, 2006, Vol. 4, No. 1 Messeguer et al.
[49] Holzer, P. (2004). TRPV1 and the gut, from a tasty receptor for apainful vanilloid to a key player in hyperalgesia. Eur. J.
Pharmacol., 500, 231-241.[50] Huang, S.M., Bisogno, T., Trevisani, M., Al-Hayani, A., De
Petrocellis, L., Fezza, F., Tognetto, M., Petros, T.J., Krey, J.F.,Chu, C.I., Miller, J.D., Davies, S.N., Geppetti, P., Walker, J.M. and
Di Marzo, V. (2002) An endogenous capsaicin-like substance withhigh potency at recombinant and native VR1 receptors. Proc. Natl.
Acad. Sci. USA, 99, 8400-8405.[51] Hunter, D.D., Myers, A.C. and Undem, B.J. (2000) Nerve-growth
factor induced phenotypic switch in guinea pig airway sensoryneurons. Am. J. Respir., 161, 1985-1990.
[52] Hwang, S.W., Cho, J., Kwak, J., Lee, S.Y., Kang, J., Jung, S., Cho,K.H., Min, Y.G., Suh, D., Kim, U. and Oh, U. (2000) Direct
activation of capsaicin receptors by products of lipooxygenases,endogenous capsaicin-like substances. Proc. Natl. Acad. Sci. USA,
97, 6155-60.
[53] Hwang, S.J. and Valtschanoff, J.G. (2003) Vanilloid receptor VR1-
positive afferents are distributed differently at different levels ofthe rat lumbar spinal cord. Neuorsci. Lett., 349, 41-44.
[54] Ichikawa, H. and Sugimoto, T. (2003) The co-expression of VR1and VRL-1 in the rat vagal sensory ganglia. Brain Res., 980, 293-
296.[55] Ilardi, J.M., Mochida, S., Sheng, Z.H. (1999) Snapin, a SNARE-
associated protein implicated in synaptic transmission. Nat.Neurosci., 2,119-124.
[56] Inoue, K., Koizumi, S., Fuziwara, S., Denda, S., Inoue, K. andDenda, M. (2002) Functional vanilloid receptors in cultured normal
human epidermal keratinocytes. Biochem. Biophys. Res. Commun.,291,124-129.
[57] Ji, R.-R., Samad, T.A., Jin, S.-X., Schmoll, R. and Woolf, C.(2002) p38 MAPK activation by NGF in primary sensory neurons
after inflammation increases TRPV1 levels and maintains heathyperalgesia. Neuron, 36, 57-68.
[58] Jin, X., Morsy, N., Winston, J., Pasricha, P.J., Garret, K. andAkbarali, H.I. (2004) Modulation of TRPV1 by nonreceptor
tyrosine kinase, c-Src kinase. Am. J. Physiol. Cell Physiol., 287,C558-C563.
[59] Jordt, S.E. and Julius, D. (2002) Molecular basis for species-specific sensitivity to hot chilli peppers. Cell, 108, 421-430.
[60] Jordt, S.E., Bautista, D.M., Chuang, H.H., McKemy, D.D.,Zygmunt, P.M., Hogestatt, E.D., Meng, I.D. and Julius, D. (2004)
Mustard oils and cannabinoids excite sensory nerve fibres throughthe TRP channel ANKTM1. Nature, 427, 260-265.
[61] Julius, D. and Basbaum, A.I. (2001) Molecular mechanisms ofnociception. Nature, 413, 203-210.
[62] Jung, J., Hwang, S.W., Kwak, J., Lee, S.Y., Kang, J.C., Kim, W.B.,Kim, D. and Oh, U. (1999) Capsaicin binds to the intracellular
domain of the capsaicin activated ion channel. J. Neurosci., 19,529-538.
[63] Jung, J., Shin, J.S., Lee, S.Y., Hwang, S.W., Koo, J., Cho, H. andOh, U. (2004) Phosphorylation of vanilloid receptor 1 by
Ca2+/calmodulin-dependent kinase II regulates its vanilloid binding.J. Biol. Chem., 279, 7048-7054.
[64] Karai, L., Brown, D.C., Mannes, A.J., Connelly, S.T., Brown, J.,Gandal, M., Wellisch, O.M., Neubert, J.K., Olah, Z. and Iadarola,
M.J. (2004) Deletion of vanilloid receptor 1-expressing primaryafferent neurons for pain control. J. Clin. Invest., 113, 1344-1352.
