deletion mutagenesis identifies a haploinsufficient role · abundance and spatial organization of...
TRANSCRIPT

Deletion Mutagenesis Identifies a Haploinsufficient Rolefor g-Zein in opaque2 Endosperm Modification1[C][W][OPEN]
Lingling Yuan, Yongchao Dou, Shahryar F. Kianian, Chi Zhang, and David R. Holding*
Center for Plant Science Innovation, Beadle Center for Biotechnology (L.Y., Y.D., C.Z., D.R.H.), Department ofAgronomy and Horticulture (L.Y., D.R.H.), and School of Biological Sciences (Y.D., C.Z.), University ofNebraska, Lincoln, Nebraska 68588; and United States Department of Agriculture-Agricultural ResearchService Cereal Disease Laboratory, University of Minnesota, St. Paul, Minnesota 55108 (S.F.K.)
Quality Protein Maize (QPM) is a hard kernel variant of the high-lysine mutant opaque2. Using g-irradiation, we created opaqueQPM variants to identify opaque2 modifier genes and to investigate deletion mutagenesis combined with Illumina sequencing asa maize (Zea mays) functional genomics tool. A K0326Y QPM deletion mutant was null for the 27- and 50-kD g-zeins and abolishedvitreous endosperm formation. Illumina exon and RNA sequencing revealed a 1.2-megabase pair deletion encompassing the27- and 50-kD g-zein genes on chromosome 7 and a deletion of at least 232 kb on chromosome 9. Protein body number wasreduced by over 90%, while protein body size is similar to the wild type. Kernels hemizygous for the g-zein deletion hadintermediate 27- and 50-kD g-zein levels and were semivitreous, indicating haploinsufficiency of these gene products in opaque2endosperm modification. The g-zein deletion further increased lysine in QPM in its homozygous and hemizygous states. Thiswork identifies 27-kD g-zein as an opaque2 modifier gene within the largest QPM quantitative trait locus and may suggest the50-kD g-zein also contributes to this quantitative trait locus. It further demonstrates that genome-wide deletions in nonreferencemaize lines can be identified through a combination of assembly of Illumina reads against the B73 genome and integration ofRNA sequencing data.
Maize (Zea mays) opaque endosperm mutants havebeen studied because of the nutritional improvementthat they often show, as well as for the insight theyprovide into the kernel hardening, an essential qualitytrait for most of the grain’s uses. The most well knownopaque mutant, opaque2 (o2), has been widely studiedbecause of its increased Lys and Trp level (Mertz et al.,1964) associated with the reduced synthesis of a-zeins.TheO2 gene encodes a transcription factor that regulatesa-zein gene expression (Schmidt et al., 1990), althoughall zein proteins are indirectly reduced. O2 also regulatespyruvate orthophosphate dikinase (Maddaloni et al.,1996) and many other genes (Jia et al., 2013). Althoughthe soft kernels and yield penalty of o2 inhibited itscommercial success, subsequent breeding projects, in-cluding those in Mexico (Vasal et al., 1980) and in SouthAfrica (Geevers and Lake, 1992), led to the development
of hard kernel o2 varieties called Quality Protein Maize(QPM). QPM kernels generally have low levels of a-zeinsand high levels of Lys and Trp, but the genetic basis of o2endosperm modification is poorly understood.
One prominent feature of QPM endosperm is accu-mulation of the 27-kD g-zein at two- to three-fold higherlevels than in the wild type and o2 (Wallace et al., 1990;Geetha et al., 1991; Lopes and Larkins, 1991). Althoughthe genetic and biochemical mechanisms responsible forthis increase are unknown, the degree of endospermvitreousness in QPM correlates with the level of 27-kDg-zein protein (Lopes and Larkins, 1991). Furthermore,the 27-kD g-zein gene, along with the closely linked50-kD g-zein, maps to the most significant quantitativetrait loci (QTL) for endosperm modification located onchromosome 7 (Lopes and Larkins, 1995; Lopes et al.,1995; Holding et al., 2008, 2011). QPM endosperm ac-cumulates larger numbers of small, g-zein-rich proteinbodies. This is proposed to allow the formation of a rigid,glassy matrix similar in texture to mature wild-type en-dosperm. RNA interference (RNAi) reduction of both27- and 16-kD g-zeins in QPM caused opaque reversionand thus supported the suggestion that 27-kD g-zeincontributes to endospermmodification (Wu et al., 2010).However, the extent to which 27-kD g-zein is alonesufficient as a modifier is unknown. Furthermore, thepossible role of the 50-kD g-zein in modification hasnot been previously addressed. The molecular charac-terization of opaque mutants has shown that vitreousendosperm formation depends on more than just correctabundance and spatial organization of zein proteins
1 This work was supported by the University of Nebraska Depart-ment of Agronomy and Horticulture and the Center for Plant ScienceInnovation.
* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:David R. Holding ([email protected]).
[C] Some figures in this article are displayed in color online but inblack and white in the print edition.
[W] The online version of this article contains Web-only data.[OPEN] Articles can be viewed online without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.113.230961
Plant Physiology�, January 2014, Vol. 164, pp. 119–130, www.plantphysiol.org � 2013 American Society of Plant Biologists. All Rights Reserved. 119 www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from
Copyright © 2014 American Society of Plant Biologists. All rights reserved.

(Holding and Larkins, 2006; Holding et al., 2007, 2010;Myers et al., 2011; Wang et al., 2012). Furthermore,mapping studies indicate there are several QTLs forendosperm modification in QPM (Lopes and Larkins,1995; Lopes et al., 1995; Holding et al., 2008, 2011).
A variety of resources for forward and reverse geneticsexist in maize, and these are based on ethyl methanesul-fonate (EMS) mutagenesis and transposon insertions(Settles, 2005; Weil et al., 2005). EMS mutagenesis createsmostly point mutations that often result in knockdownalleles, which are useful in studying gene function.Transposons that create a high frequency of knockoutalleles have been used for a variety of mutagenesisschemes (McCarty et al., 2005; Settles et al., 2007;Vollbrecht et al., 2010) and have led to various reversegenetics resources. Though transposon insertions cancreate null alleles, these can be leaky mutations, leadingto weak phenotypes. Physical deletion of genes throughirradiation is one way to remove this ambiguity. Ra-diation has been used to induce physical deletions ofgenes, and this has been shown to be a viable systemfor PCR-based reverse genetic screens in Arabidopsis(Arabidopsis thaliana; Li et al., 2001). The chromosomalalterations in Arabidopsis included point mutations,translocations, insertions, small deletions, and largedeletions ranging in size from kilobases to severalmegabases. Deletion mutagenesis in maize has potentialas a tool for investigating a number of biological pro-cesses such as endosperm maturation and other as-pects of seed development. Even without saturationmutagenesis, an indexed collection of single andmultiplegene deletions would be useful in conjunction withexisting forward and reverse genetics resources.
The potential of g-radiation for reverse genetics hasbeen investigated (Wu et al., 2005; Sato et al., 2006), andalthough the mutagenesis rate is lower than with EMSmutagenesis, g-radiation results in a higher proportionof knockout mutations (Sato et al., 2006). The dosage ofradiation must be carefully optimized to maximize themutagenesis rate while allowing an acceptable level ofviability of the M1 plants. Low and moderate radiationdoses cause a relatively high proportion of usefulmutants with normal yield (Dumanovi et al., 1969),while high radiation doses induce a greater proportionof large deletions containing genes essential for via-bility (Naito et al., 2005). In an Arabidopsis irradiationexperiment, the majority of large deletions were nottransmitted from the M1 to the M2 generation andwere assumed to result from loss of genes required forseed germination, gamete development, and other as-pects of reproductive development (Naito et al., 2005).However, in large genomes with lower gene density,the tolerance for large deletion could be higher.
