determining the specificity of pepsin for proteolytic …767/fulltext.pdf · determining the...
TRANSCRIPT

DETERMINING THE SPECIFICITY OF PEPSIN
FOR PROTEOLYTIC DIGESTION
A thesis presented
by
Melissa H. Palashoff
to The Department of Chemistry and Chemical Biology
in partial fulfillment of the requirements for the degree of Master of Science
in the field of
Chemistry
Northeastern University Boston, Massachusetts
August 2008
1

DETERMINING THE SPECIFICITY OF PEPSIN
FOR PROTEOLYTIC DIGESTION
by
Melissa H. Palashoff
ABSTRACT OF THESIS
Submitted in partial fulfillment of the requirements for the degree of Master of Science in Chemistry and Chemical Biology
in the Graduate School of Arts and Sciences of Northeastern University, August 2008
2

ABSTRACT
Pepsin is an aspartic acid protease that is commonly found in the stomach of
many organisms. Porcine pepsin is the most studied and is fully active at pH 1.9 but
inactive above pH ~7. Pepsin is known to have limited specificity and there are only
general rules about its cleavage preferences.
To further define rules regarding pepsin specificity, a database was constructed
consisting of 40 proteins and 1344 peptide cleavages from the literature. Contemporary
scientific literature was searched for all publications that involve pepsin digestion and
mass spectrometry at pH 2.5-2.7. Peptide data for 40 proteins were extracted and
combined to create a map of pepsin cleavage specificity. The frequency of cleavage for
each protein was normalized based on how many times that specific combination of
residues occurred in the protein sequence.
In addition to the literature search, nine proteins along with E.coli whole cell
lysate were digested at pH 1.0, 2.5 and 4.0. The proteins were analyzed with online
pepsin digestion using an immobilized pepsin column and UPLC/ESI-MSE. The peptides
and their fragments were identified with a combination of MSE, software analysis, and
manual inspection.
The analysis of the data indicated that pepsin maintains limited cleavage
preferences. At pH 2.5, pepsin will cleave preferentially after most bulky, hydrophobic
amino acids such as leucine and phenylalanine. Additionally, the residues that most often
occur immediately following the cleaved peptide bond are tryptophan and tyrosine. It has
also been shown that pepsin will rarely cleave at proline and histidine. Analysis
performed at pH 1.0 and 4.0 yielded similar results.
3

ACKNOLEDGEMENTS
I would like to begin by thanking my advisor Dr. John R. Engen for all of his help
in making me a better scientist. Without his guidance and advice this research would not
have been possible. I am also grateful to my other committee members, Dr. Mary Jo
Ondrechen and Dr. Paul Vouros, for helping to make this thesis the best that it could be.
I would like to sincerely thank everyone in Dr. Engen’s lab for their continuous
support over the past year. To my labmates, Dr. Thomas Wales, Dr. Roxana Iacob,
Christopher Morgan, Sean Marcsisin, Damian Houde and Susan Fang, for providing a
wonderful environment for me to work and learn in.
Finally, I would like to dedicate this to my family and friends for their constant
encouragement during the past five years. I would especially like to thank my parents,
William and Patricia, and my brother Joshua for everything they have done to support
me, both morally and materially, during my college career. Last, but certainly not least,
to Eddie for always being there to support me during the good days and the bad, thank
you.
4

TABLE OF CONTENTS
ABSTRACT ……………………………………………………………………………………….
TABLE OF CONTENTS ………………………………………………………………………….
LIST OF FIGURES ………………………………………………………………………………..
LIST OF TABLES ………………………………………………………………………………...
LIST OF ABBREVIATIONS ……………………………………………………………………
CHAPTER ONE: INTRODUCTION AND BACKGROUND TO ASPARTIC ACID
PROTEASES AND PEPSIN……………………………………………………………..
1.1 Aspartic Acid Proteases ……………………………………………………………...
1.2 Catalytic Mechanism of Aspartic Proteases ………………………………………….
1.3 Primary Structure of the Pepsin-like Family …………………………………………
1.4 Pepsin …………………………………………………………………………………
1.4.1 History of pepsin ……………………………………………………………
1.4.2 Activation of pepsin ………………………………………………………...
1.4.3 Pepsin crystal structure ……………………………………………………..
1.4.4 Activity of pepsin …………………………………………………………..
1.4.5 Pepsin and proteomics ……………………………………………………...
1.5 Research Objectives ………………………………………………………………….
1.6 References ……………………………………………………………………………
CHAPTER TWO: LITERATURE SEARCH ……………………………………………………
2.1 Introduction …………………………………………………………………………..
2.2 Materials and Methods ……………………………………………………………….
2.3 Construction of Cleavage Database …………………………………………………..
3
5
8
10
11
13
13
15
17
18
18
18
21
23
23
24
25
29
29
30
30
5

2.4 Data Normalization …………………………………………………………………..
2.5 Cleavage Data Map …………………………………………………………………..
2.6 Revised Cleavage Data Map …………………………………………………………
2.7 Summary of Literature Research …………………………………………………….
2.8 References ……………………………………………………………………………
CHAPTER 3: EXPERIMENTAL DETERMINATION OF PEPSIN SPECIFICITY AND THE
EFFECTS OF pH ………………………………………………………………………...
3.1 Introduction …………………………………………………………………………..
3.2 Instrumentation ……………………………………………………………………….
3.2.1 UPLC and online pepsin digestion …………………………………………
3.2.2 Mass Spectrometry …………………………………………………………
3.3 Materials and Methods ……………………………………………………………….
3.3.1 Protein sample analysis …………………………………………………….
3.3.1.1 Protein sample preparation ……………………………………….
3.3.1.2 UPLC methods …………………………………………………...
3.3.1.3 Mass analysis ……………………………………………………..
3.3.2 E.coli sample analysis ……………………………………………………...
3.3.2.1 E.coli sample preparation ………………………………………...
3.3.2.2 UPLC analysis ……………………………………………………
3.3.2.3 Mass analysis ……………………………………………………..
3.4 Data Analysis …………………………………………………………………………
3.4.1 Software processing ………………………………………………………..
3.4.2 Peptide analysis …………………………………………………………….
34
36
41
44
44
78
78
78
79
81
83
83
83
85
87
87
87
88
88
88
88
89
6

3.5 Results ………………………………………………………………………………..
3.6 References …………………………………………………………………………...
CHAPTER 4: PERSPECTIVES AND FUTURE DIRECTIONS ……………………………...
4.1 Discussion and Conclusions ………………………………………………………...
4.1.1 Literature search …………………………………………………………..
4.1.2 Experimental research …………………………………………………….
4.1.3 Research on pepsin specificity ……………………………………………
4.2 Future Directions ……………………………………………………………………
92
100
141
141
141
141
142
142
7

LIST OF FIGURES
Figure 1.1 Aspartic proteases family tree ………………………………………………………...
Figure 1.2 The aspartic protease catalytic mechanism …………………………………………...
Figure 1.3 Sequence alignment of porcine pepsinogen and porcine pepsin ……………………...
Figure 1.4 Crystal structure of human pepsin ……………………………………………………
Figure 2.1 Example of a peptic digest map ………………………………………………………
Figure 2.2 Example of cleavage nomenclature …………………………………………………...
Figure 2.3 Equation used for data normalization and example calculation ……………………...
Figure 2.4 Cleavage data map ……………………………………………………………………
Figure 2.5 Cleavage data map with probability defined as a percentage ………………………...
Figure 3.1 Schematic of online pepsin digestion …………………………………………………
Figure 3.2 Schematic of the operation of MSE …………………………………………………...
Figure 3.3 Example MS/MS data of hemoglobin ………………………………………………...
Figure 3.4 Example MS/MS data of myoglobin ………………………………………………….
Figure 3.5 pH 2.5 peptic digest map of Abl ……………………………………………………..
Figure 3.6 pH 2.5 peptic digest map of Albumin ……………………………………………….
Figure 3.7 pH 2.5 peptic digest map of Aldolase ……………………………………………….
Figure 3.8 pH 2.5 peptic digest map of Amyloglucosidase ……………………………………...
Figure 3.9 pH 2.5 peptic digest map of β-Lactalbumin ………………………………………....
Figure 3.10 pH 2.5 peptic digest map of Hemoglobin …………………………………………..
Figure 3.11 pH 2.5 peptic digest map of Myoglobin ……………………………………………
Figure 3.12 pH 2.5 peptic digest map of Nef ……………………………………………………
Figure 3.13 pH 2.5 peptic digest map of Ubiquitin ……………………………………………...
14
16
19
22
31
33
38
40
42
80
82
90
91
102
103
104
105
106
107
108
109
110
8

Figure 3.14 Cleavage data map pH 1.0 …………………………………………………………...
Figure 3.15 Cleavage data map pH 2.5 …………………………………………………………...
Figure 3.16 Cleavage data map pH 4.0 …………………………………………………………...
Figure 3.17 Cleavage data map with probability defined as a percentage, pH 1.0 ……………….
Figure 3.18 Cleavage data map with probability defined as a percentage, pH 2.5 ……………….
Figure 3.19 Cleavage data map with probability defined as a percentage, pH 4.0 ……………….
93
94
95
97
98
99
9

LIST OF TABLES
Table 2.1 Literature search results ………………………………………………………………..
Table 2.2 Cleavage database ……………………………………………………………………...
Table 2.3 Sum of cleavages between two residues ……………………………………………….
Table 2.4 Possible cleavages between two residues ……………………………………………...
Table 2.5 Normalized cleavage data ……………………………………………………………...
Table 2.6 Cleavage data with probability defined as a percentage ……………………………….
Table 3.1 Proteins used for digestion …………………………………………………………….
Table 3.2 Auxiliary solvents ……………………………………………………………………..
Table 3.3 pH 1.0 peptides ……………………………………………………………………….
Table 3.4 pH 4.0 peptides ……………………………………………………………………….
Table 3.5 pH 2.5 E.coli peptides ………………………………………………………………...
Table 3.6 Normalized pH 1.0 cleavage data ……………………………………………………...
Table 3.7 Normalized pH 2.5 cleavage data ……………………………………………………...
Table 3.8 Normalized pH 4.0 cleavage data ……………………………………………………...
Table 3.9 Normalized pH 1.0 cleavage data defined as a percentage …………………………….
Table 3.10 Normalized pH 2.5 cleavage data defined as a percentage …………………………...
Table 3.11 Normalized pH 4.0 cleavage data defined as a percentage …………………………...
32
51
35
37
39
43
84
86
111
121
125
93
94
95
97
98
99
10

LIST OF ABBREVIATIONS
ACN acetonitrile
DTT dithiothreitol
E.coli Escherichia coli
ESI electrospray ionization
eV electron volts
FA formic acid
fmol femtomole
HCl hydrochloric acid
Hpb hydrophobic residue
HPLC high performance liquid chromatography
kDa kilo-Dalton
kV kilovolts
L liter
LC liquid chromatography
M molar
min minute
μL microliters
μM micromolar
mM millimolar
mm millimeter
MS mass spectrometry
MS/MS tandem mass spectrometry
11

MW molecular weight
PDB protein data bank
pmol picomoles
UPLC ultra performance liquid chromatography
V volts
12

CHAPTER 1
INTRODUCTION AND BACKGROUND
TO ASPARTIC ACID PROTEASES AND PEPSIN
1.1 Aspartic Acid Proteases
Aspartic acid proteases are a large class of enzymes that are widely distributed.
They are found in a multitude of organisms such as vertebrates, retroviruses, fungi and
plants (Davies 1990). Characteristics shared by aspartic proteases are that they have an
optimum pH in the acid range and are inhibited by pepstatin (Fruton 1976). The
MEROPS database (Rawlings, Morton et al. 2008) has classified the aspartic proteases
into many different families. However, this class of enzymes is believed to have a
conserved segment of residues (-Asp-Thr-Gly-) in its primary structure (Dunn 2002).
Taking this into account, aspartic proteases can be split up into five main families (Figure
1.1). The largest of these groups is the pepsin-like family which contains about 70
members. The retroviral family contains 45 members, the cauliflower mosaic virus
family contains six members, the human spumaretroviral family contains one member
and the copia transposon family contains six members (Dunn 2001; Rawlings, Morton et
al. 2008).
Due to the fact that acid proteases are found in so many different organisms they
tend to perform a variety of functions. The most studied of these aspartic proteases
involve digestion (pepsin, gastricsin, chymosin, etc.) and protein degradation (cathepsin
D). In fungi acid proteases play an important role in sporulation. Retroviral proteases
cleave retroviruses during the activation of the virus (Dickson 1984).
13

Aspartic Proteases
Retroviral Proteases
Copia Transposons
Pepsin APepsin B
Nepenthesin
HIV-1 retropepsinHIV-2 retropepsin
Cauliflower Mosaic Viruses
Spumaretroviral
Bacilliform virus
Pepsin-like
GastricsinRenin
ChymosinCathepsin DCathepsin E
RhitopuspepsinMucorpepsin
CandidapepsinEndothiapepsinBarrierpepsin
AspergillopepsinAspergillopepsinSaccharopepsinPolyporopepsin
PhytepsinPlamepsin IPlamepsin II
YapsinMemapsin IMemapsin II
Figure 1.1. Aspartic proteases family tree
The aspartic acid protease family can be split up into five main categories. Pepsin-like contains 70 members, retroviral contains 45 members, cauliflower mosaic contains 6,
t i l t i 1 d i t t i 6 O l l ti fspumaretroviral contains 1, and copia transposon contains 6. Only a selection of enzymes are shown in this diagram. Adapted from Dunn (2001) and Rawlings, Morton et al. (2008).
14

1.2 Catalytic Mechanism of Aspartic Proteases
As previously mentioned, aspartic proteases contain a conserved segment of
residues. This amounts to about a 5% sequence identity in all of the enzymes. Two of
the residues that remain conserved are aspartic acids (hence the name aspartic acid
proteases). The aspartic residues, Asp32 and Asp215 (pepsin numbering), are located in
the active site cleft of the enzyme and are involved in the catalytic mechanism (Dunn
1989). To date the most popular mechanism of aspartic proteases (Figure 1.2) was
proposed by Northrop (Northrop 2001) and is believed to involve the controversial low-
barrier hydrogen bond.
The mechanism begins with the free enzyme wherein the two aspartate groups are
believed to form a 10-atom cyclic structure with a water molecule. This forms a loose
complex with the substrate in step 1. The catalytic process begins in step 2 when the
complex takes on the required geometry and distances. In step 3 a proton is removed
from the bound water molecule in order to generate a hydroxide ion, which attacks the
carbon atom of the substrate’s carboxyl group. A proton is then transferred to the
nitrogen of the peptide bond in step 4 and bond cleavage occurs in step 5. In step 6 the
complex opens up for release of the products in step 7. A proton is then lost in step 8 and
finally a new water molecule is bound to the complex in step 9. Steps 8 and 9 represent
the deprotonation, rehydration and restructuring of the complex to complete
isomerization. Isomerization is necessary in order to reform the low-barrier hydrogen
bond (Northrop 2001; Dunn 2002).
15

1 2 3
4 5 6
7 8 9
Figure 1.2. The aspartic protease catalytic mechanism. The first step in the mechanism is the formation of a loose complex upon substrate binding. Step 2 begins the catalytic process and in step 3 the carbonyl group is attacked following the removal of a proton. In step 4 there is the transfer of a proton and in step 5 bond cleavage occurs. The complex opens up in step 6 and the product is released in step 7. In step 8 a proton is lost and in step 9 a new water molec le binds to the comple Adapted from Northrop (2001)molecule binds to the complex. Adapted from Northrop (2001).
16

1.3 Primary Structure of the Pepsin-like Family
Pepsin-like proteases make up the largest family of acid proteases. They are
roughly 330-350 residues in length and share a common primary structure. Pepsin-like
proteases are single chain enzymes that are usually described as having two domains.
The primary structure is evidence as to why the protein is described as having two-
domains as it has a repetitive form (Orengo, Michie et al. 1997).
The sequence typically begins with 30 to 35 residues followed by two
hydrophobic residues (Hpb). The hydrophobic residues are followed by the conserved
Asp-Thr-Gly sequence found in all aspartic proteases. The Asp-Thr-Gly sequence occurs
within a wide loop known as the Psi-loop. About 45 residues after the Asp-Thr-Gly
sequence a conserved Tyr occurs. After about another 45 residues there is a Leu-Gly-Ile
sequence followed by about 90 more residues. The sequence then shows some form of
repetition. Two hydrophobic residues occur again before the Asp-Thr-Gly sequence.
There are about 85 more residues before a Leu-Gly-Asp sequence. 20 to 30 more amino
acids make up the rest of the primary structure of a pepsin-like protease (Davies 1990).
The tyrosine residue that is conserved in the sequence is important because it
helps to define the active site pockets where substrate side-chain residues bind. In the
Leu-Gly-Ile and Leu-Gly-Asp sequence the glycine occurs for structural reasons as it is
easily able to fit through the Psi-loop. The leucine, isoleucine and aspartic acid residues
are there because their bulkiness ensures that the Psi-loop is locked into place (Dunn
2001).
~ 30-35AA ~ Hpb-Hpb-Asp-Thr-Gly ~ 45-50 AA ~ Tyr ~ 45-50 AA ~ Leu-Gly-Ile ~ 90-95AA ~ Hpb-Hpb-Asp-Thr-Gly ~ 85-90AA ~ Leu-Gly-Asp ~ 20-30AA
17

1.4 Pepsin
1.4.1 History of Pepsin
In terms of evolutionary history, pepsin is considered to be the first enzyme in the
aspartic protease family. It was the first enzyme recognized as having activity (in
digestive processes) and in 1825 it was the first to be given a name (Gillespie 1898).
Porcine pepsin was also one of the first proteins to be extracted (Gillespie 1898) and to be
crystallized (Northrop 1930). It was also the first enzyme that has X-ray diffraction
patterns from crystals (Bernal and Crowfoot 1934). The extracted enzyme needs to be
acidified in order for full activity to be regained. It was noted that different acids yielded
different levels of activity. The activities were plotted against hydrogen ion
concentrations by Sorensen (Sorensen 1909). The plot had a scaling issue which
Sorensen resolved using a logarithmic abscissa, thus inventing the pH scale.
1.4.2 Activation of Pepsin
Pepsin, along with other aspartic proteases commonly found in vertebrates and
plants, is most often synthesized as an inactive zymogen. For pepsin this zymogen is
pepsinogen. Pepsinogen has the same primary structure as pepsin plus an additional 44
residues at the N-terminal of the protein (Figure 1.3). This 44 residue segment is often
referred to as a propeptide and pepsinogen is often referred to as a proenzyme (Davies
1990). The pepsinogen propeptide contains nine lysine residues, two arginine residues
and two histidine residues which make the peptide basic. The propeptide forms a helical
structure that is stabilized by electrostatic forces as six of the basic side chains form ion
pairs with the carboxylate side chains of pepsin (Perlmann 1963). The propeptide
18

Pepsinogen LVKVPLVRKKSLRQNLIKNGKLKDFLKTHKHNPASKYFPEAAALIGDEPLENYLDTEYFG Pepsin --------------------------------------------IGDEPLENYLDTEYFG
Pepsinogen TIGIGTPAQDFTVIFDTGSSNLWVPSVYCSSLACSDHNQFNPDDSSTFEATSQELSITYG Pepsin TIGIGTPAQDFTVIFDTGSSNLWVPSVYCSSLACSDHNQFNPDDSSTFEATSQELSITYG
Pepsinogen TGSMTGILGYDTVQVGGISDTNQIFGLSETEPGSFLYYAPFDGILGLAYPSISASGATPV Pepsin TGSMTGILGYDTVQVGGISDTNQIFGLSETEPGSFLYYAPFDGILGLAYPSISASGATPV
Pepsinogen FDNLWDQGLVSQDLFSVYLSSNDDSGSVVLLGGIDSSYYTGSLNWVPVSVEGYWQITLDS iPepsin FDNLWDQGLVSQDLFSVYLSSNDDSGSVVLLGGIDSSYYTGSLNWVPVSVEGYWQITLDS
Pepsinogen ITMDGETIACSGGCQAIVDTGTSLLTGPTSAIANIQSDIGASENSDGEMVISCSSIDSLP Pepsin ITMDGETIACSGGCQAIVDTGTSLLTGPTSAIANIQSDIGASENSDGEMVISCSSIDSLP
Pepsinogen DIVFTINGVQYPLSPSAYILQDDDSCTSGFEGMDVPTSSGELWILGDVFIRQYYTVFDRA Pepsin DIVFTINGVQYPLSPSAYILQDDDSCTSGFEGMDVPTSSGELWILGDVFIRQYYTVFDRA
Pepsinogen NNKVGLAPVAPepsinogen NNKVGLAPVA Pepsin NNKVGLAPVA
Figure 1.3. Sequence alignment of porcine pepsinogen and porcine pepsin. The 44 residue propeptide is highlighted in blue. This segment is cleaved off upon activation.
19

inhibits the activity of the enzyme because a segment of it blocks access to the catalytic
aspartates in the active site. Removal of the propeptide results in the activation of
pepsinogen to pepsin (James and Sielecki 1986). A loss of helical structure of the
propeptide also typically occurs during activation of the zymogen (Davies 1990).
Pepsinogen activation occurs when the pH of a solution of pepsinogen is lowered.
The lowering of the pH is believed to protonate the carboxylate side chains of pepsin
which causes the complex to break down and leads to the formation of the active enzyme.
Raising the pH can fully reverse the zymogen activation if performed in a timely manner.
However, if the pH is lowered for a prolonged period of time the activation is irreversible
(James and Sielecki 1986).
The activation of pepsinogen into pepsin is believed to occur through two
pathways, either in a one-step process or in a sequential manner. There are also two
different reactions that occur during activation. In the intramolecular reaction pepsinogen
cleaves itself to form the active pepsin, while in the intermolecular reaction pepsinogen is
cleaved by either another pepsinogen molecule, an intermediate form or an active pepsin
molecule. Kinetic experiments have shown that the intramolecular reaction is
predominant at a pH lower than 3.0 (al-Janabi, Hartsuck et al. 1972). The one-step
activation pathway appears to proceed mainly, but not exclusively, through the
intermolecular reaction (Kageyama and Takahashi 1983).
Both the one-step pathway and the stepwise pathway are believed to occur
simultaneously during the activation of pepsinogen to pepsin (Christensen, Pedersen et al.
1977). The intramolecular reaction and the intermolecular reaction are both involved in
the one-step pathway. It appears as though the intramolecular reaction is an essential part
20

for the initial activation in order to generate the active pepsin molecules. The
intermolecular reaction is important for completion of the activation (Kageyama and
Takahashi 1987).
1.4.3 Pepsin Crystal Structure
Porcine pepsin was first crystallized in 1930 by John Northrop and later refined
by Sielecki et al. in 1990 (Sielecki, Fedorov et al. 1990). Figure 1.4 illustrates the crystal
structure of human pepsin (Fujinaga, Chernaia et al. 1995). The catalytic Asp residues,
Asp32 and Asp215, are highlighted in blue while the pepsin inhibitor pepstatin is
highlighted in red. The protein can be divided up into three regions (James and Sielecki
1986). The first region consists of a six-stranded antiparallel β-sheet. This interdomain
forms the backbone of the structure and is located behind the catalytic site region. The
other two domains consist of two lobes. One lobe is the N-terminal which consists of
142 residues and the other lobe is the C-terminal which consists of 123 residues. Despite
a similar pattern in their amino acid sequences, the N-terminal and C-terminal domains
are not very similar in their secondary or tertiary structures (Sielecki, Fedorov et al.
1990).
Other elements of the pepsin crystal structure are that it consists of a short
interdomain peptide that is next to the external side of the six-stranded β-sheet (Sielecki,
Fedorov et al. 1990). There are also two strands that form a β-hairpin loop that is often
called the flap. The flap projects out at the active site cleft of the molecule (Davies
1990). Pepsin contains a large hydrophobic core at its center. This is a result of the
reassembly of three regions mentioned above. A major factor contributing to the
21

Asp215
Asp32
Figure 1.4. Crystal structure of human pepsin. The two catalytic Asp residues, Asp32 and Asp215, are highlighted in blue. The pepsin inhibitor pepstatin is highlighted in red. PDB: 1PSN
22

hydrophobic core are side chains that protrude inward from the six-stranded β-sheet
(Sielecki, Fedorov et al. 1990).
The catalytic site of pepsin is highlighted by two aspartic acid residues, Asp32 and
Asp215. There is an Asp residue located in both the N-terminal and C-terminal domain.
The two Asp residues are located towards the end of each domain and are connected
through a network of hydrogen bonds. The active site is quite rigid. However, the flap
that protrudes out above the active site is rather flexible. The flap can close around
inhibitors that are bound to the active site, thus limiting the mobility of the flap (James,
Sielecki et al. 1982).
1.4.4 Activity of Pepsin
Pepsin is an enzyme whose activity is greatly dependent on its pH. Pepsin has its
optimum enzymatic activity at a pH between 1.8 and 2.0. It is remains stable, and still
highly active, when the pH drops to as low as 1.0 (Ryle 1970). Pepsin will begin to lose
activity around pH 5 (Smith 1991) and it becomes irreversibly inactive at a pH around 7.
However, a high concentration of pepsin will not become inactive until a pH of about 8
(Jones and Landon 2002). The activity of pepsin is also dependent upon the enzyme to
protein ratio. The higher this ratio is the more efficient the enzyme becomes (Wu, Kaveti
et al. 2006).
1.4.5 Pepsin and Proteomics
Pepsin can be a very useful tool for proteomics. Pepsin has a very broad
specificity and is believed to often cleave after bulky hydrophobic residues (Fruton 1970;
23

Ryle 1970). Because of its broad specificity pepsin produces many peptides during
digestion. The multiple cleavage sites means that the peptides produced are usually
small, around 3 to 30 residues in length. The peptides are also typically overlapping
which is useful for protein mapping. Despite its broad specificity pepsin is still a very
reproducible enzyme, meaning it will yield the same peptides when digestion of a protein
is performed at identical conditions (Zhang and Smith 1993).
1.5 Research Objectives
As previously mentioned there is little known about its specificity other than the
fact that it prefers to cleave after bulky hydrophobic residues (Fruton 1970). Determining
trends in pepsin specificity is important because pepsin is an enzyme that is widely used
and without any rules about its cleavage preferences, protein characterization can be very
difficult. The reason why pepsin is used for protein characterization despite the fact it
has little known specificity is because it is one of the only enzymes to have a high
activity in the low pH range.
One of the most common uses for pepsin is when performing hydrogen /
deuterium exchange mass spectrometry (HXMS). To date, pepsin is essentially the only
enzyme that can be used for these experiments. Due to the nature of the reaction, the
digests need to be performed at a pH around 2.5 and at 0 °C. While there are other
aspartic proteases that will work at this pH requirement there are no other enzymes that
remain as active as pepsin is at this low temperature. Since pepsin is so often used to
characterize proteins and because there is so little known about pepsin cleavage
preferences, the main objective of this research is to determine if pepsin has any
24

specificity, thus advancing the use of pepsin digestion for protein characterization by
mass spectrometry.
The steps taken to determine trends in pepsin specificity consist of two main
parts. Chapter 2 outlines an extensive literature search performed in order to gather all
existing data involving pepsin cleavages. The corresponding experimental research is
discussed in chapter 3. The goal of the experimental research was to gain more data of
pepsin digests. These analyses were performed at pH 2.5 which is the most common pH
for pepsin digests. Another aspect of the experiment which is discussed in chapter 3 is
the determination of the effect of pH on the specificity of pepsin. It is known that
pepsin’s activity is highly dependent on pH (Ryle 1970) but it is not known whether or
not that loss or gain of activity is directly related to specificity.
1.6 References
al-Janabi, J., J. A. Hartsuck, et al. (1972). "Kinetics and mechanism of pepsinogen
activation." J Biol Chem 247: 4628-32.
Bernal, J. D. and D. Crowfoot (1934). "X-Ray photographs of crystalline pepsin." Nature
133: 794-795.
Christensen, K. A., V. B. Pedersen, et al. (1977). "Identification of an enzymatically
active intermediate in the activation of porcine pepsinogen." FEBS Lett 76: 214-8.
Davies, D. R. (1990). "The structure and function of the aspartic proteinases." Annu Rev
Biophys Biophys Chem 19: 189-215.
25

Dickson, C., Eisenman, R., Fan, H., Hunter, E., Teich, N. (1984). RNA Tumor Viruses.
R. Weiss, Teich, N., Varmus, H., Coffin, J. New York, Cold Spring Harbor Lab:
513-648.
Dunn, B. M. (1989). Determination of Protease Mechanism. Proteolytic Enzymes: A
Practical Approach. R. J. a. B. Beynon, J. S. Oxford, England, Information Press
Ltd.: 57-81.
Dunn, B. M. (2001). "Overview of pepsin-like aspartic peptidases." Curr Protoc Protein
Sci Chapter 21: Unit 21 3.
Dunn, B. M. (2002). "Structure and mechanism of the pepsin-like family of aspartic
peptidases." Chem Rev 102: 4431-58.
Fruton, J. S. (1970). "The specificity and mechanism of pepsin action." Adv Enzymol
Relat Areas Mol Biol 33: 401-43.
Fruton, J. S. (1976). "The mechanism of the catalytic action of pepsin and related acid
proteinases." Adv Enzymol Relat Areas Mol Biol 44: 1-36.
Fujinaga, M., M. M. Chernaia, et al. (1995). "Crystal structure of human pepsin and its
complex with pepstatin." Protein Sci 4: 960-72.
Gillespie, A. L. (1898). The Natural History of Digestion. London, W. Scott.
James, M. N., A. Sielecki, et al. (1982). "Conformational flexibility in the active sites of
aspartyl proteinases revealed by a pepstatin fragment binding to penicillopepsin."
Proc Natl Acad Sci U S A 79: 6137-41.
James, M. N. and A. R. Sielecki (1986). "Molecular structure of an aspartic proteinase
zymogen, porcine pepsinogen, at 1.8 A resolution." Nature 319(6048): 33-8.
26

Jones, R. G. and J. Landon (2002). "Enhanced pepsin digestion: a novel process for
purifying antibody F(ab')(2) fragments in high yield from serum." J Immunol
Methods 263: 57-74.
Kageyama, T. and K. Takahashi (1983). "Occurrence of two different pathways in the
activation of porcine pepsinogen to pepsin." J Biochem 93: 743-54.
Kageyama, T. and K. Takahashi (1987). "Activation mechanism of monkey and porcine
pepsinogens A. One-step and stepwise activation pathways and their relation to
intramolecular and intermolecular reactions." Eur J Biochem 165: 483-90.
Northrop, D. B. (2001). "Follow the protons: a low-barrier hydrogen bond unifies the
mechanisms of the aspartic proteases." Acc Chem Res 34: 790-7.
Northrop, J. H. (1930). "Crystalline pepsin I. Isolation and tests for purity." J. Gen.
Physiol. 13: 739-766.
Orengo, C. A., A. D. Michie, et al. (1997). "CATH--a hierarchic classification of protein
domain structures." Structure 5: 1093-108.
Perlmann, G. E. (1963). "The optical rotatory properties of pepsinogen." J Mol Biol 6:
452-64.
Rawlings, N. D., F. R. Morton, et al. (2008). "MEROPS: the peptidase database." Nucleic
Acids Res 36: D320-5.
Ryle, A. (1970). "The Porcine Pepsin and Pepsinogens." Methods Enzymol 19: 316-336.
Sielecki, A. R., A. A. Fedorov, et al. (1990). "Molecular and crystal structures of
monoclinic porcine pepsin refined at 1.8 A resolution." J Mol Biol 214: 143-70.
27

Smith, J. L., Billings, G. E., Yada, R. Y (1991). "Chemical Modification of Amino
Groups in Mucor miehei Aspartyl Proteinase, Porcine Pepsin, and Chymosin. I.
Structure and Function." Agricultural and Biological Chemistry 55: 2009-2016.
Sorensen, S. P. L. (1909). "Enzymstudien II. Mitteilung. Uber die Messung und die
Bedeutung der Wasserstoffionen-konzentration bei enzymatischen Prozessen."
Biochem. Z. 21: 201-304.
Wu, Y., S. Kaveti, et al. (2006). "Extensive deuterium back-exchange in certain
immobilized pepsin columns used for H/D exchange mass spectrometry." Anal
Chem 78: 1719-23.
Zhang, Z. and D. L. Smith (1993). "Determination of amide hydrogen exchange by mass
spectrometry: a new tool for protein structure elucidation." Protein Sci 2: 522-31.
28

CHAPTER 2
LITERATURE SEARCH
2.1. Introduction
The first part of this research project was to conduct a search of contemporary
scientific literature for publications that involve pepsin digestion and mass spectrometry.
Performing this literature search was useful because it provided a large amount of
cleavages from a diverse set of proteins. This extensive amount of data is a very good
basis for determining pepsin specificity.
A literature analysis of pepsin specificity has been performed previously (Keil
1992). However, the search performed in this book was a very broad one. There were no
limitations put on the pH at which the digestions were performed. As previously
mentioned pepsin can be greatly affected by pH. Since this search takes into account
digestions performed at a wide range of pH values, some of the specificity results could
be skewed because of it.
The literature search performed for this project puts strict limitations on the pH at
which the digestions could be performed. In order to be considered in this literature
search digestion needed to be performed within a pH range of 2.5 to 2.7. Pepsin
experiences a very high activity within these values. This pH range is also of
considerable importance because it is the range at which HXMS experiments are
conducted. As previously mentioned HXMS experiments are one of the main types of
experiments that utilize pepsin digestions. Due to this fact, there is a vast amount of
literature published containing digestions within these pH values.
29

2.2. Materials and Methods
Search engines for online databases were used to conduct the literature search.
As mentioned above the literature that was of interest involved pepsin digestions and
mass spectrometry. The initial search yielded hundreds of results. The results were
narrowed by only choosing digestions that were performed within a pH range of 2.5 to
2.7.
Once the results were limited to those containing digests performed at pH 2.5 to
2.7, all of the papers were scanned to see if they contained a peptic digest map (Figure
2.1). If two or more sources used the same peptic digest map only one was used to
extract data from. Likewise, if two or more sources contained digest maps of the same
protein from the same organism the most comprehensive map was used ensuring that a
specific protein only be considered once when gathering data. The final results yielded
peptic digest maps from 40 sources. Table 2.1 lists the references along with the
protein(s) studied in the publication.
2.3. Construction of Cleavage Database
All of the peptic digest maps retrieved from the literature search were analyzed in
order to create a database of pepsin cleavages. The database consists of the residues that
are found in the P4 to P4’ positions (see table 2.2 at end of chapter). As shown in Figure
2.2, residues P1 through P4 occur before the cleavage and residues P1’ through P4’ occur
after the cleavage. The database incorporates out to the P4 and P4’ position because there
has been some evidence that residues in these positions can effect pepsin specificity (Keil
1992).
30

Peptic peptides
RRGAISAEVY TEEDAASYVR KVIPKDYKTM
AALAKAIEKN VLFSHLDDNE RSDIFDAMFP
VSFIAGETVI QQGDEGDNFY VIDQGEMDVYVSFIAGETVI QQGDEGDNFY VIDQGEMDVY
Figure 2.1. Example of a peptic digest map. Peptic peptides are underlined in red. Pepsin will produce multiple overlapping peptides.
31

Table 2.1. Literature search results Reference Protein
Lu, Wintrode et al. 2007 Major prion protein (human) Man, Montagner et al. 2007 Myoglobin (sperm whale) Brier, Maria et al. 2007 CENP-E (human) Cheng, Cusanovich et al. 2006 Photoactive yellow protein Cheng, Wysocki et al. 2006 cytochrome c2 (rhodobacter capsulatus) Hochrein, Wales et al. 2006 HIV and SIV Nef Shi, Koeppe et al. 2006 AChBP from L. stagnalis Tsutsui, Liu et al. 2006 α1AT (human) Wales and Engen 2006 Lyn SH3 and α-spectrin SH3 Weis, Kjellen et al. 2006 Lck SH3 Yao, Zhou et al. 2006 Cks1 and Skp2 Catalina, Fischer et al. 2005 SH2 domains of Syk tSH2 Kang and Prevelige 2005 P22 capsid coat protein Lee, Hoofnagle et al. 2005 ERK2 Brier, Lemaire et al. 2004 Kinesin-like protein KIF11 (human) Casbarra, Birolo et al. 2004 Human α-LA Croy, Koeppe et al. 2004 human and bovine α-thrombin Croy, Bergqvist et al. 2004 IκBα Li, Chou et al. 2004 LR3IGF-I Mazon, Marcillat et al. 2004 Creatine kinase M-type (rabbit) Wu, Hasan et al. 2004 human rCRALBP Yan, Broderick et al. 2004 human RXRα LBD Anand, Law et al. 2003 PKA Chik and Schriemer 2003 rabbit muscle actin Cravello, Lascoux et al. 2003 PBP-2X* Rist, Jorgensen et al. 2003 σ 32 Wintrode, Friedrich et al. 2003 HSP16.9 Hasan, Smith et al. 2002 α-Crystallin (αA and αB) Yan, Zhang et al. 2002 rhM-CSFß Hughes, Mandell et al. 2001 CheB Wang, Lane et al. 2001 BMV Chen and Smith 2000 GroEL Engen, Smithgall et al. 1999 Hck SH(3 + 2) Wang, Li et al. 1999 cNTnC Resing and Ahn 1998 human MKK1 Neubert, Walsh et al. 1997 recoverin Wang, Blanchard et al. 1997 DHPR Dharmasiri and Smith 1996 horse heart cyt c Zhang, Post et al. 1996 rabbit muscle aldolase Johnson and Walsh 1994 equine myoglobin
32

P P P P P ’ P ’ P ’ P ’P4 P3 P2 P1 P1 P2 P3 P4 T E E D A A S Y
RRGAISAEVY TEEDAASYIR KVIPKDYKTM
AALAKAIEKN VLFSHLDDNE RSDIFDAMFP
VSFIAGETVI QQGDEGDNFY VIDQGEMDVY
Figure 2.2. Example of cleavage nomenclature. The P4 through P1 residues are those that occur before the cleavage site. The P1’ through P4’ residues occur after the cleavage site.
33

After the database was constructed the residues in the P1 and P1’ positions were
focused on as these are believed to have the most influence on pepsin specificity
(Hamuro, Coales et al. 2008). For the analysis, all of the cleavages between two specific
residues were tallied to construct a matrix of cleavage data (Table 2.3).
As shown in Table 2.3 the residue that occurred most often before the cleavage
site (the P1 position) is leucine. Out of the 1,344 cleavages 372, or 28%, occurred after a
leucine. The residues that occurred most often in the P1’ position are leucine and alanine.
The residues that occur least often before the cleavage point are proline, histidine and
lysine. Glycine, proline and lysine are the three residues that produce the least number of
cleavages when found in the position following the cleavage site. This matrix of
cleavage data is comprehensive of the literature search performed; however there are
some issues with it. It is difficult to extract any trends in pepsin specificity just by
looking at the table. Another problem is that this raw data does not take into account the
abundance of amino acids. To make the values more meaningful a normalization of the
data must be performed.
2.4. Data Normalization
Normalizing the data is important because it takes into account how often a
specific amino acid occurs in a protein’s sequence. Taking the data in Table 2.3 for
example, 36 cleavages occurred between a leucine and a leucine while only three
cleavages occurred between a two tryptophans. When looking at the sequences for the
proteins in the literature there are 85 times in which two leucines occur next to one
another, thus pepsin produced a cleavage between a leucine and a leucine 36 out of 85
34

Table 2.3. Sum of cleavages betw
een two residues
P1 '
A
C
D
E
F G
H
I
K
L M
N
P
Q
R
S T
V W
Y A
5
1 2
6 10
0 1
164
14 2
1 1
1 2
3 2
110
12C
1
1 1
1 0
0 0
1 0
2 1
0 2
0 0
1 1
3 0
1 D
6
1 3
4 6
1 0
140
7 1
3 4
2 2
1 2
9 3
6 E
11 1
115
7 1
2 10
6 20
6 6
2 4
101
2 16
3 10
F 17
2 13
9 10
9 3
118
14 3
6 2
5 10
115
152
10G
4
2 0
3 4
3 1
4 1
5 2
1 0
1 2
3 2
3 2
4 H
1
0 1
0 0
0 0
0 0
1 0
0 0
0 0
0 0
0 0
0 I
5 0
0 1
5 0
0 2
1 5
1 2
0 1
2 3
3 3
0 1
K
3 0
2 4
0 0
0 1
2 1
0 2
0 0
0 1
3 1
0 1
P1
L 39
9 25
2819
1313
1714
36 12
158
1415
2222
259
17M
7
1 3
106
2 1
4 9
4 4
2 0
0 0
5 1
7 2
3 N
0
1 2
1 6
2 0
5 1
2 2
1 1
0 1
1 3
7 1
4 P
0 0
1 0
0 0
0 1
0 0
0 1
0 0
0 0
1 1
0 0
Q
3 0
3 2
3 0
0 5
1 4
0 1
0 1
1 0
1 7
1 5
R
0 0
1 2
1 0
0 1
1 3
0 0
1 1
1 1
2 2
2 0
S 4
0 2
3 4
4 1
4 2
6 2
0 2
0 1
0 0
6 3
8 T
6 0
4 1
6 5
0 6
0 7
1 1
0 1
4 0
0 2
1 6
V 5
0 4
1 2
1 0
2 3
3 1
0 0
1 1
3 3
2 1
5 W
0
0 1
0 3
0 1
2 0
3 0
0 0
2 0
0 0
0 3
2 Y
3 1
0 1
4 2
0 5
1 4
2 3
0 2
1 2
2 11
2 4
Tally of cleavages between tw
o specific residues. Residues in the P
1 positions are in the left hand column and residues in the P
1 ’ positions are along the top row
. For example, cleavage betw
een a phenylalanine and glycine occurred 9 times w
hile cleavage between
a leucine and a leucine occurred 36 times.
35

possible times. Two tryptophans occurred next to each other in the primary structures
only five times. Therefore, pepsin produced a cleavage between a tryptophan and a
tryptophan three out of five times. Leucine is one of the more abundant amino acids
while tryptophan is not. This is why data normalization is needed.
The first step to normalizing the raw data was to gather the sequences of all of the
proteins from the literature search. A matrix of possible cleavages was then constructed
(Table 2.4). The total number of possible cleavages for this set of proteins is 11,240.
The data was then normalized using the equation found in Figure 2.3 (Keil 1992). The
normalized values are shown in Table 2.5.
The matrix in Table 2.5 was then transformed into cleavage data map. The cleavage
data map is a more illustrative way to represent the data than the matrix form. It is much
easier to decipher trends in pepsin specificity by glancing at the cleavage map than it is to
look at the matrix. The cleavage data map is shown in Figure 2.4.
2.5. Cleavage Data Map
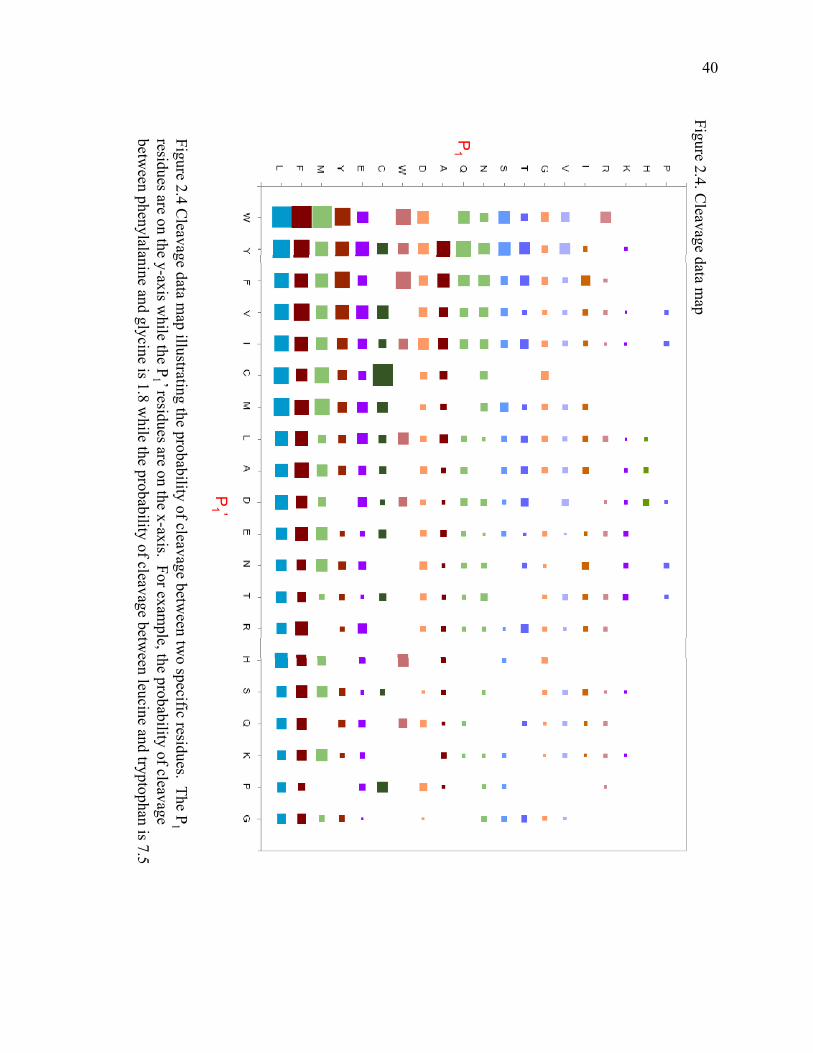
The cleavage data map in Figure 2.4 shows some trends in pepsin specificity. On
the y-axis are the P1 residues and on the x-axis are the P1’ residues. The size of the
square is the normalized cleavage value. For example, the probability that pepsin will
cleave between leucine and tryptophan is 7.5 while the probability that pepsin will cleave
between phenylalanine and glycine is 1.8. The map is arranged so that the residues
where pepsin prefers to cleave most often are nearest to the zero point on the chart and
those residues where pepsin does not prefer to cleave are farther out.
36

