development and evaluation of selected chemoprophylaxis
TRANSCRIPT

University of Arkansas, FayettevilleScholarWorks@UARK
Theses and Dissertations
5-2019
Development and Evaluation of SelectedChemoprophylaxis Candidates and a CandidateLive-Attenuated Vaccine for Prevention ofHistomoniasis in TurkeysLesleigh BeerUniversity of Arkansas, Fayetteville
Follow this and additional works at: https://scholarworks.uark.edu/etd
Part of the Animal Diseases Commons, and the Poultry or Avian Science Commons
This Thesis is brought to you for free and open access by ScholarWorks@UARK. It has been accepted for inclusion in Theses and Dissertations by anauthorized administrator of ScholarWorks@UARK. For more information, please contact [email protected].
Recommended CitationBeer, Lesleigh, "Development and Evaluation of Selected Chemoprophylaxis Candidates and a Candidate Live-Attenuated Vaccine forPrevention of Histomoniasis in Turkeys" (2019). Theses and Dissertations. 3147.https://scholarworks.uark.edu/etd/3147

Development and Evaluation of Selected Chemoprophylaxis Candidates and a Candidate Live-Attenuated Vaccine for Prevention of Histomoniasis in Turkeys
A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Poultry Science
by
Lesleigh Beer University of Arkansas
Bachelor of Science in Poultry Science, 2017
May 2019 University of Arkansas
____________________________________ Billy M. Hargis, Ph.D. Thesis Director ____________________________________ John R. Barta, Ph.D. Committee Member
____________________________________ Samuel J. Rochell, Ph.D. Committee Member
____________________________________ Guillermo Tellez, Ph.D. Ex-officio Member
____________________________________ Christine N. Vuong, Ph.D. Ex-officio Member

ABSTRACT
Histomoniasis, commonly known as blackhead disease, has increased in prevalence due
to the regulatory ban of prophylactics and therapeutics within the past 30 years. The objective of
this thesis was to evaluate the efficacy of selected dietary chemoprophylaxis candidates as well
as an in vitro live-attenuated vaccine candidate Histomonas meleagridis for prevention of
histomoniasis. Chapter one addresses deoxycholic acid and a biliogenic diet intended to
endogenously increase production of this secondary bile acid. Deoxycholic acid was effective in
vitro but failed to prevent histomoniasis when evaluated during the in vivo experimental disease
trial with turkeys. The biliogenic diet did not reduce disease prevalence. Chapter two addresses
dietary inclusion of 0.2% boric acid to prevent disease. The selected concentration of boric acid
was unsuccessful in disease prevention. Chapter three addresses the experiments conducted to
evaluate select ages, doses, and routes of a candidate live-attenuated vaccine. The live-attenuated
vaccine candidate has exhibited slight reduction in histomoniasis severity when administered
intracloacally on d14. Although the practicality of this current experimental vaccine
administration approach may be limited, further research must be conducted in order to further
elucidate conferred immune response and investigate the viability of this vaccine.

©2019 by Lesleigh Beer All Rights Reserved

ACKNOWLEDGEMENTS
My appreciation to everyone at the John Kirkpatrick Skeeles Poultry Health Laboratory
(aka U of A Poultry Health Lab) who has assisted and offered encouragement. Thank you to Dr.
Billy Hargis and my committee for their mentorship and encouragement to my research
development. I would especially like to thank my family for their continued love and support
throughout this academic endeavor.

DEDICATION
I would like to dedicate this thesis to my family and friends who have encouraged me
throughout this journey. A special dedication belongs to both my tax lady and adopted
grandmother who greatly encouraged me throughout my life. They watered the flower of
education but sadly did not get to see it bloom.

TABLE OF CONTENTS I. Introduction ..................................................................................................................................1
References ..............................................................................................................................2
II. Literature Review.........................................................................................................................4
Histomoniasis: A General Overview .....................................................................................4
Selected Chemoprophylaxis Compounds ..............................................................................8
Vaccination ............................................................................................................................18
References ..............................................................................................................................22
III. Data Chapter 1—Evaluation of deoxycholic acid as a prophylactic treatment to prevent
histomoniasis in turkeys .....................................................................................................................29
Abstract ..................................................................................................................................30
Introduction ............................................................................................................................32
Materials and Methods ...........................................................................................................33
Results ....................................................................................................................................35
Discussion ..............................................................................................................................37
References ..............................................................................................................................40
Tables .....................................................................................................................................42
Figures....................................................................................................................................44
IV. Data Chapter 2—Evaluation of boric acid as a chemoprophylaxis candidate to prevent
histomoniasis......................................................................................................................................47
Abstract ..................................................................................................................................48
Introduction ............................................................................................................................49
Materials and Methods ...........................................................................................................51

Results ....................................................................................................................................52
Discussion ..............................................................................................................................52
References ..............................................................................................................................54
Figures....................................................................................................................................56
V. Data Chapter 3—Evaluation of a candidate live-attenuated histomoniasis vaccine ....................57
Abstract ..................................................................................................................................58
Introduction ............................................................................................................................60
Materials and Methods ...........................................................................................................62
Results ....................................................................................................................................67
Discussion ..............................................................................................................................70
References ..............................................................................................................................74
Figures....................................................................................................................................76
VI. Conclusions ..................................................................................................................................86
Appendix ................................................................................................................................87

1
I. INTRODUCTION
Histomoniasis (also known as blackhead, infectious enterohepatitis, and histomonosis) is
an important disease of poultry, especially turkeys, which can be vectored by the Heterakis
gallinarum cecal worm, earthworm, or other birds such as chickens (Tyzzer and Fabyan, 1922;
van der Heijden et al., 2005; Hess et al., 2015). Caused by the protozoan parasite Histomonas
meleagridis, initial signs of disease often consist of declined feed consumption, drooping wings,
head-tucking, or inactivity (Duffy et al., 2005). Transmission occurs via cloacal drinking which
can rapidly transfer pathogens through the cloaca to the bursa of Fabricius or ceca via rhythmic
contractions that draw the material inside the vent (Hu et al., 2004; McDougald and Fuller,
2005).
Lack of research within the past 30 years, in addition to the ban of therapeutic and
prophylactic compounds, has resulted in lack of methods for controlling this disease (van der
Heijden et al., 2005; Hess et al., 2006). There is a need for evaluation of anti-protozoal
compounds (chemical and non-chemical) or vaccines for the control of histomoniasis and other
protozoal diseases (such as coccidiosis, caused by Eimeria spp.). Unfortunately, an alternative to
the previously used drugs has not yet been established, because in vitro and in vivo studies
continue to yield variable results against histomoniasis (Thøfner et al., 2012).
Previous immunological research results have been largely unsuccessful, creating doubts
for successful vaccine development (Hu and McDougald, 2004). However, in 1963, Joyner
treated H. meleagridis-infected turkeys with dimetridazole and reported recovered turkeys to be
resistant to subsequent infection, suggesting development of protective immunity. Turkeys
recovered from H. meleagridis infection and subsequently challenged to evaluate immunity have
exhibited resistance to characteristic disease symptoms and lesions even when harboring H.

2
meleagridis within the cecae (Cuckler, 1970). Intracloacal administration of in vitro-attenuated
H. meleagridis has resulted in reduced liver and cecal lesions within chickens and turkeys in
previous studies (Hess et al., 2008; Liebhart et al., 2013). In vitro passaging and subsequent
serial passaging of H. meleagridis within turkeys and chickens resulted in no reversion to
virulence (Sulejmanovic et al., 2013). Furthermore, cross-protection against heterologous
virulent isolates was demonstrated by vaccinating with an attenuated clonal strain of H.
meleagridis developed through prolonged in vitro culture methods (Sulejmanovic et al., 2016).
These successes encourage continued research in vaccine development as a solution against this
disease.
REFERENCES
Cuckler, A. 1970. Coccidiosis and histomoniasis in avian hosts. Pages 371-397 in Immunity to parasitic animals. GJ Jackson, R. Herman and I. Singer, eds., New York.
Duffy, C., M. Sims, and R. Power. 2005. Evaluation of dietary NatustatTM for control of
Histomonas meleagridis in male turkeys on infected litter. Avian Dis. 49:423–425. Van der Heijden, H. M. J. F., L. R. McDougald, and W. J. M. Landman. 2005. High yield of
parasites and prolonged in vitro culture of Histomonas meleagridis. Avian Pathol. 34:505–508.
Hess, M., T. Kolbe, E. Grabensteiner, and H. Prosl. 2006. Clonal cultures of Histomonas
meleagridis, Tetratrichomonas gallinarum and a Blastocystis sp. established through micromanipulation. Parasitology. 133:547–554.
Hess, M., D. Liebhart, I. Bilic, and P. Ganas. 2015. Histomonas meleagridis—new insights into
an old pathogen. Vet. Parasitol. 208:67–76. Hess, M., D. Liebhart, E. Grabensteiner, and A. Singh. 2008. Cloned Histomonas meleagridis
passaged in vitro resulted in reduced pathogenicity and is capable of protecting turkeys from histomonosis. Vaccine. 26:4187–4193.
Hu, J., L. Fuller, and L. R. McDougald. 2004. Infection of turkeys with Histomonas meleagridis
by the cloacal drop method. Avian Dis. 48:746–750.

3
Hu, J., and L. McDougald. 2004. The efficacy of some drugs with known antiprotozoal activity against Histomonas meleagridis in chickens. Vet. Parasitol. 121:233–238.
Liebhart, D., T. Sulejmanovic, B. Grafl, A. Tichy, and M. Hess. 2013. Vaccination against
histomonosis prevents a drop in egg production in layers following challenge. Avian Pathol. 42:79–84.
McDougald, L., and L. Fuller. 2005. Blackhead disease in turkeys: Direct transmission of
Histomonas meleagridis from bird to bird in a laboratory model. Avian Dis. 49:328–331. Sulejmanovic, T., I. Bilic, M. Hess, and D. Liebhart. 2016. An in vitro attenuated strain of
Histomonas meleagridis provides cross-protective immunity in turkeys against heterologous virulent isolates. Avian Pathol. 45:46–53.
Sulejmanovic, T., D. Liebhart, and M. Hess. 2013. In vitro attenuated Histomonas meleagridis
does not revert to virulence, following serial in vivo passages in turkeys or chickens. Vaccine. 31:5443–5450.
Thøfner, I. C. N., D. Liebhart, M. Hess, T. W. Schou, C. Hess, E. Ivarsen, X. Fretté, L. P.
Christensen, K. Grevsen, R. M. Engberg, and others. 2012. Antihistomonal effects of artemisinin and Artemisia annua extracts in vitro could not be confirmed by in vivo experiments in turkeys and chickens. Avian Pathol. 41:487–496.

4
II. LITERATURE REVIEW
The ban of nitroimidazoles, nitrofurans, and arsenical compounds for prophylaxis and
treatment in the last 30 years has led to the lack of preventative options for histomoniasis, a
disease primarily affecting turkeys. The following summarizes the disease, chemoprophylaxis
compounds focused on the mitigation of this problem, and previous immunization attempts. In
addition, an overview of the bile acid pathway and dietary inclusions to increase endogenous
production of deoxycholic acid will be summarized, because this compound was selected as an
anti-histomonal candidate.
HISTOMONIASIS: A GENERAL OVERVIEW
Histomoniasis (also known as blackhead, infectious enterohepatitis, and histomonosis) is
a protozoal disease affecting the gastrointestinal tract of turkeys, chickens, and other
gallinaceous birds (van der Heijden et al., 2005; Hess et al., 2015). High mortality occurs in
turkeys, whereas less severe damage occurs in chickens and other galliformes (Callait et al.,
2002). Considered critically and economically impactful to both turkeys and chickens,
histomoniasis is a serious and ongoing concern facing the poultry industry (Duffy et al., 2005;
Lotfi et al., 2014).
Histomonas meleagridis is included in the phylum Sarcomastigophora with further
taxonomic classification in the order Tritrichomonadida and family Dientamoebidae (Cepicka et
al., 2010; Hess and McDougald, 2013). The etiological agent of histomoniasis, H. meleagridis
penetrates the cecal epithelial lining, replicates, enters the bloodstream and parasitizes the liver
(Clarkson, 1963; Hess and McDougald, 2013). Hepatic liver lesions and tissue necrosis are
common within diagnosed birds, while individual histomonads can be observed within infected
tissues using electron microscopy. Cultivation of H. meleagridis has occurred from liver lesions

5
(Bayon and Bishop, 1937; Bishop, 1938). Overt signs of histomoniasis are often apparent at
eleven days post-infection while thickening and reddening of the mucosal layer of the ceca
begins within three days of infection (Hess and McDougald, 2013). Although commonly referred
to as blackhead within industry and laymen’s terms, the clinical sign of cyanosis of the head are
neither pathognomonic nor distinctive to histomoniasis because other diseases produce a similar
appearance (Sentíes-Cué et al., 2009; Hess and McDougald, 2013). Initial symptoms can include
declining feed consumption, drooping wings, unkempt feathers, or inactivity (Duffy et al., 2005).
Necrotic typhlitis (necrosis and inflammation of the cecae), hepatitis, sulfuric excreta, and high
mortality are characteristic pathological signs (Callait et al., 2002; Hess et al., 2015).
Severe mortality results in turkeys with an estimated annual loss of over two million
dollars and mortality approaching 80-100% in an affected flock (Callait et al., 2002; McDougald,
2005; Hess and McDougald, 2013). Although mortality is not as significant in chickens,
economic losses are estimated to exceed those of turkeys due to greater frequency of disease and
the larger number of flock infections within chickens (Callait et al., 2002). Chicken mortality can
be 10-20% with a high morbidity but an outbreak frequently is unnoticed or results in increased
condemnations at the broiler processing plant (McDougald, 2005). Mortality is most commonly
observed between nine and twelve days post-infection (Hess and McDougald, 2013).
Beginning in the 1960s, research on this organism waned because the introduction of
nitroimidazoles, nitrofurans, and arsenical compounds for prophylactic treatment against
outbreaks was made available and successfully controlled the disease (van der Heijden et al.,
2005; Hess et al., 2006). Concerns over heavy metal retention within meat products from treated
poultry resulted in the 2015 regulatory removal of Nitarsone (Histostat), the last remaining FDA-
approved drug for treatment of histomoniasis (Regmi et al., 2016). Without viable substitute

6
treatment options and no new alternatives, producers are now suffering from the losses of broiler
breeders, layer pullets, and turkeys (Hu and McDougald, 2004).
Pathogenesis
The trichomonad parasite H. meleagridis is capable of existing as either an amoeboid or
single-flagellated form, and can be transmitted directly from bird to bird or through intermediate
hosts and vectors (Lotfi et al., 2014). Lacking mitochondria, H. meleagridis possesses an
organelle called the hydrogenosome responsible for anaerobic energy metabolism (McDougald,
2005; Hauck et al., 2010; Hess and McDougald, 2013). These protozoa have a typical median
diameter of approximately 10µm, ranging from 6-20µm, and usually exist in amoeboid form but
may also exist with a single flagellum (Hess and McDougald, 2013). Recent research has
suggested the possible formation of cyst-like stages in vitro observed with detailed light and
transmission microscopy (Zaragatzki et al., 2010a; b). Isolates can be cultured from infected
birds and grown in vitro, with long-term cultivation shown to result in a different phenotype,
increased growth, and higher tenacity in adverse conditions (Gruber et al., 2017). A larger,
nonpathogenic species, Histomonas wenreichi or Parahistomonas wenrichi, was later discovered
and has a typical size of 20-30µm and 3-4 flagella (McDougald, 2005).
Although the life cycle of H. meleagridis is not fully understood, it has been
characterized as an extracellular parasite that reproduces through binary fission (Tyzzer, 1920;
Cuckler, 1970). Direct oral ingestion of histomonads has not recreated the disease reliably in
previous research, presumably due to the adverse acidity and mechanical action within the crop
and ventriculus (gizzard) of the bird (Hu et al., 2004). Histomonads are delicate in nature and
cannot withstand living long periods of time in the environment unless protected by a vector
such as the heterakid worm or earthworm (McDougald and Fuller, 2005). The cecal worm

7
Heterakis gallinarum has been identified as an important vector and reservoir for disease
transmission; H. meleagridis have been observed within the intestinal wall, larvae, and eggs of
this cecal nematode (Hess et al., 2015). Heterakis gallinarum appears to provide a protective
barrier to transport the infective material through the gastrointestinal tract of poultry, leading to
establishment of disease (Tyzzer and Fabyan, 1922; Hu et al., 2004). Ingestion of embryonated
heterakid eggs or the adult cecal worms by poultry can result in H. meleagridis infection (Hu et
al., 2004). Regular earthworms can also ingest histomonad-infected cecal worm eggs, and
thereby act as a transport host for the infected heterakid eggs (Lund et al., 1966; Hu et al., 2004).
Other protozoa and nematodes have similar transmission methods; the Enterobius vermicularis
nematode can serve as a vector of Dientamoeba fragilis, an intestinal trichomonad parasite
primarily of humans, facilitating transmission of disease (Clark et al., 2014).
Rapid transmission by direct contact between infected birds or fecal droppings can occur
without an intermediate host or vector if transmission occurs before the fragile histomonads die
(Sentíes-Cué et al., 2009). Once a turkey is infected, disease can quickly transmit to others in the
flock within two or three days, causing an outbreak (Hess and McDougald, 2013). Interestingly,
this method of transmission occurs primarily through the cloacal drinking phenomenon rather
than standard fecal-oral or respiratory-respiratory transmission. This cloacal drop or cloacal
drinking can rapidly transfer disease through the cloaca to the bursa of Fabricius or ceca via
rhythmic contractions that draw the infectious material inside the vent (Hu et al., 2004;
McDougald and Fuller, 2005). Infection has been produced with the cloacal injection of infected
tissue (liver or cecae) as well as with a suspension of histomonad culture (Berks and Neal, 1952).
Recently, oral administration of a clonal in vitro-cultured H. meleagridis to 1-day-old turkeys
followed by 5h feed restriction resulted in histomoniasis mortalities, suggesting a potential oral