With modern sequencing technologies, radiation mu-tagenesis could become a valuable approach for studyinggene and genome function. To this end, a large fastneutron population was created for functional genomicsof agriculturally important traits in soybean (Glycinemax; Bolon et al., 2011). Despite studies highlighting thepotential of deletion mutagenesis for analysis of gene
function and creating desirable changes for breedingprograms, a radiation mutagenesis approach is limitedby lack of knowledge of the extent and size range ofdeletions induced. We used g-irradiation of K0326YQPM as a means to identify genes corresponding to o2modifiers and genes generally required for vitreousendosperm formation. For identification of DNA se-quence deletions in the induced mutants, we combinedexon-capture DNA sequencing and RNA sequencing(RNA-seq). Among the opaque variants induced inQPM, one had a deletion that eliminates both the 27- and50-kD g-zein loci, which are within a significant QPMQTL on chromosome 7 (Holding et al., 2008, 2011). Bydemonstrating the dosage-dependent action of 27-kDg-zein in o2 endosperm modification, this mutant high-lights the potential of this approach for identifying othero2 modifier genes as well as genes involved in a varietyof aspects of endosperm development.
RESULTS
Identification of a 27- and 50-kD g-Zein Protein Nullamong Novel Opaque Deletion Mutants
We recovered 305M2 ears of variable seed fill, of which293 were successfully propagated to M3 families (ap-proximately 20 ears each). Although single M2 ears werepotentially affected by nongenetic environmental factors,screens of adequately filled M2 ears were conducted fordominant seed mutant phenotypes and identified morethan 30 lines having a substantial proportion of opaque,small, defective, empty pericarp, or rough kernels. All293 lines were propagated to the M3 generation, and27 lines exhibited clear segregation for opaque mutants.Of these 27 lines, opaque kernels in 17 lines were notviable and presumed to be pleiotropic mutations. Theother 10 lines had fully viable opaque kernels and werethus candidates for factors specifically involved in o2endosperm modification and/or vitreous endospermformation in general. Three of these 10 confirmed mu-tants, including line 107 (Fig. 1), line 198 (SupplementalFig. S1), and line 12 (not shown) exhibited mutantkernels in the M2 ears, while the others were first ob-served in the M3 and resulted from M2 ears that eitherappeared normal or had too few kernels to screen.Segregating M3 ears for examples of these lines is shownin Supplemental Figure S1. Analysis of zein proteinsrevealed several different phenotypic classes affectingzein composition (Supplemental Fig. S1). These includedno apparent change in zeins (line 8), generalized re-duction of all zeins (lines 66 and 112), loss of residual22-kD a-zein present in K0326Y QPM (lines 44 and 133),and loss of 19-kD a-zein (line 198). Line 107 exhibited astriking absence of the 27- and 50-kD g-zeins (Fig. 2A),which was confirmed with immunoblot analysis (Fig. 2B).Given the suggested role of 27-kD g-zein in endospermmodification in QPM, we focused on the characterizationof this mutant. As expected, 27-kD g-zein was significantlyincreased in K0326Y QPM compared with the W64A wildtype (Fig. 2A). Interestingly, although 50-kD g-zein is not
120 Plant Physiol. Vol. 164, 2014
Yuan et al.
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.
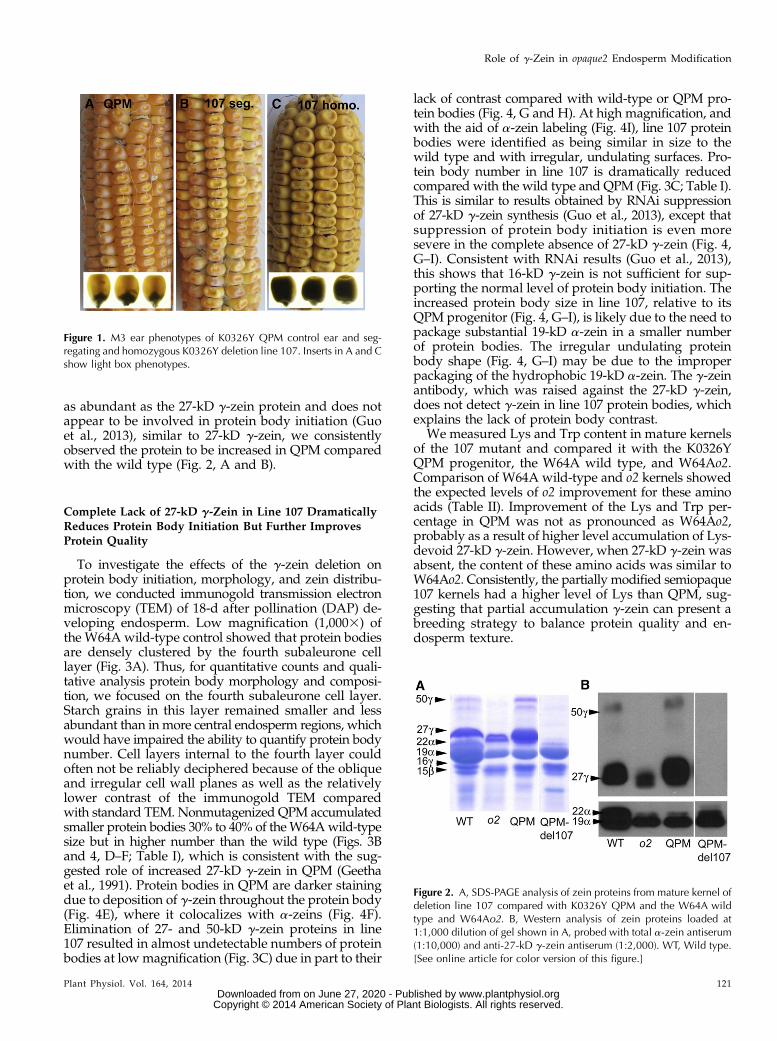
as abundant as the 27-kD g-zein protein and does notappear to be involved in protein body initiation (Guoet al., 2013), similar to 27-kD g-zein, we consistentlyobserved the protein to be increased in QPM comparedwith the wild type (Fig. 2, A and B).
Complete Lack of 27-kD g-Zein in Line 107 DramaticallyReduces Protein Body Initiation But Further ImprovesProtein Quality
To investigate the effects of the g-zein deletion onprotein body initiation, morphology, and zein distribu-tion, we conducted immunogold transmission electronmicroscopy (TEM) of 18-d after pollination (DAP) de-veloping endosperm. Low magnification (1,0003) ofthe W64A wild-type control showed that protein bodiesare densely clustered by the fourth subaleurone celllayer (Fig. 3A). Thus, for quantitative counts and quali-tative analysis protein body morphology and composi-tion, we focused on the fourth subaleurone cell layer.Starch grains in this layer remained smaller and lessabundant than in more central endosperm regions, whichwould have impaired the ability to quantify protein bodynumber. Cell layers internal to the fourth layer couldoften not be reliably deciphered because of the obliqueand irregular cell wall planes as well as the relativelylower contrast of the immunogold TEM comparedwith standard TEM. Nonmutagenized QPM accumulatedsmaller protein bodies 30% to 40% of theW64Awild-typesize but in higher number than the wild type (Figs. 3Band 4, D–F; Table I), which is consistent with the sug-gested role of increased 27-kD g-zein in QPM (Geethaet al., 1991). Protein bodies in QPM are darker stainingdue to deposition of g-zein throughout the protein body(Fig. 4E), where it colocalizes with a-zeins (Fig. 4F).Elimination of 27- and 50-kD g-zein proteins in line107 resulted in almost undetectable numbers of proteinbodies at lowmagnification (Fig. 3C) due in part to their
lack of contrast compared with wild-type or QPM pro-tein bodies (Fig. 4, G and H). At high magnification, andwith the aid of a-zein labeling (Fig. 4I), line 107 proteinbodies were identified as being similar in size to thewild type and with irregular, undulating surfaces. Pro-tein body number in line 107 is dramatically reducedcompared with the wild type and QPM (Fig. 3C; Table I).This is similar to results obtained by RNAi suppressionof 27-kD g-zein synthesis (Guo et al., 2013), except thatsuppression of protein body initiation is even moresevere in the complete absence of 27-kD g-zein (Fig. 4,G–I). Consistent with RNAi results (Guo et al., 2013),this shows that 16-kD g-zein is not sufficient for sup-porting the normal level of protein body initiation. Theincreased protein body size in line 107, relative to itsQPM progenitor (Fig. 4, G–I), is likely due to the need topackage substantial 19-kD a-zein in a smaller numberof protein bodies. The irregular undulating proteinbody shape (Fig. 4, G–I) may be due to the improperpackaging of the hydrophobic 19-kD a-zein. The g-zeinantibody, which was raised against the 27-kD g-zein,does not detect g-zein in line 107 protein bodies, whichexplains the lack of protein body contrast.