Table 2.4. Possible cleavages between tw
o residues P
1 '
A
C
D
E F
G
H
I K
L
M
N
P Q
R
S
T V
W
Y A
89
7 55
5624
7318
6155
75 22
3235
3834
4745
559
24C
8
1 13
7 7
123
7 7
14 4
5 8
105
128
9 0
4 D
47
8 47
4927
6012
4742
75 16
2127
1934
3818
449
21E
66 6
5590
3158
2148
7879
2434
19 33
4729
4950
1023
F 34
6 39
2122
4211
2431
33 6
2517
1824
3223
232
15G
56
11 47
5835
5814
4958
61 27
3422
3642
5159
5911
34H
12
4 10
1812
2544
6 18
30 13
6 21
8 14
2214
171
15I
47 8
3327
2431
2332
3761
1216
30 25
3644
4545
8 20
K
77 11
4860
2757
2739
6868
1632
37 24
3035
3456
1125
P1
L 92
16 58
8838
6128
3473
85 18
5039
5453
8270
5110
24M
25
2 19
2714
236
1228
29 7
6 11
7 13
1815
212
7 N
20
7 19
3620
268
2730
48 16
1522
1419
2924
346
14P
30 7
3456
2836
1423
3039
1216
16 12
2045
2433
1111
Q
27 3
2331
1130
1028
3247
1715
22 19
2030
2139
3 9
R
40 10
3447
2427
1234
3245
9 18
35 23
3135
3540
8 14
S 40
12 43
4831
5924
3538
65 12
2746
2937
5731
439
23T
51 5
2751
3051
1533
3868
1721
34 22
2930
4238
9 19
V 64
14 37
5731
4416
3757
69 18
4033
2138
5137
406
17W
8
2 6
6 5
113
9 11
10 1
7 2
107
6 7
7 5
7 Y
18 5
2017
7 23
4 20
2229
9 18
21 17
2219
2326
3 9
Tally of possible cleavages between tw
o specific residues. Residues in the P
1 positions are in the left hand column and residues in the
P1 ’ positions are along the top row
. For example, cleavage betw
een a phenylalanine and glycine could occur 42 times w
hile cleavage betw
een a leucine and a leucine could occur 85 times.
37

( Frequency of cleavages between two )( Frequency of cleavages between two specific residues
Total number of cleavages )( )Frequency of times two specific residues
appear next to each other in the sequences
=Normalized value ( )appear next to each other in the sequences
Total number of residues
( 9 )
Figure 2.3. Equation used for data normalization and example calculation from Keil(1993) Th li ti ti t k i t t th b d f i id
( 91344 )
( )4211240
=Normalized value = 1.8
(1993). The normalization equation takes into account the abundance of amino acids. The example calculation is for the probability of cleavage between a phenylalanine and a glycine. Cleavage between these two specific residues was observed nine times in the literature, where there was a total of 1344 cleavages. These two residues were found next to one another in the protein sequences 42 times. This means that there was possibility of cleavage between these two residues 42 times out of a possible 11240 residues. Therefore the probability of a cleavage occurring between a phenylalanine and
l i i 1 8a glycine is 1.8.
38

Table 2.5. Norm
alized cleavage data P
1 '
A
C
D
E F
G
H
I K
L
M
N
P Q
R
S
T V
W
Y A
0.5
1.2 0.3
0.93.5
0.00.5
2.20.6
1.60.8
0.3 0.2
0.20.5
0.50.4
1.70.0
4.2C
1.0
8.4 0.6
1.20.0
0.00.0
1.20.0
1.22.1
0.0 2.1
0.00.0
0.71.0
2.80.0
2.1D
1.1
1.0 0.5
0.71.9
0.10.0
2.50.0
0.80.5
1.2 1.2
0.90.5
0.20.9
1.72.8
2.4E
1.4 1.4
1.7 0.5
1.90.1
0.81.7
0.62.1
2.11.5
0.9 1.0
1.80.3
0.32.7
2.53.6
F 4.2
2.8 2.8
3.63.8
1.82.3
3.82.2
3.54.2
2.0 1.0
2.33.5
2.91.8
5.58.4
5.6G
0.6
1.5 0.0
0.41.0
0.40.6
0.70.1
0.70.6
0.2 0.0
0.20.4
0.50.3
0.41.5
1.0H
0.7
0.0 0.8
0.00.0
0.00.0
0.00.0
0.30.0
0.0 0.0
0.00.0
0.00.0
0.00.0
0.0I
0.9 0.0
0.0 0.3
1.70.0
0.00.5
0.20.7
0.71.0
0.0 0.3
0.50.6
0.60.6
0.00.4
K
0.3 0.0
0.3 0.6
0.00.0
0.00.2
0.20.1
0.00.5
0.0 0.0
0.00.2
0.70.1
0.00.3
P1
L 3.5
4.7 3.6
2.74.2
1.83.9
4.21.6
3.55.6
2.5 1.7
2.22.4
2.22.6
4.17.5
5.9M
2.3
4.2 1.3
3.13.6
0.71.4
2.82.7
1.24.8
2.8 0.0
0.00.0
2.30.6
2.88.4
3.6N
0.0
1.2 0.9
0.22.5
0.60.0
1.50.3
0.31.0
0.6 0.4
0.00.4
0.31.0
1.71.4
2.4P
0.0 0.0
0.2 0.0
0.00.0
0.00.4
0.00.0
0.00.5
0.0 0.0
0.00.0
0.30.3
0.00.0
Q
0.9 0.0
1.1 0.5
2.30.0
0.01.5
0.30.7
0.00.6
0.0 0.4
0.40.0
0.41.5
2.84.6
R
0.0 0.0
0.2 0.4
0.30.0
0.00.2
0.30.6
0.00.0
0.2 0.4
0.30.2
0.50.4
2.10.0
S 0.8
0.0 0.4
0.51.1
0.60.3
1.00.4
0.81.4
0.0 0.4
0.00.2
0.00.0
1.22.8
2.9T
1.0 0.0
1.2 0.2
1.70.8
0.01.5
0.00.9
0.50.4
0.0 0.4
1.20.0
0.00.4
0.92.6
V 0.7
0.0 0.9
0.10.5
0.20.0
0.50.4
0.40.5
0.0 0.0
0.40.2
0.50.7
0.41.4
2.5W
0.0
0.0 1.4
0.05.0
0.02.8
1.90.0
2.50.0
0.0 0.0
1.70.0
0.00.0
0.05.0
2.4Y
1.4 1.7
0.0 0.5
4.80.7
0.02.1
0.41.2
1.91.4
0.0 1.0
0.40.9
0.73.5
5.63.7
Residues in the P
1 positions are in the left hand column and residues in the P
1 ’ positions are along the top row.
39
Fi
P1
igure 2.4. Cleavage d
Figure 2.4 Cleavage
residues are on the ybetw
een phenylalan data map
e data map illustratin
y-axis while the P
1 ’ rnine and glycine
is 1.
P1 ’
ng the probability of cresidues are on the x-8 w
hile the probabili cleavage between tw
o-axis. For exam
ple, tity of cleavage betw
e o specific residues. Tthe probability of cleaeen leucine and trypto The P
1avage ophan is 7.5
40

The map shows that pepsin prefers to cleave most often after leucine,
phenylalanine, methionine and tyrosine. It prefers to cleave most often before
tryptophan, tyrosine, phenylalanine and valine. One of the more telling aspects of the
graph is where pepsin rarely cleaves. It is very obvious that pepsin will rarely cleave
after proline, histidine or lysine. Also, pepsin will rarely cleave before glycine, proline or
lysine.
2.6. Revised Cleavage Data Map
A second cleavage data map (Figure 2.5) was then produced illustrating the
probability of cleavage between two specific residues as a percentage. The percentages,
shown in Table 2.6, were calculated by dividing the number of times a cleavage occurred
between two specific amino acids by the total number of times those residues occurred
next to each other in the protein sequences. For example, cleavage between leucine and
tryptophan occurred nine out of a possible ten times, or 90% of the time while cleavage
between phenylalanine and glycine only occurred nine out of a possible 42 times, or 21%
of the time.
In the second cleavage data map presented the x-axis and y-axis are simply
ordered alphabetically. This illustration of the data makes it easier to spot trends in
pepsin specificity. While both of the cleavage data maps utilize color only in the second
version do those colors actually mean something. In the first version the colors were
arbitrarily assigned to a P1 residues while in the second version the colors represent the
percentage of time a cleavage occurred between two specific residues.
41

Figure 2.5. Cleavage data map with probability defined as a percentage
YWVTSR
P1
QPNMLKKIHGFED
Scale
A C D E F G H I K L M N P Q R S T V W Y
DC
A
P1’
100 90 80 70 60 50 40 30 20 10
Figure 2.5 Cleavage data map representing the probability of cleavage between two specific residues as a percentage. This map is read the same as in Figure 2.4 with the residue occurring before the cleavage site on the y-axis and the residue occurring following the cleavage site on the x-axis. Each colored square represents a percentagefollowing the cleavage site on the x axis. Each colored square represents a percentage range. For example, the brown square represents the range 21-30% and the orange square represents the range 81-90%. Cleavage between phenylalanine and glycineoccurred 21% of the time while cleavage between leucine and tryptophan occurred 90% of the time.
42

Table 2.6. Cleavage data w
ith probability defined as a percentage P
1 '
A
C
D
E F
G
H
I K
L
M
N
P Q
R
S
T V
W
Y A
6
14 4
11 42
0 6
26 7
19 9
3 3
3 6
6 4
20 0
50 C
13
100 8
14 0
0 0
14 0
14 25
0 25
0 0
8 13
33 0
25 D
13
13 6
8 22
2 0
30 0
9 6
14 15
11 6
3 11
20 33
29 E
17 17
20 6
23 2
10 21
8 25
25 18
11 12
21 3
4 32
30 43
F 50
33 33
43 45
21 27
46 26
42 50
24 12
28 42
34 22
65 100
67 G
7
18 0
5 11
5 7
8 2
8 7
3 0
3 5
6 3
5 18
12 H
8
0 10
0 0
0 0
0 0
3 0
0 0
0 0
0 0
0 0
0 I
11 0
0 4
21 0
0 6
3 8
8 13
0 4
6 7
7 7
0 5
K
4 0
4 7
0 0
0 3
3 1
0 6
0 0
0 3
9 2
0 4
P1
L 42
56 43
32 50
21 46
50 19
42 67
30 21
26 28
27 31
49 90
71 M
28
50 16
37 43
9 17
33 32
14 57
33 0
0 0
28 7
33 100
43 N
0
14 11
3 30
8 0
19 3
4 13
7 5
0 5
3 13
21 17
29 P
0 0
3 0
0 0
0 4
0 0
0 6
0 0
0 0
4 3
0 0
Q
11 0
13 6
27 0
0 18
3 9
0 7
0 5
5 0
5 18
33 56
R
0 0
3 4
4 0
0 3
3 7
0 0
3 4
3 3
6 5
25 0
S 10
0 5
6 13
7 4
11 5
9 17
0 4
0 3
0 0
14 33
35 T
12 0
15 2
20 10
0 18
0 10
6 5
0 5
14 0
0 5
11 32
V 8
0 11
2 6
2 0
5 5
4 6
0 0
5 3
6 8
5 17
29 W
0
0 17
0 60
0 33
22 0
30 0
0 0
20 0
0 0
0 60
29 Y
17 20
0 6
57 9
0 25
5 14
22 17
0 12
5 11
9 42
67 44
Residues in the P
1 positions are in the left hand column and residues in the P
1 ’ positions are along the top row. The values listed here
represent the percentage of time a cleavage w
as observed between tw
o specific residues.
43

2.7. Summary of Literature Search
The extensive search of contemporary scientific literature involving pepsin
digestions performed at pH 2.5 to 2.7 showed that pepsin does show some preferences in
where it cleaves. The peptic digest maps found in the literature were analyzed in order to
create a database of cleavages. The cleavages between the residues in the P1 and P1’
positions were tallied in order to create a matrix of data. These data were then
normalized in order to take into account the abundance of amino acids in the protein
sequences. The normalized data were then made into a cleavage map in order to easily
see trends in pepsin specificity. The literature shows that pepsin prefers to cleave after
bulky hydrophobic residues such as leucine and phenylalanine. It also shows that pepsin
will hardly ever cleave after proline or histidine.
The data gathered from the literature search only consists of 1,344 cleavages.
While this is a good starting point, much more data is needed to more accurately
determine trends in the specificity of pepsin.
2.8. References
Anand, G. S., D. Law, et al. (2003). "Identification of the protein kinase A regulatory
RIalpha-catalytic subunit interface by amide H/2H exchange and protein
docking." Proc Natl Acad Sci U S A 100(23): 13264-9.
Brier, S., D. Lemaire, et al. (2004). "Identification of the protein binding region of S-
trityl-L-cysteine, a new potent inhibitor of the mitotic kinesin Eg5." Biochemistry
43(41): 13072-82.
44

Brier, S., G. Maria, et al. (2007). "Purification and characterization of pepsins A1 and A2
from the Antarctic rock cod Trematomus bernacchii." Febs J 274(23): 6152-66.
Casbarra, A., L. Birolo, et al. (2004). "Conformational analysis of HAMLET, the folding
variant of human alpha-lactalbumin associated with apoptosis." Protein Sci 13(5):
1322-30.
Catalina, M. I., M. J. Fischer, et al. (2005). "Binding of a diphosphorylated-ITAM
peptide to spleen tyrosine kinase (Syk) induces distal conformational changes: a
hydrogen exchange mass spectrometry study." J Am Soc Mass Spectrom 16(7):
1039-51.
Chen, J. and D. L. Smith (2000). "Unfolding and disassembly of the chaperonin GroEL
occurs via a tetradecameric intermediate with a folded equatorial domain."
Biochemistry 39(15): 4250-8.
Cheng, G., M. A. Cusanovich, et al. (2006). "Properties of the dark and signaling states of
photoactive yellow protein probed by solution phase hydrogen/deuterium
exchange and mass spectrometry." Biochemistry 45(39): 11744-51.
Cheng, G., V. H. Wysocki, et al. (2006). "Local stability of Rhodobacter capsulatus
cytochrome c2 probed by solution phase hydrogen/deuterium exchange and mass
spectrometry." J Am Soc Mass Spectrom 17(11): 1518-25.
Chik, J. K. and D. C. Schriemer (2003). "Hydrogen/deuterium exchange mass
spectrometry of actin in various biochemical contexts." J Mol Biol 334(3): 373-
85.
45

Cravello, L., D. Lascoux, et al. (2003). "Use of different proteases working in acidic
conditions to improve sequence coverage and resolution in hydrogen/deuterium
exchange of large proteins." Rapid Commun Mass Spectrom 17(21): 2387-93.
Croy, C. H., S. Bergqvist, et al. (2004). "Biophysical characterization of the free
IkappaBalpha ankyrin repeat domain in solution." Protein Sci 13(7): 1767-77.
Croy, C. H., J. R. Koeppe, et al. (2004). "Allosteric changes in solvent accessibility
observed in thrombin upon active site occupation." Biochemistry 43(18): 5246-
55.
Dharmasiri, K. and D. L. Smith (1996). "Mass spectrometric determination of isotopic
exchange rates of amide hydrogens located on the surfaces of proteins." Anal
Chem 68(14): 2340-4.
Engen, J. R., T. E. Smithgall, et al. (1999). "Comparison of SH3 and SH2 domain
dynamics when expressed alone or in an SH(3+2) construct: the role of protein
dynamics in functional regulation." J Mol Biol 287(3): 645-56.
Hamuro, Y., S. J. Coales, et al. (2008). "Specificity of immobilized porcine pepsin in
H/D exchange compatible conditions." Rapid Commun Mass Spectrom 22(7):
1041-6.
Hasan, A., D. L. Smith, et al. (2002). "Alpha-crystallin regions affected by adenosine 5'-
triphosphate identified by hydrogen-deuterium exchange." Biochemistry 41(52):
15876-82.
Hochrein, J. M., T. E. Wales, et al. (2006). "Conformational features of the full-length
HIV and SIV Nef proteins determined by mass spectrometry." Biochemistry
45(25): 7733-9.
46

Hughes, C. A., J. G. Mandell, et al. (2001). "Phosphorylation causes subtle changes in
solvent accessibility at the interdomain interface of methylesterase CheB." J Mol
Biol 307(4): 967-76.
Johnson, R. S. and K. A. Walsh (1994). "Mass spectrometric measurement of protein
amide hydrogen exchange rates of apo- and holo-myoglobin." Protein Sci 3(12):
2411-8.
Kang, S. and P. E. Prevelige, Jr. (2005). "Domain study of bacteriophage p22 coat protein
and characterization of the capsid lattice transformation by hydrogen/deuterium
exchange." J Mol Biol 347(5): 935-48.
Keil, B. (1992). Specificity of Proteolysis. New York, Springer-Verlag.
Lee, T., A. N. Hoofnagle, et al. (2005). "Hydrogen exchange solvent protection by an
ATP analogue reveals conformational changes in ERK2 upon activation." J Mol
Biol 353(3): 600-12.
Li, X., Y. T. Chou, et al. (2004). "Integration of hydrogen/deuterium exchange and
cyanylation-based methodology for conformational studies of cystinyl proteins."
Anal Biochem 331(1): 130-7.
Lu, X., P. L. Wintrode, et al. (2007). "Beta-sheet core of human prion protein amyloid
fibrils as determined by hydrogen/deuterium exchange." Proc Natl Acad Sci U S
A 104(5): 1510-5.
Man, P., C. Montagner, et al. (2007). "Defining the interacting regions between
apomyoglobin and lipid membrane by hydrogen/deuterium exchange coupled to
mass spectrometry." J Mol Biol 368(2): 464-72.
47

Mazon, H., O. Marcillat, et al. (2004). "Hydrogen/deuterium exchange studies of native
rabbit MM-CK dynamics." Protein Sci 13(2): 476-86.
Neubert, T. A., K. A. Walsh, et al. (1997). "Monitoring calcium-induced conformational
changes in recoverin by electrospray mass spectrometry." Protein Sci 6(4): 843-
50.
Resing, K. A. and N. G. Ahn (1998). "Deuterium exchange mass spectrometry as a probe
of protein kinase activation. Analysis of wild-type and constitutively active
mutants of MAP kinase kinase-1." Biochemistry 37(2): 463-75.
Rist, W., T. J. Jorgensen, et al. (2003). "Mapping temperature-induced conformational
changes in the Escherichia coli heat shock transcription factor sigma 32 by amide
hydrogen exchange." J Biol Chem 278(51): 51415-21.
Shi, J., J. R. Koeppe, et al. (2006). "Ligand-induced conformational changes in the
acetylcholine-binding protein analyzed by hydrogen-deuterium exchange mass
spectrometry." J Biol Chem 281(17): 12170-7.
Tsutsui, Y., L. Liu, et al. (2006). "The conformational dynamics of a metastable serpin
studied by hydrogen exchange and mass spectrometry." Biochemistry 45(21):
6561-9.
Wales, T. E. and J. R. Engen (2006). "Partial unfolding of diverse SH3 domains on a
wide timescale." J Mol Biol 357(5): 1592-604.
Wang, F., J. S. Blanchard, et al. (1997). "Hydrogen exchange/electrospray ionization
mass spectrometry studies of substrate and inhibitor binding and conformational
changes of Escherichia coli dihydrodipicolinate reductase." Biochemistry 36(13):
3755-9.
48

Wang, F., W. Li, et al. (1999). "Fourier transform ion cyclotron resonance mass
spectrometric detection of small Ca(2+)-induced conformational changes in the
regulatory domain of human cardiac troponin C." J Am Soc Mass Spectrom
10(8): 703-10.
Wang, L., L. C. Lane, et al. (2001). "Detecting structural changes in viral capsids by
hydrogen exchange and mass spectrometry." Protein Sci 10(6): 1234-43.
Weis, D. D., P. Kjellen, et al. (2006). "Altered dynamics in Lck SH3 upon binding to the
LBD1 domain of Herpesvirus saimiri Tip." Protein Sci 15(10): 2402-10.
Wintrode, P. L., K. L. Friedrich, et al. (2003). "Solution structure and dynamics of a heat
shock protein assembly probed by hydrogen exchange and mass spectrometry."
Biochemistry 42(36): 10667-73.
Wu, Z., A. Hasan, et al. (2004). "Identification of CRALBP ligand interactions by
photoaffinity labeling, hydrogen/deuterium exchange, and structural modeling." J
Biol Chem 279(26): 27357-64.
Yan, X., D. Broderick, et al. (2004). "Dynamics and ligand-induced solvent accessibility
changes in human retinoid X receptor homodimer determined by hydrogen
deuterium exchange and mass spectrometry." Biochemistry 43(4): 909-17.
Yan, X., H. Zhang, et al. (2002). "Hydrogen/deuterium exchange and mass spectrometric
analysis of a protein containing multiple disulfide bonds: Solution structure of
recombinant macrophage colony stimulating factor-beta (rhM-CSFbeta)." Protein
Sci 11(9): 2113-24.
Yao, Z. P., M. Zhou, et al. (2006). "Activation of ubiquitin ligase SCF(Skp2) by Cks1:
insights from hydrogen exchange mass spectrometry." J Mol Biol 363(3): 673-86.
49

Zhang, Z. (1995). Protein Hydrogen Exchange Determined by Mass Spectrometry: A
New Tool for Probing Protein High-order Structure and Structural Changes.
Purdue. Ph.D.
Zhang, Z., C. B. Post, et al. (1996). "Amide hydrogen exchange determined by mass
spectrometry: application to rabbit muscle aldolase." Biochemistry 35(3): 779-91.
50

Table 2.2. Cleavage databaseReference Protein P4 P3 P2 P1 P1' P2' P3' P4'
Lu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) A A A G A V V GLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) G L G G Y M L GLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) Y M L G S A M SLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) L G S A M S R PLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) G S A M S R P ILu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) F G S D Y E D RLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) E D R Y Y R E NLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) Y Y R E N M H RLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) R E N M H R V PLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) Y P N Q V Y Y RLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) P N Q V Y Y R PLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) N Q V Y Y R P MLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) P M D E Y S N QLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) D C V N I T I KLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) C V N I T I K QLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) V N I T I K Q HLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) K G E N F T E TLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) F T E T D V K MLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) T E T D V K M MLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) D V K M M E R VLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) K M M E R V V ELu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) V E Q M C I T QLu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) C I T Q Y E R ELu, X., P. L. Wintrode, et al. (2007) Major prion protein (human) E S Q A Y Y Q RMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) M V L S E G EMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) S E E E W Q L VMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) E G G W Q L V LMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) Q L V L H V W AMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) V L H V W A K VMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) Q D I L I R L FMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) L I R L F K S HMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) E A E M K A S EMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) V T V L T A L GMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) L T A L G A I LMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) I P I K Y L E FMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) Y L E F I S E AMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) F I S E A I I HMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) I S E A I I H VMan, P., C. Montagner, et al. (2007) Myoglobin (sperm whale) A L E L F R K DBrier, S., E. Carletti, et al. (2006) CENP-E (human) M A E E GBrier, S., E. Carletti, et al. (2006) CENP-E (human) G A V A V C V RBrier, S., E. Carletti, et al. (2006) CENP-E (human) V A V C V R V RBrier, S., E. Carletti, et al. (2006) CENP-E (human) P L N S R E E SBrier, S., E. Carletti, et al. (2006) CENP-E (human) R P L N S R E EBrier, S., E. Carletti, et al. (2006) CENP-E (human) N S R E E S L GBrier, S., E. Carletti, et al. (2006) CENP-E (human) E T A Q V Y W KBrier, S., E. Carletti, et al. (2006) CENP-E (human) T A Q V Y W K TBrier, S., E. Carletti, et al. (2006) CENP-E (human) K T D N N V I YBrier, S., E. Carletti, et al. (2006) CENP-E (human) T D N N V I Y QBrier, S., E. Carletti, et al. (2006) CENP-E (human) N V I Y Q V D G
51

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Brier, S., E. Carletti, et al. (2006) CENP-E (human) I Y Q V D G S KBrier, S., E. Carletti, et al. (2006) CENP-E (human) K S F N F D R VBrier, S., E. Carletti, et al. (2006) CENP-E (human) S F N F D R V FBrier, S., E. Carletti, et al. (2006) CENP-E (human) T T K N V Y E EBrier, S., E. Carletti, et al. (2006) CENP-E (human) K N V Y E E I ABrier, S., E. Carletti, et al. (2006) CENP-E (human) N V Y E E I A ABrier, S., E. Carletti, et al. (2006) CENP-E (human) V Y E E I A A PBrier, S., E. Carletti, et al. (2006) CENP-E (human) E I A A P I I DBrier, S., E. Carletti, et al. (2006) CENP-E (human) P I I D S A I QBrier, S., E. Carletti, et al. (2006) CENP-E (human) I I D S A I Q GBrier, S., E. Carletti, et al. (2006) CENP-E (human) I D S A I Q G YBrier, S., E. Carletti, et al. (2006) CENP-E (human) G T I F A Y G QBrier, S., E. Carletti, et al. (2006) CENP-E (human) T I F A Y G Q TBrier, S., E. Carletti, et al. (2006) CENP-E (human) S G K T Y T M MBrier, S., E. Carletti, et al. (2006) CENP-E (human) Y T M M G S E DBrier, S., E. Carletti, et al. (2006) CENP-E (human) E D H L G V I PBrier, S., E. Carletti, et al. (2006) CENP-E (human) H D I F Q K I KBrier, S., E. Carletti, et al. (2006) CENP-E (human) R E F L L R V SBrier, S., E. Carletti, et al. (2006) CENP-E (human) E F L L R V S YBrier, S., E. Carletti, et al. (2006) CENP-E (human) R V S Y M E I YBrier, S., E. Carletti, et al. (2006) CENP-E (human) V S Y M E I Y NBrier, S., E. Carletti, et al. (2006) CENP-E (human) I T D L L C G TBrier, S., E. Carletti, et al. (2006) CENP-E (human) T D L L C G T QBrier, S., E. Carletti, et al. (2006) CENP-E (human) I I R E D V N RBrier, S., E. Carletti, et al. (2006) CENP-E (human) R N V Y V A D LBrier, S., E. Carletti, et al. (2006) CENP-E (human) Y V A D L T E EBrier, S., E. Carletti, et al. (2006) CENP-E (human) V A D L T E E VBrier, S., E. Carletti, et al. (2006) CENP-E (human) L T E E V V Y TBrier, S., E. Carletti, et al. (2006) CENP-E (human) Y T S E M A L KBrier, S., E. Carletti, et al. (2006) CENP-E (human) T S E M A L K WBrier, S., E. Carletti, et al. (2006) CENP-E (human) E T K M N Q R SBrier, S., E. Carletti, et al. (2006) CENP-E (human) H T I F R M I LBrier, S., E. Carletti, et al. (2006) CENP-E (human) R M I L E S R EBrier, S., E. Carletti, et al. (2006) CENP-E (human) P S N C E G S VBrier, S., E. Carletti, et al. (2006) CENP-E (human) V S H L N L V DBrier, S., E. Carletti, et al. (2006) CENP-E (human) S H L N L V D LBrier, S., E. Carletti, et al. (2006) CENP-E (human) H L N L V D L ABrier, S., E. Carletti, et al. (2006) CENP-E (human) A G S E R A A QBrier, S., E. Carletti, et al. (2006) CENP-E (human) A A Q T G A A GBrier, S., E. Carletti, et al. (2006) CENP-E (human) G A A G V R L KBrier, S., E. Carletti, et al. (2006) CENP-E (human) V R L K E G C NBrier, S., E. Carletti, et al. (2006) CENP-E (human) E G C N I N R SBrier, S., E. Carletti, et al. (2006) CENP-E (human) G C N I N R S LBrier, S., E. Carletti, et al. (2006) CENP-E (human) N R S L F I L GBrier, S., E. Carletti, et al. (2006) CENP-E (human) R S L F I L G QBrier, S., E. Carletti, et al. (2006) CENP-E (human) V G G F I N Y RBrier, S., E. Carletti, et al. (2006) CENP-E (human) G G F I N Y R DBrier, S., E. Carletti, et al. (2006) CENP-E (human) F I N Y R D S KBrier, S., E. Carletti, et al. (2006) CENP-E (human) D S K L T R I LBrier, S., E. Carletti, et al. (2006) CENP-E (human) N A K T R I I CBrier, S., E. Carletti, et al. (2006) CENP-E (human) R I I C T I T P
52

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Brier, S., E. Carletti, et al. (2006) CENP-E (human) I I C T I T P VBrier, S., E. Carletti, et al. (2006) CENP-E (human) I T P V S F D EBrier, S., E. Carletti, et al. (2006) CENP-E (human) P V S F D E T LBrier, S., E. Carletti, et al. (2006) CENP-E (human) T L T A L Q F ABrier, S., E. Carletti, et al. (2006) CENP-E (human) L T A L Q F A SBrier, S., E. Carletti, et al. (2006) CENP-E (human) T A L Q F A S TBrier, S., E. Carletti, et al. (2006) CENP-E (human) A L Q F A S T ABrier, S., E. Carletti, et al. (2006) CENP-E (human) F A S T A K Y MBrier, S., E. Carletti, et al. (2006) CENP-E (human) Y V N E V S T DBrier, S., E. Carletti, et al. (2006) CENP-E (human) V N E V S T D LCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein H V A F G S E DCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein F G S E D I E NCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein G S E D I E N TCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein E N T L A K M DCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein D G Q L D G L ACheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein L D G L A F G ACheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein A I Q L D G D GCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein N I L Q Y N A ACheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein I L Q Y N A A ECheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein I G K N F F K DCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein G K N F F K D VCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein S P E F Y G K FCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein S G N L N T M FCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein N L N T M F E YCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein L N T M F E Y TCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein E Y T F D Y Q MCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein Y T F D Y Q M TCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein S G D S Y W V FCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein G D S Y W V F VCheng, G., M. A. Cusanovich, et al. (2006) Photoactive yellow protein Y W V F V K R VCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus G D A A K G E KCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus H S I I A P D GCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus D G T E I V K GCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus G P N L Y G V VCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus Y P E F K Y K DCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus I V A L G A S GCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus S I V A L G A SCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus G A S G F A W TCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus A S G F A W T ECheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus W T E E D I A TCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus T E E D I A T YCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus D I A T Y V K DCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus P G A F L K E KCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus G A F L K E K LCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus T G M A F K L ACheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus E D V A A Y L ACheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus D V A A Y L A SCheng, G., V. H. Wysocki, et al. (2006) cytochrome c 2-rhodobacter capsulatus Y L A S V V KHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef G W S A I R E RHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef D G V G A V S RHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef N A D C A W L E
53

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Hochrein, J. M., T. E. Wales, et al. (2006) HIV Nef A Q E E E E V GHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef R P M T Y K A AHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef K A A L D I S HHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef I S H F L K E KHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef K G G L E G L IHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef I L D L W I Y HHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef L D L W I Y H THochrein, J. M., T. E. Wales, et al. (2006) HIV Nef Q G Y F P D W QHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef D W Q N Y T P GHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef L T F G W C F KHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef G W C F K L V PHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef K V E E A N E GHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef P M S L H G M EHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef K E V L V W R FHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef D S K L A F H HHochrein, J. M., T. E. Wales, et al. (2006) HIV Nef H P E Y Y K D CHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef H G G A I S M RHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef G D L R Q R L LHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef Y G R L L G E VHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef L S S L S C E GHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef N Q G Q Y M N THochrein, J. M., T. E. Wales, et al. (2006) SIV Nef P A E E R E K LHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef N M D D I D E EHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef D D D L V G V SHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef R T M S Y K L AHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef S Y K L A I D MHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef M S H F I K E KHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef E G I Y Y S A RHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef R I L D I Y L EHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef D I Y L E K E EHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef D W Q D Y T S GHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef F G W L W K L VHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef E A Q E D E E HHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef Q T S Q W D D PHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef Y E A Y V R Y PHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef K S G L S E E EHochrein, J. M., T. E. Wales, et al. (2006) SIV Nef L L N M A D K KShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis R A D I L Y N IShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis A D I L Y N I RShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis R P V A V S V SShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis P V A V S V S LShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis D V V F W Q Q TShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis S D R T L A W NShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis D L A A Y N A IShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis T P Q L A R V VShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis G E V L Y M P SShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis D D S E Y F S QShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis D S E Y F S Q YShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis Y S R F E I L DShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis F E I L D V T QShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis E A Y E D V E V
54
Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Shi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis E D V E V S L NShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis E V S L N F R KShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis V S L N F R K KShi, J., J. R. Koeppe, et al. (2006) AChBP from L. stagnalis S L N F R K K GTsutsui, Y., L. Liu, et al. (2006) α1AT (human) F N K I TTsutsui, Y., L. Liu, et al. (2006) α1AT (human) N L A E F A F STsutsui, Y., L. Liu, et al. (2006) α1AT (human) A F S L Y R Q LTsutsui, Y., L. Liu, et al. (2006) α1AT (human) S T N I F F S PTsutsui, Y., L. Liu, et al. (2006) α1AT (human) I A T A F A M LTsutsui, Y., L. Liu, et al. (2006) α1AT (human) A F A M L S L GTsutsui, Y., L. Liu, et al. (2006) α1AT (human) G T K A D T H DTsutsui, Y., L. Liu, et al. (2006) α1AT (human) D E I L E G L NTsutsui, Y., L. Liu, et al. (2006) α1AT (human) N F N L T E I PTsutsui, Y., L. Liu, et al. (2006) α1AT (human) Q E L L H T L NTsutsui, Y., L. Liu, et al. (2006) α1AT (human) N G L F L S E GTsutsui, Y., L. Liu, et al. (2006) α1AT (human) L K L V D K F LTsutsui, Y., L. Liu, et al. (2006) α1AT (human) H S E A F T V NTsutsui, Y., L. Liu, et al. (2006) α1AT (human) Q I N D Y V E KTsutsui, Y., L. Liu, et al. (2006) α1AT (human) I V D L V K E LTsutsui, Y., L. Liu, et al. (2006) α1AT (human) D T V F A L V NTsutsui, Y., L. Liu, et al. (2006) α1AT (human) Y I F F K G K NTsutsui, Y., L. Liu, et al. (2006) α1AT (human) E E D F H V D QTsutsui, Y., L. Liu, et al. (2006) α1AT (human) Q V T T V K V PTsutsui, Y., L. Liu, et al. (2006) α1AT (human) L G M F N I Q HTsutsui, Y., L. Liu, et al. (2006) α1AT (human) K L S S W V L LTsutsui, Y., L. Liu, et al. (2006) α1AT (human) S W V L L M K YTsutsui, Y., L. Liu, et al. (2006) α1AT (human) A T A I F F L PTsutsui, Y., L. Liu, et al. (2006) α1AT (human) I T K F L E N ETsutsui, Y., L. Liu, et al. (2006) α1AT (human) T Y D L K S V LTsutsui, Y., L. Liu, et al. (2006) α1AT (human) K S V L G Q L GTsutsui, Y., L. Liu, et al. (2006) α1AT (human) N G A D L S G VTsutsui, Y., L. Liu, et al. (2006) α1AT (human) V T E E A P L KTsutsui, Y., L. Liu, et al. (2006) α1AT (human) L K L S K A V HTsutsui, Y., L. Liu, et al. (2006) α1AT (human) A V L T I D E KTsutsui, Y., L. Liu, et al. (2006) α1AT (human) A G A M F L E ATsutsui, Y., L. Liu, et al. (2006) α1AT (human) P F V F L M I DTsutsui, Y., L. Liu, et al. (2006) α1AT (human) S P L F M G K VWales, T. E. and J. R. Engen (2006) Lyn SH3 D I V V A L Y PWales, T. E. and J. R. Engen (2006) Lyn SH3 F K K G E K M KWales, T. E. and J. R. Engen (2006) Lyn SH3 H G E W W K A KWales, T. E. and J. R. Engen (2006) Lyn SH3 E W W K A K S LWales, T. E. and J. R. Engen (2006) Lyn SH3 K S L L T K E EWales, T. E. and J. R. Engen (2006) Lyn SH3 K E G F I P S NWales, T. E. and J. R. Engen (2006) Lyn SH3 P S N Y V A K LWales, T. E. and J. R. Engen (2006) Lyn SH3 A K L N T L E TWales, T. E. and J. R. Engen (2006) α-spectrin SH3 A L Y D Y Q E KWales, T. E. and J. R. Engen (2006) α-spectrin SH3 K S P R E V T MWales, T. E. and J. R. Engen (2006) α-spectrin SH3 P R E V T M K KWales, T. E. and J. R. Engen (2006) α-spectrin SH3 G D I L T L L NWales, T. E. and J. R. Engen (2006) α-spectrin SH3 D I L T L L N SWales, T. E. and J. R. Engen (2006) α-spectrin SH3 L N S T N K D W
55

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Wales, T. E. and J. R. Engen (2006) α-spectrin SH3 T N K D W W K VWales, T. E. and J. R. Engen (2006) α-spectrin SH3 D R Q G F V P AWales, T. E. and J. R. Engen (2006) α-spectrin SH3 A A Y V K K L DWeis, D. D., P. Kjellen, et al. (2006) Lck SH3 N L V I A LWeis, D. D., P. Kjellen, et al. (2006) Lck SH3 V I A L H S Y EWeis, D. D., P. Kjellen, et al. (2006) Lck SH3 K G E Q L R I LWeis, D. D., P. Kjellen, et al. (2006) Lck SH3 G E Q L R I L EWeis, D. D., P. Kjellen, et al. (2006) Lck SH3 S G E W W K A QWeis, D. D., P. Kjellen, et al. (2006) Lck SH3 G F I P F N F VWeis, D. D., P. Kjellen, et al. (2006) Lck SH3 P F N F V A K AYao, Z. P., M. Zhou, et al. (2006) Cks1 D D E E F E Y RYao, Z. P., M. Zhou, et al. (2006) Cks1 K T H L M S E SYao, Z. P., M. Zhou, et al. (2006) Cks1 S E S E W R N LYao, Z. P., M. Zhou, et al. (2006) Cks1 Q S Q G W V H YYao, Z. P., M. Zhou, et al. (2006) Cks1 W V H Y M I H EYao, Z. P., M. Zhou, et al. (2006) Cks1 H I L L F R R PYao, Z. P., M. Zhou, et al. (2006) Skp2 G S P E F MYao, Z. P., M. Zhou, et al. (2006) Skp2 P K L N R E N FYao, Z. P., M. Zhou, et al. (2006) Skp2 P G V S W D S LYao, Z. P., M. Zhou, et al. (2006) Skp2 D E L L L G I FYao, Z. P., M. Zhou, et al. (2006) Skp2 C L P E L L K VYao, Z. P., M. Zhou, et al. (2006) Skp2 S D E S L W Q TYao, Z. P., M. Zhou, et al. (2006) Skp2 V T G R L L S QYao, Z. P., M. Zhou, et al. (2006) Skp2 V I A F R C P RYao, Z. P., M. Zhou, et al. (2006) Skp2 P L A E H F S PYao, Z. P., M. Zhou, et al. (2006) Skp2 H M D L S N S VYao, Z. P., M. Zhou, et al. (2006) Skp2 S V I E V S T LYao, Z. P., M. Zhou, et al. (2006) Skp2 H G I L S Q C SYao, Z. P., M. Zhou, et al. (2006) Skp2 N L S L E G L RYao, Z. P., M. Zhou, et al. (2006) Skp2 V N T L A K N SYao, Z. P., M. Zhou, et al. (2006) Skp2 N S N L V R L NYao, Z. P., M. Zhou, et al. (2006) Skp2 S G C S G F S EYao, Z. P., M. Zhou, et al. (2006) Skp2 F S E F A L Q TYao, Z. P., M. Zhou, et al. (2006) Skp2 Q T L L S S C SYao, Z. P., M. Zhou, et al. (2006) Skp2 S C S R L D E LYao, Z. P., M. Zhou, et al. (2006) Skp2 A H V S E T I TYao, Z. P., M. Zhou, et al. (2006) Skp2 I T Q L N L S GYao, Z. P., M. Zhou, et al. (2006) Skp2 K S D L S T L VYao, Z. P., M. Zhou, et al. (2006) Skp2 R R S P N L V HYao, Z. P., M. Zhou, et al. (2006) Skp2 L D L S D S V MYao, Z. P., M. Zhou, et al. (2006) Skp2 L K N D C F Q EYao, Z. P., M. Zhou, et al. (2006) Skp2 F Q E F F Q L NYao, Z. P., M. Zhou, et al. (2006) Skp2 E F F Q L N Y LYao, Z. P., M. Zhou, et al. (2006) Skp2 Q L N Y L Q H LYao, Z. P., M. Zhou, et al. (2006) Skp2 H L S L S R C YYao, Z. P., M. Zhou, et al. (2006) Skp2 L S R C Y D I IYao, Z. P., M. Zhou, et al. (2006) Skp2 P E T L L E L GYao, Z. P., M. Zhou, et al. (2006) Skp2 P T L K T L Q VYao, Z. P., M. Zhou, et al. (2006) Skp2 L Q V F G I V PYao, Z. P., M. Zhou, et al. (2006) Skp2 T L Q L L K E AYao, Z. P., M. Zhou, et al. (2006) Skp2 L P H L Q I N C
56

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Yao, Z. P., M. Zhou, et al. (2006) Skp2 K C R L T L Q KCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 F F F G N ICatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 I T R E E A E DCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 T R E E A E D YCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 G M S D G L Y LCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 S D G L Y L L RCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 Q S R N Y L G GCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 Y L G G F A L SCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 L G G F A L S VCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 H Y T I L N G TCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 N G T Y A I A GCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 P A D L C H Y HCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 E S D G L V C LCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 L V C L L K K PCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 F E D L K E N LCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 K E N L I R E YCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 E N L I R E Y VCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 I R E Y V K Q TCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 Q A L E Q A I LCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 A L E Q A I L SCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 Q A I L S Q K PCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 K P Q L E K L ICatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 L E K L I A T TCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 E K L I A T T ACatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 S R D E S E Q ICatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 D E S E Q I V LCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 S E Q I V L I GCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 Q I V L I G S KCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 N G K F L I R ACatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 N N G S Y A L CCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 S Y A L C L L HCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 F D T L W Q L VCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 E H Y S Y K S DCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 S D G L L R V LCatalina, M. I., M. J. Fischer, et al. (2005) SH2 domains of Syk tSH2 D G L L R V L TKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein I V T L A V D EKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein L A V D E I I EKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein A S M Q R S S NKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein I W M P V E Q EKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein P V E Q E S P TKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein W D L T D K A TKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein A T G L L E L NKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein D N D F F Q L RKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein D L R D E T A YKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein N N V E L K V AKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein N V E L K V A NKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein G S L V I T S PKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein D A W N F V A DKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein E I M F S R E LKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein G T S Y F F N PKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein D I F G R I P E
57
Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Kang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein P E E A Y R D GKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein V A G F D D V LKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein V D N R F A T VKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein T V T L S A T TKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein K I S F A G V KKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein A K N V L A Q DKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein D A T F S V V RKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein P V A L D D V SKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein A D A M A V N IKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein T N V F W A D DKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein W A D D A I R IKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein H E L F A G M KKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein F A G M K T T SKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein N G I F A T Q GKang, S. and P. E. Prevelige, Jr. (2005) P22 capsid coat protein L S G L C R I ALee, T., A. N. Hoofnagle, et al. (2005) ERK2 A G P E M V R GLee, T., A. N. Hoofnagle, et al. (2005) ERK2 G M V C S A Y DLee, T., A. N. Hoofnagle, et al. (2005) ERK2 V C S A Y D N LLee, T., A. N. Hoofnagle, et al. (2005) ERK2 Y D N L N K V RLee, T., A. N. Hoofnagle, et al. (2005) ERK2 T L R E I K I LLee, T., A. N. Hoofnagle, et al. (2005) ERK2 I K I L L R F RLee, T., A. N. Hoofnagle, et al. (2005) ERK2 G I N D I I R ALee, T., A. N. Hoofnagle, et al. (2005) ERK2 K D V Y I V Q DLee, T., A. N. Hoofnagle, et al. (2005) ERK2 V Q D L M E T DLee, T., A. N. Hoofnagle, et al. (2005) ERK2 D L M E T D L YLee, T., A. N. Hoofnagle, et al. (2005) ERK2 E T D L Y K L LLee, T., A. N. Hoofnagle, et al. (2005) ERK2 H I C Y F L Y QLee, T., A. N. Hoofnagle, et al. (2005) ERK2 A N V L H R D LLee, T., A. N. Hoofnagle, et al. (2005) ERK2 P S N L L L N TLee, T., A. N. Hoofnagle, et al. (2005) ERK2 S N L L L N T TLee, T., A. N. Hoofnagle, et al. (2005) ERK2 T T C D L K I CLee, T., A. N. Hoofnagle, et al. (2005) ERK2 T C D L K I C DLee, T., A. N. Hoofnagle, et al. (2005) ERK2 D F G L A R V ALee, T., A. N. Hoofnagle, et al. (2005) ERK2 A T R W Y R A PLee, T., A. N. Hoofnagle, et al. (2005) ERK2 P E I N L N S KLee, T., A. N. Hoofnagle, et al. (2005) ERK2 E I N L N S K GLee, T., A. N. Hoofnagle, et al. (2005) ERK2 W S V G C I L ALee, T., A. N. Hoofnagle, et al. (2005) ERK2 L A E M L S N RLee, T., A. N. Hoofnagle, et al. (2005) ERK2 P S Q E D L N CLee, T., A. N. Hoofnagle, et al. (2005) ERK2 Q E D L N C I ILee, T., A. N. Hoofnagle, et al. (2005) ERK2 I I N L K A R NLee, T., A. N. Hoofnagle, et al. (2005) ERK2 R N Y L L S L PLee, T., A. N. Hoofnagle, et al. (2005) ERK2 A L D L L D K MLee, T., A. N. Hoofnagle, et al. (2005) ERK2 Y L E Q Y Y D PLee, T., A. N. Hoofnagle, et al. (2005) ERK2 D M E L D D L PLee, T., A. N. Hoofnagle, et al. (2005) ERK2 E L I F E E T ALee, T., A. N. Hoofnagle, et al. (2005) ERK2 I C D F G L A RBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) K N I Q V V V RBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N I Q V V V R CBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) V R C R P F N LBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) P F N K A E R K
58

