depression and anxiety research article -...
TRANSCRIPT

DEPRESSION AND ANXIETY 31:880–892 (2014)
Research ArticleCHILDHOOD MALTREATMENT AND COMBAT
POSTTRAUMATIC STRESS DIFFERENTIALLY PREDICTFEAR-RELATED FRONTO-SUBCORTICAL
CONNECTIVITY
Rasmus M. Birn, Ph.D.,1,2,† Remi Patriat, M.S.,2,† Mary L. Phillips, M.D.,3 Anne Germain, Ph.D.,3and Ryan J. Herringa, M.D., Ph.D.1,3∗
Background: Adult posttraumatic stress disorder (PTSD) has been character-ized by altered fear-network connectivity. Childhood trauma is a major riskfactor for adult PTSD, yet its contribution to fear-network connectivity in PTSDremains unexplored. We examined, within a single model, the contribution ofchildhood maltreatment, combat exposure, and combat-related posttraumaticstress symptoms (PTSS) to resting-state connectivity (rs-FC) of the amygdalaand hippocampus in military veterans. Methods: Medication-free male veter-ans (n = 27, average 26.6 years) with a range of PTSS completed resting-statefMRI. Measures including the Clinician-Administered PTSD Scale (CAPS),Childhood Trauma Questionnaire (CTQ), and Combat Exposure Scale (CES)were used to predict rs-FC using multilinear regression. Fear-network seedsincluded the amygdala and hippocampus. Results: Amygdala: CTQ predictedlower connectivity to ventromedial prefrontal cortex (vmPFC), but greater an-ticorrelation with dorsal/lateral PFC. CAPS positively predicted connectivity toinsula, and loss of anticorrelation with dorsomedial/dorsolateral (dm/dl)PFC.Hippocampus: CTQ predicted lower connectivity to vmPFC, but greater anticor-relation with dm/dlPFC. CES predicted greater anticorrelation, whereas CAPSpredicted less anticorrelation with dmPFC. Conclusions: Childhood trauma,combat exposure, and PTSS differentially predict fear-network rs-FC. Childhoodmaltreatment may weaken ventral prefrontal–subcortical circuitry important inautomatic fear regulation, but, in a compensatory manner, may also strengthendorsal prefrontal–subcortical pathways involved in more effortful emotion regu-lation. PTSD symptoms, in turn, appear to emerge with the loss of connectivity inthe latter pathway. These findings suggest potential mechanisms by which devel-opmental trauma exposure leads to adult PTSD, and which brain mechanisms
1Department of Psychiatry, University of Wisconsin School ofMedicine & Public Health, Madison, Wisconsin2Department of Medical Physics, University of WisconsinSchool of Medicine & Public Health, Madison, Wisconsin3Department of Psychiatry, University of Pittsburgh School ofMedicine, Pittsburgh, Pennsylvania
Contract grant sponsor: AACAP Pilot Research Award (RJH), De-partment of Defense Congressionally Directed Medical ResearchProgram; Contract grant number; PT073961; Contract grant spon-sor: National Institute of Mental Health; Contract grant numbers:MH083035, MH076971, MH088913.
†These authors contributed equally to this work and are consideredfirst authors.
∗Correspondence to: Ryan J. Herringa, Department of Psychia-try, University of Wisconsin School of Medicine & Public Health,6001 Research Park Boulevard, Madison, WI 53719. E-mail:[email protected] for publication 15 October 2013; Revised 30 May 2014;Accepted 13 June 2014
DOI 10.1002/da.22291Published online 8 August 2014 in Wiley Online Library(wileyonlinelibrary.com).
C© 2014 Wiley Periodicals, Inc.

Research Article: Child Trauma, Combat PTSS, Brain Connectivity 881
are associated with the emergence of PTSD symptoms. Depression and Anxiety31:880–892, 2014. C© 2014 Wiley Periodicals, Inc.
Key words: abuse; maltreatment; brain imaging; functional MRI; PTSD;trauma
INTRODUCTIONNeuroimaging studies of adult posttraumatic stressdisorder (PTSD) suggest abnormalities in fronto-subcortical circuitry underlying the regulation of fear re-sponses. Relatively common findings include increasedactivation of the amygdala, insula, and dorsal ante-rior cingulate cortex (dACC), and relative hypoactiva-tion of the ventromedial prefrontal cortex (vmPFC) inresponse to emotional stimuli.[1, 2] The hippocampusshows increased activation to emotional stimuli, but im-paired recruitment during fear extinction in subjects withPTSD.[1, 3] The dACC and vmPFC appear to have op-posing roles in the regulation of fear via the amygdala(facilitation and inhibition, respectively),[4] whereas thehippocampus contextually regulates fear responses viathe amygdala and vmPFC.[5] Within this framework,neural models of PTSD posit disrupted communicationbetween the amygdala and hippocampus, and prefrontalregulatory areas, leading to exaggerated and generalizedfear responses.[2, 5]
Studies of resting-state functional brain connectivity(rs-FC) in PTSD have yielded variable results but sug-gest altered connectivity of both the amygdala and hip-pocampus (see Table 1 for a summary of prior rs-FCstudies involving the amygdala, hippocampus, and/orprefrontal areas). Compared to trauma-exposed con-trols, subjects with PTSD show greater connectivityof the amygdala with the insula,[6–8] lower connec-tivity with the hippocampus,[7] and reduced anticor-relation with pregenual (pg)/dACC.[7] Some of thesedifferences may be specific to amygdala subnuclei, as ev-idenced by a recent study showing reduced anticorrela-tion between the basolateral amygdala (BLA) and thepgACC/dorsomedial (dm)PFC and dACC, and lowerconnectivity with the inferior frontal gyrus in PTSDsubjects.[9] In contrast, there were no significant groupdifferences in centromedial amygdala (CMA) connec-tivity. To date, only one reported study has examinedrs-FC of the hippocampus in PTSD. Relative to non-trauma controls, PTSD subjects had lower connectiv-ity between the anterior hippocampus and dACC/pre-supplementary motor area, and lower connectivitybetween the posterior hippocampus and pgACC andposterior cingulate/precuneus.[10] Although the resultsof these studies vary, they appear to suggest that PTSDis characterized by lower dorsal/medial prefrontal–subcortical rs-FC, a pathway that is important in effortfulemotion regulation.[11] On the other hand, there is littleevidence for impaired ventral prefrontal–subcortical rs-
FC, a pathway important in the automatic regulation offear and emotion.[4, 11]
The contribution of childhood maltreatment to al-tered rs-FC of the amygdala and hippocampus in PTSDremains unexplored but may explain some of the abovediscrepancies. A history of childhood trauma is one ofthe largest risk factors for adult PTSD, with an effectsize comparable to the adult index trauma.[12] Althoughanalytic approaches and study samples vary, prior rs-FCstudies suggest that childhood maltreatment or early-life stress may alter amygdala and hippocampus con-nectivity in ways that create vulnerability for developingPTSD as an adult (see Table 1 for a summary). Usingseed-based approaches, childhood maltreatment/early-life stress has been associated with lower connectivity be-tween the amygdala and insula/hippocampus,[13] lowerconnectivity between the amygdala and vmPFC,[14, 15]
and lower connectivity between the hippocam-pus and vmPFC.[15] Lower amygdala–vmPFC andhippocampus–vmPFC connectivity also mediated thedevelopment of internalizing symptoms by late adoles-cence, suggesting these changes create vulnerability forpsychopathology.[14, 15] Using graph-theory approaches,childhood maltreatment/early-life stress has been asso-ciated with decreased global connectivity of the amyg-dala, hippocampus, and PFC,[16] and decreased localconnectivity and increased hub-like properties of theamygdala.[17] In contrast, resilience to childhood traumahas been associated with increased global connectivityof the hippocampus at rest.[17] Taken together, thesefindings suggest that childhood trauma exposure mayweaken fear-regulatory circuitry particularly in ventralprefrontal–subcortical pathways, creating a vulnerablebrain substrate for the development of adult PTSD.
Here we examined, using a within-subjects design,how childhood maltreatment, combat exposure, andcombat posttraumatic stress symptoms (PTSS) predictresting-state connectivity of the amygdala and hip-pocampus in medication-free, young military veterans.This study expands on prior rs-FC studies of PTSDin several important ways. First, we employed a di-mensional approach to examine the neural correlatesof PTSS (both above and below the PTSD thresh-old), which offers greater specificity to PTSD symp-toms. Second, we included continuous measures of adulttrauma (combat exposure) and childhood trauma withinthe same model to examine their relative contributionsto brain connectivity. Prior rs-FC studies of PTSDhave not included childhood or adult trauma exposureas covariates, though some have tried to control for
Depression and Anxiety

882 Birn et al.
TA
BL
E1.