8
infection route if large amounts of contaminated excreta or litter are ingested (Liebhart and Hess,
2009).
To further complicate the understanding of this disease, commensal bacterial flora have
been found to be important in disease development (McDougald, 2005). Unfortunately, the cecal
environment wherein H. meleagridis resides is constantly in a state of flux, making permanent
microflora adjustments as a method of controlling this disease difficult (Callait et al., 2002). The
nitroimidazole-class drug dimetridazole was previously an effective treatment for histomoniasis;
however, the ban of effective prophylactic and therapeutic compounds such as this have resulted
in measures focused upon prevention rather than treatment of the disease (Joyner, 1963; Hess
and McDougald, 2013; Liebhart et al., 2013). In 2015, the arsenic-based drug nitarsone
(Histostat), the last remaining FDA-approved drug for prevention of histomoniasis, was
withdrawn from the market due to concerns about inorganic arsenic levels in treated poultry
(Regmi et al., 2016). Overall effectiveness is limited, but outbreaks can be reduced with careful
animal management practices (Liebhart et al., 2017). The disease is generally more prevalent
when birds are housed in environments favoring the coexistence of the cecal nematode H.
gallinarum, the primary local reservoir for infection (Hess and McDougald, 2013). Chickens
often serve as reservoirs for the cecal worm and pose as sources for potential infection; therefore,
separate rearing should occur between poultry species to prevent resulting cross-transmission
outbreaks (Bayon and Bishop, 1937; McDougald, 2005; Hess and McDougald, 2013).
SELECTED CHEMOPROPHYLAXIS COMPOUNDS
Anti-histomonal drugs, plant-derived compounds, and vaccination attempts have been
reviewed by Liebhart et al. (2017). Recent research has focused primarily in the areas of
antiprotozoal compounds, antibiotics, vaccines, and plant products with chemical activity (Hess

9
et al., 2015). Treatment is challenging because protozoa are eukaryotic organisms; selective
toxicity is crucial so that the chemoprophylaxis compound is harmful to the parasite without
causing irreversible damage to the host. With the increasing world population, finding methods
to prevent and treat diseases such as histomoniasis are crucial for the overall well-being of
animal husbandry and food production. Additionally, the implications of discovering anti-
protozoal compounds that could have beneficial applications towards other protozoal diseases
are remarkable and warrant further research for the potential benefit of animals and humans
alike. However, identifying alternatives to the previously used drugs has not yet been
established, because in vitro and in vivo studies continue to yield variable results against
histomoniasis (Thøfner et al., 2012). The close relationship of H. meleagridis to other amoebae
and flagellates suggests any anti-protozoal compounds effective against H. meleagridis may also
have activity against related protozoal species (Hu and McDougald, 2004). If a compound were
discovered, this could revolutionize the potential for efficacious treatment of other protozoa such
as Tetratrichomonas and Entamoeba (Hess et al., 2006; Nakada-Tsukui and Nozaki, 2016).
Deoxycholic acid
Deoxycholic acid (DCA), a naturally occurring secondary bile acid, has been shown to
reduce severity of Eimeria maxima and Clostridium perfringens infections in poultry when
administered at a concentration of 1.5 g/kg in the diet by reducing intestinal villi damage (Wang
et al., 2018). A study in mice resulted in anaerobic bacteria-derived DCA protection against
colitis induced by Campylobacter jejuni (Sun et al., 2018). Considering these results, the
hypothesis that DCA might confer anti-histomonal properties is investigated in the first chapter.
Overview of bile acids. Within the small intestine, the homeostatic control of cholesterol
and the absorption of dietary fatty acids are reliant upon biliary lipid secretions that include bile

10
acids (LeBlanc et al., 1998). Cholic acid (CA) and chenodeoxycholic acid (CDCA) are the
primary bile acids (Stamp and Jenkins, 2008). Cholesterol is an important precursor for bile
acids, while bilirubin, heavy metals (such as copper and iron), and urine insoluble lipophilic
steroids are constituent components of bile acids (Stamp and Jenkins, 2008; Boyer, 2013). Liver
hepatocytes synthesize bile acids, which are water-soluble amphipathic molecules resulting from
cholesterol catabolism (Stamp and Jenkins, 2008; Winston and Theriot, 2016). According to
Lefebvre et al., (2009), the “classical pathway” for bile acid synthesis uses cholesterol 7α-
hydroxylase (CYP7A1) and contributes to approximately 75% of bile acid synthesis within the
liver. However, the alternative, “acidic pathway” initiated by sterol 27-hydroxylase (CYP27A1)
is responsible for the remaining 25% of the bile acid synthesis (Lefebvre et al., 2009). The
gallbladder functions as the storage organ for bile acids and approximately 5% of these stored
bile acids will be fecally excreted (Stamp and Jenkins, 2008). Additionally, cholecystokinin
released from the duodenum following a meal initiates gallbladder contraction, stimulating the
flow of bile (Ridlon et al., 2006; Lefebvre et al., 2009). A vast canalicular network comprised
from apical membranes of hepatocytes and bile duct cholangiocytes results in the bile-secretory
unit (Boyer, 2013).
Bile acids contribute to multiple important physiological functions and pathways
including: 1) the excretory pathway to clear harmful exogenous substrates that are not readily
processed by the kidney; 2) bile salts, which are organic solute components in bile acid that
emulsify and facilitate the digestion of fats; 3) the cholesterol elimination pathway; 4) the
excretion of IgA, inflammatory cytokines, and innate immune system stimulation; 5)
enterohepatic circulation component; and 6) excretion mechanisms for intestinal growth
hormones (Boyer, 2013). Bile acids are cytotoxic at high concentrations, and can lead to

11
carcinogenesis within some visceral organs and tissues (Stamp and Jenkins, 2008; Lefebvre et
al., 2009). The hydrophobicity of the bile acids is potentially linked to the toxicity of the
molecules, and the increased number of hydroxyl groups is inversely related to toxicity (Stamp
and Jenkins, 2008). However, bile acids function as biological detergents, aiding in the
production of antimicrobial peptides and contributing to host defense against pathogens
(Winston and Theriot, 2016).
Bile acid conjugation. Bile acids are commonly referred to as bile salts, although this
may be a more appropriate term when the conjugated bile acids are linked with sodium or
potassium salts. Largely impermeable to the intestinal enterocytes, conjugated bile acids are
usually restricted to the gastrointestinal lumen due to increased aqueous solubility, ensuring the
likelihood of distribution with digesta (Stamp and Jenkins, 2008). The increased levels of bile
acids within the lumen initiate micelle formation that contributes to the emulsification and
absorption of lipids (Lefebvre et al., 2009). The micelle aids in the diffusion of lipids as they
travel the length of the gastrointestinal tract, while also contributing to the reabsorption of bile
acids within the distal ileum (Stamp and Jenkins, 2008).
Glycine-conjugated bile acids are the most abundant and represent greater than 70% of
bile (Stamp and Jenkins, 2008). The conjugated form is denied entry to epithelial cells thereby
protecting the organs and restricting its presence to within the gastrointestinal lumen (Stamp and
Jenkins, 2008). Taurine-conjugated bile acids are present in lower abundance but contribute to
approximately 20% of bile (Stamp and Jenkins, 2008). Conjugation of CA or CDCA to either
glycine or taurine is common; conjugated bile acids are excreted into the lumen of the
gastrointestinal tract to facilitate metabolism of fats and fat-soluble vitamins (Van Eldere et al.,
1996). Passive reabsorption is prevented due to the stronger acidic nature produced from

12
conjugation as the bile acids travel through the biliary ducts (Boyer, 2013). The taurine
conjugated bile acids have detergent properties and are soluble within the normal acidic stomach
environment (Stamp and Jenkins, 2008). This solubility contributes to its potential entry into the
gastrointestinal epithelial cells. Not surprisingly, taurine supplementation has be shown to
negatively impact intestinal mucosal development through its role in toxic bile acid generation
within broiler chickens (Huang et al., 2014).
Free bile acids. Free bile acids are produced via bacterial enzymes that deconjugate
primary bile acids within the ileum and the large intestine (Cole and Fuller, 1984). Moreover,
bacterial hydroxysteroid dehydrogenases further convert free bile acids into secondary bile acids
via a 7α-dehydroxylase reaction. Other enzymatic reactions may occur through 3α- and 7α-
hydroxysteroid dehydrogenases as well, but the 7α-dehydroxylation is unique to a few anaerobic
species (Winston and Theriot, 2016). Deconjugation reactions within the small intestine and
dihydroxylation reactions within the large intestine are the two predominant methods by which
secondary bile acids are produced (Winston and Theriot, 2016). Moreover, reconjugation with
glycine or taurine can occur, renewing levels of primary and secondary bile acids during
enterohepatic recirculation.
Within the lower gastrointestinal tract, the secondary bile acids formed include DCA and
lithocholic acids (LCA) that are produced from CA and CDCA, respectively (Van Eldere et al.,
1996; Ridlon et al., 2006). Many other bile acids may also be formed via bacterial enzymatic
reactions and conjugations, but the above mentioned are the more common bile acids (Stamp and
Jenkins, 2008). Increased secondary bile acids have been associated with the etiologies of
cholesterol gallstone disease and colon carcinogenesis (Winston and Theriot, 2016). Considering
this impact against eukaryotic cells, DCA was considered to potentially exhibit toxicity to H.

13
meleagridis. High plasma cholesterol levels, particularly low-density lipoprotein cholesterols, are
associated with increased risks of cardiovascular disease (Yang et al., 2012). An inverse
relationship appears to exist for CDCA and DCA, meaning that the increasing DCA
concentrations are linked with decreasing CDCA (Ridlon et al., 2006). This is understandable in
that DCA is formed from the catabolism of CDCA.
Enterohepatic recirculation. The formation of secondary bile acids is important to the
enterohepatic circulation that can occur as often as ten times a day. Both primary (CA and
CDCA) and secondary (LCA and DCA) bile acids are circulated and reabsorbed through the
enterohepatic system (Stamp and Jenkins, 2008). This reabsorption process occurs when DCA
and LCA enter the portal veins and circulate to the liver to join the newly synthesized CA and
CDCA. Reconjugation to glycine or taurine occurs and these renewed, conjugated bile acids are
stored in the gallbladder. The rate-limiting CYP7A1 is suppressed when bile acids are returned
to the liver via the enterohepatic circulation, thereby inhibiting additional bile acid synthesis
(Stamp and Jenkins, 2008). The actual regulation of synthesis, metabolism, and transportation of
bile acids via the enterohepatic system was determined to be self-regulated by the bile acids
through activation of the Farnesoid X receptor (FXR) (Lefebvre et al., 2009). The FXR is
expressed within the liver, gastrointestinal tract, kidneys, and adrenal glands. The recirculated
bile acids bind to the FXR in the liver, resulting in negative feedback and downregulation of
transcription factors for bile acid synthesis (Stamp and Jenkins, 2008).
If DCA were capable of conferring anti-histomonal properties in vivo, use as a
commercial feed additive might be more accepted since DCA is naturally occurring and is
regenerated during enterohepatic recirculation. Moreover, certain diets have been shown to
increase bile acid output that indicates the endogenous processes can be altered by dietary

14
composition. If DCA effectively mitigates histomoniasis, an alternative to DCA inclusion in diet
would be to pursue formulation of a bile-enhancing “biliogenic” diet. This different approach
might be more economical than directly supplementing DCA into the feed. The literature
considered for formulation of this biliogenic diet is reviewed below.
High protein dietary impacts. Within chickens, the atherogenic impact (formation of
fatty plaques in the arteries) of dietary cholesterols and fats was shown to decline with the
introduction of higher protein diets (Nishida et al., 1958; Kummerow et al., 1960). Diets high in
fat are generally associated with the elevated cholesterol levels that are a precursor to bile acids
(Kokatnur et al., 1958b). An inverse linear relationship between protein intake and levels of
serum cholesterol was caused in experiments with high protein diets (Kokatnur et al., 1958b).
Interestingly, in a study conducted by Kokatnur et al. (1958b), hypercholesteremic birds that
were provided adequate or high protein diets showed a rapid decrease in serum cholesterol.
Moreover, serum cholesterol values only reached normal levels within hypercholesteremic birds
when supplemented with high dietary protein. These observations suggest that increased dietary
protein intake might be a more efficient means of reducing overall cholesterol levels in the serum
than simply eliminating or decreasing fat consumption (Kokatnur et al., 1958b).
Furthermore, dietary methionine deficiencies were found to increase serum cholesterol
(by nearly two-fold as compared to a low protein choline deficient diet), β-lipoprotein levels, and
total lipids within the sera of chicks (Kokatnur et al., 1958a; Nishida et al., 1958). These results
suggest the artherogenic effects of imbalanced cholesterol, lipid, and methionine when combined
with low protein diets (Nishida et al., 1958). Taken together, the apparent connection between
dietary protein and artherogenic effects suggest the importance of nutritional management of
dietary proteins rather than dietary fats.

15
The additional role of energy to protein ratio (E/P ratio) was considered by Kokatnur et
al., (1958a). The E/P ratio was calculated by the following equation:
100 .
Low E/P ratios were associated with significantly decreased serum cholesterol levels, presenting
an additional factor to consider when determining dietary effects (Kokatnur et al., 1958a).
Egg-enriched diets. Yang et al. (2012), conducted a study with various egg-enriched
diets to evaluate influences upon cholesterol metabolism, among other factors, in rats. Of
interest, a diet consisting of 31.25% vacuum freeze-dried egg yolk powder diet (21.78% crude
protein) and a diet consisting of 55.56% vacuum freeze-dried whole egg powder (36.09% crude
protein) were included in the study. These diets were compared to control groups provided either
a standard diet or a 0.75% cholesterol diet. Total bile acid output was higher in all groups as
compared to the controls and was particularly significant for the whole egg powder diet. Liver
cholesterol levels within both egg-enriched diets were higher than the control, although the
0.75% cholesterol diet exhibited the highest level (Yang et al., 2012). However, this indicates the
influence upon hepatic cholesterol levels within egg-enriched diets.
CYP7A1, the rate-limiting enzyme for bile acid synthesis from cholesterol, had increased
mRNA expression within the egg-yolk and whole-egg powder enriched diets as compared to
both the cholesterol and control diets (Yang et al., 2012). This increased mRNA expression
indicates that the egg-enriched diets upregulated the CYP7A1 activity, thereby promoting bile
acid synthesis. Moreover, this increased function may have contributed to the elevated output of
bile acids within the excreta (Yang et al., 2012). Taken together, the data from this study suggest
that egg-enriched diets are consistent with the lowering of cholesterol absorption as well as the
increased excretion of bile acids due to increased rate of synthesis.

16
Lecithin-enriched diets. Lecithin-enriched diets have been shown to increase biliary
excretions in both rats and chickens (Lindsay et al., 1969; LeBlanc et al., 1998). In 1969,
Lindsay et al. reported an 81% increase in bile acid excretion based upon fecal matter, and a
105% increase based upon bird body weight when chicks were fed a 2.5% lecithin diet. A 0.5%
β-sitosterol diet was also found to increase bile excretions, but to a lesser extent than the lecithin
diet. Synergistic effects resulted from the combination of the 2.5% lecithin and 0.5% β-sitosterol
diets, producing higher bile acid excretion than either diet singly (Lindsay et al., 1969)
LeBlanc et al. (1998) fed rats a 20% lecithin diet that resulted in a 25% increase in bile
flow and 61% increase in biliary bile acid output. Interestingly, the lecithin diet also contributed
to a proportional change among the bile acid composition, with DCA significantly increasing
(LeBlanc et al., 1998). This changed profile could explain why the cholesterol output was also
increased.
Considering these factors, Data Chapter 1 evaluates different concentrations of DCA
inclusion within a basal diet as well as a diet formulated with the intention of endogenously
increasing bile acid synthesis. The above data contributed to our hypothesis that DCA might be
effective as a chemoprophylaxis chemical against histomoniasis. However, the complicated
nature of histomoniasis serves to further emphasize the importance of pairing in vivo studies with
in vitro experiments to confirm that any chemoprophylaxis activity is conferred within an
animal.
Boric acid
Although boron is an essential element to humans, animals, and plants, the NRC (1994)
has no recommended level of boron for daily intake in poultry (Eren et al., 2012). Not naturally
occurring in elemental form, boron is always bound chemically with oxygen to form borates

17
(Moore et al., 1997). Boric acid, a boron compound, is used as a litter treatment for the
prevention of darkling beetles within the poultry industry, raising concerns that poultry might be
harmed by the ingestion of the boric acid litter treatment (Sander et al., 1991; Dufour et al.,
1992). Previously, boric acid was shown to be non-toxic when administered orally to 1-day-old
chicks at a dose of less than 2g/kg body weight, but levels greater than 3.89 g/kg body weight
resulted in high toxicity (Sander et al., 1991). The acute oral mean lethal dose of boric acid in 1-
day-old chicks was later determined to be 2.95 ± 0.35 g/kg of body weight, resulting in its
classification as a slightly toxic chemical (Sander et al., 1991). Toxic levels of boric acid can
result in decreased body weight, increased feed conversion, and abnormal feather growth
(Dufour et al., 1992). However, in 1992, Dufour et al. demonstrated that litter treatment with
boric acid at a rate of 0.4-0.9kg/9.3m2 did not significantly increase feed conversion rate or
decrease body weight. Previous research has suggested boron may have an important biological
role in biochemical mechanisms influencing normal growth and mineral metabolism (Kurtoğlu et
al., 2005; Çinar et al., 2015). Beginning at day-of-hatch until d21, dietary concentrations up to
240 ppm (0.024%) boron were not detrimental to broiler performance, although boron levels
within breast muscle and liver tissues increased proportionately with increased dietary
concentration (Rossi et al., 1993). Moreover, boron supplementation at 20mg/kg in the basal diet
had no impact on body weight or feed consumption in chickens; results did not suggest growth-
promotion or metabolic mineral regulation (Küçükyilmaz et al., 2017).
Boric acid has antifungal, antiseptic and antiviral properties, and it has been used as an
antifungal agent in the treatment of yeast infection (Hernandez-Patlan et al., 2018a). Boric acid
has been considered as a prophylactic measure against Saprolegnia fungal infections in Atlantic
salmon with high hatchability and survival rates following treatment (Ali et al., 2014).

