dilkes, 2001
DESCRIPTION
D.W. Dilkes. Biology Department, Erindale College, The University of Toronto at Mississauga, 3359 Mississauga Road, Mississauga, ON L5L 1C6, Canada. (e-mail: [email protected]). 1205 [Traduit par la Rédaction] 1227 Paper handled by Associate Editor B. Chatterton. Dilkes DOI: 10.1139/cjes-38-8-1205 Introduction © 2001 NRC CanadaTRANSCRIPT

An ontogenetic perspective on locomotion in theLate Cretaceous dinosaur Maiasaura peeblesorum(Ornithischia: Hadrosauridae)
David W. Dilkes
Abstract: Ontogenetic growth in the forelimb and hindlimb of the dinosaurMaiasaura peeblesorum(Ornithischia:Hadrosauridae) from the Upper Cretaceous (Campanian–Maastrichtian) Two Medicine Formation of Montana is investi-gated by multivariate and bivariate morphometrics and the biomechanics of beam theory. Results support a hypothesisof an age-dependent selection of stance. Juveniles walked primarily as bipeds. As an individual matured, its predomi-nant stance shifted to quadrupedality. Within the forelimb, morphometric results show a probable allometric enlarge-ment of postural muscles, an allometric increase in the lever arms of protractor muscles, and an increased robustness ofthe humerus to enhance its resistance to bending stresses. In contrast, the hindlimb is characterized by a relative de-crease in the resistance of the femur and tibia to bending stresses. In addition, there is an allometric enlargement of thefemoral fourth trochanter and positive allometry of the lengths of metatarsals III and IV. The most likely explanationfor the different growth patterns is that the hindlimb was sufficiently robust at a young age to accommodate increasedpostural and locomotory stresses through largely isometric growth, whereas a behavioral shift to quadrupedality in olderindividuals necessitated an allometric response in the forelimb. Osteological adaptations for weight-bearing in themanus include metacarpals that are united firmly with a resultant cross-sectional shape that is resistant to bending andhyperextendable joints between the metacarpals and first phalanges. It is probable that flexor muscles that attached tothe caudal surface of the metacarpals reinforced the reduced carpus and lessened the likelihood of collapse duringquadrupedality.
Résumé: La croissance ontogénétique des membres antérieurs et des membres postérieurs du dinosaureMaiasaurapeeblesorum(Ornithischia: Hadrosauridae) de la Formation de Two Medicine du Montana, du Crétacé supérieur (Cam-panienBMaastrichtien), est examinée en utilisant la morphométrie multivariée et à deux variables ainsi que la bioméca-nique de la théorie de la poutre. Les résultats appuient l’hypothèse d’un choix de posture reliée à l’âge. Les jeunesimmatures avaient surtout avec une démarche bipède. À mesure que l’individu vieillissait, sa posture prédominante al-lait vers une démarche à quatre pattes. Pour le membre antérieur, les résultats morphométriques montrent un agrandis-sement allométrique probable des muscles de posture, une augmentation allométrique dans les bras de levier ou dansles muscles d’élongation et une augmentation de la robustesse de l’humérus pour accroître sa résistance aux contraintesde flexion. Par contre, le membre postérieur est caractérisé par une réduction de la résistance du fémur et du tibia auxcontraintes de flexion. De plus, on voit un agrandissement allométrique du quatrième trochanter fémoral et une allo-métrie positive de la longueur des 3e et 4e métatarses. L’explication la plus plausible pour les patrons de croissance di-vergents est que le membre postérieur était assez robuste à un jeune âge pour permettre une augmentation descontraintes de posture et de locomotion par une croissance surtout isométrique, alors qu’un changement de comporte-ment vers une démarche à quatre pattes chez les individus plus âgés exigeait une réponse allométrique du membre an-térieur. Des adaptations ostéologiques pour la portance comprennent, dans la paume, des métacarpes qui sontfermement reliés pour donner une forme transversale résistant à la flexion et des articulations capablesd’hyperextension entre les métacarpes et les premières phalanges. Il est probable que les muscles fléchisseurs qui serattachaient à la surface caudale renforçaient le carpus réduit et amoindrissaient le risque d’affaissement lors de la dé-marche à quatre pattes.