Sum
mar
yof
prio
rre
stin
g-st
ate
func
tion
alco
nnec
tivi
tyst
udie
sof
child
hood
stre
ss/t
raum
a,an
dad
ult
PT
SDsy
mpt
oms
Ref
.Se
edor
netw
ork
anal
ysis
Seed
/net
wor
kap
proa
chSt
ress
/tra
uma
type
Subj
ects
(n)
Mai
nfin
ding
s(o
rigi
nal
conn
ectiv
itysi
gn)
Not
es
Chi
ldho
odst
ress
/tra
uma
expo
sure
[13]
Seed
Am
ygda
ladA
CC
PC
Cdm
PFC
Chi
ldho
odem
otio
nal
mal
trea
tmen
t(C
EM
)–
emot
iona
lab
use/
negl
ect
Adu
ltsw
ithC
EM
(44)
Adu
ltsw
ithou
tCE
M(4
4)↓A
myg
dala
-ins
ula/
hipp
ocam
pus
(+)
↓Am
ygda
la-p
recu
neus
(−)
↓dA
CC
-vm
PFC
(+)
↓dA
CC
-pre
cune
us(−
)
Psy
chia
tric
med
icat
ion
use
not
repo
rted
.Gro
ups
mat
ched
for
psyc
hopa
thol
ogy.
Phy
sica
l/se
xual
abus
eex
clud
ed.
[14]
Seed
Am
ygda
laM
ater
nal/
fam
ilyst
ress
upto
1ye
arof
age
Ado
lesc
entc
omm
unity
sam
ple
(57)
↓Am
ygda
la-v
mP
FC(+
)E
ffect
sfo
und
infe
mal
eson
lyan
dm
edia
ted
bych
ildco
rtis
olle
vels
atag
e4.
5ye
ars.
[15]
Seed
Am
ygda
laH
ippo
cam
pus
Chi
ldho
odm
altr
eatm
ent
–al
ltyp
esco
mbi
ned
Ado
lesc
entc
omm
unity
sam
ple
(64)
↓Am
ygda
la-s
gAC
C(+
)↓H
ippo
cam
pus-
sgA
CC
(+)
Am
ygda
laef
fect
sin
fem
ales
only
,hi
ppoc
ampu
sef
fect
sin
both
sexe
s.[3
7]Se
edA
myg
dala
PC
CC
hild
hood
mal
trea
tmen
t(C
M)–
phys
ical
,sex
ual,
orem
otio
nala
buse
Hea
lthy
adul
tsw
ithC
M(1
2)H
ealth
yad
ults
with
outC
M(9
)
↓PC
C-v
mP
FC(+
)↓P
CC
-IT
C(+
)↑A
myg
dala
-vm
PFC
(tre
ndon
ly)
Apr
iori
sear
chre
gion
sw
ere
used
.A
llsu
bjec
tsw
ere
free
ofcu
rren
tps
ychi
atri
cill
ness
.Fou
rad
ults
with
CM
repo
rted
past
psyc
hiat
ric
illne
ss.
[16]
Net
wor
kG
raph
theo
ryC
hild
hood
negl
ect(
CN
)–
emot
iona
l/ph
ysic
alA
dult
MD
Dw
ithC
N(1
8)A
dult
MD
Dw
ithou
tCN
(20)
Hea
lthy
adul
tsw
ithou
tCN
(20)
MD
Dw
ithvs
.with
outC
N:
↓glo
balc
onne
ctiv
ityin
dl/v
l/dm
PFC
,ins
ula,
caud
ate,
thal
amus
,par
ahip
poca
mpa
lgy
rus,
amyg
dala
,hip
poca
mpu
s
All
MD
Dpa
tient
sw
ere
rece
ivin
gan
tidep
ress
antm
edic
atio
ns.
Pri
mar
yan
alys
isus
edgl
obal
sign
alre
gres
sion
.Mos
tC
N-a
ssoc
iate
dfin
ding
sdi
sapp
ear
whe
ngl
obal
sign
alre
gres
sion
was
notu
sed.
Glo
bal
conn
ectiv
ity=
degr
ee.
[17]
Net
wor
kG
raph
theo
ryE
arly
-life
stre
ss(E
LS)
such
asab
use,
negl
ect,
care
give
rde
ath,
acci
dent
“Sus
cept
ible
”ad
ults
with
curr
ento
rpa
stM
DD
/PT
SDan
dE
LS
(19)
“Res
ilien
t”ad
ults
(no
MD
D/P
TSD
)with
EL
S(7
)“C
ontr
ol”
adul
tsw
ithou
tE
LS
(12)
Susc
eptib
levs
.oth
ergr
oups
:↓l
ocal
conn
ectiv
ityan
d↑h
ubpr
oper
ties
ofam
ygda
la,↓
hub
prop
ertie
sof
dAC
C,↓
loca
lco
nnec
tivity
ofle
ftvl
PFC
Res
ilien
tvs.
othe
rgr
oups
:↓g
loba
lcon
nect
ivity
and
hub
prop
ertie
sof
righ
tvlP
FC,↓
loca
lco
nnec
tivity
ofdA
CC
,↑lo
cal
conn
ectiv
ityof
mP
FC,↑
glob
alco
nnec
tivity
ofhi
ppoc
ampu
s
All-
fem
ale
sam
ple.
Onl
y↓h
ubpr
oper
ties
ofri
ghtv
lPFC
for
the
resi
lient
grou
psu
rviv
em
ultip
leco
mpa
riso
nco
rrec
tion.
Loc
alco
nnec
tivity
=ef
ficie
ncy.
Hub
prop
ertie
s=
betw
eenn
ess
cent
ralit
y.G
loba
lcon
nect
ivity
=de
gree
.
(Con
tinue
d)
Depression and Anxiety

Research Article: Child Trauma, Combat PTSS, Brain Connectivity 883
TA
BL
E1.
Con
tinu
ed
Ref
.Se
edor
netw
ork
anal
ysis
Seed
/net
wor
kap
proa
chSt
ress
/tra
uma
type
Subj
ects
(n)
Mai
nfin
ding
s(o
rigi
nal
conn
ectiv
itysi
gn)
Not
es
Adu
ltP
TSD
[58]
Seed
PC
Cm
PFC
Chi
ldho
odab
use
PT
SD(1
7)N
TC
(15)
↓PC
Cto
mP
FC,p
arie
talc
orte
x,m
iddl
ete
mpo
ralg
yrus
,pa
rahi
ppoc
ampa
lgyr
us,i
nsul
a,hi
ppoc
ampu
s,am
ygda
la↓m
PFC
topa
riet
alco
rtex
,PC
C
All-
fem
ale
sam
ple.
13P
TSD
subj
ects
wer
ere
ceiv
ing
psyc
hiat
ric
med
icat
ions
.
[6]
Seed
Am
ygda
laC
omba
tP
TSD
(17)
TE
C(1
7)↑A
myg
dala
-ins
ula
(+)
All-
mal
esa
mpl
e.P
TSD
subj
ects
wer
eun
med
icat
edat
time
ofst
udy.
[7]
Seed
Am
ygda
laC
omba
tP
TSD
(15)
TE
C(1
4)↑A
myg
dala
-ins
ula
(+)
↓Am
ygda
la-h
ippo
cam
pus(
+)↓A
myg
dala
-ros
tral
/dor
salA
CC
(−)
Apr
iori
sear
chre
gion
sw
ere
used
.A
ll-m
ale
sam
ple.
Tw
oP
TSD
subj
ects
wer
eta
king
traz
odon
e.[8
]Se
edP
CC
vmP
FCA
nter
ior
insu
la
Com
bat
PT
SD(1
5)T
EC
(15)
NT
C,c
omm
unity
sam
ple
(15)
PT
SDvs
.TE
C:
↑PC
C-a
myg
dala
and
-ins
ula
(−)
*↑In
sula
-am
ygda
la(+
)*↑
Insu
la-h
ippo
cam
pus
(−)
PT
SDvs
.NT
C:
↓PC
C-h
ippo
cam
pus
(+)
↓vm
PFC
-ros
tral
AC
C(+
)
Apr
iori
sear
chre
gion
sw
ere
used
.A
ll-m
ale
sam
ple.