18
Brittingham and Wilson, (2014) showed that growth rate of Trichomonas vaginalis, the
protozoan causative agent of trichomoniasis in humans, was reduced with low boric acid
concentrations (0.2%) and exhibited lethality to trichomonads at higher concentrations (≤0.4%),
independent of environmental acidification. In an in vitro gastrointestinal model, boric acid
appeared to decrease concentration of Salmonella Enteritidis within the intestinal compartment
(Hernandez-Patlan et al., 2018a). However, a concentration of 0.1%, boric acid within the basal
diet had no significant reduction in Salmonella Enteritidis during an in vivo study (Hernandez-
Patlan et al., 2018b). Dietary supplementation of boron is considered economical in that a
100mg/kg diet was estimated to cost 0.5 US $ per one ton of prepared feed (Bozkurt and
Kucukyilmaz, 2015). As bacterial flora are an important factor in development of histomoniasis
development, the reduction of bacteria in vitro in addition to the antifungal properties exhibited
by boric acid contributed to the interest in evaluating this compound for anti-histomonal
properties (Hernandez-Patlan et al., 2018a). Considering these experiments, we hypothesized that
boric acid might be efficacious against the trichomonad parasite H. meleagridis at the selected
dietary concentration of 0.2%; this hypothesis is addressed in Data Chapter 2.
VACCINATION
Vaccinations are important for the induction of a host-immune response to protect against
disease (Mitra et al., 2018). Previous immunological research results with histomoniasis have
largely been unsuccessful; therefore, the likelihood of vaccine development for prevention of this
disease has been considered unlikely (Hu and McDougald, 2004). However, turkeys recovered
from histomoniasis and subsequently challenged to evaluate immunity have exhibited resistance
to characteristic clinical signs and lesions even when harboring H. meleagridis within the cecae,
suggesting the presence of immune response (Cuckler, 1970). Moreover, H. meleagridis-infected

19
turkeys rescued with dimetridazole were shown to be resistant to subsequent infection,
suggesting an acquired protective immunity (Joyner, 1963).
In the early 1900s, intravenous inoculation was found to eventually reproduce
histomoniasis when injection was repeated (Tyzzer et al., 1921). Oral administration of fresh
subcutaneous, liver, and lung lesions did not reproduce histomoniasis within turkeys (Tyzzer et
al., 1921). Tyzzer (1921) stated that turkeys recovered from H. meleagridis infection were not
able to be re-infected, suggesting immunity. Furthermore, histomoniasis resistance increased
with age with only a few mature birds exhibiting clinical signs of infection (Tyzzer et al., 1921).
However, turkeys recovered from histomoniasis only exhibited a degree of immunity that
appeared transient rather than permanent due to the reappearance of clinically apparent disease
several months following recovery from disease challenge (Tyzzer and Fabyan, 1922).
Passive immunization with injection of antiserum from immune into susceptible poultry
did not confer protection against histomoniasis upon challenge (Clarkson, 1963; Bleyen et al.,
2009). Intramuscular injection of an inactivated clonal H. meleagridis also failed to produce
effective protection (Hess et al., 2008). These data suggest acquired immune protection is largely
cell-mediated rather than humoral (antibody-based). Previous studies with chickens have shown
that feed deprivation and an alkaline pH prior to oral challenge were required in order to develop
lesions consistent with histomoniasis, suggesting that oral transfer of the parasite should not be
overlooked (Cuckler, 1970). Liebhart et al. (2010) demonstrated a protective effect of an in vitro
attenuated H. meleagridis administered orally to 1-day-old turkeys, further suggesting that oral
transfer should not be disregarded. A rectally inoculated nonpathogenic strain of Histomonas
was shown to afford some protection against challenge with pathogenic histomonads three to six
weeks post-vaccination, but the effectiveness declined when challenge occurred via infected

20
heterakid eggs (Lund, 1959). In regards to this outcome, Lund (1959) suggested that the immune
barrier was limited to the cecal mucosa, which could be infiltrated by the cecal worms, thus
allowing disease development. Reduced albumin and increased γ–globulin are the primary serum
protein changes associated with H. meleagridis infection (Clarkson, 1966). Recently, vaccination
with attenuated histomonads reduced T and B cell subset deviation; mortality in turkeys
suffering from histomoniasis was demonstrated to be associated with higher cellular immune
response when compared to chickens (Mitra et al., 2017). Co-infection of H. gallinarum with H.
meleagridis resulted in an increased mRNA expression of Th1 cytokine IFN-γ (Schwarz et al.,
2011).
Attenuated H. meleagridis
Following propagation for two years, an H. meleagridis culture originally pathogenic to
chickens was found to have lost pathogenicity and induce protection against pathogenic strains
when allowed to multiply within the chicken’s cecae (Tyzzer, 1932). Early studies by Tyzzer
(1934) reported reduced virulence of H. meleagridis that was cultivated in vitro, although
immunization attempts yielded conflicting success. Histomonads passaged in vitro more than
1000 times over a period of seven years were shown to be nonpathogenic and considered to have
lost efficacy as an immunizing strain capable of protecting against pathogenic H. meleagridis
strains (Lund and Chute, 1967). Stable attenuation has been shown in a H. meleagridis that was
passaged 295 times in vitro and subsequently serially passaged in vivo within turkeys and
chickens with no reversion to virulence (Sulejmanovic et al., 2013). More recent studies within
chickens and turkeys have shown reduction of liver and cecal lesions following intracloacal
administration of clonal in vitro attenuated H. meleagridis utilized as a vaccine strain (Hess et
al., 2008; Liebhart et al., 2013). Nguyen Pham et al. (2013) cloacally inoculated turkeys with a

21
low-virulence H. meleagridis strain that was obtained via serial passage in turkeys and showed
induced protection in the face of subsequent challenge by a virulent H. meleagridis. Furthermore,
an attenuated clonal strain of H. meleagridis developed through prolonged in vitro culture
methods demonstrated a cross-protective capability against heterologous virulent isolates
(Sulejmanovic et al., 2016). Pullets vaccinated at 18-week-of-age with an in vitro-attenuated,
clonal culture exhibited reduced pathology and prevention of a severe drop in egg production as
compared to pullets challenged without prior vaccination (Liebhart et al., 2013). Taken together,
these data suggest that a protective immune response against histomoniasis may be induced.
Our laboratory obtained a virulent field isolate of H. meleagridis that was able to be
cultivated and preserved based upon previous methods (van der Heijden et al., 2005; van der
Heijden and Landman, 2007). This virulent H. meleagridis isolate was propagated for ~80
passages and evaluated as a live-attenuated vaccine candidate. Based upon the rapid in vitro
growth and perceived virulence reduction within turkeys, we believed that it may be a worthy
vaccine candidate, a topic that will be further explored in Data Chapter 3 within a disease
challenge study.

22
REFERENCES
Ali, S. E., E. Thoen, O. Evensen, and I. Skaar. 2014. Boric acid inhibits germination and colonization of Saprolegnia spores in vitro and in vivo. PLoS ONE. 9:e91878.
Bayon, H., and A. Bishop. 1937. Cultivation of Histomonas meleagridis from the liver lesions of a hen. Nature. 139:370-371.
Berks, G., and R. Neal. 1952. The effect of some drugs upon Histomonas meleagridis in vitro. Ann. Trop. Med. Parasitol. 46:68–71.
Bishop, A. 1938. Histomonas meleagridis in domestic fowls (Gallus gallus). Cultivation and experimental infection. Parasitology. 30:181–194.
Bleyen, N., E. Ons, M. De Gussem, and B. M. Goddeeris. 2009. Passive immunization against Histomonas meleagridis does not protect turkeys from an experimental infection. Avian Pathol. 38:71–76.
Boyer, J. L. 2013. Bile formation and secretion. Compr. Physiol. 3:1035–1078.
Bozkurt, M., and K. Kucukyilmaz. 2015. The role of boron in poultry nutrition Part II: Compositional and mechanical properties of bone and egg quality. Worlds Poult. Sci. J. 71:483–492.
Brittingham, A., and W. A. Wilson. 2014. The antimicrobial effect of boric acid on Trichomonas vaginalis. Sex. Transm. Dis. 41:718–722.
Callait, M., C. Granier, C. Chauve, and L. Zenner. 2002. In vitro activity of therapeutic drugs against Histomonas meleagridis (Smith, 1895). Poult. Sci. 81:1122–1127.
Cepicka, I., Hampl, V., Kulda, J., 2010. Critical taxonomic revision of parabasalids with
description of one new genus and three new species. Protist. 161: 400–433.
Çinar, M., K. Küçükyilmaz, M. Bozkurt, A. Çatli, E. Bintaş, H. Akşit, R. Konak, Ç. Yamaner, and K. Seyrek. 2015. Effects of dietary boron and phytase supplementation on growth performance and mineral profile of broiler chickens fed on diets adequate or deficient in calcium and phosphorus. Br. Poult. Sci. 56:576–589.
Clark, C. G., D. Röser, and C. R. Stensvold. 2014. Transmission of Dientamoeba fragilis:
Pinworm or cysts?. Trends Parasitol. 30: 136-140. Clarkson, M. 1963. Immunological responses to Histomonas meleagridis in the turkey and fowl.
Immunology. 6:156-168.

23
Clarkson, M. 1966. Progressive serum protein changes in turkeys infected with Histomonas meleagridis. J. Comp. Pathol. 76:387–IN9.
Cole, C., and R. Fuller. 1984. Bile acid deconjugation and attachment of chicken gut bacteria: their possible role in growth depression. Br. Poult. Sci. 25:227–231.
Cuckler, A. 1970. Coccidiosis and histomoniasis in avian hosts. Pages 371-397 in Immunity to
parasitic animals. GJ Jackson, R. Herman and I. Singer, eds., New York. Duffy, C., M. Sims, and R. Power. 2005. Evaluation of dietary NatustatTM for control of
Histomonas meleagridis in male turkeys on infected litter. Avian Dis. 49:423–425.
Dufour, L., J. E. Sander, R. D. Wyatt, G. N. Rowland, and R. Page. 1992. Experimental exposure of broiler chickens to boric acid to assess clinical signs and lesions of toxicosis. Avian Dis. 36: 1007–1011.
Van Eldere, J., P. Celis, G. De Pauw, E. Lesaffre, and H. Eyssen. 1996. Tauroconjugation of cholic acid stimulates 7 alpha-dehydroxylation by fecal bacteria. Appl. Environ. Microbiol. 62:656–661.
Eren, M., F. Uyanik, B. K. Guclu, and M. Cinar. 2012. Effects of dietary boric acid and borax supplementation on growth performance and some biochemical parameters in broilers. Revue Méd. Vét. 163:546–551.
Gruber, J., P. Ganas, and M. Hess. 2017. Long-term in vitro cultivation of Histomonas meleagridis coincides with the dominance of a very distinct phenotype of the parasite exhibiting increased tenacity and improved cell yields. Parasitology. 144:1253–1263.
Hauck, R., P. L. Armstrong, and L. R. McDougald. 2010. Histomonas meleagridis (Protozoa: Trichomonadidae): Analysis of growth requirements in vitro. J. Parasitol. 96:1–7.
Van der Heijden, H. M. J. F., and W. J. M. Landman. 2007. Improved culture of Histomonas
meleagridis in a modification of Dwyer medium. Avian Dis. 51:986–988.
Van der Heijden, H. M. J. F., L. R. McDougald, and W. J. M. Landman. 2005. High yield of parasites and prolonged in vitro culture of Histomonas meleagridis. Avian Pathol. 34:505–508.
Hernandez-Patlan, D., B. Solis-Cruz, A. Méndez-Albores, J. D. Latorre, X. Hernandez-Velasco, G. Tellez, and R. López-Arellano. 2018a. Comparison of PrestoBlue® and plating method to evaluate antimicrobial activity of ascorbic acid, boric acid and curcumin in an in vitro gastrointestinal model. J. Appl. Microbiol. 124:423–430.

24
Hernandez-Patlan, D., B. Solis-Cruz, K. P. Pontin, J. D. Latorre, M. F. Baxter, X. Hernandez-Velasco, R. Merino-Guzman, A. Méndez-Albores, B. M. Hargis, R. Lopez-Arellano, and others. 2018b. Evaluation of a solid dispersion of curcumin with polyvinylpyrrolidone and boric acid against Salmonella Enteritidis infection and intestinal permeability in broiler chickens: A pilot study. Front. Microbiol. 9: 1289.
Hess, M., T. Kolbe, E. Grabensteiner, and H. Prosl. 2006. Clonal cultures of Histomonas
meleagridis, Tetratrichomonas gallinarum and a Blastocystis sp. established through micromanipulation. Parasitology. 133:547–554.
Hess, M., D. Liebhart, I. Bilic, and P. Ganas. 2015. Histomonas meleagridis—new insights into
an old pathogen. Vet. Parasitol. 208:67–76. Hess, M., D. Liebhart, E. Grabensteiner, and A. Singh. 2008. Cloned Histomonas meleagridis
passaged in vitro resulted in reduced pathogenicity and is capable of protecting turkeys from histomonosis. Vaccine. 26:4187–4193.
Hess, M., and L. McDougald. 2013. Histomoniasis (blackhead) and other protozoan diseases of the intestinal tract. Pages 1172–1178 in Diseases of Poultry.13th ed. E. Swayne, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. L. Suarez, and V. L. Nair, eds., Wiley-Blackwell, Ames, IA.
Hu, J., L. Fuller, and L. R. McDougald. 2004. Infection of turkeys with Histomonas meleagridis
by the cloacal drop method. Avian Dis. 48:746–750.
Hu, J., and L. McDougald. 2004. The efficacy of some drugs with known antiprotozoal activity against Histomonas meleagridis in chickens. Vet. Parasitol. 121:233–238.
Huang, C., Y. Guo, and J. Yuan. 2014. Dietary taurine impairs intestinal growth and mucosal structure of broiler chickens by increasing toxic bile acid concentrations in the intestine. Poult. Sci. 93:1475–1483.
Joyner, L. 1963. Immunity to histomoniasis in turkeys following treatment with dimetridazole. J. Comp. Pathol. Ther. 73:201–207.
Kokatnur, M., N. Rand, and F. Kummerow. 1958a. Effect of the energy to protein ratio on serum and carcass cholesterol levels in chicks. Circ. Res. 6:424–431.
Kokatnur, M., N. Rand, F. Kummerow, and H. Scott. 1958b. Effect of dietary protein and fat on changes of serum cholesterol in mature birds. J. Nutr. 64:177–184.
Küçükyilmaz, K., M. Bozkurt, M. Çinar, and A. E. Tüzün. 2017. Evaluation of the boron and phytase, alone or in combination, in broiler diets. J. Poult. Sci. 54:26–33.

25
Kummerow, F., A. Ueno, T. Nishida, and M. Kokatnur. 1960. Unsaturated fatty acids and plasma lipids. Am. J. Clin. Nutr. 8:62–67.
Kurtoğlu, F., V. Kurtoğlu, I. Çelik, T. Keçeci, and M. Nizamlioğlu. 2005. Effects of dietary boron supplementation on some biochemical parameters, peripheral blood lymphocytes, splenic plasma cells and bone characteristics of broiler chicks given diets with adequate or inadequate cholecalciferol (vitamin D3) content. Br. Poult. Sci. 46:87–96.
LeBlanc, M. J., V. Gavino, A. Pérea, I. M. Yousef, E. Lévy, and B. Tuchweber. 1998. The role of dietary choline in the beneficial effects of lecithin on the secretion of biliary lipids in rats. Biochim. Biophys. Acta. 1393:223–234.
Lefebvre, P., B. Cariou, F. Lien, F. Kuipers, and B. Staels. 2009. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 89:147–191.
Liebhart, D., P. Ganas, T. Sulejmanovic, and M. Hess. 2017. Histomonosis in poultry: Previous
and current strategies for prevention and therapy. Avian Pathol. 46:1–18.
Liebhart, D., and M. Hess. 2009. Oral infection of turkeys with in vitro-cultured Histomonas meleagridis results in high mortality. Avian Pathol. 38:223–227.
Liebhart, D., T. Sulejmanovic, B. Grafl, A. Tichy, and M. Hess. 2013. Vaccination against histomonosis prevents a drop in egg production in layers following challenge. Avian Pathol. 42:79–84.
Liebhart, D., M. Windisch, and M. Hess. 2010. Oral vaccination of 1-day-old turkeys with in vitro attenuated Histomonas meleagridis protects against histomonosis and has no negative effect on performance. Avian Pathol. 39:399–403.
Lindsay, O., J. Biely, and B. March. 1969. Excretion of bile acids by cockerels fed different lipids. Poult. Sci. 48:1216–1222.
Lotfi, A., R. Hauck, P. Olias, and H. M. Hafez. 2014. Pathogenesis of histomonosis in experimentally infected specific-pathogen-free (SPF) layer-type chickens and SPF meat-type chickens. Avian Dis. 58:427–432.
Lund, E. E. 1959. Immunizing action of a nonpathogenic strain of Histomonas against blackhead in turkeys. J. Protozool. 6:182–185.
Lund, E. E. A. P. C., and A. M. Chute. 1967. Histomonas meleagridis after one thousand in vitro passages. J. Eukaryot. Microbiol. 14:349–351.
Lund, E. E., E. E. Wehr, and D. J. Ellis. 1966. Earthworm transmission of Heterakis and Histomonas to turkeys and chickens. J. Parasitol. 52: 899–902.