We measured Lys and Trp content in mature kernelsof the 107 mutant and compared it with the K0326YQPM progenitor, the W64A wild type, and W64Ao2.Comparison of W64A wild-type and o2 kernels showedthe expected levels of o2 improvement for these aminoacids (Table II). Improvement of the Lys and Trp per-centage in QPM was not as pronounced as W64Ao2,probably as a result of higher level accumulation of Lys-devoid 27-kD g-zein. However, when 27-kD g-zein wasabsent, the content of these amino acids was similar toW64Ao2. Consistently, the partially modified semiopaque107 kernels had a higher level of Lys than QPM, sug-gesting that partial accumulation g-zein can present abreeding strategy to balance protein quality and en-dosperm texture.
Figure 1. M3 ear phenotypes of K0326Y QPM control ear and seg-regating and homozygous K0326Y deletion line 107. Inserts in A and Cshow light box phenotypes.
Figure 2. A, SDS-PAGE analysis of zein proteins from mature kernel ofdeletion line 107 compared with K0326Y QPM and the W64A wildtype and W64Ao2. B, Western analysis of zein proteins loaded at1:1,000 dilution of gel shown in A, probed with total a-zein antiserum(1:10,000) and anti-27-kD g-zein antiserum (1:2,000). WT, Wild type.[See online article for color version of this figure.]
Plant Physiol. Vol. 164, 2014 121
Role of g-Zein in opaque2 Endosperm Modification
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Line 107 Shows Absence of 27- and 50-kD g-Zein Genesand Haploinsufficiency for o2 Endosperm Modification
We sought to determine if the absence of 27- and 50-kDg-zein proteins resulted from an indirect effect on zeinsynthesis in general, a regulatory defect, or the physicalloss of the genes. Reverse transcription (RT)-PCR showedthat transcripts of both genes were undetectable inendosperm RNA of line 107 (Fig. 5A). Levels of theunlinked 16-kD g-zein were normal, ruling out the pos-sibility that zein gene expression was generally affected.Genomic PCR did not detect the 27- or 50-kD zein genes,
while the 16-kD g-zein gene was present (Fig. 5B),showing that the protein absence was not a regulatoryeffect but due rather to the loss of these genes. Giventhat the 27- and 50-kD g-zein genes are closely linked(being separated by approximately 27 kb; Holding et al.,2008), the most likely scenario was that a deletion coveredboth genes, which was confirmed by sequencing exon-captured genomic DNA (below).
While most of the heritable opaque mutants we re-covered appeared to segregate according to Mendelianratios in the M3 (Supplemental Fig. S1), line 107 hadmore than 25% opaque kernels and obvious variability
Figure 3. Low-magnification TEM images fromimmunogold labeling showing distribution of pro-tein bodies in representative 18-DAP endosperm ofthe wild type, K0326Y QPM, and QPM deletionline 107. Subaleurone cell layers are labeled with2nd, 3rd, and 4th. Cell walls are labeled withcw., starch grains are labeled with S, and selectedprotein bodies are labeled with vertical arrows.Depicted protein bodies in C show connections tothe endoplasmic reticulum. Bar = 5 mm. WT, Wildtype.
Figure 4. Immunogold TEM analysis showinga- and g-zein distribution in protein bodies of fourthsubaleurone cell layer of 18-DAP endosperm of thewild type, K0326Y QPM, and QPM deletion line107. Bar = 1 mm. WT, Wild type.
122 Plant Physiol. Vol. 164, 2014
Yuan et al.
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

in the level of opacity (Fig. 1B). With careful examina-tion on a light box, the number of fully opaque kernels(Fig. 6) was found to be approximately 25% (46/190,9/38, and 14/56 in three M3 segregating ears). Kernelsin this class were homozygous for the deletion mutationbecause they gave rise to 100% opaque kernels whenpropagated (Fig. 1C). Another class of kernels, makingup less than 50%, was opaque with a vitreous cap (Fig. 6).These were determined to be hemizygous for the g-zeindeletion, because they gave rise to segregating ears whenpropagated (Fig. 1B). The remainder of the expected 50%hemizygous kernels from these ears comprised lesssevere semiopaques, which could not be reliably scored.SDS-PAGE analysis of zein proteins from vitreous,semiopaque, and fully opaque kernels from a segregatingear showed that partial opacity results from reducedaccumulation of the 27- and 50-kD g-zein proteins (Fig. 6).This suggests that expression from a single allele ofthese two genes is haploinsufficient for full o2 endo-sperm modification. Importantly, unlike the majorityof mutants that first segregated in the M3 ears, the M2ear for line 107 segregated 1:1 for normal and opaquewith vitreous cap (hemizygous) kernels, which supportsthe haploinsufficiency explanation and is consistentwith the ontology of the mature embryonic shoot apicalmeristem (Johri and Coe, 1983; Poethig et al., 1986); see“Discussion”).
Exon- and RNA-Seq Identifies a Deletion Spanning the27- and 50-kD g-Zein Genes
To confirm that the 27- and 50-kD g-zeins were withina single deletion and define the size of that deletion,we used Illumina sequencing of exon-enriched genomicDNA. Deletions were called according to an algorithmthat considered both the number of reads for a givenexon in the nonmutagenized QPM and the ratio of readsbetween nonmutagenized QPM and a mutagenized line(see “Materials andMethods”). The relatively nonstringent
criteria returned deletion candidates on all chromo-somes apart from chromosome 10 (Supplemental TableS1). We tested all of the candidates using genomic PCRand RT-PCR (data not shown) and established that allexcept those genes within the chromosome 7 and 9deletions described below were false positives. As furtherconfirmation, none of the genes identified as false posi-tives were identified as differentially expressed usingRNA-seq (Supplemental Tables S1 and S2).
By combining exon read data with genomic and RT-PCR, we identified two deletions in line 107. One ofthese is in chromosome 7, Bin 7.02, and is approximately1,236,000 bp (Fig. 7). Using genomic PCR on exon de-letion candidates, we showed that there are at least 11predicted genes in this region that are absent in line 107.RT-PCR revealed that at least seven of these genes areexpressed in normal developing endosperm but notdetectable in RNA of line 107 (Fig. 7). Two genes closeto the 59 end of the deletion encode the 27- and 50-kDg-zeins. Four of the 11 genes are either endospermnonexpressed or expressed at a low level or are pseu-dogenes. Four other genes were confirmed as deleted inthis region by comparing exon sequencing (exon-seq)and RNA-seq reads (Supplemental Table S2). We usedprimers within flanking genes (Fig. 7) as well as inter-genic primers to estimate the extremities of this dele-tion. A smaller deletion of at least 231,968 bp wasidentified on chromosome 9, Bin 9.03 (Fig. 8). This re-gion contains five genes, four of which have expressionabolished based on RT-PCR. Expression of the fifthgene was not detectable in the nonmutagenized control.Genomic and RT-PCR tests of other exon deletion can-didates (Supplemental Table S1) identified elsewherein the genome by low or absent read number did notconfirm additional single or multiple exon deletions.
