cyclopropyl sterol phospholipid composition membrane ... · cyclopropyl sterol andphospholipid...
TRANSCRIPT

Plant Physiol. (1989) 90, 591-5970032-0889/89/90/0591 /07/$01 .00/0
Received for publication October 12, 1988and in revised form January 27, 1989
Cyclopropyl Sterol and Phospholipid Composition ofMembrane Fractions from Maize Roots Treated with
Fenpropimorph
Anne Grandmougin, Pierrette Bouvier-Nave, Pascaline Ullmann, Pierre Benveniste, andMarie-Andree Hartmann*
Laboratoire de Biochimie V6getale, URA CNRS No. 1182, Universite Louis Pasteur, Institut de Botanique, 28 rueGoethe, 67083 Strasbourg Cedex, France
ABSTRACT
Maize (Zea mays L.) caryopses were grown in the presence offenpropimorph, a systemic fungicide, for 7 days in the dark.Membrane fractions enriched, respectively, in endoplasmic retic-ulum, plasma membrane, and mitochondria were isolated fromcontrol and treated maize roots and analyzed for their free sterol,phospholipid, and fatty acid composition. In treated plants, theintracellular distribution of free sterols was dramatically modifiedboth qualitatively and quantitatively. The normally occurring A5-sterols disappeared almost completely and were replaced by9,B,19-cyclopropyl sterols, mainly cycloeucalenol and 24-methylpollinastanol. These new compounds were found to accumulatein all the membrane fractions in such a way that the endoplasmicreticulum-rich fraction became the richest one in free sterolsinstead of the plasma membrane. In contrast, the fenpropimorphtreatment of maize roots was shown not to affect either therelative proportions or the amounts of the individual phospho-lipids, but an increase in the unsaturation index of phospholipid-fatty acyl chains of the endoplasmic reticulum-rich fraction wasobserved. The present data suggest that, in higher plant mem-branes, cyclopropyl sterols could play a structural role similar tothat of the bulk of A5-sterols.
Whereas animal and fungal cells generally contain onemajor sterol, respectively, cholesterol (1)1 and ergosterol (17),
' Chemical nomenclature: cholesterol (1), cholest-5-en-3,B-ol; 24-methyl cholesterol (2), (24r)-24-methylcholest-5-en-3(3-ol; stigmas-terol (3), (24S)24-ethylcholesta-5,22E-dien-33-ol; sitosterol (4), (24R)-24-ethylcholest-5-en-3f3-ol; isofucosterol (5), stigmasta-5,24(28)Z-dien-3f-ol; 24-methyl pollinastanol (6), (24r)-24,14a-di-methyl-9fl, 19-cyclo-5a-cholestan-3#-ol; 24-methylene pollinastanol(7), 14a-methyl-9f,19-cyclo-5a-ergost-24(28)-en-33-ol; 24-ethyl pol-linastanol (8), 14a-methyl-9j3, 19-cyclo-5a-stigmastan-3f-ol; obtusi-foliol (9), 4a, 14a-dimethyl-5a-ergosta-8,24(28)-dien-3(-ol; cycloeu-calenol (10), 4a, 14a-dimethyl-9(, 19-cyclo-5a-ergost-24(28)-en-3(-ol;24-dihydrocycloeucalenol (11), (24R)-24,4a, 14a-trimethyl-9,B, 19-cy-clo-5a-cholestan-3,B-ol; 31-norcyclobranol (12), 4a,14a-dimethyl-9,3,19-cyclo-5a-ergost-24(25)-en-3fl-ol; cyclofuntumienol (13), 4a,14a-dimethyl-93,19-cyclo-5a-stigmast-24(28)Z-en-3,3-ol; cycloar-tenol (14), 4,4,14a-trimethyl-9,B, 19-cyclo-5a-cholest-24-en-3,-ol; 24-methylene cycloartanol (15), 4,4,14a-trimethyl-93, 19-cyclo-5a-er-gost-24(28)-en-33-ol; f-amyrin (16), olean-12-en-3,B-ol; ergosterol(17), (24R)-24-methylcholesta-5,7,22E-trien-3,B-ol; fenpropimorph,(R,S)-4-[3-(4-tert-butylphenyl)-2-methyl-propyl]-2,6-dimethylmor-pholine.
