differentiated regions of human placental …jcs.biologists.org/content/joces/25/1/293.full.pdf ·...
TRANSCRIPT

y. Cell Sci. 25, 293-312 (1977) 293
Printed in Great Britain
DIFFERENTIATED REGIONS OF HUMAN
PLACENTAL CELL SURFACE ASSOCIATED
WITH EXCHANGE OF MATERIALS BETWEEN
MATERNAL AND FOETAL BLOOD:
COATED VESICLES
C. D.OCKLEFORD AND A. WHYTE
Department of Pathology, The University of Cambridge, Tennis Court Road,Cambridge CBz 1QP, England
SUMMARY
Coated vesicles may be an important component of the micropinocytic system of the humanplacenta. Regions of very dense reaction with glycocalyx stains are restricted to membraneswithin forming and fully formed coated vesicles. This is interpreted as evidence againstpermanently grouped specific binding sites having a role in the selective uptake of materials bymicropinocytosis, and as support for theories of coated-vesicle formation which take intoaccount the dynamic nature of membrane components. The pyroantimonate precipitationtechnique which was employed in an attempt to localize cations in placental tissue at termresulted in the deposition of electron-dense material in coated vesicles and basement membrane.
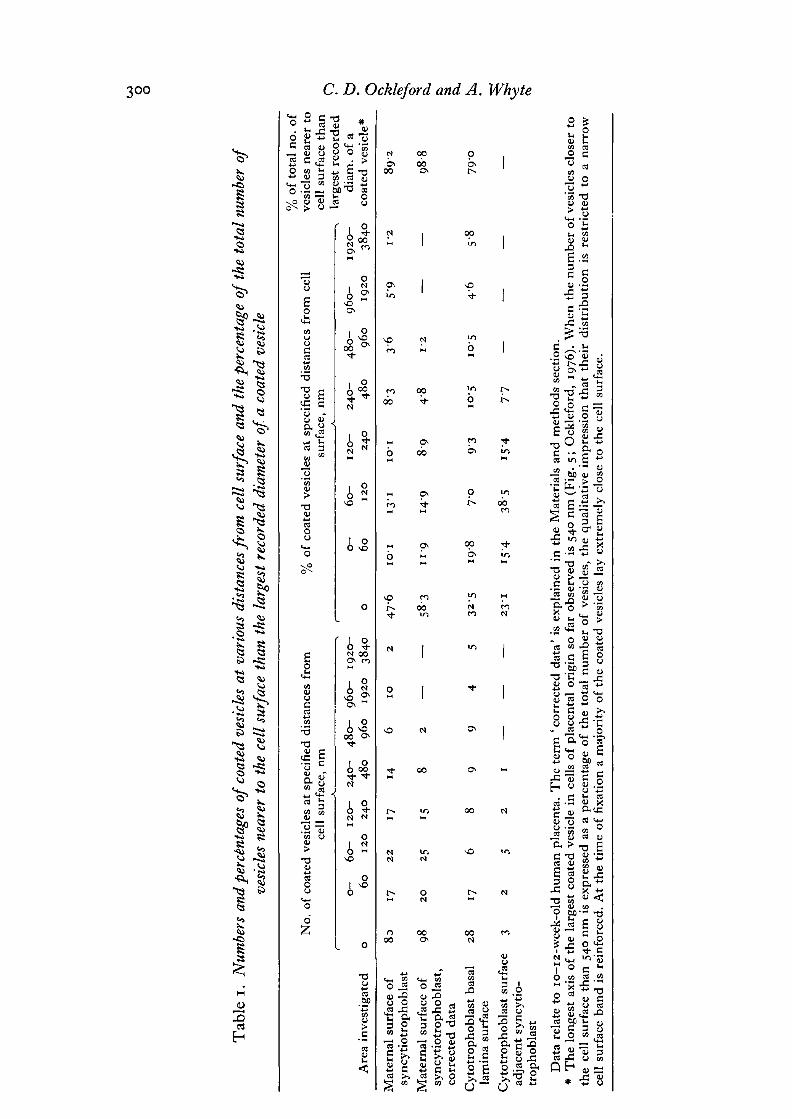
Examination of the distribution of coated vesicles in placental tissue explants at 8-13 weeksof gestation revealed a restricted distribution of these organelles. Probably more than 89 % ofcoated vesicles lie within the largest vesicles' diameter from the cell surface.
Placental coated vesicles were isolated and examined using negative staining. A polygonallypatterned structure was apparent on their surfaces. Analysis of the isolated fraction of coatedvesicles using sodium dodecyl sulphate polyacrylamide gel electrophoresis shows the presenceof a major protein of molecular weight 180000. This is the same molecular weight that has beengiven for clathrin, the major protein of the raised polygonally patterned structure on thecytoplasmic surface of coated vesicles from other sources.
INTRODUCTION
Much is known of the physiology of transport of materials into the human placenta(Miller & Berndt, 1975) but there is a lack of data on the organization of the dynamicstructures involved in uptake. Two major questions arise from the physiological work.Firstly, the processes of uptake are unexplained. Secondly, the placenta is able tocollect certain materials from their relatively dilute state in maternal blood and main-tain them at a higher concentration in the foetal circulation. Materials taken up andconcentrated in this way include proteins, polysaccharides, fats, certain vitamins andinorganic ions (Brambell, 1970; Llewellyn-Jones, 1969).
Allison & Davies (1974) have gathered a consensus of support for their view thatendocytic organelles are separable by their different structural natures. These differ-ences in structure may correlate with the physiologically distinct classes of endocyticactivity. For example, there are drugs which differentially suppress certain classes of

294 C. D. Ockleford and A. Whyte
activity (Wills, Davies, Allison & Haswell, 1972). In addition, different forms ofendocytosis have differing energy requirements (Simpson & Spicer, 1973). Micro-pinocytosis by coated vesicle formation represents an area where understanding of theunderlying motility is developing (Kanaseki & Kadota, 1969; Ockleford, 1976).
One approach to the problems of the motile processes of uptake in the placenta andthe phenomenon of selection is to study particular endocytic organelles. Coatedvesicles are a common constituent of cells and are found in the placenta. They occurwith great abundance in cells which selectively absorb protein (Lloyd & Beck, 1974).Since the human placenta takes up proteins selectively, there is a comparative cellularanatomical argument for making coated vesicles the organelles of choice in this study.
This research has been undertaken at a favourable moment in the developmentof techniques for the study of these organelles. Pearse (1975) has recently developeda method for the isolation of porcine brain coated vesicles. This has proved adaptableto the placenta and has enabled us to complement our fine-structural data fromsectioned material with structural data from negatively stained, isolated vesicles andwith data from polyacrylamide gel electrophoresis. Using this range of techniques wehave sought information that could be used to gain an understanding of the dynamicevents involved in selective uptake by coated-vesicle formation in the transportprocesses which support foetal development.
MATERIALS AND METHODS
Trophoblast tissue
Portions of human trophoblast tissue were recovered shortly after therapeutic terminationsof pregnancy performed in the 8-12th week. Groups of about 20 chorionic villi were isolatedby dissection and washed 3 times in Hanks' Balanced Salts Solution (BSS) (Flow Laboratories,Irvine, Scotland). Unless stated specifically to the contrary, this was the tissue used. Termtrophoblast was obtained post-partum and was treated similarly. Trophoblast cells werecultured as described by Loke & Borland (1970).
Transmission election microscopy
The methods routinely used for fixation and embedding and for ultrathin sectioning aredescribed fully in a previous publication (Ockleford, 1975).
Samples containing coated vesicles were negatively stained using 1 % uranyl acetate (Huxley,1963). They were supported on carbon-coated copper grids.
Chorionic villi were stained with ruthenium red by the method of Luft (1965); by adding tothe tissue a mixture of 1-8 ml of 10 % ruthenium red together with 23 ml of a mixture of thesolutions A and B described earlier (Ockleford, 1975); and by preincubating the tissue for10 min in i-8 ml of 10% ruthenium red and 23 ml of Hanks' BSS prior to fixation.
Chorionic villi were stained with Alcian blue by using a 1 % solution of the dye (Behnke &Zelander, 1970; Rothman, 1970).
The tissue was stained with colloidal iron using the method first used for isolated membraneby Marx, Graf & Wesemann (1973). Vibrio comma (cholerae) neuraminidase (Behringwerke A.G.Marburg Lahn) at activities of 50 and 500 U/ml was used to predigest one group of tissuesamples. Of these, some samples were esterified using CVIN HC1 dissolved in methanol andothers were esterified and later saponified using a 1 % solution of KOH in 80 % ethanol asthese authors have recommended (Marx et al. 1973).
For tannic acid fixation tissue-cultured trophoblast cells were fixed using 8 % tannic acid and