Tw
oP
TSD
subj
ects
wer
eta
king
traz
odon
e.T
hetw
oco
mpa
riso
ngr
oups
wer
eco
mbi
ned
for
prim
ary
anal
ysis
.
[10]
Seed
Ant
erio
rhi
ppoc
ampu
sP
oste
rior
hipp
ocam
pus
Var
ious
–in
terp
erso
nal
viol
ence
,acc
iden
t,co
mba
t,ch
ildho
odab
use,
rape
,nat
ural
disa
ster
PT
SD(1
7)G
AD
(39)
NT
C(6
0)
PT
SDvs
.NT
C:
↓Ant
erio
rhi
ppoc
ampu
sto
dAC
C/p
re-S
MA
(+)
↓Pos
teri
orhi
ppoc
ampu
sto
PC
C,
PC
,pgA
CC
(+)
Apr
iori
sear
chre
gion
san
dgl
obal
sign
alre
gres
sion
wer
eus
ed.F
our
PT
SDsu
bjec
tsw
ere
rece
ivin
gps
ychi
atri
cm
edic
atio
ns.
Pos
teri
orhi
ppoc
ampa
lfind
ings
wer
esp
ecifi
cto
PT
SD.
[9]
Seed
Bas
olat
eral
amyg
dala
(BL
A)
Cen
trom
edia
lam
ygda
la(C
MA
)
Com
bat
PT
SD(2
0)T
EC
(22)
↓BL
A-p
gAC
C/d
mP
FC(−
)↓B
LA
-dA
CC
(−)
↓BL
Ato
IFG
(+)
12P
TSD
and
two
TE
Csu
bjec
tsw
ere
rece
ivin
gps
ychi
atri
cm
edic
atio
ns.
Stud
ies
incl
uded
here
asse
ssed
one
orm
ore
key
fear
-net
wor
kno
des
incl
udin
gth
eam
ygda
la,h
ippo
cam
pus,
and/
orpr
efro
ntal
cort
ex.N
ote
that
decr
ease
sin
conn
ectiv
ityth
ator
igin
ally
have
ane
gativ
esi
gnre
flect
alo
ssof
antic
orre
latio
n.
Depression and Anxiety

884 Birn et al.
adult trauma (combat exposure) by contrasting with ahealthy combat-exposed group. Finally, this study uses awithin-subjects design that is less likely to be affectedby confounding variables that may influence groupcontrasts. Our primary hypotheses were that child-hood trauma would predict lower amygdala–vmPFCand hippocampal-vmPFC connectivity, whereas adulttrauma would predict greater hippocampal-vmPFC con-nectivity as an adaptive mechanism.[18] We expected thatPTSS, and possibly adult trauma, would predict lossof anticorrelation between the amygdala/hippocampusand dorsal prefrontal areas including pgACC, dmPFC,and dACC, and greater amygdala–insula connectivitygiven the findings from previous resting-state studies ofPTSD.
METHODSPARTICIPANTS
Twenty-eight combat veterans (all males, right-handed; 26.6 ± 2.6years, and previously described[19]) from Operations Enduring andIraqi Freedom were recruited from ongoing studies to participate inthis study. Prior military service information was obtained by DDForm 214 documentations. One subject was unable to complete thescan due to back discomfort, leaving 27 subjects in the final analysis.All participants were nonmedicated for at least 3 weeks (2 months forfluoxetine) at the time of the study. Written informed consent wasobtained following the University of Pittsburgh Institutional ReviewBoard guidelines. Exclusion criteria included active substance abuse(past month), suicidality, psychotic or bipolar disorder, MRI con-traindication, or neurological disease. In addition, participants wereexcluded if they were currently receiving treatment for traumatic braininjury (TBI) or had concussive symptoms at the time of the study. Ofthe 27 participants, 12 (44.4%) reported exposure to blast, fire, or ex-plosion. Eight participants (29.6%) reported a history of closed headinjury, and two of them also endorsed a loss of consciousness during de-ployment. Six of these participants endorsed being dazed, confused or“seeing stars,” two endorsed not remembering the injury, and four en-dorsed having symptoms of concussion afterward. Symptoms includedheadache, dizziness, and irritability. However, none had concussivesymptoms or were being treated for any sequelae of TBI at the timeof participation. Seventeen subjects met full criteria for current PTSD(past month) using the F1/I2 criteria of the Clinician-AdministeredPTSD Scale (CAPS).[20,21] PTSD symptoms for all subjects were re-lated to combat and not childhood maltreatment experiences.
BEHAVIORAL AND CLINICAL MEASURESIn addition to the CAPS, the Structured Clinical Interview for
DSM-IV Axis I Disorders (SCID-I) was conducted to examine cur-rent and past psychiatric diagnoses.[22] Three subjects met criteria forcurrent major depressive disorder. Subjects were free of other cur-rent comorbid psychiatric disorders. Past DSM-IV diagnoses includedmajor depressive disorder (n = 4), bulimia nervosa (n = 1), alcoholabuse/dependence (n = 14), cannabis abuse/dependence (n = 4), andcocaine abuse/dependence (n = 1). The prevalence of these diagnosesdid not significantly differ between PTSD and non-PTSD subjects(Chi-squared test, P � .09). The Beck Depression Inventory (BDI)[23]
was utilized to measure depressive-symptom severity. The Combat Ex-posure Scale (CES)[24] was used to assess the level of combat exposure.Childhood maltreatment history was quantified using the ChildhoodTrauma Questionnaire (CTQ),[25] which yields a continuous measureof childhood maltreatment experiences based on a Likert rating scale
for each item. IQ was estimated using the National Adult Reading Test(NART).[26] Supporting Information Table S1 summarizes the differ-ent demographic, behavioral, clinical measures, and their correlations.
DATA ACQUISITIONFunctional and structural scans were acquired using a 3T Siemens
Trio MRI scanner. Functional data were acquired using a 5.5-minFAIR-QUIPSSII sequence: oblique acquisition, TE = 18 ms, TR =4,000 ms, TI1 = 700 ms, TI2 = 1,000 ms, FOV = 240 × 240 mm, 3.75× 3.75 mm2 in-plane resolution, slice thickness = 6 mm, 21 slices. Thispulse sequence can provide simultaneous estimates of cerebral bloodflow and blood oxygenation level–dependent (BOLD) signal. For thecurrent study, we use only the BOLD signal, processed in a man-ner described below. Recent work has demonstrated that BOLD mea-sures derived from arterial spin labeling scans provide similar measuresof functional connectivity as data acquired with conventional BOLDfMRI acquisition.[27] Structural brain data were obtained using an axialT1-weighted MPRAGE sequence: TR = 2.2 s, TE = 3.29 ms, TI =1 s, FOV = 256 × 192mm, flip angle = 9°, voxel size = 1 × 1 × 1 mm,192 slices.
PREPROCESSINGAll preprocessing and analysis were carried out using Analysis
of Functional NeuroImages (AFNI),[28] unless otherwise specified.BOLD signal was extracted from the acquired ASL data by addingto each time point the average ASL signal from the neighboring timepoints (3dcalc).[29]
For preprocessing, the EPI images were first despiked(3dDespike),[30] and image time series were registered in timeusing a six-parameter (rigid-body) alignment to correct for subjectmotion (3dvolreg). Resulting BOLD resting scans were alignedto the anatomical scan and the outcome was checked visually toensure that no obvious misalignment was present (align epi anat.pyand @Align Centers). The anatomy was converted to standardTalairach-Tournoux space and resampled to a 2 × 2 × 2 mm grid(@auto tlrc). This transformation was applied to the BOLD restingscans (adwarp). The first four volumes of the resting-state scans wereignored to account for T1 saturation effects. Cerebrospinal fluid(CSF), gray and white matter segmentation were performed on eachsubject’s anatomical scan using the “fast” routine from the FMRIBSoftware Library (FSL).[31–33] White matter and CSF masks wereresampled to the resting scan’s grid (3dresample). The CSF mask waseroded once in each direction to ensure that the voxels present in themask would only overlap with voxels representing primarily CSF onthe resting scan. The white matter mask was eroded twice to makesure that very little gray matter signal would be included in the finalwhite matter mask. The final white matter and CSF eroded maskswere used to create nuisance regressors by extracting the averagetime series over each of those masks (3dROIstats). These averagetime series along with the six motion parameters were entered asnuisance variables in a regression model to remove their effect onthe resting data, keeping the residual time series (3dDeconvolve).Resting data were then temporally filtered (3dBandpass) to ignoresignal frequencies <0.01Hz and >0.1Hz. Finally, resting scans werespatially smoothed with a FWHM of 6 mm (3dmerge).