in most higher plants, sterols are present as mixtures in which24-methyl cholesterol (2), stigmasterol (3), and sitosterol (4)often predominate (20). These A5-sterols mostly accumulatein the plasma membrane (pM)2 (13), where they are believedto regulate the membrane fluidity and, consequently, theactivity of membrane-bound enzymes. In addition to thisstructural role, a few sterol molecules might be involved inmetabolic functions. To obtain a better insight into sterolfunctions in plants, we have adopted a strategy consisting ofmodifying qualitatively and quantitatively the sterol compo-sition of higher plant membranes by using sterol biosynthesisinhibitors. Thus, when maize caryopses are grown in thepresence of tridemorph or fenpropimorph, two N-substitutedmorpholines known to interfere with ergosterol biosynthesis(15), the A5-sterols disappear almost completely and are re-placed by 93, 19-cyclopropyl sterols, mainly cycloeucalenol(10) and 24-methyl pollinastanol (6) (Fig. 1). Such plantsconstitute the most suitable material for studying structuraland functional roles of sterols. Because the structural differ-ences between A5-sterols and 9(,19-cyclopropyl sterols couldinduce some changes in the membrane lipid organization, thenext important questions to be answered were the following:(a) can these unusual sterols serve as membrane components,and (b) how are these cyclopropyl sterols distributed amongthe different cell membrane systems? For that purpose, mem-brane fractions enriched in ER, PM, and MIT were isolatedfrom control and fenpropimorph-treated maize roots andanalyzed for their sterol composition. The present paperprovides evidence that treatment of maize roots with thefungicide leads to dramatic changes in both the qualitativeand quantitative intracellular distribution of free sterols. Analmost complete disappearance of the normally occurring A5-sterols and their replacement by 9(,B19-cyclopropyl sterolswere observed in all the fractions, especially in the ER-richfraction. Because a retailoring of the PL molecular species(i.e. changes in the head group composition or in the degreeof unsaturation ofFA chains) in response to the altered sterol
2 Abbreviations: PM, plasma membrane; MIT, mitochondria;AdoMet-CMT, S-adenosyl-L-methionine-cycloartenol-C24-methyltransferase; COI, cycloeucalenol-obtusifoliol isomerase; UDPG-SGT,UDP-glucose:sterol-f3-E-glucosyl transferase; PL, phospholipids; PA,phosphatidic acid; PC, phosphatidylcholine; PE, phosphatidyletha-nolamine; PG, phosphatidylglycerol; PI, phosphatidylinositol; PS,phosphatidylserine; FA, fatty acids; UI, unsaturation index.
591 www.plantphysiol.orgon January 7, 2020 - Published by Downloaded from
Copyright © 1989 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 90,1989
I -
,g,-'C'--e-*' 3-
Figure 1. Biosynthesis of sterols in control and fenpropimorph-treated maize roots with the COI as the major target. COI, Cycloeucalenol-obtusifoliol isomerase; CMT, AdoMet-cycloartenol-C24-methyl transferase; Si, A8__A7-sterol isomerase.
pattern could be expected to take place, both PL and FAcontents of the different membranes were also determined. Itwas observed that the fenpropimorph treatment of maizeroots did not affect either the relative proportions or theamounts of the individual PL. But a significant increase inthe UI of PL-FA chains was found in the ER-rich fraction.Some preliminary aspects ofthis work related to sterol analysiswere previously published elsewhere (3, 12).
MATERIALS AND METHODS
Plant Material
Maize (Zea mays L., cv LG 11) caryopses were allowed togerminate in moist vermiculite in the dark at 25°C for 1 week.The vermiculite was soaked daily with water with or without20 mg/L of (R,S) fenpropimorph (0.5 L for 85 g, i.e. 300grains). Fenpropimorph was first dissolved in ethanol, but thefinal concentration of ethanol never exceeded 0.1 %.
Isolation of Membranes
Roots from 7-d-old control and treated plants were excised,chopped into small pieces, then disrupted in a blender withan appropriate buffered medium: 0.5 M mannitol, 5 mMEDTA, 10 mm 2-mercaptoethanol, 0.5% (w/v) BSA, and 0.1M Tris-HCl (pH 8.0) (1 mL/g of fresh material). Membranefractions were isolated by differential and isopycnic sucrose
density gradient centrifugation as described previously (1 1).Three purified membrane fractions were used in the presentwork: (a) a light fraction (density: 1.10 g/mL) rich in ER; (b)a heavy fraction (1.17 g/mL) that mainly comprised PMvesicles; and (c) a fraction enriched in mitochondria (1.18 g/mL). Membranes were resuspended in 0.1 M Tris-HCl (pH8.0) containing 1 mm 2-mercaptoethanol. Specific subcellularcomponents (ER, PM, and MIT) were identified by assays forthe following markers: ER, antimycin A-insensitive NADHCyt c reductase, AdoMet-CMT and COI; PM, vanadate-sensitive H+-ATPase (pH 6.5) and UDPG-SGT; MIT, anti-mycin A-sensitive NADH Cyt c reductase.
Enzyme AssaysNADH Cyt c reductase was determined spectrophotomet-
rically (1 1). ATPase activity of membrane fractions (30-50,gg of protein) was measured in the presence of 3 mm Tris-ATP, 3 mM MgS04, 25 mM KCI, 25 mM K2S04, 0.25 Msucrose, 1 mm NaN3, 0.05% (w/v) Triton X-100, and 20 mMTris-Mes (pH 6.5) in a total volume of 0.5 mL. Sodium azidewas introduced in all the assays to inhibit any contaminatingactivity from the mitochondrial ATPase. After a 25-minincubation at 30°C, ATP hydrolysis was stopped by additionof 1 mL of 5% (w/v) SDS. The Pi released was determined asdescribed by Hartmann and Benveniste (11). The ATPasesensitivity toward Na orthovanadate (100 gM) was routinelytested.
592 GRANDMOUGIN ET AL.
tI -1
i
www.plantphysiol.orgon January 7, 2020 - Published by Downloaded from Copyright © 1989 American Society of Plant Biologists. All rights reserved.