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Brier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) K A S A H S I VBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S A H S I V E CBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S I V E C D P VBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) V E C D P V R KBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) K E V S V R T GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) V R T G G L A DBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) T G G L A D K SBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S S R K T Y T FBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) T Y T F D M V FBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Y T F D M V F GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) F D M V F G A SBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Q I D V Y R S VBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S V V C P I L DBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) C P I L D E V IBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) P I L D E V I MBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) I L D E V I M GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) D E V I M G Y NBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) M G Y N C T I FBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N C T I F A Y GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) C T I F A Y G QBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Y G Q T G T G KBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) P N E E Y T W EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N E E Y T W E EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) E E Y T W E E DBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Y T W E E D P LBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) I I P R T L H QBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) P R T L H Q I FBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) T L H Q I F E KBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) H Q I F E K L TBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Q I F E K L T DBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) K L T D N G T EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N G T E F S V KBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) G T E F S V K VBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) T E F S V K V SBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) K V S L L E I YBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) V S L L E I Y NBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N E E L F D L LBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N P S S D V S EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) D V S E R L Q MBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) R L Q M F D D PBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) K R G V I I K GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) G L E E I T V HBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) V H N K D E V YBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) D E V Y Q I L EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) E V Y Q I L E KBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Y Q I L E K G ABrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Q I L E K G A ABrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) A A T L M N A YBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) A T L M N A Y SBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) L M N A Y S S RBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S S R S H S V F
59

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Brier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) H S V F S V T IBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) T I H M K E T TBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) T I D G E E L VBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) D G E E L V K IBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) G E E L V K I GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) E E L V K I G KBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) K L N L V D L ABrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) L N L V D L A GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N I G R S G A VBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) R E A G N I N QBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) N Q S L L T L GBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Q S L L T L G RBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) L L T L G R V IBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) L T L G R V I TBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) R V I T A L V EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) V I T A L V E RBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) I T A L V E R TBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) R E S K L T R IBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) E S K L T R I LBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) L Q D S L G G RBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) Q D S L G G R TBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) D S L G G R T RBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) G G R T R T S IBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) R T S I I A T IBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) T S I I A T I SBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S I I A T I S PBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) I I A T I S P ABrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) I A T I S P A SBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) I S P A S L N LBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S P A S L N L EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S L N L E E T LBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) L N L E E T L SBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) E E T L S T L EBrier, S., D. Lemaire, et al. (2004) Kinesin-like protein KIF11 (human) S T L E Y A G RCasbarra, A., L. Birolo, et al. (2004) Human α-LA L S Q L L K D ICasbarra, A., L. Birolo, et al. (2004) Human α-LA G I A L P E L ICasbarra, A., L. Birolo, et al. (2004) Human α-LA C T M F H T S GCasbarra, A., L. Birolo, et al. (2004) Human α-LA Y G L F Q I S NCasbarra, A., L. Birolo, et al. (2004) Human α-LA S N K L W C K SCasbarra, A., L. Birolo, et al. (2004) Human α-LA K I L D I K G ICasbarra, A., L. Birolo, et al. (2004) Human α-LA G I D Y W L A HCasbarra, A., L. Birolo, et al. (2004) Human α-LA L E Q W L C E KCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin R P L F E K K SCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin T E R E L L E SCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin L E S Y I D G RCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin E G S D A E I GCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin Q V M L F R K SCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin Q E L L C G A SCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin N D L L V R I GCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin D L L V R I G KCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin E R N I E K I S
60
Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Croy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin E K I S M L E KCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin I S M L E K I YCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin S M L E K I Y ICroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin Y N W R E N L DCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin R E N L D R D ICroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin D I A L M K L KCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin F S D Y I H P VCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin A A S L L Q A GCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin A S L L Q A G YCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin G N L K E T W TCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin G G P F V M K SCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin P F V M K S P FCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin F N N R W Y Q MCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin K Y G F Y T H VCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin T H V F R K K KCroy, C. H., J. R. Koeppe, et al. (2004) human α-thrombin I D Q F G ECroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin Q P F F N E K TCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin V M L F R K S PCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin Q E L L C G A SCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin A S L I S D R WCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin W V L T A A H CCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin D D L L V R I GCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin K E N L D R D ICroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin D I A L L K L KCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin R P I E L S D YCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin L S D Y I H P VCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin H A G F K G R VCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin T T S V A E V QCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin T S V A E V Q PCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin K A S T R I R ICroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin T D N M F C A GCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin D N M F C A G YCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin G G P F V M K SCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin P F V M K S P YCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin K S P Y N N R WCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin Y N N R W Y Q MCroy, C. H., J. R. Koeppe, et al. (2004) bovine α-thrombin G I V S W G E GCroy, C. H., S. Bergqvist, et al. (2004) IκBα D S F L H L A ICroy, C. H., S. Bergqvist, et al. (2004) IκBα F L H L A I I HCroy, C. H., S. Bergqvist, et al. (2004) IκBα A L T M E V I RCroy, C. H., S. Bergqvist, et al. (2004) IκBα L T M E V I R QCroy, C. H., S. Bergqvist, et al. (2004) IκBα M E V I R Q V KCroy, C. H., S. Bergqvist, et al. (2004) IκBα K G D L A F L NCroy, C. H., S. Bergqvist, et al. (2004) IκBα D L A F L N F QCroy, C. H., S. Bergqvist, et al. (2004) IκBα L A F L N F Q NCroy, C. H., S. Bergqvist, et al. (2004) IκBα F L N F Q N N LCroy, C. H., S. Bergqvist, et al. (2004) IκBα P L H L A V I TCroy, C. H., S. Bergqvist, et al. (2004) IκBα E I A E A L L GCroy, C. H., S. Bergqvist, et al. (2004) IκBα A G C D P E L RCroy, C. H., S. Bergqvist, et al. (2004) IκBα D P E L R D F RCroy, C. H., S. Bergqvist, et al. (2004) IκBα E L R D F R G N
61

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Croy, C. H., S. Bergqvist, et al. (2004) IκBα G N T R L H L ACroy, C. H., S. Bergqvist, et al. (2004) IκBα T P K H L A C ECroy, C. H., S. Bergqvist, et al. (2004) IκBα P L H L A C E QCroy, C. H., S. Bergqvist, et al. (2004) IκBα E Q G C L A S VCroy, C. H., S. Bergqvist, et al. (2004) IκBα L A S V G V L TCroy, C. H., S. Bergqvist, et al. (2004) IκBα L H S I L K A TCroy, C. H., S. Bergqvist, et al. (2004) IκBα G H T L L H L ACroy, C. H., S. Bergqvist, et al. (2004) IκBα H T L L H L A SCroy, C. H., S. Bergqvist, et al. (2004) IκBα G I V E L L V SCroy, C. H., S. Bergqvist, et al. (2004) IκBα I V E L L V S KCroy, C. H., S. Bergqvist, et al. (2004) IκBα G R T A L H L ACroy, C. H., S. Bergqvist, et al. (2004) IκBα A L H L A V D KCroy, C. H., S. Bergqvist, et al. (2004) IκBα R P S T R I Q QCroy, C. H., S. Bergqvist, et al. (2004) IκBα Q L G Q L T L ECroy, C. H., S. Bergqvist, et al. (2004) IκBα E S E F T E F TCroy, C. H., S. Bergqvist, et al. (2004) IκBα Y D D C V F G GLi, X., Y. T. Chou, et al. (2004) LR3IGF-I M P S L F V N GLi, X., Y. T. Chou, et al. (2004) LR3IGF-I P S L F V N G PLi, X., Y. T. Chou, et al. (2004) LR3IGF-I C G A E L V D ALi, X., Y. T. Chou, et al. (2004) LR3IGF-I G A E L V D A LLi, X., Y. T. Chou, et al. (2004) LR3IGF-I L V D A L Q F VLi, X., Y. T. Chou, et al. (2004) LR3IGF-I V D A L Q F V CLi, X., Y. T. Chou, et al. (2004) LR3IGF-I D A L Q E V C GLi, X., Y. T. Chou, et al. (2004) LR3IGF-I D R G F Y F N KLi, X., Y. T. Chou, et al. (2004) LR3IGF-I R G F Y F N K PLi, X., Y. T. Chou, et al. (2004) LR3IGF-I F Y F N K P T GLi, X., Y. T. Chou, et al. (2004) LR3IGF-I Q T G I V D E CLi, X., Y. T. Chou, et al. (2004) LR3IGF-I V D E C C F R SLi, X., Y. T. Chou, et al. (2004) LR3IGF-I R L E M Y C A PMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) Y P D L S K H NMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) T P D L Y K K LMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) E T P S G F T LMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) P S G F T L D DMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) F T L D D V I QMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) T L D D V I Q TMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) F I M T V G C VMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) M T V G C V A GMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) D E E S Y T V FMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) Y T V F K D L FMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) D L F D P I I QMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) H E N L K G G DMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) H Y V L S S R VMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) K L S V E A L NMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) V E A L N S L TMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) Q Q Q L I D D HMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) D D H F L F D KMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) D H F L F D K PMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) S P L L L A S GMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) P L L L A S G MMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) N K S F L V W VMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) K S F L V W V N
62

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Mazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) E D H L R V I SMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) V I S M E K Q GMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) K E V F R R F CMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) F C V G L Q K IMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) K I E E I F K KMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) H P F M W N E HMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) H L G Y V L T CMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) G Y V L T C P SMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) V L T C P S N LMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) P S N L G T G LMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) H P K F E E I LMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) E E I L T R L RMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) L T R L R L Q KMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) A A V G S V F DMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) G S V F D I S NMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) S E V E Q V Q LMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) E V E Q V Q L VMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) Q V Q L V V D GMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) G V K L M V E MMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) L M V E M E K KMazon, H., O. Marcillat, et al. (2004) Creatine kinase M-type (rabbit) M V E M E K K LWu, Z., A. Hasan, et al. (2004) human rCRALBP D K H M S E G VWu, Z., A. Hasan, et al. (2004) human rCRALBP Y V N F R L Q YWu, Z., A. Hasan, et al. (2004) human rCRALBP Q A E N T A FYan, X., D. Broderick, et al. (2004) human RXRα LBD M P V E R I L EYan, X., D. Broderick, et al. (2004) human RXRα LBD P V E R I L E AYan, X., D. Broderick, et al. (2004) human RXRα LBD E R I L E A E LYan, X., D. Broderick, et al. (2004) human RXRα LBD I L E A E L A VYan, X., D. Broderick, et al. (2004) human RXRα LBD L E A E L A V EYan, X., D. Broderick, et al. (2004) human RXRα LBD E A E L A V E PYan, X., D. Broderick, et al. (2004) human RXRα LBD K T E T Y V E AYan, X., D. Broderick, et al. (2004) human RXRα LBD V E A N M G L NYan, X., D. Broderick, et al. (2004) human RXRα LBD N M G L N P S SYan, X., D. Broderick, et al. (2004) human RXRα LBD N I C Q A A D KYan, X., D. Broderick, et al. (2004) human RXRα LBD Q L F T L V F WYan, X., D. Broderick, et al. (2004) human RXRα LBD L F T L V E W AYan, X., D. Broderick, et al. (2004) human RXRα LBD T L V E W A K RYan, X., D. Broderick, et al. (2004) human RXRα LBD I P H F S E L PYan, X., D. Broderick, et al. (2004) human RXRα LBD P H F S E L P LYan, X., D. Broderick, et al. (2004) human RXRα LBD F S E L P L D DYan, X., D. Broderick, et al. (2004) human RXRα LBD P L D D Q V I LYan, X., D. Broderick, et al. (2004) human RXRα LBD L D D Q V I L LYan, X., D. Broderick, et al. (2004) human RXRα LBD Q V I L L R A GYan, X., D. Broderick, et al. (2004) human RXRα LBD W N E L L I A SYan, X., D. Broderick, et al. (2004) human RXRα LBD N E L L I A S FYan, X., D. Broderick, et al. (2004) human RXRα LBD L I A S F S H RYan, X., D. Broderick, et al. (2004) human RXRα LBD I A S F S H R SYan, X., D. Broderick, et al. (2004) human RXRα LBD R S I A V K D GYan, X., D. Broderick, et al. (2004) human RXRα LBD D G I L L A T GYan, X., D. Broderick, et al. (2004) human RXRα LBD G I L L A T G LYan, X., D. Broderick, et al. (2004) human RXRα LBD A T G L H V H R
63

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Yan, X., D. Broderick, et al. (2004) human RXRα LBD A G V G A I F DYan, X., D. Broderick, et al. (2004) human RXRα LBD G A I F D R V LYan, X., D. Broderick, et al. (2004) human RXRα LBD A I F D R V L TYan, X., D. Broderick, et al. (2004) human RXRα LBD V L T E L V S KYan, X., D. Broderick, et al. (2004) human RXRα LBD L T E L V S K MYan, X., D. Broderick, et al. (2004) human RXRα LBD D M Q M D K T EYan, X., D. Broderick, et al. (2004) human RXRα LBD K T E L G C L RYan, X., D. Broderick, et al. (2004) human RXRα LBD L G C L R A I VYan, X., D. Broderick, et al. (2004) human RXRα LBD C L R A I V L FYan, X., D. Broderick, et al. (2004) human RXRα LBD A I V L F N P DYan, X., D. Broderick, et al. (2004) human RXRα LBD N P A E V E A LYan, X., D. Broderick, et al. (2004) human RXRα LBD A E V E A L R EYan, X., D. Broderick, et al. (2004) human RXRα LBD E V E A L R E KYan, X., D. Broderick, et al. (2004) human RXRα LBD A L R E K V Y AYan, X., D. Broderick, et al. (2004) human RXRα LBD Y A S L E A Y CYan, X., D. Broderick, et al. (2004) human RXRα LBD S L E A Y C K HYan, X., D. Broderick, et al. (2004) human RXRα LBD L E A Y C K H KYan, X., D. Broderick, et al. (2004) human RXRα LBD P G R F A K L LYan, X., D. Broderick, et al. (2004) human RXRα LBD F A K L L L R LYan, X., D. Broderick, et al. (2004) human RXRα LBD A K L L L R L PYan, X., D. Broderick, et al. (2004) human RXRα LBD L L R L P A L RYan, X., D. Broderick, et al. (2004) human RXRα LBD L P A L R S I GYan, X., D. Broderick, et al. (2004) human RXRα LBD A L R S I G L KYan, X., D. Broderick, et al. (2004) human RXRα LBD R S I G L K C LYan, X., D. Broderick, et al. (2004) human RXRα LBD S I G L K C L EYan, X., D. Broderick, et al. (2004) human RXRα LBD G L K C L E H LYan, X., D. Broderick, et al. (2004) human RXRα LBD K C L E H L F FYan, X., D. Broderick, et al. (2004) human RXRα LBD H L F F F K L IYan, X., D. Broderick, et al. (2004) human RXRα LBD P I D T F L M EYan, X., D. Broderick, et al. (2004) human RXRα LBD T F L M E M L EAnand, G. S., D. Law, et al. (2003) PKA - R1α A I S A E V Y TAnand, G. S., D. Law, et al. (2003) PKA - R1α D A A S Y V R KAnand, G. S., D. Law, et al. (2003) PKA - R1α K D Y K T M A AAnand, G. S., D. Law, et al. (2003) PKA - R1α T M A A L A K AAnand, G. S., D. Law, et al. (2003) PKA - R1α E K N V L F S HAnand, G. S., D. Law, et al. (2003) PKA - R1α K N V L F S H LAnand, G. S., D. Law, et al. (2003) PKA - R1α F S H L D D N EAnand, G. S., D. Law, et al. (2003) PKA - R1α L D D N E R S DAnand, G. S., D. Law, et al. (2003) PKA - R1α D D N E R S D IAnand, G. S., D. Law, et al. (2003) PKA - R1α R S D I F D A MAnand, G. S., D. Law, et al. (2003) PKA - R1α D E G D N F Y VAnand, G. S., D. Law, et al. (2003) PKA - R1α S F G E L A L IAnand, G. S., D. Law, et al. (2003) PKA - R1α E K A L I Y G TAnand, G. S., D. Law, et al. (2003) PKA - R1α N V K L W G I DAnand, G. S., D. Law, et al. (2003) PKA - R1α D S Y R R I L MAnand, G. S., D. Law, et al. (2003) PKA - R1α M G S T L R K RAnand, G. S., D. Law, et al. (2003) PKA - R1α G S T L R K R KAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit S V K E F L A KAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit A K E D L L K KAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit N T A Q D D Q FAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit T A Q L D Q F D
64

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Anand, G. S., D. Law, et al. (2003) PKA - C-Subunit L D Q F F R I KAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit G T G S G G R VAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit H K E S V N H YAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit K Q K V K K L KAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit Q K V V Q L K QAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit V K L K K I E HAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit T L N E P R I LAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit A V N F M F L VAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit A G G E R F S HAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit F S H L S R I GAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit I G R F Y E P HAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit H A R F Y A A QAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit L D L I L R D LAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit K P E N D L I DAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit N L L I S Q Q GAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit E I I L W K G YAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit A V D W F A L GAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit F P S H D S S DAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit H F S S L L K DAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit F S S D T K D LAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit G V D L I K R FAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit T T D W F A I YAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit D T S N D D D YAnand, G. S., D. Law, et al. (2003) PKA - C-Subunit T S N F D D Y EChik, J. K. and D. C. Schriemer (2003) PKA - C-Subunit T T A L V C D NChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin G S G L V K A GChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin R A V F P S I VChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin Q G V M V G M GChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin V M V G M G Q KChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin R G I L T L K YChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin G I L T L K Y PChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin W D D M E K I WChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin F Y N E L R V AChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin Y N E L R V A PChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin H P T L L T E AChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin K M T Q I M F EChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin T Q I M F E T FChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin Q I M F E T F NChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin V L S L Y A S GChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin G I V L D S G DChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin I Y E G Y A K PChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin I M R L D L A GChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin T D Y L M K I LChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin R S Y S F V T TChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin S Y S F V T T AChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin T T A E R E I VChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin A E R E I V R DChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin K L C Y V A L DChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin I G N E R F R CChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin Q P S F I G M EChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin N S I M K C D I
65

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Chik, J. K. and D. C. Schriemer (2003) rabbit muscle actin I M K C D I D IChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin Y A N N V M S GChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin K E I T A L A PChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin P S T M K I K IChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin Y S V W I G G SChik, J. K. and D. C. Schriemer (2003) rabbit muscle actin F Q Q M W I T KCravello, L., D. Lascoux, et al. (2003) PBP-2X* S P E F G T G TCravello, L., D. Lascoux, et al. (2003) PBP-2X* F G T D L A K ECravello, L., D. Lascoux, et al. (2003) PBP-2X* G T D L A K E ACravello, L., D. Lascoux, et al. (2003) PBP-2X* K E A K K V H QCravello, L., D. Lascoux, et al. (2003) PBP-2X* T V P A K R G TCravello, L., D. Lascoux, et al. (2003) PBP-2X* I A E D A T S YCravello, L., D. Lascoux, et al. (2003) PBP-2X* Y N V Y A V I DCravello, L., D. Lascoux, et al. (2003) PBP-2X* N V Y A V I D ECravello, L., D. Lascoux, et al. (2003) PBP-2X* V I D E N Y K SCravello, L., D. Lascoux, et al. (2003) PBP-2X* I D E N Y K S ACravello, L., D. Lascoux, et al. (2003) PBP-2X* E N Y K S A T GCravello, L., D. Lascoux, et al. (2003) PBP-2X* G K I L Y V E KCravello, L., D. Lascoux, et al. (2003) PBP-2X* K I L Y V E K TCravello, L., D. Lascoux, et al. (2003) PBP-2X* K T Q F N K V ACravello, L., D. Lascoux, et al. (2003) PBP-2X* K V A E V F H KCravello, L., D. Lascoux, et al. (2003) PBP-2X* V A E V F H K YCravello, L., D. Lascoux, et al. (2003) PBP-2X* Y L D M E E S YCravello, L., D. Lascoux, et al. (2003) PBP-2X* M E E S Y V R ECravello, L., D. Lascoux, et al. (2003) PBP-2X* E E S Y V R E QCravello, L., D. Lascoux, et al. (2003) PBP-2X* Q P N L K Q V SCravello, L., D. Lascoux, et al. (2003) PBP-2X* A K G N G I T YCravello, L., D. Lascoux, et al. (2003) PBP-2X* G N G I T Y A NCravello, L., D. Lascoux, et al. (2003) PBP-2X* Y A N M M S I KCravello, L., D. Lascoux, et al. (2003) PBP-2X* K K E L E A A ECravello, L., D. Lascoux, et al. (2003) PBP-2X* K E L E A A E VCravello, L., D. Lascoux, et al. (2003) PBP-2X* E L E A A E V KCravello, L., D. Lascoux, et al. (2003) PBP-2X* L E A A E V K GCravello, L., D. Lascoux, et al. (2003) PBP-2X* E A A E V K G ICravello, L., D. Lascoux, et al. (2003) PBP-2X* K G I D F T T SCravello, L., D. Lascoux, et al. (2003) PBP-2X* G I D F T T S PCravello, L., D. Lascoux, et al. (2003) PBP-2X* S Y P N G Q F ACravello, L., D. Lascoux, et al. (2003) PBP-2X* Y P N G Q F A SCravello, L., D. Lascoux, et al. (2003) PBP-2X* P N G Q F A S SCravello, L., D. Lascoux, et al. (2003) PBP-2X* A S S F I G L ACravello, L., D. Lascoux, et al. (2003) PBP-2X* S F I G L A Q LCravello, L., D. Lascoux, et al. (2003) PBP-2X* I G L Q A L H ECravello, L., D. Lascoux, et al. (2003) PBP-2X* L A Q L H E N ECravello, L., D. Lascoux, et al. (2003) PBP-2X* Q L H E N E D GCravello, L., D. Lascoux, et al. (2003) PBP-2X* K S L L G T S GCravello, L., D. Lascoux, et al. (2003) PBP-2X* M E S S L N L ICravello, L., D. Lascoux, et al. (2003) PBP-2X* N S I L A G T DCravello, L., D. Lascoux, et al. (2003) PBP-2X* L A G T D G I ICravello, L., D. Lascoux, et al. (2003) PBP-2X* G I I T Y E K DCravello, L., D. Lascoux, et al. (2003) PBP-2X* P G T E Q V S QCravello, L., D. Lascoux, et al. (2003) PBP-2X* G T E Q V S Q R
66

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Cravello, L., D. Lascoux, et al. (2003) PBP-2X* M D G K D V Y TCravello, L., D. Lascoux, et al. (2003) PBP-2X* D G K D V Y T TCravello, L., D. Lascoux, et al. (2003) PBP-2X* K D V Y T T I SCravello, L., D. Lascoux, et al. (2003) PBP-2X* L Q S E M E T QCravello, L., D. Lascoux, et al. (2003) PBP-2X* Q S E M E T Q MCravello, L., D. Lascoux, et al. (2003) PBP-2X* Q M D A F Q F KCravello, L., D. Lascoux, et al. (2003) PBP-2X* F Q F K V K G KCravello, L., D. Lascoux, et al. (2003) PBP-2X* T A T L V S A KCravello, L., D. Lascoux, et al. (2003) PBP-2X* S A K T G E I LCravello, L., D. Lascoux, et al. (2003) PBP-2X* G E I L A T T QCravello, L., D. Lascoux, et al. (2003) PBP-2X* L A T T Q R P TCravello, L., D. Lascoux, et al. (2003) PBP-2X* T Q R P T F D ACravello, L., D. Lascoux, et al. (2003) PBP-2X* R P T F D A D TCravello, L., D. Lascoux, et al. (2003) PBP-2X* T K E G I T E DCravello, L., D. Lascoux, et al. (2003) PBP-2X* I T E D F V W RCravello, L., D. Lascoux, et al. (2003) PBP-2X* T E D F V W R DCravello, L., D. Lascoux, et al. (2003) PBP-2X* R D I L Y Q S NCravello, L., D. Lascoux, et al. (2003) PBP-2X* G S T M K V M MCravello, L., D. Lascoux, et al. (2003) PBP-2X* M K V M M L A ACravello, L., D. Lascoux, et al. (2003) PBP-2X* K V M M L A A ACravello, L., D. Lascoux, et al. (2003) PBP-2X* V M M L A A A ICravello, L., D. Lascoux, et al. (2003) PBP-2X* I D N N T F P GCravello, L., D. Lascoux, et al. (2003) PBP-2X* G E V F N S S ECravello, L., D. Lascoux, et al. (2003) PBP-2X* N S S E L K I ACravello, L., D. Lascoux, et al. (2003) PBP-2X* I A D A T I R DCravello, L., D. Lascoux, et al. (2003) PBP-2X* T I R D W D V NCravello, L., D. Lascoux, et al. (2003) PBP-2X* A T I R D W D VCravello, L., D. Lascoux, et al. (2003) PBP-2X* I R D W D V N ECravello, L., D. Lascoux, et al. (2003) PBP-2X* E G L T G G R MCravello, L., D. Lascoux, et al. (2003) PBP-2X* G G R M M T F SCravello, L., D. Lascoux, et al. (2003) PBP-2X* G R M M T F S QCravello, L., D. Lascoux, et al. (2003) PBP-2X* M M T F S Q G FCravello, L., D. Lascoux, et al. (2003) PBP-2X* G M T L L E Q KCravello, L., D. Lascoux, et al. (2003) PBP-2X* D A T W L D Y LCravello, L., D. Lascoux, et al. (2003) PBP-2X* A T W L D Y L NCravello, L., D. Lascoux, et al. (2003) PBP-2X* F G V P T R F GCravello, L., D. Lascoux, et al. (2003) PBP-2X* P T R F G L T DCravello, L., D. Lascoux, et al. (2003) PBP-2X* L T D E Y A G QCravello, L., D. Lascoux, et al. (2003) PBP-2X* A D N I V N I ACravello, L., D. Lascoux, et al. (2003) PBP-2X* V N I A Q S S FCravello, L., D. Lascoux, et al. (2003) PBP-2X* A F T A I A N DCravello, L., D. Lascoux, et al. (2003) PBP-2X* A N D G V M L ECravello, L., D. Lascoux, et al. (2003) PBP-2X* N D G V M L E PCravello, L., D. Lascoux, et al. (2003) PBP-2X* F I S A I Y D PCravello, L., D. Lascoux, et al. (2003) PBP-2X* I Y D P N D Q TCravello, L., D. Lascoux, et al. (2003) PBP-2X* D Q T A R K S QCravello, L., D. Lascoux, et al. (2003) PBP-2X* Q T A R K S Q KCravello, L., D. Lascoux, et al. (2003) PBP-2X* R K S Q K E I VCravello, L., D. Lascoux, et al. (2003) PBP-2X* K S Q K E I V GCravello, L., D. Lascoux, et al. (2003) PBP-2X* I V G N P V S KCravello, L., D. Lascoux, et al. (2003) PBP-2X* N P V S K D A A
67

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Cravello, L., D. Lascoux, et al. (2003) PBP-2X* A A S L T R T NCravello, L., D. Lascoux, et al. (2003) PBP-2X* S L T R T N M VCravello, L., D. Lascoux, et al. (2003) PBP-2X* T N M V L V G TCravello, L., D. Lascoux, et al. (2003) PBP-2X* N M V L V G T DCravello, L., D. Lascoux, et al. (2003) PBP-2X* D P V Y G T M YCravello, L., D. Lascoux, et al. (2003) PBP-2X* Y G T M Y N H SCravello, L., D. Lascoux, et al. (2003) PBP-2X* G T M Y N H S TCravello, L., D. Lascoux, et al. (2003) PBP-2X* P G Q N V A L KCravello, L., D. Lascoux, et al. (2003) PBP-2X* G T A Q I A D ECravello, L., D. Lascoux, et al. (2003) PBP-2X* Q I A D E K N GCravello, L., D. Lascoux, et al. (2003) PBP-2X* G G Y L V G L TCravello, L., D. Lascoux, et al. (2003) PBP-2X* V G L T D Y I FCravello, L., D. Lascoux, et al. (2003) PBP-2X* D Y I F S A V SCravello, L., D. Lascoux, et al. (2003) PBP-2X* Y I F S A V S MCravello, L., D. Lascoux, et al. (2003) PBP-2X* I F S A V S M SCravello, L., D. Lascoux, et al. (2003) PBP-2X* A V S M S P A ECravello, L., D. Lascoux, et al. (2003) PBP-2X* N P D F I L Y VCravello, L., D. Lascoux, et al. (2003) PBP-2X* D F I L Y V T VCravello, L., D. Lascoux, et al. (2003) PBP-2X* I L Y V T V Q QCravello, L., D. Lascoux, et al. (2003) PBP-2X* V T V Q Q P E HCravello, L., D. Lascoux, et al. (2003) PBP-2X* Y S G I Q L G ECravello, L., D. Lascoux, et al. (2003) PBP-2X* I Q L G E F A NCravello, L., D. Lascoux, et al. (2003) PBP-2X* L G E F A N P ICravello, L., D. Lascoux, et al. (2003) PBP-2X* A N P I L E R ACravello, L., D. Lascoux, et al. (2003) PBP-2X* N P I L E R A SCravello, L., D. Lascoux, et al. (2003) PBP-2X* A S A M K D S LCravello, L., D. Lascoux, et al. (2003) PBP-2X* S L N L Q T T ACravello, L., D. Lascoux, et al. (2003) PBP-2X* L N L Q T T A KCravello, L., D. Lascoux, et al. (2003) PBP-2X* A K A L E Q V SCravello, L., D. Lascoux, et al. (2003) PBP-2X* P G D L A E E LCravello, L., D. Lascoux, et al. (2003) PBP-2X* G D L A E E L RCravello, L., D. Lascoux, et al. (2003) PBP-2X* L V Q P I V V GCravello, L., D. Lascoux, et al. (2003) PBP-2X* V G T G T K I KCravello, L., D. Lascoux, et al. (2003) PBP-2X* T G T K I K N SCravello, L., D. Lascoux, et al. (2003) PBP-2X* T K I K N S S ACravello, L., D. Lascoux, et al. (2003) PBP-2X* A E E G K N L ACravello, L., D. Lascoux, et al. (2003) PBP-2X* V L I L S D K ACravello, L., D. Lascoux, et al. (2003) PBP-2X* E E V P D M Y GCravello, L., D. Lascoux, et al. (2003) PBP-2X* P D M Y G W T KCravello, L., D. Lascoux, et al. (2003) PBP-2X* G W T K E T A ECravello, L., D. Lascoux, et al. (2003) PBP-2X* T A E T L A K WCravello, L., D. Lascoux, et al. (2003) PBP-2X* A E T L A K W LCravello, L., D. Lascoux, et al. (2003) PBP-2X* K W L N I E L ECravello, L., D. Lascoux, et al. (2003) PBP-2X* N I E L E F Q GCravello, L., D. Lascoux, et al. (2003) PBP-2X* D V R A N T A ICravello, L., D. Lascoux, et al. (2003) PBP-2X* A I K D I K K ICravello, L., D. Lascoux, et al. (2003) PBP-2X* T L T L G DRist, W., T. J. Jorgensen, et al. (2003) σ 32 M Q S L A L A PRist, W., T. J. Jorgensen, et al. (2003) σ 32 V G N L D S Y IRist, W., T. J. Jorgensen, et al. (2003) σ 32 N L D S Y I R ARist, W., T. J. Jorgensen, et al. (2003) σ 32 W P M L S A D E
68

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Rist, W., T. J. Jorgensen, et al. (2003) σ 32 D L E A A K T LRist, W., T. J. Jorgensen, et al. (2003) σ 32 A A K T L I L SRist, W., T. J. Jorgensen, et al. (2003) σ 32 H L R F V V H IRist, W., T. J. Jorgensen, et al. (2003) σ 32 Q A D L I Q E GRist, W., T. J. Jorgensen, et al. (2003) σ 32 N I G L M K A VRist, W., T. J. Jorgensen, et al. (2003) σ 32 L M K A V R R FRist, W., T. J. Jorgensen, et al. (2003) σ 32 P E V G V R L VRist, W., T. J. Jorgensen, et al. (2003) σ 32 G V R L V S F ARist, W., T. J. Jorgensen, et al. (2003) σ 32 R L V S F A V HRist, W., T. J. Jorgensen, et al. (2003) σ 32 L V S F A V H WRist, W., T. J. Jorgensen, et al. (2003) σ 32 E I H E Y V L RRist, W., T. J. Jorgensen, et al. (2003) σ 32 V L R N W R I VRist, W., T. J. Jorgensen, et al. (2003) σ 32 K L F F N L R KRist, W., T. J. Jorgensen, et al. (2003) σ 32 W F N Q D E V ERist, W., T. J. Jorgensen, et al. (2003) σ 32 N Q D E V E M VRist, W., T. J. Jorgensen, et al. (2003) σ 32 D E V E M V A RRist, W., T. J. Jorgensen, et al. (2003) σ 32 E V E M V A R ERist, W., T. J. Jorgensen, et al. (2003) σ 32 E S R M A A Q DRist, W., T. J. Jorgensen, et al. (2003) σ 32 Q D M T F D L SRist, W., T. J. Jorgensen, et al. (2003) σ 32 D M T F D L S SRist, W., T. J. Jorgensen, et al. (2003) σ 32 A P V L Y L Q DRist, W., T. J. Jorgensen, et al. (2003) σ 32 K S S N F A D GRist, W., T. J. Jorgensen, et al. (2003) σ 32 S S N F A D G IRist, W., T. J. Jorgensen, et al. (2003) σ 32 I E D D N W E ERist, W., T. J. Jorgensen, et al. (2003) σ 32 R S Q D I I R ARist, W., T. J. Jorgensen, et al. (2003) σ 32 R A R W L D E DRist, W., T. J. Jorgensen, et al. (2003) σ 32 K S T L Q E L ARist, W., T. J. Jorgensen, et al. (2003) σ 32 L Q E L A D R YRist, W., T. J. Jorgensen, et al. (2003) σ 32 Q E L A D R Y GRist, W., T. J. Jorgensen, et al. (2003) σ 32 Y G V S A E R VRist, W., T. J. Jorgensen, et al. (2003) σ 32 Q L E K N A M KWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 M S I V RWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 F A D L W A D PWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 P F D T F R S IWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 F D T F R S I VWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 E T A A F A N AWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 N A R M D W K EWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 A R M D W K E TWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 A H V F K A D LWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 G N V L V V S GWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 N D K W H R V EWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 S G K F V R R FWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 L L E D A K V EWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 K V E E V K A GWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 N G V L T V T VWintrode, P. L., K. L. Friedrich, et al. (2003) HSP16.9 E V K A I Q I SHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA M D V T I QHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA P S R L F D Q FHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA D Q F F G E G LHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA E G L F E Y D LHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA Y D L L P F L S
69

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Hasan, A., D. L. Smith, et al. (2002) α-Crystallin αA R Q S L P R T VHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA R T V L D S G IHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA S G I S G V R SHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA R D K F V I F LHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA F V I F L D V KHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA I F L D V K H FHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA P E D L T V K VHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA Q D D F V E I HHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA E R Q D D H G YHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA D D H G Y I S RHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA V D Q S A L S CHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA Q S A L S C S LHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA S C S L S A D GHasan, A., D. L. Smith, et al. (2002) α-Crystallin αA D G M L T F C GHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ M D I A I HHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ P S R L F D Q FHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ R L F D Q F F GHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ F D Q F F G E HHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ E S D L F P T SHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ P P S F L R A PHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ S W F D T G L SHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ G L S E M R L EHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ K D R F S V N LHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ S V N L D V K HHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ P E E L K V K VHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ D V I E V H G KHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ I T S S L S S DHasan, A., D. L. Smith, et al. (2002) α-Crystallin αΒ D G V L T V N GYan, X., H. Zhang, et al. (2002) rhM-CSFß S G H L Q S L QYan, X., H. Zhang, et al. (2002) rhM-CSFß L Q S L Q R L IYan, X., H. Zhang, et al. (2002) rhM-CSFß D S Q M E T S CYan, X., H. Zhang, et al. (2002) rhM-CSFß I T F E F V D QYan, X., H. Zhang, et al. (2002) rhM-CSFß K A F L L V Q DYan, X., H. Zhang, et al. (2002) rhM-CSFß D I M E D T M RYan, X., H. Zhang, et al. (2002) rhM-CSFß T M R F R D N TYan, X., H. Zhang, et al. (2002) rhM-CSFß T P N A I A I VYan, X., H. Zhang, et al. (2002) rhM-CSFß N A I A I V Q KYan, X., H. Zhang, et al. (2002) rhM-CSFß I V Q L Q E L SYan, X., H. Zhang, et al. (2002) rhM-CSFß Q L Q E L S L RYan, X., H. Zhang, et al. (2002) rhM-CSFß E L S L R L K SYan, X., H. Zhang, et al. (2002) rhM-CSFß C V R T F Y E TYan, X., H. Zhang, et al. (2002) rhM-CSFß V R T F Y E T PYan, X., H. Zhang, et al. (2002) rhM-CSFß P L Q L L E K VYan, X., H. Zhang, et al. (2002) rhM-CSFß K N L L D K D WYan, X., H. Zhang, et al. (2002) rhM-CSFß W N I F S K N CYan, X., H. Zhang, et al. (2002) rhM-CSFß C S S Q D V V TYan, X., H. Zhang, et al. (2002) rhM-CSFß C N C L Y P K AYan, X., H. Zhang, et al. (2002) rhM-CSFß V A Q L T W E DYan, X., H. Zhang, et al. (2002) rhM-CSFß L T W E D S E GYan, X., H. Zhang, et al. (2002) rhM-CSFß S S L L P G E QYan, X., H. Zhang, et al. (2002) rhM-CSFß H T V D P G S A
70

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Hughes, C. A., J. G. Mandell, et al. (2001) CheB M S K I RHughes, C. A., J. G. Mandell, et al. (2001) CheB D S A L M R Q IHughes, C. A., J. G. Mandell, et al. (2001) CheB I M T E I I N SHughes, C. A., J. G. Mandell, et al. (2001) CheB D M E M V A T AHughes, C. A., J. G. Mandell, et al. (2001) CheB P L V A R D L IHughes, C. A., J. G. Mandell, et al. (2001) CheB V A R D L I K KHughes, C. A., J. G. Mandell, et al. (2001) CheB V L T L D V E MHughes, C. A., J. G. Mandell, et al. (2001) CheB L T L D V E M PHughes, C. A., J. G. Mandell, et al. (2001) CheB D G L D F L E KHughes, C. A., J. G. Mandell, et al. (2001) CheB G L D F L E K LHughes, C. A., J. G. Mandell, et al. (2001) CheB L D F L E K L MHughes, C. A., J. G. Mandell, et al. (2001) CheB L E K L M R L RHughes, C. A., J. G. Mandell, et al. (2001) CheB P V V M V S S LHughes, C. A., J. G. Mandell, et al. (2001) CheB A L E L G A I DHughes, C. A., J. G. Mandell, et al. (2001) CheB A I D F V T K PHughes, C. A., J. G. Mandell, et al. (2001) CheB E G M L A Y S EHughes, C. A., J. G. Mandell, et al. (2001) CheB G M L A Y S E MHughes, C. A., J. G. Mandell, et al. (2001) CheB Y S E M I A E KHughes, C. A., J. G. Mandell, et al. (2001) CheB P T T L K A G PHughes, C. A., J. G. Mandell, et al. (2001) CheB S E K L I A I GHughes, C. A., J. G. Mandell, et al. (2001) CheB G G T E A I R HHughes, C. A., J. G. Mandell, et al. (2001) CheB G T E A I R H VHughes, C. A., J. G. Mandell, et al. (2001) CheB P L S S P A V IHughes, C. A., J. G. Mandell, et al. (2001) CheB P A V I I T Q HHughes, C. A., J. G. Mandell, et al. (2001) CheB P P G F T R S FHughes, C. A., J. G. Mandell, et al. (2001) CheB A E R L N K L CHughes, C. A., J. G. Mandell, et al. (2001) CheB V L P G H A Y IHughes, C. A., J. G. Mandell, et al. (2001) CheB P G H A Y I A PHughes, C. A., J. G. Mandell, et al. (2001) CheB G H A Y I A P GHughes, C. A., J. G. Mandell, et al. (2001) CheB H M E L A R S GHughes, C. A., J. G. Mandell, et al. (2001) CheB S G A N Y Q I KHughes, C. A., J. G. Mandell, et al. (2001) CheB P S V D V L F HHughes, C. A., J. G. Mandell, et al. (2001) CheB V D V L F H S VHughes, C. A., J. G. Mandell, et al. (2001) CheB D V L F H S V AHughes, C. A., J. G. Mandell, et al. (2001) CheB G V I L T G M GHughes, C. A., J. G. Mandell, et al. (2001) CheB E A S C V V F GHughes, C. A., J. G. Mandell, et al. (2001) CheB M P R E A I N MHughes, C. A., J. G. Mandell, et al. (2001) CheB G V S E V V D LHughes, C. A., J. G. Mandell, et al. (2001) CheB V S E V V D L SHughes, C. A., J. G. Mandell, et al. (2001) CheB Q M L A K I S AWang, L., L. C. Lane, et al. (2001) BMV A I A G Y S I SWang, L., L. C. Lane, et al. (2001) BMV A S S D A I T AWang, L., L. C. Lane, et al. (2001) BMV S D A I T A K AWang, L., L. C. Lane, et al. (2001) BMV A T N A M S I TWang, L., L. C. Lane, et al. (2001) BMV N A M S I T L PWang, L., L. C. Lane, et al. (2001) BMV G R V L L W L GWang, L., L. C. Lane, et al. (2001) BMV R V L L W L G LWang, L., L. C. Lane, et al. (2001) BMV L L W L G L L PWang, L., L. C. Lane, et al. (2001) BMV L G L L P S V AWang, L., L. C. Lane, et al. (2001) BMV R I K A C V A EWang, L., L. C. Lane, et al. (2001) BMV K A C V A E K Q
71