The RNA-seq comparison of K0326Y QPM and line107 had three purposes: confirmation of deletions iden-tified with exon-seq; identification of additional deletionsthat exon-seq may have missed; and potential identifi-cation of indirect transcriptional affects of putative genedeletions, as a consequence of defective transcriptionfactors. For this, we mixed RNA in equal proportionsextracted from whole developing seeds (endospermand embryo), juvenile leaf, and juvenile root. Thispotentially allowed us to identify transcript alterationsresulting in sporophytic tissues as well as seed tissues.Because our goal was to identify complete losses orsubstantial changes in expression, rather than minorfold changes, a single RNA-seq library was preparedfrom K0326Y QPM and line 107, and this allowed barcoding and pooling with other deletion mutants. Thegenes with transcripts that were decreased or increased
Table I. Protein body number in fourth subaleurone layer of the wildtype, K0326Y QPM, and K0326Y line 107
Protein bodies per 100-mm square of fourth subaleurone cell layerwere counted from two independent kernels and two independentembedments of each kernel. P values show that counts are significantlydifferent from the wild type.
W64A Wild Type K0326Y QPM K0326Y Line 107
Protein body no. 50.0 87.6 6.2SD 63.0 68.4 60.8P value — 1.6 3 10–5 4.4 3 10–9
Table II. Lys and Trp content in mature kernels
Mature kernel amino acid contents for Lys and Trp are shown in grams per 100 g protein 6 SD (n = 3).
W64A Wild Type W64A o2 K0326Y QPM K0326Y Line 107 Opaque K0326Y 107 Semiopaque
Lys 2.39 6 0.08 4.96 6 0.04 4.09 6 0.00 4.93 6 0.17 4.42 6 0.01Trp 0.61 6 0.05 1.00 6 0.00 0.92 6 0.00 0.97 6 0.13 0.92 6 0.05
Plant Physiol. Vol. 164, 2014 123
Role of g-Zein in opaque2 Endosperm Modification
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

in line 107 based on RNA-seq, along with their exon-seqreads, are shown in Supplemental Table S2. Twenty geneswere increased in abundance more than 4-fold in line 107compared with K0326Y QPM, whereas 70 genes hadtranscripts more than 4-fold reduced. Within these 70were genes shown to be within the chromosome 7 and 9deletions (Supplemental Tables S3 and S4). Of the fivegenes within the chromosome 9 deletion, only one gene,GRMZM2G099648, a ubiquitin ligase, had RNA-seqreads (Supplemental Table S4). Of the other differen-tially expressed genes with chromosome coordinatesnot within the chromosome 7 and 9 deletions, nonequalified as candidate deletions. However, severaldifferentially expressed genes identified with RNA-seqwere not detected with exon-seq in either K0326Y orline 107 (Supplemental Table S2, pink highlights). Wetested all of these genes with at least two pairs of ge-nomic PCR primers each and showed that they werenot gene deletions missed during exon-seq (not shown).
Zein genes are expressed at high levels, and this wasreflected in the reads measured for the 27- and 50-kDg-zein genes in K0326Y QPM (386,441 and 180,567reads, respectively). As expected, reads for these genesin line 107 were drastically reduced to 0.2% and 0.1%of control, respectively, but surprisingly, they were notcompletely absent as might be expected for deletion ofsingle copy genes (869 and 284 reads, respectively).These genes share extensive identity with the 16-kDg-zein, but at a lower level than our allowable thresholdfor RNA-seq reads (98-bp perfect match of the 100-bpread length). However, some nucleotides within theg-zein reads had suboptimal scores, and if some hadbeen incorrectly called for 16-kD g-zein reads, thiscould give rise to reads incorrectly annotated as 50- or27-kD reads. Alignment of K0326Y sequences to theB73 reference genome is also a possible source of errors.In addition, because K0326Y and line 107 were se-quenced in the same pool in the same flow cell lane,these reads could, in theory, be the result of very low-level incorrect parsing of bar code sequences.
Three a-zein genes were around six- to 16-fold down-regulated in line 107 (Supplemental Table S2). One ofthese is annotated as a Z1A (19-kD) family member andtwo are Z1C (22-kD) family members. Z1A genes arenot regulated by O2, and the Z1C genes identified here
are also likely O2 independent. The basis for thisconclusion is that their expression in K0326Y (modifiedo2/o2) is still substantial (Supplemental Table S2), andthe O2 transcript in both K0326Y and line 107 wasundetectable by RT-PCR and quantitative RT-PCR(data not shown). Consequently, these a-zeins may berepressed through an unknown feedback mechanismthat results from reduced levels of g-zein synthesis.Such a mechanism was also proposed as a result of therecent observations of indirect suppression of a-zeingene expression caused by various g-zein RNAi con-structs (Guo et al., 2013).
DISCUSSION
The 27-kD g-Zein Acts in a Dosage-Dependent Manner toModify o2 Endosperm
The feasibility of radiation mutagenesis to identifyo2 modifier genes was demonstrated because one lineof a population of 293 M2 families had a deletion of asuspected modifier gene, the 27-kD g-zein as well thenearby 50-kD g-zein, and caused complete opaquereversion in QPM. Null mutant alleles have not beenpreviously reported for either of these genes, althoughknockdown lines have been generated with RNAi byourselves and others (Wu and Messing, 2010; Guoet al., 2013). RNAi studies have allowed dissection ofthe overlapping and distinct roles of the differentg-zeins, but their interpretation is not simple becausethe transgenic lines are not null. However, their phe-notypes have suggested that the 27-kD g-zein has aspecialized role in protein body initiation (Guo et al.,2013). Crossing an RNAi event that reduces both the27- and 16-kD g-zeins to K0326Y QPM caused completeopaque kernel reversion and thus is consistent withabundant previous data suggesting that 27-kD g-zeinplays a key role in endosperm modification (Geethaet al., 1991; Lopes and Larkins, 1991, 1995; Lopes et al.,1995; Wu et al., 2010).
Figure 6. SDS-PAGE and kernel opacity analysis showing hap-loinsufficiency of g-zein for endosperm modification. Selected kernelsfrom a segregating ear are shown. [See online article for color versionof this figure.]
Figure 5. A, RT-PCR showing g-zein transcript accumulation. B, Ge-nomic PCR analysis of g-zein genes.
124 Plant Physiol. Vol. 164, 2014
Yuan et al.
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

The 27- and 50-kD g-zein null line reported here pro-vides more definitive proof of essentiality of the 27-kDg-zein for QPM development. It also invokes the pos-sibility that the 50-kD g-zein is involved in endospermmodification. Despite the relatively lower abundance ofthe 50-kD g-zein, it is readily detectable in wild-typeendosperm and becomes almost undetectable in o2endosperm (Fig. 2). Furthermore, during endospermmodification in QPM, this protein is not only restoredbut is increased relative to the wild type. Like that ofincreased 27-kD g-zein transcript and protein (Holdinget al., 2011), the mechanism causing this increase isunknown. It is possible that pre- or posttranscriptionalregulation could affect these genes independently or
that they could be coregulated. In either case, thesegenes are so closely linked that it is not currently possibleto separate their contributions to the QTL on chromo-some 7.02.