RESTING-STATE CONNECTIVITY ANALYSISResting-state connectivity analyses were performed using a seed-
based (region-of-interest or ROI) approach. For our primary analy-ses, we used four ROIs: left and right amygdala (Talairach LPI co-ordinates: −23, −6, −20 and 21, −6, −20, respectively), and left andright hippocampus (−31, −25, −11 and 29, −25, −11, respectively).AFNI was used to generate 4 mm spheres centered on the above
Depression and Anxiety

Research Article: Child Trauma, Combat PTSS, Brain Connectivity 885
Figure 1. Functional connectivity maps showing regions that are significantly connected to (A) left amygdala and (B) left hippocampus.Seed regions are shown on the left, with connectivity maps on the right. Connectivity maps for the right amygdala and hippocampusshowed similar patterns. Positive connectivity is indicated by orange-yellow overlays, and negative connectivity by blue overlays. Foraxial and coronal views, the right side of the brain faces left. Maps are displayed at voxelwise P � .005.
co-ordinate locations provided by the Talairach Daemon.[34,35] Allseeds were resampled to the resolution of the resting-state EPIs(3dresample) and average time series were computed for each seed(3dmaskave). Connectivity maps were generated for each subject andeach seed using a linear regression (3dDeconvolve). To facilitate com-parison with prior rs-FC studies of PTSD, we also conducted supple-mentary seed-based analyses using amygdala[9] and hippocampal[10]
subdivisions, as well as anterior insula.[8] These analyses are furtherdescribed in the Supporting Information.
At the group level and for each seed independently, a multivariateregression analysis was conducted (3dttest++) to generate group-levelconnectivity maps for the above-mentioned seeds including CAPS,CES, and CTQ scores. Correlation coefficients were converted toZ-scores using Fisher’s-Z transformation. Multiple comparison cor-rection was performed using Monte Carlo simulation (3dClustSim),which incorporates the estimated smoothness of the data to establishthe likelihood of false positives of different cluster sizes (i.e., clustersize thresholding).[36] The cluster threshold was 357 voxels (2 × 2 ×2 mm3) at an individual voxel threshold of P � .005, resulting in acorrected α � 0.05.
RESULTSFor the following multivariate regression analyses, re-
lationships between the variable of interest and residu-alized connectivity (adjusted for other variables in themodel) are shown in the figures as indicated. Relation-ships between the variable of interest and raw con-nectivity scores can be seen in Supporting InformationFig. S1.
RESTING-STATE CONNECTIVITYFigure 1 shows the connectivity maps for left amygdala
and hippocampus (threshold: P = .005). Connectivitymaps for right amygdala and hippocampus were similar.The left and right amygdala showed positive functionalconnectivity to each other as well as the insula, ven-
tral striatum, hippocampus, parahippocampal gyrus, andmedial frontal gyrus, and negative connectivity to poste-rior cingulate and occipital cortex. The left and right hip-pocampus showed positive connectivity with each otheras well as other regions including the lentiform nucleus,insula, and thalamus. Connectivity maps for amygdalaand hippocampal subdivisions are described in the sup-plemental text and displayed in Supporting InformationFigs. S2 and S3, respectively.
CORRELATION WITH CHILDHOODMALTREATMENT (CTQ TOTAL SCORE)
Amygdala Connectivity. CTQ negatively pre-dicted connectivity from left and right amygdala tomPFC including vmPFC/rostral (r)ACC and dorsome-dial (dm)PFC (BA 9, 10, 32; Fig. 2A), indicating morenegative residualized connectivity with higher CTQ (seescatter plots, Fig. 2B). In addition, CTQ negatively pre-dicted connectivity between left amygdala and lateralPFC including both ventrolateral (vl) and dorsolateral(dl) PFC (BA 9, 10, 47; Table 2). Like the mPFC clus-ters, this association indicated more negative residu-alized connectivity with higher CTQ scores. Finally,CTQ positively predicted connectivity from left andright amygdala to cerebellum (Table 2). Supplementaryanalyses of the CMA and BLA seeds revealed overallsimilar patterns to the primary amygdala seed. In thiscase, CTQ negatively predicted connectivity from CMAand BLA to mPFC areas including vmPFC/rACC anddmPFC (Supporting Information Fig. S4a-b, Table S2).However, the negative association of CTQ with connec-tivity to lateral PFC (specifically dlPFC) was only presentfor the BLA (Supporting Information Table S2).
Hippocampus Connectivity. CTQ negativelypredicted connectivity from left and right hippocampus
Depression and Anxiety
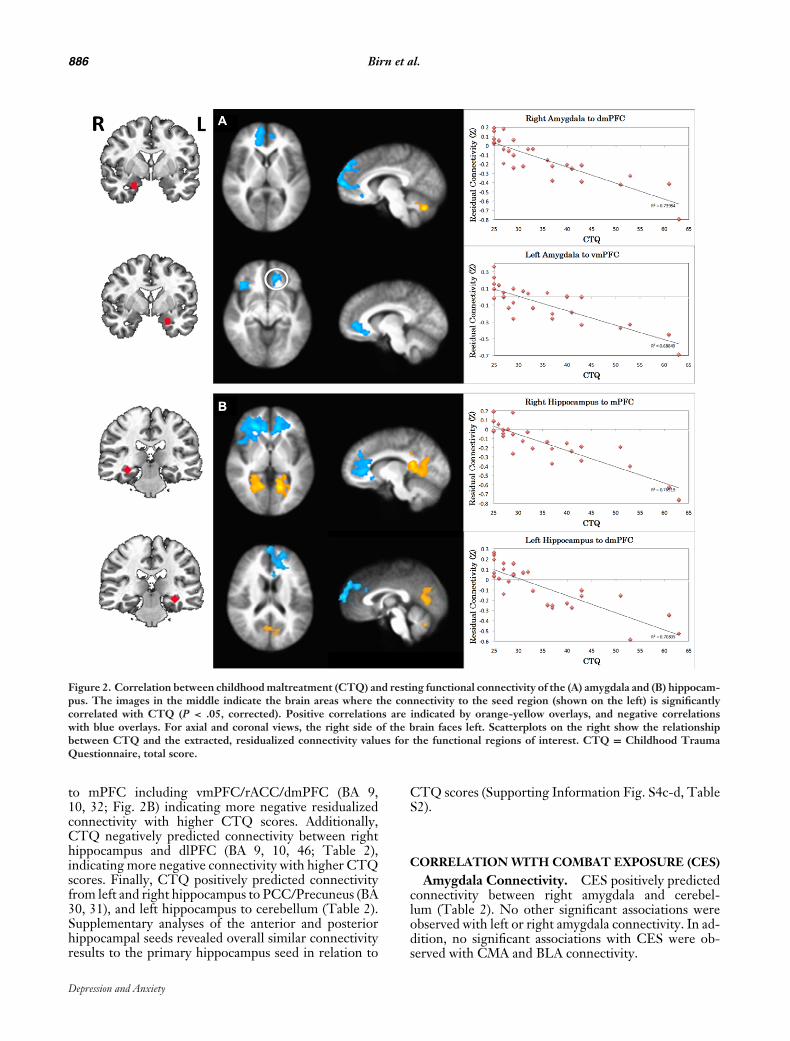
886 Birn et al.
Figure 2. Correlation between childhood maltreatment (CTQ) and resting functional connectivity of the (A) amygdala and (B) hippocam-pus. The images in the middle indicate the brain areas where the connectivity to the seed region (shown on the left) is significantlycorrelated with CTQ (P < .05, corrected). Positive correlations are indicated by orange-yellow overlays, and negative correlationswith blue overlays. For axial and coronal views, the right side of the brain faces left. Scatterplots on the right show the relationshipbetween CTQ and the extracted, residualized connectivity values for the functional regions of interest. CTQ = Childhood TraumaQuestionnaire, total score.
to mPFC including vmPFC/rACC/dmPFC (BA 9,10, 32; Fig. 2B) indicating more negative residualizedconnectivity with higher CTQ scores. Additionally,CTQ negatively predicted connectivity between righthippocampus and dlPFC (BA 9, 10, 46; Table 2),indicating more negative connectivity with higher CTQscores. Finally, CTQ positively predicted connectivityfrom left and right hippocampus to PCC/Precuneus (BA30, 31), and left hippocampus to cerebellum (Table 2).Supplementary analyses of the anterior and posteriorhippocampal seeds revealed overall similar connectivityresults to the primary hippocampus seed in relation to
CTQ scores (Supporting Information Fig. S4c-d, TableS2).