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Wang, L., L. C. Lane, et al. (2001) BMV A C V A E K Q AWang, L., L. C. Lane, et al. (2001) BMV A E A A F Q V AWang, L., L. C. Lane, et al. (2001) BMV E A A F Q V A LWang, L., L. C. Lane, et al. (2001) BMV A F Q V A L A VWang, L., L. C. Lane, et al. (2001) BMV F Q V A L A V AWang, L., L. C. Lane, et al. (2001) BMV Q V A L A V A DWang, L., L. C. Lane, et al. (2001) BMV S S K E V V A AWang, L., L. C. Lane, et al. (2001) BMV V A A M Y T D AWang, L., L. C. Lane, et al. (2001) BMV Y T D A F R G AWang, L., L. C. Lane, et al. (2001) BMV G D L L N L Q IWang, L., L. C. Lane, et al. (2001) BMV L L N L Q I Y LWang, L., L. C. Lane, et al. (2001) BMV Q I Y L Y A S EWang, L., L. C. Lane, et al. (2001) BMV P A K A V V H LWang, L., L. C. Lane, et al. (2001) BMV V V H L E V E HWang, L., L. C. Lane, et al. (2001) BMV F D D F F T P VChen, J. and D. L. Smith (2000) GroEL R G V N V L A DChen, J. and D. L. Smith (2000) GroEL K G R N V V L DChen, J. and D. L. Smith (2000) GroEL D G V S V A R EChen, J. and D. L. Smith (2000) GroEL E I E L E D K FChen, J. and D. L. Smith (2000) GroEL G A Q M V K E VChen, J. and D. L. Smith (2000) GroEL A T V L A Q A IChen, J. and D. L. Smith (2000) GroEL V T A A V E E LChen, J. and D. L. Smith (2000) GroEL E L K A L S V PChen, J. and D. L. Smith (2000) GroEL D S K A I A Q VChen, J. and D. L. Smith (2000) GroEL V G K L I A E AChen, J. and D. L. Smith (2000) GroEL A E A M D K V GChen, J. and D. L. Smith (2000) GroEL Q D E L D V V EChen, J. and D. L. Smith (2000) GroEL D E L D V V E GChen, J. and D. L. Smith (2000) GroEL E G M Q F D R GChen, J. and D. L. Smith (2000) GroEL S P Y F I N K PChen, J. and D. L. Smith (2000) GroEL N K P E T G A VChen, J. and D. L. Smith (2000) GroEL G A V E L E S PChen, J. and D. L. Smith (2000) GroEL P F I L L A D KChen, J. and D. L. Smith (2000) GroEL F I L L A D K KChen, J. and D. L. Smith (2000) GroEL R E M L P V L EChen, J. and D. L. Smith (2000) GroEL L P V L E A V AChen, J. and D. L. Smith (2000) GroEL K P L L I I A EChen, J. and D. L. Smith (2000) GroEL I I A E D V E GChen, J. and D. L. Smith (2000) GroEL G E A L A T L VChen, J. and D. L. Smith (2000) GroEL L A T L V V N TChen, J. and D. L. Smith (2000) GroEL T M R G I V K VChen, J. and D. L. Smith (2000) GroEL M L Q D I A T LChen, J. and D. L. Smith (2000) GroEL I S E E I G M EChen, J. and D. L. Smith (2000) GroEL E E I G M E L EChen, J. and D. L. Smith (2000) GroEL K A T L E D L GChen, J. and D. L. Smith (2000) GroEL D T T T I I D GChen, J. and D. L. Smith (2000) GroEL Q I E E A T S DChen, J. and D. L. Smith (2000) GroEL K L Q E R V A KChen, J. and D. L. Smith (2000) GroEL G G V A V I K VChen, J. and D. L. Smith (2000) GroEL A A T E V E M KChen, J. and D. L. Smith (2000) GroEL E D A L H A T R
72
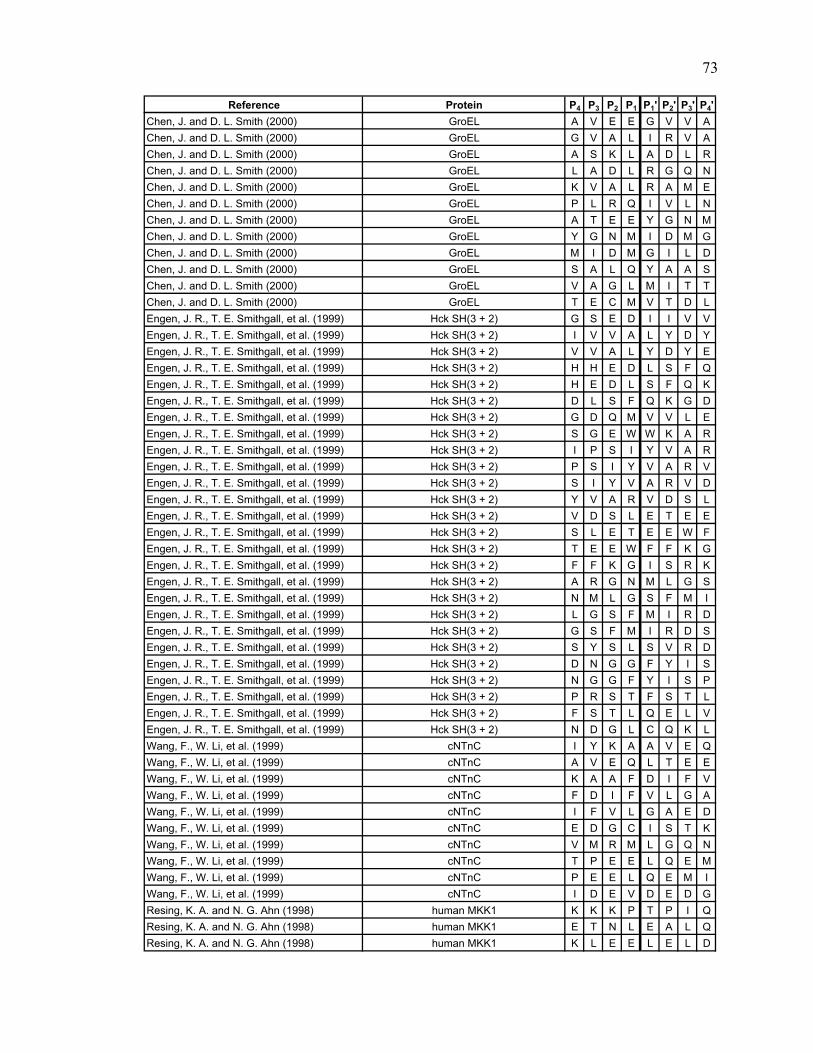
Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Chen, J. and D. L. Smith (2000) GroEL A V E E G V V AChen, J. and D. L. Smith (2000) GroEL G V A L I R V AChen, J. and D. L. Smith (2000) GroEL A S K L A D L RChen, J. and D. L. Smith (2000) GroEL L A D L R G Q NChen, J. and D. L. Smith (2000) GroEL K V A L R A M EChen, J. and D. L. Smith (2000) GroEL P L R Q I V L NChen, J. and D. L. Smith (2000) GroEL A T E E Y G N MChen, J. and D. L. Smith (2000) GroEL Y G N M I D M GChen, J. and D. L. Smith (2000) GroEL M I D M G I L DChen, J. and D. L. Smith (2000) GroEL S A L Q Y A A SChen, J. and D. L. Smith (2000) GroEL V A G L M I T TChen, J. and D. L. Smith (2000) GroEL T E C M V T D LEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) G S E D I I V VEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) I V V A L Y D YEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) V V A L Y D Y EEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) H H E D L S F QEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) H E D L S F Q KEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) D L S F Q K G DEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) G D Q M V V L EEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) S G E W W K A REngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) I P S I Y V A REngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) P S I Y V A R VEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) S I Y V A R V DEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) Y V A R V D S LEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) V D S L E T E EEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) S L E T E E W FEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) T E E W F F K GEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) F F K G I S R KEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) A R G N M L G SEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) N M L G S F M IEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) L G S F M I R DEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) G S F M I R D SEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) S Y S L S V R DEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) D N G G F Y I SEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) N G G F Y I S PEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) P R S T F S T LEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) F S T L Q E L VEngen, J. R., T. E. Smithgall, et al. (1999) Hck SH(3 + 2) N D G L C Q K LWang, F., W. Li, et al. (1999) cNTnC I Y K A A V E QWang, F., W. Li, et al. (1999) cNTnC A V E Q L T E EWang, F., W. Li, et al. (1999) cNTnC K A A F D I F VWang, F., W. Li, et al. (1999) cNTnC F D I F V L G AWang, F., W. Li, et al. (1999) cNTnC I F V L G A E DWang, F., W. Li, et al. (1999) cNTnC E D G C I S T KWang, F., W. Li, et al. (1999) cNTnC V M R M L G Q NWang, F., W. Li, et al. (1999) cNTnC T P E E L Q E MWang, F., W. Li, et al. (1999) cNTnC P E E L Q E M IWang, F., W. Li, et al. (1999) cNTnC I D E V D E D GResing, K. A. and N. G. Ahn (1998) human MKK1 K K K P T P I QResing, K. A. and N. G. Ahn (1998) human MKK1 E T N L E A L QResing, K. A. and N. G. Ahn (1998) human MKK1 K L E E L E L D
73

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Resing, K. A. and N. G. Ahn (1998) human MKK1 R L E A F L T QResing, K. A. and N. G. Ahn (1998) human MKK1 L E A F L T Q KResing, K. A. and N. G. Ahn (1998) human MKK1 K D D D F E K IResing, K. A. and N. G. Ahn (1998) human MKK1 D D D F E K I SResing, K. A. and N. G. Ahn (1998) human MKK1 K I S E L G A GResing, K. A. and N. G. Ahn (1998) human MKK1 G V V F K V S HResing, K. A. and N. G. Ahn (1998) human MKK1 G L V M A R K LResing, K. A. and N. G. Ahn (1998) human MKK1 L I H L E I K PResing, K. A. and N. G. Ahn (1998) human MKK1 I I R E L Q V LResing, K. A. and N. G. Ahn (1998) human MKK1 I V G F Y G A FResing, K. A. and N. G. Ahn (1998) human MKK1 Y G A F Y S D GResing, K. A. and N. G. Ahn (1998) human MKK1 G E I S I C M EResing, K. A. and N. G. Ahn (1998) human MKK1 I S I C M E H MResing, K. A. and N. G. Ahn (1998) human MKK1 G G S L D Q V LResing, K. A. and N. G. Ahn (1998) human MKK1 V S I A V I K GResing, K. A. and N. G. Ahn (1998) human MKK1 L T Y L R E K HResing, K. A. and N. G. Ahn (1998) human MKK1 K P S N I L V NResing, K. A. and N. G. Ahn (1998) human MKK1 E I K L C D V GResing, K. A. and N. G. Ahn (1998) human MKK1 S G Q L I D S MResing, K. A. and N. G. Ahn (1998) human MKK1 L I D S M A N SResing, K. A. and N. G. Ahn (1998) human MKK1 M A N S F V G TResing, K. A. and N. G. Ahn (1998) human MKK1 A N S F V G T RResing, K. A. and N. G. Ahn (1998) human MKK1 G T H Y S V Q SResing, K. A. and N. G. Ahn (1998) human MKK1 W S M G L S L VResing, K. A. and N. G. Ahn (1998) human MKK1 G L S L V E M AResing, K. A. and N. G. Ahn (1998) human MKK1 D A K E L E L MResing, K. A. and N. G. Ahn (1998) human MKK1 E L E L M F G CResing, K. A. and N. G. Ahn (1998) human MKK1 E L M F G C Q VResing, K. A. and N. G. Ahn (1998) human MKK1 F G C Q V E G DResing, K. A. and N. G. Ahn (1998) human MKK1 E G D A A E T PResing, K. A. and N. G. Ahn (1998) human MKK1 M A I F E L L DResing, K. A. and N. G. Ahn (1998) human MKK1 E L L D Y I V NResing, K. A. and N. G. Ahn (1998) human MKK1 S G V F S L E FResing, K. A. and N. G. Ahn (1998) human MKK1 F S L E F Q D FResing, K. A. and N. G. Ahn (1998) human MKK1 M V H A F I K RResing, K. A. and N. G. Ahn (1998) human MKK1 D A E E V D F AResing, K. A. and N. G. Ahn (1998) human MKK1 A G W L C S T IResing, K. A. and N. G. Ahn (1998) human MKK1 L C S T I G L NNeubert, T. A., K. A. Walsh, et al. (1997) recoverin L S K E I L E ENeubert, T. A., K. A. Walsh, et al. (1997) recoverin I L E E L Q L NNeubert, T. A., K. A. Walsh, et al. (1997) recoverin E L Q L N T K FNeubert, T. A., K. A. Walsh, et al. (1997) recoverin E E E L S S W YNeubert, T. A., K. A. Walsh, et al. (1997) recoverin L S S W Y Q S FNeubert, T. A., K. A. Walsh, et al. (1997) recoverin Y Q S F L K E CNeubert, T. A., K. A. Walsh, et al. (1997) recoverin Q S F L K E C PNeubert, T. A., K. A. Walsh, et al. (1997) recoverin Q T I Y S K F FNeubert, T. A., K. A. Walsh, et al. (1997) recoverin Y S K F F P E ANeubert, T. A., K. A. Walsh, et al. (1997) recoverin D P K A Y A Q HNeubert, T. A., K. A. Walsh, et al. (1997) recoverin Q H V F R S F DNeubert, T. A., K. A. Walsh, et al. (1997) recoverin D G T L D F K ENeubert, T. A., K. A. Walsh, et al. (1997) recoverin D F K E Y V I A
74
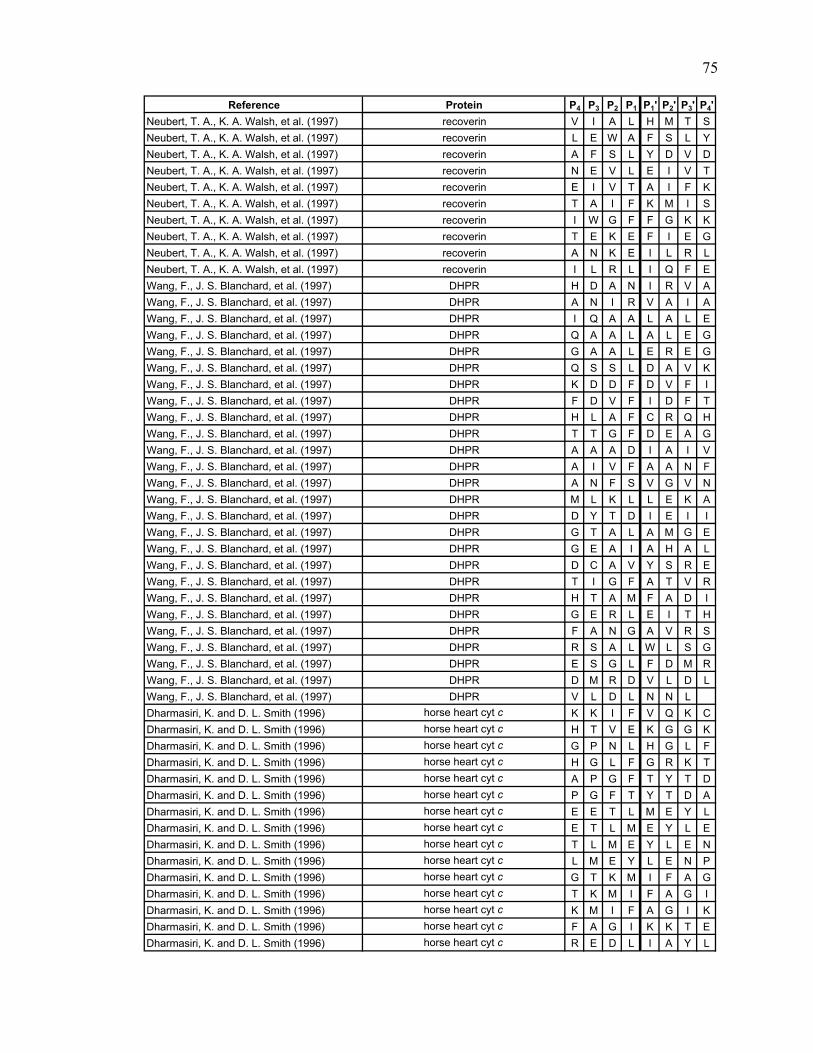
Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Neubert, T. A., K. A. Walsh, et al. (1997) recoverin V I A L H M T SNeubert, T. A., K. A. Walsh, et al. (1997) recoverin L E W A F S L YNeubert, T. A., K. A. Walsh, et al. (1997) recoverin A F S L Y D V DNeubert, T. A., K. A. Walsh, et al. (1997) recoverin N E V L E I V TNeubert, T. A., K. A. Walsh, et al. (1997) recoverin E I V T A I F KNeubert, T. A., K. A. Walsh, et al. (1997) recoverin T A I F K M I SNeubert, T. A., K. A. Walsh, et al. (1997) recoverin I W G F F G K KNeubert, T. A., K. A. Walsh, et al. (1997) recoverin T E K E F I E GNeubert, T. A., K. A. Walsh, et al. (1997) recoverin A N K E I L R LNeubert, T. A., K. A. Walsh, et al. (1997) recoverin I L R L I Q F EWang, F., J. S. Blanchard, et al. (1997) DHPR H D A N I R V AWang, F., J. S. Blanchard, et al. (1997) DHPR A N I R V A I AWang, F., J. S. Blanchard, et al. (1997) DHPR I Q A A L A L EWang, F., J. S. Blanchard, et al. (1997) DHPR Q A A L A L E GWang, F., J. S. Blanchard, et al. (1997) DHPR G A A L E R E GWang, F., J. S. Blanchard, et al. (1997) DHPR Q S S L D A V KWang, F., J. S. Blanchard, et al. (1997) DHPR K D D F D V F IWang, F., J. S. Blanchard, et al. (1997) DHPR F D V F I D F TWang, F., J. S. Blanchard, et al. (1997) DHPR H L A F C R Q HWang, F., J. S. Blanchard, et al. (1997) DHPR T T G F D E A GWang, F., J. S. Blanchard, et al. (1997) DHPR A A A D I A I VWang, F., J. S. Blanchard, et al. (1997) DHPR A I V F A A N FWang, F., J. S. Blanchard, et al. (1997) DHPR A N F S V G V NWang, F., J. S. Blanchard, et al. (1997) DHPR M L K L L E K AWang, F., J. S. Blanchard, et al. (1997) DHPR D Y T D I E I IWang, F., J. S. Blanchard, et al. (1997) DHPR G T A L A M G EWang, F., J. S. Blanchard, et al. (1997) DHPR G E A I A H A LWang, F., J. S. Blanchard, et al. (1997) DHPR D C A V Y S R EWang, F., J. S. Blanchard, et al. (1997) DHPR T I G F A T V RWang, F., J. S. Blanchard, et al. (1997) DHPR H T A M F A D IWang, F., J. S. Blanchard, et al. (1997) DHPR G E R L E I T HWang, F., J. S. Blanchard, et al. (1997) DHPR F A N G A V R SWang, F., J. S. Blanchard, et al. (1997) DHPR R S A L W L S GWang, F., J. S. Blanchard, et al. (1997) DHPR E S G L F D M RWang, F., J. S. Blanchard, et al. (1997) DHPR D M R D V L D LWang, F., J. S. Blanchard, et al. (1997) DHPR V L D L N N LDharmasiri, K. and D. L. Smith (1996) horse heart cyt c K K I F V Q K CDharmasiri, K. and D. L. Smith (1996) horse heart cyt c H T V E K G G KDharmasiri, K. and D. L. Smith (1996) horse heart cyt c G P N L H G L FDharmasiri, K. and D. L. Smith (1996) horse heart cyt c H G L F G R K TDharmasiri, K. and D. L. Smith (1996) horse heart cyt c A P G F T Y T DDharmasiri, K. and D. L. Smith (1996) horse heart cyt c P G F T Y T D ADharmasiri, K. and D. L. Smith (1996) horse heart cyt c E E T L M E Y LDharmasiri, K. and D. L. Smith (1996) horse heart cyt c E T L M E Y L EDharmasiri, K. and D. L. Smith (1996) horse heart cyt c T L M E Y L E NDharmasiri, K. and D. L. Smith (1996) horse heart cyt c L M E Y L E N PDharmasiri, K. and D. L. Smith (1996) horse heart cyt c G T K M I F A GDharmasiri, K. and D. L. Smith (1996) horse heart cyt c T K M I F A G IDharmasiri, K. and D. L. Smith (1996) horse heart cyt c K M I F A G I KDharmasiri, K. and D. L. Smith (1996) horse heart cyt c F A G I K K T EDharmasiri, K. and D. L. Smith (1996) horse heart cyt c R E D L I A Y L
75

Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Dharmasiri, K. and D. L. Smith (1996) horse heart cyt c E D L I A Y L KDharmasiri, K. and D. L. Smith (1996) horse heart cyt c D L I A Y L K KDharmasiri, K. and D. L. Smith (1996) horse heart cyt c L I A Y L K K AZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase H P A L T P E QZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase E L S D I A H RZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase K G I L A A D EZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase G S I A K R L QZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase I G T E N T E EZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase N T E E N R R FZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase N R R F Y R Q LZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase R G L L L T A DZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase Q L L L T A D DZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase L L L T A D D RZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase L T A D D R V NZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase T A D D R V N PZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase P C I G G V I LZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase G V I L F H E TZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase H E T L Y Q K AZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase G V V G I K V DZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase L D G L S E R CZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase D G L S E R C AZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase D G A D F A K WZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase R C V L K I G EZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase T P S A L A I MZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase P S A L A I M EZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase S A L A I M E NZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase A I M E N A N VZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase A N V L A R Y AZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase L A R Y A S I CZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase I C Q Q N G I VZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase P I V E P E I LZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase R C Q Y V T E KZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase Y V T E K V L AZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase E K V L A A V YZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase V L A A V Y K AZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase L A A V Y K A LZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase A A V Y K A L SZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase H I Y L E G T LZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase Y L E G T L L KZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase Y S H E E I A MZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase E I A M A T V TZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase M A T V T A L RZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase T V T A L R R TZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase V T A L R R T VZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase P A V T G V T FZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase S E E E A S I NZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase E E E A S I N LZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase S I N L N A I NZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase N L N A I N K CZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase W A L T F S Y GZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase A L Q A S A L K
76
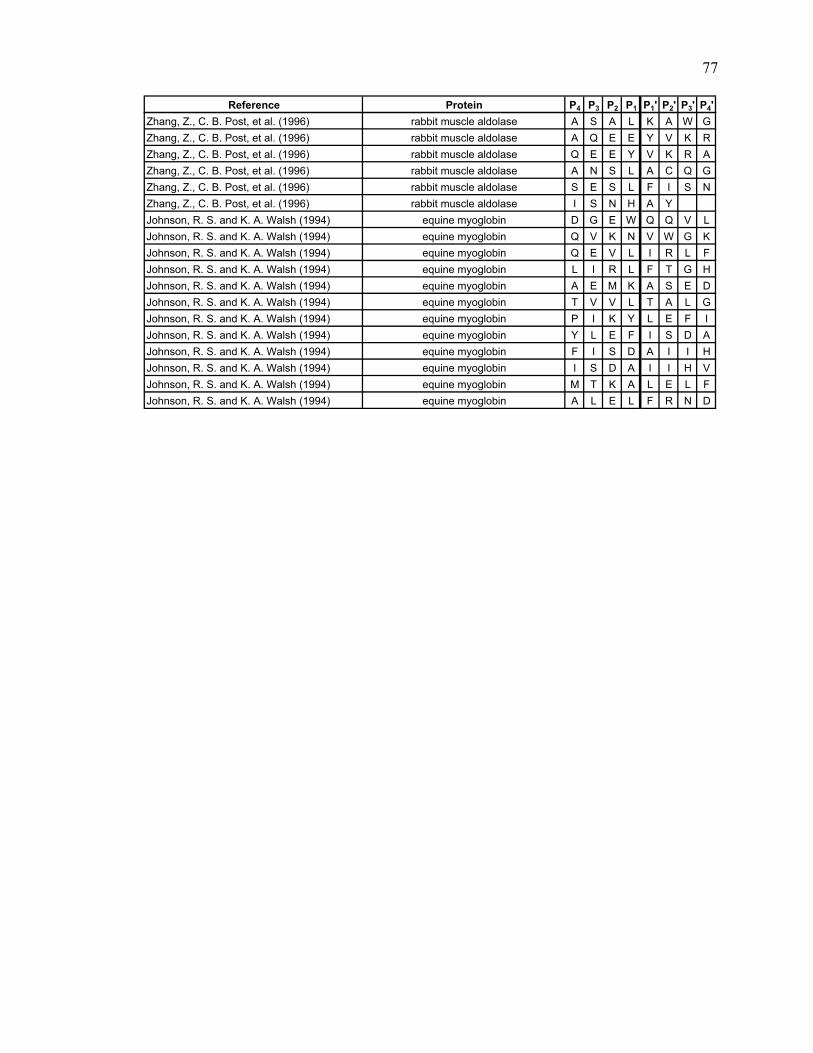
Reference Protein P4 P3 P2 P1 P1' P2' P3' P4'Zhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase A S A L K A W GZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase A Q E E Y V K RZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase Q E E Y V K R AZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase A N S L A C Q GZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase S E S L F I S NZhang, Z., C. B. Post, et al. (1996) rabbit muscle aldolase I S N H A YJohnson, R. S. and K. A. Walsh (1994) equine myoglobin D G E W Q Q V LJohnson, R. S. and K. A. Walsh (1994) equine myoglobin Q V K N V W G KJohnson, R. S. and K. A. Walsh (1994) equine myoglobin Q E V L I R L FJohnson, R. S. and K. A. Walsh (1994) equine myoglobin L I R L F T G HJohnson, R. S. and K. A. Walsh (1994) equine myoglobin A E M K A S E DJohnson, R. S. and K. A. Walsh (1994) equine myoglobin T V V L T A L GJohnson, R. S. and K. A. Walsh (1994) equine myoglobin P I K Y L E F IJohnson, R. S. and K. A. Walsh (1994) equine myoglobin Y L E F I S D AJohnson, R. S. and K. A. Walsh (1994) equine myoglobin F I S D A I I HJohnson, R. S. and K. A. Walsh (1994) equine myoglobin I S D A I I H VJohnson, R. S. and K. A. Walsh (1994) equine myoglobin M T K A L E L FJohnson, R. S. and K. A. Walsh (1994) equine myoglobin A L E L F R N D
77

CHAPTER 3
EXPERIMENTAL DETERMINATION OF
PEPSIN SPECIFICITY AND THE EFFECTS OF pH
3.1 Introduction
The second part of this project involved the experimental research, the goal of
which was to increase the data set of pepsin cleavages. The literature search provided a
good starting point, but as stated previously, the amount of data produced was not enough
to accurately determine the trends in pepsin specificity.
Another goal of the experimental research was to determine if the specificity of
pepsin is affected by a change in pH. It is known that pepsin activity is directly affected
by pH (Jones and Landon 2002). Pepsin experiences its highest activity around pH 1.9.
It remains very active down to pH 1 and begins to lose activity around pH 5 (Brier, Maria
et al. 2007).
To address the two main goals of the experimental project various proteins were
digested with pepsin and analyzed using LC/MS/MS. The proteins were digested at three
different pH conditions, pH 1.0, pH 2.5 and pH 4.0. In addition to digesting a selection
of proteins, the bacterium Escherichia coli (E.coli) whole cell lysate was also digested at
the aforementioned pH points and analyzed using LC/MS/MS.
3.2 Instrumentation
Online digestion and sample analysis was performed using UPLC/ESI-MSE. This
method was chosen because it allows for fast and accurate determination of peptides
(Chakraborty, Berger et al. 2007).
78

3.2.1 UPLC and online pepsin digestion
UPLC was chosen for analysis over HPLC because it allows for a faster analysis
time while still obtaining a very high-resolution separation (Swartz 2005). UPLC uses a
smaller particle size (less than 2 μm diameter) which creates very high operating
pressures, up to 15,000 psi. Another plus to using UPLC over HPLC is that UPLC has a
greater sensitivity therefore a smaller sample size can be used (Plumb, Castro-Perez et al.
2004; Swartz 2005).
Online pepsin is digestion preferred over a solution digest for various reasons.
One reason is that online digests allow for greater pepsin efficiency as pepsin becomes
more efficient as the enzyme to protein ratio increases. Compared to a solution digest,
with online digests much more pepsin can be packed into a column, therefore the
concentration of pepsin will be greater, thus increasing efficiency (Wu, Kaveti et al.
2006). The time needed for digestion can also be decreased when performing online
digestions because the efficiency of pepsin is increased. A solution digest that would
take five minutes can be decreased to 20 seconds in a pepsin column (Jones and Landon
2002). One final bonus to performing online digestion is the ability to automate the
process. This was especially important with this project because of the number of
samples that needed to be analyzed.
A schematic of the UPLC setup for online digestion can be found in Figure 3.1.
The sample is injected into valve A. It travels through the loop and is then pushed
through the pepsin column via the auxiliary solvent manager (ASM) which contains a pH
buffer in water. Following digestion the peptides are desalted and concentrated on the
peptide trap. Valve B then switches the trap inline with the binary solvent manager
79

Binary Solvent Manger
Auxiliary Solvent Manager: Pump B
A B
MSC18 Column
A ili S l t M
Pepsin Column
Peptide Trap
Auxiliary Solvent Manager:Pump A
Figure 3.1. Schematic of online pepsin digestion adapted from Wang (2002). The sample is injected into the loop of valve A and is then pushed through the pepsin column with Pump A of the auxiliary solvent manager (ASM) containing H b ff i H O Th ti tid th d lt d d t t d thpH buffer in H2O. The peptic peptides are then desalted and concentrated on the
peptide trap. Valve B then switches to turn the trap inline with the binary solvent manager (BSM). The BSM creates the correct mixture of A and B solvents, where A is 0.05% FA in H2O and B is 0.05% FA in ACN, dependent on the gradient. The sample is then separated on a C18 column with the eluant flowing to the mass spectrometer. Pump B of the ASM delivers the reference material to the mass spectrometer.
80

(BSM). The BSM will then make up the needed mixture of A and B solvents, where A is
0.05% formic acid (FA) in water and B is 0.05%FA in acetonitrile (ACN), according to
the desired gradient. The peptic peptides are then separated on a C18 analytical column.
Following separation the eluant flows into the mass spectrometer for analysis (Wang, Pan
et al. 2002).
3.2.2 Mass spectrometry
For these experiments the technique of MSE was used for mass spectral analyses.
MSE is a data independent method of performing MS and MS/MS analyses. With MSE
both fragment and precursor ion information can be obtained relatively quickly and with
accurate mass in one analysis (Plumb, Johnson et al. 2006).
A diagram of the operation of MSE is displayed in Figure 3.2 (Plumb, Johnson et
al. 2006). The peptides that are separated during the UPLC analysis are introduced into
the mass spectrometer using an electrospray ionization (ESI) source and then enter the
first quadruple where the peptides are separated. The peptic peptides then move into the
collision cell where they are bombarded with either low or elevated energy. After this
step they move into the second quadruple which is operated in V mode (Chakraborty,
Berger et al. 2007).
The collision energy in the gas cell continuously alternates between low and
elevated energies at a specified rate. At the low collision energy a classic MS spectrum is
obtained generating information about the precursor ions. When hit with the elevated
collision energy, information about the fragment ions is obtained. When the elevated
energy is used, the high voltage causes the formation of both the precursor ions and its
81

MS
6
1 2 3 4
5 7
MSE
Figure 3.2. Schematic of the operation of MSE adapted from Plumb (2006). Peptides are separated during the UPLC separation, in this example there are two peptides that co-elute (1), and are introduced into the mass spectrometer via an ESI source (2). From there the peptides are separated in the first quadruple (3) then move into the collision cell (4) then into the second quadruple (5) which is operated in V mode. The collision energy in the gas cell alternates between low and high energy. At low energy precursor ion information is obtained yielding a classic mass spectrum (6). At the elevated energy information is collected for all of the fragment ions (7).
82

fragments. Therefore, throughout one analysis, information is constantly being gathered
about both the precursor and fragment ions. During data processing, the precursor ions
are matched up with their fragment ions based on their common retention times.
(Chakraborty, Berger et al. 2007).
3.3 Materials and Methods
3.3.1 Protein sample analysis
The proteins used for these experiments are commercially available from Sigma-
Aldrich or were obtained in-house. A list of the proteins used for digestion can be found
in Table 3.1. The proteins encompass a wide range of sizes, from 8.6 kDa (Ubiquitin) to
68.0 kDa (Amyloglucosidase).
3.3.1.1 Protein sample preparation
All of the proteins were dissolved into 50 mM TRIS buffer, pH 8.315, to a
concentration of 10 μM. These stock solutions were kept in a -20 °C freezer. 100 μL of
each protein stock solution were diluted into 100 μL of TRIS buffer to a concentration of
5 μM. Abelson tyrosine kinase (Abl), Nef (a small HIV accessory protein) and Ubiquitin
were heated at 60 °C for ten minutes to help reduce the disulfide bonds present. A small
amount of 1 M guanidine-HCl was also added to all samples to aid in digestion. For pH
1.0 analysis the samples were adjusted to pH 1.0 by adding 0.1 M HCl. The samples
were also analyzed at pH 2.5 and 4.0. Fresh aliquots of sample were adjusted to these pH
points by adding a buffer consisting of 50 mM potassium phosphate monobasic and 50
83

Table 3.1. Proteins used for digestion
Protein SwissProt ID and Ascension Number
Molecular Weight (kDa)
Number of Residues
Abelson tyrosine kinase (Abl)*
57.4 501
Albumin Albu_Bovin (P02769) 66.0 583 Aldolase Aldoa_Rabit (P00883) 39.5 364
Amyloglucosidase Amyg_Aspng (P69328) 68.0 640 β-Lactoglobulin Lacb_Bovin (P02754) 18.4 162
Hemoglobin
Hba_Bovin (P01966) Hbb_Bovin (P02070) Hbe2_Bovin (P06642) Hbe4_Bovin (P06643)
64.5 576
Myoglobin Myg_Horse (P68082) 17.0 154 Nef* 25.5 226
Ubiquitin Ubiq_Bovin (P62990) 8.6 76 * Proteins obtained in-house, all others available from Sigma-Aldrich
84

mM potassium phosphate dibasic for pH 1.0 and 2.5 analyses, and 50 mM citric acid for
pH 4.0 analysis.
3.3.1.2 UPLC methods
Online pepsin digestion and UPLC separation was performed on a Waters
nanoAcquitiy UPLC system. All samples were analyzed in triplicate. Pepsin digestion
was carried out using a 2.1 mm x 50 mm stainless steel column (Alltech) packed with
pepsin immobilized onto POROS-20R2 support (Applied Biosystems) as described
(Wang, Pan et al. 2002). Online digestion was essentially performed as described (Wang,
Pan et al. 2002). 10 μL of protein (50 pmol) was loaded onto a 20 μL loop and pushed
through the pepsin column by pump A of the ASM (see table 3.2 for solvents used) at a
flow rate of 100 μL/min. Digestion, trapping and desalting was a total of 3 minutes. A
vanGuard BEH C18 trap (Waters) containing 1.7 μm particles was used. The switch
valve then moved to align the peptide trap with the BSM. Chromatography was then
carried out using a nanoAcquity Symmetry C18, 180 μm x 20 mm column (Waters)
containing 5 μm particles. The elution gradient was 8% to 40% acetonitrile (ACN) in 6
minutes followed by a ramp to 85% ACN over 30 seconds with a flow rate of 40 μL/min.
0.05% formic acid (FA) was added to both mobile phases. The sample manager was held
at a temperature of 4 °C, the pepsin column was at room temperature and the analytical
column was held at 35 °C.
85
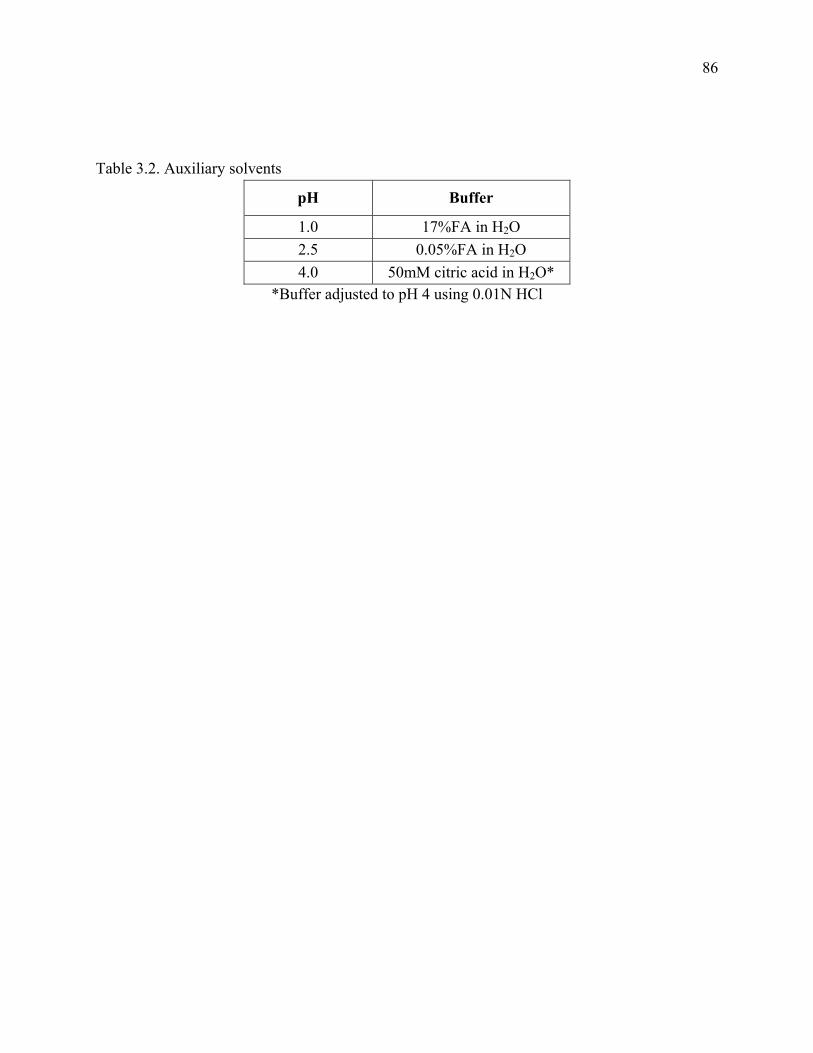
Table 3.2. Auxiliary solvents
pH Buffer
1.0 17%FA in H2O 2.5 0.05%FA in H2O 4.0 50mM citric acid in H2O*
*Buffer adjusted to pH 4 using 0.01N HCl
86

3.3.1.3 Mass analysis
All mass spectral analyses were carried out using a Waters Synapt HDMS system
equipped with a standard ESI source. All measurements were taken in positive ion V
mode with a capillary voltage of 3.2 kV, cone voltage of 37 V, source temperature of 100
°C, desolvation temperature of 250 °C and desolvation gas flow of 600 L/hour. The low
collision energy was 5 eV and the elevated collision energy was 25 eV. Each 0.4 second
scan spanned an m/z of 50-1700 with an interscan delay time of 0.02 seconds. To ensure
accurate mass all analyses were carried out using Waters LockSpray technology. (Glu1)-
fibrinopeptide B (GFP) was used as a reference. The 250 fmol/μL solution of GFP was
delivered to the mass spectrometer at a rate of 2.5 μL/min using pump B of the ASM.
Sampling of the reference solution was performed every 20 seconds.
3.3.2 E.coli sample analysis
E.coli whole cell lysate was obtained from Waters Corporation. The sample was
prepared from E.coli strain DH5alpha (Q-BIOgene) as described by Millea et al. (Millea,
Krull et al. 2006). The sample was measured to have a final protein concentration of 95
mg/mL using a modified Coomassie dyebinding assay (BioRad).
3.3.2.1 E.coli sample preparation
10.4 μL of E.coli was added to 187.5 μL of H2O and 6.2 μL of DTT. The
solution was then heated at 60 °C for 10 minutes. Following heating, the sample was
cooled back to room temperature and briefly centrifuged. 6.2 μL of iodoacetic acid was
then added and the sample was vortexed then allowed to sit in the dark at room
87

temperature for 30 minutes. After being stored in darkness, 40 μL of H2O was added to
the sample followed by 1 M guanidine HCl. The sample was then adjusted to the
appropriate pH as described in section 3.3.1.1. Finally, the sample was vortexed for
about one minute.
3.3.2.2 UPLC analysis
Online digestion and UPLC separation were performed essentially as described in
section 3.3.1.2. Changes made to the methods include a total digestion, trapping and
desalting time of 18 minutes with a flow rate of 15 μL/min. The elution gradient was
also changed to 8% to 40% ACN in 90 minutes followed by a ramp to 85% ACN over 6
minutes with a flow rate of 40 μL/min. All other parameters remained unchanged.
3.3.2.3 Mass analysis
Mass spectral analyses were carried out as described in section 3.3.1.3 with the
only change being that the elevated collision energy was set at 15 eV.
3.4 Data Analysis
3.4.1 Software processing
Acquired data was processed using Waters ProteinLynx Global Server software
(PLGS version 2.3). The low energy data and elevated energy data were lockmass
corrected and the fragment ions were aligned with their corresponding precursor ion
based on their retention times. In the case of the proteins, processed data was searched
against the known protein sequence. The processed E.coli data were searched against a
88

database of over 8700 proteins found in E.coli. The search was set for non-specific
enzymes in order to identify the maximum number of peptides, allowing for one missed
cleavage and no modifications in the search parameters.
3.4.2 Peptide analysis
All peptides generated from the software search were further refined to ensure
accuracy. MS/MS data for resulting peptides was investigated. Example MS/MS data
can be found in Figures 3.3 and 3.4. These examples show two different peptides
identified from hemoglobin and myoglobin respectively. In the spectra the red peaks
represent y-ion fragments while the blue peaks represent b-ion fragments. The green
peaks are those representing neutral loss of water, ammonia or immonium. All remaining
peaks are grey and were not assigned to one of the above groups. These ions along with
internal fragments are not labeled.
For a peptide to be considered reliable it must have been found in at least two out
of the three replicates. Also, the standard deviation retention time must have been less
than ten seconds and a minimum of five fragment ions must have been generated. Once
the reliable peptides were established they were then mapped onto their corresponding
protein sequence. Peptic peptide maps of the nine proteins analyzed at pH 2.5 can be
found in Figures 3.5-3.13 at the end of this chapter. With the exception of β-lactalbumin
and amyloglucosidase the sequence coverage percent for the proteins was very high.
89
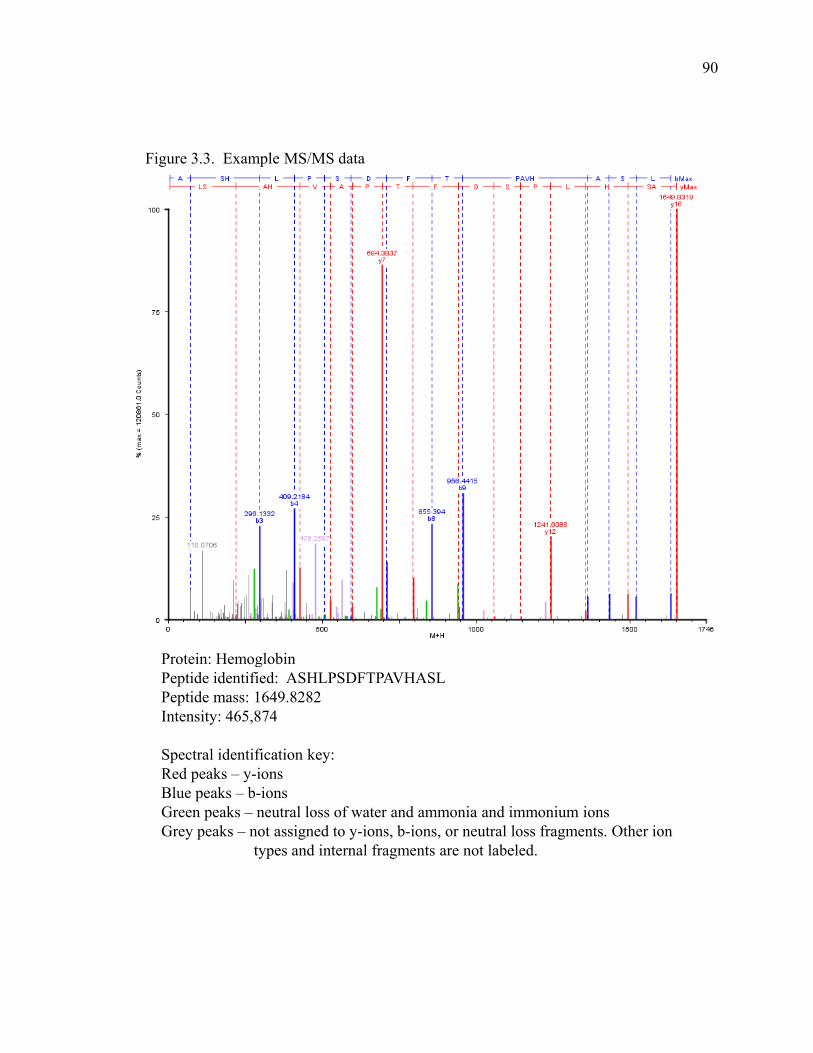
Figure 3.3. Example MS/MS data
Protein: HemoglobinProtein: HemoglobinPeptide identified: ASHLPSDFTPAVHASLPeptide mass: 1649.8282Intensity: 465,874
Spectral identification key:Red peaks – y-ionsBl k b iBlue peaks – b-ionsGreen peaks – neutral loss of water and ammonia and immonium ionsGrey peaks – not assigned to y-ions, b-ions, or neutral loss fragments. Other ion
types and internal fragments are not labeled.
90
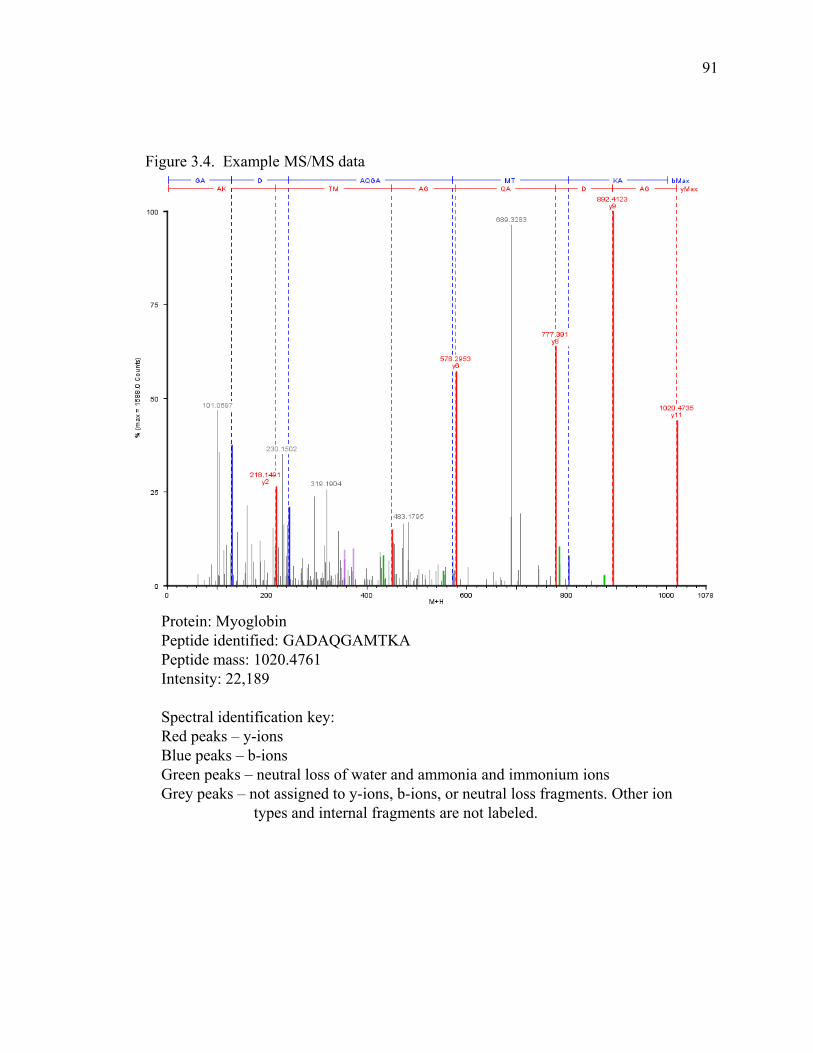
Figure 3.4. Example MS/MS data
Protein: MyoglobinPeptide identified: GADAQGAMTKAPeptide mass: 1020 4761Peptide mass: 1020.4761Intensity: 22,189
Spectral identification key:Red peaks – y-ionsBlue peaks – b-ionsGreen peaks – neutral loss of water and ammonia and immonium ionsG k t i d t i b i t l l f t Oth iGrey peaks – not assigned to y-ions, b-ions, or neutral loss fragments. Other ion
types and internal fragments are not labeled.
91