The 27-kD g-zein deletion also extends our knowledgeof its role in protein body initiation. While protein bodynumber is reduced to about 40% of the wild-type numberby 27-kD g-zein RNAi (Guo et al., 2013), the knockoutmutant reduced the protein body number to about 12%of the wild-type and about 7% of the QPM protein bodynumber. Vitreous endosperm formation in modified o2 isthought to be driven, at least in part, by the initiation of alarger number of protein bodies as a result of increased27-kD g-zein accumulation (Dannenhoffer et al., 1995;
Figure 7. Exon-seq and genomic and RT-PCR verification of deletion on chromosome 7.02. Illumina reads column and PCRcolumns show nonmutagenized K0326Y versus line 107. Most gene identifications show absence of Illumina reads in morethan one exon (denoted by multiple sets of coordinates). GRMZM2G117230 and 429842 are the 59 and 39 flanking intactgenes. Genes with an asterisk are either low-abundance transcripts, endosperm nonexpressed genes, or pseudogenes.
Plant Physiol. Vol. 164, 2014 125
Role of g-Zein in opaque2 Endosperm Modification
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Figure 8. Exon-seq and genomic and RT-PCR verification of deletion on chromosome 9.03. Illumina reads column and PCRcolumns show nonmutagenized K0326Y versus line 107. Most gene identifications show absence of Illumina reads in morethan one exon (denoted by multiple sets of coordinates). GRMZM2G398628 and 435380 are the two nondeleted genes flankingthe 59 end of the deletion, and GRMZM2G499214 and 108302 are the two nearest nondeleted genes flanking the 39 end of thedeletion. Genes with an asterisk are either low-abundance transcripts, endosperm nonexpressed genes, or pseudogenes.
126 Plant Physiol. Vol. 164, 2014
Yuan et al.
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Gibbon and Larkins, 2005). These protein bodies remainsmall, with g-zein located throughout (Fig. 4), and havean even round shape due to their appropriate packag-ing of residual a-zein (Fig. 4). However, in the absenceof the 27- and 50-kD g-zeins, the protein body size issimilar to the wild type, with highly irregular shape.The size increase likely results from the packaging ofresidual a-zein in less than 10% of the number of pro-tein bodies (Fig. 4), and the distortions likely result fromhydrophobic a-zeins, especially the abundant 19-kDa-zeins, not being correctly packaged by the remainingg-zeins. Protein body shape distortions also result fromdominant negative a- and g-zein mutants, which in-terfere with correct zein trafficking into the lumen of theendoplasmic reticulum (Coleman et al., 1997; Kim et al.,2004, 2006). Similarly, low-level accumulation of anunprocessed a-kafirin protein causes protein body re-ticulation in the high digestibility high lysine mutant ofSorghum bicolor (Wu et al., 2013).Unlike the highly duplicated a-zein genes, whose
expression collectively results in massive levels ofa-zein protein, the g-zeins are encoded by single copygenes and, in the case of 27-kD g-zein, protein levelsapproach those of a-zeins. What drives this high-levelexpression as well as it’s even greater expression inQPM is not known. The degree to which g-zein proteinlevels are limited by pre- and posttranscriptional factorsis also not known. Our results show that one copy of the27-kD g-zein gene is not sufficient for full modificationbecause hemizygous seeds accumulate intermediateamounts of the protein and are only partially modified.This haploinsufficiency suggests that maximal tran-scription from both 27-kD g-zein gene copies is requiredto create a fully vitreous endosperm. However, thesynthesis of 27-kD g-zein is also likely limited by aminoacid availability and translational capacity because27-kD g-zein overexpression constructs do not furtherincrease the protein (D. Holding, unpublished data).The g-zein deletion is currently being introgressed intothe B73 wild type to separate it from the effects of theo2 mutation and the modifier genes. This will provideinsight into whether this same critical threshold forg-zein abundance also exists for normal vitreous en-dosperm formation as it does for endosperm modifi-cation in QPM.The increased Lys and Trp content of the QPM
g-zein deletion emphasizes one of the drawbacks ofQPM in relation to unmodified o2. Because 27-kDg-zein is devoid of Lys, its increased level in QPMreduces the nutritional quality compared with starchyunmodified o2. Our results show that this particularQPM inbred has Lys and Trp levels intermediate be-tween W64Ao2 and the wild type. However, hemizy-gous g-zein deletion kernels had similar amounts ofthese amino acids to unmodified o2 while exhibitinggreater levels of vitreousness. This may suggest thatselection of lines with partially accumulating g-zeinmay be a novel strategy for breeding maize lines withboth increased Lys and an acceptably vitreous kerneltexture.
The g-Zein Deletion Line Demonstrates the Potential forIdentifying Other o2 Modifiers as Well as for GeneralMaize Seed Functional Genomics
We used g-irradiation mutagenesis to study genesaffecting vitreous endosperm formation (modification)in the hard kernel o2 variant K0236Y QPM. In additionto o2 modifier genes, this approach potentially allowsfunctional genomic dissection of genes more generallyinvolved in endosperm formation as well as seed andplant development. In a population of 293 M3 families,we identified 27 mutants segregating for opaque kernelphenotypes. Although M2 screens did identify at leastthree dominant opaque kernel mutants, which were truebreeding from the M2 to the M3 and M4, the majority ofopaque mutants were first observed segregating in theM3 ears. The likely reason for this is that differentpopulations of cells within the mature maize embryoshoot apical meristem give rise to the ear and tassel(Johri and Coe, 1983; Poethig et al., 1986). A hemizy-gous DNA deletion in an ear progenitor cell within theshoot apical meristem of a mature embryo will ulti-mately result in a mixture of hemizygous and wild-typekernels in an M2 ear because the tassel progenitor cellsgiving rise to the pollen would not have the mutation.Thus, only dominant mutations would be visible in theM2 ears. The number of tassel and ear progenitor cellsin the meristem was given as four (61) for the tasseland one to three cells for the ear (Johri and Coe, 1983).These numbers would be expected to produce variablephenotypic ratios for dominant mutants in M2 ears.Line 107 and line 12 (not shown) had apparent 1:1 ratiosof normal to mutant seeds (19:19 and 164:174, respec-tively) and may suggest that the mutation arose in onemeristem initial cell that gave rise to the whole ear. Theother dominant mutant (line 198) had a lower ratio ofM2mutant kernels (230:52), perhaps suggesting that themutation arose in one of several ear or tassel initial cells.
Using mutants that had viable kernels taken fromsegregating M3 ears, and gave rise to 100% mutant M4ears, we were able to assume homozygosity and directlyassay for gene loss using Illumina sequencing of genomicDNA. To simplify the sequencing template, we enrichedfor the exon fraction, using exon capture hybridization toa Nimblegen B73 maize exome array. This simplificationof sequencing template enabled pooling of five mutantsand sequencing in a single Genome Analyzer II flow celllane. Despite this suboptimal Illumina platform (com-pared with the Hi-Seq 2000 platform) and assemblingK0326Y QPM sequences against the nonisogenic B73genome, we were able to call and test deletions rangingfrom single to multiple genes. RNA-seq allowed verifi-cation of the deleted genes and identified genes withexpression that was indirectly affected by DNA deletions.