[Traduit par la Rédaction] 1227
Dilkes
Can. J. Earth Sci.38: 1205–1227 (2001) © 2001 NRC Canada
1205
DOI: 10.1139/cjes-38-8-1205
Received July 7, 2000. Accepted January 22, 2001. Published on the NRC Research Press Web site at http://cjes.nrc.ca onAugust 20, 2001.
Paper handled by Associate Editor B. Chatterton.
D.W. Dilkes. Biology Department, Erindale College, The University of Toronto at Mississauga, 3359 Mississauga Road,Mississauga, ON L5L 1C6, Canada. (e-mail: [email protected]).
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:47:51 AM
Color profile: DisabledComposite Default screen

Introduction
Few published studies of the relative bipedality orquadrupedality of non-avian dinosaurs have incorporatedjuveniles. Although numerous fossils of juvenile dinosaursare known (Carpenter and Alf 1994) and new discoveries arebecoming increasingly frequent (e.g., Dal Sasso and Signore1998), many of these juveniles are incomplete and make uponly a very small portion of a growth sequence. As a conse-quence, many studies of postcranial growth in non-aviandinosaurs are restricted to simple comparisons of the propor-tions of limbs in the juveniles and adults (Russell 1970;Dodson 1980; Norman 1980) or, if statistical, then deal withobligatory quadrupedal (Lehman 1990) and bipedal (Smith1998) dinosaurs. Only the study onIguanodonbernissartensis(Norman 1980) presented evidence of anontogenetic shift from bipedality to quadrupedality. However,the sample size in this study consisted of only four individuals,and the conclusions were based solely upon limb ratios. Amore recent attempt to examine ontogeny and locomotion inthe iguanodontian Dryosaurus lettowvorbecki from abiomechanic approach (Heinrich et al. 1993) purportedlyrevealed an ontogenetic shift from a quadrupedal to a bipedstance. The methodology in this paper was a significantimprovement upon the comparison of limb ratios. Thesestudies demonstrate clearly that ontogeny provides a newand highly informative view on locomotion in the adult.
The inclusion of juveniles in an ontogenetic study of loco-motion is particularly relevant for those groups of dinosaurswhose stance is debated currently. Consideration of anontogenetic dimension may reveal patterns that can explainthe morphology of the adult better than comparisons of simpleratios between adults of different species. The relativebipedality or quadrupedality is uncertain for manyprosauropods, the hadrosaurids, and basal iguanodontianssensu Weishampel and Heinrich (1992), such asIguanodonand Dryosaurus. Many prosauropods have been interpretedas facultatively bipedal, defined as quadrupedal stance atslow speeds with a shift to a bipedal stance occurring onlywhen moving quickly. Smaller prosauropods such asThecodontosaurusmay have been bipedal (Galton 1990;Benton et al. 2000), and larger prosauropods such asRiojasauruswere perhaps obligatory quadrupeds (Heerdenand Galton 1997). Comparisons of limb ratios and proportionsof the trunk and tail between hadrosaurs and quadrupedaldinosaurs and the slenderness of the metacarpals, the reduc-tion of the carpals to only two small bones, and the mitten-like manus of a “mummified” hadrosaur (Osborn 1912)appeared to demonstrate that hadrosaurs were strict bipeds(Galton 1970). Acceptance of hadrosaurs as strict bipedswas crucial for an interpretation of the stance of theiguanodontianIguanodon. Comparisons of limb proportionsand hindlimb to forelimb ratios of hadrosaurs and theornithopodTenontosaurus(a probable quadruped) with thoseof Iguanodon mantelliand the largerI. bernissartensissug-gested thatI. mantelli was predominantly a biped, whereasI. bernissartensiswas probably a quadruped (Norman 1980).Quadrupedality among iguanodontians is supported stronglyby the presence of numerous adaptations for weight-bearingin the carpus and manus (Norman 1980) and trackways(Norman 1980; Moratalla et al. 1992, 1994; Pérez-Lorente et
al. 1997). However, this interpretation is challenged bybiomechanical arguments that suggest a virtually habitualbipedal stance forI. bernissartensis(Alexander 1985;Christian and Preuschoft 1996). Trackways of large,quadrupedally walking hadrosaurs (Currie et al. 1991) haverefuted the conclusion that these dinosaurs were strictlybipedal. Hadrosaurs are interpreted currently as facultativelybipedal.