3.5 Results
Digestion of the nine proteins at pH 1.0 generated 582 peptides (see Table 3.3 at
end of chapter) at 695 cleavage points. pH 2.5 protein digestions produced 482 peptides
(shown in Figures 3.5-3.13 at end of chapter) with 591 different cleavage points and pH
4.0 digestions yielded 235 (see Table 3.4 at end of chapter) peptides and 366 cleavage
points. The E.coli digest at pH 2.5 produced 991 peptides (see Table 3.5 at end of
chapter) at 1368 different cleavage points. There were 49 proteins that were identified in
the E.coli lysate. The sum of data for all digests performed at pH 2.5 is 1473 peptides
and 1959 cleavage points.
While the 49 proteins identified by digesting E.coli lysate seems like a lot, this
number was much lower than expected as E.coli contains thousands of proteins. One
thing that could help to increase the number of proteins and peptides generated is to
further optimize the digestion and chromatography conditions. By increasing the
digestion time and running the analysis with a longer gradient there is a possibility that
more proteins and peptides will be generated.
After the peptic peptide maps were constructed as described in section 3.4 the
data was treated as described in sections 2.3-2.5. A database of cleavages was produced,
followed by data normalization and then the creation of a peptide cleavage map for each
pH. The cleavage data maps along with the corresponding matrix of normalized data
representing the probability of a cleavage occurring between two specific amino acids
can be found in Figures 3.14-3.16 and Tables 3.6-3.8 for pH 1.0, 2.5 and 4.0 respectively.
The peptide cleavage maps show that at all pH points pepsin will often cleave
after most bulky hydrophobic residues such as leucine and phenylalanine. The data also
92

Figure 3.14. Cleavage data map pH 1.0
P1
A C D E F G H I K L M N P Q R S T V W YA 1.6 0.9 0.7 0.3 0.9 0.8 0.6 1.7 0.7 1.1 2.7 1.0 0.0 0.8 1.1 1.3 0.7 1.5 1.5 1.5C 0.0 0.0 0.5 2.3 0.0 0.0 0.0 0.0 0.0 1.5 0.0 0.0 1.1 1.8 0.0 0.0 1.0 0.0 0.0 1.5D 0.8 0.0 0.3 1.0 1.7 0.3 0.0 2.0 0.5 0.8 0.0 0.0 0.4 0.0 0.0 0.3 0.5 0.0 0.0 2.3E 1.4 0.6 0.0 1.4 2.0 0.5 2.3 0.5 0.5 1.7 4.5 1.1 0.9 0.6 0.5 1.3 1.2 0.8 2.3 2.1
Table 3.6. Normalized pH 1.0 cleavage dataP1’
F 2.7 0.0 1.2 0.0 0.9 1.4 0.0 2.7 2.0 1.8 0.0 1.5 1.3 1.7 0.8 1.9 1.9 2.6 3.0 2.3G 0.6 0.0 0.0 0.2 1.8 0.3 2.6 0.9 0.8 0.8 0.0 0.9 0.9 0.6 1.0 1.0 0.7 0.7 3.0 1.1H 0.4 1.1 0.0 2.3 0.0 1.4 0.6 1.5 0.0 1.7 0.0 0.0 0.6 0.0 0.0 1.3 0.0 0.6 0.0 0.0I 0.7 0.0 0.0 0.0 2.3 0.6 0.0 1.5 1.0 1.1 4.5 1.1 0.0 1.1 1.5 0.8 1.3 0.7 0.0 0.0K 0.2 1.5 1.0 0.5 0.4 1.5 1.5 0.6 0.6 1.0 0.0 0.0 0.5 0.6 0.9 0.6 0.3 0.9 0.9 0.8L 1.6 1.8 1.2 2.4 3.2 0.7 1.5 2.9 1.7 2.3 0.0 1.9 1.5 1.3 1.7 2.0 1.4 1.7 0.9 2.7M 1.1 0.0 2.3 1.9 4.5 2.3 0.0 2.3 1.8 0.0 0.0 0.0 2.3 2.3 3.0 0.0 1.1 4.5 0.0 0.0
P1
N 1.7 1.5 0.0 0.0 2.8 0.3 0.0 0.0 1.5 0.5 0.0 0.0 1.0 0.0 0.8 0.0 1.1 1.7 0.0 0.9P 1.8 0.0 0.5 0.8 1.5 1.4 1.5 0.0 0.4 0.9 2.3 0.0 0.0 0.6 2.3 0.4 0.0 2.3 0.0 2.3Q 1.4 1.5 0.0 1.7 0.0 0.5 0.0 0.0 0.4 0.5 4.5 0.0 0.0 0.9 0.0 0.0 0.9 0.8 0.0 1.5R 1.8 0.9 1.8 0.5 0.0 0.0 4.5 1.5 2.3 0.3 0.0 1.0 1.5 0.0 0.5 0.0 0.0 0.9 0.0 0.0S 0.3 0.0 0.7 0.7 0.8 1.0 1.5 0.6 0.0 0.9 0.0 0.6 0.6 0.0 0.6 0.7 0.5 0.0 0.8 0.0T 0.7 0.0 0.6 0.4 0.5 0.7 1.1 0.0 0.4 0.6 0.9 4.5 1.3 0.5 0.0 0.5 0.4 1.3 0.0 0.8V 0.9 0.0 1.2 0.8 0.0 1.9 0.0 2.3 0.6 1.6 1.5 2.3 0.8 0.0 1.5 1.0 1.2 0.5 3.4 1.8W 0 0 1 5 0 0 0 0 0 0 0 6 0 0 0 0 0 0 3 0 0 0 0 0 1 5 1 1 0 0 0 0 0 8 2 3 0 0 1 5W 0.0 1.5 0.0 0.0 0.0 0.6 0.0 0.0 0.0 3.0 0.0 0.0 1.5 1.1 0.0 0.0 0.8 2.3 0.0 1.5Y 1.8 0.0 0.0 0.6 0.8 0.9 1.5 1.1 1.9 1.7 0.0 0.0 0.0 0.0 1.1 0.9 0.0 1.9 0.0 2.3
93

Figure 3.15. Cleavage data map pH 2.5
P1
P1’
A C D E F G H I K L M N P Q R S T V W YA 1.4 2.2 0.8 0.7 0.4 0.6 1.1 2.2 0.9 1.3 0.4 0.7 1.0 1.0 0.8 1.1 1.2 1.3 1.2 1.0C 0.0 0.0 0.0 0.0 1.3 0.0 0.0 1.9 0.0 0.0 1.6 0.0 0.6 0.0 1.3 0.0 1.2 0.8 0.0 1.9D 1.4 1.6 0.2 0.7 1.6 0.4 0.0 1.5 0.7 1.6 1.3 0.7 0.5 0.0 1.1 0.5 0.2 1.5 1.3 1.1E 1.5 0.0 0.7 1.4 2.4 0.7 1.3 2.1 1.3 1.1 1.8 1.0 0.8 0.6 1.5 0.7 0.7 2.1 1.0 2.3
Table 3.7. Normalized pH 2.5 cleavage data
P1
F 2.9 2.6 2.7 0.8 1.6 2.1 3.1 3.0 1.4 1.8 2.6 2.2 1.8 1.8 2.1 1.8 1.8 3.3 1.1 2.4G 0.5 0.0 0.1 1.0 1.5 0.5 0.9 1.0 0.9 0.4 0.6 0.5 0.5 0.8 0.5 0.8 0.5 0.8 3.0 0.5H 0.7 1.9 0.0 1.7 0.0 1.4 0.0 1.2 0.3 0.4 0.0 0.0 1.2 0.0 0.0 1.0 1.5 1.9 7.8 1.0I 1.0 2.2 0.6 0.4 1.6 1.1 0.6 1.0 0.6 0.9 1.3 0.4 0.2 0.4 0.3 0.9 0.8 0.7 0.0 0.6K 0.5 1.6 0.1 0.8 0.4 0.8 0.5 0.5 0.5 0.2 0.4 0.3 0.5 0.6 0.9 0.9 0.6 0.8 0.5 0.5L 1.7 0.6 2.9 1.8 3.5 1.2 2.0 2.4 2.0 2.1 1.6 2.2 0.8 1.9 1.9 1.5 2.1 2.2 1.8 2.6M 1.8 0.0 0.7 1.8 1.9 1.3 3.9 3.4 0.5 1.8 0.0 2.3 1.7 1.6 1.9 1.8 0.7 1.7 3.9 0.0N 0.8 0.0 0.2 0.8 1.8 0.3 0.0 1.3 0.7 1.0 0.5 0.8 0.8 0.0 1.6 0.6 0.0 1.9 1.6 1.8P 0.6 0.0 0.2 0.3 0.7 0.7 0.0 0.5 0.6 0.4 0.0 0.3 1.9 0.5 0.6 0.0 0.9 0.6 0.0 1.2Q 1.7 0.0 0.4 0.6 1.2 0.7 0.0 1.1 0.6 0.6 0.0 1.5 0.3 1.6 0.0 0.8 0.8 0.8 0.0 2.7R 0.6 0.9 0.0 0.9 0.3 0.5 0.8 0.5 1.2 0.3 0.5 0.8 1.0 0.7 0.4 0.8 0.2 0.4 0.0 0.7S 1.2 0.0 0.2 1.5 0.8 0.7 0.9 0.7 0.6 0.7 0.8 0.5 1.1 0.9 1.1 0.4 0.0 1.1 1.7 0.5T 1.1 1.0 0.3 1.2 2.2 0.5 0.8 1.6 0.8 1.0 0.5 0.6 1.1 0.5 1.1 1.0 0.5 1.3 0.6 0.3V 0.7 0.0 1.0 1.0 1.1 0.7 0.0 1.6 0.6 0.7 1.1 0.4 0.7 0.7 0.8 0.9 0.8 0.9 1.3 1.0W 0 6 2 6 1 2 0 0 0 0 0 0 1 6 2 6 0 0 0 5 2 6 0 0 0 0 5 5 0 0 3 9 1 4 1 9 3 9 5 2W 0.6 2.6 1.2 0.0 0.0 0.0 1.6 2.6 0.0 0.5 2.6 0.0 0.0 5.5 0.0 3.9 1.4 1.9 3.9 5.2Y 0.5 3.9 1.2 0.5 1.2 0.8 0.0 0.4 1.2 1.2 1.9 0.5 1.0 0.0 0.5 0.0 0.7 2.9 0.0 0.6
94

Figure 3.16. Cleavage data map pH 4.0
P1
P1’
A C D E F G H I K L M N P Q R S T V W YA 2.0 0.0 1.0 0.9 0.0 0.0 0.9 1.8 1.2 2.2 0.0 3.1 1.4 1.6 0.7 1.8 0.0 1.2 0.0 0.0C 1.0 0.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.8 1.4 0.0 0.0 1.2 0.0 0.0 0.0D 3.5 0.0 0.6 1.1 1.0 0.9 0.0 0.9 0.4 0.0 0.0 0.0 1.8 0.0 0.0 1.4 1.2 0.0 0.0 0.0E 1.3 0.0 0.0 1.9 0.8 0.0 1.8 0.9 0.0 0.7 3.5 1.2 0.9 3.5 1.6 0.0 2.0 1.0 2.3 1.6
Table 3.8. Normalized pH 4.0 cleavage data
P1
F 1.6 0.0 2.6 1.8 0.0 1.8 0.0 2.3 0.9 3.9 0.0 0.0 2.0 1.0 0.0 3.0 1.2 2.8 7.0 0.0G 2.9 0.0 0.0 0.5 0.0 0.5 1.0 3.5 0.0 1.2 2.3 2.3 0.0 0.0 1.2 3.0 0.0 0.5 2.3 0.0H 0.7 3.5 0.0 0.0 0.0 2.1 0.0 2.3 0.9 1.0 0.0 0.0 1.8 0.0 1.4 3.0 0.0 0.0 0.0 3.5I 0.6 2.3 0.0 0.0 0.0 0.0 0.0 2.3 0.8 0.0 3.5 0.0 0.0 3.5 0.0 1.0 2.3 1.0 0.0 0.0K 0.4 0.0 0.0 0.4 0.0 0.0 2.3 0.0 0.5 0.4 0.0 1.8 0.0 0.0 0.0 0.0 2.0 1.1 1.4 1.9L 1.8 0.0 2.3 1.9 4.7 0.8 1.5 2.8 0.8 1.7 0.0 2.8 3.5 4.7 2.1 2.1 1.4 1.6 0.0 2.3M 0.0 0.0 0.0 1.0 3.5 1.8 0.0 0.0 1.4 0.0 0.0 7.0 3.5 0.0 0.0 0.0 3.5 3.5 0.0 0.0N 1.3 2.3 0.0 0.0 0.0 0.0 0.0 0.0 2.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.9 0.0 1.8P 1.0 0.0 1.0 0.0 2.3 0.8 0.0 0.0 0.7 0.0 0.0 0.0 0.0 1.2 0.0 0.0 0.0 0.9 0.0 0.0Q 1.0 0.0 0.0 0.9 0.0 0.0 0.0 3.5 0.6 2.8 0.0 0.0 0.0 1.8 0.0 0.0 0.0 0.0 0.0 1.8R 1.2 1.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 2.8 0.0 0.8 0.0 0.0 1.4 0.0 1.2S 1.2 0.0 0.6 1.2 1.8 0.0 1.2 0.8 0.0 0.6 0.0 0.0 0.0 0.0 0.0 1.2 1.0 2.3 7.0 0.0T 0.5 0.0 0.0 0.6 1.2 0.0 0.0 0.0 0.6 1.6 0.0 0.0 2.3 0.0 0.0 1.8 0.0 0.8 0.0 2.0V 0.0 0.0 1.9 0.5 2.3 1.6 1.8 2.3 0.0 1.7 2.3 1.0 0.0 0.0 1.4 1.3 2.5 0.5 2.3 0.7W 0 0 3 5 0 0 0 0 0 0 1 4 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 3 5W 0.0 3.5 0.0 0.0 0.0 1.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 3.5Y 1.8 0.0 0.0 0.0 0.0 0.8 0.0 2.3 1.2 2.3 0.0 0.0 0.0 0.0 1.8 0.0 1.8 2.8 0.0 0.0
95

show that pepsin has a low probability of cleaving after the residues arginine, cysteine,
proline and histidine. The pH 2.5 data incorporates all of the E.coli data so there is a
large data set for this pH point. The sizes of the squares in the pH 2.5 cleavage data map
are also generally smaller than those found on the other two maps. This is because the
sequence coverage percent of the peptides identified in the E.coli lysate was on the lower
end. The lower the sequence coverage the lower the probability of cleavage appears to
be.
A revised version of the cleavage data maps were made as described in section
2.6 with the data representing the probability of cleavage between two specific residues
as a percentage. The data maps and corresponding percentage values can be found in
Figures 3.17-3.19 and Tables 3.9-3.11 for pH 1.0, 2.5 and 4.0 respectively. One of the
differences in this set of cleavage data maps is that the pH 2.5 map only takes into
consideration the data that was produced from the digestion of the nine standard proteins
and not the data produced from the digestion of E.coli whole cell lysate. The benefit to
only showing this selected data is that now the maps for all three pH points can be
directly compared because they contain digestion of the same proteins, therefore they
have the same number of possible cleavages. Overall, there were not many distinct
differences between the digestions at the different pH points. The trends in the
specificity of pepsin were similar at pH 1.0, 2.5 and 4.0.
One of the things that was affected by the change in pH was in the number of
peptides generated from each digestion. The low number of peptides produced by the
digestions at pH 4.0 is a reason why the pH 4.0 cleavage data map looks sparse compared
to those of pH 1.0 and 2.5. pH 1.0 produced the most amount of data, while pH 4.0
96

Figure 3.17. Cleavage data map with probability defined as a percentage, pH 1.0
YWVTSRQP
P1
NMLKIHGFFEDCA
A C D E F G H I K L M N P Q R S T V W YP1’Scale
Table 3.9. Normalized pH 1.0 cleavage data defined as a percentage
100 90 80 70 60 50 40 30 20 10
A C D E F G H I K L M N P Q R S T V W YA 31 20 15 6 11 18 13 33 8 25 60 14 0 18 25 24 15 27 33 33C 0 0 11 33 0 0 0 0 0 33 0 0 25 40 0 0 22 0 0 33
P1’
D 18 0 14 23 25 7 0 44 5 14 0 0 0 0 0 0 10 0 0 50E 33 13 0 26 56 11 40 11 12 39 100 24 20 14 11 26 27 11 50 45F 36 0 25 0 20 31 0 60 33 40 100 33 29 43 17 29 33 57 67 50G 20 0 0 5 40 7 57 20 12 19 0 8 20 13 0 24 14 25 67 25H 9 25 0 50 14 40 13 33 0 29 0 0 13 0 0 29 0 17 0 0I 15 0 0 20 50 14 0 33 22 25 33 25 0 25 33 18 29 13 0 0K 9 33 22 15 9 33 33 14 8 15 0 0 11 0 20 14 8 21 20 8L 33 40 31 52 67 11 40 54 40 47 100 38 33 17 38 44 30 37 17 62P1M 25 0 33 43 100 40 0 33 40 0 0 100 50 33 67 0 25 50 0 0N 46 33 0 0 63 8 0 0 33 9 0 0 22 0 17 0 25 9 0 20P 40 0 11 18 33 30 33 0 0 20 50 0 0 11 50 9 0 50 0 50Q 20 33 0 38 0 10 0 0 8 13 0 0 0 20 0 0 20 17 0 33R 40 20 40 10 0 0 100 33 67 6 25 33 33 0 11 0 0 20 0 0S 6 0 20 21 18 23 33 13 10 15 0 14 0 0 13 17 11 0 14 0T 20 0 14 8 18 17 20 0 9 13 20 100 28 10 0 12 0 29 0 17V 19 0 14 16 0 33 0 50 23 40 33 33 17 0 33 21 25 14 75 40
1
W 0 33 0 0 0 29 0 0 100 67 0 0 33 20 0 0 0 50 0 33Y 40 0 0 14 33 20 33 25 43 36 0 0 0 0 25 9 0 43 0 50
97
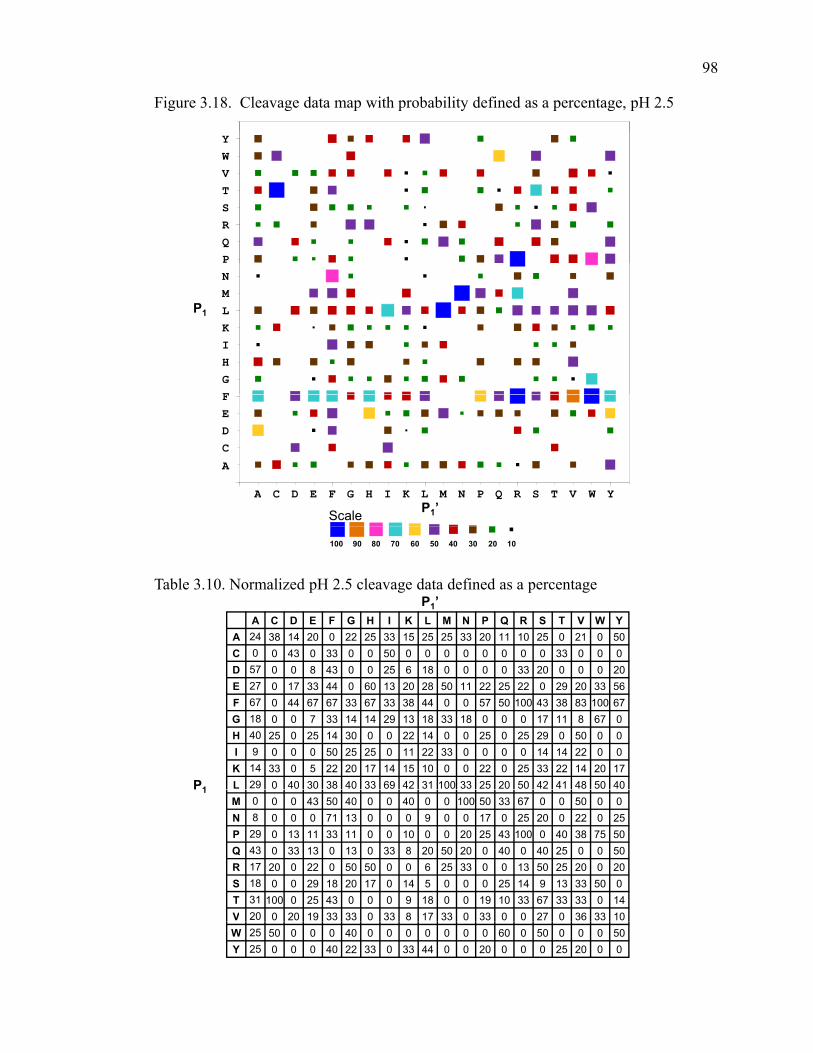
Figure 3.18. Cleavage data map with probability defined as a percentage, pH 2.5
YWVTSRQPNMLKIHGF
P1
A C D E F G H I K L M N P Q R S T V W Y
FEDC
A
P1’Scale
Table 3.10. Normalized pH 2.5 cleavage data defined as a percentage
100 90 80 70 60 50 40 30 20 10
P1’A C D E F G H I K L M N P Q R S T V W Y
A 24 38 14 20 0 22 25 33 15 25 25 33 20 11 10 25 0 21 0 50C 0 0 43 0 33 0 0 50 0 0 0 0 0 0 0 0 33 0 0 0
P1
D 57 0 0 8 43 0 0 25 6 18 0 0 0 0 33 20 0 0 0 20E 27 0 17 33 44 0 60 13 20 28 50 11 22 25 22 0 29 20 33 56F 67 0 44 67 67 33 67 33 38 44 0 0 57 50 100 43 38 83 100 67G 18 0 0 7 33 14 14 29 13 18 33 18 0 0 0 17 11 8 67 0H 40 25 0 25 14 30 0 0 22 14 0 0 25 0 25 29 0 50 0 0I 9 0 0 0 50 25 25 0 11 22 33 0 0 0 0 14 14 22 0 0K 14 33 0 5 22 20 17 14 15 10 0 0 22 0 25 33 22 14 20 17L 29 0 40 30 38 40 33 69 42 31 100 33 25 20 50 42 41 48 50 401M 0 0 0 43 50 40 0 0 40 0 0 100 50 33 67 0 0 50 0 0N 8 0 0 0 71 13 0 0 0 9 0 0 17 0 25 20 0 22 0 25P 29 0 13 11 33 11 0 0 10 0 0 20 25 43 100 0 40 38 75 50Q 43 0 33 13 0 13 0 33 8 20 50 20 0 40 0 40 25 0 0 50R 17 20 0 22 0 50 50 0 0 6 25 33 0 0 13 50 25 20 0 20S 18 0 0 29 18 20 17 0 14 5 0 0 0 25 14 9 13 33 50 0T 31 100 0 25 43 0 0 0 9 18 0 0 19 10 33 67 33 33 0 14V 20 0 20 19 33 33 0 33 8 17 33 0 33 0 0 27 0 36 33 10W 25 50 0 0 0 40 0 0 0 0 0 0 0 60 0 50 0 0 0 50Y 25 0 0 0 40 22 33 0 33 44 0 0 20 0 0 0 25 20 0 0
98

Figure 3.19. Cleavage data map with probability defined as a percentage, pH 4.0
YWVTSRQP
P1
NMLKIHGF
P1’Scale
FEDCA
A C D E F G H I K L M N P Q R S T V W Y
Table 3.11. Normalized pH 4.0 cleavage data defined as a percentage
100 90 80 70 60 50 40 30 20 10
P1’A C D E F G H I K L M N P Q R S T V W Y
A 20 0 14 13 0 0 0 22 15 25 0 22 20 22 10 19 0 16 0 0C 14 13 0 0 0 0 0 0 0 0 0 0 25 20 0 0 17 0 0 0
P1
D 57 0 8 17 14 11 0 13 6 0 0 0 25 0 0 20 14 0 0 0E 20 0 0 28 22 0 40 13 0 11 50 16 11 50 22 0 29 13 33 22F 22 0 33 33 33 25 0 33 13 44 0 0 29 17 0 43 25 33 100 0G 36 0 0 20 0 7 14 43 0 18 33 27 0 0 0 33 0 8 33 0H 20 25 0 0 0 30 0 33 22 0 0 0 25 0 25 43 0 17 0 50I 9 33 0 0 0 0 0 33 11 0 33 0 0 25 0 14 29 11 0 0K 5 0 0 5 0 0 33 0 8 5 0 25 0 0 0 0 22 14 20 33L 25 0 40 25 63 13 7 31 13 21 0 33 50 60 25 35 18 24 0 301M 0 0 0 14 50 20 0 0 20 0 0 100 50 0 0 0 50 25 0 0N 23 33 0 0 0 0 0 0 33 0 0 0 0 0 0 0 0 22 0 25P 14 0 13 0 33 11 0 0 10 0 0 0 0 14 0 0 0 13 0 0Q 14 0 0 13 0 0 0 33 8 40 0 0 0 20 0 0 0 0 0 25R 17 20 0 0 0 0 0 0 33 0 0 0 40 0 13 0 0 20 0 0S 9 0 8 18 27 0 17 11 0 9 0 0 0 0 0 18 13 33 50 0T 8 0 0 8 14 0 0 0 9 18 0 0 31 0 0 0 0 11 0 29V 5 0 30 6 33 22 20 33 0 17 33 13 0 0 20 18 33 9 33 10W 0 50 0 0 0 20 0 0 0 0 0 0 0 20 0 0 0 0 0 50Y 25 0 0 0 0 11 17 33 17 33 0 0 0 0 25 0 25 40 0 0
99

produced the least amount. This is most likely because at pH 1.0 the proteins are
denatured, therefore it is easier for the pepsin to digest them. Another reason for the
lower number of cleavages at pH 4.0 is the fact that the activity of pepsin is much lower
at this pH than at the lower pH values. If pepsin is not as active then it will not cleave as
often therefore producing less peptides.
3.6 References
Brier, S., G. Maria, et al. (2007). "Purification and characterization of pepsins A1 and A2
from the Antarctic rock cod Trematomus bernacchii." Febs J 274: 6152-66.
Chakraborty, A. B., S. J. Berger, et al. (2007). "Use of an integrated MS--multiplexed
MS/MS data acquisition strategy for high-coverage peptide mapping studies."
Rapid Commun Mass Spectrom 21(5): 730-44.
Jones, R. G. and J. Landon (2002). "Enhanced pepsin digestion: a novel process for
purifying antibody F(ab')(2) fragments in high yield from serum." J Immunol
Methods 263: 57-74.
Millea, K. M., I. S. Krull, et al. (2006). "Integration of multidimensional chromatographic
protein separations with a combined "top-down" and "bottom-up" proteomic
strategy." J Proteome Res 5(1): 135-46.
Plumb, R., J. Castro-Perez, et al. (2004). "Ultra-performance liquid chromatography
coupled to quadrupole-orthogonal time-of-flight mass spectrometry." Rapid
Commun Mass Spectrom 18(19): 2331-7.
100

Plumb, R. S., K. A. Johnson, et al. (2006). "UPLC/MS(E); a new approach for generating
molecular fragment information for biomarker structure elucidation." Rapid
Commun Mass Spectrom 20: 1989-94.
Swartz, M. E. (2005). "Ultra performance liquid chromatography (UPLC): An
introduction." Lc Gc North America: 8-14.
Swartz, M. E. (2005). "UPLC (TM): An introduction and review." Journal of Liquid
Chromatography & Related Technologies 28(7-8): 1253-1263.
Wang, L., H. Pan, et al. (2002). "Hydrogen exchange-mass spectrometry: optimization of
digestion conditions." Mol Cell Proteomics 1: 132-8.
Wu, Y., S. Kaveti, et al. (2006). "Extensive deuterium back-exchange in certain
immobilized pepsin columns used for H/D exchange mass spectrometry." Anal
Chem 78: 1719-23.
101

Figure 3.5. pH 2.5 peptic digest map of Abl
1 GQQPGKVLGD QRREPQGLSE AARWNBKENL LAGPSENDPN LFVALYDFVA
51 SGDNTLSITK GEKLRVLGYN HNGEWCEAQT KNGQGWVPSN YITPVNSLEK
101 HSWYHGPVSR NAAEYLLSSG INGSFLVRES ESSPGQRSIS LRYEGRVYHY
151 RINTASDGKL YVSSESRFNT LAELVHHHST VADGLITTLH YPAPKRNKPT
201 VYGVSPNYDK WEMERTDITM KHKLGGGQYG EVYEGVWKKY SLTVAVKTLK
251 EDTMEVEEFL KEAAVMKEIK HPNLVQLLGV CTREPPFYII TEFMTYGNLL
301 DYLRECNRQE VNAVVLLYMA TQISSAMEYL EKKNFIHRDL AARNCLVGEN
351 HLVKVADFGL SRLMTGDTYT AHAGAKFPIK WTAPESLAYN KFSIKSDVWA
401 FGVLLWEIAT YGMSPYPGID LSQVYELLEK DYRMERPEGC PEKVYELMRA
451 CWQWNPSDRP SFAEIHQAFE TMFQESSISD EVEKELGKEN LYFQGHHHHHQ Q Q Q
501 H
102

Figure 3.6. pH 2.5 peptic digest map of Albumin
1 DTHKSEIAHR FKDLGEEHFK GLVLIAFSQY LQQCPFDEHV KLVNELTEFA
51 KTCVADESHA GCEKSLHTLF GDELCKVASL RETYGDMADC CEKQEPERNE
101 CFLSHKDDSP DLPKLKPDPN TLCDEFKADE KKFWGKYLYE IARRHPYFYA
151 PELLYYANKY NGVFQECCQA EDKGACLLPK IETMREKVLA SSARQRLRCA
201 SIQKFGERAL KAWSVARLSQ KFPKAEFVEV TKLVTDLTKV HKECCHGDLL
251 ECADDRADLA KYICDNQDTI SSKLKECCDK PLLEKSHCIA EVEKDAIPEN
301 LPPLTADFAE DKDVCKNYQE AKDAFLGSFL YEYSRRHPEY AVSVLLRLAK
351 EYEATLEECC AKDDPHACYS TVFDKLKHLV DEPQNLIKQN CDQFEKLGEY
401 GFQNALIVRY TRKVPQVSTP TLVEVSRSLG KVGTRCCTKP ESERMPCTED
451 YLSLILNRLC VLHEKTPVSE KVTKCCTESL VNRRPCFSAL TPDETYVPKA
501 FDEKLFTFHA DICTLPDTEK QIKKQTALVE LLKHKPKATE EQLKTVMENF
551 VAFVDKCCAA DDKEACFAVE GPKLVVSTQT ALA
103

Figure 3.7. pH 2.5 peptic digest map of Aldolase
1 MPHSHPALTP EQKKELSDIA HRIVAPGKGI LAADESTGSI AKRLQSIGTE
51 NTEENRRFYR QLLLTADDRV NPCIGGVILF HETLYQKADD GRPFPQVIKS
101 KGGVVGIKVD KGVVPLAGTN GETTTQGLDG LSERCAQYKK DGADFAKWRC
151 VLKIGEHTPS ALAIMENANV LARYASICQQ NGIVPIVEPE ILPDGDHDLK
201 RCQYVTEKVL AAVYKALSDH HIYLEGTLLK PNMVTPGHAC TQKYSHEEIA
251 MATVTALRRT VPPAVTGVTF LSGGQSEEEA SINLNAINKC PLLKPWALTF
301 SYGRALQASA LKAWGGKKEN LKAAQEEYVK RALANSLACQ GKYTPSGQAG
351 AAASESLFIS NHAY
104

Figure 3.8. pH 2.5 peptic digest map of Amyloglucosidase
1 MSFRSLLALS GLVCTGLANV ISKRATLDSW LSNEATVART AILNNIGADG
51 AWVSGADSGI VVASPSTDNP DYFYTWTRDS GLVLKTLVDL FRNGDTSLLS
101 TIENYISAQA IVQGISNPSG DLSSGAGLGE PKFNVDETAY TGSWGRPQRD
151 GPALRATAMI GFGQWLLDNG YTSTATDIVW PLVRNDLSYV AQYWNQTGYD
201 LWEEVNGSSF FTIAVQHRAL VEGSAFATAV GSSCSWCDSQ APEILCYLQS
251 FWTGSFILAN FDSSRSGKDA NTLLGSIHTF DPEAACDDST FQPCSPRALA
301 NHKEVVDSFR SIYTLNDGLS DSEAVAVGRY PEDTYYNGNP WFLCTLAAAE
351 QLYDALYQWD KQGSLEVTDV SLDFFKALYS DAATGTYSSS SSTYSSIVDA
401 VKTFADGFVS IVETHAASNG SMSEQYDKSD GEQLSARDLT WSYAALLTAN
451 NRRNSVVPAS WGETSASSVP GTCAATSAIG TYSSVTVTSW PSIVATGGTT
501 TTATPTGSGS VTSTSKTTAT ASKTSTSTSS TSCTTPTAVA VTFDLTATTT
551 YGENIYLVGS ISQLGDWETS DGIALSADKY TSSDPLWYVT VTLPAGESFE
601 YKFIRIESDD SVEWESDPNR EYTVPQACGT STATVTDTWR
105

Figure 3.9. pH 2.5 peptic digest map of β-Lactalbumin
1 LIVTQTMKGL DIQKVAGTWY SLAMAASDIS LLDAQSAPLR VYVEELKPTP
51 EGDLEILLQK WENGECAQKK IIAEKTKIPA VFKIDALNEN KVLVLDTDYK
101 KYLLFCMENS AEPEQSLACQ CLVRTPEVDD EALEKFDKAL KALPMHIRLS
151 FNPTQLEEQC HI
106

Figure 3.10. pH 2.5 peptic digest map of Hemoglobin
1 MVLSAADKGN VKAAWGKVGG HAAEYGAEAL ERMFLSFPTT KTYFPHFDLS
51 HGSAQVKGHG AKVAAALTKA VEHLDDLPGA LSELSDLHAH KLRVDPVNFK
Alpha chain
101 LLSHSLLVTL ASHLPSDFTP AVHASLDKFL ANVSTVLTSK YR
P02070|HBB_BOVIN Coverage Map
1 MLTAEEKAAV TAFWGKVKVD EVGGEALGRL LVVYPWTQRF FESFGDLSTA
Beta chain1 MLTAEEKAAV TAFWGKVKVD EVGGEALGRL LVVYPWTQRF FESFGDLSTA
51 DAVMNNPKVK AHGKKVLDSF SNGMKHLDDL KGTFAALSEL HCDKLHVDPE
101 NFKLLGNVLV VVLARNFGKE FTPVLQADFQ KVVAGVANAL AHRYH
1 MVHFTTEENV AVASLWAKVN VEVVGGESLA RLLIVCPWTQ RFFDSFGNLY
51 SESAIMGNPK VKVYGRKVLN SFGNAIKHMD DLKGTFADLS ELHCDKLHVD
Epsilon-2 chain
101 PENFRLLGNM ILIVLATHFS KEFTPQMQAA WQKLTNAVAN ALTHKYH
1 MVHFTTEEKA AVASLWAKVN VEVVGGESLA RLLIVYPWTQ RFFDSFGNLYEpsilon-4 chain
51 SESAIMGNPK VKAHGRKVLN SFGNAIEHMD DLKGTFADLS ELHCDKLHVD
101 PENFRLLGNM ILIVLATHFS KEFTPQMQAS WQKLTNAVAN ALAHKYH
107

Figure 3.11. pH 2.5 peptic digest map of Myoglobin
1 MGLSDGEWQQ VLNVWGKVEA DIAGHGQEVL IRLFTGHPET LEKFDKFKHL
51 KTEAEMKASE DLKKHGTVVL TALGGILKKK GHHEAELKPL AQSHATKHKI
101 PIKYLEFISD AIIHVLHSKH PGDFGADAQG AMTKALELFR NDIAAKYKEL
151 GFQG
108

Figure 3.12. pH 2.5 peptic digest map of Nef
1 MGSSHHHHHH SSGLVPRGSH MGGKWSKSSV IGWPAVRERM RRAEPAADGV
51 GAVSRDLEKH GAITSSNTAA NNAACAWLEA QEEEEVGFPV TPQVPLRPMT
101 YKAAVDLSHF LKEKGGLEGL IHSQRRQDIL DLWIYHTQGY FPDWQNYTPG
151 PGVRYPLTFG WCYKLVPVEP DKVEEANKGE NTSLLHPVSL HGMDDPEREV
201 LEWRFDSRLA FHHVARELHP EYFKNC
109

Figure 3.13. pH 2.5 peptic digest map of Ubiquitin
1 MQIFVKTLTG KTITLEVEPS DTIENVKAKI QDKEGIPPDQ QRLIFAGKQL
5151 EDGRTLSDYN IQKESTLHLV LRLRGG
110

Table 3.3. pH 1.0 peptides Protein Residues Peptide Sequence
Abl 1-20 GQQPGKVLGDQRREPQGLSE Abl 29-57 NLLAGPSENDPNLFVALYDFVASGDNTLS Abl 31-41 LAGPSENDPNL Abl 31-42 LAGPSENDPNLF Abl 32-50 AGPSENDPNLFVALYDFVA Abl 42-56 FVALYDFVASGDNTL Abl 46-64 YDFVASGDNTLSITKGEKL Abl 46-74 YDFVASGDNTLSITKGEKLRVLGYNHNGE Abl 46-75 YDFVASGDNTLSITKGEKLRVLGYNHNGEW Abl 48-75 FVASGDNTLSITKGEKLRVLGYNHNGEW Abl 48-77 FVASGDNTLSITKGEKLRVLGYNHNGEWCE Abl 49-63 VASGDNTLSITKGEK Abl 49-64 VASGDNTLSITKGEKL Abl 49-75 VASGDNTLSITKGEKLRVLGYNHNGEW Abl 50-64 ASGDNTLSITKGEKL Abl 51-64 SGDNTLSITKGEKL Abl 57-75 SITKGEKLRVLGYNHNGEW Abl 65-77 RVLGYNHNGEWCE Abl 75-84 WCEAQTKNGQ Abl 76-90 CEAQTKNGQGWVPSN Abl 76-98 CEAQTKNGQGWVPSNYITPVNSL Abl 77-114 EAQTKNGQGWVPSNYITPVNSLEKHSWYHGPVSRNAAE Abl 78-90 AQTKNGQGWVPSN Abl 85-131 GWVPSNYITPVNSLEKHSWYHGPVSRNAAEYLLSSGINGSFLVRESE Abl 86-104 WVPSNYITPVNSLEKHSWY Abl 88-114 PSNYITPVNSLEKHSWYHGPVSRNAAE Abl 91-114 YITPVNSLEKHSWYHGPVSRNAAE Abl 92-114 ITPVNSLEKHSWYHGPVSRNAAE Abl 93-102 TPVNSLEKHS Abl 93-114 TPVNSLEKHSWYHGPVSRNAAE Abl 98-110 LEKHSWYHGPVSR Abl 99-114 EKHSWYHGPVSRNAAE Abl 107-122 PVSRNAAEYLLSSGIN Abl 107-125 PVSRNAAEYLLSSGINGSF Abl 110-123 RNAAEYLLSSGING Abl 111-123 NAAEYLLSSGING Abl 115-125 YLLSSGINGSF Abl 115-126 YLLSSGINGSFL Abl 117-127 LSSGINGSFLV Abl 125-141 FLVRESESSPGQRSISL Abl 126-140 LVRESESSPGQRSIS Abl 126-141 LVRESESSPGQRSISL Abl 126-160 LVRESESSPGQRSISLRYEGRVYHYRINTASDGKL Abl 127-141 VRESESSPGQRSISL Abl 140-160 SLRYEGRVYHYRINTASDGKL Abl 141-160 LRYEGRVYHYRINTASDGKL Abl 142-160 RYEGRVYHYRINTASDGKL Abl 154-163 TASDGKLYVS Abl 159-185 KLYVSSESRFNTLAELVHHHSTVADGL Abl 161-171 YVSSESRFNTL Abl 161-173 YVSSESRFNTLAE Abl 172-185 AELVHHHSTVADGL Abl 172-202 AELVHHHSTVADGLITTLHYPAPKRNKPTVY Abl 172-213 AELVHHHSTVADGLITTLHYPAPKRNKPTVYGVSPNYDKWEM Abl 174-185 LVHHHSTVADGL Abl 186-213 ITTLHYPAPKRNKPTVYGVSPNYDKWEM Abl 198-212 KPTVYGVSPNYDKWE Abl 203-213 GVSPNYDKWEM Abl 214-236 ERTDITMKHKLGGGQYGEVYEGV Abl 214-237 ERTDITMKHKLGGGQYGEVYEGVW
111
Abl 214-243 ERTDITMKHKLGGGQYGEVYEGVWKKYSLT Abl 214-245 ERTDITMKHKLGGGQYGEVYEGVWKKYSLTVA Abl 214-255 ERTDITMKHKLGGGQYGEVYEGVWKKYSLTVAVKTLKEDTME Abl 215-230 RTDITMKHKLGGGQYG Abl 219-246 TMKHKLGGGQYGEVYEGVWKKYSLTVAV Abl 222-244 HKLGGGQYGEVYEGVWKKYSLTV Abl 233-245 YEGVWKKYSLTVA Abl 244-255 VAVKTLKEDTME Abl 246-255 VKTLKEDTME Abl 256-274 VEEFLKEAAVMKEIKHPNL Abl 258-276 EFLKEAAVMKEIKHPNLVQ Abl 259-274 FLKEAAVMKEIKHPNL Abl 261-274 KEAAVMKEIKHPNL Abl 275-292 VQLLGVCTREPPFYIITE Abl 278-292 LGVCTREPPFYIITE Abl 278-293 LGVCTREPPFYIITEF Abl 282-292 TREPPFYIITE Abl 282-293 TREPPFYIITEF Abl 293-313 FMTYGNLLDYLRECNRQEVNA Abl 294-313 MTYGNLLDYLRECNRQEVNA Abl 302-313 YLRECNRQEVNA Abl 316-342 LLYMATQISSAMEYLEKKNFIHRDLAA Abl 317-346 LYMATQISSAMEYLEKKNFIHRDLAARNCL Abl 318-346 YMATQISSAMEYLEKKNFIHRDLAARNCL Abl 321-335 TQISSAMEYLEKKNF Abl 327-338 MEYLEKKNFIHR Abl 328-346 EYLEKKNFIHRDLAARNCL Abl 329-346 YLEKKNFIHRDLAARNCL Abl 330-346 LEKKNFIHRDLAARNCL Abl 331-351 EKKNFIHRDLAARNCLVGENH Abl 339-354 DLAARNCLVGENHLVK Abl 347-358 VGENHLVKVADF Abl 347-360 VGENHLVKVADFGL Abl 347-363 VGENHLVKVADFGLSRL Abl 348-358 GENHLVKVADF Abl 348-360 GENHLVKVADFGL Abl 348-363 GENHLVKVADFGLSRL Abl 364-387 MTGDTYTAHAGAKFPIKWTAPESL Abl 365-377 TGDTYTAHAGAKF Abl 366-377 GDTYTAHAGAKF Abl 369-388 YTAHAGAKFPIKWTAPESLA Abl 378-388 PIKWTAPESLA Abl 380-398 KWTAPESLAYNKFSIKSDV Abl 383-392 APESLAYNKF Abl 383-393 APESLAYNKFS Abl 394-424 IKSDVWAFGVLLWEIATYGMSPYPGIDLSQV Abl 397-424 DVWAFGVLLWEIATYGMSPYPGIDLSQV Abl 406-421 WEIATYGMSPYPGIDL Abl 416-425 YPGIDLSQVY Abl 416-430 YPGIDLSQVYELLEK Abl 428-446 LEKDYRMERPEGCPEKVYE Abl 429-437 EKDYRMERP Abl 432-448 YRMERPEGCPEKVYELM Abl 447-462 LMRACWQWNPSDRPSF Abl 448-462 MRACWQWNPSDRPSF Abl 458-499 DRPSFAEIHQAFETMFQESSISDEVEKELGKENLYFQGHHHH Abl 463-472 AEIHQAFETM Abl 472-495 MFQESSISDEVEKELGKENLYFQG Abl 473-491 FQESSISDEVEKELGKENL Abl 474-491 QESSISDEVEKELGKENL Abl 476-489 SSISDEVEKELGKE Abl 481-495 EVEKELGKENLYFQG Abl 485-495 ELGKENLYFQG
112