[65] Karlsson, J.A. (1993) A role for capsaicin sensitive, tachykinincontaining nerves in chronic coughing and sneezing but not in
asthma, a hypothesis. Thorax, 48, 396-400.[66] Kim, H., Neubert, J.K., San Miguel, A., Xu, K., Krishnaraju, R.K.,
Iadarola, M.J., Goldman, D. and Dionne, R.A. (2004) Geneticinfluence on variability in human acute experimental pain
sensitivity associated with gender, ethnicity and psychologicaltemperament. Pain, 109, 488-496.
[67] Kyle, D.J., Sun, Q., Fatesse, L., Zhang, C. and Zhou, X. (2004)
Therapeutic agents useful for treating pain. WO 04/002983.
[68] Lee, J., Kang, M., Shin, M., Kim, J.M., Kang, S.U., Lim, J.O.,Choi, H.K., Suh, Y.G., Park, H. G., Oh, U., Kim, H.D., Park, Y.
H., Ha, H.J., Kim, Y.H., Toth, A., Wang, Y., Tran, R., Pearce,L.V., Lundberg, D.J. and Blumberg, P.M. (2003) N-(3-Acyloxy-
2-benzylpropyl)-N[4-(methylsulfonylamino) benzyl]thiourea anal-ogues, novel potent and high affinity antagonists and partial
antagonists of the vanilloid receptor. J. Med. Chem., 46, 3116-3126.
[69] Lee, C.-H., Bayburt, E.K., Didomenico, D.S., Drizin, I., Gomtsyan,A.R., Koenig, J.R., Perner, R.J., Schmidt, R.G., Turner, S.C.,
White, T.K. and Zheng, X. (2003) Fused azabicyclic compoundsthat inhibit VR-1. WO 03/070247.
[70] Lipinski, C.A., Lombardo, F., Dominy, B.W. and Feeney, P.J.(2001) Experimental and computational approaches to estimate
solubility and permeability in drug discovery and developmentsettings. Adv.Drug Deliv. Rev., 46, 3-26.
[71] Marinelli, S., Vaughan, C.W., MacDonald, J.C. and Connor, M.(2002) Capsaicin activation of glutamatergic synaptic transmission
in the rat locus coeruleus in vitro. J. Physiol., 543.2, 531-540.
[72] Marinelli, S., Pascucci, T., Bernardi, G., Puglisi-Allegra, S. and
Mercuri, N.B. (2005) Activation of TRPV1 in the VTA excitesdopaminergic neurons and increases chemical- and noxious-
induced dopamine release in the nucleus accumbens.Neuropsychopharmacology, 30, 864-870.
[73] McDonnel, M.E., Zhang, S.P., Dubin, A.E. and Dax, S.L. (2002)Synthesis and in vitro evaluation of a novel iodinated
resiniferatoxin derivative that is an agonist at the human vanilloidVR1 receptor. Bioorg. Med. Chem. Lett., 12, 11189-11192.
[74] McKemy, D.D., Neuhasser, W.M. and Julius, D. (2002)Identification of a cold receptor reveals a general role for TRP
channels in thermosensation. Nature, 416, 52-58.[75] Melck, D., Bisogno, T., De Petrocellis, L., Chuang, H., Julius, D.,
Bifulco, M. and Di Marzo, V. (1999) Unsaturated long chain N-acyl-vanillyl-amides (N-AVAMS), vanilloid receptor ligands that
inhibit anandamide-facilitated transpor and bind to CB1cannabinoid receptors. Biochem. Biophys. Res. Commun., 262, 275-
284.[76] Mellor, I.R., Ogilvie, J., Pluteanu, F., Clothier, R.H., Parker, T.L.,
Rosini, M., Minarini, A., Tumiatti, V. and Melchiorre, C. (2004)Methoctramine analogues inhibit responses to capsaicin and
protons in rat dorsal root ganglion neurons. Eur. J. Pharmacol.,505; 37-50.
[77] Minke, B. and Cook, B. (2002) TRP channel proteins and signaltransduction. Physiol. Rev., 82, 429-472.