STEROLS AND PHOSPHOLIPIDS IN FENPROPIMORPH-TREATED MAIZE
COI was assayed by incubating, at 30°C for 1 h, 0.4 mL ofmembrane preparation (0.3-0.6 mg of protein) with 100 AMcycloeucalenol and 0.1 % (w/v) Tween-80 in a total volumeof 0.5 mL. The reaction was stopped by the addition of 0.5mL of 6% ethanolic KOH. The neutral lipids were extractedthree times with 3 volumes of hexane and subjected to TLCwith dichloromethane as the solvent (two runs). The 4a-methyl sterols (RF 0.45) were scraped off, eluted, and analyzedby GLC as described previously (13). Quantitative determi-nations were made by measuring peak areas ofcycloeucalenoland obtusifoliol.AdoMet-CMT was assayed at pH 8.0 by incubating 0.4 mL
of membrane fraction (0.3-0.6 mg of protein) with 60 Mmcycloartenol, 0.12% (w/v) Tween-80, and 85 uM [methyl-3H]AdoMet (0.6 ,uCi) in a total volume of 0.5 mL. After a 1-hincubation at 30C, the reaction was stopped by the additionof0.5 mL of6% ethanolic KOH. The mixtures were extractedthree times with three volumes of hexane, and the radioactiv-ity incorporated into 24-methylene cycloartanol wasmeasured.UDPG-SGT was assayed as described previously (23).
Membrane Protein Determination
Membrane protein was determined according to Schacterleand Pollack (25) with BSA as a standard (Fraction V, SigmaChemical Co).
Lipid Analyses
Sterols were extracted and analyzed according to Hartmannand Benveniste (13). Phospholipids were extracted and quan-tified as described previously (33).
Fatty Acids
FA methyl esters from total PL were prepared by transes-terification with 0.1 M KOH in anhydrous methanol at 37°Cfor 30 min. They were analyzed by GLC on a Superox glasscapillary column (60 m) at 165°C. Quantitative determina-tions were made with heptadecanoic (C 17:0) methyl ester asan internal reference.
Chemicals
UDP-[U-'4C]glucose and S-adenosyl-L-[methyl-3H]methi-onine were purchased from Amersham (U.K.). All otherchemicals were from Sigma Chemical Co. Fenpropimorphwas kindly supplied by BASF Co. (F.R.G.).
RESULTS
Enzymic Characterization of Membrane Fractions fromControl and Fenpropimorph-treated Maize Roots
Several membrane fractions were isolated from control andfenpropimorph-treated maize roots by differential and sucrose
density gradient centrifugation according to a procedure pre-viously tested with maize coleoptiles (11). Three types offractions were prepared: (a) purified mitochondria (d: 1.18 g/mL) from a 10,000g pellet, (b) a light membrane fraction (d:
1.10 g/mL), and (c) a heavy membrane fraction (d: 1.17 g/mL) from a 100,000g pellet.
Fractions from Control Maize Roots
The activities of the antimycin A-insensitive NADH-Cyt coxidoreductase, a classical marker for ER membranes, and ofCOI and AdoMet-CMT, two enzy-mes involved in free sterolbiosynthesis, were shown to be mainly associated with thelight fraction (Table I). The same fraction was also found tocontain high activities of cinnamic acid-4 hydroxylase and inchain-laurate hydroxylase, two Cyt P450 monooxygenases(data not shown), indicating the presence in this fraction ofmembranes derived from the ER. The occurrence of impor-tant activities ofUDPG-SGT and H+-ATPase (pH 6.5) in theheavy fraction suggests that this fraction is enriched in PMvesicles (11, 28). ATPase activity was assayed in the presenceof sodium vanadate or sodium nitrate; it was found to be60% inhibited by 100 Mm sodium vanadate but totally resistantto 0.1 M sodium nitrate, suggesting the absence of contami-nating tonoplast membranes (28). A high sterol to PL molarratio is also shown to be associated with the heavy fraction(Table II), and this constitutes an additional criterion foridentifyingPM vesicles. Mitochondrial membranes were char-acterized by a NADH-Cyt c oxidoreductase activity, whichwas sensitive to both antimycin A and KCN; no significantsuccinate dehydrogenase activity, another marker for internalmitochondrial membranes, could be measured in thisfraction.
Fractions from Treated Maize Roots
The treatment of maize seedlings with fenpropimorph wasfound to induce a modification of the morphology of rootsconsisting of a strong decrease in the length, an increase inthe thickness of the main root, and a reduction in the numberof adventitious roots. As shown in Table I, such a modifica-tion did not affect the distribution of enzyme markers. Theantimycin A-insensitive NADH-Cyt c oxidoreductase andAdoMet-CMT activities remained mainly localized in the ER-rich fraction. The heavy membrane fraction retained highH+-ATPase and UDPG-SGT activities. However, the UDPG-SGT activity was significantly lower than that of control PM.When exogenous A5-sterols were added to the incubationmedium, the activity increased, indicating that the enzymeitselfwas not inactivated, but could not be fully active becauseof the absence of sufficient amounts of A5-sterols (see TableII) and of the poor ability of cyclopropyl sterols to serve assubstrates for the UDPG-SGT (32).