CORRELATION WITH COMBAT EXPOSURE (CES)Amygdala Connectivity. CES positively predicted
connectivity between right amygdala and cerebel-lum (Table 2). No other significant associations wereobserved with left or right amygdala connectivity. In ad-dition, no significant associations with CES were ob-served with CMA and BLA connectivity.
Depression and Anxiety
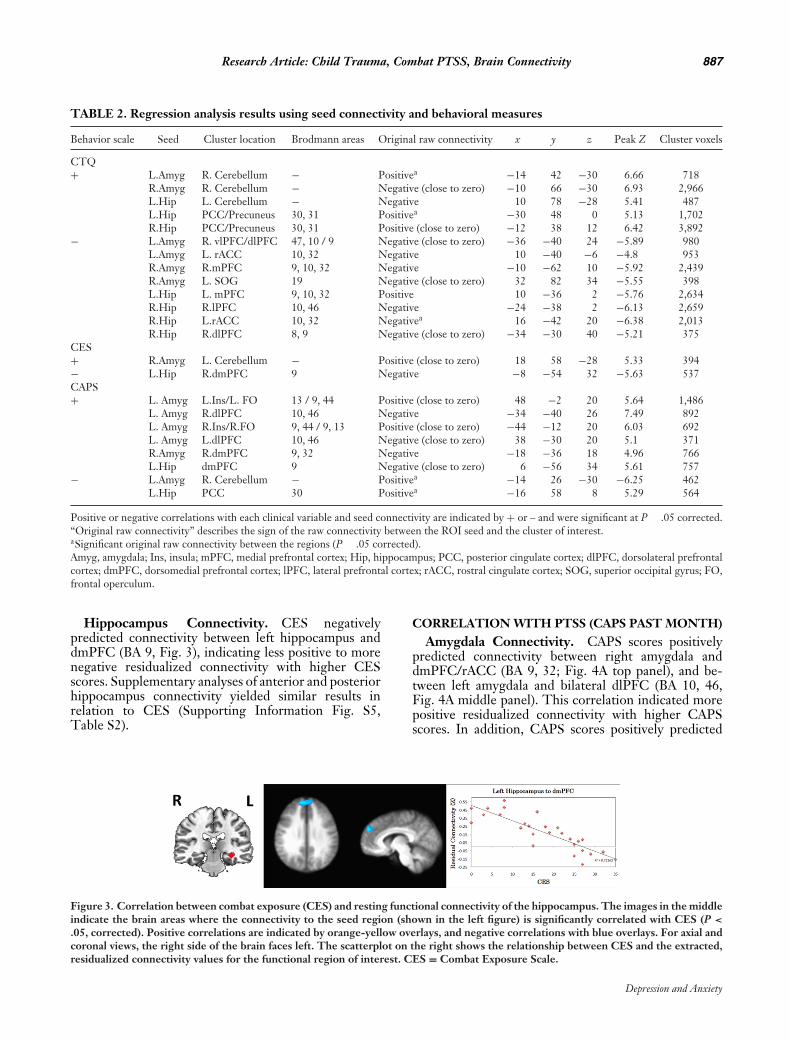
Research Article: Child Trauma, Combat PTSS, Brain Connectivity 887
TABLE 2. Regression analysis results using seed connectivity and behavioral measures
Behavior scale Seed Cluster location Brodmann areas Original raw connectivity x y z Peak Z Cluster voxels
CTQ+ L.Amyg R. Cerebellum − Positivea −14 42 −30 6.66 718
R.Amyg R. Cerebellum − Negative (close to zero) −10 66 −30 6.93 2,966L.Hip L. Cerebellum − Negative 10 78 −28 5.41 487L.Hip PCC/Precuneus 30, 31 Positivea −30 48 0 5.13 1,702R.Hip PCC/Precuneus 30, 31 Positive (close to zero) −12 38 12 6.42 3,892
− L.Amyg R. vlPFC/dlPFC 47, 10 / 9 Negative (close to zero) −36 −40 24 −5.89 980L.Amyg L. rACC 10, 32 Negative 10 −40 −6 −4.8 953R.Amyg R.mPFC 9, 10, 32 Negative −10 −62 10 −5.92 2,439R.Amyg L. SOG 19 Negative (close to zero) 32 82 34 −5.55 398L.Hip L. mPFC 9, 10, 32 Positive 10 −36 2 −5.76 2,634R.Hip R.lPFC 10, 46 Negative −24 −38 2 −6.13 2,659R.Hip L.rACC 10, 32 Negativea 16 −42 20 −6.38 2,013R.Hip R.dlPFC 8, 9 Negative (close to zero) −34 −30 40 −5.21 375
CES+ R.Amyg L. Cerebellum − Positive (close to zero) 18 58 −28 5.33 394− L.Hip R.dmPFC 9 Negative −8 −54 32 −5.63 537CAPS+ L. Amyg L.Ins/L. FO 13 / 9, 44 Positive (close to zero) 48 −2 20 5.64 1,486
L. Amyg R.dlPFC 10, 46 Negative −34 −40 26 7.49 892L. Amyg R.Ins/R.FO 9, 44 / 9, 13 Positive (close to zero) −44 −12 20 6.03 692L. Amyg L.dlPFC 10, 46 Negative (close to zero) 38 −30 20 5.1 371R.Amyg R.dmPFC 9, 32 Negative −18 −36 18 4.96 766L.Hip dmPFC 9 Negative (close to zero) 6 −56 34 5.61 757
− L.Amyg R. Cerebellum − Positivea −14 26 −30 −6.25 462L.Hip PCC 30 Positivea −16 58 8 5.29 564
Positive or negative correlations with each clinical variable and seed connectivity are indicated by + or – and were significant at P � .05 corrected.“Original raw connectivity” describes the sign of the raw connectivity between the ROI seed and the cluster of interest.aSignificant original raw connectivity between the regions (P � .05 corrected).Amyg, amygdala; Ins, insula; mPFC, medial prefrontal cortex; Hip, hippocampus; PCC, posterior cingulate cortex; dlPFC, dorsolateral prefrontalcortex; dmPFC, dorsomedial prefrontal cortex; lPFC, lateral prefrontal cortex; rACC, rostral cingulate cortex; SOG, superior occipital gyrus; FO,frontal operculum.
Hippocampus Connectivity. CES negativelypredicted connectivity between left hippocampus anddmPFC (BA 9, Fig. 3), indicating less positive to morenegative residualized connectivity with higher CESscores. Supplementary analyses of anterior and posteriorhippocampus connectivity yielded similar results inrelation to CES (Supporting Information Fig. S5,Table S2).
CORRELATION WITH PTSS (CAPS PAST MONTH)Amygdala Connectivity. CAPS scores positively
predicted connectivity between right amygdala anddmPFC/rACC (BA 9, 32; Fig. 4A top panel), and be-tween left amygdala and bilateral dlPFC (BA 10, 46,Fig. 4A middle panel). This correlation indicated morepositive residualized connectivity with higher CAPSscores. In addition, CAPS scores positively predicted
Figure 3. Correlation between combat exposure (CES) and resting functional connectivity of the hippocampus. The images in the middleindicate the brain areas where the connectivity to the seed region (shown in the left figure) is significantly correlated with CES (P <
.05, corrected). Positive correlations are indicated by orange-yellow overlays, and negative correlations with blue overlays. For axial andcoronal views, the right side of the brain faces left. The scatterplot on the right shows the relationship between CES and the extracted,residualized connectivity values for the functional region of interest. CES = Combat Exposure Scale.