[78] Mohapatra, D.P. and Nau, C. (2005) Regulation of Ca2+-dependentdesensitization in the vanilloid receptor TRPV1 by calcineurin and
cAMP-dependent protein kinase. J. Biol. Chem., 280 , 13424-13432.
[79] Montell, C., Birnbaumer, L. and Flockerzi, V. (2002) The TRPchannels, a remarkably functional family. Cell, 108; 595-598.
[80] Montell, C. (2003) Thermosensation, Hot findings make TRPNsvery cool. Curr. Biol., 13, R476-R478.
[81] Moqrich, A., Hwang, S.W., Earley, T.J., Petrus, M.J., Murray,A.N., Spencer, K.S., Andahazy, M., Story, G.M. and Patapoutian,
A. (2005) Impaired thermosensation in mice lacking TRPV3, a heatand camphor sensor in the skin. Science, 307, 1468-1472.
[82] Moran, M.M., Xu, H. and Clapham, D.E. (2004) TRP ion channelsin the nervous system. Curr. Opin. Neurobiol., 14, 362-369.
[83] Morenilla-Palao, C., Planells-Cases, R., García-Sanz, N. andFerrer-Montiel, A. (2004) Regulated exocytosis contributes to
protein kinase C potentiation of vanilloid receptor activity. J. Biol.Chem., 279, 25665-25672.
[84] Nagy, I., Sántha, P., Jancsó, G. and Urbán, L. (2004) The role ofvanilloid (capsaicin) receptor (TRPV1) in physiology and
pathology. Eur. J. Pharmacol., 500, 351-369.[85] Numazaki, M., Tominaga, T., Takeuchi, K., Murayama, N.,
Toyooka, H. and Tominaga, M. (2003) Structural determinant ofTRPV1 desensitization interacts with calmodulin. Proc. Natl. Acad.
Sci. USA, 100, 8002-8006.[86] Obreja, O., Rathee, K.P., Lips, K.S., Distler, C. and Kress, M.
(2002) IL-1 potentiates heat-activated currents in rat sensoryneurons, involvement of IL-1R1, tyrosine kinase and protein kinase
C. FASEB J., 16, 1497-1503.[87] Olah, Z., Karai, L., and Iadarola, M.J. (2002) Protein kinase
C(alpha) is required for vanilloid receptor 1 activation. Evidencefor multiple signalling pathways. J. Biol. Chem., 277, 35752-
35759.[88] Pare, M., Elde, R., Mazurkiewicz, J.E., Smith, A.M. and Rice, F.L.
(2001) The Meissner corpuscule revised, a multiafferentmechanoreceptor with nociceptor immunological properties. J.
Neurosci., 21, 7236-7246.

Physiology and Pharmacology of the Vanilloid Receptor Current Neuropharmacology, 2006, Vol. 4, No. 1 15
[89] Peier, A.M., Moqrich, A., Hergarden, A.C., Reeve, A.J., Anderson,D.A., Story, G.M., Earley, T.J., Dragoni, I., McIntyre, P., Bevan, S.
and Patapoutian, A. (2002) A TRP channel that senses cold stimuliand menthol. Cell, 108, 705-715.
[90] Planells-Cases, R., Aracil, A., Merino, J., Gallar, J., Pérez-Payá, E.,Belmonte, C. and Ferrer-Montiel, A.V.(2000) Arginine rich
peptides are blockers of VR-1 channels with analgesic activity.FEBS Lett., 481, 131-36.
[91] Planells-Cases, R., Garcia-Martinez, C., Royo, M., Pérez-Payá, E.,Carreño, C., Albericio, F., Messeguer, A. and Ferrer-Montiel, A.
(2003) Small molecules targeting the vanilloid receptor complex asdrugs for inflammatory pain. Drugs Future, 28, 787-795.
[92] Pomonis, J.D., Harrison, J.E., Mark, L., Bristol, D.R., Valenzano,K.J. and Walker, K. (2003) N-(4-Tertiarybutylphenyl)-4-(3-
cholorphyridin-2-yl)tetrahydropyrazine-1(2H)-carboxi-amide(BCTC), a novel, orally effective vanilloid receptor 1 antagonist
with analgesic properties, II. in vivo characterization in rat modelsof inflammatory and neuropathic pain. J. Pharmacol. Exp. Ther.,
306, 387-393.[93] Premkumar, L.S. and Ahern, G.P. (2000) Induction of vanilloid
receptor channel activity by protein kinase C. Nature, 408, 985-990.