Prospects and Limitations for Maize Seed FunctionalGenomics Using Radiation Mutagenesis
The data presented here strengthen genetic, biochemi-cal, and transgenic evidence suggesting that the 27-kD
Plant Physiol. Vol. 164, 2014 127
Role of g-Zein in opaque2 Endosperm Modification
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

g-zein, possibly acting with the 50-kD g-zein, is an o2modifier gene. As with transposon or EMS mutagenesis,in which multiple genes are mutated, deletion muta-genesis requires reverse genetics to formally link muta-tions to phenotype. Typically, this involves isolation ofindependent mutant alleles in the gene suspected ofcausing the phenotype or complementation or knock-out using transgenic constructs. The knockout mutantsof the 27- and 50-kD g-zeins described herein are theonly known mutants in these genes. However, weshowed the essentiality of 27-kD g-zein in modificationof K0326Y QPM endosperm using RNAi (Wu et al.,2010). Furthermore, RNAi lines that dramatically re-duce the level of 27-kD g-zein support the role of thisprotein in vitreous endosperm formation in a normalvitreous endosperm formation (Wu and Messing, 2010;Guo et al., 2013).
In addition to identifying genes involved in vitreousendosperm formation, characterizing more opaque mu-tants in this QPM population will allow appraisal of theheterogeneity and frequency of radiation-induced de-letions and their utility for maize functional genomicswhen combined with current reverse genetics resourcesand ever-improving sequencing capabilities. In additionto large deletions, radiation can also cause subgenescale mutations (Naito et al., 2005), but we could notdetect these without a K0326Y QPM reference genome.A second, much larger deletion population has beencreated that has 1,784 mutagenized families in the B73reference genetic background. Many apparent viableopaque and small kernel seed mutants are currentlybeing tested for heritability. Because of the isogenicityof these mutants with the B73 reference genome, wewill be able to detect subgene mutations in this pop-ulation, and exon-seq and RNA-seq reads can be as-sembled and compared for identification and appraisalof the power of deletion mutagenesis for creating andutilizing mutations ranging from a few base pairs tomega base pairs.
MATERIALS AND METHODS
Radiation Dosage Testing and g-Radiation Mutagenesis
K0326Y QPM seed were equilibrated to 13% moisture as previously de-scribed (Hossain et al., 2004) and treated in batches of approximately 50 seedswith various dosages of g-radiation (15–35 Gray [Gy]). After irradiation, theseeds were immediately planted in the greenhouse in potting soil. Plant survivalwas determined as the proportion of seedlings compared with nonirradiatedcontrol seeds 1 month after sowing. Radiation dosage was set at 25 Gy based ona 75% survival rate and was used for full-scale mutagenesis of 1,909 seeds. Theseseeds were planted in the field at the University of Nebraska East Campus farmin summer 2010.
A population of 1,909 K0326Y QPM seeds, treated with 25 Gy of g-radiation,were propagated in the field in 2010 and 1,108 germinated (58%). The radiationdosage was previously optimized to produce 75% survival rate under controlledgreenhouse condition using a small-scale (approximately 50 seeds per dosage)experiment.
Screening of M2 and Heritability Testing in M3
Adequately filledM2 ears (more than approximately 30 kernels) that did nothave fungal contamination were screened for dominant kernel phenotypes
before shelling and after shelling on light box. Zein profiles were obtained fromputative opaque mutant kernels to prioritize lines for more urgent greenhouseheritability testing. Twenty kernels from all M2 ears were propagated in thefield in summer 2011. This enabled heritability testing of dominant M2 phe-notypes, rescue of M2 ears with few kernels, and identification of recessivemutants appearing in the M3 generation.
Genotyping, RT-PCR, Zein Protein Analysis, and Fixationfor Microscopy
Plants or seeds were selected for the g-zein deletion using leaf or embryogenomic PCR using DNA extracted according to the urea method (Holdinget al., 2008). For RT-PCR, RNA was extracted and DNase-treated complemen-tary DNA synthesized as previously described (Holding et al., 2011). All primersused for genotyping and RT-PCR are shown in Supplemental Table S5. Zeinswere extracted from mature and developing endosperm as previously described(Wallace et al., 1990). Endosperms of kernels genotyped for the deletion wereprocessed as previously described (Guo et al., 2013). For the 18 DAP stage,kernels were removed, without damaging the base of the kernels, using a razorblade and treated as follows. Kernels were placed embryo-side down on a cleanglass plate and sliced in half longitudinally through the embryo. The embryohalves were placed immediately into DNA extraction buffer on ice, one-half ofthe endosperm was frozen in liquid nitrogen for protein analysis, and the otherhalf kernel was dissected for fixation. Keeping the pericarp intact, the half kernelwas placed cut-side down, and a 1- to 2-mm longitudinal central section wastaken and placed into 5 mL fixative (2% [w/v] Suc, 0.1 M sodium cacodylate, pH7.4, and 5% [v/v] glutaraldehyde). Samples were fixed at 4°C for at least 1 weekwhile genotyping, and SDS-PAGE of zeins was conducted to identify transgenickernels for further processing. Embryo DNA was extracted using microcentrifugescale urea DNA extraction.
Immunogold TEM and Protein Body Counts
Selected kernel segments were further dissected following fixation to removethe apical and basal kernel portions leaving two 1-mm midkernel pieces withpericarp still attached for orientation. Samples were infiltrated in LR White resin(Electron Microscopy Sciences) before sectioning. Ninety-nanometer sectionswere cut using a Diatome diamond knife and a LKBUltrotome III microtome andattached to carbon and Formvar-coated grids (EMS). For immunogold labeling,grids were hybridized in 50-mL drops on Parafilm strips as follows. The gridswere incubated for 30 min in blocking solution, pH 8.2, containing 0.2% (w/v)bovine serum albumin and 0.06% (v/v) Tween 20 in 20 mM Tris-HCl and500 mM NaCl. Rabbit primary antisera were added (a-zein at 1:2,000 andg-zein at 1:100) and incubated overnight at 4°C before three 10-min washes in100 mL blocking solution. Secondary antibody (goat anti-rabbit, 15 nm, goldconjugated from EMS) was added at 1:20 dilution for 1 h at room temperature.The grids were washed three times, rinsed three times in deionized water andair (Holding et al., 2008), and viewed on a Hitachi 7500 TEM.
Protein body images were standardized by viewing the fourth subaleuronestarchy endosperm cell layer. At the 18-DAP kernel stage, the first starchyendosperm cell layer had few protein bodies and increased in size and numberin the layers internal to this. Estimations of protein body number were stan-dardized by making counts within 100-mm square regions within fourth sub-aleurone cell layer cells away from the cell wall and nucleus.
Exon-Seq
Genomic DNA was extracted from leaf tissue using a modified urea ex-traction protocol (Holding et al., 2008). DNA concentration and purity wasassayed with NanoDrop and 100 mg crude DNA purified using Qiagen DNAclean kit. Ten-microgram samples of purified DNA was used for exon capture.DNA library construction, exome capture, and quality control validation werecarried out as a paid service by the Genomic Technologies Facility at IowaState University. For DNA library construction, 3-mg DNA samples weresheared with nebulizers and prepared incorporating bar codes according toIllumina TruSeq DNA Sample Prep kit’s protocol (catalog no. FC–121–2001).For quality control of the precapture libraries, a Bioanalyzer 2100 was used toestablish library size and quantity. For hybridization, each library was dilutedto 30 ng mL–1. Fragmented DNA from four opaque deletion mutants and theprogenitor K0326Y QPM parent were used to make Illumina TruSeq DNAlibraries, which allowed incorporation of bar codes within the adaptors andpooling of the five samples. The five libraries (300 ng per library) were
128 Plant Physiol. Vol. 164, 2014
Yuan et al.