Coated vesicles of human placenta 295
a 1 % solution of the penetration agent digitonin (Tilney et al. 1973). The cells were embeddedusing Spurr's (1969) low viscosity resin.
Cation precipitation was accomplished using the method of Tandler, Libanati & Sanchis(1970). To increase the insolubility of the cationic precipitate the tissue was heated for 5 minat 95 CC in a half-saturated solution of potassium pyroantimonate and then rinsed twice in ice-cold distilled water as these authors recommended. Term placental tissue was used for cationprecipitation.
Distribution of coated vesicles
The surface of the syncytiotrophoblast where caveolae occur is highly convoluted. There-fore, it is not obvious whether some membrane-bounded electron-lucent areas represent truevacuoles within the cell or transverse sections through invaginating membrane pockets whichare still continuous with the cell surface. This complicates an analysis of the distribution ofcoated vesicles based upon measured distance from the cell surface. Consequently the data havebeen treated in 2 ways. Firstly, measurements have been made of the distance of coatedvesicles from the nearest definitely recognizable portion of the cell surface. This type of treat-ment gives an overestimate of the real average distance from the cell surface but gives a firmupper limit to the value. The second more uncertain method used any available indicator (forexample faint traces of contents or vesicle shape and size) to designate some ' apparent vesicles'as sections through invaginations of the cell surface membrane. Then, if coated vesicle profileswere closer to these than they were to the definitely recognizable cell surface, this new lowervalue was substituted in an attempt to achieve a more accurate estimate of the average distancefrom the cell surface. Only sections that were approximately perpendicular to the long axis ofthe villi were used. Any variation from the vertical in the section plane would be likely toincrease the apparent average distance of coated vesicles from the cell surface obtained usingeither method. Both potential errors lead to overestimation of the average distance of coatedvesicles from the cell surface.
A comparison of distribution and number of coated vesicles in cultured trophoblast cellswith and without 0-02 % ethylenediamine tetra-acetic acid (EDTA) added to the culturemedium was undertaken. The cells were incubated in the presence of EDTA at 37 °C for15 min; they were removed from the culture flasks by shaking or by means of a rubber police-man, fixed as described earlier and embedded in low-viscosity resin (Spurr, 1969).
Isolation of coated vesicles
Coated vesicles were isolated using the technique devised by Pearse (1975) with the followingmodifications. Tissue was disrupted (3 x 5-s periods) using an MSE homogenizer operated athalf speed. Samples were examined electron microscopically at all stages of purification.Coated vesicles were collected after the first sucrose density gradient step (5-60 %) for sub-sequent examination by electron microscopy and polyacrylamide gel electrophoresis.
SDS polyacrylamide gel electrophoresis
Electrophoresis of the proteins of the coated vesicle fraction was undertaken using 7-5 %polyacrylamide gels without stacking gels (Laemmli, 1970). Runs were initiated at 1 mA per gel.Once the dye front had migrated 1 cm into the gel the current was increased to 2 mA per gel.Gels were fixed for 30 min using 10 % glacial acetic acid in 50 % aqueous methanol. Protein-containing bands were stained for 30 min using a 1-25 % solution of Coomassie brilliant blue inthe same solvent. Gels were destained at 60 °C using several changes of 7 % acetic acid in 10 %aqueous methanol. Mobilities of the major protein bands were expressed according to theformula of Weber & Osborn (1969). Molecular weights of proteins from the coated-vesiclefraction were assessed from a calibration graph. This was plotted using the mobilities of proteinsof known molecular weight determined from gels run simultaneously with those containingcoated-vesicle protein. The proteins used for calibration were pepsin (Sigma), mol. wt, 35000;bovine serum albumin fraction V (Sigma), mol. wt, 68000; and /?-galactosidase (Sigma gradem ) , mol. wt of oligomer, 130000. Gels were scanned using a Pye Unicam SP1800 ultravioletrecording spectrophotometer operated at a wavelength of 540 nm and a slit height of 2 mm.
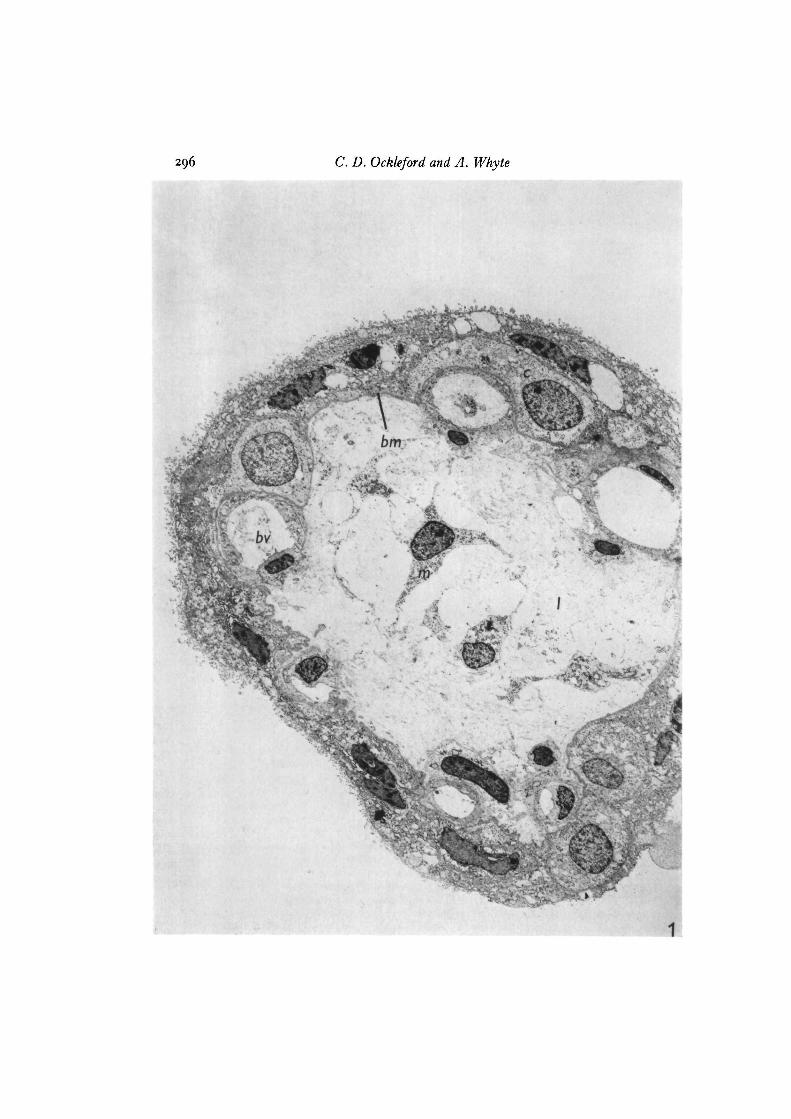
296 C. D. Ockleford and A. Whyte
\ •'- . \S- /•W'
''-•-^•J*&**&SJM