Depression and Anxiety

888 Birn et al.
Figure 4. Correlation between posttraumatic stress symptoms (PTSS) and resting functional connectivity of the (A) amygdala and (B)hippocampus. The images in the middle indicate the brain areas where the connectivity to the seed region (shown on the left) issignificantly correlated with PTSS (P < .05, corrected). Positive correlations are indicated by orange-yellow overlays, and negativecorrelations with blue overlays. For axial and coronal views, the right side of the brain faces left. Scatterplots on the right show therelationship between CAPS and the extracted, residualized connectivity values for the functional regions of interest. PTSS were basedon the past-month total score of the Clinician-Administered PTSD Scale (CAPS).
connectivity between left amygdala and bilateral an-terior insula (BA 13; Fig. 4A bottom panel), indicat-ing more positive residualized connectivity with higherCAPS scores. Finally, CAPS scores negatively predictedconnectivity between left amygdala and cerebellum (Ta-ble 2). Supplementary analyses of the CMA and BLAyielded similar connectivity results in association withCAPS scores, with no clear differences between the am-gydala subdivisions (Supporting Information Fig. S6a-c,Table S2).
Hippocampus Connectivity. CAPS scores posi-tively predicted connectivity to dmPFC (BA 9, Fig. 4B)indicating more positive residualized connectivity withhigher CAPS scores. CAPS scores negatively predictedconnectivity to PCC (BA 30, Table 2). Supplementaryanalyses of anterior and posterior hippocampus con-nectivity yielded similar results in relation to CAPS.However, the association with anterior hippocampusconnectivity was more widespread in the mPFC withextension into dlPFC (Supporting Information Fig. S6d,Table S2).
Depression and Anxiety

Research Article: Child Trauma, Combat PTSS, Brain Connectivity 889
COVARIATION FOR DEPRESSIVE SYMPTOMSAND TBI HISTORY
To examine potentially confounding effects of depres-sive symptoms and TBI history in the current results, weconducted a secondary analysis of our primary variablesof interest and resting-state connectivity, with depressivesymptoms (BDI) and lifetime TBI history (present or ab-sent) included as an additional covariates in our model.Inclusion of these variables did not change the patternof results with our primary variables.
DISCUSSIONIn this study, we investigated resting-state functional
connectivity of the amygdala and hippocampus, two keyfear-network nodes, in young combat veterans with awide range of PTSD symptoms. Importantly, we ex-amined the relation of this connectivity not only toPTSD symptom severity but also to both adult and childtrauma exposure within a single model. This representsan important step in identifying potential mechanisms bywhich developmental trauma exposure may lead to adultPTSD symptoms, and in turn which circuitry changesdistinguish PTSD symptoms from trauma exposure it-self.
Amygdala. We found that childhood maltreatmentnegatively predicted connectivity between the amygdalaand vmPFC, as well as connectivity between the amyg-dala and lateral PFC regions. The vmPFC and its rat ho-mologue IL are notable for their role in the recall of fearextinction and downregulation of amygdala-dependentfear responses.[4] The vmPFC is recruited during the re-call of fear extinction in healthy individuals,[39, 40] andthe magnitude of activation is correlated with extinctionretention.[41] In addition, resting amygdala metabolismnegatively predicts vmPFC activation during extinctionrecall,[42] which suggests that communication betweenthese regions plays an important role in human fear ex-tinction. Consistent with this regulatory role, greaterfunctional connectivity between amygdala and vmPFCis linked to lower anxiety levels during presentation offearful faces[43] and at rest[44] in healthy individuals. Inagreement with the current results, our prior work re-vealed that early-life stress and childhood maltreatmentare associated with weaker amygdala–vmPFC connectiv-ity, which in turn predicts the development of internal-izing symptoms.[14, 15] Thus, our data support a model inwhich childhood maltreatment experiences may weakenthis key automatic fear-regulatory circuit, which maycontribute risk toward the development of PTSDfollowing subsequent trauma in adulthood. At the sametime, a weakening of amygdala–vmPFC connectivity in-duced by childhood maltreatment does not appear suf-ficient in and of itself to account for the emergence ofPTSD symptoms, which instead are associated with aloss of amygdala connectivity to more dorsal prefrontalareas as described below.
In contrast to childhood maltreatment exposure,PTSS predicted a loss of anticorrelation betweenthe amygdala and dorsal PFC regions includingdmPFC/rACC and dlPFC. These dorsal PFC areasare notable for their role in effortful top-down emo-tion regulation, and appear to exert their regulatoryinfluence on the amygdala via the vmPFC.[11, 45] Inhealthy individuals, dorsal PFC regions are anticorre-lated with the amygdala during task and rest.[44, 46, 47]
Furthermore, successful emotion regulation is associatedwith greater amygdala–dm/dlPFC anticorrelation dur-ing task,[48] and higher anxiety levels are associated witha loss of resting amygdala–dmPFC anticorrelation.[44]
PTSD has also been characterized by reduced amygdala–dmPFC/rACC anticorrelation both during symptomprovocation[47] and at rest.[7, 9] Our results are consis-tent with these studies and, when examined in relationto raw (nonresidualized) amygdala–dmPFC connectiv-ity, suggest a loss of anticorrelation and a switch to pos-itive connectivity with higher CAPS scores (see Sup-porting Information Fig. S1). Within this framework,childhood maltreatment may create a vulnerable brainsubstrate by weakening amygdala–vmPFC connectiv-ity, yet PTSD symptoms may only emerge with a sub-sequent loss of amygdala–dorsal PFC anticorrelation.Interestingly, childhood maltreatment negatively pre-dicted amygdala–dorsal PFC connectivity (i.e., greateranticorrelation). This could be attributable to sharedvariance with vmPFC–amygdala connectivity (also nega-tively predicted by CTQ). Alternatively, our results sug-gest that greater amygdala–dorsal PFC anticorrelationrepresents a compensatory brain mechanism employ-ing effortful emotion regulation following childhoodmaltreatment, the failure of which is associated withthe emergence of PTSD symptoms. Ultimately, lon-gitudinal studies examining both childhood and adulttrauma exposure and adult PTSS, along with causal-ity modeling[49] will be needed to tease apart thesepossibilities.
PTSS also positively correlated with restingamygdala–anterior insula connectivity. The ante-rior insula is involved in the representation of visceralemotional states[50] and has strong structural andfunctional connectivity with the amygdala.[51, 52] Asdetailed in previous studies, greater connectivity at restbetween these areas may represent heightened integra-tion of threat and interoceptive processing that couldunderlie hyperarousal and re-experiencing phenomenaof PTSD.[6, 7] As such, these findings remain consistentwith prior resting-state studies of PTSD and suggestspecificity of changes in this circuit to PTSD symptoms.
Hippocampus. We found that childhood maltreat-ment negatively predicted connectivity between the hip-pocampus and vmPFC, as well as dm/dlPFC. Combatexposure also negatively predicted hippocampus–PFCconnectivity, though this was localized to the dmPFC.The hippocampus is known to play an important rolein the regulation of fear by contextually limiting fearresponses via connections to both the amygdala and
Depression and Anxiety

890 Birn et al.
vmPFC.[5] In rats, chronic stress impairs hippocampus-mPFC long-term potentiation, which is required forthe proper gating of conditioned fear.[53, 54] In hu-mans, both the vmPFC and hippocampus are recruitedduring the recall of fear extinction, for example,[40, 41] insupport of their co-ordinated role in gating fear. Con-sistent with the current findings, prior work from ourgroup has shown that childhood maltreatment is asso-ciated with weaker hippocampus–vmPFC connectivity,which in turn mediates the development of internal-izing symptoms.[15] Within this framework, our find-ings suggest that childhood maltreatment in particularmay weaken the contextual regulation of fear by im-pairing communication between the hippocampus andvmPFC, though this would require further testing ina fear-conditioning paradigm. Of note, a longitudinalstudy revealed that adult trauma (combat exposure) ac-tually increases hippocampus–vmPFC connectivity toemotional stimuli, and failure to increase this connec-tivity was associated with the development of PTSDsymptoms.[18] Our results suggest that childhood mal-treatment exposure could compromise the ability of thiscircuit to adaptively upregulate following adult trauma,placing an individual at risk for developing fear-relatedpsychopathology as an adult.