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

combined together for one array hybridization using the Nimblegen Zm B73HX3 exome capture array (1.5mg total per array). The procedure of hybridi-zation followed NimbleGen Array User’s Guide (Plant Sequence CaptureIllumina Optimized protocol (version 1, 2010). The eluted libraries weresubjected to 10 cycles of amplification using the TruSeq enrichment PCR kit.For validation of hybridization efficacy, the ratios of five random genes weretested by real-time quantitative RT-PCR in the pre- and postcaptured librariesas part of the service at the Iowa State University genomics facility (notshown). Libraries were then sequenced (100-bp single-end reads) using theIllumina Genome Analyzer 2 service at the University of Nebraska genomicscore facility. Short reads were discarded if their average quality scores weresmaller than 30. The adapter sequence on 59 end and six nucleotides on the 59end of 7-base bar codes were used to determine the trimming position anddeconvolute the reads. After trimming the adapter sequence and bar codes,the grouped short reads were mapped against the B73 genome (ZmB73_Ref-Gen_v2 from http://www.maizesequence.org) using Bowtie (Langmeadet al., 2009), allowing up to two base mismatches per read. Reads mapped tomultiple locations were discarded. Numbers of reads in exons were countedusing the HTSeq-count tool using B73 gene annotations (ZmB73_5b_FGS fromhttp://www.maizesequence.org), and the ‘union’ resolution mode was used.For pairwise comparisons, the edgeR package (Robinson et al., 2010) withtrimmed mean of M-values normalization method (Robinson and Oshlack, 2010)was used to analyze the numbers of reads aligned to exons.
The use of a Nimblegen exome array to purge repetitive and intron sequencesand specifically select for exon sequences resulted in the template size of eachmutantbeing reduced from 3.2 gigabase pairs to approximately 50 megabase pairs (64-foldsimplification) and allowed us to sequence this pool in a single Genome Analyzer IIflow cell lane, which has a coverage of 3.6 gigabase pairs. After deconvolution ofreads from the five lines, they were assembled against the B73 genome. The non-isogenic nature of the QPM background compared with B73 precluded the pos-sibility to assemble reads and infer subexon mutations in this population, but wewere able to effectively score for presence/absence of exons. Deletions were calledaccording to an algorithm that considered both the number of reads for a givenexon in the nonmutagenized QPM and the ratio of reads between nonmutagenizedQPM and a mutagenized line. An exon was automatically called a deletion can-didate if its P value was smaller than 10–15 (30% of candidates fit this criterion). Theother 70% of called candidates had P values ranging between 10–15 and 10–5. Thesecandidates were only called when the number of reads for the mutagenized linewere not larger than 0, 1, 2, 3, and 5 when the numbers of reads for the non-mutagenized reference line were in ranges of (30–50), (50–70), (70–100), (100–150),and (150 and above), respectively. For example, a candidate with 125 reads innonmutagenized line must have three or less reads in the mutagenized line.
RNA-Seq
Even though we are mostly interested in the loss of endosperm-specifictranscripts, total RNA was extracted as previously described (Holding et al.,2011) from leaf, root, and whole seed. RNA was purified using Qiagen RNeasycleanup kits, and a pool was made for RNA-seq to identify transcripts abol-ished from multiple tissues. Ten micrograms of RNA was used for preparationof Illumina complementary DNA libraries, and 100-bp reads were obtainedfrom Illumina Genome Analyzer 2 platform using RNA from nonmutagenizedK0326Y, line 107, and one other deletion mutant per flow cell lane. For theRNA-seq reads, the same method as exon reads was used to deconvolute thebar codes, trim the adapter sequences, map to the reference genome, andanalyze the abundance of reads in each gene. A threshold value for foldchange differential expression was set at log2 greater than 1 (2-fold actualvalues) and adjusted P values , 0.001 for the null hypothesis, although weshow gene changes at least log2 greater than 2 (4-fold actual values). Differ-entially expressed genes were annotated using the Gene Ontology database.
Sequence data from this article can be found in the GenBank/EMBL datalibraries under accession numbers AF371263, DQ400403, AF371261, andNM001174192 for primer design in the 50-, 16-, and 27-kD gamma-zein genesand the tubulin control gene, respectively. All other genes are listed by theirmaize genome descriptors (e.g. GRMZM2G398628).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. M3 phenotypes of selected segregating K0326YQPM opaque deletion mutants.
Supplemental Table S1. Candidate gene deletions in deletion line 107from exon-seq.
Supplemental Table S2. RNA-seq showing genes identified as at least 4-folddifferentially expressed between K0326Y QPM and deletion line 107.
Supplemental Table S3. Exon reads and RNA-seq reads for genes withinthe chromosome 7 deletion.
Supplemental Table S4. Exon reads and RNA-seq reads for genes withinthe chromosome 9 deletion.
Supplemental Table S5. Primers for genomic and RT-PCR.
ACKNOWLEDGMENTS
We thank Justin Hegstad and Allen Peckrul at North Dakota StateUniversity for their technical support in determining the appropriate radiationdosage for the seed mutagenesis, Kyla Morton and Dr. Xiaomei Guo atUniversity of Nebraska-Lincoln for assisting with field work, Dr. Han Chen ofthe University of Nebraska Microscopy Core Facility for help with immunogoldlabeling, and anonymous reviewers for their constructive comments.
Received October 21, 2013; accepted November 6, 2013; published November8, 2013.
LITERATURE CITED
Bolon YT, Haun WJ, Xu WW, Grant D, Stacey MG, Nelson RT, GerhardtDJ, Jeddeloh JA, Stacey G, Muehlbauer GJ, et al (2011) Phenotypic andgenomic analyses of a fast neutron mutant population resource in soy-bean. Plant Physiol 156: 240–253
Coleman CE, Clore AM, Ranch JP, Higgins R, Lopes MA, Larkins BA(1997) Expression of a mutant a-zein creates the floury2 phenotype intransgenic maize. Proc Natl Acad Sci USA 94: 7094–7097
Dannenhoffer JM, Bostwick DE, Or E, Larkins BA (1995) opaque-15, amaize mutation with properties of a defective opaque-2 modifier. ProcNatl Acad Sci USA 92: 1931–1935
Dumanovi J, Denic M, Ehrenber L, Bergstra K (1969) Radiation-inducedheritable variation of quantitative characters in wheat. Hereditas-Genetiskt Arkiv 62: 221–227
Geetha KB, Lending CR, Lopes MA, Wallace JC, Larkins BA (1991)opaque-2 modifiers increase g-zein synthesis and alter its spatial dis-tribution in maize endosperm. Plant Cell 3: 1207–1219
Geevers HO, Lake JK (1992) Development of modified opaque-2 maize inSouth Africa. In ET Mertz, ed, Quality Protein Maize. American Societyof Cereal Chemists, St. Paul, pp 49–78
Gibbon BC, Larkins BA (2005) Molecular genetic approaches to develop-ing quality protein maize. Trends Genet 21: 227–233
Guo X, Yuan L, Chen H, Sato SJ, Clemente TE, Holding DR (2013)Nonredundant function of zeins and their correct stoichiometric ratiodrive protein body formation in maize endosperm. Plant Physiol 162:1359–1369
Holding DR, Hunter BG, Chung T, Gibbon BC, Ford CF, Bharti AK,Messing J, Hamaker BR, Larkins BA (2008) Genetic analysis of opaque2modifier loci in quality protein maize. Theor Appl Genet 117: 157–170
Holding DR, Hunter BG, Klingler JP, Wu S, Guo X, Gibbon BC, Wu R,Schulze JM, Jung R, Larkins BA (2011) Characterization of opaque2modifier QTLs and candidate genes in recombinant inbred lines derivedfrom the K0326Y quality protein maize inbred. Theor Appl Genet 122:783–794
Holding DR, Larkins BA (2006) The development and importance of zeinprotein bodies in maize endosperm. Maydica 51: 243–254
Holding DR, Meeley RB, Hazebroek J, Selinger D, Gruis F, Jung R,Larkins BA (2010) Identification and characterization of the maize arogenatedehydrogenase gene family. J Exp Bot 61: 3663–3673
Holding DR, Otegui MS, Li BL, Meeley RB, Dam T, Hunter BG, Jung R,Larkins BA (2007) The maize floury1 gene encodes a novel endoplasmicreticulum protein involved in zein protein body formation. Plant Cell 19:2569–2582
Hossain KG, Riera-Lizarazu O, Kalavacharla V, Vales MI, Maan SS,Kianian SF (2004) Radiation hybrid mapping of the species cytoplasm-specific (scsae) gene in wheat. Genetics 168: 415–423
Plant Physiol. Vol. 164, 2014 129
Role of g-Zein in opaque2 Endosperm Modification
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.