26
McDougald, L. R. 2005. Blackhead disease (histomoniasis) in poultry: A critical review. Avian Dis. 49:462–476.
McDougald, L., and L. Fuller. 2005. Blackhead disease in turkeys: Direct transmission of
Histomonas meleagridis from bird to bird in a laboratory model. Avian Dis. 49:328–331.
Mitra, T., W. Gerner, F. A. Kidane, P. Wernsdorf, M. Hess, A. Saalmüller, and D. Liebhart. 2017. Vaccination against histomonosis limits pronounced changes of B cells and T-cell subsets in turkeys and chickens. Vaccine. 35:4184–4196.
Mitra, T., Kidane, F. A., Hess, M., & Liebhart, D. 2018. Unravelling the immunity of poultry
against the extracellular protozoan parasite Histomonas meleagridis is a cornerstone for vaccine development: A review. Front. Immunol. 9: 2518.
Moore, J. A., E. S. Committee, and others. 1997. An assessment of boric acid and borax using the IEHR evaluative process for assessing human developmental and reproductive toxicity of agents. Reproductive Toxicol. 11:123–160.
Nakada-Tsukui, K., and T. Nozaki. 2016. Immune response of amebiasis and immune evasion by Entamoeba histolytica. Front. Immunol. 7:175.
Nguyen Pham, A. D., J. K. De Gussem, and B. M. Goddeeris. 2013. Intracloacally passaged low-virulent Histomonas meleagridis protects turkeys from histomonosis. Vet. Parasitol. 196:307–313.
Nishida, T., F. Takenaka, and F. Kummerow. 1958. Effect of dietary protein and heated fat on serum cholesterol and beta-lipoprotein levels, and on the incidence of experimental atherosclerosis in chicks. Circ. Res. 6:194–202.
Regmi, P. R., A. L. Shaw, L. L. Hungerford, J. R. Messenheimer, T. Zhou, P. Pillai, A. Omer, and J. M. Gilbert. 2016. Regulatory considerations for the approval of drugs against histomoniasis (blackhead disease) in turkeys, chickens, and game birds in the United States. Avian Dis. 60:725–730.
Ridlon, J. M., D.-J. Kang, and P. B. Hylemon. 2006. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 47:241–259.
Rossi, A., R. Miles, B. Damron, and L. Flunker. 1993. Effects of dietary boron supplementation
on broilers. Poult. Sci. 72:2124–2130.
Sander, J. E., L. Dufour, R. D. Wyatt, P. B. Bush, and R. K. Page. 1991. Acute toxicity of boric acid and boron tissue residues after chronic exposure in broiler chickens. Avian Dis. 35: 745–749.

27
Schwarz, A., M. Gauly, H. Abel, G. Daş, J. Humburg, A. T. A. Weiss, G. Breves, and S. Rautenschlein. 2011. Pathobiology of Heterakis gallinarum mono-infection and co-infection with Histomonas meleagridis in layer chickens. Avian Pathol. 40:277–287.
Sentíes-Cué, G., R. Chin, and H. Shivaprasad. 2009. Systemic histomoniasis associated with high mortality and unusual lesions in the bursa of Fabricius, kidneys, and lungs in commercial turkeys. Avian Dis. 53:231–238.
Stamp, D., and G. Jenkins. 2008. An overview of bile-acid synthesis, chemistry and function. Bile acids: Toxicology and bioactivity. Cambridge, UK: Royal Society of Chemistry: pp. 1–13.
Sulejmanovic, T., I. Bilic, M. Hess, and D. Liebhart. 2016. An in vitro attenuated strain of Histomonas meleagridis provides cross-protective immunity in turkeys against heterologous virulent isolates. Avian Pathol. 45:46–53.
Sulejmanovic, T., D. Liebhart, and M. Hess. 2013. In vitro attenuated Histomonas meleagridis does not revert to virulence, following serial in vivo passages in turkeys or chickens. Vaccine. 31:5443–5450.
Sun, X., K. Winglee, R. Z. Gharaibeh, J. Gauthier, Z. He, P. Tripathi, D. Avram, S. Bruner, A. Fodor, and C. Jobin. 2018. Microbiota-derived metabolic factors reduce Campylobacteriosis in mice. Gastroenterology. 154:1751–1763.
Thøfner, I. C. N., D. Liebhart, M. Hess, T. W. Schou, C. Hess, E. Ivarsen, X. Fretté, L. P. Christensen, K. Grevsen, R. M. Engberg, and others. 2012. Antihistomonal effects of artemisinin and Artemisia annua extracts in vitro could not be confirmed by in vivo experiments in turkeys and chickens. Avian Pathol. 41:487–496.
Tyzzer, E. E. 1920. The flagellate character and reclassification of the parasite producing“ Blackhead” in turkeys: Histomonas (gen. nov.) meleagridis (Smith). J. Parasitol. 6:124–131.
Tyzzer, E. E. 1932. Problems and observations concerning the transmission of blackhead infection in turkeys. Proc. Am. Philos. Soc. 71:407–410.
Tyzzer, E. E. 1934. Studies on histomoniasis, or“ blackhead” infection, in the chicken and the turkey. Daedalus. 69: 189–264.
Tyzzer, E. E., and M. Fabyan. 1922. A inquiry into the source of the virus in blackhead of turkeys, together with observations on the administration of ipecac and of sulfur. J. Exp. Med. 35:791–812.
Tyzzer, E. E., M. Fabyan, and N. C. Foot. 1921. Further observations on“ blackhead” in turkeys. J. Infect. Dis. 29:268–286.

28
Wang, H., J. D. L. Cardenas, M. Bansal, B. Al-Rubaye, G. Tellez, B. Hargis, and X. Sun. 2018. Microbiota metabolic product deoxycholic acid controls chicken necrotic enteritis. bioRxiv:215640.
Winston, J. A., and C. M. Theriot. 2016. Impact of microbial derived secondary bile acids on colonization resistance against Clostridium difficile in the gastrointestinal tract. Anaerobe. 41:44–50.
Yang, F., M. Ma, J. Xu, X. Yu, and N. Qiu. 2012. An egg-enriched diet attenuates plasma lipids and mediates cholesterol metabolism of high-cholesterol fed rats. Lipids. 47:269–277.
Zaragatzki, E., M. Hess, E. Grabensteiner, F. Abdel-Ghaffar, K. A. S. Al-Rasheid, and H. Mehlhorn. 2010a. Light and transmission electron microscopic studies on the encystation of Histomonas meleagridis. Parasitol. Res. 106:977–983.
Zaragatzki, E., H. Mehlhorn, F. Abdel-Ghaffar, K. A. S. Rasheid, E. Grabensteiner, and M. Hess. 2010b. Experiments to produce cysts in cultures of Histomonas meleagridis-the agent of histomonosis in poultry. Parasitol. Res. 106:1005–1007.

29
III. DATA CHAPTER 1
Evaluation of deoxycholic acid as a prophylactic treatment to prevent histomoniasis in
turkeys
L.C. Beer*, C.N. Vuong*, J.D. Latorre*, S.J. Rochell*, X. Sun*, G. Tellez*, and B. M. Hargis*
*Center of Excellence for Poultry Science, University of Arkansas Division of Agriculture,
Fayetteville, AR 72701
This manuscript will be submitted to Poultry Science

30
ABSTRACT
Deoxycholic acid (DCA) is a naturally occurring secondary bile acid, originating from
intestinal bacterial metabolic conversion of cholate, a primary bile acid. DCA has been shown to
have anti-histomonal properties in vitro, leading to our hypothesis that DCA inclusion within the
feed may prevent histomoniasis in turkeys. Selected concentrations of DCA within a basal starter
diet were evaluated for effects on body weight gain (BWG), lesions, and mortality of H.
meleagridis-challenged turkeys. Treatments consisted of Negative Control, 0.25% DCA diet,
0.5% DCA diet, 1% DCA diet, or Wild-Type (WT) Positive Control. The basal turkey starter
diet was fed to all groups until d7, at which time DCA diets were administered to respective
groups. Via intracloacal inoculation, 2x105 WT H. meleagridis cells/turkey were administered on
d14, and lesions were evaluated d13 post-challenge. Pre-challenged d0-14 BWG in the 0.25%
DCA group was higher (p≤0.05) than the 1% DCA group. There were no significant differences
in pre-challenge d0-14 BWG between any of the other groups. No significant differences in
mortalities from histomoniasis occurred in DCA treatment groups as compared to the WT
Positive Control. No lesions or mortalities characteristics of histomoniasis were observed at any
time in the Negative Control poults. Presence of classic Histomoniasis-related liver lesions was
statistically higher in the 0.5% DCA diet as compared to the WT Positive Control. Utilizing the
same controls and experimental timeline, an additional group was included to evaluate a
biliogenic diet that was formulated to encourage endogenous bile acid production. The biliogenic
diet had no statistical impact on pre-challenge d0-14 BWG, but this diet did not reduce mortality
or lesions related to histomoniasis. Taken together, these data suggest DCA inclusion within the
feed at these concentrations and under these experimental conditions does not prevent
Histomoniasis. Although DCA treatment reduced H. meleagridis cells in vitro, the in vivo trial

31
resulted in no reduction of mortalities or lesion presence from histomoniasis within the DCA
diets as compared to the WT Positive Control.
Key Words: blackhead, deoxycholic acid, histomoniasis, Histomonas meleagridis, turkey

32
INTRODUCTION
Histomoniasis, also known as blackhead, is an important disease particularly affecting
turkeys in addition to other gallinaceous birds (van der Heijden et al., 2005; Hess et al., 2015).
Caused by the protozoan parasite Histomonas meleagridis, mortality can approach 80-100% of
the flock with significant economic damage incurred (Callait et al., 2002; McDougald, 2005;
Hess and McDougald, 2013). Nitroimidazoles were previously an effective treatment for
histomoniasis; however, regulatory action resulted in the removal of effective prophylactic and
therapeutic compounds such as this without any alternatives introduced for disease treatment
(Joyner, 1963; Hess and McDougald, 2013; Liebhart et al., 2013).
Deoxycholic acid (DCA) is a naturally occurring secondary bile acid produced through
intestinal bacterial metabolic conversion of cholate (Van Eldere et al., 1996; Ridlon et al., 2006).
DCA has been shown to reduce severity of Eimeria maxima and Clostridium perfringens poultry
infections when administered in a dietary concentration of 1.5 g/kg with an associated reduction
of damaged intestinal villi (Wang et al., 2018). A study in mice demonstrated that anaerobic
bacteria-derived DCA protected against colitis induced by Campylobacter jejuni (Sun et al.,
2018). The primary purpose of the present study was to evaluate DCA as a chemoprophylaxis
candidate against histomoniasis. In addition, dietary composition has been shown to influence
endogenous production of bile acids within the host (Yang et al., 2012). Therefore, a biliogenic
diet was formulated to encourage endogenous bile acid formation in the turkey with the
hypothesis that severity of histomoniasis would be reduced.

33
MATERIALS AND METHODS
In vitro Assessment of Deoxycholic Acid
Three in vitro assays were completed to evaluate selected concentrations of high purity
DCA sodium salt (VWR International LLC, USA). Wild-type (WT), virulent H. meleagridis
were added at a ratio of 100µL histomonads: 50µL DCA treatment into a 96-well, sterile
microtiter plate. Each treatment was performed in pentaplicate. Incubation occurred at 40C with
wells capped and parafilm utilized to maintain anaerobic conditions. Following incubation,
viable histomonads were enumerated by trypan blue 0.4% vital dye exclusion using a
hemocytometer, and cell counts were expressed as viable histomonads/mL.
In Assay 1, a concentration of 2.01x106 histomonads/mL of WT H. meleagridis was
added according to the method above. Treatments included sterile phosphate-buffered saline
(PBS) as a negative control or selected final DCA concentrations of 0.4, 2, and 4mM. The plate
was incubated 7-8.5 hours before viable histomonads were enumerated as described above. In
Assay 2, a concentration of 6.88x105 histomonads/mL was used and treatments included either
PBS control or 0.5, 1, 2, or 4mM DCA concentrations. The plate was incubated 6-8 hours before
viable histomonads were enumerated. In Assay 3, a concentration of 6.35x105 histomonads/mL
was added and treatments included either PBS or DCA concentrations of 0.5, 1, or 2mM. The
plate was incubated and viable histomonads were enumerated at two time periods of 4-6 hours
and 27-29 hours.
Animal Source and Diet
A total of 140 day-of-hatch female turkey poults were obtained from a local commercial
hatchery. Poults were neck-tagged individually and randomly allocated to floor pens at the
University of Arkansas Poultry Health Laboratory. All animal handling procedures were in

34
compliance with the Institutional Animal Care and Use Committee (IACUC protocol #18113) of
the University of Arkansas. A corn-soy-based starter feed that met or exceeded nutrient
requirements for poultry (NRC, 1994) and water were provided ad libitum. Early poult
mortalities unrelated to histomoniasis were recorded and altered group numbers reported in the
experiment.
DCA diet. On d7, DCA was included in the diet at selected concentrations of either 0.25,
0.5, or 1%. Treatments consisted of Negative Control (n=59), 0.25% DCA diet (n=20), 0.5%
DCA diet (n=20), 1% DCA diet (n=20), and WT Positive Control (n=20). Turkeys remained on
treatment diets for the remainder of the experiment.
Biliogenic diet. Utilizing the same positive and negative controls as above, a Biliogenic
diet treatment group (n=20) consisting of 20% whole egg powder (Heartland Supply Co., USA)
inclusion within a basal turkey starter was evaluated. This treatment’s objective purpose was to
physiologically upregulate natural bile acid synthesis to potentially increase endogenous DCA
production within the turkey (Table 1). This group was subjected to the same experimental
timeline and evaluation methods as the DCA treatment groups mentioned previously (Figure 1).
Histomonas meleagridis Challenge
To initiate disease challenge, all poults other than the Negative Control received a total
dose of 2x105 WT, virulent H. meleagridis cells/poult administered intracloacally with an animal
gavage needle on d14. Inoculation occurred twice (at half total dosage) with 1h between
inoculations to ensure each bird received an infectious dose.
Lesion Scores and Body Weight Gain
All poults were weighed individually on d0 and d14 for calculation of pre-challenge body
weight gain (BWG). Presence liver and cecal lesions associated with histomoniasis was recorded

35
from all mortalities following challenge. On d13, all remaining poults were necropsied for the
presence or absence of liver and cecal lesions typical of histomoniasis.
Statistical Analysis
In vitro data were computed using JMP Pro 14 software. Significant differences between
viable histomonads/mL in treatment groups were determined using ANOVA, and means were
further separated using Tukey’s multiple comparison post hoc test with values of p≤0.05
considered significant. Pre-challenged BWG data were also analyzed using JMP Pro 14 software,
with significant differences between BWG in treatment groups determined using ANOVA.
Where applicable, means were further separated using Tukey’s multiple comparison post hoc test
with values of p≤0.05 considered significant. Mortalities and lesion presence related to
histomoniasis were compared against the WT Positive Control using chi-square test with a
difference of p≤0.05 considered significant.
RESULTS
In vitro Cell Viability Assays
In assay 1, mean viable histomonads/mL (Log10) for PBS, 0.4mM DCA, 2mM DCA and
4mM DCA treatments following 7-8.5h incubation were 6.22, 6.25, 0.00, and 0.00, respectively
(Table 2). The treatments of 2mM and 4mM DCA significantly reduced the concentration of
viable histomonads as compared to either the PBS negative control or the 0.4mM DCA.
In assay 2, mean viable histomonads/mL (Log10) for PBS, 0.5mM DCA, 1mM DCA,
2mM DCA, and 4mM DCA treatments following 6-8h incubation were 6.11, 6.12, 4.71, 0.00,
and 0.00, respectively. The treatments of 1mM, 2mM, and 4mM DCA significantly reduced the
concentration of viable histomonads as compared to either the PBS or the 0.5mM DCA.