Coated vesicles of human placenta 297
RESULTS
Transmission electron microscopy of sectioned material
Coated vesicles occur at or near the cell surface of the syncytiotrophoblast of thehuman placenta. They are frequently observed near microvillous areas of the cellsurface (Figs, i, 2) but are rare enough to have escaped detection in areas underlyingcell surface which is devoid of microvilli. The microvillous border of the cell ispopulated with profiles of uncoated vesicles of similar size to the profiles of coatedvesicles (Fig. 2). The microvilli and the surface region of the cell contain actin-likemicrofilaments with a diameter of about 9 nm. The rather disordered nature of thesemicrovilli compared for example with those of intestine is reflected at the macro-molecular level (Fig. 2). The microfilaments in placental microvilli are not packed inparallel in the regular hexagonal arrays seen in microvilli of intestine. Immediatelybelow the cell surface of the syncytiotrophoblast, which in vivo is in contact withmaternal blood, are a number of microtubules about 24 nm in diameter. Data on thedistribution of coated vesicles relative to the nearest cell surface are presented inTable 1 and in Figs. 3-5. These illustrate the fact that a very large proportion (atleast 89 %) of coated vesicles lie less than 540 nm from the cell surface. The distance540 nm was chosen because it is the longest axis across the largest coated vesicle yetmeasured (Ockleford, 1976). The fact that a higher proportion of vesicles are indirect contact with the cell surface than are present in the cytoplasm immediatelybelow the surface might suggest that the process of vesicle formation is slow comparedwith the rate of their subsequent movement away from the cell surface.
The image of the polygonally patterned structure on the surface of coated vesiclesobtained using tannic acid-fixed material was less informative than that obtained usingconventional fixation techniques. The electron-dense product surrounding the vesicleswas much thicker than the height of the ridges of the polygonally patterned lattice.
The 3 cell surface glycocalyx stains all reacted positively with the maternal surfaceof the syncytiotrophoblast (Figs. 1, 6, 7). However, the electron-dense regions wereconsiderably wider in the lumen of vesicles and in the caveolae of forming vesicles thanelsewhere on the surface when both ruthenium red (Fig. 6) and Alcian blue (Fig. 7)were used as stains. Within vesicles and caveolae this layer was up to about 40 nm inwidth and sometimes completely occluded the lumen of the vesicle. On other areasof the cell surface the layer of stain was usually less than 10 nm wide. Staining withcolloidal iron hydroxide at low pH was positive and remained so after neuraminidasedigestion, after esterification and after esterification followed by saponification.
The potassium pyroantimonate cation-precipitation technique produced a rather
Fig. 1. Transmission electron micrograph of a transverse section through a hollowfinger-shaped chorionic villus stained with Alcian blue. The outermost layer ofcytoplasm is the continuous syncytiotrophoblast (s). In places this contacts the base-ment membrane (bm). An incomplete layer is formed of pale-staining cytotrophoblastcells (c) which are frequently interposed between the basement membrane and thesyncytiotrophoblast. The lumen (/) of the villus is for the most part structureless butcontains occasional mesenchymal cells (m) and developing blood vessels (bv) x 2400.