Lipid Analysis of Membrane Fractions from Control andFenpropimorph-treated Maize Roots
Fractions from Control Maize Roots
As shown in Table II, major sterols of the three membranefractions from control plants were the typical plant sterols:24-methyl cholesterol (2), sitosterol (3), and stigmasterol (4),with minute quantities of cholesterol (1) and isofucosterol(5). They accounted for at least 98% of total free sterols.
593
www.plantphysiol.orgon January 7, 2020 - Published by Downloaded from Copyright © 1989 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 90,1989
Table I. Enzymic Characterization of Membrane Fractions from Control and Fenpropimorph-TreatedMaize Roots
Membrane Fraction
Enzyme PM ER MIT
Ca Tb C T c T
NADH Cyt c reductasec- Antimycin A 0.2 0.16 2.2 1.9 0.34 0.37+ Antimycin A (1 ,uM) 0.13 0.12 2.2 1.9 0.14 0.15
AdoMet-CMTd 1.2 0.6 33.5 24 1.8 NDeCOld ND ND 470 ND 140 NDUDPG-SGTr-A5-sterols 3.1 1.1 1.1 0.13 0.19 0.06+ A,5-sterols 2.5 2.5 1.2 0.20 0.22 0.20
H+-ATPase (pH 6.5)' 280 320 ND ND ND NDa C, control plants.; b T, treated plants; c Results expressed as lsmol/min/mg protein. d Results
expressed as pmol/min/mg protein.; 6 Not determined.; 'Results expressed as nmoles/min/mg pro-tein.
Table II. Sterols of Membrane Fractions from Control and Fenpropimorph-Treated Maize Roots
Membrane Fraction
Sterol PM ER MIT
ca Tb C T C T
% of total sterols
4-DesmethylsterolsCholesterol (1) 2 1 1 1 2 124-Methyl cholesterol (2) 27 Trc 25 0 26 TrStigmasterol (3) 55 4 55 2 58 3Sitosterol (4) 14 1 15 Tr 12 Trlsofucosterol (5) Tr Tr 2 0 Tr Tr24-Methyl pollinastanol (6p 0 44 0 38 0 3824-Ethyl pollinastanol (8) 0 1 0 1 0 Tr
4a-Methyl sterolsObtusifoliol (9) Tr Tr Tr Tr Tr TrCycloeucalenol (10) 1 25 1 33 Tr 3124-Dihydrocycloeucalenol (1 1) 0 21 0 19 0 2231 -Norcyclobranol (12) 0 0 0 1 0 0Cyclofuntumienol (13) 0 1 0 2 0 2
4,4-Dimethyl sterolsCycloartenol (14) Tr 1 1 1 1 Tr24-Methylene cycloartanol (15) Tr 1 Tr 1 Tr 1B-Amyrine (16) Tr Tr Tr 1 Tr 1
A5-Sterols 99 5 98 2 98 59#,19-Cyclopropyl sterols 0.7 94 1 97 0.3 94Total sterols (Ag/mg protein) 60 85 30 95 20 30Sterols/PL molar ratio 0.45 0.62 0.06 0.21 0.12 0.18
a C, Control plants. b T, treated plants. C Tr = trace, amounts <0.5%. d 24-methyl pollinas-tanol + 5% 24-methylene pollinastanol (7).
Sterol precursors (4a-methyl- and 4,4-dimethyl sterols) werepresent only in trace amounts. The sterol profile was almostidentical in the three membrane systems and was similar tothat of the whole tissue. However, on a protein basis, the PM-rich fraction appeared to contain the highest amount of freesterols (60 ug/mg of protein).The three fractions contained the same classes of PL, with
PC and PE as major compounds (Table III). PA, PG, PS, andPI were also identified. The ER-rich fraction was found to beenriched in total PL (about 1 mmol/mg of protein), especially
in PC. Our data concerning the PL composition ofPM frommaize roots are in agreement with those reported by Grone-wald et al. (10). PL fatty acyl chains ofthe different membranefractions were also analyzed (Table IV). The two major com-pounds, linoleic acid (18:2) and palmitic acid (16:0), ac-counted for about 90% of total FA in all the fractions. Othercompounds were stearic acid (18:0), oleic acid (18:1), andlinolenic acid (18:3). The UI was found to be very similar inthe three fractions.Although the same classes of lipids are represented in the
594 GRANDMOUGIN ET AL.
www.plantphysiol.orgon January 7, 2020 - Published by Downloaded from Copyright © 1989 American Society of Plant Biologists. All rights reserved.