36
In assay 3, mean viable histomonads/mL (Log10) for PBS, 0.5mM DCA, 1mM DCA,
and 2mM DCA following 4-6h incubation were 6.30, 6.31, 6.16, and 1.34, respectively. The
1mM DCA treatment significantly reduced the viable histomonads as compared to PBS or
0.5mM DCA. The 2mM DCA had lower viable histomonads as compared to all other treatments.
Following 27-29h incubation, histomonads from assay 3 were enumerated with mean counts of
6.48, 6.18, 4.46, and 0.00, respectively. The 0.5mM DCA treatment had lower viable histomonad
counts as compared to the PBS. The 1mM and 2mM DCA significantly reduced viable
histomonads as compared to either the PBS or 0.5mM DCA treatments.
Pre-challenge BWG from d0-14
The selected dietary concentrations of DCA had no statistically negative impact on d0-14
pre-challenged BWG as compared to the basal control diet. The 0.25% DCA diet had higher
(p≤0.05) pre-challenge BWG as compared to the 1% DCA diet (Table 3). The 1% DCA diet was
lower at pre-challenge BWG than the Negative Control but was not statistically different. No
differences were observed in pre-challenge BWG between any of the other DCA treatments.
Histomoniasis Infection Response and Lesions
Differences in mortalities associated with histomoniasis were not significant between any
of the DCA treatments as compared to the WT Positive Control (Figure 2). Presence liver and
cecal lesions associated with histomoniasis was significantly higher in the 0.5% DCA diet as
compared to the WT Positive Control (Figure 3). No classical lesions associated with
histomoniasis were observed at any time in the Negative Control.
Biliogenic Dietary Impact on Histomoniasis
The biliogenic diet did not reduce histomoniasis. No statistical difference was detected in
the d0-14 pre-challenge BWG (Figure 4A). Mortalities and lesions related to histomoniasis were

37
not reduced in the biliogenic diet group as compared to the WT Positive Control (Figures 4B
and 4C, respectively).
DISCUSSION
In vitro testing of DCA effectively reduced the viability of H. meleagridis; viable
histomonad counts decreased with increased concentration of DCA. However, dietary inclusion
at the selected concentrations neither prevented nor mitigated the disease in H. meleagridis-
challenged turkeys. Although not statistically different, BWG during the pre-challenge phase
decreased with increased dietary DCA as compared to the basal diet. This could indicate that the
maximum acceptable concentration of DCA inclusion was approached, and higher DCA
inclusion rates may not be useful if any further research is conducted. Again, although not
statistically significant, the 0.25% and 0.5% DCA groups had higher mortalities as compared to
the WT Positive Control. The higher lesion presence within the 0.5% DCA group as compared to
the WT Positive Control further contributes to the conclusion from this study that DCA was not
effective against histomoniasis under these experimental conditions. A higher inclusion rate of
DCA would be discouraged unless further evaluation was completed to evaluate physiological
impact. In addition, a higher concentration of DCA would likely be economically unfeasible.
Mortalities and lesions characteristic of histomoniasis were not reduced by the biliogenic diet.
Similarly to the DCA diets, the biliogenic diet numerically (not statistically) reduced pre-
challenge BWG as compared to the basal diet. Mortalities and lesions were not statistically
different, although the biliogenic diet marginally increased the rates of both. If future studies
were to be conducted, it might be beneficial to include measurement of feed intake to evaluate
whether the poults were ingesting adequate amounts of feed compared to the basal diet.

38
According to Lefebvre et al. (2009), the “classical pathway” for bile acid synthesis
contains the cholesterol 7α-hydroxylase (CYP7A1) and contributes to approximately 75% of bile
acid synthesis within the liver. Cholesterol, an important precursor for bile acids, has recently
been shown to enhance H. meleagridis growth in vitro (Gruber et al., 2018). Based on this recent
finding by Gruber et al. (2018), the biliogenic diet formulation, which included 20% whole egg
powder, may have increased normal cholesterol levels and inadvertently aided the parasite. Both
lecithin-enriched and egg-enriched diets have been associated with increased total bile acid
output (Lindsay et al., 1969; LeBlanc et al., 1998; Yang et al., 2012), but no measurements of
bile acids were evaluated within the present study. Although DCA displayed significant
reduction of histomonads in vitro, the ineffectiveness to prevent histomoniasis in vivo is
consistent with the variable results of other candidates evaluated against this disease. Extracts of
the medicinal herb Artemisia annua as well as artemisinin (a main active compound) previously
exhibited anti-histomonal properties in vitro but failed to prevent infection within experimental
challenged turkeys and chickens (Thøfner et al., 2012). The same H. meleagridis culture was
utilized by Thøfner et al. (2012), but in vitro susceptibility did not translate to in vivo results.
These findings are similar to the DCA results, further emphasizing the importance of
incorporating in vivo evaluation of anti-histomonal chemoprophylaxis compounds rather than
relying only upon positive in vitro data. Further confounding this already complicated disease,
the cecal environment wherein H. meleagridis resides is constantly in a state of flux,
demonstrating the difficulties in controlling histomoniasis. Bacterial flora contribute an
important role in disease development; therefore, influence of chemoprophylaxis candidates on
the gastrointestinal environment as well as the parasite should be considered (Callait et al., 2002;
McDougald, 2005). In conclusion, DCA and the formulated biliogenic diet were evaluated for

39
the first known time against histomoniasis but none of the dietary treatments were effective in
the mitigation of disease.

40
REFERENCES
Callait, M., C. Granier, C. Chauve, and L. Zenner. 2002. In vitro activity of therapeutic drugs against Histomonas meleagridis (Smith, 1895). Poult. Sci. 81:1122–1127.
Van Eldere, J., P. Celis, G. De Pauw, E. Lesaffre, and H. Eyssen. 1996. Tauroconjugation of cholic acid stimulates 7 alpha-dehydroxylation by fecal bacteria. Appl. Environ. Microbiol. 62:656–661.
Gruber, J., A. Pletzer, and M. Hess. 2018. Cholesterol supplementation improves growth rates of Histomonas meleagridis in vitro. Exp. Parasitol. 185: 53-61
Van der Heijden, H. M. J. F., L. R. McDougald, and W. J. M. Landman. 2005. High yield of parasites and prolonged in vitro culture of Histomonas meleagridis. Avian Pathol. 34:505–508.
Hess, M., D. Liebhart, I. Bilic, and P. Ganas. 2015. Histomonas meleagridis—new insights into an old pathogen. Vet. Parasitol. 208:67–76.
Hess, M., and L. McDougald. 2013. Histomoniasis (blackhead) and other protozoan diseases of the intestinal tract. Pages 1172–1178 in Diseases of Poultry.13th ed. E. Swayne, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. L. Suarez, and V. L. Nair, eds., Wiley-Blackwell, Ames, IA.
Joyner, L. 1963. Immunity to histomoniasis in turkeys following treatment with dimetridazole. J.
Comp. Pathol. Ther. 73:201–207.
LeBlanc, M. J., V. Gavino, A. Pérea, I. M. Yousef, E. Lévy, and B. Tuchweber. 1998. The role of dietary choline in the beneficial effects of lecithin on the secretion of biliary lipids in rats. Biochim. Biophys. Acta. 1393:223–234.
Lefebvre, P., B. Cariou, F. Lien, F. Kuipers, and B. Staels. 2009. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 89:147–191.
Liebhart, D., T. Sulejmanovic, B. Grafl, A. Tichy, and M. Hess. 2013. Vaccination against histomonosis prevents a drop in egg production in layers following challenge. Avian Pathol. 42:79–84.
Lindsay, O., J. Biely, and B. March. 1969. Excretion of bile acids by cockerels fed different lipids. Poult. Sci. 48:1216–1222.
McDougald, L. R. 2005. Blackhead disease (histomoniasis) in poultry: A critical review. Avian Dis. 49:462–476.
NRC. 1994. Nutrient Requirements of Poultry. 9th rev. ed. Natl. Acad. Press, Washington, DC.

41
R Ridlon, J. M., D.-J. Kang, and P. B. Hylemon. 2006. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 47:241–259.
Sun, X., K. Winglee, R. Z. Gharaibeh, J. Gauthier, Z. He, P. Tripathi, D. Avram, S. Bruner, A.
Fodor, and C. Jobin. 2018. Microbiota-derived metabolic factors reduce Campylobacteriosis in mice. Gastroenterology. 154:1751–1763.
Thøfner, I. C. N., D. Liebhart, M. Hess, T. W. Schou, C. Hess, E. Ivarsen, X. Fretté, L. P. Christensen, K. Grevsen, R. M. Engberg, and others. 2012. Antihistomonal effects of artemisinin and Artemisia annua extracts in vitro could not be confirmed by in vivo experiments in turkeys and chickens. Avian Pathol. 41:487–496.
Wang, H., J. D. L. Cardenas, M. Bansal, B. Al-Rubaye, G. Tellez, B. Hargis, and X. Sun. 2018. Microbiota metabolic product deoxycholic acid controls chicken necrotic enteritis. bioRxiv:215640.
Yang, F., M. Ma, J. Xu, X. Yu, and N. Qiu. 2012. An egg-enriched diet attenuates plasma lipids
and mediates cholesterol metabolism of high-cholesterol fed rats. Lipids. 47:269–277.

42
TABLES
Table 1. Ingredient composition of the biliogenic diet for induction of endogenous bile acids.
Item1 % as-fed
Corn 45.7
Soybean meal 28.2
Spray-dried egg powder2 20.0
Dicalcium phosphate 2.75
Limestone 1.52
Poultry Fat 1.00
L-lysine HCl 0.23
Salt 0.20
DL-methionine 0.15
CT Tky Starter VTM 0.15
Choline chloride (60%) 0.07
L-threonine 0.01 168mg of biotin was added to the premix 2Spray-dried egg powder, Heartland Supply Co., Fayetteville, AR.

43
Table 2. In vitro viability assays evaluating selected concentrations of deoxycholic acid for anti-histomonal properties.1,2
ASSAY 1 Viable Cells/mL (Log10)
Treatment After 7-8.5 hr. incubation PBS 6.22 ± 0.02
a
0.4 mM DCA 6.25 ± 0.03a
2 mM DCA 0.00 ± 0.00b
4 mM DCA 0.00 ± 0.00b
ASSAY 2 Viable Cells/mL (Log10) Treatment After 6-8 hr. incubation PBS 6.11 ± 0.02
a
0.5 mM DCA 6.12 ± 0.05a
1 mM DCA 4.71 ± 0.25b
2 mM DCA 0.00 ± 0.00b
4 mM DCA 0.00 ± 0.00b
ASSAY 3 Viable Cells/mL (Log10) Treatment After 4-6hr. Incubation After 27-29 hr. incubation PBS 6.30 ± 0.05
a 6.48 ± 0.01
a
0.5 mM DCA 6.31 ± 0.03a 6.18 ± 0.07
b
1 mM DCA 6.16 ± 0.04b 4.46 ± 1.25
c
2 mM DCA 1.34 ± 0.92c 0.00 ± 0.00
c
a-cData are expressed as the Mean ± SEM, n=5 samples. Statistical evaluation using ANOVA followed by Tukey’s multiple post-hoc test. Means with no common superscript differ significantly (p≤0.05). 1Assay 1-3 began with concentrations of 2.01x106, 6.88x105, and 6.35x105 histomonads/mL of the wild-type H. meleagridis, respectively, added at a ratio of 100µL cells: 50µL treatment and incubated under anaerobic conditions at 40°C. 2DCA=Deoxycholic acid Table 3. Effect of dietary inclusion of selected concentrations of deoxycholic acid on d0-14 BWG during pre-challenge phase.
Treatment d0-14 Pre-Challenge BWG (g)1
Wild-Type Positive Control d14 270 ± 9.36ab 0.25% DCA 289 ± 14.0a 0.5% DCA 257 ± 13.1ab 1% DCA 242 ± 12.4b Negative Control 265 ± 6.34ab
a,bMeans ± SEM with no common superscript differ significantly (p≤0.05). 1Statistical evaluation using ANOVA followed by Tukey’s multiple post-hoc test. 2DCA=Deoxycholic Acid.
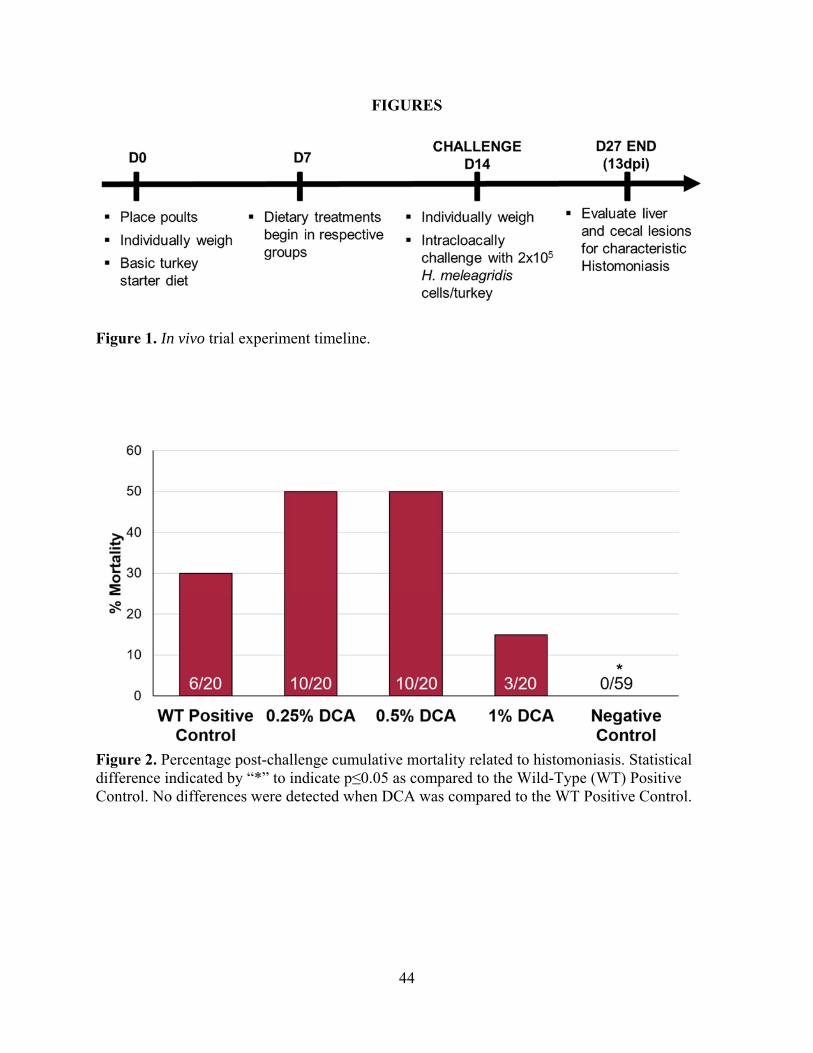
44
FIGURES
Figure 1. In vivo trial experiment timeline.
Figure 2. Percentage post-challenge cumulative mortality related to histomoniasis. Statistical difference indicated by “*” to indicate p≤0.05 as compared to the Wild-Type (WT) Positive Control. No differences were detected when DCA was compared to the WT Positive Control.

45
Figure 3. Percentage cumulative liver and cecal lesions associated with histomoniasis beginning from 9d post-challenge until d13 experiment termination. Lesions were determined based on the presence or absence of classic histomoniasis that is characterized by target-like liver lesions and cecal cores. Statistical differences indicated by “*” to indicate p≤0.05 as compared to the Wild-Type (WT) Positive Control. No lesions or mortalities associated with histomoniasis were detected at any time within the Negative Control.

46
Figure 4. A) Biliogenic dietary effect on d0-14 BWG during pre-challenge phase. No significant differences were found between BWG. B) Post-challenge cumulative mortality associated with histomoniasis. Statistical difference indicated by “*” to indicate p≤0.05 as compared to Wild-Type (WT) Positive Control. C) Percentage cumulative liver and cecal lesions associated with histomoniasis beginning from d6 post-challenge until d13 experiment termination. No significant differences in lesions associated with histomoniasis were found between the biliogenic diet group and WT Positive Control following WT H. meleagridis-challenge.

47
IV. DATA CHAPTER 2
Evaluation of boric acid as a chemoprophylaxis candidate to prevent histomoniasis
L. C. Beer*, C. N. Vuong*, T. L. Barros*, J. D. Latorre*, G. Tellez*, and B. M. Hargis*
*University of Arkansas: Division of Agriculture, Poultry Science Dept., Fayetteville, AR 72701
To be submitted as a research note to Poultry Science

48
ABSTRACT
Histomoniasis, caused by the protozoan parasite Histomonas meleagridis, is a disease to
which turkeys are especially susceptible. Currently, no chemoprophylaxis compounds are
available to mitigate this disease. Boric acid exhibits antifungal, antiseptic and antiviral
properties, and it has been used in the treatment of yeast infections. Based upon these
characteristics, an experiment was conducted to evaluate whether boric acid might be efficacious
against H. meleagridis. Treatments consisted of Negative Control, 0.2% Boric Acid diet, and
Wild-Type Positive (WT) Challenged Control. The 0.2% Boric Acid diet was administered to the
respective group beginning on day-of-hatch. On d21, challenge with 2x105 H. meleagridis
cells/turkey occurred intracloacally, and lesions were evaluated on d14 post-challenge.
Individual body weights were recorded on d0, 21, and 35 to calculate the pre-challenge and post-
challenge body weight gain (BWG). The 0.2% Boric Acid group resulted in lower pre-challenge
d0-21 BWG (p≤0.05) than the Negative Control. Post-challenge d21-35 BWG was not
statistically lower (p=0.0567) than the wild-type positive control. No differences were detected
in mortalities associated with histomoniasis between the 0.2% Boric Acid group and the WT
Positive Control. Liver and cecal lesions were not statistically different between the 0.2% Boric
Acid group and the WT Positive Control. Taken together, these data suggest that boric acid was
not efficacious in the prevention or reduction of disease severity when provided at this dietary
concentration under these experimental conditions.
Key words: blackhead, boric acid, boron, histomoniasis, Histomonas meleagridis

49
INTRODUCTION
Histomoniasis, also commonly known as blackhead, is a disease of turkeys associated
with high mortality (Callait et al., 2002). Considered critically and economically impactful to
both turkeys and chickens, histomoniasis is a serious concern facing the poultry industry (Duffy
et al., 2005; Lotfi et al., 2014). Histomonas meleagridis, the etiological agent of histomoniasis,
penetrates the cecal epithelial lining, replicates, enters the bloodstream, and parasitizes the liver
(Clarkson, 1963; Hess and McDougald, 2013). Research on this organism waned in the 1960s
following the introduction of nitroimidazoles, nitrofurans, and arsenical compounds for
prophylaxis and treatment of outbreaks; compounds have since been banned due to regulatory
action (van der Heijden et al., 2005; Hess et al., 2006). In 2015, the arsenic-based drug nitarsone
(Histostat), the last remaining FDA-approved drug for prevention of histomoniasis, was
withdrawn from the market due to concerns about inorganic arsenic residues in treated poultry
(Regmi et al., 2016). Unfortunately, no alternative to the previously used drugs has been
identified; in vitro and in vivo studies continue to yield variable results against histomoniasis
(Thøfner et al., 2012).
Boron is an essential element to humans, animals, and plants (Eren et al., 2012). The NRC
(1994) has no recommended level of boron for daily intake in poultry. Within the poultry
industry, application of boric acid, a boron compound, to the litter is used in the prevention of
darkling beetles (Sander et al., 1991; Dufour et al., 1992). In 1992, Dufour et al. showed that
boric acid litter treatment at a rate of 0.4-0.9kg/9.3m2 did not significantly increase feed
conversion or decrease body weight. Previous studies have suggested the important biological
role that boron may have on the biochemical mechanisms influencing mineral metabolism and
normal growth (Kurtoğlu et al., 2005; Çinar et al., 2015). Dietary supplementation of boron is

50
considered economical in that a 100mg/kg diet was estimated to cost 0.5 USD per ton of
prepared feed (Bozkurt and Kucukyilmaz, 2015). Beginning at day-of-hatch until 21d, up to 240
ppm (0.024%) boron within the diet was not detrimental to broiler performance, although boron
levels within breast muscle and liver tissues increased in proportion boron dietary concentration
(Rossi et al., 1993). Boron (20mg/kg) supplementation in a basal diet had no impact on body
weight or feed consumption in chickens; results did not suggest growth-promotion or metabolic
mineral regulation (Küçükyilmaz et al., 2017). However, the acute oral mean lethal dose of boric
acid in 1-day-old chicks was determined to be 2.95 ± 0.35 g/kg of body weight, resulting in the
classification of boron as a slightly toxic chemical (Sander et al., 1991).
Containing antifungal, antiseptic and antiviral properties, boric acid has also been used to
treat yeast infections (Hernandez-Patlan et al., 2018a). Within an in vitro model, boric acid
decreased concentration of Salmonella Enteritidis within the intestinal compartment (Hernandez-
Patlan et al., 2018a). However, during an in vivo study, a concentration of 0.1% boric acid within
the basal diet had no significant reduction in Salmonella Enteritidis (Hernandez-Patlan et al.,
2018b). The growth rate of Trichomonas vaginalis, protozoan causative agent of trichomoniasis
in humans, was reduced with low boric acid concentrations (0.2%) and exhibited lethality to
trichomonads at higher concentrations (≤0.4%), independent of environmental acidification
(Brittingham and Wilson, 2014). Bacterial flora are important in development of histomoniasis,
contributing to the interest in boric acid as a potential chemoprophylactic compound against the
disease. Considering these experiments, the antifungal properties, and the potential cost-
effectiveness, we hypothesized that boric acid might be efficacious against the trichomonad H.
meleagridis at the selected dietary concentration of 0.2%.