298 C. D. Ockleford and A. Whyte
755*

Coated vesicles of human placenta 299
general distribution of an electron-dense precipitate in the syncytiotrophoblast(Fig. 8). Marked local increases in the concentration of precipitate, possibly indicatinglocally increased densities of cations at these sites in vivo, were observed in severalpositions. Heaviest precipitation occurred in the basement membrane. Granules inthe basement membrane (see Ockleford & Menon, 1977, fig. 2) were also covered byprecipitate. Irregularly shaped areas within syncytiotrophoblast nuclei were muchmore heavily covered with pyroantimonate than was the remainder of the nucleoplasm.Several very large cytoplasmic vacuoles were unlabelled. Precipitate was frequentlyobserved in coated vesicles where there was an apparent tendency for it to be localizedunderlying the luminal surface of the vesicle wall (Fig. 8).
The distribution and apparent number of coated vesicles were not substantiallyaltered by treatment of tissue-cultured trophoblast cells with the divalent cationchelator EDTA at a concentration of 0-02 %. However, the surface morphology of themajority of cells was altered. These became more convoluted in outline, forming amicrovillous pattern similar to that of the syncytiotrophoblast cell surface.
Isolated coated vesicles
Coated vesicles were isolated successfully as judged by electron microscopy(Fig. 9 A, B). The polygonal pattern visualized in these negatively stained preparationscorresponds to that predicted from a prior 3-dimensional reconstruction based onmicrographs of sectioned material (Ockleford, 1976). As has been pointed out fornegatively stained coated vesicles isolated from other sources (Crowther, Finch &Pearse, 1976), the vertices of the polygons (Fig. 9A, inset) are particularly prominent.Occasionally the walls of the polygons have a beaded appearance (Fig. 9 A, lowestvesicle in inset). The centre-to-centre spacing of the swollen regions is about 3-0 nm.
The overall size of these vesicles varies. The range from smallest to largest shownin Fig. 9 A is 70-120 nm across their longest axis. The shape of the vesicles is alsovariable (Fig. 9B). Both these features are described in detail elsewhere (Ockleford,Whyte & Bowyer, 1977). The membranous portion is apparently absent from manyof the polygonally patterned structures. It appears in a collapsed form within others.In Fig. 9 A the lattice nearest the centre of the field contains a large electron-lucentstructure which is probably the collapsed phospholipid bilayer.
Fig. 2. Electron micrograph of section through the microvillous surface of thesyncytiotrophoblast stained with lead citrate and uranyl acetate. These are similar toother microvilli (mv) in that they apparently contain microfilaments (mf) of the samediameter as that usually given for actin microfilaments (9 nm). The microvilli are lessuniform in size and shape and less regularly ordered than in other microvillous cellsurfaces. Microtubules {mi) about 24 nm in diameter are preserved using this methodof fixation (see Materials and methods of Ockleford & Menon, 1977). Coated vesicles(cv), caveolae (cav) and smooth-surfaced vesicles (v) are commonly found componentsin this region, x 38800.
Tab
le I
. N
umbe
rs a
nd p
erce
ntag
es o
f co
ated
ves
icle
s at
var
ious
dis
tanc
es fr
om c
ell
surf
ace
and
the
perc
enta
ge o
f th
e to
tal n
umbe
r of
ve
sicl
es n
eare
r to
the
cell
surf
ace
than
the
lar
gest
rec
orde
d di
amet
er o
f a
coat
ed v
esic
le
-
% o
f to
tal
no.
of
No
. of
coa
ted
vesi
cles
at
spec
ifie
d di
stan
ces
fro
m
% o
f co
ated
ves
icle
s at
spe
cifi
ed d
ista
nccs
fro
m c
ell
vesi
cles
nea
rer
to
cell
sur
face
. nm
su
rfac
e. n
m
cell
su
rfac
e th
an
A
c
, la
rges
t re
cord
ed
0-
60-
120-
240-
4
80
- 96
0-
1920
- 0-
60-
12
0-
240-
48
0-
960-
19
20-
diam
. of
a
Are
a in
vest
igat
cd
o 60
1
20
24
0 48
0 96
0 19
20
3840
o
60
12
0
240
480
960
1920
38
40
coat
ed v
esic
le*
Mat
ern
al s
urf
ace
of
8s
17
22
17
14
6
10
2
47.6
10.1
13
.1
10.1
8.
3 3 6
5'9
1'2
89
.2
syn
cyti
otr
op
ho
bla
st
Mat
ern
al s
urfa
ce o
f 98
20
25
15
8 2
- -
58.3
I 1.
9 14
.9
8.9
4.
8 1
'2
-
-
98.8
sy
ncy
tio
tro
ph
ob
last
, co
rrec
ted
dat
a
Cy
totr
op
ho
bla
st b
asal
28
17
6
8 9
9 4
5 32
.5
19.8
7.
0 9
'3
10.5
10
.5
4'6
5.
8 79
'0
lam
ina
surf
ace
Cy
totr
op
ho
bla
st s
urfa
ce
3 2
5 2
I - - -
23.1
15
.4
38.5
15
.4
7'7
-
-
-
-
adja
cent
syn
cyti
o-
tro
ph
ob
last
Dat
a re
late
to
10-
12-w
eek-
old
hu
man
pla
cent
a. T
he
term
'co
rrec
ted
dat
a' i
s ex
plai
ned
in t
he
Mat
eria
ls a
nd
met
ho
ds
sect
ion.
*
Th
e lo
nges
t ax
is o
f th
e la
rges
t co
ated
ves
icle
in
cel
ls o
f pl
acen
tal
orig
in s
o f
ar o
bser
ved
is 5
40 n
m (
Fig
. 5
; O
ckle
ford
, 19
76).
Wh
en t
he
nu
mb
er o
f ve
sicl
es c
lose
r to
th
e ce
ll s
urfa
ce t
han
540
nm
is
expr
esse
d as
a p
erce
ntag
e of
th
e to
tal
nu
mb
er o
f ve
sicl
es,
the
qual
itat
ive
impr
essi
on t
hat
th
eir
dis
trib
uti
on
is
rest
rict
ed t
o a
nar
row
ce
ll s
urfa
ce b
and
is
rein
forc
ed.
At
the
tim
e of
fix
atio
n a
maj
orit
y of
th
e co
ated
ves
icle
s la
y ex
trem
ely
clo
se t
o t
he
cell
sur
face
.
Coated vesicles of human placenta 301
75
>-oS 50
o
25
0 0-60 60-120 120-240.240-480 480-960 960- 1920-1920 3840
Distance from cell surface, nm
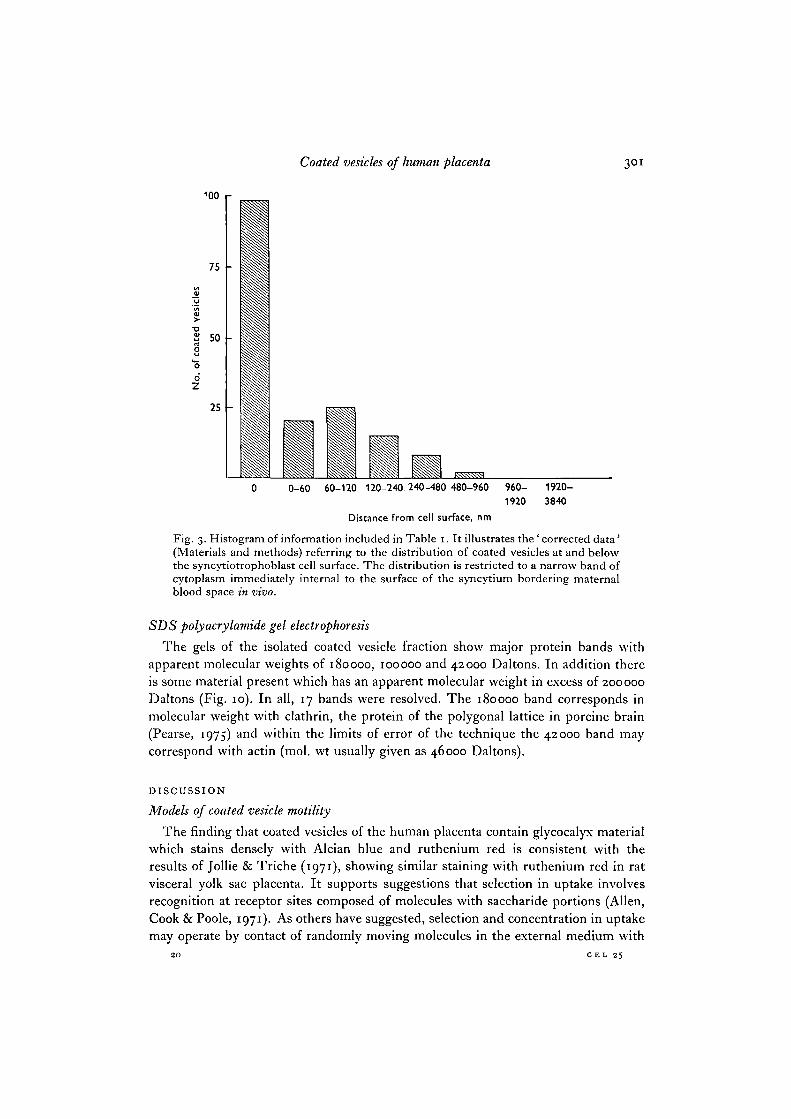
Fig. 3. Histogram of information included in Table 1. It illustrates the ' corrected data'(Materials and methods) referring to the distribution of coated vesicles at and belowthe syncytiotrophoblast cell surface. The distribution is restricted to a narrow band ofcytoplasm immediately internal to the surface of the syncytium bordering maternalblood space in vivo.
SDS polyaerylamide gel electrophoresis
The gels of the isolated coated vesicle fraction show major protein bands withapparent molecular weights of 180000, 100000 and 42000 Daltons. In addition thereis some material present which has an apparent molecular weight in excess of 200000Daltons (Fig. 10). In all, 17 bands were resolved. The 180000 band corresponds inmolecular weight with clathrin, the protein of the polygonal lattice in porcine brain(Pearse, 1975) and within the limits of error of the technique the 42000 band maycorrespond with actin (mol. wt usually given as 46000 Daltons).
DISCUSSION
Models of coated vesicle motility
The finding that coated vesicles of the human placenta contain glycocalyx materialwhich stains densely with Alcian blue and ruthenium red is consistent with theresults of Jollie & Triche (1971), showing similar staining with ruthenium red in ratvisceral yolk sac placenta. It supports suggestions that selection in uptake involvesrecognition at receptor sites composed of molecules with saccharide portions (Allen,Cook & Poole, 1971). As others have suggested, selection and concentration in uptakemay operate by contact of randomly moving molecules in the external medium with
20 CEL 25