In contrast to childhood maltreatment and similar tothe amygdala findings, PTSS predicted a loss of anti-correlation between the hippocampus and dorsal PFC,specifically to dmPFC. In healthy individuals, the hip-pocampus and dmPFC show inverse patterns of acti-vation in signaling safety versus threat (i.e., hippocam-pus activation and dmPFC deactivation to safety cues,and vice versa to conditioned threat).[55] Furthermore,PTSD subjects display hippocampus deactivation anddACC activation to extinguished threat cues,[3] suggest-ing a reversal of the pattern seen in healthy individuals.In light of this, the loss of resting dmPFC–hippocampusanticorrelation (and switch to positive connectivity) as-sociated with PTSS in the current study could reflectimpaired communication between the hippocampus anddmPFC in predicting safety versus threat. This suggeststhat the ability of the hippocampus to contextually reg-ulate fear is further compromised in PTSD, involving adisruption in communication with dorsal prefrontal ar-eas. Like the amygdala connectivity findings, childhoodmaltreatment may create a vulnerable brain substrate byreducing vmPFC–hippocampal connectivity; however,PTSD symptoms may only emerge with a subsequentloss of dmPFC–hippocampus anticorrelation. Interest-ingly, both childhood maltreatment and combat expo-sure negatively predicted dorsal PFC–hippocampus con-nectivity (i.e., greater anticorrelation), suggesting thesemay be compensatory attempts that are lost with the de-velopment of PTSS. As with the amygdala connectivityfindings, longitudinal studies will ultimately be needed,along with causality modeling, to tease apart these pos-sibilities.
Of note, we did not observe any marked dissociationsof amygdala or hippocampal subdivisions in our results,
in contrast to prior studies.[9, 10] The reasons for this arenot entirely clear but likely reflect different experimen-tal designs (within-subjects vs. between-group analysis),analytic approaches (e.g., use of global signal regressionand a priori search regions[10]), and the inclusion of theadditional trauma variables in our study. Our study sam-ple also differs from these prior studies in that we did notallow any current psychiatric medication usage. Furtherwork will be needed, likely with larger sample sizes, toexamine whether there are consistent and dissociable rs-FC findings among amygdala and hippocampal subdivi-sions in PTSD.
The results of this study underscore the utility of us-ing resting-state fMRI to detect the neurobiological cor-relates of trauma exposure and PTSS. However, theseresults are not without limitations. First, these findingsare correlational only and do not prove causation. It isthus difficult to tease out whether observed changes rep-resent preexisting vulnerabilities, state-related changes,or compensatory changes. Second, these findings maynot generalize to other populations (particularly womengiven the all-male sample), trauma types, or to indi-viduals with more severe PTSS. In addition, resting-state connectivity may not necessarily equate to task-related connectivity, an area which requires furtherstudy. Third, although subjects were free of current al-cohol or substance abuse, this study did not have mea-sures of cumulative, lifelong alcohol use, which couldaffect the current results. However, PTSD and non-PTSD subjects had similar frequencies of past alcoholabuse/dependence that makes this less likely. Finally,this is a moderate number of subjects with results con-sidered preliminary, and would merit replication in alarger sample and using longer duration acquisitions. Al-though 5 min of resting-state data have been used suc-cessfully in other studies to characterize differences infunctional connectivity (e.g.,[56]) and have moderate test–retest reliability,[57] more recent studies suggest that thereliability of these results would improve even furtherwith longer duration acquisitions.[58]
CONCLUSIONThe results from this study suggest that childhood
trauma, combat exposure, and combat PTSS differen-tially predict resting-state connectivity of the amygdalaand hippocampus, two key nodes in the fear network.Childhood trauma in particular may create a vulnerablebrain substrate by weakening ventromedial prefrontal–subcortical pathways involved in the extinction and con-textual gating of fear, yet may also be accompaniedby compensatory strengthening of dorsal prefrontal–subcortical circuits involved in effortful emotionregulation. PTSS, in turn, may emerge upon this back-ground with the loss of anticorrelation between dor-sal prefronto-subcortical pathways, as well as greaterfear/interoceptive processing involving the amygdalaand insula. These findings highlight the importance ofexamining developmental trauma exposure and adult
Depression and Anxiety

Research Article: Child Trauma, Combat PTSS, Brain Connectivity 891
PTSS within a single model. They also represent animportant step in identifying potential mechanisms bywhich developmental trauma exposure may lead to adultPTSD, and which mechanisms are associated with theemergence of PTSD symptoms themselves.
Acknowledgments. The authors would like tothank Benjamin Paul, Ashlee Filippone, and RyanStocker for their assistance with participant screening,MRI procedures, and data organization. This work wassupported by the AACAP Pilot Research Award (RJH),Department of Defense Congressionally Directed Med-ical Research Program (PT073961 (A.G.)), and the Na-tional Institute of Mental Health (MH083035 (A.G.),MH076971 (M.L.P.), MH088913 (M.L.P.)).
Conflict of Interest. Dr. Germain has served as aconsultant for Concurrent Technologies Corporation.The other authors report no biomedical financial inter-ests or potential conflicts of interest.
REFERENCES1. Patel R, Spreng RN, Shin LM, Girard TA. Neurocircuitry
models of posttraumatic stress disorder and beyond: a meta-analysis of functional neuroimaging studies. Neurosci BiobehavRev 2012;36(9):2130–2142.
2. Pitman RK, Rasmusson AM, Koenen KC, et al. Biologicalstudies of post-traumatic stress disorder. Nat Rev Neurosci2012;13(11):769–787.
3. Milad MR, Pitman RK, Ellis CB, et al. Neurobiological basis offailure to recall extinction memory in posttraumatic stress disor-der. Biol Psychiatry 2009;66(12):1075–1082.
4. Milad MR, Quirk GJ. Fear extinction as a model for transla-tional neuroscience: ten years of progress. Annu Rev Psychol2012;63:129–1251.
5. Maren S, Phan KL, Liberzon I. The contextual brain: implicationsfor fear conditioning, extinction and psychopathology. Nat RevNeurosci 2013;14(6):417–428.
6. Rabinak CA, Angstadt M, Welsh RC, et al. Altered amygdalaresting-state functional connectivity in post-traumatic stress dis-order. Front Psychiatry 2011;2(62):1–8.
7. Sripada RK, King AP, Garfinkel SN, et al. Altered resting-stateamygdala functional connectivity in men with posttraumatic stressdisorder. J Psychiatry Neurosci 2012;37(2):241–249.
8. Sripada RK, King AP, Welsh RC, et al. Neural dysregulation inposttraumatic stress disorder: evidence for disrupted equilibriumbetween salience and default mode brain networks. PsychosomMed 2012;74(9):904–911.
9. Brown VM, Labar KS, Haswell CC, et al. Altered resting-statefunctional connectivity of basolateral and centromedial amygdalacomplexes in posttraumatic stress disorder. Neuropsychopharma-cology 2014;39(2):361–369.
10. Chen AC, Etkin A. Hippocampal network connectivity and activa-tion differentiates post-traumatic stress disorder from generalizedanxiety disorder. Neuropsychopharmacology 2013;38(10):1889–1898.
11. Phillips ML, Ladouceur CD, Drevets WC. A neural model of vol-untary and automatic emotion regulation: implications for under-standing the pathophysiology and neurodevelopment of bipolardisorder. Mol Psychiatry 2008;13(9):829, 833–857.
12. Brewin CR, Andrews B, Valentine JD. Meta-analysis of risk fac-tors for posttraumatic stress disorder in trauma-exposed adults. JConsult Clin Psychol 2000;68(5):748–766.
13. van der Werff SJA, Pannekoek JN, Veer IM, et al. Resting-statefunctional connectivity in adults with childhood emotional mal-treatment. Psychol Med 2013;43(9):1825–1836.
14. Burghy CA, Stodola DE, Ruttle PL, et al. Developmental path-ways to amygdala-prefrontal function and internalizing symptomsin adolescence. Nat Neurosci 2012;15(12):1736–1741.
15. Herringa RJ, Birn RM, Ruttle PL, et al. Childhood maltreatmentis associated with altered fear circuitry and increased internal-izing symptoms by late adolescence. Proc Natl Acad Sci USA2013;110(47):19119–19124.
16. Wang L, Dai Z, Peng H, et al. Overlapping and segregated resting-state functional connectivity in patients with major depressivedisorder with and without childhood neglect. Hum Brain Mapp2014;35(4):1154–1166.
17. Cisler JM, James GA, Tripathi S, et al. Differential functionalconnectivity within an emotion regulation neural network amongindividuals resilient and susceptible to the depressogenic effects ofearly life stress. Psychol Med 2013;43(03):507–518.
18. Admon R, Lubin G, Stern O, et al. Human vulnerability to stressdepends on amygdala’s predisposition and hippocampal plasticity.Proc Natl Acad Sci USA 2009;106(33):14120–14125.
19. Herringa RJ, Phillips ML, Fournier JC, Kronhaus DM, GermainA. Childhood and adult trauma both correlate with dorsal anteriorcingulate activation to threat in combat veterans. Psychol Med2013;43(7):1533–1542.