Albumin 1-21 DTHKSEIAHRFKDLGEEHFKG Albumin 1-24 DTHKSEIAHRFKDLGEEHFKGLVL Albumin 2-24 THKSEIAHRFKDLGEEHFKGLVL Albumin 3-24 HKSEIAHRFKDLGEEHFKGLVL Albumin 21-43 GLVLIAFSQYLQQCPFDEHVKLV Albumin 23-35 VLIAFSQYLQQCP Albumin 25-38 IAFSQYLQQCPFDE Albumin 25-42 IAFSQYLQQCPFDEHVKL Albumin 25-45 IAFSQYLQQCPFDEHVKLVNE Albumin 27-38 FSQYLQQCPFDE Albumin 30-45 YLQQCPFDEHVKLVNE Albumin 31-45 LQQCPFDEHVKLVNE Albumin 31-77 LQQCPFDEHVKLVNELTEFAKTCVADESHAGCEKSLHTLFGDELCKV Albumin 34-81 CPFDEHVKLVNELTEFAKTCVADESHAGCEKSLHTLFGDELCKVASLR Albumin 59-82 HAGCEKSLHTLFGDELCKVASLRE Albumin 107-121 DDSPDLPKLKPDPNT Albumin 112-133 LPKLKPDPNTLCDEFKADEKKF Albumin 113-133 PKLKPDPNTLCDEFKADEKKF Albumin 125-142 EFKADEKKFWGKYLYEIA Albumin 126-144 FKADEKKFWGKYLYEIARR Albumin 138-154 LYEIARRHPYFYAPELL Albumin 141-153 IARRHPYFYAPEL Albumin 141-154 IARRHPYFYAPELL Albumin 141-155 IARRHPYFYAPELLY Albumin 145-156 HPYFYAPELLYY Albumin 154-164 LYYANKYNGVF Albumin 169-191 QAEDKGACLLPKIETMREKVLAS Albumin 170-188 AEDKGACLLPKIETMREKV Albumin 177-191 LLPKIETMREKVLAS Albumin 177-208 LLPKIETMREKVLASSARQRLRCASIQKFGER Albumin 199-211 CASIQKFGERALK Albumin 212-227 AWSVARLSQKFPKAEF Albumin 213-227 WSVARLSQKFPKAEF Albumin 275-293 KECCDKPLLEKSHCIAEVE Albumin 276-290 ECCDKPLLEKSHCIA Albumin 277-300 CCDKPLLEKSHCIAEVEKDAIPEN Albumin 279-319 DKPLLEKSHCIAEVEKDAIPENLPPLTADFAEDKDVCKNYQ Albumin 292-303 VEKDAIPENLPP Albumin 328-338 SFLYEYSRRHP Albumin 331-345 YEYSRRHPEYAVSVL Albumin 334-354 SRRHPEYAVSVLLRLAKEYEA Albumin 337-357 HPEYAVSVLLRLAKEYEATLE Albumin 355-399 TLEECCAKDDPHACYSTVFDKLKHLVDEPQNLIKQNCDQFEKLGE Albumin 369-390 YSTVFDKLKHLVDEPQNLIKQN Albumin 402-422 FQNALIVRYTRKVPQVSTPTL Albumin 406-422 LIVRYTRKVPQVSTPTL Albumin 407-419 IVRYTRKVPQVST Albumin 407-421 IVRYTRKVPQVSTPT Albumin 407-422 IVRYTRKVPQVSTPTL Albumin 408-421 VRYTRKVPQVSTPT Albumin 418-441 STPTLVEVSRSLGKVGTRCCTKPE Albumin 419-432 TPTLVEVSRSLGKV Albumin 419-440 TPTLVEVSRSLGKVGTRCCTKP Albumin 443-463 ERMPCTEDYLSLILNRLCVLH Albumin 448-466 TEDYLSLILNRLCVLHEKT Albumin 460-470 CVLHEKTPVSE Albumin 460-511 CVLHEKTPVSEKVTKCCTESLVNRRPCFSALTPDETYVPKAFDEKLFTFHAD Albumin 464-482 EKTPVSEKVTKCCTESLVN Albumin 490-505 LTPDETYVPKAFDEKL Albumin 496-515 YVPKAFDEKLFTFHADICTL Albumin 529-543 VELLKHKPKATEEQL Albumin 529-545 VELLKHKPKATEEQLKT Albumin 529-547 VELLKHKPKATEEQLKTVM
113

Albumin 529-549 VELLKHKPKATEEQLKTVMEN Albumin 529-550 VELLKHKPKATEEQLKTVMENF Albumin 531-545 LLKHKPKATEEQLKT Albumin 532-550 LKHKPKATEEQLKTVMENF Albumin 555-579 DKCCAADDKEACFAVEGPKLVVSTQ Albumin 568-580 AVEGPKLVVSTQT Aldolase 2-31 PHSHPALTPEQKKELSDIAHRIVAPGKGIL Aldolase 3-15 HSHPALTPEQKKE Aldolase 4-11 SHPALTPE Aldolase 5-15 HPALTPEQKKE Aldolase 5-22 HPALTPEQKKELSDIAHR Aldolase 7-16 ALTPEQKKEL Aldolase 9-35 TPEQKKELSDIAHRIVAPGKGILAADE Aldolase 10-34 PEQKKELSDIAHRIVAPGKGILAAD Aldolase 31-63 LAADESTGSIAKRLQSIGTENTEENRRFYRQLL Aldolase 32-54 AADESTGSIAKRLQSIGTENTEE Aldolase 32-58 AADESTGSIAKRLQSIGTENTEENRRF Aldolase 32-63 AADESTGSIAKRLQSIGTENTEENRRFYRQLL Aldolase 32-65 AADESTGSIAKRLQSIGTENTEENRRFYRQLLLT Aldolase 32-67 AADESTGSIAKRLQSIGTENTEENRRFYRQLLLTAD Aldolase 33-58 ADESTGSIAKRLQSIGTENTEENRRF Aldolase 34-58 DESTGSIAKRLQSIGTENTEENRRF Aldolase 34-64 DESTGSIAKRLQSIGTENTEENRRFYRQLLL Aldolase 35-58 ESTGSIAKRLQSIGTENTEENRRF Aldolase 35-74 ESTGSIAKRLQSIGTENTEENRRFYRQLLLTADDRVNPCI Aldolase 50-70 ENTEENRRFYRQLLLTADDRV Aldolase 52-71 TEENRRFYRQLLLTADDRVN Aldolase 55-64 NRRFYRQLLL Aldolase 55-92 NRRFYRQLLLTADDRVNPCIGGVILFHETLYQKADDGR Aldolase 64-79 LTADDRVNPCIGGVIL Aldolase 65-79 TADDRVNPCIGGVIL Aldolase 66-79 ADDRVNPCIGGVIL Aldolase 68-79 DRVNPCIGGVIL Aldolase 78-102 ILFHETLYQKADDGRPFPQVIKSKG Aldolase 80-95 FHETLYQKADDGRPFP Aldolase 85-106 YQKADDGRPFPQVIKSKGGVVG Aldolase 85-131 YQKADDGRPFPQVIKSKGGVVGIKVDKGVVPLAGTNGETTTQGLDGL Aldolase 96-106 QVIKSKGGVVG Aldolase 107-131 IKVDKGVVPLAGTNGETTTQGLDGL Aldolase 108-125 KVDKGVVPLAGTNGETTT Aldolase 112-128 GVVPLAGTNGETTTQGL Aldolase 120-147 NGETTTQGLDGLSERCAQYKKDGADFAK Aldolase 139-166 KKDGADFAKWRCVLKIGEHTPSALAIME Aldolase 148-164 WRCVLKIGEHTPSALAI Aldolase 154-170 IGEHTPSALAIMENANV Aldolase 154-171 IGEHTPSALAIMENANVL Aldolase 163-188 AIMENANVLARYASICQQNGIVPIVE Aldolase 169-188 NVLARYASICQQNGIVPIVE Aldolase 171-188 LARYASICQQNGIVPIVE Aldolase 171-207 LARYASICQQNGIVPIVEPEILPDGDHDLKRCQYVTE Aldolase 171-210 LARYASICQQNGIVPIVEPEILPDGDHDLKRCQYVTEKVL Aldolase 172-188 ARYASICQQNGIVPIVE Aldolase 172-207 ARYASICQQNGIVPIVEPEILPDGDHDLKRCQYVTE Aldolase 175-188 ASICQQNGIVPIVE Aldolase 179-207 QQNGIVPIVEPEILPDGDHDLKRCQYVTE Aldolase 184-204 VPIVEPEILPDGDHDLKRCQY Aldolase 188-199 EPEILPDGDHDL Aldolase 189-203 PEILPDGDHDLKRCQ Aldolase 189-204 PEILPDGDHDLKRCQY Aldolase 189-207 PEILPDGDHDLKRCQYVTE Aldolase 193-204 PDGDHDLKRCQY Aldolase 194-207 DGDHDLKRCQYVTE Aldolase 208-226 KVLAAVYKALSDHHIYLEG
114

Aldolase 211-229 AAVYKALSDHHIYLEGTLL Aldolase 211-239 AAVYKALSDHHIYLEGTLLKPNMVTPGHA Aldolase 211-247 AAVYKALSDHHIYLEGTLLKPNMVTPGHACTQKYSHE Aldolase 212-229 AVYKALSDHHIYLEGTLL Aldolase 212-233 AVYKALSDHHIYLEGTLLKPNM Aldolase 212-247 AVYKALSDHHIYLEGTLLKPNMVTPGHACTQKYSHE Aldolase 213-229 VYKALSDHHIYLEGTLL Aldolase 213-247 VYKALSDHHIYLEGTLLKPNMVTPGHACTQKYSHE Aldolase 213-251 VYKALSDHHIYLEGTLLKPNMVTPGHACTQKYSHEEIAM Aldolase 214-226 YKALSDHHIYLEG Aldolase 214-239 YKALSDHHIYLEGTLLKPNMVTPGHA Aldolase 214-250 YKALSDHHIYLEGTLLKPNMVTPGHACTQKYSHEEIA Aldolase 215-247 KALSDHHIYLEGTLLKPNMVTPGHACTQKYSHE Aldolase 227-247 TLLKPNMVTPGHACTQKYSHE Aldolase 227-250 TLLKPNMVTPGHACTQKYSHEEIA Aldolase 229-247 LKPNMVTPGHACTQKYSHE Aldolase 230-247 KPNMVTPGHACTQKYSHE Aldolase 230-251 KPNMVTPGHACTQKYSHEEIAM Aldolase 236-251 PGHACTQKYSHEEIAM Aldolase 238-254 HACTQKYSHEEIAMATV Aldolase 240-251 CTQKYSHEEIAM Aldolase 240-280 CTQKYSHEEIAMATVTALRRTVPPAVTGVTFLSGGQSEEEA Aldolase 248-260 EIAMATVTALRRT Aldolase 250-265 AMATVTALRRTVPPAV Aldolase 252-267 ATVTALRRTVPPAVTG Aldolase 252-270 ATVTALRRTVPPAVTGVTF Aldolase 252-276 ATVTALRRTVPPAVTGVTFLSGGQS Aldolase 255-285 TALRRTVPPAVTGVTFLSGGQSEEEASINLN Aldolase 257-270 LRRTVPPAVTGVTF Aldolase 258-276 RRTVPPAVTGVTFLSGGQS Aldolase 264-292 AVTGVTFLSGGQSEEEASINLNAINKCPL Aldolase 265-287 VTGVTFLSGGQSEEEASINLNAI Aldolase 265-289 VTGVTFLSGGQSEEEASINLNAINK Aldolase 267-304 GVTFLSGGQSEEEASINLNAINKCPLLKPWALTFSYGR Aldolase 269-294 TFLSGGQSEEEASINLNAINKCPLLK Aldolase 271-284 LSGGQSEEEASINL Aldolase 285-299 NAINKCPLLKPWALT Aldolase 285-300 NAINKCPLLKPWALTF Aldolase 287-299 INKCPLLKPWALT Aldolase 289-316 KCPLLKPWALTFSYGRALQASALKAWGG Aldolase 290-316 CPLLKPWALTFSYGRALQASALKAWGG Aldolase 300-311 FSYGRALQASAL Aldolase 300-326 FSYGRALQASALKAWGGKKENLKAAQE Aldolase 300-327 FSYGRALQASALKAWGGKKENLKAAQEE Aldolase 309-327 SALKAWGGKKENLKAAQEE Aldolase 312-326 KAWGGKKENLKAAQE Aldolase 318-345 KENLKAAQEEYVKRALANSLACQGKYTP Aldolase 322-330 KAAQEEYVK Aldolase 326-338 EEYVKRALANSLA Aldolase 327-337 EYVKRALANSL Aldolase 328-337 YVKRALANSL Aldolase 328-352 YVKRALANSLACQGKYTPSGQAGAA Aldolase 328-355 YVKRALANSLACQGKYTPSGQAGAAASE Aldolase 328-357 YVKRALANSLACQGKYTPSGQAGAAASESL Aldolase 332-347 ALANSLACQGKYTPSG Aldolase 337-357 LACQGKYTPSGQAGAAASESL Aldolase 338-355 ACQGKYTPSGQAGAAASE Aldolase 338-357 ACQGKYTPSGQAGAAASESL Aldolase 338-358 ACQGKYTPSGQAGAAASESLF Aldolase 347-362 GQAGAAASESLFISNH Aldolase 350-358 GAAASESLF
Amyloglucosidase 25-43 ATLDSWLSNEATVARTAIL Amyloglucosidase 31-43 LSNEATVARTAIL
115

Amyloglucosidase 43-58 LNNIGADGAWVSGADS Amyloglucosidase 53-73 VSGADSGIVVASPSTDNPDYF Amyloglucosidase 54-69 SGADSGIVVASPSTDN Amyloglucosidase 55-67 GADSGIVVASPST Amyloglucosidase 60-73 IVVASPSTDNPDYF Amyloglucosidase 74-84 YTWTRDSGLVL Amyloglucosidase 109-133 QAIVQGISNPSGDLSSGAGLGEPKF Amyloglucosidase 110-133 AIVQGISNPSGDLSSGAGLGEPKF Amyloglucosidase 111-133 IVQGISNPSGDLSSGAGLGEPKF Amyloglucosidase 134-154 NVDETAYTGSWGRPQRDGPAL Amyloglucosidase 134-156 NVDETAYTGSWGRPQRDGPALRA Amyloglucosidase 160-177 IGFGQWLLDNGYTSTATD Amyloglucosidase 178-189 IVWPLVRNDLSY Amyloglucosidase 212-230 TIAVQHRALVEGSAFATAV Amyloglucosidase 309-324 FRSIYTLNDGLSDSEA Amyloglucosidase 310-324 RSIYTLNDGLSDSEA Amyloglucosidase 325-343 VAVGRYPEDTYYNGNPWFL Amyloglucosidase 389-413 SSSSTYSSIVDAVKTFADGFVSIVE β-Lactoglobulin 1-11 LIVTQTMKGLD β-Lactoglobulin 1-13 LIVTQTMKGLDIQ β-Lactoglobulin 1-19 LIVTQTMKGLDIQKVAGTW β-Lactoglobulin 10-19 LDIQKVAGTW β-Lactoglobulin 13-38 QKVAGTWYSLAMAASDISLLDAQSAP β-Lactoglobulin 32-42 LDAQSAPLRVY β-Lactoglobulin 42-54 YVEELKPTPEGDL β-Lactoglobulin 42-57 YVEELKPTPEGDLEIL β-Lactoglobulin 43-54 VEELKPTPEGDL β-Lactoglobulin 83-95 KIDALNENKVLVL β-Lactoglobulin 132-149 ALEKFDKALKALPMHIRL β-Lactoglobulin 147-156 IRLSFNPTQL
Hemoglobin-alpha 1-32 MVLSAADKGNVKAAWGKVGGHAAEYGAEALER Hemoglobin-alpha 2-24 VLSAADKGNVKAAWGKVGGHAAE Hemoglobin-alpha 2-25 VLSAADKGNVKAAWGKVGGHAAEY Hemoglobin-alpha 2-28 VLSAADKGNVKAAWGKVGGHAAEYGAE Hemoglobin-alpha 2-30 VLSAADKGNVKAAWGKVGGHAAEYGAEAL Hemoglobin-alpha 3-22 LSAADKGNVKAAWGKVGGHA Hemoglobin-alpha 3-29 LSAADKGNVKAAWGKVGGHAAEYGAEA Hemoglobin-alpha 4-24 SAADKGNVKAAWGKVGGHAAE Hemoglobin-alpha 6-41 ADKGNVKAAWGKVGGHAAEYGAEALERMFLSFPTTK Hemoglobin-alpha 7-18 DKGNVKAAWGKV Hemoglobin-alpha 7-43 DKGNVKAAWGKVGGHAAEYGAEALERMFLSFPTTKTY Hemoglobin-alpha 9-22 GNVKAAWGKVGGHA Hemoglobin-alpha 16-26 GKVGGHAAEYG Hemoglobin-alpha 30-43 LERMFLSFPTTKTY Hemoglobin-alpha 30-57 LERMFLSFPTTKTYFPHFDLSHGSAQVK Hemoglobin-alpha 34-47 FLSFPTTKTYFPHF Hemoglobin-alpha 34-49 FLSFPTTKTYFPHFDL Hemoglobin-alpha 34-68 FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALT Hemoglobin-alpha 34-74 FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHL Hemoglobin-alpha 34-81 FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGAL Hemoglobin-alpha 34-84 FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSEL Hemoglobin-alpha 35-60 LSFPTTKTYFPHFDLSHGSAQVKGHG Hemoglobin-alpha 35-72 LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVE Hemoglobin-alpha 35-81 LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGAL Hemoglobin-alpha 35-83 LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSE Hemoglobin-alpha 35-84 LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSEL Hemoglobin-alpha 36-47 SFPTTKTYFPHF Hemoglobin-alpha 38-58 PTTKTYFPHFDLSHGSAQVKG Hemoglobin-alpha 38-65 PTTKTYFPHFDLSHGSAQVKGHGAKVAA Hemoglobin-alpha 38-81 PTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGAL Hemoglobin-alpha 38-83 PTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSE Hemoglobin-alpha 48-67 DLSHGSAQVKGHGAKVAAAL Hemoglobin-alpha 48-72 DLSHGSAQVKGHGAKVAAALTKAVE
116

Hemoglobin-alpha 48-81 DLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGAL Hemoglobin-alpha 48-84 DLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSEL Hemoglobin-alpha 49-78 LSHGSAQVKGHGAKVAAALTKAVEHLDDLP Hemoglobin-alpha 51-76 HGSAQVKGHGAKVAAALTKAVEHLDD Hemoglobin-alpha 53-86 SAQVKGHGAKVAAALTKAVEHLDDLPGALSELSD Hemoglobin-alpha 60-78 GAKVAAALTKAVEHLDDLP Hemoglobin-alpha 65-81 AALTKAVEHLDDLPGAL Hemoglobin-alpha 66-81 ALTKAVEHLDDLPGAL Hemoglobin-alpha 67-81 LTKAVEHLDDLPGAL Hemoglobin-alpha 67-92 LTKAVEHLDDLPGALSELSDLHAHKL Hemoglobin-alpha 67-99 LTKAVEHLDDLPGALSELSDLHAHKLRVDPVNF Hemoglobin-alpha 68-99 TKAVEHLDDLPGALSELSDLHAHKLRVDPVNF Hemoglobin-alpha 69-97 KAVEHLDDLPGALSELSDLHAHKLRVDPV Hemoglobin-alpha 81-116 LSELSDLHAHKLRVDPVNFKLLSHSLLVTLASHLPS Hemoglobin-alpha 82-99 SELSDLHAHKLRVDPVNF Hemoglobin-alpha 84-99 LSDLHAHKLRVDPVNF Hemoglobin-alpha 85-98 SDLHAHKLRVDPVN Hemoglobin-alpha 85-99 SDLHAHKLRVDPVNF Hemoglobin-alpha 86-104 DLHAHKLRVDPVNFKLLSH Hemoglobin-alpha 87-96 LHAHKLRVDP Hemoglobin-alpha 88-99 HAHKLRVDPVNF Hemoglobin-alpha 94-105 VDPVNFKLLSHS Hemoglobin-alpha 94-106 VDPVNFKLLSHSL Hemoglobin-alpha 94-107 VDPVNFKLLSHSLL Hemoglobin-alpha 94-124 VDPVNFKLLSHSLLVTLASHLPSDFTPAVHA Hemoglobin-alpha 103-136 SHSLLVTLASHLPSDFTPAVHASLDKFLANVSTV Hemoglobin-alpha 105-126 SLLVTLASHLPSDFTPAVHASL Hemoglobin-alpha 106-135 LLVTLASHLPSDFTPAVHASLDKFLANVST Hemoglobin-alpha 107-137 LVTLASHLPSDFTPAVHASLDKFLANVSTVL Hemoglobin-alpha 108-126 VTLASHLPSDFTPAVHASL Hemoglobin-alpha 108-129 VTLASHLPSDFTPAVHASLDKF Hemoglobin-alpha 108-133 VTLASHLPSDFTPAVHASLDKFLANV Hemoglobin-alpha 108-134 VTLASHLPSDFTPAVHASLDKFLANVS Hemoglobin-alpha 108-137 VTLASHLPSDFTPAVHASLDKFLANVSTVL Hemoglobin-alpha 109-131 TLASHLPSDFTPAVHASLDKFLA Hemoglobin-alpha 109-133 TLASHLPSDFTPAVHASLDKFLANV Hemoglobin-alpha 110-126 LASHLPSDFTPAVHASL Hemoglobin-alpha 110-128 LASHLPSDFTPAVHASLDK Hemoglobin-alpha 110-129 LASHLPSDFTPAVHASLDKF Hemoglobin-alpha 111-126 ASHLPSDFTPAVHASL Hemoglobin-alpha 111-129 ASHLPSDFTPAVHASLDKF Hemoglobin-alpha 111-133 ASHLPSDFTPAVHASLDKFLANV Hemoglobin-alpha 111-134 ASHLPSDFTPAVHASLDKFLANVS Hemoglobin-alpha 111-137 ASHLPSDFTPAVHASLDKFLANVSTVL Hemoglobin-alpha 112-141 SHLPSDFTPAVHASLDKFLANVSTVLTSKY Hemoglobin-alpha 119-133 TPAVHASLDKFLANV Hemoglobin-alpha 121-133 AVHASLDKFLANV Hemoglobin-beta 3-13 TAEEKAAVTAF Hemoglobin-beta 12-30 AFWGKVKVDEVGGEALGRL Hemoglobin-beta 14-30 WGKVKVDEVGGEALGRL Hemoglobin-beta 28-52 GRLLVVYPWTQRFFESFGDLSTADA Hemoglobin-beta 31-44 LVVYPWTQRFFESF Hemoglobin-beta 31-84 LVVYPWTQRFFESFGDLSTADAVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 41-84 FESFGDLSTADAVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 45-84 GDLSTADAVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 48-80 STADAVMNNPKVKAHGKKVLDSFSNGMKHLDDL Hemoglobin-beta 48-84 STADAVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 50-84 ADAVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 51-84 DAVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 51-85 DAVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTFA Hemoglobin-beta 52-62 AVMNNPKVKAH Hemoglobin-beta 52-84 AVMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 53-84 VMNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF
117

Hemoglobin-beta 54-65 MNNPKVKAHGKK Hemoglobin-beta 54-69 MNNPKVKAHGKKVLDS Hemoglobin-beta 54-84 MNNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 55-84 NNPKVKAHGKKVLDSFSNGMKHLDDLKGTF Hemoglobin-beta 59-90 VKAHGKKVLDSFSNGMKHLDDLKGTFAALSEL Hemoglobin-beta 60-91 KAHGKKVLDSFSNGMKHLDDLKGTFAALSELH Hemoglobin-beta 72-90 NGMKHLDDLKGTFAALSEL Hemoglobin-beta 75-95 KHLDDLKGTFAALSELHCDKL Hemoglobin-beta 77-91 LDDLKGTFAALSELH Hemoglobin-beta 85-101 AALSELHCDKLHVDPEN Hemoglobin-beta 85-109 AALSELHCDKLHVDPENFKLLGNVL Hemoglobin-beta 88-109 SELHCDKLHVDPENFKLLGNVL Hemoglobin-beta 91-101 HCDKLHVDPEN Hemoglobin-beta 95-108 LHVDPENFKLLGNV Hemoglobin-beta 112-131 VLARNFGKEFTPVLQADFQK Hemoglobin-beta 113-123 LARNFGKEFTP Hemoglobin-beta 113-124 LARNFGKEFTPV Hemoglobin-beta 113-132 LARNFGKEFTPVLQADFQKV Hemoglobin-beta 114-125 ARNFGKEFTPVL Hemoglobin-beta 114-127 ARNFGKEFTPVLQA Hemoglobin-beta 114-132 ARNFGKEFTPVLQADFQKV Hemoglobin-beta 116-133 NFGKEFTPVLQADFQKVV Hemoglobin-beta 118-133 GKEFTPVLQADFQKVV Hemoglobin-beta 118-138 GKEFTPVLQADFQKVVAGVAN Hemoglobin-beta 120-135 EFTPVLQADFQKVVAG Hemoglobin-beta 126-139 QADFQKVVAGVANA
Hemoglobin-epsilon 2 4-30 FTTEENVAVASLWAKVNVEVVGGESLA Hemoglobin-epsilon 2 8-49 ENVAVASLWAKVNVEVVGGESLARLLIVCPWTQRFFDSFGNL Hemoglobin-epsilon 2 37-73 PWTQRFFDSFGNLYSESAIMGNPKVKVYGRKVLNSFG Hemoglobin-epsilon 2 67-83 KVLNSFGNAIKHMDDLK Hemoglobin-epsilon 2 93-103 HCDKLHVDPEN Hemoglobin-epsilon 2 107-140 LGNMILIVLATHFSKEFTPQMQAAWQKLTNAVAN Hemoglobin-epsilon 2 112-136 LIVLATHFSKEFTPQMQAAWQKLTN Hemoglobin-epsilon 2 123-146 FTPQMQAAWQKLTNAVANALTHKY Hemoglobin-epsilon 2 125-138 PQMQAAWQKLTNAV Hemoglobin-epsilon 2 128-135 QAAWQKLT Hemoglobin-epsilon 4 10-18 AAVASLWAK Hemoglobin-epsilon 4 21-33 VEVVGGESLARLL Hemoglobin-epsilon 4 21-64 VEVVGGESLARLLIVYPWTQRFFDSFGNLYSESAIMGNPKVKAH Hemoglobin-epsilon 4 47-76 GNLYSESAIMGNPKVKAHGRKVLNSFGNAI Hemoglobin-epsilon 4 52-80 ESAIMGNPKVKAHGRKVLNSFGNAIEHMD Hemoglobin-epsilon 4 54-79 AIMGNPKVKAHGRKVLNSFGNAIEHM Hemoglobin-epsilon 4 62-85 KAHGRKVLNSFGNAIEHMDDLKGT Hemoglobin-epsilon 4 67-92 KVLNSFGNAIEHMDDLKGTFADLSEL Hemoglobin-epsilon 4 93-103 HCDKLHVDPEN Hemoglobin-epsilon 4 106-140 LLGNMILIVLATHFSKEFTPQMQASWQKLTNAVAN Hemoglobin-epsilon 4 115-143 LATHFSKEFTPQMQASWQKLTNAVANALA Hemoglobin-epsilon 4 121-135 KEFTPQMQASWQKLT
Myoglobin 2-12 GLSDGEWQQVL Myoglobin 2-14 GLSDGEWQQVLNV Myoglobin 2-15 GLSDGEWQQVLNVW Myoglobin 2-20 GLSDGEWQQVLNVWGKVEA Myoglobin 2-30 GLSDGEWQQVLNVWGKVEADIAGHGQEVL Myoglobin 8-30 WQQVLNVWGKVEADIAGHGQEVL Myoglobin 9-30 QQVLNVWGKVEADIAGHGQEVL Myoglobin 10-30 QVLNVWGKVEADIAGHGQEVL Myoglobin 13-30 NVWGKVEADIAGHGQEVL Myoglobin 13-31 NVWGKVEADIAGHGQEVLI Myoglobin 15-30 WGKVEADIAGHGQEVL Myoglobin 15-31 WGKVEADIAGHGQEVLI Myoglobin 16-30 GKVEADIAGHGQEVL Myoglobin 19-34 EADIAGHGQEVLIRLF Myoglobin 24-47 GHGQEVLIRLFTGHPETLEKFDKF
118

Myoglobin 25-45 HGQEVLIRLFTGHPETLEKFD Myoglobin 30-69 LIRLFTGHPETLEKFDKFKHLKTEAEMKASEDLKKHGTVV Myoglobin 31-56 IRLFTGHPETLEKFDKFKHLKTEAEM Myoglobin 34-55 FTGHPETLEKFDKFKHLKTEAE Myoglobin 34-70 FTGHPETLEKFDKFKHLKTEAEMKASEDLKKHGTVVL Myoglobin 40-50 TLEKFDKFKHL Myoglobin 42-70 EKFDKFKHLKTEAEMKASEDLKKHGTVVL Myoglobin 45-59 DKFKHLKTEAEMKAS Myoglobin 48-65 KHLKTEAEMKASEDLKKH Myoglobin 49-65 HLKTEAEMKASEDLKKH Myoglobin 49-66 HLKTEAEMKASEDLKKHG Myoglobin 54-74 AEMKASEDLKKHGTVVLTALG Myoglobin 57-66 KASEDLKKHG Myoglobin 57-70 KASEDLKKHGTVVL Myoglobin 71-106 TALGGILKKKGHHEAELKPLAQSHATKHKIPIKYLE Myoglobin 71-107 TALGGILKKKGHHEAELKPLAQSHATKHKIPIKYLEF Myoglobin 73-104 LGGILKKKGHHEAELKPLAQSHATKHKIPIKY Myoglobin 73-106 LGGILKKKGHHEAELKPLAQSHATKHKIPIKYLE Myoglobin 74-104 GGILKKKGHHEAELKPLAQSHATKHKIPIKY Myoglobin 84-97 EAELKPLAQSHATK Myoglobin 107-135 FISDAIIHVLHSKHPGDFGADAQGAMTKA Myoglobin 107-138 FISDAIIHVLHSKHPGDFGADAQGAMTKALEL Myoglobin 108-135 ISDAIIHVLHSKHPGDFGADAQGAMTKA Myoglobin 108-137 ISDAIIHVLHSKHPGDFGADAQGAMTKALE Myoglobin 108-138 ISDAIIHVLHSKHPGDFGADAQGAMTKALEL Myoglobin 109-136 SDAIIHVLHSKHPGDFGADAQGAMTKAL Myoglobin 111-135 AIIHVLHSKHPGDFGADAQGAMTKA Myoglobin 111-137 AIIHVLHSKHPGDFGADAQGAMTKALE Myoglobin 111-138 AIIHVLHSKHPGDFGADAQGAMTKALEL Myoglobin 112-135 IIHVLHSKHPGDFGADAQGAMTKA Myoglobin 112-138 IIHVLHSKHPGDFGADAQGAMTKALEL Myoglobin 113-142 IHVLHSKHPGDFGADAQGAMTKALELFRND Myoglobin 115-140 VLHSKHPGDFGADAQGAMTKALELFR Myoglobin 115-145 VLHSKHPGDFGADAQGAMTKALELFRNDIAA Myoglobin 122-146 GDFGADAQGAMTKALELFRNDIAAK Myoglobin 124-145 FGADAQGAMTKALELFRNDIAA Myoglobin 124-146 FGADAQGAMTKALELFRNDIAAK Myoglobin 126-143 ADAQGAMTKALELFRNDI Myoglobin 129-146 QGAMTKALELFRNDIAAK Myoglobin 132-147 MTKALELFRNDIAAKY Myoglobin 139-152 FRNDIAAKYKELGF
Nef 3-30 SSHHHHHHSSGLVPRGSHMGGKWSKSSV Nef 9-21 HHSSGLVPRGSHM Nef 9-32 HHSSGLVPRGSHMGGKWSKSSVIG Nef 17-27 RGSHMGGKWSK Nef 17-32 RGSHMGGKWSKSSVIG Nef 28-52 SSVIGWPAVRERMRRAEPAADGVGA Nef 29-41 SVIGWPAVRERMR Nef 34-48 PAVRERMRRAEPAAD Nef 35-64 AVRERMRRAEPAADGVGAVSRDLEKHGAIT Nef 41-62 RRAEPAADGVGAVSRDLEKHGA Nef 42-62 RAEPAADGVGAVSRDLEKHGA Nef 47-79 ADGVGAVSRDLEKHGAITSSNTAANNAACAWLE Nef 47-81 ADGVGAVSRDLEKHGAITSSNTAANNAACAWLEAQ Nef 49-77 GVGAVSRDLEKHGAITSSNTAANNAACAW Nef 50-72 VGAVSRDLEKHGAITSSNTAANN Nef 56-90 DLEKHGAITSSNTAANNAACAWLEAQEEEEVGFPV Nef 58-89 EKHGAITSSNTAANNAACAWLEAQEEEEVGFP Nef 60-72 HGAITSSNTAANN Nef 60-93 HGAITSSNTAANNAACAWLEAQEEEEVGFPVTPQ Nef 77-97 WLEAQEEEEVGFPVTPQVPLR Nef 82-108 EEEEVGFPVTPQVPLRPMTYKAAVDLS Nef 99-118 MTYKAAVDLSHFLKEKGGLE
119

Nef 106-119 DLSHFLKEKGGLEG Nef 106-120 DLSHFLKEKGGLEGL Nef 108-119 SHFLKEKGGLEG Nef 140-164 YFPDWQNYTPGPGVRYPLTFGWCYK Nef 152-173 GVRYPLTFGWCYKLVPVEPDKV Nef 154-166 RYPLTFGWCYKLV Nef 173-187 VEEANKGENTSLLHP Nef 183-209 SLLHPVSLHGMDDPEREVLEWRFDSRL Nef 201-219 LEWRFDSRLAFHHVARELH Nef 202-219 EWRFDSRLAFHHVARELH Nef 206-220 DSRLAFHHVARELHP
Ubiquitin 1-15 MQIFVKTLTGKTITL Ubiquitin 4-15 FVKTLTGKTITL Ubiquitin 25-45 NVKAKIQDKEGIPPDQQRLIF Ubiquitin 44-58 IFAGKQLEDGRTLSD Ubiquitin 46-58 AGKQLEDGRTLSD Ubiquitin 46-67 AGKQLEDGRTLSDYNIQKESTL Ubiquitin 59-69 YNIQKESTLHL
120

Table 3.4. pH 4.0 peptides Protein Residues Peptide Sequence
Abl 31-41 LAGPSENDPNL Abl 31-42 LAGPSENDPNLF Abl 49-64 VASGDNTLSITKGEKL Abl 65-75 RVLGYNHNGEW Abl 76-90 CEAQTKNGQGWVPSN Abl 78-90 AQTKNGQGWVPSN Abl 93-102 TPVNSLEKHS Abl 93-103 TPVNSLEKHSW Abl 126-141 LVRESESSPGQRSISL Abl 127-141 VRESESSPGQRSISL Abl 150-160 YRINTASDGKL Abl 172-185 AELVHHHSTVADGL Abl 186-202 ITTLHYPAPKRNKPTVY Abl 187-202 TTLHYPAPKRNKPTVY Abl 198-210 KPTVYGVSPNYDK Abl 203-213 GVSPNYDKWEM Abl 378-387 PIKWTAPESL Abl 482-491 VEKELGKENL
Albumin 1-21 DTHKSEIAHRFKDLGEEHFKG Albumin 28-58 SQYLQQCPFDEHVKLVNELTEFAKTCVADES Albumin 32-79 QQCPFDEHVKLVNELTEFAKTCVADESHAGCEKSLHTLFGDELCKVAS Albumin 71-78 GDELCKVA Albumin 77-105 VASLRETYGDMADCCEKQEPERNECFLSH Albumin 103-137 LSHKDDSPDLPKLKPDPNTLCDEFKADEKKFWGKY Albumin 113-133 PKLKPDPNTLCDEFKADEKKF Albumin 130-159 EKKFWGKYLYEIARRHPYFYAPELLYYANK Albumin 146-158 PYFYAPELLYYAN Albumin 154-164 LYYANKYNGVF Albumin 179-192 PKIETMREKVLASS Albumin 189-202 LASSARQRLRCASI Albumin 231-262 TKLVTDLTKVHKECCHGDLLECADDRADLAKY Albumin 235-271 TDLTKVHKECCHGDLLECADDRADLAKYICDNQDTIS Albumin 240-277 VHKECCHGDLLECADDRADLAKYICDNQDTISSKLKEC Albumin 241-277 HKECCHGDLLECADDRADLAKYICDNQDTISSKLKEC Albumin 253-271 ADDRADLAKYICDNQDTIS Albumin 261-289 KYICDNQDTISSKLKECCDKPLLEKSHCI Albumin 269-290 TISSKLKECCDKPLLEKSHCIA Albumin 278-287 CDKPLLEKSH Albumin 287-327 HCIAEVEKDAIPENLPPLTADFAEDKDVCKNYQEAKDAFLG Albumin 291-306 EVEKDAIPENLPPLTA Albumin 292-306 VEKDAIPENLPPLTA Albumin 292-307 VEKDAIPENLPPLTAD Albumin 292-308 VEKDAIPENLPPLTADF Albumin 308-323 FAEDKDVCKNYQEAKD Albumin 373-386 FDKLKHLVDEPQNL Albumin 374-386 DKLKHLVDEPQNL Albumin 378-390 HLVDEPQNLIKQN Albumin 381-396 DEPQNLIKQNCDQFEK Albumin 384-399 QNLIKQNCDQFEKLGE Albumin 395-406 EKLGEYGFQNAL Albumin 407-419 IVRYTRKVPQVST Albumin 466-483 TPVSEKVTKCCTESLVNR Albumin 485-512 PCFSALTPDETYVPKAFDEKLFTFHADI Albumin 488-501 SALTPDETYVPKAF Albumin 493-528 DETYVPKAFDEKLFTFHADICTLPDTEKQIKKQTAL Albumin 496-514 YVPKAFDEKLFTFHADICT Albumin 506-521 FTFHADICTLPDTEKQ Albumin 516-528 PDTEKQIKKQTAL Albumin 529-542 VELLKHKPKATEEQ Albumin 542-555 QLKTVMENFVAFVD
121

Albumin 544-560 KTVMENFVAFVDKCCAA Albumin 545-572 TVMENFVAFVDKCCAADDKEACFAVEGP Aldolase 2-18 PHSHPALTPEQKKELSD Aldolase 2-21 PHSHPALTPEQKKELSDIAH Aldolase 2-25 PHSHPALTPEQKKELSDIAHRIVA Aldolase 2-31 PHSHPALTPEQKKELSDIAHRIVAPGKGIL Aldolase 4-11 SHPALTPE Aldolase 10-34 PEQKKELSDIAHRIVAPGKGILAAD Aldolase 32-44 AADESTGSIAKRL Aldolase 32-46 AADESTGSIAKRLQS Aldolase 32-50 AADESTGSIAKRLQSIGTE Aldolase 32-54 AADESTGSIAKRLQSIGTENTEE Aldolase 33-50 ADESTGSIAKRLQSIGTE Aldolase 37-53 TGSIAKRLQSIGTENTE Aldolase 60-79 RQLLLTADDRVNPCIGGVIL Aldolase 62-82 LLLTADDRVNPCIGGVILFHE Aldolase 64-76 LTADDRVNPCIGG Aldolase 71-93 NPCIGGVILFHETLYQKADDGRP Aldolase 85-97 YQKADDGRPFPQV Aldolase 85-106 YQKADDGRPFPQVIKSKGGVVG Aldolase 107-121 IKVDKGVVPLAGTNG Aldolase 107-122 IKVDKGVVPLAGTNGE Aldolase 107-128 IKVDKGVVPLAGTNGETTTQGL Aldolase 107-130 IKVDKGVVPLAGTNGETTTQGLDG Aldolase 107-131 IKVDKGVVPLAGTNGETTTQGLDGL Aldolase 107-132 IKVDKGVVPLAGTNGETTTQGLDGLS Aldolase 107-133 IKVDKGVVPLAGTNGETTTQGLDGLSE Aldolase 109-134 VDKGVVPLAGTNGETTTQGLDGLSER Aldolase 142-164 GADFAKWRCVLKIGEHTPSALAI Aldolase 175-188 ASICQQNGIVPIVE Aldolase 180-188 QNGIVPIVE Aldolase 183-204 IVPIVEPEILPDGDHDLKRCQY Aldolase 189-203 PEILPDGDHDLKRCQ Aldolase 189-204 PEILPDGDHDLKRCQY Aldolase 191-202 ILPDGDHDLKRC Aldolase 193-203 PDGDHDLKRCQ Aldolase 193-204 PDGDHDLKRCQY Aldolase 203-221 QYVTEKVLAAVYKALSDHH Aldolase 212-224 AVYKALSDHHIYL Aldolase 213-224 VYKALSDHHIYL Aldolase 215-224 KALSDHHIYL Aldolase 225-234 EGTLLKPNMV Aldolase 241-253 TQKYSHEEIAMAT Aldolase 257-270 LRRTVPPAVTGVTF Aldolase 258-270 RRTVPPAVTGVTF Aldolase 285-299 NAINKCPLLKPWALT Aldolase 287-299 INKCPLLKPWALT Aldolase 301-310 SYGRALQASA Aldolase 305-325 ALQASALKAWGGKKENLKAAQ Aldolase 309-321 SALKAWGGKKENL Aldolase 320-334 NLKAAQEEYVKRALA Aldolase 320-342 NLKAAQEEYVKRALANSLACQGK Aldolase 327-343 EYVKRALANSLACQGKY Aldolase 327-348 EYVKRALANSLACQGKYTPSGQ Aldolase 329-357 VKRALANSLACQGKYTPSGQAGAAASESL Aldolase 337-357 LACQGKYTPSGQAGAAASESL Aldolase 338-357 ACQGKYTPSGQAGAAASESL
Hemoglogin-alpha 2-24 VLSAADKGNVKAAWGKVGGHAAE Hemoglogin-alpha 3-21 LSAADKGNVKAAWGKVGGH Hemoglogin-alpha 3-22 LSAADKGNVKAAWGKVGGHA Hemoglogin-alpha 3-24 LSAADKGNVKAAWGKVGGHAAE Hemoglogin-alpha 4-24 SAADKGNVKAAWGKVGGHAAE Hemoglogin-alpha 27-36 AEALERMFLS
122