Individuals of different ontogenetic ages are required totest a hypothesis of an age-dependent stance. Furthermore,there should be a combination of biomechanic andmorphometric analyses that extend beyond simple ratios oflengths of the forelimb and hindlimb or selected bones of thelimbs. Limb ratios are an unsatisfactory method to examinepostcranial growth because they reveal little biomechanicalinformation and fail to capture the majority of shape infor-mation that may reveal allometric patterns. Mass accumula-tions with individuals at different growth stages are knownfor Iguanodonfrom the locality at Bernissart, Belgium (Nor-man 1986), hadrosaurids from Montana (Varricchio and Hor-ner 1993), Alberta (Dodson 1971), and Alaska (Nelms1989), and neoceratopsians such asChasmosaurusmariscalensisfrom Texas (Lehman 1990) and several generafrom Alberta and Montana (Sampson et al. 1997). The largesample sizes obtained from these bone beds provide data onintraspecific variation needed for an assessment ofontogenetic changes. A particularly impressive growth se-quence is known for the hadrosaurMaiasaura peeblesorumfrom the Upper Cretaceous Two Medicine Formation ofMontana (Fig. 1). The age of this formation has been datedrecently to between 82.6 and 74.0 million years (Rogers etal. 1993), indicating that it straddles the Campanian–Maastrichtian boundary.Maiasaura first gained scientificnotice in 1978 when a nest with egg shell and thedisarticulated remains of more than a dozen nestlings werediscovered in the Willow Creek Anticline near Choteau,Montana (Horner and Makela 1979). Soon afterwards, sevenother nests were discovered on the same horizon as the nestof 1978, one of which contained the remains of additional,younger juveniles approximately 1 m in body length (Horner1982). Three nesting horizons are currently known forMaiasaura in the Willow Creek Anticline (Horner 1994).Additional discoveries ofMaiasaura in the Willow CreekAnticline include a virtually monospecific bone bed knownas Camposaur on the eastern flank of the anticline (Fig. 1C).The Camposaur bone bed is one portion of a significantlylarger deposit that is estimated at 3 km from east to west and0.5 km from north to south and is one of the largestmonospecific bone beds of dinosaurs in the world. This bonebed is an apparently catastrophic thanatocoenosis of thou-sands of individuals that span a three-fold size range from3 m to over 7 m (Varricchio and Horner 1993; Schmitt et al.1998). Other localities such as the Brandvold Site on thewestern flank of the Willow Creek Anticline and theChildren’s Dig on the eastern flank are part of this larger de-posit. Density of bones in the Camposaur bone bed is excep-tionally high, yet all are disarticulated and unassociated andthe majority are fragmented. When the Camposaur speci-mens are combined with the nestlings, a growth sequenceexists for Maiasaura that occupies conservatively an orderof magnitude in size.
© 2001 NRC Canada
1206 Can. J. Earth Sci. Vol. 38, 2001
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:47:51 AM
Color profile: DisabledComposite Default screen

This paper tests the hypothesis that there is an ontogeneticshift in stance for the hadrosaurM. peeblesorumby investi-gating the morphometric and biomechanic aspects of growthof the forelimb and hindlimb from nestling to adult.Morphometrics of external dimensions and cortical bonethickness of limb bones are based upon the bivariate model
of simple allometry (Huxley 1932; Shea 1985) and principalcomponent analysis (PCA; Jolicoeur 1963). The forelimband hindlimb are modelled as beams to identify anybiomechanical changes during growth. Biomechanics of theadult manus are also studied to test the hypothesis that itwould not have been capable of supporting even a small
© 2001 NRC Canada
Dilkes 1207
Fig. 1. Geographic location of the Camposaur site. (A) Orthographic map showing state of Montana (hatched). (B) Location of theWillow Creek Anticline (hatched rectangle) in the Two Medicine Formation near the town of Choteau. (C) Topographic map of theWillow Creek Anticline corresponding to the rectangle in (B). The topographic map is adapted from the U.S. Geological Survey7.5 min topographic map Watson Flats.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:47:56 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
1208 Can. J. Earth Sci. Vol. 38, 2001
fraction of the body weight during locomotion, constitutingthe strongest morphologic evidence against quadrupedalityin hadrosaurs.