20. Blake DD, Weathers FW, Nagy LM, et al. The develop-ment of a Clinician-Administered PTSD Scale. J Trauma Stress1995;8(1):75–90.
21. Weathers FW, Keane TM, Davidson JR. Clinician-AdministeredPTSD Scale: a review of the first ten years of research. DepressAnxiety 2001;13(3):132–1356.
22. First MB, Spitzer RL, Gibbon M, Williams JBW. StructuredClinical Interview for DSM-IV-TR Axis I Disorders, ResearchVersion, Patient Edition. (SCID-I/P). New York: Biometrics Re-search, New York State Psychiatric Institute; 2002.
23. Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J. An inven-tory for measuring depression. Arch Gen Psychiatry 1961;4:561–571.
24. Keane T, Fairbank J, Caddell J, Zimering R, Taylor K, MoraC. Clinical evaluation of a measure to assess combat exposure.Psychol Assess 1989;1(1):53–55.
25. Scher CD, Stein MB, Asmundson GJ, McCreary DR, FordeDR. The childhood trauma questionnaire in a community sam-ple: psychometric properties and normative data. J Trauma Stress2001;14(4):843–857.
26. Blair JR, Spreen O. Predicting premorbid IQ: a revision ofthe national adult reading test. Clin Neuropsychol 1989;3(2):129–136.
27. Zhu S, Fang Z, Hu S, Wang Z, Rao H. Resting state brain functionanalysis using concurrent BOLD in ASL perfusion fMRI. PLoSOne 2013;8(6):e65884.
28. Cox RW. AFNI: software for analysis and visualization of func-tional magnetic resonance neuroimages. Comput Biomed Res1996;29(3):162–173.
29. Wong EC, Buxton RB, Frank LR. Implementation of quantitativeperfusion imaging techniques for functional brain mapping usingpulsed arterial spin labeling. NMR Biomed 1997;10(4–5):237–249.
30. Jo HJ, Gotts SJ, Reynolds RC, et al. Effective preprocessing pro-cedures virtually eliminate distance-dependent motion artifacts inresting state FMRI. J Appl Math 2013;2013.
Depression and Anxiety

892 Birn et al.
31. Smith SM, Jenkinson M, Woolrich MW, et al. Advances in func-tional and structural MR image analysis and implementation asFSL. Neuroimage 2004;23 Suppl 1:S208–219.
32. Woolrich MW, Jbabdi S, Patenaude B, et al. Bayesian analysis ofneuroimaging data in FSL. Neuroimage 2009;45(1 Suppl):S173–S186.
33. Zhang Y, Brady M, Smith S. Segmentation of brain MR im-ages through a hidden Markov random field model and theexpectation-maximization algorithm. IEEE Trans Med Imaging2001;20(1):45–57.
34. Lancaster JL, Rainey LH, Summerlin JL, et al. Automated label-ing of the human brain: a preliminary report on the developmentand evaluation of a forward-transform method. Hum Brain Mapp1997;5(4):238–242.
35. Lancaster JL, Woldorff MG, Parsons LM, et al. Automated Ta-lairach Atlas labels for functional brain mapping. Hum Brain Mapp2000;10(3):120–131.
36. Forman SD, Cohen JD, Fitzgerald M, Eddy WF, Mintun MA,Noll DC. Improved assessment of significant activation in func-tional magnetic resonance imaging (fMRI): use of a cluster-sizethreshold. Magn Reson Med 1995;33(5):636–647.
37. Philip NS, Sweet LH, Tyrka AR, Price LH, Bloom RF, Carpen-ter LL. Decreased default network connectivity is associated withearly life stress in medication-free healthy adults. Eur Neuropsy-chopharmacol 2013;23(1):24–32.
38. Bluhm RL, Williamson PC, Osuch EA, et al. Alterations in defaultnetwork connectivity in posttraumatic stress disorder related toearly-life trauma. J Psychiatry Neurosci 2009;34(3):187–194.
39. Phelps EA, Delgado MR, Nearing KI, LeDoux JE. Extinctionlearning in humans: role of the amygdala and vmPFC. Neuron2004;43(6):897–905.
40. Kalisch R, Korenfeld E, Stephan KE, Weiskopf N, Seymour B,Dolan RJ. Context-dependent human extinction memory is me-diated by a ventromedial prefrontal and hippocampal network. JNeurosci 2006;26(37):9503–9511.
41. Milad MR, Wright CI, Orr SP, Pitman RK, Quirk GJ, RauchSL. Recall of fear extinction in humans activates the ventrome-dial prefrontal cortex and hippocampus in concert. Biol Psychiatry2007;62(5):446–454.
42. Linnman C, Zeidan MA, Furtak SC, Pitman RK, Quirk GJ, MiladMR. Resting amygdala and medial prefrontal metabolism predictsfunctional activation of the fear extinction circuit. Am J Psychiatry2012;169(4):415–423.
43. Pezawas L, Meyer-Lindenberg A, Drabant EM, et al. 5-HTTLPRpolymorphism impacts human cingulate-amygdala interactions: agenetic susceptibility mechanism for depression. Nat Neurosci2005;8(6):828–834.
44. Kim MJ, Gee DG, Loucks RA, Davis FC, Whalen PJ. Anxi-ety dissociates dorsal and ventral medial prefrontal cortex func-
tional connectivity with the amygdala at rest. Cereb Cortex2011;21(7):1667–1673.
45. Ghashghaei HT, Hilgetag CC, Barbas H. Sequence of infor-mation processing for emotions based on the anatomic dia-logue between prefrontal cortex and amygdala. Neuroimage2007;34(3):905–923.
46. Roy AK, Shehzad Z, Margulies DS, et al. Functional connectivityof the human amygdala using resting state fMRI. Neuroimage2009;45(2):614–626.
47. Gilboa A, Shalev AY, Laor L, et al. Functional connectivity ofthe prefrontal cortex and the amygdala in posttraumatic stressdisorder. Biol Psychiatry 2004;55(3):263–272.
48. Lee H, Heller AS, van Reekum CM, Nelson B, Davidson RJ.Amygdala-prefrontal coupling underlies individual differences inemotion regulation. Neuroimage 2012;62(3):1575–1581.
49. Sridharan D, Levitin DJ, Menon V. A critical role forthe right fronto-insular cortex in switching between central-executive and default-mode networks. Proc Natl Acad Sci USA2008;105(34):12569–12574.
50. Garfinkel SN, Critchley HD. Interoception, emotion and brain:new insights link internal physiology to social behaviour. Com-mentary on: “Anterior insular cortex mediates bodily sensibilityand social anxiety” by Terasawa et al. (2012). Soc Cogn AffectNeurosci 2013;8(3):231–234.
51. Reynolds SM, Zahm DS. Specificity in the projections of pre-frontal and insular cortex to ventral striatopallidum and the ex-tended amygdala. J Neurosci 2005;25(50):11757–11767.
52. Salvador R, Martınez A, Pomarol-Clotet E, et al. A simple viewof the brain through a frequency-specific functional connectivitymeasure. Neuroimage 2008;39(1):279–289.
53. Garcia R, Spennato G, Nilsson-Todd L, Moreau J-L, DeschauxO. Hippocampal low-frequency stimulation and chronic mildstress similarly disrupt fear extinction memory in rats. NeurobiolLearn Mem. 2008;89(4):560–566.
54. Sotres-Bayon F, Sierra-Mercado D, Pardilla-Delgado E, QuirkGJ. Gating of fear in prelimbic cortex by hippocampal and amyg-dala inputs. Neuron 2012;76(4):804–812.
55. Lissek S, Bradford DE, Alvarez RP, et al. Neural substrates ofclassically conditioned fear-generalization in humans: a paramet-ric fMRI study. Soc Cogn Affect Neurosci 2013.
56. Greicius MD, Flores BH, Menon V, et al. Resting-state functionalconnectivity in major depression: abnormally increased contribu-tions from subgenual cingulate cortex and thalamus. Biol Psychi-atry 2007;62(5):429–437.
57. Shehzad Z, Kelly AMC, Reiss PT, et al. The resting brain: un-constrained yet reliable. Cereb Cortex 2009;19(10):2209–2229.
58. Birn RM, Molloy EK, Patriat R, et al. The effect of scan length onthe reliability of resting-state fMRI connectivity estimates. Neu-roimage 2013;83:550–558.
Depression and Anxiety