302 C. D. Ockleford and A. Whyte
100
75
2 50
oZ
25
0 0-60 60-120 120-240 240-480 480-960 960- 1920-1920 3840
Distance from cell surface, nm
Fig. 4. Histogram of some of the data in Table 1. Showing the uncorrected distribu-tion of the same coated vesicles as in Fig. 3. It sets a firm upper limit to the distancethese coated vesicles may be from the syncytial cell surface.
receptor sites lying on the external surface of the membrane (Lloyd & Beck, 1974).It is easy to see how such a system may be compatible with Kanaseki & Kadota's (1969)model of vesicle motility. Their hypothesis depends upon the transformation ofparticular hexagons existing in an extended hexagonal lattice under the cytoplasmicsurface of the membrane into pentagons with the same length of side; an effect whichis presumed to generate curvature in the membrane, ultimately causing a vesicle topinch off. Selective uptake would simply require coextensiveness of the internalpolygonal lattice and the external glycomolecule receptor region.
Because the number of coated vesicles at the surface of cells does not diminish withtime, and because they are continually being internalized, it is necessary to assumethat they or their components are being recycled into the membrane or that themembrane is continually repopulated with newly synthesized vesicles or components.Initially when vesicle components enter cell surface membranes, and prior to thetransformation of the first hexagon to a pentagon according to Kanaseki & Kadota's(1969) model, there should be a flat hexagonal lattice underlying the membrane anda dense but flat thickened area of glycocalyx on the exterior surface. No observationsof flat or convex regions of membrane with thickened glycocalyces similar to thosefound in caveolae and fully formed vesicles have been made in this study. Unless theflattened stage is extremely short-lived, these data detract from confidence in the fixeddifferentiated area model. On the other hand, the observations are consistent with alater view (Ockleford, 1976) that suggests movement together in a manner analogous
Coated vesicles of human placenta 3°3
oZ
100
75
SO
25
0 0-60 60-120 120-240 240-480 480-960 960- 1920-1920 3840
Distance from cell surface, nm
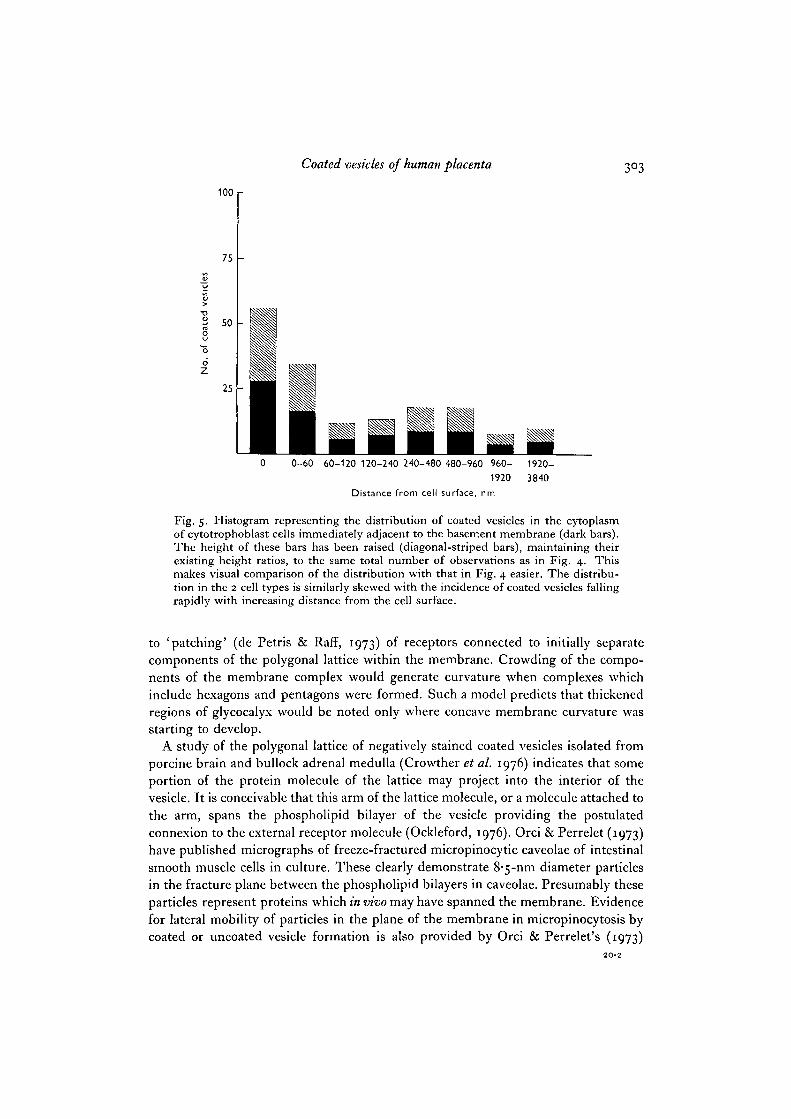
Fig. 5. Histogram representing the distribution of coated vesicles in the cytoplasmof cytotrophoblast cells immediately adjacent to the basement membrane (dark bars).The height of these bars has been raised (diagonal-striped bars), maintaining theirexisting height ratios, to the same total number of observations as in Fig. 4. Thismakes visual comparison of the distribution with that in Fig. 4 easier. The distribu-tion in the 2 cell types is similarly skewed with the incidence of coated vesicles fallingrapidly with increasing distance from the cell surface.
to 'patching' (de Petris & Raff, 1973) of receptors connected to initially separatecomponents of the polygonal lattice within the membrane. Crowding of the compo-nents of the membrane complex would generate curvature when complexes whichinclude hexagons and pentagons were formed. Such a model predicts that thickenedregions of glycocalyx would be noted only where concave membrane curvature wasstarting to develop.
A study of the polygonal lattice of negatively stained coated vesicles isolated fromporcine brain and bullock adrenal medulla (Crowther et al. 1976) indicates that someportion of the protein molecule of the lattice may project into the interior of thevesicle. It is conceivable that this arm of the lattice molecule, or a molecule attached tothe arm, spans the phospholipid bilayer of the vesicle providing the postulatedconnexion to the external receptor molecule (Ockleford, 1976). Orci & Perrelet (1973)have published micrographs of freeze-fractured micropinocytic caveolae of intestinalsmooth muscle cells in culture. These clearly demonstrate 8-5-nm diameter particlesin the fracture plane between the phospholipid bilayers in caveolae. Presumably theseparticles represent proteins which in vivo may have spanned the membrane. Evidencefor lateral mobility of particles in the plane of the membrane in micropinocytosis bycoated or uncoated vesicle formation is also provided by Orci & Perrelet's (1973)
3°4 C. D. Ockleford and A. Whyte
i • > v*V
' \ 6