Hemoglogin-alpha 34-47 FLSFPTTKTYFPHF Hemoglogin-alpha 34-49 FLSFPTTKTYFPHFDL Hemoglogin-alpha 35-47 LSFPTTKTYFPHF Hemoglogin-alpha 35-54 LSFPTTKTYFPHFDLSHGSA Hemoglogin-alpha 36-47 SFPTTKTYFPHF Hemoglogin-alpha 45-68 PHFDLSHGSAQVKGHGAKVAAALT Hemoglogin-alpha 48-67 DLSHGSAQVKGHGAKVAAAL Hemoglogin-alpha 52-61 GSAQVKGHGA Hemoglogin-alpha 53-72 SAQVKGHGAKVAAALTKAVE Hemoglogin-alpha 62-79 KVAAALTKAVEHLDDLPG Hemoglogin-alpha 62-82 KVAAALTKAVEHLDDLPGALS Hemoglogin-alpha 66-81 ALTKAVEHLDDLPGAL Hemoglogin-alpha 67-81 LTKAVEHLDDLPGAL Hemoglogin-alpha 68-83 TKAVEHLDDLPGALSE Hemoglogin-alpha 71-81 VEHLDDLPGAL Hemoglogin-alpha 71-83 VEHLDDLPGALSE Hemoglogin-alpha 73-87 HLDDLPGALSELSDL Hemoglogin-alpha 80-95 ALSELSDLHAHKLRVD Hemoglogin-alpha 81-95 LSELSDLHAHKLRVD Hemoglogin-alpha 82-94 SELSDLHAHKLRV Hemoglogin-alpha 85-99 SDLHAHKLRVDPVNF Hemoglogin-alpha 88-99 HAHKLRVDPVNF Hemoglogin-alpha 94-105 VDPVNFKLLSHS Hemoglogin-alpha 94-106 VDPVNFKLLSHSL Hemoglogin-alpha 108-123 VTLASHLPSDFTPAVH Hemoglogin-alpha 109-124 TLASHLPSDFTPAVHA Hemoglogin-alpha 110-124 LASHLPSDFTPAVHA Hemoglogin-alpha 111-123 ASHLPSDFTPAVH Hemoglogin-alpha 111-124 ASHLPSDFTPAVHA Hemoglogin-alpha 111-126 ASHLPSDFTPAVHASL Hemoglogin-alpha 117-131 DFTPAVHASLDKFLA Hemoglogin-alpha 120-140 PAVHASLDKFLANVSTVLTSK Hemoglogin-beta 5-22 EEKAAVTAFWGKVKVDEV Hemoglogin-beta 6-22 EKAAVTAFWGKVKVDEV Hemoglogin-beta 14-27 WGKVKVDEVGGEAL Hemoglogin-beta 14-30 WGKVKVDEVGGEALGRL Hemoglogin-beta 48-69 STADAVMNNPKVKAHGKKVLDS Hemoglogin-beta 52-69 AVMNNPKVKAHGKKVLDS Hemoglogin-beta 52-73 AVMNNPKVKAHGKKVLDSFSNG Hemoglogin-beta 53-69 VMNNPKVKAHGKKVLDS Hemoglogin-beta 54-69 MNNPKVKAHGKKVLDS Hemoglogin-beta 55-69 NNPKVKAHGKKVLDS Hemoglogin-beta 65-77 KVLDSFSNGMKHL Hemoglogin-beta 68-84 DSFSNGMKHLDDLKGTF Hemoglogin-beta 70-84 FSNGMKHLDDLKGTF Hemoglogin-beta 79-89 DLKGTFAALSE Hemoglogin-beta 86-97 ALSELHCDKLHV Hemoglogin-beta 89-108 ELHCDKLHVDPENFKLLGNV Hemoglogin-beta 114-125 ARNFGKEFTPVL Hemoglogin-beta 118-133 GKEFTPVLQADFQKVV Hemoglogin-beta 118-138 GKEFTPVLQADFQKVVAGVAN Hemoglogin-beta 123-131 PVLQADFQK Hemoglogin-beta 125-140 LQADFQKVVAGVANAL Hemoglogin-beta 126-139 QADFQKVVAGVANA Hemoglogin-beta 128-140 DFQKVVAGVANAL Hemoglogin-beta 131-144 KVVAGVANALAHRY
Hemoglogin-epsilon 2 10-26 VAVASLWAKVNVEVVGG Hemoglogin-epsilon 2 22-36 EVVGGESLARLLIVC Hemoglogin-epsilon 2 58-71 NPKVKVYGRKVLNS Hemoglogin-epsilon 2 109-122 NMILIVLATHFSKE Hemoglogin-epsilon 2 125-142 PQMQAAWQKLTNAVANAL Hemoglogin-epsilon 4 25-42 GGESLARLLIVYPWTQRF Hemoglogin-epsilon 4 27-40 ESLARLLIVYPWTQ
123

Hemoglogin-epsilon 4 67-107 KVLNSFGNAIEHMDDLKGTFADLSELHCDKLHVDPENFRLL Hemoglogin-epsilon 4 99-131 VDPENFRLLGNMILIVLATHFSKEFTPQMQASW Hemoglogin-epsilon 4 101-113 PENFRLLGNMILI Hemoglogin-epsilon 4 107-123 LGNMILIVLATHFSKEF Hemoglogin-epsilon 4 109-122 NMILIVLATHFSKE Hemoglogin-epsilon 4 124-144 TPQMQASWQKLTNAVANALAH Hemoglogin-epsilon 4 137-145 AVANALAHK
Myoglobin 2-12 GLSDGEWQQVL Myoglobin 14-37 VWGKVEADIAGHGQEVLIRLFTGH Myoglobin 15-30 WGKVEADIAGHGQEVL Myoglobin 16-30 GKVEADIAGHGQEVL Myoglobin 26-36 GQEVLIRLFTG Myoglobin 30-41 LIRLFTGHPETL Myoglobin 41-52 LEKFDKFKHLKT Myoglobin 41-55 LEKFDKFKHLKTEAE Myoglobin 42-56 EKFDKFKHLKTEAEM Myoglobin 49-65 HLKTEAEMKASEDLKKH Myoglobin 58-72 ASEDLKKHGTVVLTA Myoglobin 69-84 VLTALGGILKKKGHHE Myoglobin 71-87 TALGGILKKKGHHEAEL Myoglobin 73-87 LGGILKKKGHHEAEL Myoglobin 85-111 AELKPLAQSHATKHKIPIKYLEFISDA Myoglobin 88-104 KPLAQSHATKHKIPIKY Myoglobin 92-102 QSHATKHKIPI Myoglobin 98-110 HKIPIKYLEFISD Myoglobin 105-117 LEFISDAIIHVLH Myoglobin 108-127 ISDAIIHVLHSKHPGDFGAD Myoglobin 109-124 SDAIIHVLHSKHPGDF Myoglobin 109-136 SDAIIHVLHSKHPGDFGADAQGAMTKAL Myoglobin 111-132 AIIHVLHSKHPGDFGADAQGAM Myoglobin 112-127 IIHVLHSKHPGDFGAD Myoglobin 112-130 IIHVLHSKHPGDFGADAQG Myoglobin 112-135 IIHVLHSKHPGDFGADAQGAMTKA Myoglobin 113-121 IHVLHSKHP
Nef 4-38 SHHHHHHSSGLVPRGSHMGGKWSKSSVIGWPAVRE Nef 11-32 SSGLVPRGSHMGGKWSKSSVIG Nef 35-53 AVRERMRRAEPAADGVGAV Nef 52-68 AVSRDLEKHGAITSSNT Nef 60-72 HGAITSSNTAANN Nef 85-100 EVGFPVTPQVPLRPMT Nef 98-115 PMTYKAAVDLSHFLKEKG Nef 100-112 TYKAAVDLSHFLK Nef 154-167 RYPLTFGWCYKLVP Nef 189-202 SLHGMDDPEREVLE Nef 207-224 SRLAFHHVARELHPEYFK
Ubiquitin 1-15 MQIFVKTLTGKTITL Ubiquitin 16-45 EVEPSDTIENVKAKIQDKEGIPPDQQRLIF Ubiquitin 25-45 NVKAKIQDKEGIPPDQQRLIF Ubiquitin 44-58 IFAGKQLEDGRTLSD
124

Table 3.5. pH 2.5 E.coli peptides Protein Peptide Sequence
ACP_ECOLI (P02901) AAIDYINGHQA ACP_ECOLI (P02901) AIDYINGHQA ACP_ECOLI (P02901) ALEEEF ACP_ECOLI (P02901) DTEIPDEEA ACP_ECOLI (P02901) DTEIPDEEAEKITTVQ ACP_ECOLI (P02901) FDTEIPDEE ACP_ECOLI (P02901) FDTEIPDEEA ACP_ECOLI (P02901) GVKQEEVTNNAS ACP_ECOLI (P02901) GVKQEEVTNNASF ACP_ECOLI (P02901) LGADSLDTVE ACP_ECOLI (P02901) RVKKIIGEQL ACP_ECOLI (P02901) SFVEDLGADSLDTVELVM
AHPC_ECOLI (P26427) AEGIGRDASDL AHPC_ECOLI (P26427) EGRWSVFFFYPADFTFVCPTELGDV AHPC_ECOLI (P26427) FKNGEF AHPC_ECOLI (P26427) GDPTGALTRNFDNMR AHPC_ECOLI (P26427) GDVADHYEEL AHPC_ECOLI (P26427) GEVCPAKWKEGE AHPC_ECOLI (P26427) GLADRATFVVDPQGIIQ AHPC_ECOLI (P26427) IEITEKDTEGRW AHPC_ECOLI (P26427) IEVTAEGIGRDASDL AHPC_ECOLI (P26427) IGDPTGAL AHPC_ECOLI (P26427) IGDPTGALTRN AHPC_ECOLI (P26427) IGDPTGALTRNFDNM AHPC_ECOLI (P26427) IGRDASDLLR AHPC_ECOLI (P26427) ITEKDTEGRW AHPC_ECOLI (P26427) QYVASHPGEVCPAKWKE AHPC_ECOLI (P26427) REDEGLADRATF AHPC_ECOLI (P26427) TAEGIGRDASDL AHPC_ECOLI (P26427) TAEGIGRDASDLL AHPC_ECOLI (P26427) VTAEGIGRDASDL ALKH_ECOLI (P10177) AISPGLTEPLL ALKH_ECOLI (P10177) AIVGAGTVLNPQQL ALKH_ECOLI (P10177) FAISPGLTEPLL ALKH_ECOLI (P10177) FKFFPAEANGGVKALQA ALKH_ECOLI (P10177) GTVLNPQQLAEVTEAGAQ ALKH_ECOLI (P10177) IAKEVPEAIVGAGTVLNPQQLAE ALKH_ECOLI (P10177) KAATEGTIPLIPGIST ALKH_ECOLI (P10177) LVAGGVRVLE ALKH_ECOLI (P10177) PFSQVRFCPTGGISP ALKH_ECOLI (P10177) PQQLAEVTEAGAQFAI ALKH_ECOLI (P10177) PQQLAEVTEAGAQFAISPGL ALKH_ECOLI (P10177) QLAEVTEAGAQFAISPGLT ALKH_ECOLI (P10177) SVLCIGGSWLVPADALEA ALKH_ECOLI (P10177) TTGPVVPVIV ALKH_ECOLI (P10177) YLALKSVLCIGGS ARTI_ECOLI (P30859) AGMDITPEREKQVL ARTI_ECOLI (P30859) DLQNGRIDGVF ARTI_ECOLI (P30859) DLQNGRIDGVFGDTA ARTI_ECOLI (P30859) IAVRQGNTEL ARTI_ECOLI (P30859) IMDKHPEITTVPYDSYQNAKL ARTI_ECOLI (P30859) NAKLDLQNG ARTI_ECOLI (P30859) QALCKEIDATC ARTI_ECOLI (P30859) QNGRIDGVFGDTA ARTI_ECOLI (P30859) SIDANNQIVGFDVDLAQ CH10_ECOLI (P05380) AAKSTRGEVL CH10_ECOLI (P05380) AVGNGRILENG CH10_ECOLI (P05380) AVGNGRILENGEVKPLD CH10_ECOLI (P05380) AVGNGRILENGEVKPLDV CH10_ECOLI (P05380) AVGNGRILENGEVKPLDVKVGD
125
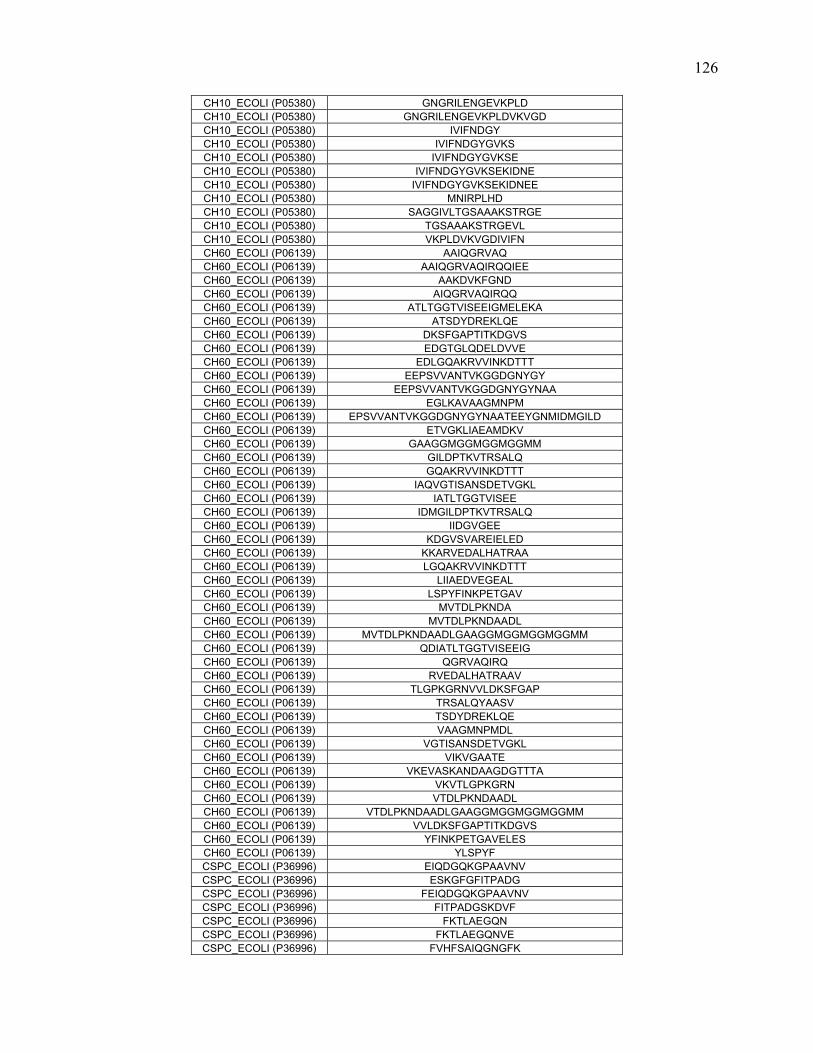
CH10_ECOLI (P05380) GNGRILENGEVKPLD CH10_ECOLI (P05380) GNGRILENGEVKPLDVKVGD CH10_ECOLI (P05380) IVIFNDGY CH10_ECOLI (P05380) IVIFNDGYGVKS CH10_ECOLI (P05380) IVIFNDGYGVKSE CH10_ECOLI (P05380) IVIFNDGYGVKSEKIDNE CH10_ECOLI (P05380) IVIFNDGYGVKSEKIDNEE CH10_ECOLI (P05380) MNIRPLHD CH10_ECOLI (P05380) SAGGIVLTGSAAAKSTRGE CH10_ECOLI (P05380) TGSAAAKSTRGEVL CH10_ECOLI (P05380) VKPLDVKVGDIVIFN CH60_ECOLI (P06139) AAIQGRVAQ CH60_ECOLI (P06139) AAIQGRVAQIRQQIEE CH60_ECOLI (P06139) AAKDVKFGND CH60_ECOLI (P06139) AIQGRVAQIRQQ CH60_ECOLI (P06139) ATLTGGTVISEEIGMELEKA CH60_ECOLI (P06139) ATSDYDREKLQE CH60_ECOLI (P06139) DKSFGAPTITKDGVS CH60_ECOLI (P06139) EDGTGLQDELDVVE CH60_ECOLI (P06139) EDLGQAKRVVINKDTTT CH60_ECOLI (P06139) EEPSVVANTVKGGDGNYGY CH60_ECOLI (P06139) EEPSVVANTVKGGDGNYGYNAA CH60_ECOLI (P06139) EGLKAVAAGMNPM CH60_ECOLI (P06139) EPSVVANTVKGGDGNYGYNAATEEYGNMIDMGILD CH60_ECOLI (P06139) ETVGKLIAEAMDKV CH60_ECOLI (P06139) GAAGGMGGMGGMGGMM CH60_ECOLI (P06139) GILDPTKVTRSALQ CH60_ECOLI (P06139) GQAKRVVINKDTTT CH60_ECOLI (P06139) IAQVGTISANSDETVGKL CH60_ECOLI (P06139) IATLTGGTVISEE CH60_ECOLI (P06139) IDMGILDPTKVTRSALQ CH60_ECOLI (P06139) IIDGVGEE CH60_ECOLI (P06139) KDGVSVAREIELED CH60_ECOLI (P06139) KKARVEDALHATRAA CH60_ECOLI (P06139) LGQAKRVVINKDTTT CH60_ECOLI (P06139) LIIAEDVEGEAL CH60_ECOLI (P06139) LSPYFINKPETGAV CH60_ECOLI (P06139) MVTDLPKNDA CH60_ECOLI (P06139) MVTDLPKNDAADL CH60_ECOLI (P06139) MVTDLPKNDAADLGAAGGMGGMGGMGGMM CH60_ECOLI (P06139) QDIATLTGGTVISEEIG CH60_ECOLI (P06139) QGRVAQIRQ CH60_ECOLI (P06139) RVEDALHATRAAV CH60_ECOLI (P06139) TLGPKGRNVVLDKSFGAP CH60_ECOLI (P06139) TRSALQYAASV CH60_ECOLI (P06139) TSDYDREKLQE CH60_ECOLI (P06139) VAAGMNPMDL CH60_ECOLI (P06139) VGTISANSDETVGKL CH60_ECOLI (P06139) VIKVGAATE CH60_ECOLI (P06139) VKEVASKANDAAGDGTTTA CH60_ECOLI (P06139) VKVTLGPKGRN CH60_ECOLI (P06139) VTDLPKNDAADL CH60_ECOLI (P06139) VTDLPKNDAADLGAAGGMGGMGGMGGMM CH60_ECOLI (P06139) VVLDKSFGAPTITKDGVS CH60_ECOLI (P06139) YFINKPETGAVELES CH60_ECOLI (P06139) YLSPYF CSPC_ECOLI (P36996) EIQDGQKGPAAVNV CSPC_ECOLI (P36996) ESKGFGFITPADG CSPC_ECOLI (P36996) FEIQDGQKGPAAVNV CSPC_ECOLI (P36996) FITPADGSKDVF CSPC_ECOLI (P36996) FKTLAEGQN CSPC_ECOLI (P36996) FKTLAEGQNVE CSPC_ECOLI (P36996) FVHFSAIQGNGFK
126
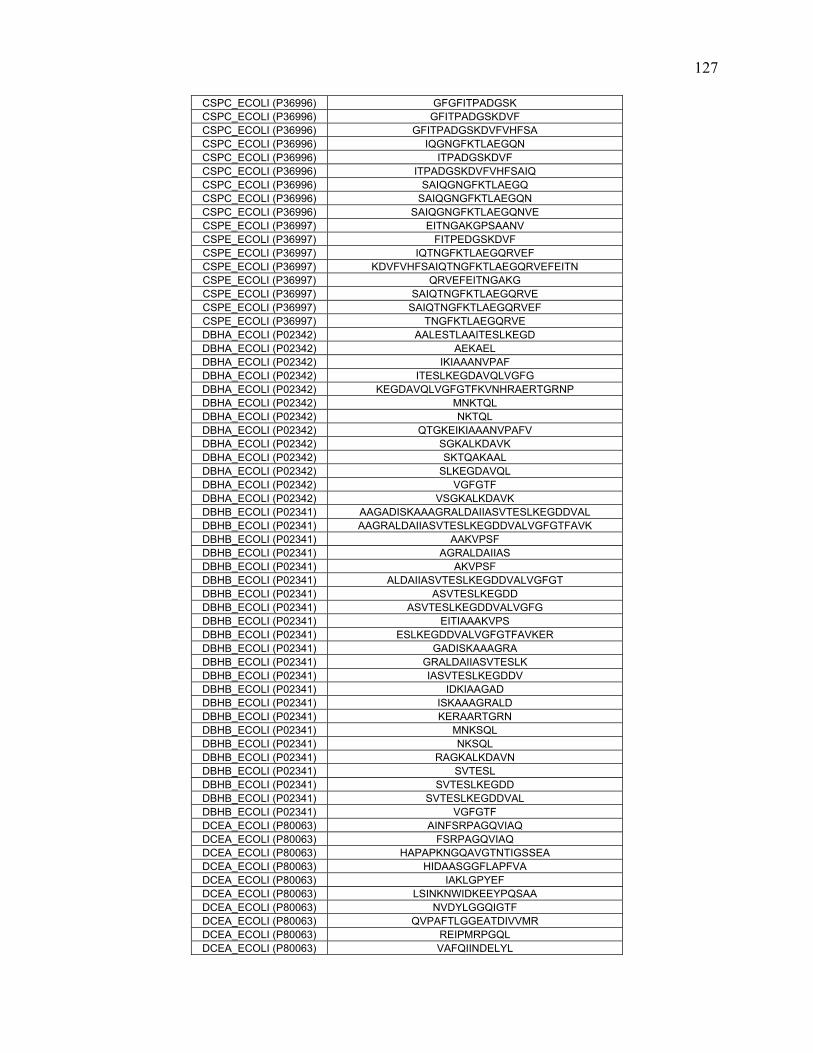
CSPC_ECOLI (P36996) GFGFITPADGSK CSPC_ECOLI (P36996) GFITPADGSKDVF CSPC_ECOLI (P36996) GFITPADGSKDVFVHFSA CSPC_ECOLI (P36996) IQGNGFKTLAEGQN CSPC_ECOLI (P36996) ITPADGSKDVF CSPC_ECOLI (P36996) ITPADGSKDVFVHFSAIQ CSPC_ECOLI (P36996) SAIQGNGFKTLAEGQ CSPC_ECOLI (P36996) SAIQGNGFKTLAEGQN CSPC_ECOLI (P36996) SAIQGNGFKTLAEGQNVE CSPE_ECOLI (P36997) EITNGAKGPSAANV CSPE_ECOLI (P36997) FITPEDGSKDVF CSPE_ECOLI (P36997) IQTNGFKTLAEGQRVEF CSPE_ECOLI (P36997) KDVFVHFSAIQTNGFKTLAEGQRVEFEITN CSPE_ECOLI (P36997) QRVEFEITNGAKG CSPE_ECOLI (P36997) SAIQTNGFKTLAEGQRVE CSPE_ECOLI (P36997) SAIQTNGFKTLAEGQRVEF CSPE_ECOLI (P36997) TNGFKTLAEGQRVE DBHA_ECOLI (P02342) AALESTLAAITESLKEGD DBHA_ECOLI (P02342) AEKAEL DBHA_ECOLI (P02342) IKIAAANVPAF DBHA_ECOLI (P02342) ITESLKEGDAVQLVGFG DBHA_ECOLI (P02342) KEGDAVQLVGFGTFKVNHRAERTGRNP DBHA_ECOLI (P02342) MNKTQL DBHA_ECOLI (P02342) NKTQL DBHA_ECOLI (P02342) QTGKEIKIAAANVPAFV DBHA_ECOLI (P02342) SGKALKDAVK DBHA_ECOLI (P02342) SKTQAKAAL DBHA_ECOLI (P02342) SLKEGDAVQL DBHA_ECOLI (P02342) VGFGTF DBHA_ECOLI (P02342) VSGKALKDAVK DBHB_ECOLI (P02341) AAGADISKAAAGRALDAIIASVTESLKEGDDVAL DBHB_ECOLI (P02341) AAGRALDAIIASVTESLKEGDDVALVGFGTFAVK DBHB_ECOLI (P02341) AAKVPSF DBHB_ECOLI (P02341) AGRALDAIIAS DBHB_ECOLI (P02341) AKVPSF DBHB_ECOLI (P02341) ALDAIIASVTESLKEGDDVALVGFGT DBHB_ECOLI (P02341) ASVTESLKEGDD DBHB_ECOLI (P02341) ASVTESLKEGDDVALVGFG DBHB_ECOLI (P02341) EITIAAAKVPS DBHB_ECOLI (P02341) ESLKEGDDVALVGFGTFAVKER DBHB_ECOLI (P02341) GADISKAAAGRA DBHB_ECOLI (P02341) GRALDAIIASVTESLK DBHB_ECOLI (P02341) IASVTESLKEGDDV DBHB_ECOLI (P02341) IDKIAAGAD DBHB_ECOLI (P02341) ISKAAAGRALD DBHB_ECOLI (P02341) KERAARTGRN DBHB_ECOLI (P02341) MNKSQL DBHB_ECOLI (P02341) NKSQL DBHB_ECOLI (P02341) RAGKALKDAVN DBHB_ECOLI (P02341) SVTESL DBHB_ECOLI (P02341) SVTESLKEGDD DBHB_ECOLI (P02341) SVTESLKEGDDVAL DBHB_ECOLI (P02341) VGFGTF DCEA_ECOLI (P80063) AINFSRPAGQVIAQ DCEA_ECOLI (P80063) FSRPAGQVIAQ DCEA_ECOLI (P80063) HAPAPKNGQAVGTNTIGSSEA DCEA_ECOLI (P80063) HIDAASGGFLAPFVA DCEA_ECOLI (P80063) IAKLGPYEF DCEA_ECOLI (P80063) LSINKNWIDKEEYPQSAA DCEA_ECOLI (P80063) NVDYLGGQIGTF DCEA_ECOLI (P80063) QVPAFTLGGEATDIVVMR DCEA_ECOLI (P80063) REIPMRPGQL DCEA_ECOLI (P80063) VAFQIINDELYL
127

DCEA_ECOLI (P80063) WHAPAPKNGQAVGTNTIGSSE DCEA_ECOLI (P80063) WVIWRDEEAL DCEA_ECOLI (P80063) YEFPQPLHDAL DCEA_ECOLI (P80063) YLDGNARQNLAT DCEB_ECOLI (P28302) AGKPTDKPN DCEB_ECOLI (P28302) AINFSRPAGQVIAQ DCEB_ECOLI (P28302) APKNGQAVGTNTIG DCEB_ECOLI (P28302) AYLADEIAKLGPYEFI DCEB_ECOLI (P28302) ELLDSRFGAKSIST DCEB_ECOLI (P28302) ELVFNVDYLGGQIGT DCEB_ECOLI (P28302) ELYLDGNARQNLATF DCEB_ECOLI (P28302) FSRPAGQVIAQ DCEB_ECOLI (P28302) GGQIGTFAINFSR DCEB_ECOLI (P28302) GTFAINFSRPAGQV DCEB_ECOLI (P28302) HIDAASGGFLAPFVA DCEB_ECOLI (P28302) IAESKRFPLHE DCEB_ECOLI (P28302) IAKLGPYEF DCEB_ECOLI (P28302) KEEYPQSAAIDLRCVNM DCEB_ECOLI (P28302) KLKDGEDPGYTL DCEB_ECOLI (P28302) LAPFVAPDIVWD DCEB_ECOLI (P28302) LRSELLDSRFGAKSIST DCEB_ECOLI (P28302) LSINKNWIDKEEYPQSAA DCEB_ECOLI (P28302) NTIGVVPTF DCEB_ECOLI (P28302) NVDYLGGQIGTF DCEB_ECOLI (P28302) NVDYLGGQIGTFAINFSRPAGQVIAQYYEFL DCEB_ECOLI (P28302) QVPAFTLGGEATDIVVMR DCEB_ECOLI (P28302) REIPMRPGQL DCEB_ECOLI (P28302) RPAGQVIAQYYEFL DCEB_ECOLI (P28302) RPGQLFMDPKRM DCEB_ECOLI (P28302) RSELLDSRFGAKS DCEB_ECOLI (P28302) SASGHKFGLAPLGCGWVIWRDE DCEB_ECOLI (P28302) TDLRSELLDSRF DCEB_ECOLI (P28302) VADLWHAPAPKNGQA DCEB_ECOLI (P28302) VAFQIINDELYL DCEB_ECOLI (P28302) VDYLGGQIGTFAIN DCEB_ECOLI (P28302) WHAPAPKNGQAVGTNTIGSSE DCEB_ECOLI (P28302) WIDKEEYPQSAAID DCEB_ECOLI (P28302) WQVPAFTLGGEA DCEB_ECOLI (P28302) WVIWRDEEAL DCEB_ECOLI (P28302) YEFPQPLHDAL DCEB_ECOLI (P28302) YLDGNARQNLAT DGAL_ECOLI (P02927) AKNLADGKGAADGTNWKIDNKV DGAL_ECOLI (P02927) ALVKSGALAGTVL DGAL_ECOLI (P02927) AMAMGAVEALKAHN DGAL_ECOLI (P02927) AMAMGAVEALKAHNKSSIPVFGVDA DGAL_ECOLI (P02927) ATFDLAKNLADG DGAL_ECOLI (P02927) DTAQAKDKMDA DGAL_ECOLI (P02927) ESGIIQGDLIAKH DGAL_ECOLI (P02927) FMSVVRKAIEQDA DGAL_ECOLI (P02927) IAKHWAANQGWDL DGAL_ECOLI (P02927) IEKARGQNVPVV DGAL_ECOLI (P02927) IEQDAKAAPDVQLLMNDSQ DGAL_ECOLI (P02927) INLVDPAAAGTVI DGAL_ECOLI (P02927) INLVDPAAAGTVIEKARGQNVPVVFFNKE DGAL_ECOLI (P02927) IQGDLIAKHWA DGAL_ECOLI (P02927) KARGQNVPVVF DGAL_ECOLI (P02927) KATFDLAKNLADGKGAA DGAL_ECOLI (P02927) KELNDKGIKTEQ DGAL_ECOLI (P02927) KGEPGHPDAEARTTY DGAL_ECOLI (P02927) LFGAAAHAADTRIG DGAL_ECOLI (P02927) LMNDSQNDQSKQNDQIDVL DGAL_ECOLI (P02927) LSGPNANKIE DGAL_ECOLI (P02927) LTLSAVMASMLFGAAAHAA
128

DGAL_ECOLI (P02927) NDANNQAKATF DGAL_ECOLI (P02927) TFDLAKNLADGKGAADGT DGAL_ECOLI (P02927) VDPAAAGTVIEKARGQNVPVVFFNKEPSR DGAL_ECOLI (P02927) VFFNKEPSRKALDSYDKA DGAL_ECOLI (P02927) VIEKARGQNVPVVF DGAL_ECOLI (P02927) VIKELNDKGIKTEQ DGAL_ECOLI (P02927) VKSGALAGTVL DGAL_ECOLI (P02927) VRKAIEQDAKAAPDVQL DGAL_ECOLI (P02927) VVIANNDA DGAL_ECOLI (P02927) WDTAQAKDKMDA DGAL_ECOLI (P02927) WLSGPNANKIE DGAL_ECOLI (P02927) YVGTDSKESGIIQGDL DGAL_ECOLI (P02927) YYVGTDSKESGIIQGDL DNAK_ECOLI (P04475) ADNKSLGQFNLDGINPAPRGMPQ DNAK_ECOLI (P04475) AGADASANNAKDDD DNAK_ECOLI (P04475) AGLSVSDIDDVILVGG DNAK_ECOLI (P04475) AIMDGTTPRVLE DNAK_ECOLI (P04475) AIMDGTTPRVLENAEGDRTTPSI DNAK_ECOLI (P04475) AIMDGTTPRVLENAEGDRTTPSII DNAK_ECOLI (P04475) AIMDGTTPRVLENAEGDRTTPSIIA DNAK_ECOLI (P04475) ALAYGLDKGTGNRT DNAK_ECOLI (P04475) AQQTDVNLPYITA DNAK_ECOLI (P04475) AQVSQKLME DNAK_ECOLI (P04475) ATNGDTHLGGED DNAK_ECOLI (P04475) ATNGDTHLGGEDF DNAK_ECOLI (P04475) AVITVPAY DNAK_ECOLI (P04475) AVITVPAYF DNAK_ECOLI (P04475) AYGLDKGTGNRTIA DNAK_ECOLI (P04475) AYGLDKGTGNRTIAV DNAK_ECOLI (P04475) DDKTAIESALTALETALKG DNAK_ECOLI (P04475) DDVILVGGQTRMPMVQ DNAK_ECOLI (P04475) DGINPAPRGMPQIE DNAK_ECOLI (P04475) DGTTPRVLENAEGDRTTPSIIA DNAK_ECOLI (P04475) DIDADGIL DNAK_ECOLI (P04475) DKGTGNRTIAVY DNAK_ECOLI (P04475) DLVNRSIEPLKVAL DNAK_ECOLI (P04475) EAVAIGAAVQGGVLTGDVKD DNAK_ECOLI (P04475) ELVQTRNQGDHLLHSTRKQVEEA DNAK_ECOLI (P04475) EPTAAALAYGLDKGT DNAK_ECOLI (P04475) EPTAAALAYGLDKGTGN DNAK_ECOLI (P04475) EVKRIINEPTAAAL DNAK_ECOLI (P04475) FGKEPRKDVNPDEA DNAK_ECOLI (P04475) FNDAQRQATKDAGRIAGL DNAK_ECOLI (P04475) GDHLLHSTRKQVEE DNAK_ECOLI (P04475) GGQTRMPMVQKKVAEF DNAK_ECOLI (P04475) GIETMGGVMTTLIAKNTT DNAK_ECOLI (P04475) GKEPRKDVNPDEAVAIGAAVQGGVLTGDV DNAK_ECOLI (P04475) GQFNLDGINPAPRGM DNAK_ECOLI (P04475) IAKNTTIPTKHSQV DNAK_ECOLI (P04475) IAKNTTIPTKHSQVF DNAK_ECOLI (P04475) IAKNTTIPTKHSQVFSTAEDNQSAVT DNAK_ECOLI (P04475) IAQQQHAQQQTAGADASANNAKDDDVVD DNAK_ECOLI (P04475) IAQQQHAQQQTAGADASANNAKDDDVVDAE DNAK_ECOLI (P04475) IAVYDLGGGTFDISIIE DNAK_ECOLI (P04475) IDEVDGEKT DNAK_ECOLI (P04475) IDEVDGEKTFE DNAK_ECOLI (P04475) IMDGTTPRVLE DNAK_ECOLI (P04475) IMDGTTPRVLENAEGDRTTPSIIA DNAK_ECOLI (P04475) INPAPRGMPQIEVTFDI DNAK_ECOLI (P04475) KASSGLNEDEIQKMV DNAK_ECOLI (P04475) KQVEEAGDKLPADDKT DNAK_ECOLI (P04475) KTAIESALTALETALK DNAK_ECOLI (P04475) LAQVSQKLME
129

DNAK_ECOLI (P04475) LDVTPLSL DNAK_ECOLI (P04475) LDVTPLSLGIETMGG DNAK_ECOLI (P04475) LLDVTPLSL DNAK_ECOLI (P04475) LVNRSIEPLKVAL DNAK_ECOLI (P04475) MEIAQQQHAQQQTAGADASANNAKDDDVVD DNAK_ECOLI (P04475) MEIAQQQHAQQQTAGADASANNAKDDDVVDAE DNAK_ECOLI (P04475) NAEADRKFEEL DNAK_ECOLI (P04475) NAEGDRTTPSIIA DNAK_ECOLI (P04475) NEPTAAALAYGLDK DNAK_ECOLI (P04475) NGDAWVEVKGQKM DNAK_ECOLI (P04475) NLDGINPAPRGMPQI DNAK_ECOLI (P04475) NLDGINPAPRGMPQIE DNAK_ECOLI (P04475) NLDGINPAPRGMPQIEVT DNAK_ECOLI (P04475) NPAPRGMPQIEVTF DNAK_ECOLI (P04475) PRVLENAEGDRTTPSIIA DNAK_ECOLI (P04475) PYITA DNAK_ECOLI (P04475) PYITADATGPKHMN DNAK_ECOLI (P04475) QGGVLTGDVKDV DNAK_ECOLI (P04475) RAADNKSLGQFNL DNAK_ECOLI (P04475) RKQVEEAGDKLPADDK DNAK_ECOLI (P04475) RKQVEEAGDKLPADDKT DNAK_ECOLI (P04475) RRFQDEEVQRD DNAK_ECOLI (P04475) RSIEPLKVAL DNAK_ECOLI (P04475) SLGIETMGGVMTT DNAK_ECOLI (P04475) SSAQQTDVNL DNAK_ECOLI (P04475) SSAQQTDVNLPYITA DNAK_ECOLI (P04475) SVSDIDD DNAK_ECOLI (P04475) TGPKHMNIKVTRAKL DNAK_ECOLI (P04475) VAIMDGTTPRVLENAEGDRTTPSIIA DNAK_ECOLI (P04475) VEVKGQKMAPPQISA DNAK_ECOLI (P04475) VGGQTRMPMVQKKVAEF DNAK_ECOLI (P04475) VGQPAKRQAVTNPQNTL DNAK_ECOLI (P04475) VGQPAKRQAVTNPQNTLF DNAK_ECOLI (P04475) VLATNGDTHLGGED DNAK_ECOLI (P04475) VLATNGDTHLGGEDF DNAK_ECOLI (P04475) VNPDEAVAIGAAVQGG DNAK_ECOLI (P04475) VNRSIEPLKVAL DNAK_ECOLI (P04475) VQRDVSIMPFKIIA DNAK_ECOLI (P04475) VSAKDKNSGKEQK DNAK_ECOLI (P04475) VSIMPFKIIA DNAK_ECOLI (P04475) VSIMPFKIIAADNGDA DNAK_ECOLI (P04475) WVEVKGQKMAPPQISA DNAK_ECOLI (P04475) WVEVKGQKMAPPQISAE DNAK_ECOLI (P04475) YDLGGGTF DNAK_ECOLI (P04475) YDLGGGTFDISIIEIDEVD EFTS_ECOLI (P02997) ADKVLDAAVAGKITDVEVLKAQFE EFTS_ECOLI (P02997) AGFQAFADKVL EFTS_ECOLI (P02997) AMQSGKPKEIAEKMVEG EFTS_ECOLI (P02997) CKKALTEANGDIEL EFTS_ECOLI (P02997) EGDVLGSYQHGARIGVL EFTS_ECOLI (P02997) HVAASKPEF EFTS_ECOLI (P02997) ILEVNCQTDFVAKDA EFTS_ECOLI (P02997) RFEVGEGIEKVET EFTS_ECOLI (P02997) TGQPFVMEPSKTVGQL EFTS_ECOLI (P02997) VSLTGQPFVMEPSKTVGQL EFTU_ECOLI (P02990) AAQMDGAILVVAATDGPMPQTREHIL EFTU_ECOLI (P02990) AIREGGRTVGAGVVA EFTU_ECOLI (P02990) AKTYGGAARAF EFTU_ECOLI (P02990) DQIDNAPEEKARG EFTU_ECOLI (P02990) DYVKNMITGAAQMDGAIL EFTU_ECOLI (P02990) EKFERTKPHVNVG EFTU_ECOLI (P02990) EKFERTKPHVNVGTIGH EFTU_ECOLI (P02990) FDQIDNAPEEKARG
130
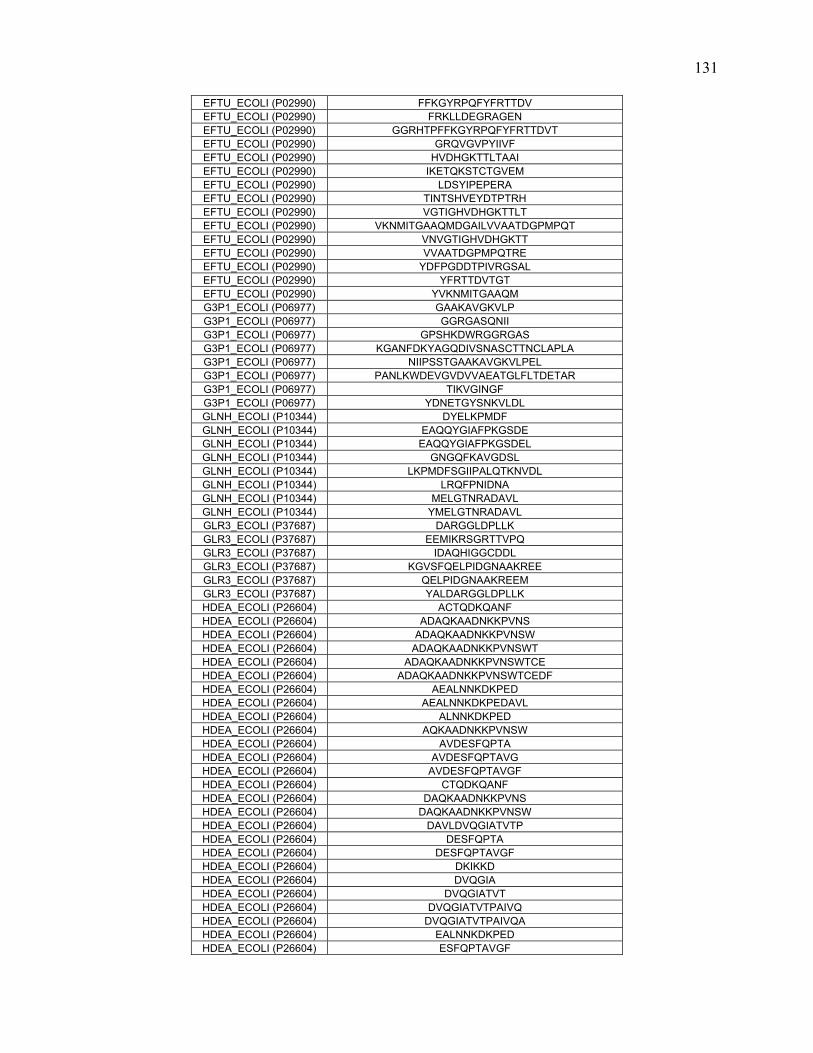
EFTU_ECOLI (P02990) FFKGYRPQFYFRTTDV EFTU_ECOLI (P02990) FRKLLDEGRAGEN EFTU_ECOLI (P02990) GGRHTPFFKGYRPQFYFRTTDVT EFTU_ECOLI (P02990) GRQVGVPYIIVF EFTU_ECOLI (P02990) HVDHGKTTLTAAI EFTU_ECOLI (P02990) IKETQKSTCTGVEM EFTU_ECOLI (P02990) LDSYIPEPERA EFTU_ECOLI (P02990) TINTSHVEYDTPTRH EFTU_ECOLI (P02990) VGTIGHVDHGKTTLT EFTU_ECOLI (P02990) VKNMITGAAQMDGAILVVAATDGPMPQT EFTU_ECOLI (P02990) VNVGTIGHVDHGKTT EFTU_ECOLI (P02990) VVAATDGPMPQTRE EFTU_ECOLI (P02990) YDFPGDDTPIVRGSAL EFTU_ECOLI (P02990) YFRTTDVTGT EFTU_ECOLI (P02990) YVKNMITGAAQM G3P1_ECOLI (P06977) GAAKAVGKVLP G3P1_ECOLI (P06977) GGRGASQNII G3P1_ECOLI (P06977) GPSHKDWRGGRGAS G3P1_ECOLI (P06977) KGANFDKYAGQDIVSNASCTTNCLAPLA G3P1_ECOLI (P06977) NIIPSSTGAAKAVGKVLPEL G3P1_ECOLI (P06977) PANLKWDEVGVDVVAEATGLFLTDETAR G3P1_ECOLI (P06977) TIKVGINGF G3P1_ECOLI (P06977) YDNETGYSNKVLDL GLNH_ECOLI (P10344) DYELKPMDF GLNH_ECOLI (P10344) EAQQYGIAFPKGSDE GLNH_ECOLI (P10344) EAQQYGIAFPKGSDEL GLNH_ECOLI (P10344) GNGQFKAVGDSL GLNH_ECOLI (P10344) LKPMDFSGIIPALQTKNVDL GLNH_ECOLI (P10344) LRQFPNIDNA GLNH_ECOLI (P10344) MELGTNRADAVL GLNH_ECOLI (P10344) YMELGTNRADAVL GLR3_ECOLI (P37687) DARGGLDPLLK GLR3_ECOLI (P37687) EEMIKRSGRTTVPQ GLR3_ECOLI (P37687) IDAQHIGGCDDL GLR3_ECOLI (P37687) KGVSFQELPIDGNAAKREE GLR3_ECOLI (P37687) QELPIDGNAAKREEM GLR3_ECOLI (P37687) YALDARGGLDPLLK HDEA_ECOLI (P26604) ACTQDKQANF HDEA_ECOLI (P26604) ADAQKAADNKKPVNS HDEA_ECOLI (P26604) ADAQKAADNKKPVNSW HDEA_ECOLI (P26604) ADAQKAADNKKPVNSWT HDEA_ECOLI (P26604) ADAQKAADNKKPVNSWTCE HDEA_ECOLI (P26604) ADAQKAADNKKPVNSWTCEDF HDEA_ECOLI (P26604) AEALNNKDKPED HDEA_ECOLI (P26604) AEALNNKDKPEDAVL HDEA_ECOLI (P26604) ALNNKDKPED HDEA_ECOLI (P26604) AQKAADNKKPVNSW HDEA_ECOLI (P26604) AVDESFQPTA HDEA_ECOLI (P26604) AVDESFQPTAVG HDEA_ECOLI (P26604) AVDESFQPTAVGF HDEA_ECOLI (P26604) CTQDKQANF HDEA_ECOLI (P26604) DAQKAADNKKPVNS HDEA_ECOLI (P26604) DAQKAADNKKPVNSW HDEA_ECOLI (P26604) DAVLDVQGIATVTP HDEA_ECOLI (P26604) DESFQPTA HDEA_ECOLI (P26604) DESFQPTAVGF HDEA_ECOLI (P26604) DKIKKD HDEA_ECOLI (P26604) DVQGIA HDEA_ECOLI (P26604) DVQGIATVT HDEA_ECOLI (P26604) DVQGIATVTPAIVQ HDEA_ECOLI (P26604) DVQGIATVTPAIVQA HDEA_ECOLI (P26604) EALNNKDKPED HDEA_ECOLI (P26604) ESFQPTAVGF
131