Jia M, Wu H, Clay KL, Jung R, Larkins BA, Gibbon BC (2013) Identificationand characterization of lysine-rich proteins and starch biosynthesis genes inthe opaque2 mutant by transcriptional and proteomic analysis. BMC PlantBiol 13: 60
Johri MM, Coe EH Jr (1983) Clonal analysis of corn plant development. I.The development of the tassel and the ear shoot. Dev Biol 97: 154–172
Kim CS, Gibbon BC, Gillikin JW, Larkins BA, Boston RS, Jung R (2006)The maize Mucronate mutation is a deletion in the 16-kDa g-zein genethat induces the unfolded protein response. Plant J 48: 440–451
Kim CS, Hunter BG, Kraft J, Boston RS, Yans S, Jung R, Larkins BA(2004) A defective signal peptide in a 19-kD a-zein protein causes theunfolded protein response and an opaque endosperm phenotype in themaize De*-B30 mutant. Plant Physiol 134: 380–387
Langmead B, Trapnell C, Pop M, Salzberg SL (2009) Ultrafast andmemory-efficient alignment of short DNA sequences to the human ge-nome. Genome Biol 10: R25
Li X, Song Y, Century K, Straight S, Ronald P, Dong X, Lassner M, ZhangY (2001) A fast neutron deletion mutagenesis-based reverse geneticssystem for plants. Plant J 27: 235–242
Lopes MA, Larkins BA (1991) g-Zein content is related to endospermmodification in Quality Protein Maize. Crop Sci 31: 1655–1662
Lopes MA, Larkins BA (1995) Genetic analysis of opaque2 modifier geneactivity in maize endosperm. Theor Appl Genet 91: 274–281
Lopes MA, Takasaki K, Bostwick DE, Helentjaris T, Larkins BA (1995)Identification of two opaque2 modifier loci in quality protein maize. MolGen Genet 247: 603–613
Maddaloni M, Donini G, Balconi C, Rizzi E, Gallusci P, Forlani F,Lohmer S, Thompson R, Salamini F, Motto M (1996) The transcrip-tional activator Opaque-2 controls the expression of a cytosolic form ofpyruvate orthophosphate dikinase-1 in maize endosperms. Mol GenGenet 250: 647–654
McCarty DR, Settles AM, Suzuki M, Tan BC, Latshaw S, Porch T, RobinK, Baier J, Avigne W, Lai JS, et al (2005) Steady-state transposon mu-tagenesis in inbred maize. Plant J 44: 52–61
Mertz ET, Bates LS, Nelson OE (1964) Mutant gene that changes proteincomposition and increases lysine content of maize endosperm. Science145: 279–280
Myers AM, James MG, Lin QH, Yi G, Stinard PS, Hennen-Bierwagen TA,Becraft PW (2011) Maize opaque5 encodes monogalactosyldiacylgly-cerol synthase and specifically affects galactolipids necessary for amy-loplast and chloroplast function. Plant Cell 23: 2331–2347
Naito K, Kusaba M, Shikazono N, Takano T, Tanaka A, Tanisaka T,Nishimura M (2005) Transmissible and nontransmissible mutationsinduced by irradiating Arabidopsis thaliana pollen with g-rays and carbonions. Genetics 169: 881–889
Poethig RS, Coe EH, Johri MM (1986) Cell lineage patterns in maize em-bryogenesis: a clonal analysis. Dev Biol 117: 392–404
Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bioconductorpackage for differential expression analysis of digital gene expressiondata. Bioinformatics 26: 139–140
Robinson MD, Oshlack A (2010) A scaling normalization method for dif-ferential expression analysis of RNA-seq data. Genome Biol 11: R25
Sato Y, Shirasawa K, Takahashi Y, Nishimura M, Nishio T (2006) Mutantselection from progeny of g-ray-irradiated rice by DNA heteroduplexcleavage using Brassica petiole extract. Breed Sci 56: 179–183
Schmidt RJ, Burr FA, Aukerman MJ, Burr B (1990) Maize regulatory geneopaque-2 encodes a protein with a “leucine-zipper” motif that binds tozein DNA. Proc Natl Acad Sci USA 87: 46–50
Settles AM (2005) Maize community resources for forward and reversegenetics. Maydica 50: 405–414
Settles AM, Holding DR, Tan BC, Latshaw SP, Liu J, Suzuki M, Li L,O’Brien BA, Fajardo DS, Wroclawska E, et al (2007) Sequence-indexedmutations in maize using the UniformMu transposon-tagging popula-tion. BMC Genomics 8: 116
Vasal SK, Villegas E, Bjarnason M, Gela W, Goertz P (1980) Geneticmodifiers and breeding strategies in developing hard endosperm opa-que2 materials. In WG Pollmer, RH Phillips, eds, Quality Traits of Maizefor Grain and Silage Use. Martinus Nijhoff, London, pp 37–73
Vollbrecht E, Duvick J, Schares JP, Ahern KR, Deewatthanawong P, Xu L,Conrad LJ, Kikuchi K, Kubinec TA, Hall BD, et al (2010) Genome-widedistribution of transposed Dissociation elements in maize. Plant Cell 22:1667–1685
Wallace JC, Lopes MA, Paiva E, Larkins BA (1990) New methods for ex-traction and quantitation of zeins reveal a high content of g-zein inmodified opaque-2 maize. Plant Physiol 92: 191–196
Wang GF, Wang F, Wang G, Wang F, Zhang XW, Zhong MY, Zhang J, LinDB, Tang YP, Xu ZK, et al (2012) Opaque1 encodes a myosin XI motorprotein that is required for endoplasmic reticulum motility and proteinbody formation in maize endosperm. Plant Cell 24: 3447–3462
Weil C, Monde R, Till B, Comai L, Henikoff S (2005) Mutagenesis andfunctional genomics in maize. Maydica 50: 415–424
Wu JL, Wu CJ, Lei CL, Baraoidan M, Bordeos A, Madamba MRS, Ramos-Pamplona M, Mauleon R, Portugal A, Ulat VJ, et al (2005) Chemical-and irradiation-induced mutants of indica rice IR64 for forward andreverse genetics. Plant Mol Biol 59: 85–97
Wu Y, Holding DR, Messing J (2010) g-Zeins are essential for endospermmodification in quality protein maize. Proc Natl Acad Sci USA 107:12810–12815
Wu Y, Messing J (2010) RNA interference-mediated change in protein bodymorphology and seed opacity through loss of different zein proteins.Plant Physiol 153: 337–347
Wu Y, Yuan L, Guo X, Holding DR, Messing J (2013) Mutation in the seedstorage protein kafirin creates a high-value food trait in sorghum. NatCommun 4: 2217
130 Plant Physiol. Vol. 164, 2014
Yuan et al.
www.plantphysiol.orgon June 27, 2020 - Published by Downloaded from Copyright © 2014 American Society of Plant Biologists. All rights reserved.