STEROLS AND PHOSPHOLIPIDS IN FENPROPIMORPH-TREATED MAIZE
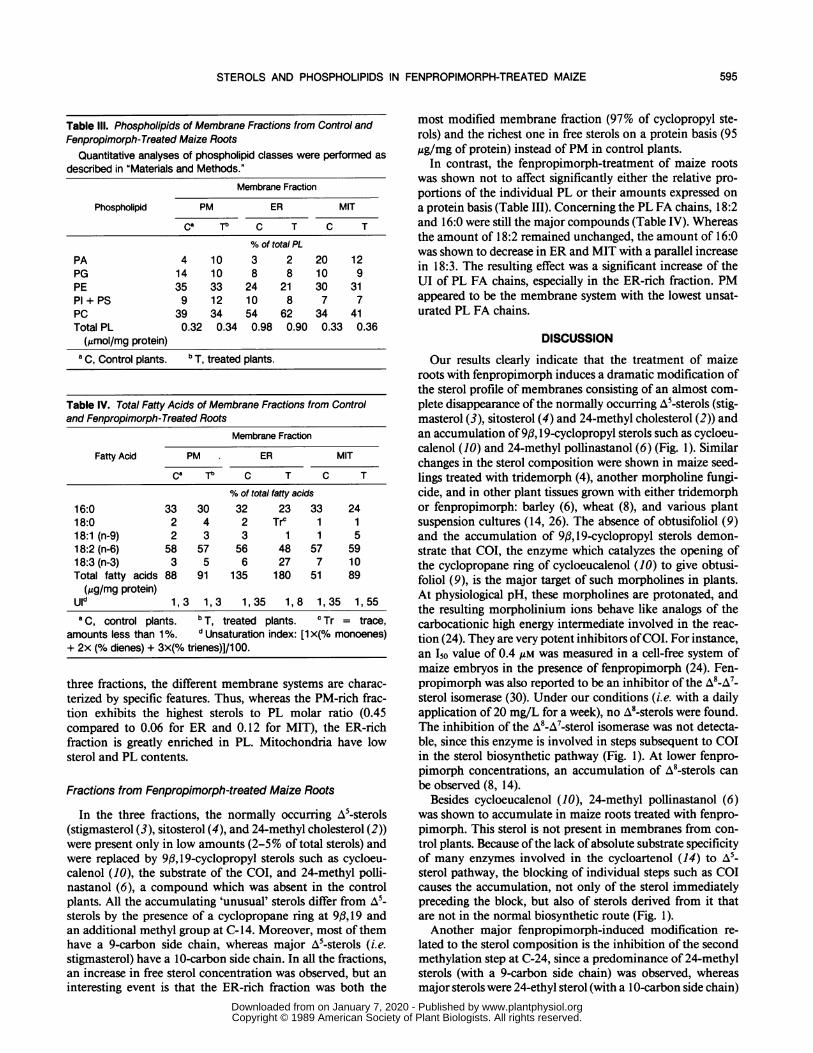
Table IlIl. Phospholipids of Membrane Fractions from Control andFenpropimorph-Treated Maize Roots
Quantitative analyses of phospholipid classes were performed asdescribed in "Materials and Methods."
Membrane Fraction
Phospholipid PM ER MIT
ca Tb C T C T
% of total PL
PA 4 10 3 2 20 12PG 14 10 8 8 10 9PE 35 33 24 21 30 31Pi + PS 9 12 10 8 7 7PC 39 34 54 62 34 41Total PL 0.32 0.34 0.98 0.90 0.33 0.36(gmol/mg protein)
a C, Control plants. b T, treated plants.
Table IV. Total Fatty Acids of Membrane Fractions from Controland Fenpropimorph-Treated Roots
Membrane Fraction
Fatty Acid PM ER MIT
Ca Tb C T c T
% of total fatty acids
16:0 33 30 32 23 33 2418:0 2 4 2 Trc 1 118:1 (n-9) 2 3 3 1 1 518:2 (n-6) 58 57 56 48 57 5918:3 (n-3) 3 5 6 27 7 10Total fatty acids 88 91 135 180 51 89(ug/mg protein)
Uld 1, 3 1, 3 1, 35 1, 8 1, 35 1, 55a C, control plants. b T, treated plants. c Tr = trace,
amounts less than 1%. d Unsaturation index: [1 x(% monoenes)+ 2x (% dienes) + 3x(% trienes)]/1 00.
three fractions, the different membrane systems are charac-terized by specific features. Thus, whereas the PM-rich frac-tion exhibits the highest sterols to PL molar ratio (0.45compared to 0.06 for ER and 0.12 for MIT), the ER-richfraction is greatly enriched in PL. Mitochondria have lowsterol and PL contents.
Fractions from Fenpropimorph-treated Maize Roots
In the three fractions, the normally occurring A5-sterols(stigmasterol (3), sitosterol (4), and 24-methyl cholesterol (2))were present only in low amounts (2-5% of total sterols) andwere replaced by 9#,,19-cyclopropyl sterols such as cycloeu-calenol (10), the substrate of the COI, and 24-methyl polli-nastanol (6), a compound which was absent in the controlplants. All the accumulating 'unusual' sterols differ from A5-
sterols by the presence of a cyclopropane ring at 93, 19 andan additional methyl group at C- 14. Moreover, most ofthemhave a 9-carbon side chain, whereas major A5-sterols (i.e.stigmasterol) have a 10-carbon side chain. In all the fractions,an increase in free sterol concentration was observed, but an
interesting event is that the ER-rich fraction was both the
most modified membrane fraction (97% of cyclopropyl ste-rols) and the richest one in free sterols on a protein basis (95,gg/mg of protein) instead ofPM in control plants.
In contrast, the fenpropimorph-treatment of maize rootswas shown not to affect significantly either the relative pro-portions of the individual PL or their amounts expressed ona protein basis (Table III). Concerning the PL FA chains, 18:2and 16:0 were still the major compounds (Table IV). Whereasthe amount of 18:2 remained unchanged, the amount of 16:0was shown to decrease in ER and MIT with a parallel increasein 18:3. The resulting effect was a significant increase of theUI of PL FA chains, especially in the ER-rich fraction. PMappeared to be the membrane system with the lowest unsat-urated PL FA chains.