51
MATERIALS AND METHODS
Animal Source and Diet
A total of 120 day-of-hatch female turkey poults were obtained from a local commercial
hatchery. Poults were neck-tagged individually and randomly allocated to floor pens at the
University of Arkansas Poultry Health Laboratory. Early poult mortalities unrelated to
histomoniasis were recorded and the altered group numbers are reported in the experiment. All
animal handling procedures were in compliance with the Institutional Animal Care and Use
Committee (IACUC protocol #18113) of the University of Arkansas. A corn-soy-based starter
feed that met or exceeded nutrient requirements of poultry (NRC, 1994) and water were provided
ad libitum. Treatments consisted of Negative Control (n=34), 0.2% Boric Acid diet (n=27), and
Wild-Type (WT) Positive Challenged Control (n=20). Beginning on day-of-hatch, boric acid
(Sigma-Aldrich, St. Louis, MO) was incorporated into the basal diet at a concentration of 0.2%
for the treatment group receiving the boric acid diet.
Histomonas meleagridis
On d21, all poults other than the Negative Control received a total dosage of 2x105 WT,
virulent H. meleagridis cells/turkey administered intracloacally with an animal gavage needle.
Inoculation occurred twice with a 1h period between each inoculation.
Lesion Scores and Body Weight Gain
All poults were individually weighed on d0, 21, and 35 for calculation of pre-challenge
and post-challenge body weight gain (BWG). Liver and cecal lesions associated with
histomoniasis were tabulated from all mortalities. On d14 post-challenge, all remaining poults
were necropsied to evaluate liver and cecal lesions. Classical lesions associated with
histomoniasis were separately recorded on a scale of 0-3, with 3 being the most severe.

52
Statistical Analysis
BWG data were analyzed using JMP Pro 14 software, with significant differences
between treatment groups determined using a Student’s t-test with p≤0.05 considered significant.
Differences in mortalities associated with histomoniasis were analyzed using chi square test.
Lesion score data were analyzed using the Proc Mixed Procedure in SAS 9.4 software with
significance between mean lesion score values considered at p≤0.05 in comparison to the WT
Positive Control.
RESULTS
Pre-challenge BWG from d0-21 was significantly lower in poults fed the 0.2% Boric
Acid diet as compared to poults fed the basal diet in the Negative Control group (Figure 1A).
Post-challenge BWG from d21-35 in the 0.2% Boric Acid group was numerically lower than the
WT Positive Control but not statistically different with p=0.0567 (Figure 1B). No differences
were detected in mortalities associated with histomoniasis in the 0.2% Boric Acid group as
compared to the WT Positive Control (Figure 1C). Moreover, neither liver nor cecal lesions were
reduced in the 0.2% Boric Acid group as compared to the WT Positive Control following H.
meleagridis-challenge (Figures 1D and 1E, respectively).
DISCUSSION
At this selected dietary concentration of boric acid, the significantly lower pre-challenged
BWG suggests that the level of boric acid should not exceed 0.2% in the diet fed to turkeys.
Lowered body weight gain is consistent with previous literature indicating that toxic levels of
boric acid can induce increased feed conversion and decreased body weight (Dufour et al.,
1992). Boric acid previously resulted in high toxicity when orally administered to 1-day-old
chicks at levels greater than 3.89 g/kg body weight (Sander et al., 1991). Efficacy of alternative

53
chemoprophylaxis compounds against related protozoa is suggested due to the close relationship
of H. meleagridis to other amoebae and flagellates (Hu and McDougald, 2004). However,
selective toxicity against protozoa is crucial to ensure that the chemoprophylaxis compound is
harmful to the parasite without causing irreversible damage to the host. The early mortalities
excluded from the data set could potentially have resulted from boric acid toxicity, but tissue
levels and lesions other than those characteristic to histomoniasis were not considered within this
experimental design. Overall, these data suggest that boric acid at this selected dietary
concentration and under these experimental conditions is not effective in the mitigation of
histomoniasis. If continued, future research should consider boric acid concentrations less than
0.2% within the diet.

54
REFERENCES
Bozkurt, M., and K. Kucukyilmaz. 2015. The role of boron in poultry nutrition Part II: Compositional and mechanical properties of bone and egg quality. Worlds Poult. Sci. J. 71:483–492.
Brittingham, A., and W. A. Wilson. 2014. The antimicrobial effect of boric acid on Trichomonas vaginalis. Sex. Transm. Dis. 41:718–722.
Callait, M., C. Granier, C. Chauve, and L. Zenner. 2002. In vitro activity of therapeutic drugs against Histomonas meleagridis (Smith, 1895). Poult. Sci. 81:1122–1127.
Çinar, M., K. Küçükyilmaz, M. Bozkurt, A. Çatli, E. Bintaş, H. Akşit, R. Konak, Ç. Yamaner,
and K. Seyrek. 2015. Effects of dietary boron and phytase supplementation on growth performance and mineral profile of broiler chickens fed on diets adequate or deficient in calcium and phosphorus. Br. Poult. Sci. 56:576–589.
Clarkson, M. 1963. Immunological responses to Histomonas meleagridis in the turkey and fowl.
Immunology. 6:156-168.
Duffy, C., M. Sims, and R. Power. 2005. Evaluation of dietary NatustatTM for control of Histomonas meleagridis in male turkeys on infected litter. Avian Dis. 49:423–425.
Dufour, L., J. E. Sander, R. D. Wyatt, G. N. Rowland, and R. Page. 1992. Experimental exposure of broiler chickens to boric acid to assess clinical signs and lesions of toxicosis. Avian Dis. 36: 1007–1011.
Eren, M., F. Uyanik, B. K. Guclu, and M. Cinar. 2012. Effects of dietary boric acid and borax supplementation on growth performance and some biochemical parameters in broilers. Revue Méd. Vét. 163:546–551.
Van der Heijden, H. M. J. F., L. R. McDougald, and W. J. M. Landman. 2005. High yield of parasites and prolonged in vitro culture of Histomonas meleagridis. Avian Pathol. 34:505–508.
Hernandez-Patlan, D., B. Solis-Cruz, A. Méndez-Albores, J. D. Latorre, X. Hernandez-Velasco, G. Tellez, and R. López-Arellano. 2018a. Comparison of PrestoBlue® and plating method to evaluate antimicrobial activity of ascorbic acid, boric acid and curcumin in an in vitro gastrointestinal model. J. Appl. Microbiol. 124:423–430.
Hernandez-Patlan, D., B. Solis-Cruz, K. P. Pontin, J. D. Latorre, M. F. Baxter, X. Hernandez-
Velasco, R. Merino-Guzman, A. Méndez-Albores, B. M. Hargis, R. Lopez-Arellano, and others. 2018b. Evaluation of a solid dispersion of curcumin with polyvinylpyrrolidone and boric acid against Salmonella Enteritidis infection and intestinal permeability in broiler chickens: A pilot study. Front. Microbiol. 9: 1289.

55
Hess, M., T. Kolbe, E. Grabensteiner, and H. Prosl. 2006. Clonal cultures of Histomonas meleagridis, Tetratrichomonas gallinarum and a Blastocystis sp. established through micromanipulation. Parasitology. 133:547–554.
Hess, M., and L. McDougald. 2013. Histomoniasis (blackhead) and other protozoan diseases of
the intestinal tract. Pages 1172–1178 in Diseases of Poultry.13th ed. E. Swayne, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. L. Suarez, and V. L. Nair, eds., Wiley-Blackwell, Ames, IA.
Hu, J., and L. McDougald. 2004. The efficacy of some drugs with known antiprotozoal activity
against Histomonas meleagridis in chickens. Vet. Parasitol. 121:233–238.
Küçükyilmaz, K., M. Bozkurt, M. Çinar, and A. E. Tüzün. 2017. Evaluation of the boron and phytase, alone or in combination, in broiler diets. J. Poult. Sci. 54:26–33.
Kurtoğlu, F., V. Kurtoğlu, I. Çelik, T. Keçeci, and M. Nizamlioğlu. 2005. Effects of dietary boron supplementation on some biochemical parameters, peripheral blood lymphocytes, splenic plasma cells and bone characteristics of broiler chicks given diets with adequate or inadequate cholecalciferol (vitamin D3) content. Br. Poult. Sci. 46:87–96.
Lotfi, A., R. Hauck, P. Olias, and H. M. Hafez. 2014. Pathogenesis of histomonosis in experimentally infected specific-pathogen-free (SPF) layer-type chickens and SPF meat-type chickens. Avian Dis. 58:427–432.
NRC. 1994. Nutrient Requirements of Poultry. 9th rev. ed. Natl. Acad. Press, Washington, DC.
Regmi, P. R., A. L. Shaw, L. L. Hungerford, J. R. Messenheimer, T. Zhou, P. Pillai, A. Omer, and J. M. Gilbert. 2016. Regulatory considerations for the approval of drugs against histomoniasis (blackhead disease) in turkeys, chickens, and game birds in the United States. Avian Dis. 60:725–730.
Rossi, A., R. Miles, B. Damron, and L. Flunker. 1993. Effects of dietary boron supplementation on broilers. Poult. Sci. 72:2124–2130.
Sander, J. E., L. Dufour, R. D. Wyatt, P. B. Bush, and R. K. Page. 1991. Acute toxicity of boric acid and boron tissue residues after chronic exposure in broiler chickens. Avian Dis. 35: 745–749.
Thøfner, I. C. N., D. Liebhart, M. Hess, T. W. Schou, C. Hess, E. Ivarsen, X. Fretté, L. P. Christensen, K. Grevsen, R. M. Engberg, and others. 2012. Antihistomonal effects of artemisinin and Artemisia annua extracts in vitro could not be confirmed by in vivo experiments in turkeys and chickens. Avian Pathol. 41:487–496.

56
FIGURES
Figure 1. BWG at A) Pre-challenge from d0-21 and B) post-challenge from d21-35. BWG data expressed as mean ± SEM and were analyzed using Student’s t-test in JMP Pro 14. C) Percentage of mortalities associated with histomoniasis post-challenge. No difference was detected when analyzed with chi square test. Cumulative lesion scores associated with histomoniasis beginning from d10-d14 post-challenge for D) liver and E) cecae. Lesions score of “0” indicative of a healthy bird with no disease whereas a score of “3” indicates classically severe histomoniasis. No differences between mean lesion scores were detected with SAS Proc Mixed Procedure for either liver or cecal lesions. Numbers within columns indicate the total poults per evaluated lesion score.

57
V. DATA CHAPTER 3
Evaluation of a candidate live-attenuated histomoniasis vaccine
L.C. Beer*, J.D. Latorre*, G. Tellez*, B.M. Hargis*, and C.N. Vuong*
*University of Arkansas: Division of Agriculture, Poultry Science Dept, Fayetteville, AR 72701
This manuscript will be submitted to Poultry Science

58
ABSTRACT
No vaccines are currently available to alleviate histomoniasis, a protozoal disease
primarily affecting turkeys. Three experiments evaluated a live-attenuated vaccine candidate
(VC) H. meleagridis. At selected dosages and time-points, turkeys (n=~40/group) were
vaccinated via different routes (oral vs. cloacal) and subsequently challenged with a virulent
wild-type (WT) H. meleagridis.
Experiment 1 included Negative Control and intracloacally-administered VC or WT on
d14 (2x105 Histomonas cells/poult). On d29, a subset of Negative Control and all VC were WT-
challenged. On d11 post-challenge, liver and cecal lesions were tabulated. Mortalities and
lesions were significantly lower (p≤0.05) in the VC compared to the WT Positive Control.
Experiment 2 included Negative Control, VC Oral (doses: 2 x103 or 2x104), VC Cloacal
(doses: 2x103, 2x104, or 2x105), or WT Cloacal Positive Control (2x105). On day-of-hatch,
vaccinations were administered pre-feeding via their respective routes and doses. On d21, VC
groups were challenged intracloacally with 2x105 WT. Two challenged-only control groups were
introduced on d21, receiving the 2x105 WT either cloacally or orally for the first time. Lesions
were evaluated d14 post-challenge. Mortalities and lesions were higher in all groups as compared
to the orally-administered WT. No differences in lesions were observed between the VC groups
and the Cloacal WT Positive Control.
Experiment 3 included Negative Control, VC Cloacal (d0 or d14), VC Oral (d0), or WT
Cloacal Positive Control (d0, d14, or d28). A dose of 2x105 was used for both the VC and WT
challenge. On d28, all treatments were challenged intracloacally with WT, and lesions evaluated
d14 post-challenge. Mortalities and liver lesions were lower in the d14 VC Cloacal as compared

59
to WT Cloacal Positive Control. Cecal lesions were not different in any VC treatment group as
compared to the WT Cloacal Positive Control.
Although histomoniasis was not completely prevented, d14 intracloacal administration of
the VC reduced of lesions and mortalities in experiments 1 and 3. Taken together, these data
encourage further research with the possibility that an attenuated strain could be efficacious for
lessening the impact of histomoniasis in turkeys.
Key Words: blackhead, histomoniasis, Histomonas meleagridis, turkey, vaccination

60
INTRODUCTION
Histomoniasis, also commonly known as blackhead, is an important protozoal disease
pertaining primarily to turkeys from the etiological origin of Histomonas meleagridis (Clarkson,
1963; Hess and McDougald, 2013). Flock mortality due to histomoniasis can approach 80-100%,
indicating the substantial economic importance of this affliction (Callait et al., 2002;
McDougald, 2005; Hess and McDougald, 2013). Direct contact between infected poultry or fecal
droppings results in rapid transmission and can occur without an intermediate host or vector if
transmission occurs before environmental degradation of the histomonad (Sentíes-Cué et al.,
2009). The primary method of transmission is considered to be through cloacal drinking that can
quickly transfer disease through the cloaca into the bursa of Fabricius or ceca via rhythmic
contractions (Hu et al., 2004; McDougald and Fuller, 2005). Recently, oral administration of a
clonal in vitro cultivated H. meleagridis administered to 1-day-old turkeys followed by 5h feed
withdrawal resulted in mortalities associated with histomoniasis; the oral route of poultry in
contact with large amounts of contaminated excreta or litter should be further considered
(Liebhart and Hess, 2009).
Limited research within the past 30 years combined with the ban of therapeutic and
prophylactic compounds has led to the absence of effective methods for controlling this disease
(van der Heijden et al., 2005; Hess et al., 2006). Nitarsone (Histostat), the last remaining FDA-
approved drug for treatment of histomoniasis, was withdrawn from the market in 2015 due to
concerns over detectable levels of heavy metals retained in meat products from treated poultry
(Regmi et al., 2016). In vitro and in vivo studies continue to yield variable results with no
alternatives introduced to replace the recently banned effective compounds such as the

61
nitroimidazoles, nitrofurans, and arsenical compounds (Thøfner et al., 2012). Moreover, no
vaccines are currently available to mitigate this disease.
Results from previous immunological research has been unsuccessful, contributing to
doubts over the possibility of vaccine development for prevention of histomoniasis (Hu and
McDougald, 2004). However, in 1963, Joyner treated turkeys suffering from acute histomoniasis
with dimetridazole and reported resistance to subsequent infection in the recovered turkeys,
suggesting the development of protective immunity. Cuckler (1970) reported turkeys recovered
from H. meleagridis-infection were resistant to subsequent challenge, even with maintained
presence of H. meleagridis within the cecae. Early studies by Tyzzer (1934) expressed the
reduction of virulence within long-term in vitro cultivated H. meleagridis, although
immunization results yielded conflicting success. More recent studies with chickens and turkeys
have indicated reduced liver and cecal lesions following intracloacal administration of clonal in
vitro attenuated H. meleagridis lines (Hess et al., 2008; Liebhart et al., 2013). Stable attenuation
with no reversion to virulence was demonstrated with a H. meleagridis that was passaged 295
times in vitro and subsequently serially passaged 5 times in vivo in turkeys and chickens
(Sulejmanovic et al., 2013). Furthermore, cross-protection against heterologous virulent isolates
was demonstrated by vaccinating with an attenuated clonal strain of H. meleagridis developed
through prolonged in vitro culture methods (Sulejmanovic et al., 2016). Nguyen Pham et al.
(2013) cloacally inoculated turkeys with a low-virulent H. meleagridis strain that was obtained
via serial passage in turkeys and showed induced protection following subsequent challenge with
a virulent H. meleagridis. Moreover, Liebhart et al. (2010) demonstrated a protective effect of an
in vitro attenuated H. meleagridis administered orally to 1-day-old turkeys. Development of a
vaccine against histomoniasis is encouraged by these immunological research advances.