Coated vesicles of human placenta 305
observation that particles similar in size to those close-packed in caveolae occur inthe surrounding membrane. In the syncytiotrophoblast the distribution of coatedvesicles at the cell surface appears to be linked with the distribution of microvilli. Thefunctions of the 2 organelles may therefore be interdependent. Usually the suggestedselective advantage of an irregular or microvillous border is that the increase insurface provides a larger area for exchange of materials with the extracellular environ-ment. If it is assumed that because of their common distribution the increased cellsurface area created by microvilli is important to the selective uptake function of coatedvesicles, then it seems probable that the microvillous surface provides an increased areaof receptor sites. Thus the observation that coated vesicles form only at and between thebases of microvilli is significant, because it suggests loaded receptors move downwardsto the bases of microvilli before they and their adherent extracellular material areenclosed in vesicles. This correlation may therefore be taken as evidence for thelateral mobility of receptors for selective uptake.
Because the net charge on the surface of most cells is negative, and because manyof the receptors are presumably chemically similar, it is reasonable to suppose thatthere may be a repulsion between predominantly negatively charged receptor mole-cules at close range. Multivalent molecules such as immunoglobulins may act asagents which bind receptors into the close-packed membrane complex and overcomethe hypothetical tendency for receptors to repel each other. Rohlich & Allison (personalcommunication) have demonstrated the presence of Fc receptors in the caveolae ofcoated vesicles, and this hypothesis is further supported by the observation of Contractor& Krakauer (1976) showing a stimulatory effect of serum in the uptake of protein bycultured human trophoblast cells. Presumably a divalent cation such as calciumcould also perform this 'bridging' role. The deposition of pyroantimonate in placentalcoated vesicles would seem to indicate the presence of a divalent cation. The observa-tion of a calcium-binding component in isolated brain coated vesicles may consequentlybe significant (Blitz & Fine, 1976). The fact that EDTA does not obviously alter theoccurrence or distribution of coated vesicles in cultured trophoblast cells does notrule out such a potential function.
Clathrin
The successful isolation of human placental coated vesicles by a similar method tothat which Pearse (1975, 1976) developed for porcine brain and bullock adrenalmedulla suggests that the organelles from the different sources share commonproperties. The ultrastructural similarity in the polygonal patterns on the cytoplasmic
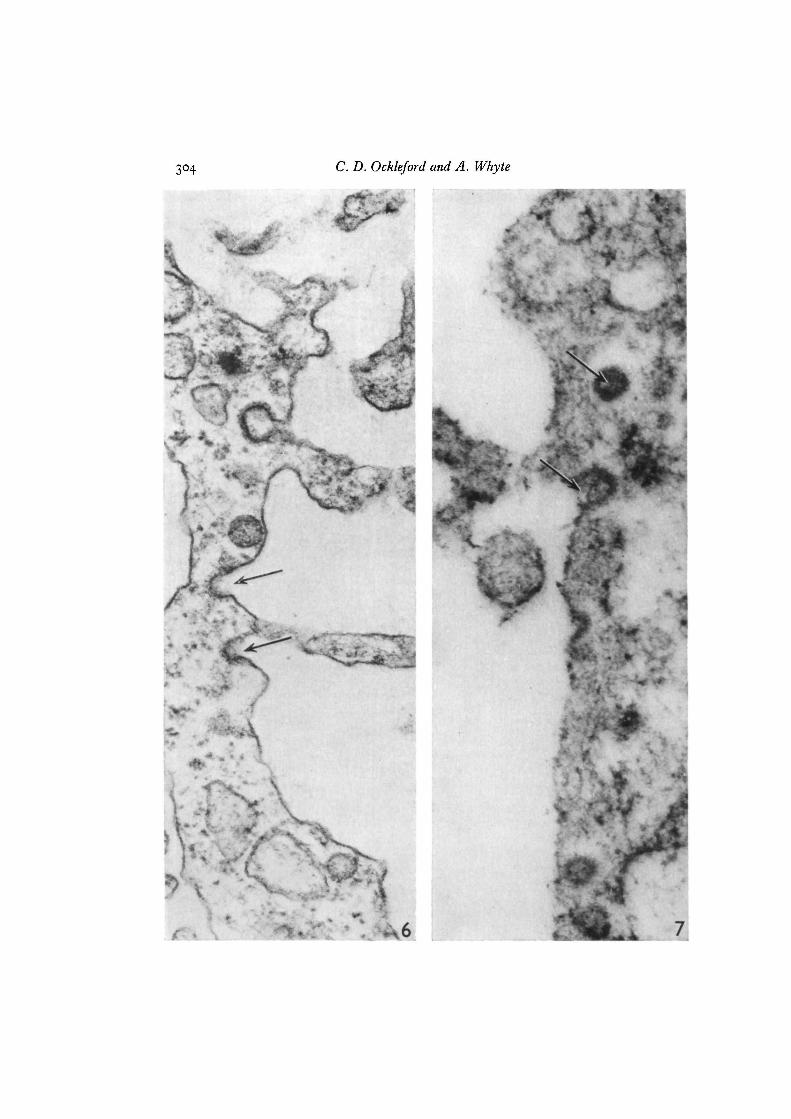
Fig. 6. This section through the cell surface of syncytiotrophoblast which in vivo is incontact with maternal blood, has been stained with ruthenium red. The lining of theluminal surfaces of caveolae (arrows) and coated vesicles exhibit increased electrondensity using this technique for the demonstration of glycocalyces. x IOIOOO.Fig. 7. Transmission electron micrograph of a section through the cell surface ofsyncytiotrophoblast stained with Alcian blue. The lining of the luminal surfaces ofcaveolae and coated vesicles (arrows) stain positively with this cationic dye. x 70000.

306 C. D. Ockleford and A. Whyte
/z
,'*

Coated vesicles of human placenta 307
surface of the vesicles suggests a similarity at the molecular level. The similarity of theSDS gel electrophoresis protein-banding patterns of the isolated placental coatedvesicle fractions to those of porcine brain points to the same conclusion. The proteinof greatest interest in this respect is the major protein band in the coated-vesiclefraction, which has an apparent molecular weight of 180000 Daltons. This is identicalwith the weight of the protein clathrin which Pearse (1975) has shown forms the poly-gonally patterned network on the cytoplasmic surface of porcine brain coated vesicles.
Coated vesicle function
Various, sometimes apparently contradictory, roles have been proposed as primaryfunctions for coated vesicles. However, it is possible to discern similarities betweenprocesses previously classified as separate when more attention was given to theproduct of coated-vesicle formation than to its mechanism, (a) Coated vesicles areknown to take up materials by micropinocytosis. There is evidence for this functionfrom both comparative anatomy and more directly from tracer studies which indicatethe uptake of proteins (Rodewald, 1973; King & Enders, 1971; Roth & Porter, 1964;Bowers, 1964; Lagunoff & Curran, 1972). (b) They may also remove membrane fromthe cell surface, thus facilitating membrane turnover in secretory cells. Because thesurface of these cells does not grow larger as a result of incorporating exocytic vesiclemembranes during secretion, the cell must possess a method for internalization ofmembrane at the same approximate rate as incorporation. This may (Henser & Reese,1973; Fried & Blaustein, 1976) or may not (Nordmann & Morris, 1976; Pearse, 1976)be mainly the result of coated-vesicle action. The main difference between membranerecycling and selective uptake may simply be the absence of functional receptors,(c) Although it was not the case for syncytiotrophoblast, coated vesicles have beendescribed forming in association with Golgi membrane. It has therefore been suggestedthat some coated vesicles are specialized exocytic vesicles, which discharge materialsprocessed by the Golgi to the exterior (Friend & Farquhar, 1967; Jamieson & Palade,1971). That coated vesicles may function in both endocytosis and exocytosis is ap-parently paradoxical. One function of the Golgi is to package and condense cellularproducts. Crowding of clathrin molecules attached through the Golgi membrane tomolecules which may be secreted would concentrate them in much the same wayas the process of concentration in selective uptake by micropinocytosis has been
Fig. 8. The most electron-dense portions of the image in this transverse sectionthrough a term placental chorionic villus represent precipitated cation pyroantimonates.Ultrastructure is generally poorly preserved using this technique, but mitochondria{mi) and coated vesicles {cv) are recognizable. The basement membrane {bin), which isoften multilaminous later in pregnancy, is in 2 parts in this section. Both layers are sitesof rather heavier precipitation than in other areas. It is important to observe thatthe precipitate is absent from large parts of electron-lucent vacuoles (fa). The smallquantities of precipitate over coated vesicles may not therefore simply be the result ofcoincidences in distribution between one group of randomly distributed small particlesoccurring with a high frequency superimposing on a lesser number of larger structureswhich are also randomly distributed, x 125000.