DISCUSSION
Our results clearly indicate that the treatment of maizeroots with fenpropimorph induces a dramatic modification ofthe sterol profile of membranes consisting of an almost com-plete disappearance of the normally occurring A5-sterols (stig-masterol (3), sitosterol (4) and 24-methyl cholesterol (2)) andan accumulation of 9,B, 1 9-cyclopropyl sterols such as cycloeu-calenol (10) and 24-methyl pollinastanol (6) (Fig. 1). Similarchanges in the sterol composition were shown in maize seed-lings treated with tridemorph (4), another morpholine fungi-cide, and in other plant tissues grown with either tridemorphor fenpropimorph: barley (6), wheat (8), and various plantsuspension cultures (14, 26). The absence of obtusifoliol (9)and the accumulation of 9f3, 19-cyclopropyl sterols demon-strate that COI, the enzyme which catalyzes the opening ofthe cyclopropane ring of cycloeucalenol (10) to give obtusi-foliol (9), is the major target of such morpholines in plants.At physiological pH, these morpholines are protonated, andthe resulting morpholinium ions behave like analogs of thecarbocationic high energy intermediate involved in the reac-tion (24). They are very potent inhibitors ofCOI. For instance,an I50 value of 0.4 tLM was measured in a cell-free system ofmaize embryos in the presence of fenpropimorph (24). Fen-propimorph was also reported to be an inhibitor of the A8-A7sterol isomerase (30). Under our conditions (i.e. with a dailyapplication of 20 mg/L for a week), no .A'-sterols were found.The inhibition of the A8-A7-sterol isomerase was not detecta-ble, since this enzyme is involved in steps subsequent to COIin the sterol biosynthetic pathway (Fig. 1). At lower fenpro-pimorph concentrations, an accumulation of A8-sterols canbe observed (8, 14).
Besides cycloeucalenol (10), 24-methyl pollinastanol (6)was shown to accumulate in maize roots treated with fenpro-pimorph. This sterol is not present in membranes from con-trol plants. Because ofthe lack ofabsolute substrate specificityof many enzymes involved in the cycloartenol (14) to A5-sterol pathway, the blocking of individual steps such as COIcauses the accumulation, not only of the sterol immediatelypreceding the block, but also of sterols derived from it thatare not in the normal biosynthetic route (Fig. 1).Another major fenpropimorph-induced modification re-
lated to the sterol composition is the inhibition of the secondmethylation step at C-24, since a predominance of 24-methylsterols (with a 9-carbon side chain) was observed, whereasmajor sterols were 24-ethyl sterol (with a 10-carbon side chain)
595
www.plantphysiol.orgon January 7, 2020 - Published by Downloaded from Copyright © 1989 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 90, 1989
in control roots. This accumulation of such 24-methylatedcyclopropyl sterols induced by the fenpropimorph treatmentcorresponds to a situation different from that occurring insunflower suspension cultures grown in the presence of 25-epiminolanosterol, an inhibitor of the AdoMet-CMT (29). Inthis case, only 24-desalkylsterols accumulated, with cycloar-tenol as a major compound, and the increase in these sterolswas found to be correlated with a growth inhibition (29). Suchresults emphasize the functional significance of the alkylgroup at C-24.Our data clearly demonstrate that 9A, 1 9-cyclopropyl sterols,
which accumulate following treatment of maize roots withfenpropimorph, are actually incorporated into well character-ized membranes: ER, PM, and MIT. In membranes fromfenpropimorph-treated tissues, the remaining A5-sterols arepresent only in very low amounts (3-5% of total sterols). It ispossible that these trace levels of usual sterols play nonstruc-tural, specific roles and are involved in some metabolic func-tions that remain to be investigated (21). About half of theaccumulating cyclopropyl sterols are 4a-methyl sterols,whereas such sterols, which are precursors of A5-sterols, areminor compounds in control plants. Interestingly, the ER-rich fraction becomes the richest membrane fraction in freesterols instead ofPM in nontreated plants, indicating that thefungicide causes not only a qualitative change in the sterolcomposition of membranes but also a quantitative one. Per-haps, in fenpropimorph-treated maize root cells, the intracel-lular transfer of cyclopropyl sterols through the ER-GOLGI-PM pathway is less efficient than that of A5-sterols in controlcells.
Specific PL-sterol associations in biological membranes arevery common. Thus, it has been suggested that the PL ofmarine organisms (sponges) contain unique structural featurescomplementary to the side chain-alkylated sterols, leading tothe exclusion of cholesterol from liposomes made with suchPL (2). Relationships between sterol structure and PL-FAcomposition were also shown in yeast (7). In maize roots, thealmost complete replacement of A5-sterols by cyclopropylsterols, triggered by the fungicide, could induce an adaptationconsisting of changes in PL composition. The present datashow that no major modification either in the relative pro-portions or in the amounts of the individual PL of the threemembrane fractions was detected after the treatment of rootswith fenpropimorph. However, some changes in the relativeproportions of membrane PL-FA chains were observed. Inparticular, a significant increase of the unsaturation index ofFA of the ER-rich fraction was found probably as a responseto the very important accumulation of 9B,19-cyclopropylsterols in this fraction. The PL molecular species compositionof the different membranes was not investigated. But varia-tions in the intramolecular positioning of the FA chains onthe glycerol backbone can also have profound effects on thephysical and functional properties of membranes (27). Suchlipid changes were reported to occur in response to environ-mental stress (low temperature acclimation, for instance) (16).