Materials and methods
MaterialsSpecimens used in this study consist of the nestlings from
the 1978 nest (Peabody Museum of Natural History at YaleUniversity, housed formerly at Princeton University, desig-nated by YPM-PU 22400), and excavations from Camposaur(Museum of the Rockies 005, designated by MOR-005). The1978 nest and Camposaur are situated in Teton County,Montana, in section 30, T24N, R6W. Specimens from theBrandvold Site were too fragmentary to be included in thestudy. All specimens are housed in the Museum of the Rock-ies in Bozeman, Montana.
Ideally, a thorough study of appendicular ontogeny shouldinclude the girdles as well as the limbs. Unfortunately, withfew exceptions, available specimens of the girdles ofMaiasaura are incomplete, usually distorted, and few innumber. Accordingly, statistical analysis of postcranialontogeny inMaiasaura is restricted to the humerus, ulna,femur, tibia, and metatarsals. Nestlings are combined withmaterial from the bone bed for the humerus, all three meta-tarsals, femur, and tibia, but excluded for the ulna becauseavailable specimens are incomplete. The majority ofpostcranial elements from the Camposaur locality sufferfrom varying degrees of taphonomic distortion, often in dif-ferent planes. External and internal features were examinedfor any gross distortions caused by permineralization of the
bone or compaction of the sediments. Any specimensdisplaying deformations that would clearly compromisemeasurements were eliminated.
Methods
Selection of measurementsMeasurements used in the morphometric analyses are of
two types: (i) distances between homologous, three-dimensionally defined landmarks; and (ii ) distances betweenextremal points. Extremal points are the traditional sort usedin morphometrics and include such variables as diameters ofshafts or openings (e.g., orbits), minimum circumference,maximum width of condyles, and points farthest from otherpoints. Extremal points are kept to a minimum becauseinformation on their displacement is limited to the singledimension of the line that connects them, whereas displace-ments of homologous landmarks can be detected in two orthree dimensions (Bookstein 1990). The variables used inthe morphometric analyses excluding cortical bone thicknessare shown in Fig. 2. Variables 11 for the humerus, 4 for thefemur, 5 for the tibia, and 12 for metatarsal III are minimumcircumferences of the shafts. For the ulna, variable 1 is theminimum diameter of the shaft, and the outer point on theolecranon for variable 4 is an extremal point. Additionalextremal points are variables 2 and 3 for the femur and thepoint separating variables 2 and 3 for the tibia. All otherpoints are landmarks of category 2 (sharp corners and maximaof curvatures) as defined by Bookstein (1990). As the proxi-mal and distal ends of humeri, femora, tibia, and metatarsalsare curved and were subject to biomechanical forces, great-