cv
•7
C. D. Ockleford and A. Whyte
m
i m
9A
b
4

Coated vesicles of human placenta 309
suggested to occur (Ockleford, 1976). The functions in both processes to which theclathrin lattice is related may be concentration of selected molecules and invaginationof a membrane, (d) It has been suggested that coated vesicles carry their clathrinlattices in several processes purely as a 'fender', which prevents vesicular contact andfusion with lysosomal membranes (Wild, 1976). In this hypothetical manner thecontents of the vesicle are protected from degradation by lysosomal enzymes. Thereare apparently no cases recorded in the literature of vesicles still bearing clathrin coatsfusing with lysosomes.
Loss of coats
Kanaseki & Kadota (1969) have provided evidence for the loss of clathrin latticesby vesicles. If clathrin loss occurred very shortly after vesicle formation in thesyncytiotrophoblast then an obvious explanation for the apparently restricted distri-bution of coated vesicles in this tissue follows. If coats are not lost then some otherexplanation of the restricted distribution of coated vesicles at the syncytiotrophoblastcell surface must be sought.
Intracellular motility of coated vesicles
The protein of isolated coated vesicles appearing as a band on SDS polyacrylamidegels with an apparent molecular weight of 42000 may be actin. If this is the case, it ispossible that actin is a contaminant of the preparation, reflecting the close cellularassociation of coated vesicles and microfilaments. Equally, actin may be attached tocoated vesicles in a more specific and meaningful way, as it apparently is to chromaffingranules (Burridge & Phillips, 1975), where it is a proposed link in the chain oftransport processes causing intracellular movement of the granules.
The physical association of coated vesicles with actin-like microfilaments andmicrotubules may not be fortuitous. Both these structures have been considerednecessary for proper motile function. Either or both may be necessary componentsfor the transport in the syncytiotrophoblast of coated vesicles, vesicles which have losttheir clathrin coats, or of empty coats.
The nature of the remaining Coomassie-staining bands on the polyacrylamide gelsmust remain a matter for speculation, though it is perhaps not without significance thata major component of apparent molecular weight 100000 Daltons has been found by
Fig. 9 A. Electron micrograph of coated vesicles (cv) isolated from human placenta andnegatively stained using uranyl acetate. It illustrates the polygonally patterned net-work of the protein clathrin on the surface of the vesicles. There are some smooth-surfaced membrane structures (s) in the samples. Some of the coated vesicles (arrow)contain an electron-lucent portion thought to represent the membrane of the vesicleretracted into the central region. On occasion (arrowhead in inset) the walls of polygonsappear to exhibit beaded substructure. The longest axis across images of vesicles in thisfigure varies between 70 and 120 nm. x 55000; inset x 140000.Fig. OB. A similar sample to that shown in Fig. 9 A which illustrates variability invesicle shape over and above the variability in size already indicated. One vesicle(arrow) is definitely non-spherical, x 55000.

3io C. D. Ockleford and A. Whyte
230 180 145 100
Daltons x10~3
42
\
Fig. io. The bands of the isolated coated-vesicle fraction on this 75 % SDS poly-acrylamide gel indicate the major proteins present. The molecular weights of some ofthese proteins determined from their mobilities are also shown. The arrow indicates anartifactual band situated just behind the dye front. The band with an apparent molecularweight of 180000 Daltons, the same molecular weight as that of clathrin from pig brain(Pearse, 1975), is one of the most prominent.
Blitz & Fine (1976) in a coated-vesicle preparation isolated from mammalian brainhaving high Ca2+-activated ATPase activity. In sarcoplasmic reticulum Scales & Inesi(1976) have described a Ca2+-dependent ATPase with a molecular weight of 106000Daltons. In contrast, however, Blitz & Fine (1976) demonstrated in their preparationa component with a molecular weight of 60000 Daltons which was not apparent on gelsof placental coated-vesicle proteins.