[94] Rami, H.K., Thompson, M., Wyman, P., Jerman, J.C., Egerton, J.,Brough, S., Stevens, A.J., Randall, A.D., Smart, D., Gunthorpe,
M.J. and Davis, J.B. (2004) Discovery of small moleculeantagonists of TRPV1. Bioorg. Med. Chem. Lett., 14, 3631-3634.
[95] Rosenbaum, T., Gordon-Shaag, A., Munari, M. and Gordon, S.E.(2004) Ca2+/calmodulin modulates TRPV1 activation by capsaicin.
Brain Res., 996, 176-183.[96] Schaible, H.G. and Richter, F. (2004) Pathophysiology of pain.
Langenbecks Arch. Sur., 389, 237-243.[97] Scotland, R.S., Chauhan, S., Davis, C., DeFelipe, C., Hunt, S.,
Kabir, J., Kotsonis, P., Oh, U. and Ahluwalia, A. (2004) Vanilloidreceptor TRPV1, sensory C fibers, and vascular autoregulation, a
novel mechanism involved in myogenic constriction. Circ. Res.,95, 1027-1034.
[98] Smith, G.D., Gunthorpe, M.J., Kelsell, R.E., Hayes, P.D., Reilly,P., Facer, P., Wright, J.E., Jerman, J.C., Walhin, J.P., Ooi, L.,
Egerton, J., Charles, K.J., Smart, D., Randall, A.D., Anand, P. andDavis, J.B. (2002) TRPV3 is a temperature-sensitive vanilloid
receptor-like protein. Nature, 418, 186-190.[99] Stander, S., Moormann, C., Schumacher, M., Buddenkotte, J.,
Artuc, M., Shpacovitc, V., Brzoska, T., Lippert, U., Henz, B.M.,Luger, T.A., Metze, D. and Steinhoff, M. (2004) Expression of
vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mastcells, and epithelial cells of appendage structures. Exp. Dermatol.,
13, 129-139.[100] Story, G.M., Peier, A.M., Reeve, A.J., Eid, S.R., Mosbacher, J.,
Hricik, T.R., Earley, T.J., Hergarden, A.C., Anderesson, D.A.,Hwang, S.W., McIntyre, P., Jegla, T., Bevan, S. and Patapoutian,
A. (2003) ANKTM1, a TRP-like channel expressed in nociceptiveneurons, is activated by cold temperatures. Cell, 112, 819-829.
[101] Strecker, T., Messlinger, K., Weyand, M. and Reeh, P.W. (2005)Role of different proton sensitive channels in releasing calcitonin
gen related peptide from isolated hearts and mutant mice.Cardiovascular Res., 5, 405-410.
[102] Sugiura, T., Tominaga, M., Katsuya, H. and Mizumura, K. (2002)Bradykinin lowers the threshold temperature for heat activation of
vanilloid receptor 1. J, Neurophysiol., 88, 544-548.[103] Szallasi, A. and Blumberg, P.M. (1999) Vanilloid (Capsaicin)
receptors and mechanisms. Pharmacol. Rev., 51, 159-212.[104] Szallasi, A. and Di Marzo, V. (2000) New perspectives on
enigmatic vanilloid receptors. Trends Neurosci., 23, 491-497.[105] Szallasi, A. (2001) Vanilloid receptor ligands, hopes and realities
for the future. Drugs Aging, 18, 561-573.
[106] Szallasi, A. and Appendino, G. (2004) Vanilloid receptor TRPV1antagonists as the next generation of painkillers. Are we putting the
cart before the horse. J. Med. Chem., 47, 2717-2723.[107] Tominaga, M., Caterina, M.J., Malmberg, A.B., Rosen, T.A.,
Gilbert, H., Skinner, K., Raumann, B.E., Basbaum, A.I. and Julius,D. (1998) The cloned capsaicin receptor integrates multiple pain-
producing stimuli. Neuron, 21, 531-543[108] Tympanidis, P., Casula, M.A., Yiangou, Y., Terenghi, G., Dowd, P.
and Anand, P. (2004) Increased vanilloid receptor VR1 innervationin vulvodynia. Eur. J. Pain, 8, 129-133.