HDEA_ECOLI (P26604) FAEALNNKDKPED HDEA_ECOLI (P26604) FAEALNNKDKPEDAVL HDEA_ECOLI (P26604) GFAEALNNKD HDEA_ECOLI (P26604) IATVTPAIVQ HDEA_ECOLI (P26604) KDKVKGEW HDEA_ECOLI (P26604) LAVDE HDEA_ECOLI (P26604) LAVDESFQPTA HDEA_ECOLI (P26604) LAVDESFQPTAVG HDEA_ECOLI (P26604) LAVDESFQPTAVGF HDEA_ECOLI (P26604) NKDKPEDAVLDVQGIAT HDEA_ECOLI (P26604) NKKPVNSW HDEA_ECOLI (P26604) PAIVQACTQDKQANFKDK HDEA_ECOLI (P26604) PTAVGFAEALNNKDKPE HDEA_ECOLI (P26604) QGIATVTPAIVQ HDEA_ECOLI (P26604) QPTAVGF HDEA_ECOLI (P26604) SFQPTA HDEA_ECOLI (P26604) SFQPTAVGF HDEA_ECOLI (P26604) SFQPTAVGFAEALNN HDEA_ECOLI (P26604) TAVGFAEALNNKDK HDEA_ECOLI (P26604) TCEDF HDEA_ECOLI (P26604) TVTPAIVQ HDEA_ECOLI (P26604) TVTPAIVQA HDEA_ECOLI (P26604) VDESFQPTAVGF HDEA_ECOLI (P26604) VTPAIVQ HDEA_ECOLI (P26604) WTCEDF HDEB_ECOLI (P26605) ANESAKDMTCQEF HDEB_ECOLI (P26605) ETVYKGGDT HDEB_ECOLI (P26605) ETVYKGGDTVT HDEB_ECOLI (P26605) ETVYKGGDTVTL HDEB_ECOLI (P26605) FIDLNPKAMT HDEB_ECOLI (P26605) FIDLNPKAMTPVAW HDEB_ECOLI (P26605) FKNQASNDLPN HDEB_ECOLI (P26605) HEETVYKGGDTVT HDEB_ECOLI (P26605) IDLNPKAMT HDEB_ECOLI (P26605) IDLNPKAMTPVAW HDEB_ECOLI (P26605) IDLNPKAMTPVAWW HDEB_ECOLI (P26605) IDLNPKAMTPVAWWML HDEB_ECOLI (P26605) KAFIFMGAVAALS HDEB_ECOLI (P26605) LNETDL HDEB_ECOLI (P26605) LTQIPKVIE HDEB_ECOLI (P26605) LTQIPKVIEY HDEB_ECOLI (P26605) MTPVAW HDEB_ECOLI (P26605) NETDL HDEB_ECOLI (P26605) NETDLTQIPKVIE HDEB_ECOLI (P26605) NISSLRKAFIFM HDEB_ECOLI (P26605) NPKAMTPVAW HDEB_ECOLI (P26605) TDLTQIPKVIE HDEB_ECOLI (P26605) TQIPKVIE HDEB_ECOLI (P26605) TQIPKVIEY HDEB_ECOLI (P26605) VYKGGDTVTL HDEB_ECOLI (P26605) WMLHEET HLPA_ECOLI (P11457) ADKIAIVNMGSLFQQVAQKTGVS HLPA_ECOLI (P11457) ADKIAIVNMGSLFQQVAQKTGVSNT HLPA_ECOLI (P11457) ADKIAIVNMGSLFQQVAQKTGVSNTL HLPA_ECOLI (P11457) ADKIAIVNMGSLFQQVAQKTGVSNTLE HLPA_ECOLI (P11457) ADKIAIVNMGSLFQQVAQKTGVSNTLENE HLPA_ECOLI (P11457) AIVNMGSLFQQVAQKTG HLPA_ECOLI (P11457) FQQVAQKTGVSNTLE HLPA_ECOLI (P11457) FQQVAQKTGVSNTLENE HLPA_ECOLI (P11457) GRASELQRMETDLQAK HLPA_ECOLI (P11457) NEFKGRASELQRM HLPA_ECOLI (P11457) VDANAVAYNSSDVKDITAD HNS_ECOLI (P08936) DPNELL
132

HNS_ECOLI (P08936) EKLEV HNS_ECOLI (P08936) EVVVNE HNS_ECOLI (P08936) IADGIDPNEL HNS_ECOLI (P08936) IADGIDPNELL HNS_ECOLI (P08936) IADGIDPNELLNS HNS_ECOLI (P08936) IKKAMDEQGKSLDD HNS_ECOLI (P08936) IKKAMDEQGKSLDDF HNS_ECOLI (P08936) IRTLRA HNS_ECOLI (P08936) LEKLEV HNS_ECOLI (P08936) LKILNN HNS_ECOLI (P08936) LNSLA HNS_ECOLI (P08936) SEALKILNN HNS_ECOLI (P08936) TKTWTGQGRTPAVIK HNS_ECOLI (P08936) VDENGETKTWTGQGRTPAV HNS_ECOLI (P08936) VDENGETKTWTGQGRTPAVI HNS_ECOLI (P08936) WTGQGRTPAV HNS_ECOLI (P08936) WTGQGRTPAVI IPYR_ECOLI (P17288) AKAEIVASFERAKN IPYR_ECOLI (P17288) EAAKAEIVASFER IPYR_ECOLI (P17288) IEIPANADPIKY IPYR_ECOLI (P17288) IEIPANADPIKYEIDKESGALFVDRF IPYR_ECOLI (P17288) SLLNVPAGKDLPED IPYR_ECOLI (P17288) SLLNVPAGKDLPEDI IPYR_ECOLI (P17288) SLLNVPAGKDLPEDIY IPYR_ECOLI (P17288) VIEIPANADPIKY IPYR_ECOLI (P17288) VLVPTPYPLQPGSV IPYR_ECOLI (P17288) VVIEIPANADPIKY IPYR_ECOLI (P17288) VVIEIPANADPIKYEID IPYR_ECOLI (P17288) VVIEIPANADPIKYEIDKESGAL IPYR_ECOLI (P17288) YDHIKDVNDLPEL IPYR_ECOLI (P17288) YGYINHTLSL KDUI_ECOLI (Q46938) ASIDTGTPAKF KDUI_ECOLI (Q46938) EIGHRDALYVGKGA KDUI_ECOLI (Q46938) SIHSGVGTKAYT KDUI_ECOLI (Q46938) VTPDEVSPVTLGDNLTS KDUI_ECOLI (Q46938) YEIGHRDAL MALE_ECOLI (P02928) AVINAASGRQTVDE MALE_ECOLI (P02928) KFPQVAATGDGPDIIFWAHDR MALE_ECOLI (P02928) PDKLEEKFPQVAATGDGPDII MALE_ECOLI (P02928) TVDEALKDAQTRI MALE_ECOLI (P02928) VIWINGDKGYNGL OSMY_ECOLI (P27291) AKAVDGVKSVKNDL OSMY_ECOLI (P27291) AVKVAKGVE OSMY_ECOLI (P27291) AVKVAKGVEGVTSVSDKL OSMY_ECOLI (P27291) DGVKSVKNDLKTK OSMY_ECOLI (P27291) DHDNIKSTD OSMY_ECOLI (P27291) DIVPSRHVKVETTDG OSMY_ECOLI (P27291) DTATTSEIKAKLLADDIVPS OSMY_ECOLI (P27291) ENNAQTTNESAG OSMY_ECOLI (P27291) ENNAQTTNESAGQKVDSSM OSMY_ECOLI (P27291) ENNAQTTNESAGQKVDSSMNKVGNF OSMY_ECOLI (P27291) FVESQAQAEE OSMY_ECOLI (P27291) FVESQAQAEEA OSMY_ECOLI (P27291) GVTSVSDKL OSMY_ECOLI (P27291) GVTSVSDKLHVRDAKEG OSMY_ECOLI (P27291) HVRDAKEGSVKGYAG OSMY_ECOLI (P27291) IAKAVDGVKSVKNDL OSMY_ECOLI (P27291) IAKAVDGVKSVKNDLKTK OSMY_ECOLI (P27291) ISVKTDQKVVT OSMY_ECOLI (P27291) ISVKTDQKVVTLSG OSMY_ECOLI (P27291) KVETTDGVVQLSGTVDSQA OSMY_ECOLI (P27291) LADDI OSMY_ECOLI (P27291) LADDIVPSRHVKVETTDG
133

OSMY_ECOLI (P27291) LSGFVESQAQAEEAVKVAKGVEGVTS OSMY_ECOLI (P27291) NKVGNF OSMY_ECOLI (P27291) PSRHVKVETTDG OSMY_ECOLI (P27291) PSRHVKVETTDGVVQLS OSMY_ECOLI (P27291) RDAKEGSVKGYA OSMY_ECOLI (P27291) SAGQKVDSSM OSMY_ECOLI (P27291) SGTVDSQAQSDRAES OSMY_ECOLI (P27291) SGTVDSQAQSDRAESIA OSMY_ECOLI (P27291) SQAQSDRAESIAKA OSMY_ECOLI (P27291) SVKTDQKVVT OSMY_ECOLI (P27291) SVSDKL OSMY_ECOLI (P27291) TDGVVQLSGTVDSQA OSMY_ECOLI (P27291) VDHDNIKSTD OSMY_ECOLI (P27291) VDHDNIKSTDIS OSMY_ECOLI (P27291) VKVAKGVEGVTSVSDKL OSMY_ECOLI (P27291) VQLSGTVDSQAQSDRAES OSMY_ECOLI (P27291) VVQLSGTVDSQAQSD OSMY_ECOLI (P27291) VVQLSGTVDSQAQSDRAES OSMY_ECOLI (P27291) VVQLSGTVDSQAQSDRAESIA PTGA_ECOLI (P08837) DGTIGKIFETNHAFSI PTGA_ECOLI (P08837) GETPVIRIKK PTGA_ECOLI (P08837) IEDVPDVVF PTGA_ECOLI (P08837) IEFDLPLLE PTGA_ECOLI (P08837) IIAPLSGE PTGA_ECOLI (P08837) IVNIEDVPDVVF PTGA_ECOLI (P08837) SGVELFVHFGIDTVELKG PTGA_ECOLI (P08837) TGTIEIIAPLSGEI PTHP_ECOLI (P07006) EAKGFTSEITVTSNGKSA PTHP_ECOLI (P07006) FKLQTLGL PTHP_ECOLI (P07006) FKLQTLGLTQGTVVT PTHP_ECOLI (P07006) FVKEAKGF PTHP_ECOLI (P07006) FVKEAKGFTSEITV PTHP_ECOLI (P07006) ISAEGEDEQKAVEHL PTHP_ECOLI (P07006) ITAPNGLHTRPAAQ PTHP_ECOLI (P07006) ITAPNGLHTRPAAQF PTHP_ECOLI (P07006) ITVTSNGKSASAKSL PTHP_ECOLI (P07006) ITVTSNGKSASAKSLF PTHP_ECOLI (P07006) KGFTSEITVTSNGK PTHP_ECOLI (P07006) KLQTLGLTQGTVVT PTHP_ECOLI (P07006) MFQQEVTITAPNG PTHP_ECOLI (P07006) QEVTITAPNGLHT PTHP_ECOLI (P07006) SLFKLQTLGLTQGTVVTISAEGEDEQKAVEHLV PTHP_ECOLI (P07006) TVTSNGKSASAKSL PTHP_ECOLI (P07006) VTITAPNGL PTHP_ECOLI (P07006) VTITAPNGLHTR PTHP_ECOLI (P07006) VTITAPNGLHTRPAAQ PTHP_ECOLI (P07006) VTITAPNGLHTRPAAQF PTHP_ECOLI (P07006) VTSNGKSASAKSL RBSB_ECOLI (P02925) AAHKFNVL RBSB_ECOLI (P02925) AAHKFNVLASQPADFDRIKGLNVM RBSB_ECOLI (P02925) AATIAQLPDQIGAKGVE RBSB_ECOLI (P02925) AATIAQLPDQIGAKGVETA RBSB_ECOLI (P02925) AKKAGEGAKV RBSB_ECOLI (P02925) ALSATVSANAMAKDTIALVVSTLNNPFFVSLKD RBSB_ECOLI (P02925) ANIPVITL RBSB_ECOLI (P02925) ARERGEGFQQA RBSB_ECOLI (P02925) ASQPADF RBSB_ECOLI (P02925) AVAAHKFNVL RBSB_ECOLI (P02925) AVALSATVSANAMAKDTIALV RBSB_ECOLI (P02925) DGTPDGEKAVNDGKL RBSB_ECOLI (P02925) DGTPDGEKAVNDGKLAAT RBSB_ECOLI (P02925) DKVLKGEKVQAKYPVD RBSB_ECOLI (P02925) DRIKGLNV
134

RBSB_ECOLI (P02925) DRQATKGEVVSHIASD RBSB_ECOLI (P02925) DRQATKGEVVSHIASDN RBSB_ECOLI (P02925) DRQATKGEVVSHIASDNVL RBSB_ECOLI (P02925) DRQATKGEVVSHIASDNVLGGKIAGD RBSB_ECOLI (P02925) DSQNNPAKE RBSB_ECOLI (P02925) DSQNNPAKELANVQD RBSB_ECOLI (P02925) DSQNNPAKELANVQDL RBSB_ECOLI (P02925) FDRIKGLNV RBSB_ECOLI (P02925) FQQAVAAHKFNVL RBSB_ECOLI (P02925) GAKGVETADKVLKGEKVQAKYPVD RBSB_ECOLI (P02925) GAKGVETADKVLKGEKVQAKYPVDL RBSB_ECOLI (P02925) GGKIAGDYIAKKAG RBSB_ECOLI (P02925) GIAGTSAARERGEGF RBSB_ECOLI (P02925) GIAGTSAARERGEGFQQAVAA RBSB_ECOLI (P02925) HIASDNVLGGKIAGD RBSB_ECOLI (P02925) IAKKAGEGAKVIE RBSB_ECOLI (P02925) IAQLPDQIGAKG RBSB_ECOLI (P02925) IAQLPDQIGAKGVE RBSB_ECOLI (P02925) IAQLPDQIGAKGVET RBSB_ECOLI (P02925) IAQLPDQIGAKGVETA RBSB_ECOLI (P02925) IAQLPDQIGAKGVETAD RBSB_ECOLI (P02925) IAQLPDQIGAKGVETADKVL RBSB_ECOLI (P02925) IAQLPDQIGAKGVETADKVLKGEKVQ RBSB_ECOLI (P02925) IAQLPDQIGAKGVETADKVLKGEKVQAKYPVD RBSB_ECOLI (P02925) IASDNVLGGKIAGD RBSB_ECOLI (P02925) INPTDSDAVGNA RBSB_ECOLI (P02925) INPTDSDAVGNAVKMANQANIPVITL RBSB_ECOLI (P02925) KDGAQKEADKL RBSB_ECOLI (P02925) KDGAQKEADKLGYNL RBSB_ECOLI (P02925) KDTIAL RBSB_ECOLI (P02925) KGEVVSHIASDNVLGG RBSB_ECOLI (P02925) KVIELQGIAGTSAARERG RBSB_ECOLI (P02925) LANVQD RBSB_ECOLI (P02925) LANVQDL RBSB_ECOLI (P02925) LANVQDLTVRGT RBSB_ECOLI (P02925) LGGKIAGDY RBSB_ECOLI (P02925) LGYNLVVLDSQNNPAKELANVQD RBSB_ECOLI (P02925) LINPTDSDAVGNA RBSB_ECOLI (P02925) LKDGAQKEADK RBSB_ECOLI (P02925) LLINPTDSDAVGNA RBSB_ECOLI (P02925) LLTAHPDVQAVFAQNDEMA RBSB_ECOLI (P02925) LNNPFFVSLKDGAQ RBSB_ECOLI (P02925) LQGIAGTSA RBSB_ECOLI (P02925) LQGIAGTSAARERGEG RBSB_ECOLI (P02925) LQTAGKSDVM RBSB_ECOLI (P02925) LRALQTAGKSDV RBSB_ECOLI (P02925) LRALQTAGKSDVM RBSB_ECOLI (P02925) LTAHPDVQ RBSB_ECOLI (P02925) LTVRGTKIL RBSB_ECOLI (P02925) LTVRGTKILL RBSB_ECOLI (P02925) LVVSTL RBSB_ECOLI (P02925) LVVSTLNNPFF RBSB_ECOLI (P02925) MQNLLTAHPDVQ RBSB_ECOLI (P02925) MVVGFDGTPDGEKAVNDGKL RBSB_ECOLI (P02925) MVVGFDGTPDGEKAVNDGKLAAT RBSB_ECOLI (P02925) NLLTAHPDVQ RBSB_ECOLI (P02925) NPFFVSLKDGAQKE RBSB_ECOLI (P02925) NVLGGKIAGD RBSB_ECOLI (P02925) NVLGGKIAGDY RBSB_ECOLI (P02925) PDGEKAVNDGKL RBSB_ECOLI (P02925) PDGEKAVNDGKLAAT RBSB_ECOLI (P02925) QGIAGTSA RBSB_ECOLI (P02925) QGIAGTSAARERGEG
135
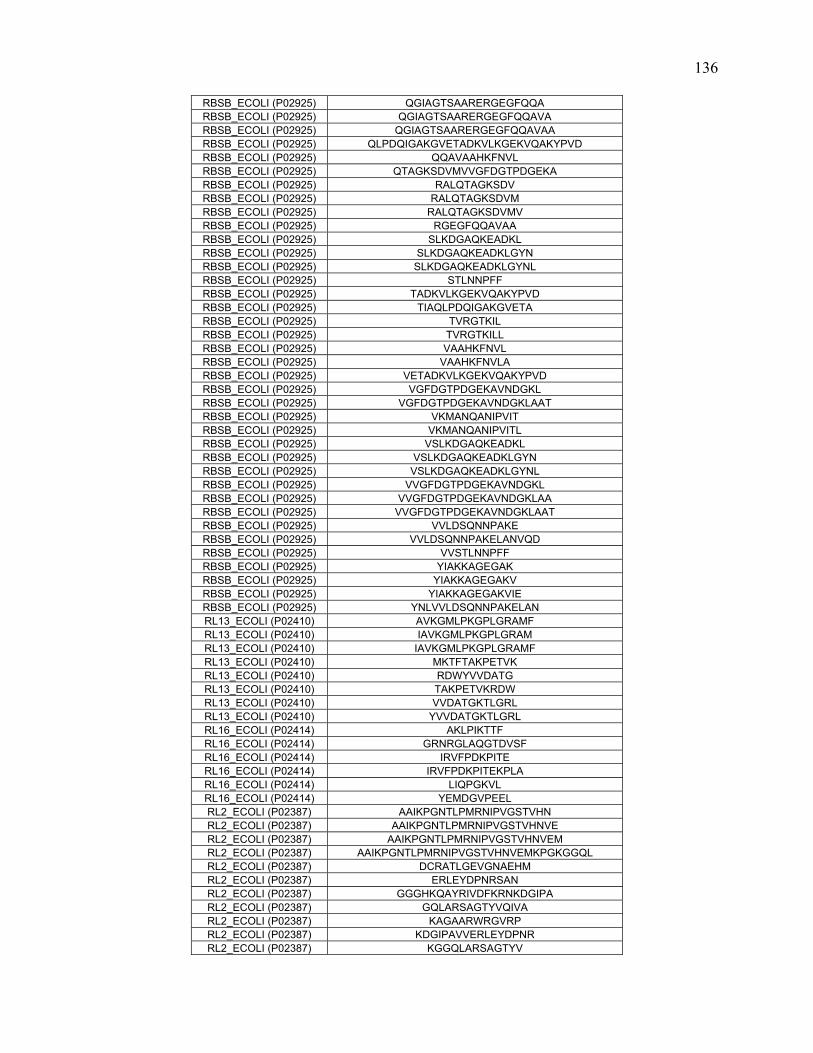
RBSB_ECOLI (P02925) QGIAGTSAARERGEGFQQA RBSB_ECOLI (P02925) QGIAGTSAARERGEGFQQAVA RBSB_ECOLI (P02925) QGIAGTSAARERGEGFQQAVAA RBSB_ECOLI (P02925) QLPDQIGAKGVETADKVLKGEKVQAKYPVD RBSB_ECOLI (P02925) QQAVAAHKFNVL RBSB_ECOLI (P02925) QTAGKSDVMVVGFDGTPDGEKA RBSB_ECOLI (P02925) RALQTAGKSDV RBSB_ECOLI (P02925) RALQTAGKSDVM RBSB_ECOLI (P02925) RALQTAGKSDVMV RBSB_ECOLI (P02925) RGEGFQQAVAA RBSB_ECOLI (P02925) SLKDGAQKEADKL RBSB_ECOLI (P02925) SLKDGAQKEADKLGYN RBSB_ECOLI (P02925) SLKDGAQKEADKLGYNL RBSB_ECOLI (P02925) STLNNPFF RBSB_ECOLI (P02925) TADKVLKGEKVQAKYPVD RBSB_ECOLI (P02925) TIAQLPDQIGAKGVETA RBSB_ECOLI (P02925) TVRGTKIL RBSB_ECOLI (P02925) TVRGTKILL RBSB_ECOLI (P02925) VAAHKFNVL RBSB_ECOLI (P02925) VAAHKFNVLA RBSB_ECOLI (P02925) VETADKVLKGEKVQAKYPVD RBSB_ECOLI (P02925) VGFDGTPDGEKAVNDGKL RBSB_ECOLI (P02925) VGFDGTPDGEKAVNDGKLAAT RBSB_ECOLI (P02925) VKMANQANIPVIT RBSB_ECOLI (P02925) VKMANQANIPVITL RBSB_ECOLI (P02925) VSLKDGAQKEADKL RBSB_ECOLI (P02925) VSLKDGAQKEADKLGYN RBSB_ECOLI (P02925) VSLKDGAQKEADKLGYNL RBSB_ECOLI (P02925) VVGFDGTPDGEKAVNDGKL RBSB_ECOLI (P02925) VVGFDGTPDGEKAVNDGKLAA RBSB_ECOLI (P02925) VVGFDGTPDGEKAVNDGKLAAT RBSB_ECOLI (P02925) VVLDSQNNPAKE RBSB_ECOLI (P02925) VVLDSQNNPAKELANVQD RBSB_ECOLI (P02925) VVSTLNNPFF RBSB_ECOLI (P02925) YIAKKAGEGAK RBSB_ECOLI (P02925) YIAKKAGEGAKV RBSB_ECOLI (P02925) YIAKKAGEGAKVIE RBSB_ECOLI (P02925) YNLVVLDSQNNPAKELAN RL13_ECOLI (P02410) AVKGMLPKGPLGRAMF RL13_ECOLI (P02410) IAVKGMLPKGPLGRAM RL13_ECOLI (P02410) IAVKGMLPKGPLGRAMF RL13_ECOLI (P02410) MKTFTAKPETVK RL13_ECOLI (P02410) RDWYVVDATG RL13_ECOLI (P02410) TAKPETVKRDW RL13_ECOLI (P02410) VVDATGKTLGRL RL13_ECOLI (P02410) YVVDATGKTLGRL RL16_ECOLI (P02414) AKLPIKTTF RL16_ECOLI (P02414) GRNRGLAQGTDVSF RL16_ECOLI (P02414) IRVFPDKPITE RL16_ECOLI (P02414) IRVFPDKPITEKPLA RL16_ECOLI (P02414) LIQPGKVL RL16_ECOLI (P02414) YEMDGVPEEL RL2_ECOLI (P02387) AAIKPGNTLPMRNIPVGSTVHN RL2_ECOLI (P02387) AAIKPGNTLPMRNIPVGSTVHNVE RL2_ECOLI (P02387) AAIKPGNTLPMRNIPVGSTVHNVEM RL2_ECOLI (P02387) AAIKPGNTLPMRNIPVGSTVHNVEMKPGKGGQL RL2_ECOLI (P02387) DCRATLGEVGNAEHM RL2_ECOLI (P02387) ERLEYDPNRSAN RL2_ECOLI (P02387) GGGHKQAYRIVDFKRNKDGIPA RL2_ECOLI (P02387) GQLARSAGTYVQIVA RL2_ECOLI (P02387) KAGAARWRGVRP RL2_ECOLI (P02387) KDGIPAVVERLEYDPNR RL2_ECOLI (P02387) KGGQLARSAGTYV
136

RL2_ECOLI (P02387) NIPVGSTVHN RL2_ECOLI (P02387) PMRNIPVGSTVHN RL2_ECOLI (P02387) PMRNIPVGSTVHNVE RL2_ECOLI (P02387) RATLGEVGNAEHM RL2_ECOLI (P02387) RVLGKAGAARWRGV RL2_ECOLI (P02387) TTRHIGGGHKQAYRIVDFK RL2_ECOLI (P02387) VQIVARDGAY RL2_ECOLI (P02387) VRGTAMNPVDHPHGGGEGR
RL24_ECOLI (P02425) AATGKADRVGFRFEDG RL24_ECOLI (P02425) AIFNAATGKADRVGF RL24_ECOLI (P02425) EAAIQVSNVAIFNAAT RL24_ECOLI (P02425) FKSNSETIK RL24_ECOLI (P02425) FNAATGKADRVGFRFEDG RL24_ECOLI (P02425) NAATGKADRVGF RL24_ECOLI (P02425) RFFKSNSETI RL29_ECOLI (P02429) AASGQLQQSHLLK RL29_ECOLI (P02429) GQLQQSHLLKQ RL29_ECOLI (P02429) LREKSVEEL RL29_ECOLI (P02429) QAASGQLQQSHL RL29_ECOLI (P02429) RMQAASGQLQQSHL RL3_ECOLI (P02386) ADVKKVDVTGTSKGKGF RL3_ECOLI (P02386) AGQMGNERVTVQSL RL3_ECOLI (P02386) DVTGTSKGKGFAGTVKRWNFRTQDA RL3_ECOLI (P02386) ELFADVKKVDVT RL3_ECOLI (P02386) FADVKKVDVTGTSKGKGF RL3_ECOLI (P02386) GLWEFRLAEGEEF RL3_ECOLI (P02386) GYRAIQVTTGAKKANRVTKPEAGHFA RL3_ECOLI (P02386) KKMAGQMGNERVTVQSL RL3_ECOLI (P02386) LVKGAVPGATGSDL RL3_ECOLI (P02386) NSLSHRVPG RL3_ECOLI (P02386) NSLSHRVPGSI RL3_ECOLI (P02386) RIFTEDGVSIPVTVIEVEANRVTQV RL3_ECOLI (P02386) RTQDATHGNSLSHRVPGSIGQNQTP RL3_ECOLI (P02386) TGAKKANRVTKPEAG RL3_ECOLI (P02386) TGTSKGKGFAGTVKRWNFRT RL3_ECOLI (P02386) TVGQSISVELFADV RL6_ECOLI (P02390) GPRDGY RL6_ECOLI (P02390) KGADKQVIGQVA RL6_ECOLI (P02390) QAGTARALLNSMVIGVTEG RL6_ECOLI (P02390) QVIGQVAADLRAY RL6_ECOLI (P02390) SLGFSHPVDHQLPAGIT RL6_ECOLI (P02390) SLGFSHPVDHQLPAGITAECPTQTE RL6_ECOLI (P02390) SRVAKAPVVVPAGVD RL7_ECOLI (P02392) AAAVAVAAGPVEAAEEKT RL7_ECOLI (P02392) AVAAGPVEAAEEKTEFDVI RL7_ECOLI (P02392) FGVSAAAAVAVAAGPVEAAE RL7_ECOLI (P02392) IKAVRGATGLGL RL7_ECOLI (P02392) KAAGANKVAV RL7_ECOLI (P02392) KAAGANKVAVIKA RL7_ECOLI (P02392) LISAMEEKFGVSAAAAVAVAAGPVEAAEE RL7_ECOLI (P02392) LVESAPAAL RL7_ECOLI (P02392) VAVAAGPVE RL7_ECOLI (P02392) VAVAAGPVEA
RS15_ECOLI (P02371) AQINHLQGHF RS15_ECOLI (P02371) FGRDANDTGSTEVQ RS15_ECOLI (P02371) GRDANDTGSTEVQVA RS15_ECOLI (P02371) SLSTEA RS15_ECOLI (P02371) SLSTEATAKIVSE RS15_ECOLI (P02371) TAQINHLQGHF RS6_ECOLI (P02358) AHYVLMNVEAPQEVID RS6_ECOLI (P02358) FRFNDAVIRSMVMRT RS6_ECOLI (P02358) ITGAEGKIHRLED RS6_ECOLI (P02358) LMNVEAPQEVI
137

RS6_ECOLI (P02358) MVHPDQSEQVPGM RS6_ECOLI (P02358) QVPGMIERYTAAIT
SODC_ECOLI (P53635) EGPEGAGHLGDLPALVVN SODC_ECOLI (P53635) IGSVTITETDKGLEFS SODC_ECOLI (P53635) IHAKGSCQPATKDGKASAAE SODC_ECOLI (P53635) ITETDKGLEFSPDL SODC_ECOLI (P53635) LVTSQGVGQSIGSVT SODC_ECOLI (P53635) MSDQPKPLGGGGERYACG SODC_ECOLI (P53635) MSDQPKPLGGGGERYACGV SODC_ECOLI (P53635) NLVTSQGVGQSIGS SODC_ECOLI (P53635) NLVTSQGVGQSIGSVT SODC_ECOLI (P53635) QGVGQSIGSVTITETDK SODC_ECOLI (P53635) TGAQAASEKVEMNLVT SODC_ECOLI (P53635) TKDGKASAAESAGGHL SODC_ECOLI (P53635) VATGAQAASEKVE SODC_ECOLI (P53635) VATGAQAASEKVEMNLVT SODC_ECOLI (P53635) VHVGGDNMSDQPKPLGGGGERYACG SODC_ECOLI (P53635) VTSQGVGQSIGSVT SODC_ECOLI (P53635) VTSQGVGQSIGSVTITET SODC_ECOLI (P53635) VVATGAQAASEK STFR_ECOLI (P76072) AAAQSKSTAESAATRAETAAKRAEDIASAVALEDA STFR_ECOLI (P76072) AAARSASAAKTSETNAKASET STFR_ECOLI (P76072) AAASSASSAASSASSASASKDEA STFR_ECOLI (P76072) AAKSSETNASSSASSAASSAT STFR_ECOLI (P76072) AARSASAAKTSETN STFR_ECOLI (P76072) AASTSAGQASASATAAG STFR_ECOLI (P76072) AATSAGAAKTSETNASASLQSAATSASTATTK STFR_ECOLI (P76072) ADKRGMRYVRVNAP STFR_ECOLI (P76072) AGAAKTSETNASASLQSA STFR_ECOLI (P76072) AGSATAAAQSKSTAESAATRA STFR_ECOLI (P76072) APGADLVVNDTTY STFR_ECOLI (P76072) AQNTAAAKKSASDA STFR_ECOLI (P76072) ARNASAVAQNTAAAKKSASD STFR_ECOLI (P76072) ARSSETAAGQSASAAAGSKT STFR_ECOLI (P76072) ASAGAHAHTVGIGAH STFR_ECOLI (P76072) ASSAASSASSASASKDEATRQASAAKSSATTASTKATEA STFR_ECOLI (P76072) ASSASSAASSASSASASKDEATR STFR_ECOLI (P76072) ASSASSASASKDE STFR_ECOLI (P76072) ASSSAGTASTKATEASKSAAAA STFR_ECOLI (P76072) ASSTDLGTKTTSSFDY STFR_ECOLI (P76072) ATAAGNSAKAAK STFR_ECOLI (P76072) ATSASTATTKASEAATS STFR_ECOLI (P76072) AVAQNTAAAK STFR_ECOLI (P76072) EEVARNASAVAQNTAAAK STFR_ECOLI (P76072) ETAAKRAEDIASAVA STFR_ECOLI (P76072) GATNPATECIAADV STFR_ECOLI (P76072) GATSGKYYPVV STFR_ECOLI (P76072) GHTITVNAAGNAENTVK STFR_ECOLI (P76072) HSASASSTDLGTKTTSSFDYGTKSTNNTGAHTHSVS STFR_ECOLI (P76072) KSAAAAESSKSAAATSAG STFR_ECOLI (P76072) KSSATTASTKATEA STFR_ECOLI (P76072) KSSATTASTKATEAAGS STFR_ECOLI (P76072) LLTNQGDVYGGWNTLR STFR_ECOLI (P76072) LQSAATSASTATTKA STFR_ECOLI (P76072) NAAGNAENTVKN STFR_ECOLI (P76072) NAPAGATSGKYYP STFR_ECOLI (P76072) NTAAAKKSASDAST STFR_ECOLI (P76072) NVNTASANSGAGSASTRLSVVHNQNYATSSAGAHTHS STFR_ECOLI (P76072) PAGATSGKYYPVVVM STFR_ECOLI (P76072) PESYPVGAPIPWPSDTVPSGYALMQGQAFDKSAY STFR_ECOLI (P76072) PSHAGTITVYEDSQPGT STFR_ECOLI (P76072) PVFAFIEDGLSI STFR_ECOLI (P76072) QASASATAAGKSAESAA
138

STFR_ECOLI (P76072) QASASATAAGKSAESAASS STFR_ECOLI (P76072) QSASAAAGSKTAAASSA STFR_ECOLI (P76072) RAASTSAGQAASSAQSASSS STFR_ECOLI (P76072) RSASAAKTSETNAKA STFR_ECOLI (P76072) SAGAAKTSET STFR_ECOLI (P76072) SANSGAGSASTRLS STFR_ECOLI (P76072) SASLNGNALTAT STFR_ECOLI (P76072) SKSAAATSAGAAKTS STFR_ECOLI (P76072) SKTAAASSASSAASSA STFR_ECOLI (P76072) SLIVNDNLSCKKL STFR_ECOLI (P76072) SSASASKDEATRQASAAKSS STFR_ECOLI (P76072) SSASSASASKDEA STFR_ECOLI (P76072) SSETAAGQSASAA STFR_ECOLI (P76072) STKATEAAGSATAAAQ STFR_ECOLI (P76072) THSASASSTDLGTKTTSSFDYGTKSTNNTGAHTHSV STFR_ECOLI (P76072) THSLANVNTASAN STFR_ECOLI (P76072) TSKNLPPESYPVGAP STFR_ECOLI (P76072) TTKAGEATEQASAAARS STFR_ECOLI (P76072) TTKASEAATSARDAA STFR_ECOLI (P76072) TVGIGAHTHSVA TNAA_ECOLI (P00913) AGGQPVSLANLKAMYSIAKKYDIPVVMDSARFAEN TNAA_ECOLI (P00913) DFKGNFDLEGLERGIEE TNAA_ECOLI (P00913) EDVFIDLLTDSGTGAVTQS TNAA_ECOLI (P00913) ERLAVGLYDGMNL TNAA_ECOLI (P00913) FDLEGLERGIEEVGPNNV TNAA_ECOLI (P00913) GLEGGAMERLA TNAA_ECOLI (P00913) GLERGIEEVGPNNV TNAA_ECOLI (P00913) IIKSGMNPFL TNAA_ECOLI (P00913) LAMSAKKDAMVPMGGL TNAA_ECOLI (P00913) LEGLERGIEEVGPNNV TNAA_ECOLI (P00913) LRLTIPRATY TNAA_ECOLI (P00913) LTDSGTGAVTQSMQ TNAA_ECOLI (P00913) TITSNSAGGQPVSL TNAA_ECOLI (P00913) VATITSNSAGGQPVSLANL TNAA_ECOLI (P00913) VDAGKLLPHIPADQF TNAA_ECOLI (P00913) VDAGKLLPHIPADQFPAQAL TNAA_ECOLI (P00913) VSLANLKAMYSIAKKY TTDT_ECOLI (P39414) AAALAMPEIP TTDT_ECOLI (P39414) ELILAPVTPSNSARG TTDT_ECOLI (P39414) EQLAQPGFKFTAK TTDT_ECOLI (P39414) FCLMVGAAIGLGSI TTDT_ECOLI (P39414) GDQVPRWAETELQAMGP TTDT_ECOLI (P39414) GFKFTAKSLSWAVSG TTDT_ECOLI (P39414) MVGAAIGLGSILTPYA TTDT_ECOLI (P39414) PEIPLPVFCLMVG TTDT_ECOLI (P39414) SLSWAVSGF TTDT_ECOLI (P39414) TGYEKTGLGR YBGS_ECOLI (P75758) AADAGQVAPDARE YBGS_ECOLI (P75758) AALAADSGAQTNNGQANAAADAGQVAPDARENVA YBGS_ECOLI (P75758) ADSGAQTNNGQANAAADAGQVAPDARE YBGS_ECOLI (P75758) ALAADSGAQTNNGQANAAADAGQ YBGS_ECOLI (P75758) KVQTGDGINNDVDTKTDGTTQ YBGS_ECOLI (P75758) NVAPNNVDNNGVNTGSGGTM YBGS_ECOLI (P75758) NVAPNNVDNNGVNTGSGGTML YBGS_ECOLI (P75758) NVAPNNVDNNGVNTGSGGTMLH YBGS_ECOLI (P75758) PDARENVAPNNVDNNGVNTG YBGS_ECOLI (P75758) SGAQTNNGQANAAADAGQVAPDAR YBGS_ECOLI (P75758) SGAQTNNGQANAAADAGQVAPDARE YBGS_ECOLI (P75758) VDNNGVNTGSGGT YCII_ECOLI (P31070) LTAGPMPAVDSNDPGAAGF YCII_ECOLI (P31070) LTAGPMPAVDSNDPGAAGFTGSTV
YGIW_ECOLI (P52083) AAVIAVMALCSAPVMA YGIW_ECOLI (P52083) AAVIAVMALCSAPVMAAEQG
139

YGIW_ECOLI (P52083) AEQGGFSGPSATQSQAGGFQGPNGSVT YGIW_ECOLI (P52083) ALCSAPVMAAEQG YGIW_ECOLI (P52083) CSAPVMAAEQGGFS YGIW_ECOLI (P52083) ESAKSLRDDTW YGIW_ECOLI (P52083) IQGEVDKDWNSVE YGIW_ECOLI (P52083) KDASGTINVDIDHKRWNGVTVTPKDTVEIQG YGIW_ECOLI (P52083) MAAEQGGFSGPSAT YGIW_ECOLI (P52083) RISDDL YGIW_ECOLI (P52083) TVESAKSLRDDTW YGIW_ECOLI (P52083) VDIDHKRWNGVTVTPKDTVE YJBJ_ECOLI (P32691) DWETRN YJBJ_ECOLI (P32691) GKIQERYGYQKDQAEKE YJBJ_ECOLI (P32691) IEGKRDQ YJBJ_ECOLI (P32691) IEGKRDQL YJBJ_ECOLI (P32691) IEGKRDQLVG YJBJ_ECOLI (P32691) IIEGKRDQLVG YJBJ_ECOLI (P32691) KLTDDDMTIIEGK YJBJ_ECOLI (P32691) MTIIEGKRDQL YJBJ_ECOLI (P32691) RYGYQKDQAEKE YJBJ_ECOLI (P32691) TIIEGKRDQL YJBJ_ECOLI (P32691) TIIEGKRDQLVG YJBJ_ECOLI (P32691) VGKIQE YJBJ_ECOLI (P32691) VGKIQERYGYQKDQAEKE YJBJ_ECOLI (P32691) VVDWETRN YJBJ_ECOLI (P32691) VVDWETRNE YJBJ_ECOLI (P32691) VVDWETRNEYRW YJGF_ECOLI (P39330) AAGLKVGDIVKT YJGF_ECOLI (P39330) ENAPAAIGPYVQGVDLG YJGF_ECOLI (P39330) FTEHNATFPARSCVE YJGF_ECOLI (P39330) IATENAPAAIGPYVQG YJGF_ECOLI (P39330) IITSGQIPVNPKTGEVPAD YJGF_ECOLI (P39330) IITSGQIPVNPKTGEVPADVAA YJGF_ECOLI (P39330) IITSGQIPVNPKTGEVPADVAAQARQSL YJGF_ECOLI (P39330) IITSGQIPVNPKTGEVPADVAAQARQSLDNVKA YJGF_ECOLI (P39330) QIPVNPKTGEVPADVAAQ YJGF_ECOLI (P39330) SKTIATENAPAAIGPYVQG YJGF_ECOLI (P39330) SKTIATENAPAAIGPYVQGVD YJGF_ECOLI (P39330) SKTIATENAPAAIGPYVQGVDLGNMIITSGQIPVNPKTGE YJGF_ECOLI (P39330) VKDLNDFATVN YJGF_ECOLI (P39330) YVQGVDLGNMIITSGQIPVNP YJGK_ECOLI (P39335) EPYEARRAEYHA YJGK_ECOLI (P39335) IAFLPEGVDEKT YJGK_ECOLI (P39335) IIGNIHNLQPWLPQE YJGK_ECOLI (P39335) ILNEGDFVVFY YJGK_ECOLI (P39335) KGKHDIEGNR YJGK_ECOLI (P39335) MIIGNIHNLQPWLPQE YJGK_ECOLI (P39335) MIIGNIHNLQPWLPQEL YJGK_ECOLI (P39335) QPWLPQELR YJGK_ECOLI (P39335) WLADKDIAFLPEGV
The left column is the protein identified, named according to its SwissProt entry name.
The SwissProt accession number is listed in parentheses.
140

CHAPTER 4
PERSPECTIVES AND FUTURE DIRECTIONS
4.1 Discussion and Conclusions
4.1.1 Literature search
The analysis of the data that have been gathered from the literature search
indicated that although pepsin is not as specific as other proteases, it does maintain
limited cleavage preferences. At pH 2.5, pepsin will cleave preferentially after most
bulky, hydrophobic amino acids such as leucine and phenylalanine. Additionally, the
residues that most often occur immediately following the cleaved peptide bond are
tryptophan and tyrosine. It has also been shown that pepsin will rarely cleave at proline
and histidine.
4.1.2 Experimental research
The results yielded by the corresponding experimental research support the initial
trends established in the literature search. The data collected for the digest of the proteins
and E.coli samples at pH 2.5 show that pepsin cleaves most often after leucine and
phenylalanine and least often after arginine, cysteine, proline and histidine.
The data obtained from performing digestion of the samples at pH 1.0 and pH 4.0
also yielded similar results. There were no distinct differences of pepsin specificity
between the three pH points. Due to the fact that all three pH points yielded similar
results of pepsin specificity it has been shown that the activity of pepsin is not directly
related to its specificity.
141

4.1.3 Research on pepsin specificity
The research presented in this thesis does not represent the first time that the
specificity of pepsin has been investigated. A study performed by Zhang (Zhang 1995)
examines pepsin specificity by analyzing the cleavages produced from digesting three
standard proteins. Another more recent study into the specificity of pepsin was
performed by Hamuro et al. (Hamuro, Coales et al. 2008). This research is more
comprehensive than that of Zhang as it consists of cleavages produced from the digestion
of 39 proteins.
The work presented by Hamuro et al. is comparable to the work performed for
this thesis. The results were similar as pepsin will cleave preferentially after most bulky
hydrophobic residues. However, there were some differences in the research performed.
The research by Hamuro et al. only considered the digestion of proteins at a pH around
2.5. They also did not strictly control the pH of the sample before digestion. The pH of
the samples that were digested ranged from 1.3 to 2.5.
The research for this thesis puts strict limitations on the pH of both the samples
and the digestion of the samples. This produces a better understanding of pepsin
specificity at a given pH. This work also looks into the specificity of pepsin at other pH
points, thus expanding current knowledge beyond the specificity that is known at pH 2.5.
4.2 Future Directions
All of the research presented here makes for a very good start at determining the
preferences of where pepsin will cleave. However, there is still more work that needs to
be done. In order to get a more solid perspective on the specificity of pepsin much more
142

data needs to be acquired. This research yielded a total of 1473 peptides generated from
58 proteins at pH 2.5. While this appears to be a lot of data, it is not yet enough for this
type of study.
Hundreds of proteins and tens of thousands of cleavages should be gathered in
order to verify the trends that were established with this work. However, while this
sounds like a good idea it would be rather difficult to actually complete. One way to
possibly gain this much data is to optimize digestion and chromatography conditions of
E.coli whole cell lysate. Only 49 proteins were identified during the E.coli digestion in
this research. If the conditions were optimized the digestion of E.coli could produce
hundreds of proteins, yielding the data set that is needed.
Another aspect of pepsin specificity that still needs to be investigated is how the
residues in positions P4 through P2 and P2’ through P4’ affect where pepsin will cleave.
This research just focused on the residues in the P1 and P1’ positions as they have the
most effect on pepsin specificity. However, from these studies there was the creation of a
cleavage database that incorporates out to the P4 and P4’ residues for the proteins
analyzed. This database will allow for further examination of what residues have an
effect on the specificity of pepsin.
4.3 References
Hamuro, Y., S. J. Coales, et al. (2008). "Specificity of immobilized porcine pepsin in
H/D exchange compatible conditions." Rapid Commun Mass Spectrom 22: 1041-
6.
143

Zhang, Z. (1995). Protein Hydrogen Exchange Determined by Mass Spectrometry: A
New Tool for Probing Protein High-order Structure and Structural Changes.,
Purdue. Ph.D.
144