Fig. 2. Variables selected from forelimb and hindlimb ofMaiasaura peeblesorumfor morphometric analysis. (A) Left humerus in caudaland proximal (above) views. (B) Left ulna. (C) Left femur in medial view. (D) Left tibia in caudal view. (E) Left metatarsal II in medialand proximal (above) views. (F) Left metatarsal III in (from left to right and top to bottom) cranial, caudal, proximal, and distal views.(G) Left metatarsal IV in (from left to right and top to bottom) cranial, caudal, proximal, and distal views. Not drawn to scale.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:47:58 AM
Color profile: DisabledComposite Default screen
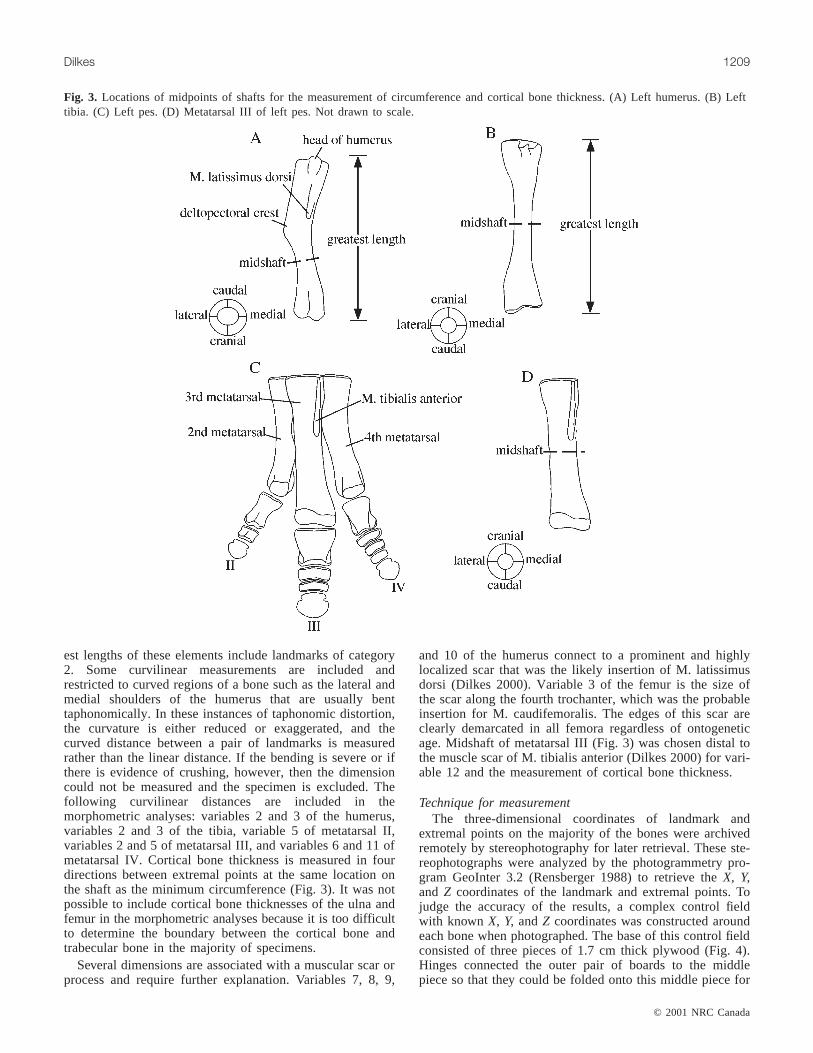
est lengths of these elements include landmarks of category2. Some curvilinear measurements are included andrestricted to curved regions of a bone such as the lateral andmedial shoulders of the humerus that are usually benttaphonomically. In these instances of taphonomic distortion,the curvature is either reduced or exaggerated, and thecurved distance between a pair of landmarks is measuredrather than the linear distance. If the bending is severe or ifthere is evidence of crushing, however, then the dimensioncould not be measured and the specimen is excluded. Thefollowing curvilinear distances are included in themorphometric analyses: variables 2 and 3 of the humerus,variables 2 and 3 of the tibia, variable 5 of metatarsal II,variables 2 and 5 of metatarsal III, and variables 6 and 11 ofmetatarsal IV. Cortical bone thickness is measured in fourdirections between extremal points at the same location onthe shaft as the minimum circumference (Fig. 3). It was notpossible to include cortical bone thicknesses of the ulna andfemur in the morphometric analyses because it is too difficultto determine the boundary between the cortical bone andtrabecular bone in the majority of specimens.
Several dimensions are associated with a muscular scar orprocess and require further explanation. Variables 7, 8, 9,
and 10 of the humerus connect to a prominent and highlylocalized scar that was the likely insertion of M. latissimusdorsi (Dilkes 2000). Variable 3 of the femur is the size ofthe scar along the fourth trochanter, which was the probableinsertion for M. caudifemoralis. The edges of this scar areclearly demarcated in all femora regardless of ontogeneticage. Midshaft of metatarsal III (Fig. 3) was chosen distal tothe muscle scar of M. tibialis anterior (Dilkes 2000) for vari-able 12 and the measurement of cortical bone thickness.