62
Therefore, the objective of this study was to evaluate an H. meleagridis strain that was passaged
in vitro for immune protection in an experimental histomoniasis challenge model and to further
elucidate the possible routes and age for administration.
MATERIALS AND METHODS
Animal Source
Poults were tagged individually and randomly allocated to floor pens at the University of
Arkansas Poultry Health Laboratory on day-of-hatch. All animal handling procedures were in
compliance with the Institutional Animal Care and Use Committee (IACUC protocol #18113 and
#19032) of the University of Arkansas. A corn-soy based starter feed that met or exceeded
nutrient requirements of poultry (NRC, 1994) and water were provided ad libitum. Early poult
mortalities unrelated to histomoniasis were recorded and the altered group numbers are reported
in the experiments.
Histomonas meleagridis Isolate and Culture
A virulent wild-type (WT) H. meleagridis was obtained and cultivated based upon
previously published methods (van der Heijden et al., 2005; van der Heijden and Landman,
2007). The WT was passaged approximately 80 times and chosen for evaluation as a live-
attenuated vaccine candidate (VC). Modified Dwyer’s Media (MDM) was used for H.
meleagridis cultivation (Gibco, Life Technologies Corporation, USA; Lonza Walkersville Inc.;
USA), supplemented with 10% heat-inactivated horse serum (Gibco) and 1.6mg/mL white rice
flour. Histomonads were incubated anaerobically at 40°C for 48-72h before 1mL was sub-
cultured into 12.5mL of fresh, supplemented MDM. Growth was confirmed by observation with
an inverted microscope or enumeration on a hemocytometer. For long-term preservation of H.
meleagridis, 10% dimethylsulfoxide (OmniSolv, MilliporeSigma, USA) was added as a

63
cryoprotectant. The suspension was then distributed into cryogenic vials (VWR, USA) and
allowed to freeze at approximately -1°C/min under controlled conditions until reaching -80°C, at
which time vials were transferred to long-term storage in a liquid nitrogen tank. Approximately 6
days before needed for challenge or vaccination, the desired H. meleagridis aliquot was retrieved
from the liquid nitrogen and cultured into fresh supplemented media. Passages occurred every
48-72h. Following incubation, viable histomonads/mL were enumerated on a hemocytomer.
Within each experiment, MDM was utilized as the diluent to prepare the proper dosage
concentration.
Lesion Scoring System
The lesion score system was developed and initiated during “phase 2” of experiment 1
and continued throughout the remainder of the study. Classic lesions associated with
histomoniasis were evaluated on a scale of “0-3”, with “3” being the most severe (Figure 1). The
individuals determining the lesion scores were blinded to the treatment groups. All mortalities
were evaluated for liver and cecal lesions. According to this scale, a liver score of: “0” presents
with no detectable H. meleagridis-related lesions, “1” indicates detectible lesions that are not
clinically relevant (not significant ongoing pathology), “2” signifies intermediate lesions
suggesting significant pathology but not imminent mortality, while “3” denotes confluent or
nearly confluent lesions deemed likely to be fatal. The cecae were observed and palpated from
the serosal surface (the mucosa was not evaluated). According to the scale, a cecal score of: “0”
indicates no detectable H. meleagridis-related lesions observed from serosal inspection and
palpation, “1” denotes thickening (not clinically significant) of the cecae, “2” indicates clinically
meaningful cecal wall thickening without cecal cores, “3” specifies classic typhlitis with
thickened cecal walls, inflammation, and cecal cores.

64
Experiment 1
Phase 1 (d14-29). A total of 120 day-of-hatch female turkey poults were obtained from a
local commercial hatchery. Groups included a non-challenged Negative Control (n=59), the
Vaccine Candidate (VC) (n=39), and the Wild-Type (WT) Challenged Positive Control (n=20).
On d14, the VC group was inoculated with an in vitro attenuated (passage ~80) H. meleagridis at
a total dose of 2x105 histomonads/poult, and the WT Positive Control group received a total dose
of 2x105 virulent WT histomonads/poult (Figure 2). Inoculations were administered
intracloacally with an animal gavage needle and occurred twice with approximately 1h between
each inoculation. On d27 (13d post-challenge), the WT Positive Control group from “phase 1”
was euthanized to evaluate characteristic disease lesions and used for comparison against the
VC-induced lesions. On d28 (d14 post-vaccination), a subset of n=5 poults was sampled from
both the Negative Control and VC groups to evaluate for lesions associated with histomoniasis.
Phase 2 (d29-40). On d29 (d15 post-vaccination), all poults except the Negative Control
were challenged with the WT using the previously mentioned procedure. A newly introduced
d29 WT Cloacal Positive Control was created from a subset of the Negative Control group to
serve as concurrent reference against the VC group. All mortalities were evaluated for
characteristic liver and cecal lesions pertaining to histomoniasis. On d40 (d11 post-challenge), all
remaining poults were euthanized by CO2 inhalation and lesions were tabulated according to the
method described above.
Experiment 2
A total of 280 day-of-hatch female turkey poults were obtained from a local commercial
hatchery and allocated to groups. Treatments included non-challenged Negative Control (n=34),
VC Oral 2x103 (n=36), VC Oral 2x104 (n=38), VC Cloacal 2x103 (n=36), VC Cloacal 2x104

65
(n=37), VC Cloacal 2x105 (n=30), and WT Cloacal Positive Control 2x105 histomonads/poult
(n=36). In addition, a total of 30 day-of-hatch poults were humanely euthanized for pH
measurement of the combined proventriculus-ventriculus region using pH indicator strips
(Sigma-Aldrich, St. Louis, MO, USA).
Phase 1 (d0-21). On day-of-hatch, poults were vaccinated prior to feeding with selected
doses of either 2x103, 2x104, or 2x105 histomonads/poult with in vitro attenuated (passage ~80)
H. meleagridis administered either orally or intracloacally (Figure 4). The WT Cloacal Positive
Control received 2x105 histomonads/poult of WT, virulent H. meleagridis via intracloacal
administration according to the method described in experiment 1. On d15, a sample from each
group was evaluated for lesions to compare the VC to the WT group, leaving a remaining subset
of n=20 from each VC treatment group for the following challenge “phase 2.”
Phase 2 (d21-35). A WT Oral Positive Control (n=14) and WT Cloacal Positive Control
(n=20) were introduced on d21 and were created from reallocation of the Negative Control
poults. On d21, all poults received intracloacal challenge with WT H. meleagridis at 2x105 total
histomonads/poult in a pair of inoculations, with the exception of the WT Oral Positive Control
which received this dose orally in a single administration. On d35 (d14 post-challenge), all
remaining poults were euthanized by CO2 inhalation and lesion scores were tabulated according
to the method described above.
Experiment 3
A total of 480 day-of-hatch female turkey poults were obtained from a local commercial
hatchery and allocated to groups. Treatments included Negative Control (n=217), d0 VC Oral
(n=53), d0 VC Cloacal (n=52), d0 WT Oral Positive Control d0 (n=60), and d0 WT Cloacal
Positive Control d0 (n=60). In addition, a total of 60 day-of-hatch poults were humanely

66
euthanized and the pH of the combined proventriculus-ventriculus region was measured in each
poult using pH indicator strips (Sigma-Aldrich, St. Louis, MO, USA).
Phase 1 (d0-14). On day-of-hatch and prior to feeding, poults received selected doses of
2x105 histomonads/poult of either the WT challenge or the in vitro attenuated (passage ~80) VC
H. meleagridis administered either orally or cloacally according to the method described above
(Figure 7). On d14, all poults from the WT Oral Positive Control and WT Cloacal Positive
Control were evaluated for liver and cecal lesions. A total of n=10 poults/group were likewise
evaluated from the Negative Control, d0 VC Oral, and d0 VC Cloacal groups to compare to the
WT lesion presence.
Phase 2 (d14-28). Treatments included Negative Control (n=108), d0 VC Oral (n=43), d0
VC Cloacal (n=42), d14 VC Cloacal (n=56), and d14 WT Cloacal Positive Control (n=43). On
d14, the newly introduced WT Cloacal Positive Control and VC Cloacal group received 2x105
histomonads/poult of WT or VC H. meleagridis strain administered intracloacally, respectively.
On d28, all poults from the d14 WT Cloacal Positive Control were evaluated for liver and cecal
lesions. A total of n=10 poults/group were likewise evaluated from the Negative Control and d14
VC Cloacal groups. A total of n=5 poults/group were evaluated from the d0 VC Oral and d0 VC
Cloacal groups.
Phase 3 (d28-42). Treatments included Negative Control (n=53), d0 VC Oral (n=38), d0
VC Cloacal (n=37), d14 VC Cloacal (n=45), and d28 WT Cloacal Positive Control (n=45). On
d28, all poults except for the Negative Control, received 2x105 cells/poult of the WT H.
meleagridis challenge via intracloacal administration. On d42, all remaining poults were
evaluated for liver and cecal lesions.

67
Statistical Analysis
Differences in mortalities associated with histomoniasis were analyzed using the chi
square test with p≤0.05 considered significant. Fisher’s Exact test (2018 GraphPad Software,
LLC) was performed to further examine the mortality relationship between the Cloacal VC
2x105 group and the Cloacal WT Positive Control when difference was not apparent via the chi
square test. Lesion score data were analyzed using the Proc Mixed Procedure in SAS 9.4
software with significance between mean lesion score values denoted at p-values of ≤0.05,
≤0.005, or ≤0.0005 in comparison to the WT Positive Control.
RESULTS
Experiment 1
Phase 1 (d14-28). No mortalities associated with histomoniasis occurred in the Negative
Control or VC, whereas the WT Positive Control reached 30% mortality before euthanasia on
13d post-challenge (d27) (Figure 3A). No indications of histomoniasis were observed in the
Negative Controls at any time. Cumulative lesions from mortalities and scheduled termination
date of the WT Positive Control revealed 75% liver and 80% cecal lesions characteristic of
histomoniasis. On d28 (d14 post-vaccination), among the n=5 VC subset examined, one VC
poult exhibited normal liver and cecae under gross examination, whereas two poults had
presented with normal livers but relatively normal cecae with the exception of small, button-like
lesions. One poult had target-like liver lesions with the cecae feeling hard, thickened, and
exhibiting larger bumps and scalloping. The fifth VC poult exhibited paling edges to the liver,
narrow and thin margins, and was possibly beginning to develop liver lesions. In addition, the
cecae were large, with the presence of thickened walls and scalloping.

68
Phase 2 (d29-40). Mortalities relating to histomoniasis within the Negative Control, VC,
and WT Positive Control were 0.00, 2.94, and 22.2%, respectively (Figure 3B). The Negative
Control and VC mortalities were different (p≤0.05) as compared to the WT Positive Control. The
VC displayed lower liver (p≤0.0005) and cecal (p≤0.005) lesions than the WT Positive Control
(Figures 3C and 3D). No lesions or mortalities associated with histomoniasis were observed in
the Negative Control.
Experiment 2
Phase 1 (d0-21). A mean pH of 4.4 was determined from the proventriculus-ventriculus
region from the day-of-hatch poults. Mortalities related to histomoniasis in the Negative Control,
VC Oral 2x103, VC Oral 2x104, VC Cloacal 2x103, VC Cloacal 2x104, VC Cloacal 2x105, and
WT Cloacal Positive Control were 0.00, 2.78, 2.63, 0.00, 5.41, 3.33, and 22.2%, respectively
(Figure 5A). Mortalities in the Negative Control and all VC groups were lower (p≤0.05) than the
WT Cloacal Positive Control. Liver and cecal lesions were lower (p≤0.0005) in all VC Oral and
VC Cloacal groups as compared to the WT Cloacal Positive Control (Figures 5B and 5C).
Phase 2 (d21-35). Mortalities related to histomoniasis in the WT Oral Positive Control,
VC Oral 2x103, VC Oral 2x104, VC Cloacal 2x103, VC Cloacal 2x104, VC Cloacal 2x105, and
WT Cloacal Positive Control were 0.00, 50.0, 50.0, 60.0, 55.0, 35.0, and 55.0%, respectively
(Figure 6A). The WT Oral Positive Control had lower (p≤0.05) mortalities as compared to the
WT Cloacal Positive Control. No differences in mortalities were found between any of the VC
treatments as compared to the WT Cloacal Positive Control when evaluated with a chi square
test. To further elucidate any possible difference between the VC Cloacal 2x105 group as
compared to the WT Cloacal Positive Control, a Fisher’s Exact test was computed, resulting in
p=0.3406. The WT Oral Positive Control resulted in lower (p≤0.0005) liver and cecal lesions as

69
compared to the WT Cloacal Positive Control (Figures 6B and 6C). The WT Oral Positive
Control received only lesion scores of “0”, indicating no detectable lesions associated with
histomoniasis. No differences in liver or cecal lesions were found between any of the VC groups
as compared to the WT Cloacal Positive Control. The p-values for the VC Cloacal 2x105 as
compared to the WT Cloacal Positive Control for liver and cecal lesions were p=0.1610 and
p=0.6793, respectively.
Experiment 3
Phase 1 (d0-14). A mean pH of 5.0 was determined from the proventriculus-ventriculus
region from the day-of-hatch poults. No mortalities associated with histomoniasis occurred in the
Negative Control, VC Oral, or VC Cloacal groups (Figure 8A). The d0 WT Oral Positive
Control and d0 WT Cloacal Positive Control resulted in 16.7 and 15% mortalities by d14 post-
challenge, respectively. The d0 WT Oral Positive Control was not different in liver or cecal
lesions (p=0.0908 and p=0.2360, respectively) as compared to the d0 WT Cloacal Positive
Control (Figures 8B and 8C). The d0 VC Oral and d0 VC Cloacal were lower in liver (p≤0.005)
lesions and cecal (p≤0.0005) lesions as compared to the d0 WT Cloacal Positive Control.
Negative Control was lower (p≤0.0005) in liver and cecal lesions as compared to the d0 WT
Cloacal Positive Control.
Phase 2 (d14-28). No mortalities associated with histomoniasis occurred in the Negative
Control, d0 VC Oral, d0 VC Cloacal, or d14 VC Cloacal groups by 28-days-of-age. The d14
introduced WT Cloacal Positive Control reached 48.8% mortality by d14 post-challenge (Figure
9A). The d0 VC Oral, d0 VC Cloacal, d14 VC Cloacal, and Negative Control were lower
(p≤0.05) in liver and cecal lesions as compared to the d14 WT Cloacal Positive Control (Figures
9B and 9C).