[109] Van der Stelt, M. and Di Marzo, V. (2004) Endovanilloids,Putative endogenous ligands of transient receptor potential
vanilloid 1 channels. Eur. J. Biochem., 271, 1827-1834.[110] Vellani, V., Mapplebeck, S., Moriondo, A., Davis, J.B. and
McNaughton, P.A. (2001) Protein kinase C activation potentiatesgating of the vanilloid receptor VR1 by capsaicin, protons, heat and
anandamide. J. Physiol., 534, 813-825.[111] Voets, T., Droogmans, G., Wissenbach, U., Janssens, A., Flockerzi,
V. and Nilius, B. (2004) The principle of temperature–dependentgating in cold- and heat-sensitive TRP channels. Nature, 430, 748-
754.[112] Wahl, P., Foged, C., Tullin, S. and Thomsen, C. (2001) Iodo-
Resiniferatoxin, a New Potent Vanilloid Receptor Antagonist. Mol.Pharmacol., 59, 9–15.
[113] Walker, K.M., Urban, L., Medhurst, S.J., Patel, S., Panesar, M.,Fox, A.J. and McIntyre, P. (2003) The VR1 antagonist capsazepine
reverses mechanical hyperalgesia in models of inflammatory andneuropathic pain. J. Pharmacol. Exp. Ther., 304, 56-62.
[114] Wang, Y., Szabo, T., Welter, J.D., Toth, A., Tran, R., Lee, J.,Kang, S.U., Suh, Y.G., Blumberg, P.M., and Lee, J. (2002) High
affinity antagonists of the vanilloid receptor. Mol. Pharmacol.,62,947-956.
[115] Wang, Y., Kedei, N., Wang, M., Wang, Q.J., Huppler, A.R., Toth,A., Tran, R. and Blumberg, P.M. (2004) Interaction between
protein kinase C-mu and the vanilloid receptor type 1. J. Biol.Chem., 279, 53674-53682.
[116] Ward, S.M., Bayguinov, J., Won, K.J., Grundy, D. and Berthoud,H.R. (2003) Distribution of the vanilloid receptor (VR1) in the
gastrointestinal track. J. Comp. Neurol., 465, 121-135.[117] Woodbury, C.J., Zwick, M., Wang, S., Lawson, J.J., Caterina, M.J.,
Koltzenburg, M., Albers, K.M., Koerber, H.R. and Davis, B.M.(2004) Nociceptors lacking TRPV1 and TRPV2 have normal heat
responses. J. Neurosci., 24, 6410-6415.[118] Yiangou, Y., Facer, P., Dyer, N.H., Chan, C.L., Knowles, C.,
Williams, N.S. and Anand, P. (2001) Vanilloid receptor 1immunoreativity in inflamed human bowel. Lancet, 357, 1338-
1339.[119] Yura, T., Mogi, M., Ikegami, Y., Masuda, T., Kokubo, T., Urbahns,
K., Yoshida, N., Marumo, N., Shiroo, M., Masaomi, T., Takeshita,K., Moriwaki, T. and Tusikimi, Y. (2003) Urea derivatives WO
03/055484.[120] Yura, T., Mogi, M., Ikegami, Y., Masuda, T., Kokubo, T., Urbahns,
K., Yoshida, N., Marumo, N., Shiroo, M., Masaomi, T., Takeshita,K., Moriwaki, T. and Tusikimi, Y. (2003) Hydroxytetrahydro-
naphtalenylurea derivatives. WO 03/095420.[121] Zheng, J., Dai, C., Steyger, P.S., Kim, Y., Vass, Z., Ren, T. and
Nuttal, A.L. (2003) Vanilloid receptors in hearing, altered cochlearsensitivity by vanilloids and expression of TRPV1 in the organ of
corti. J. Neurophysiol., 90, 444-455.[122] Zymunt, P.M., Peterson, J., Anderson, D.A., Chuang, H., Sorgard,
M., Di Marzo, V., Julius, D. and Hogestatt, E.D. (1999) Vanilloidreceptors on sensory nerves mediates the vasodilator action of
anandamide. Nature, 400, 452-457.
Received: April 28, 2005 Revised: July 18, 2005 Accepted: August 18, 2005