Technique for measurementThe three-dimensional coordinates of landmark and
extremal points on the majority of the bones were archivedremotely by stereophotography for later retrieval. These ste-reophotographs were analyzed by the photogrammetry pro-gram GeoInter 3.2 (Rensberger 1988) to retrieve theX, Y,and Z coordinates of the landmark and extremal points. Tojudge the accuracy of the results, a complex control fieldwith known X, Y, andZ coordinates was constructed aroundeach bone when photographed. The base of this control fieldconsisted of three pieces of 1.7 cm thick plywood (Fig. 4).Hinges connected the outer pair of boards to the middlepiece so that they could be folded onto this middle piece for
© 2001 NRC Canada
Dilkes 1209
Fig. 3. Locations of midpoints of shafts for the measurement of circumference and cortical bone thickness. (A) Left humerus. (B) Lefttibia. (C) Left pes. (D) Metatarsal III of left pes. Not drawn to scale.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:02 AM
Color profile: DisabledComposite Default screen

transportation. Holes were drilled in the boards at specificintervals. Aluminum pins 5, 8, 10, and 15 cm high weremanufactured to fit into these holes. TheX, Y, andZ coordi-nates of the tops of the pins provided the control fieldaround the fossil. Stereophotographs were taken with a35 mm Nikon FG camera equipped with a 50 mm lens andmounted on a tripod. Kodak Technical Pan (ASA 25) and T-Max (ASA 100) black and white films were used.
Landmarks, extremal points, helping points along curvedsurfaces, and the top of each aluminum pin were marked onblack and white stereophotographs and theirx andy coordi-nates digitized with a Summagraphics Pad. Helping pointsare nonhomologous point selected to describe a curved surfaceand were not included in any morphometric analysis. Aftercalculation of X, Y, and Z coordinates of the landmarks,extremal points, and helping points, distances were calculatedbetween selected pairs of points by Euclidean geometry.Lengths along curved surfaces were approximated by addingdistances between helping points. Errors of calculated distancesbased upon three-dimensional coordinates of landmarksvaried between 1 and 3% of the same distances measuredwith dial calipers. This margin of error is deemed to bewithin an acceptable range of repeatability errors.
In view of the limited number of dimensions that could bemeasured on the femora, it was decided to use a set of largedial calipers to take all linear measurements for this bone.Minimal circumferences of the humerus, femur, tibia, and
metatarsal III were determined by measuring with a set ofsmall dial calipers a thin nylon string wrapped around theshaft. Cortical bone thickness was also measured with dialcalipers.
MorphometricsGiven the disarticulated nature of the nestlings and individuals
from the Camposaur bone bed, it was not possible to iden-tify any ontogenetic changes to the relative proportions ofthe skeleton. Accordingly, limb ratios cannot be calculatedfor Maiasaura. Data were transformed logarithmically toequalize variances of differently sized variables, render thedata independent of scale, and linearize allometric relation-ships (Bookstein et al. 1985). Bivariate expressions ofallometric growth were generated for the femur and tibia ofMaiasaurabecause these bones tend to have greater damageand offer the fewest landmarks. Circumference of thehumerus was analyzed bivariately because the sample sizewas considerably larger than that for the multivariate dataset. Accurate measurements of the circumference of metatar-sal III could be made for only a small subset of availablespecimens, so this dimension was also studied bivariately.Each variable to be analyzed bivariately, with the exceptionof the circumference of metatarsal III, was regressed againstthe maximum length of its respective bone. Circumferenceand cortical bone thicknesses of metatarsal III wereregressed against the scores on the first principal componentof a PCA because greatest length is not isometric (seeResults). Growth lines for all bivariate data were calculatedby reduced major axis regression, a method preferred (e.g.,Rayner 1985; McCardle 1988) for reasons such as scaleinvariance, robustness in situations where data are notbivariately normal, and less bias when error variances areunknown. Isometry (size change without accompanyingshape change) is interpreted when the regression coefficient(represented byk in the bivariate formulay = bxk) is 1.00.Isometry, in the sense employed in this paper, refers to thoseinstances where the organ and body have the same rates ofgrowth. Positive and negative allometry (shape changeaccompanying size change) exists ifk > 1.00 andk < 1.00,respectively. Significance of the calculatedk values wasdetermined byt test. As multiplet tests were conducted forindividual bones, the sequential Bonferroni procedure wasapplied to reduce possible type I errors (Rice 1989). Theresults were accepted as significant at thea/c level, whereais alpha (0.05) andc is the total number of comparisons.