70
Phase 3 (d28-42). No mortalities or lesions associated with histomoniasis occurred in the
Negative Control. Mortalities in the d0 VC Oral, d0 VC Cloacal, d14 VC Cloacal, and d28 WT
Cloacal Positive Control reached 44.7, 32.4, 22.2, and 42.2%, respectively (Figure 10A). The
d14 VC Cloacal group resulted in lower (p≤0.05) mortalities than the d28 WT Cloacal Positive
Control. Liver and cecal lesions were different (p≤0.0005) between the Negative Control and
d28 WT Cloacal Positive Control. The d14 VC Cloacal group exhibited lower (p≤0.0005)
cumulative liver lesions than the d28 WT Cloacal Positive Control (Figure 10B). Although not
statistically significant (p=0.0899), the d14 VC Cloacal group somewhat reduced cecal lesions as
compared to the d28 WT Cloacal Positive Control (Figure 10C). No differences were detected
in liver or cecal lesions between any of the other VC groups as compared to the d28 WT Cloacal
Positive Control.
DISCUSSION
The VC lessened the severity of histomoniasis when administered at a total dosage of
2x105 histomonads/turkey on d14 in experiment 1. The decreased mortalities following WT
challenge and the lack of vaccine-related mortalities suggested that this passage strain of H.
meleagridis might be attenuated enough to initiate immune response within the turkey without
resulting in lethality due to the VC. Although disease was not completely prevented, the lowered
lesions and mortalities as compared to the WT Cloacal Positive Control suggest that some
protection was offered via the VC. Within all of the experiments, the Negative Controls never
exhibited mortalities or lesions associated with histomoniasis, further confirming that disease can
be prevented by management and absence of exposure.
In experiment 2, low vaccine-related mortalities associated with histomoniasis occurred
in the oral (doses of 2x103 and 2x104) and cloacal (doses of 2x103 and 2x105) VC which could

71
potentially be explained since the H. meleagridis VC is not a clonal population. Variation may
be occurring each time the cells are propagated, with greater or fewer virulent cells present at
various concentrations depending upon the passage. During phase 1 of experiment 2, lesion
presence was lower in the subsets of VC turkeys regardless of dose or route when compared to
the WT Cloacal Positive Control, suggesting that the VC H. meleagridis was still capable of
invading the tissues without causing characteristic rampant lesions. Although prolonged in vitro
passaging of H. meleagridis has been reported to decrease vaccination efficacy against
pathogenic strains, stable attenuation occurred in other studies without reversion to virulence
(Lund and Chute, 1967; Sulejmanovic et al., 2013).
Oral transfer of H. meleagridis should not be overlooked as previous studies with
chickens have demonstrated that feed deprivation and an alkaline pH prior to oral challenge were
required in order to develop lesions characteristic with histomoniasis (Cuckler, 1970).
Interestingly, WT oral challenge on d21 resulted in no mortalities or lesions associated with
histomoniasis, consistent with the prevailing thoughts that H. meleagridis cannot survive the low
pH within the proventriculus and ventriculus typically present following feed consumption.
Conversely, in experiment 3, turkeys that were WT challenged orally on d0 were equally
susceptible to H. meleagridis infection with no differences noted in mortalities or lesions as
compared to the WT Cloacal Positive Control. Following feed ingestion, the average pH in the
proventriculus/ventriculus has been reported as 3.5 in broiler chickens with variability between
1.9 and 4.5 (Svihus, 2011). The susceptibility of the poults on day-of-hatch pre-feeding could
likely be explained by the higher pH (measured as 4.4 and 5.0 in experiments 2 and 3,
respectively) within the proventriculus/ventriculus region, indicating a more basic environment
for which the protozoa could potentially survive until parasitizing the cecae. If repeated in future

72
studies, pH should be measured in a subset of turkeys post-feeding to compare to pre-feeding
measurements. Unexpectedly, the subset of the d0 VC Oral (dose 2x105) in experiment 3
exhibited some mild to severe lesions characteristic with histomoniasis when evaluated on d28
post-vaccine, indicating the ability of the VC H. meleagridis to invade tissue although no
mortalities related to the disease occurred during this timeframe.
Some turkeys within the WT Cloacal Positive Control group in each experiment
exhibited lesion scores of “0”, indicating no abnormality or pathology of histomoniasis. The
absence of lesions within a subset of the WT Cloacal Positive Control could potentially be
explained in that some turkeys may be less susceptible to challenge, cecal retrograde of the
parasite inoculum could vary between turkeys, or some turkeys may excrete the inoculum before
cloacal uptake occurs. Moreover, the absence of clonal culture could contribute to prevalence of
certain cell populations with greater or lower degrees of virulence. Although passages of the WT
H. meleagridis are minimized to prevent in vitro attenuation in culture, a chance of virulence or
population changes are present with each propagation. To reduce this concern, both the WT and
VC should be single-cell cloned and evaluated to ensure the same genetic population is being
evaluated in subsequent experiments.
Liebhart et al. (2010) demonstrated protectiveness of an in vitro attenuated H.
meleagridis administered to 1-day-old turkeys. However, under the conditions of these
experiments, the cloacal route appears to be the more efficacious route as compared to the oral
route for administration, especially at an older age. Administration of the VC H. meleagridis
intracloacally on d14 resulted in lower mortalities and lesions in both experiments 1 and 3,
suggesting that this might be an efficacious alternative for the prevention of histomoniasis.
Intracloacal inoculation with attenuated H. meleagridis has shown induced protection as well as

73
cross-protective capability against virulent isolates (Nguyen Pham et al., 2013; Sulejmanovic et
al., 2016). Further research should be conducted to elucidate the most efficacious route and
dosage of the VC H. meleagridis as well as the proper age for administration. It should be noted
that other passage isolates of H. meleagridis have been evaluated by other research groups and
shown to induce protection against subsequent challenge, as outlined above. Additionally, if our
selected VC is efficacious at alleviating histomoniasis and is successfully single-cell cloned, we
will need to address a major limiting factor to larger scale production of the live-attenuated H.
meleagridis strain, specifically more efficient methods to feasibly propagate cells to meet
commercial production needs. The current cell culture methods seem unfeasible for mass
production, as the cells grow at varied rates and the media is relatively costly. Moreover, for
introduction to the industry, the intracloacal route would not be the most practical technique for
large-scale application. The d0 administration would be preferable for industry application for
incorporation within the hatchery. Presumably, oral route at day-of-hatch seems the most
efficient, if possible. However, if the cloacal route proves to be the only effective administration
to induce a robust immune response, then potentially an alternative method could be developed
for cloacal application during the beak trimming, toenail removal, or sex determination
procedure at the hatchery. Certainly, a main consideration is the identification of an effective
prophylaxis for this life-threatening disease of poultry. The current lack of any approved
prophylactic measures against histomoniasis suggests that any contribution to the mitigation of
this disease is substantial to improving animal food production and reducing this emerging
problem.

74
REFERENCES
Callait, M., C. Granier, C. Chauve, and L. Zenner. 2002. In vitro activity of therapeutic drugs against Histomonas meleagridis (Smith, 1895). Poult. Sci. 81:1122–1127.
Clarkson, M. 1963. Immunological responses to Histomonas meleagridis in the turkey and fowl.
Immunology. 6:156-168.
Cuckler, A. 1970. Coccidiosis and histomoniasis in avian hosts. Pages 371-397 in Immunity to parasitic animals. GJ Jackson, R. Herman and I. Singer, eds., New York.
Van der Heijden, H. M. J. F., and W. J. M. Landman. 2007. Improved culture of Histomonas
meleagridis in a modification of Dwyer medium. Avian Dis. 51:986–988.
Van der Heijden, H. M. J. F., L. R. McDougald, and W. J. M. Landman. 2005. High yield of parasites and prolonged in vitro culture of Histomonas meleagridis. Avian Pathol. 34:505–508.
Hess, M., D. Liebhart, E. Grabensteiner, and A. Singh. 2008. Cloned Histomonas meleagridis
passaged in vitro resulted in reduced pathogenicity and is capable of protecting turkeys from histomonosis. Vaccine. 26:4187–4193.
Hess, M., and L. McDougald. 2013. Histomoniasis (blackhead) and other protozoan diseases of the intestinal tract. Pages 1172–1178 in Diseases of Poultry.13th ed. E. Swayne, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. L. Suarez, and V. L. Nair, eds., Wiley-Blackwell, Ames, IA.
Hu, J., L. Fuller, and L. R. McDougald. 2004. Infection of turkeys with Histomonas meleagridis
by the cloacal drop method. Avian Dis. 48:746–750.
Hu, J., and L. McDougald. 2004. The efficacy of some drugs with known antiprotozoal activity against Histomonas meleagridis in chickens. Vet. Parasitol. 121:233–238.
Liebhart, D., and M. Hess. 2009. Oral infection of turkeys with in vitro-cultured Histomonas meleagridis results in high mortality. Avian Pathol. 38:223–227.
Liebhart, D., T. Sulejmanovic, B. Grafl, A. Tichy, and M. Hess. 2013. Vaccination against histomonosis prevents a drop in egg production in layers following challenge. Avian Pathol. 42:79–84.
Liebhart, D., M. Windisch, and M. Hess. 2010. Oral vaccination of 1-day-old turkeys with in vitro attenuated Histomonas meleagridis protects against histomonosis and has no negative effect on performance. Avian Pathol. 39:399–403.
Lund, E. E. A. P. C., and A. M. Chute. 1967. Histomonas meleagridis after one thousand in vitro
passages. J. Eukaryot. Microbiol. 14:349–351.

75
McDougald, L. R. 2005. Blackhead disease (histomoniasis) in poultry: A critical review. Avian Dis. 49:462–476.
McDougald, L., and L. Fuller. 2005. Blackhead disease in turkeys: Direct transmission of
Histomonas meleagridis from bird to bird in a laboratory model. Avian Dis. 49:328–331.
Nguyen Pham, A. D., J. K. De Gussem, and B. M. Goddeeris. 2013. Intracloacally passaged low-virulent Histomonas meleagridis protects turkeys from histomonosis. Vet. Parasitol. 196:307–313.
NRC. 1994. Nutrient Requirements of Poultry. 9th rev. ed. Natl. Acad. Press, Washington, DC.
Regmi, P. R., A. L. Shaw, L. L. Hungerford, J. R. Messenheimer, T. Zhou, P. Pillai, A. Omer,
and J. M. Gilbert. 2016. Regulatory considerations for the approval of drugs against histomoniasis (blackhead disease) in turkeys, chickens, and game birds in the United States. Avian Dis. 60:725–730.
Sentíes-Cué, G., R. Chin, and H. Shivaprasad. 2009. Systemic histomoniasis associated with high mortality and unusual lesions in the bursa of Fabricius, kidneys, and lungs in commercial turkeys. Avian Dis. 53:231–238.
Sulejmanovic, T., I. Bilic, M. Hess, and D. Liebhart. 2016. An in vitro attenuated strain of Histomonas meleagridis provides cross-protective immunity in turkeys against heterologous virulent isolates. Avian Pathol. 45:46–53.
Sulejmanovic, T., D. Liebhart, and M. Hess. 2013. In vitro attenuated Histomonas meleagridis does not revert to virulence, following serial in vivo passages in turkeys or chickens. Vaccine. 31:5443–5450.
Svihus, B. 2011. The gizzard: Function, influence of diet structure and effects on nutrient
availability. Worlds Poult. Sci. J. 67:207-224.
Thøfner, I. C. N., D. Liebhart, M. Hess, T. W. Schou, C. Hess, E. Ivarsen, X. Fretté, L. P. Christensen, K. Grevsen, R. M. Engberg, and others. 2012. Antihistomonal effects of artemisinin and Artemisia annua extracts in vitro could not be confirmed by in vivo experiments in turkeys and chickens. Avian Pathol. 41:487–496.
Tyzzer, E. E. 1934. Studies on histomoniasis, or“ blackhead” infection, in the chicken and the turkey. Daedalus. 69:189–264.
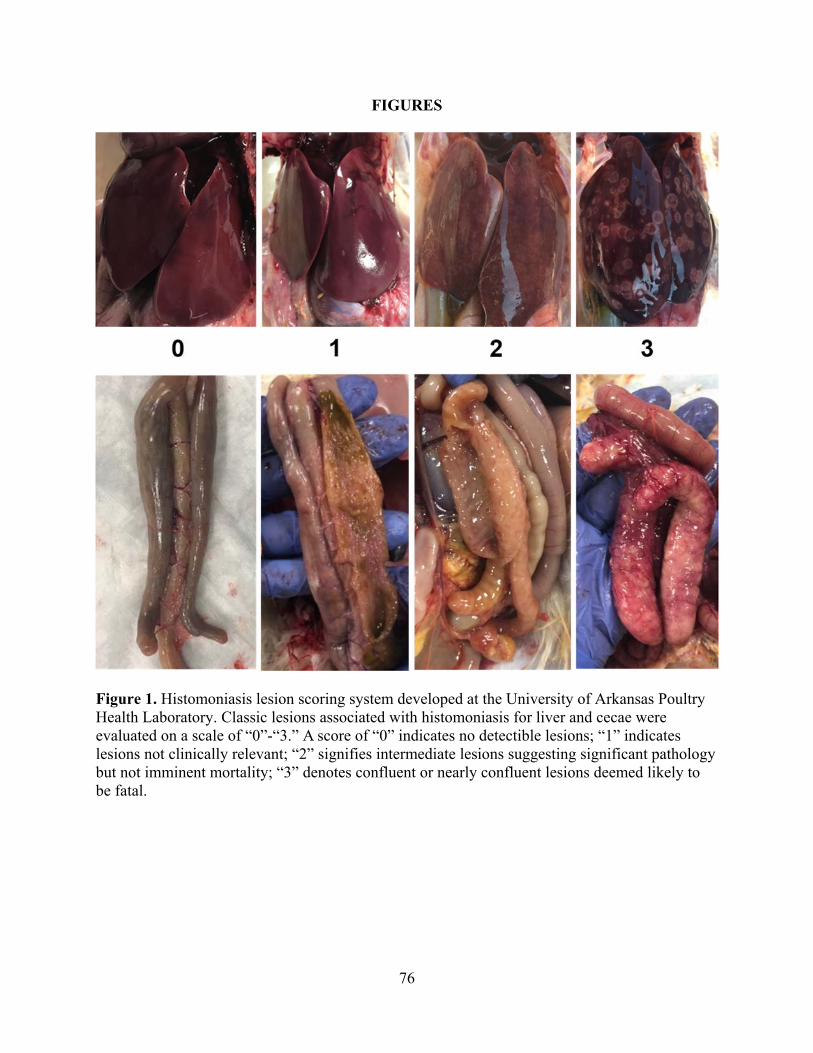
76
FIGURES
Figure 1. Histomoniasis lesion scoring system developed at the University of Arkansas Poultry Health Laboratory. Classic lesions associated with histomoniasis for liver and cecae were evaluated on a scale of “0”-“3.” A score of “0” indicates no detectible lesions; “1” indicates lesions not clinically relevant; “2” signifies intermediate lesions suggesting significant pathology but not imminent mortality; “3” denotes confluent or nearly confluent lesions deemed likely to be fatal.

77
Figure 2. Timeline for Experiment 1. VC=Vaccine Candidate; WT=Wild-Type.

78
Figure 3. Experiment 1 cumulative percentage mortalities associated with histomoniasis at A) Phase 1 (d14-29) and B) Phase 2 (d29-40). Lesion scores on 11d post-challenge for C) liver and D) cecae. Statistical difference detected by the SAS Proc Mixed Procedure between mean lesion scores as compared to the Wild-Type (WT) Positive Control group is indicated by “*” for p≤0.05, “**” for p≤0.005, and “***” for p≤0.0005. Numbers within columns indicate the total poults per evaluated lesion score. VC=Vaccine Candidate.

79
Figure 4. Experiment 2 timeline. VC=Vaccine Candidate; WT=Wild-Type.

80
Figure 5. Experiment 2 response during Phase 1 (d0-21) for A) cumulative percentage of mortalities associated with histomoniasis, lesion scores of B) liver and C) cecae at 15d post-administration of Vaccine Candidate (VC) or Wild-Type (WT) Histomonas meleagridis. Statistical difference detected by the SAS Proc Mixed Procedure between mean lesion scores as compared to the Wild-Type (WT) Positive Control group is indicated by “*” for p≤0.05, “**” for p≤0.005, and “***” for p≤0.0005. Numbers within columns indicate the total poults per evaluated lesion score. VC=Vaccine Candidate.

81
Figure 6. Experiment 2 response during Phase 2 (d21-35) for A) cumulative percentage of mortalities associated with histomoniasis, lesion scores of B) liver and C) cecae beginning from d9-14 post-WT challenge. Statistical difference detected by the SAS Proc Mixed Procedure between mean lesion scores as compared to the Wild-Type (WT) Positive Control group is indicated by “*” for p≤0.05, “**” for p≤0.005, and “***” for p≤0.0005. Numbers within columns indicate the total poults per evaluated lesion score. VC=Vaccine Candidate.

82
Figure 7. Experiment 3 timeline. VC=Vaccine Candidate; WT=Wild-Type.

83
Figure 8. Experiment 3 response during Phase 1 (d0-14) for A) cumulative percentage of mortalities associated with histomoniasis, lesions scores of B) liver and C) cecae on d14 post-administration of Vaccine Candidate (VC) or Wild-Type (WT) Histomonas meleagridis. Statistical difference detected by the SAS Proc Mixed Procedure between mean lesion scores as compared to the Wild-Type (WT) Positive Control group is indicated by “*” for p≤0.05, “**” for p≤0.005, and “***” for p≤0.0005. Numbers within columns indicate the total poults per evaluated lesion score. VC=Vaccine Candidate.

84
Figure 9. Experiment 3 response during Phase 2 (d14-28) for A) cumulative percentage of mortalities associated with histomoniasis, lesion scores of B) liver and C) cecae. Statistical difference detected by the SAS Proc Mixed Procedure between mean lesion scores as compared to the Wild-Type (WT) Positive Control group is indicated by “*” for p≤0.05, “**” for p≤0.005, and “***” for p≤0.0005. Numbers within columns indicate the total poults per evaluated lesion score. VC=Vaccine Candidate.

85
Figure 10. Experiment 3 response during Phase 3 (d28-42) for A) cumulative percentage of mortalities associated with histomoniasis, lesion scores of B) liver and C) cecae. Statistical difference detected by the SAS Proc Mixed Procedure between mean lesion scores as compared to the Wild-Type (WT) Positive Control group is indicated by “*” for p≤0.05, “**” for p≤0.005, and “***” for p≤0.0005. Numbers within columns indicate the total poults per evaluated lesion score. VC=Vaccine Candidate.

86
VI. CONCLUSIONS
None of the dietary treatments of deoxycholic acid (DCA), biliogenic formulation, or
boric acid reduced disease within H. meleagridis-challenged turkeys. Although DCA exhibited
anti-histomonal properties in vitro, efficacy was not transferred in vivo under the experimental
conditions of this study. These results further confirm the complicated nature of this disease as
well as the importance of in vivo evaluation rather than reliance only upon in vitro methods.
Under the conditions of experiment 1 within the vaccination study, the Vaccine
Candidate (VC) Histomonas meleagridis resulted in lowered disease-related mortalities and
lesions as compared to the Wild-Type Cloacal Positive Control. Although histomoniasis was not
completely prevented, d14 intracloacal administration of the VC lowered lesions and mortalities
in experiments 1 and 3. Taken together, these data suggest the potential efficacy of the VC to
protect from histomoniasis. Further research should be conducted to elucidate the most effective
dosage, route, and age for administration of this VC.

87
APPENDIX

88