Coated vesicles of human placenta 311
Kate Barber's technical assistance in the course of this work is highly appreciated. We aregrateful to Dr Brooks of the Maternity Hospital, Cambridge and the Gynaecologists ofAddenbrooke's Hospital for their clinical coordination and to Drs E. A. Munn and Y. W. Lokefor encouragement. At the time of this work C.D.O. was funded by a Medical ResearchCouncil Project grant (G973/112/B to Dr Y. W. Loke), and A. W. was holder of a MedicalResearch Council Research Studentship.
REFERENCES
ALLEN, J. M., COOK, G. M. W. & POOLE, A. R. (1971). Action of concanavalin A on theattachment stage of phagocytosis by macrophages. Expl Cell Res. 68, 466-471.
ALLISON, A. C. & DAVIS, P. (1974). Mechanisms of endocytosis and exocytosis. In Transport atthe Cellular Level, Symp. Soc. exp. Biol. (ed. M. A. Sleigh & D. H. Jennings), pp. 521-546.Cambridge University Press.
BEHNKE, O. & ZELANDER, T. (1970). Preservation of intercellular substances by the cationic dyealcian blue in preparative procedures for electron microscopy. J. Ultrastruct.Res. 31,424-438.
BLITZ, A. L. & FINE, R. E. (1976). Functional and compositional similarities between coatedvesicles, synaptic vesicles and sarcoplasmic reticulum fragments. J. Cell Biol. 70 (2) 204A.
BOWERS, B. (1964). Coated vesicles in the pericardial cells of the aphid (Myzus persicae Sulz).Protoplasma 59, 351-367.
BRAMBELL, F. W. (1970). The transmission of passive immunity from mother to young.In Frontiers of Biology, 18, pp. 242-245. Amsterdam: North-Holland Publishing. New York:American Elsevier.
BURRIDGE, K. & PHILLIPS, J. H. (1975). Association of actin and myosin with secretory granulemembranes. Nature, Lond. 254, 526-529.
CONTRACTOR, S. F. & KRAKAUER, K. (1976). Pinocytosis and intracellular digestion of 125I-labelled haemoglobin by trophoblastic cells in tissue culture in the presence and absence ofserum. J. Cell Set. z i , 595-607.
CROWTHER, R. A., FINCH, J. T. & PEARSE, B. M. F. (1976). On the structure of coated vesicles.J. molec. Biol. 103, 785-798.
DE PETRIS, S. & RAFF, M. C. (1973). Fluidity of the plasma membrane and its implications forcell movement. In Locomotion of Tissue Cells, Ciba Fdn Symp. 14 (new series), pp. 27-41.Cambridge University Press.
FRIED, R. C. & BLAUSTEIN, M. P. (1976). Synaptic vesicle recycling in synaptosomes in vitro.Nature, Lond. 261, 255-256.
FRIEND, D. S. & FARQUHAR, M. G. (1969). Functions of coated vesicles during proteinabsorption in the rat vas deferens. J. Cell Biol. 35, 357-376.
HENSER, J. E. & REESE, T. S. (1973). Evidence for recycling of synaptic vesicle membraneduring transmitter release at the frog neuromuscular junction. J. Cell Biol. 57, 315-344.
HUXLEY, H. E. (1963). Electron microscope studies on the structure of natural and syntheticprotein filaments from striated muscle. ,7. molec. Biol. 7, 281-308.
JAMIESON, J. D. & PALADE, G. E. (1971). Synthesis, intracellular transport, and discharge ofsecretory proteins in stimulated pancreatic exocrine cells. J. Cell Biol. 35, 357-376.
JOLLIE, W. P. & TRICHE, T. J. (1971). Ruthenium labelling of micropinocytotic activity in therat visceral yolk sac placenta. J. Ultrastruct. Res. 35, 541-553.
KANASEKI, T. & KADOTA, K. (1969). The 'vesicle in a basket'. A morphological study of thecoated vesicle isolated from the nerve endings of the guinea pig brain with special referenceto the mechanism of membrane movement. J. Cell Biol. 42, 202-220.
KING, B. F. & ENDERS, A. C. (1971). Protein absorption by the guinea pig chorioallantoicplacenta. Am. J. Anat. 130, 409-430.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the assembly of the head of thebacteriophage T4. Nature, Lond. 227, 680-685.
LAGUNOFF, D. & CURRAN, D. E. (1972). Role of bristle coated membrane in the uptake offcrritin by rat macrophages. Expl Cell Res. 75, 337-346.
LLEWELLYN-JONES, D. (1969). Infectious diseases in pregnancy. In Fundamentals of Obstetricsand Gynaecology, pp. 216-217. London: Faber & Faber.

312 CD. Ockleford and A. Whyte
LLOYD, J. B. & BECK, F. (1974). Lysosomes. In The Cell in Medical Science, vol. 1, The Cell andIts Organelles (ed. F. Beck & J. B. Lloyd), pp. 272-313. London and New York: AcademicPress.
LOKE, Y. W. & BORLAND, R. (1970). Immunofluorescent localisation of chorionic gonado-trophin in monolayer cultures of human trophoblast cells. Nature, Lond. 228, 561-562.
LUFT, J. H. (1965). Ruthenium red and violet 1. Chemistry, purification, methods of use forelectron microscopy and mechanism of action. Anat. Rec. 171, 347-368.
MARX, R., GRAF, E. & WESEMANN, W. (1973). Histochemical and biochemical demonstration ofsialic acid and sulphate in vesicles and membranes isolated from nerve endings of rat brain.J. Cell Sci. 13, 237-255.
MILLER, R. K. & BERNDT, W. O. (1975). Mechanisms of transport across the placenta: anin vitro approach. Life Sci. 16, 7-30.
NORDMANN, J. J. & MORRIS, J. F. (1976). Membrane retrieval at neurosecretory axon endings.Nature, Lond. 261, 723-724.
OCKLEFORD, C. D. (1975). Redundancy of washing in the preparation of biological specimensfor transmission electron microscopy. J. Microscopy 105, part 2, 193-203.
OCKLEFORD, C. D. (1976). A three-dimensional reconstruction of the polygonal pattern onplacental coated-vesicle membranes. J. Cell Sci. 21, 83-91.
OCKLEFORD, C. D. & MENON, G. (1977). Differentiated regions of human placental cell surfaceassociated with exchange of materials between maternal and foetal blood. J. Cell Sci. 25,279-291.
OCKLEFORD, C. D., WHYTE, A. & BOWYER, D. E. (1977). Variation in the volume of coatedvesicles isolated from human placenta. Cell Biology int. Rep. (in Press).
ORCI, L. & PERRELET, A. (1973). Membrane-associated particles: Increase at sites of pino-cytosis demonstrated by freeze-etching. Science, N.Y. 181, 868-869.
PEARSE, B. M. F. (1975). Coated vesicles from pig brain. Purification and biochemicalcharacterization. J. violec. Biol. 97, 93-98.
PEARSE, B. M. F. (1976). Clathrin: a unique protein associated with intracellular transfer ofmembrane by coated vesicles. Proc. natn. Acad. Sci. U.S.A. 73, 1255-1259.
RODEWALD, R. B. (1973). Intestinal transport of antibodies in the new-born rat. J. Cell Biol.58, 189-211.
ROTH, T. F. & PORTER, K. R. (1964). Yolk protein uptake in the oocyte of the mosquitoAedes aegypti L. J. Cell Biol. 20, 313-331.
ROTHMAN, A. H. (1970). Alcian blue as an electron stain. Expl Cell Res. 58, 177-179.SCALES, D. & INESI, G. (1976). Assembly of ATPase protein in sarcoplasmic reticulum mem-
branes. Biophys. J. 16, 735-751.SIMPSON, J. V. & SPICER, S. S. (1973). Activities of specific cell constituents in phagocytosis
(endocytosis). Int. Rev. exp. Path. 12, 79-118.SPURR, A. R. (1969). A low-viscosity epoxy resin embedding medium for electron microscopy.
J. Ultrastruct. Res. 26, 31-43.TANDLER, C. J., LIBANATI, C. M. & SANCHIS, C. A. (1970). The intracellular localisation of
inorganic cations with potassium pyroantimonate: electron probe microanalysis. J. Cell Biol.45, 3SS-366.
TIGHE, J. R., GARROD, P. R. & CURRAN, R. C. (1967). The trophoblast of the human chorionicvillus. J. Path. Bact. 93, 559-567.
TILNEY, L. G., BRYAN, J., BUSH, D. J., FUJIWARA, K., MOOSEKER, M. S., MURPHY, D. B. &SNYDER, D. H. (1973). Microtubules: evidence for 13 protofilaments. J. Cell Biol. 59267-275-
WEBER, K. & OSBORN, M. (1969). The reliability of molecular weight determinations bydodecyl sulfate-polyacrylamide gel electrophoresis. J. biol. Chem. 244 (16), 4406-4412.
WILD, A. E. (1976). Mechanism of protein transport across the rabbit yolk sac endoderm. InMaternofoetal Transmission of Immunoglobulins (ed. W. A. Hemmings), pp. 155-165. London,New York and Melbourne: Cambridge University Press.
WILLS, E. J., DAVIES, P., ALLISON, A. C. & HASWELL, A. D. (1972). The failure of cytochalasinB to inhibit pinocytosis by macrophages. Nature, Netv Biol. 240, 58-60.
(Received 6 September 1976)