Current models of PL(PC)-sterol interactions emphasizethe importance of some structural features for the sterolmolecule, such as the presence of a free 33-hydroxyl group, aquasi-plane tetracyclic ring, and an 8- to 10-carbon aliphatic
side chain (9). Whereas the first and the third requirementsare satisfied, the planeness of the tetracyclic nucleus of cyclo-propyl sterols can be questioned. Cycloartenol, a precursor ofA5-sterols, was previously believed to have a bent conforma-tion (rings: A-chair/B-boat/C-chair) (5). However, x-ray dif-fraction studies (1) as well as recent NMR experiments (19,22) indicate that such a molecule can also acquire a pseudo-planar (flat shape) conformation (A-chair/B-half-chair/C-1,3-diplanar). These studies also show that the conformationassumed by Bloch (5) does not occur in solution or in thecrystal state. According to Milon et al. (19), another con-former in which ring C is in the chair conformation wouldexist in solution; it would be more stable by 6.3 kJ/mol thanthe pseudoplanar one and would be in fast equilibrium withit, especially at low temperatures. Although until now no dataabout the conformation of cyclopropyl sterols embedded inmembranes were available, these results indicate that thecyclopropane ring of such compounds gives some flexibilityto these molecules at the level of the C, which may beimportant for their membrane properties. Finally, we haveshown recently that cyclopropyl sterols can form complexeswith filipin as well as plant A5-sterols do ( 18), indicating that,for these compounds, sterol structural requirements neededfor the interaction with PC are, at least partially, fulfilled.
In conclusion, our results indicate that 93, 19-cyclopropylsterols with a methyl group at C-24 can readily replace A5-sterols as plant membrane components and give an effectivepacking with the FA chains of PL. Despite a reduced growth,9,3, 19-cyclopropyl sterols seem to be well tolerated by maizeseedlings, which remain apparently healthy when they aregrown in the dark. The growth of Rubus fruticosus cell sus-pensions treated with fenpropimorph or N-(1,5,9-trimethyl-decyl)-4a, 10-dimethyl-8-aza-trans-decal-3(3-ol, another inhib-itor ofthe COI, is not significantly impaired, even after severalsubcultures (26). In certain plant tissues, cyclopropyl sterolsmay have biological functions of their own. Their presence inpollen is well known (31), but they were also detected in othermaterials such as cotton floral buds and anthers (17). Inter-estingly, most of these cyclopropyl sterols were found only assteryl esters (17, 31). The physiological functions of such'natural' cyclopropyl sterols remain to be investigated.
ACKNOWLEDGMENTS
We thank Dr. C. Leray (Centre de Neurochimie de Strasbourg) forproviding facilities for fatty acid analysis.
LITERATURE CITED
1. Allen FH, Trotter J (1971) Crystal and molecular structure of abis-p-bromobenzoate derivative of cyclograndisolide ([3a-methoxy-9, l9-cyclo-903-lanost-24-en-27,23(R)]-olide). J ChemSoc B 1079-1084
2. Ayanoglu E, Dizgunes N, Wijekoon WMD, Djerassi C (1986)Biophysical properties of unusual phospholipids and sterolsfrom marine invertebrates. Biochim Biophys Acta 863:110-114
3. Benveniste P, Bladocha M, Costet MF, Ehrhardt A (1984) Useofinhibitors of sterol biosynthesis to study plasmalemma struc-ture and function. In A Boudet, G Alibert, G Marigo, P Lea,eds, Journal of the Annual Proceedings of the PhytochemicalSociety of Europe. Oxford University Press, Oxford, pp283-300
596 GRANDMOUGIN ET AL.
www.plantphysiol.orgon January 7, 2020 - Published by Downloaded from Copyright © 1989 American Society of Plant Biologists. All rights reserved.

STEROLS AND PHOSPHOLIPIDS IN FENPROPIMORPH-TREATED MAIZE
4. Bladocha M, Benveniste P (1983) Manipulation by tridemorph,a systemic fungicide, of the sterol composition of maize leavesand roots. Plant Physiol 71: 756-762
5. Bloch KE (1983) Sterol structure and membrane function. CRCCrit Rev Biochem 14: 47-92
6. Burden RS, Clark T, Holloway PJ (1987) Effects of sterol bio-synthesis inhibiting fungicides and plant growth regulators onthe sterol composition of barley plants. Pest Biochem Physiol27: 289-300
7. Buttke TM, Jones SD, Bloch K (1980) Effect of sterol side chainson growth and membrane fatty acid composition of Saccha-romyces cerevisiae. J Bacteriol 144: 124-130
8. Costet-Corio MF, Benveniste P (1988) Sterol metabolism inwheat treated by N-substituted morpholines. Pest Sci 22:343-357
9. Demel RA, De Kruyff B (1976) The function of sterols inmembranes. Biochim Biophys Acta 457: 109-132
10. Gronewald JW, Abou-Khalil W, Weber EJ, Hanson JB (1982)Lipidic composition of a plasma membrane enriched fractionof maize roots. Phytochemistry 21: 859-862
11. Hartmann MA, Benveniste P (1978) Sterol biosynthetic capabil-ity of purified membrane fractions from maize coleoptiles.Phytochemistry 17: 1037-1042
12. Hartmann MA, Grandmougin A, Benveniste P (1985) Modifica-tion du profil sterolique de la membrane plasmique par lefenpropimorphe. C R Acad Sci Paris 301 (Ser III): 601-605
13. Hartmann MA, Benveniste P (1987) Plant membrane sterols:isolation, identification and biosynthesis. Methods Enzymol148: 632-650
14. Hosokawa G, Patterson GW, Lusby WR (1984) Effect of triari-mol, tridemorph and triparanol on sterol biosynthesis in carrot,tobacco and soybean suspension cultures. Lipids 19: 449-456
15. Kerkenaar A (1983) Mode of action of tridemorph and relatedcompounds. In J Miyamoto, KC Kearney, eds, PesticideChemistry: Human Welfare and the Environment, Vol 3.Pergamon Press, Oxford, pp 123-127
16. Lynch DV, Thompson GA (1984) Microsomal phospholipid mo-lecular species alterations during low temperature acclimationin Dunaliella. Plant Physiol 74: 193-197
17. Lusby WR, Oliver JE, McKibben GH, Thompson MJ (1987)Free and esterified sterols of cotton buds and anthers. Lipids22: 80-83
18. Milhaud J, Bolard J, Benveniste P, Hartmann MA (1988) Inter-action of the polyene antibiotic filipin with model and naturalmembranes containing plant sterols. Biochim Biophys Acta943: 315-325
19. Milon A, Nakatani Y, Kintzinger JP, Ourisson G (1989) The
conformation of cycloartenol investigated by NMR and mo-lecular mechanics. Helv Chim Acta 72: 1-13
20. Nes WR (1977) The biochemistry of plant sterols. Adv Lip Res15: 233-324
21. Nes WD, Heftmann E (1981) A comparison of triterpenoids withsterols as membrane components. J Nat Prod 44: 377-400
22. Nes WD, Benson M, Lundin RE, Le PH (1988) Conformationalanalysis of 9j#, 19-cyclopropyl sterols: detection of the pseudo-planar conformer by nuclear Overhauser effects and its func-tional implications. Proc Natl Acad Sci USA 85: 5759-5763
23. Quantin E, Hartmann MA, Schuber F, Benveniste P (1980)Latency of uridine diphosphate glucose-sterol-fl-D-glucosyl-transferase, a plasma membrane-bound enzyme of etiolatedmaize coleoptiles. Plant Sci Lett 17: 193-199
24. Rahier A, Schmitt P, Huss B, Benveniste P, Pommer EH (1986)Chemical structure-activity relationships of the inhibition ofsterol biosynthesis by N-substituted morpholines in higherplants. Pest Biochem Physiol 25: 112-124
25. Schacterle GR, Pollack RL (1973) A simplified method for thequantitative assay of small amounts of protein in biologicalmaterial. Anal Biochem 51: 654-655
26. Taton M, Benveniste P, Rahier A (1987) Comparative study ofthe inhibition of sterol biosynthesis in Rubusfruticosus suspen-sion cultures and Zea mays seedlings by N-(1,5,9-trimethyl-decyl)-4a, 10-dimethyl-8-aza-trans-decal-3fB-ol and derivatives.Phytochemistry 26: 385-392
27. Stubbs CD, Smith AD (1984) The modification of mammalianmembrane polyunsaturated fatty acid composition in relationto membrane fluidity and function. Biochim Biophys Acta779: 89-137
28. Sze H (1985) H+-translocating ATPases: advances using mem-brane vesicles. Annu Rev Plant Physiol 36: 175-208
29. Tal B, Nes WD (1987) Regulation of sterol biosynthesis: impor-tance ofthe C-24 alkyl group to growth ofsunflower suspensioncultures (abstract 969). Plant Physiol 83: S-161
30. Taton M, Benveniste P, Rahier A (1987) Mechanism of inhibi-tion of sterol biosynthesis enzymes by N-substituted morpho-lines. Pest Sci 21: 269-280
31. Thompson MJ, Dutky SR, Lehner Y, Standifier LN, HerbertEW (1978) 24-methylene pollinastanol and 24-dehydropolli-nastanol, two sterols of pollen. Phytochemistry 17: 1053-1056
32. Ullmann P, Rimmele D, Benveniste P, Bouvier-Nave P (1984)Phospholipid-dependence of plant UDP-glucose-sterol-j-D-glucosyl transferase. II. Acetone-mediated delipidation andkinetic studies. Plant Sci Lett 36: 29-36
33. Ullmann P, Bouvier-Nave P, Benveniste P (1987) Regulation byphospholipids and kinetic studies of plant membrane-boundUDP-glucose:sterol-,B-D-glucosyl transferase. Plant Physiol 85:51-55
597
www.plantphysiol.orgon January 7, 2020 - Published by Downloaded from Copyright © 1989 American Society of Plant Biologists. All rights reserved.