PCA (Jolicoeur 1963) was selected for multivariate analysisof the humerus, ulna, and three metatarsals. In themultivariate generalization of the bivariate formulay = bxk,the relative growth of a biological structure is described bythe eigenvector (vector matrix of coefficients) of the firstprincipal component of a variance–covariance matrix of thelogarithmically transformed dimensions of that structure.The coefficients of this eigenvector are proportional to thespecific relative growth rates of the original dimensions. In amultivariate context, size is represented best as a linear com-bination of variables (Bookstein et al. 1985) with high posi-tive correlations between the variables and the component.Body size of each individual is represented by its score onthe first principal component: larger individuals will havehigher scores on this axis. All components after the first
© 2001 NRC Canada
1210 Can. J. Earth Sci. Vol. 38, 2001
Fig. 4. Schematic drawing of board used for stereophotography.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:04 AM
Color profile: DisabledComposite Default screen
principal component represent allometry-free biologic shape,random variation, and error (measurement and taphonomic).Thus, only the coefficients of the first component describethe implications of size change for shape, and the first com-ponent is described best as an allometric size variable(Bookstein 1989).
In the present study, eigenvectors were normalized (i.e.,summation of squared coefficients equals 1.00). Coefficientsof the first eigenvector were rescaled by [1/(p)½]–1, wherepis the number of variables, to create multivariate allometriccoefficients (Leamy and Bradley 1982; Shea 1985). As withthe bivariate formula, a multivariate allometric coefficient of1.00 indicates isometry, a coefficient greater than 1.00 ispositively allometric, and a coefficient less than 1.00 is nega-tively allometric.
To judge the significance of components and the coeffi-cients of an eigenvector, means and standard errors of thenormalized allometric coefficients were estimated by thejackknife resampling method (Mosteller and Tukey 1977), atechnique that has been shown to be useful in a PCA (Gibsonet al. 1984; McGillivray 1985; Diniz-Filho et al. 1994; Peres1994). A jackknife analysis entails the repeated computationof a sample statistic (e.g., variance) following the randomomission of a single operational taxonomic unit (OTU).Hence, for n OTUs the sample statistic is recalculatedntimes with a sample size ofn – 1. Pseudovalues, defined asyp = nyall – (n – 1)yi, are calculated, whereyall is the samplestatistic calculated with the full sample, andyi is the samplestatistic calculated on the sample size ofn – 1. The estimateof the variance is obtained by dividing the variance of thepseudovalues byn. The standard error is the square root ofthis variance estimate. The mean and standard error of thesepseudovalues can aid in determining the accuracy of thesample statistic based upon the full sample. Significance ofthe principal components was assessed by the ratio of thejackknife estimate and its standard error for each of the coef-ficients of a component (Gibson et al. 1984). This ratio isdesignated byT (Marcus 1990). As the distribution ofjackknifed variances is oftenhighly skewed (Efron 1982),jackknifed estimates andT valueswill also be skewed. Thus,the t distribution suggested by Mosteller and Tukey (1977)and followed by Gibson et al. (1984) for testing theT statis-tic is incorrect. A value forT of 3.00 was chosen in the latterstudy to indicate a significant coefficient. A more conserva-tive probability distribution is recommended by Marcus(1990), and followed in this paper, in which the significantTvalue is raised to 5.00. Only those components with a largenumber of significant coefficients were retained (Gibson etal. 1984). The standard errors for the coefficients of the firstprincipal component allowed tests of isometry byt test.
PCA was carried out with the statistical package NTSYS-pc(Rohlf 1990). Parameters for the bivariate allometric formulawere calculated by a reduced major axis program written byK. Baia, formerly of the Biology Department, Erindale College,University of Toronto.
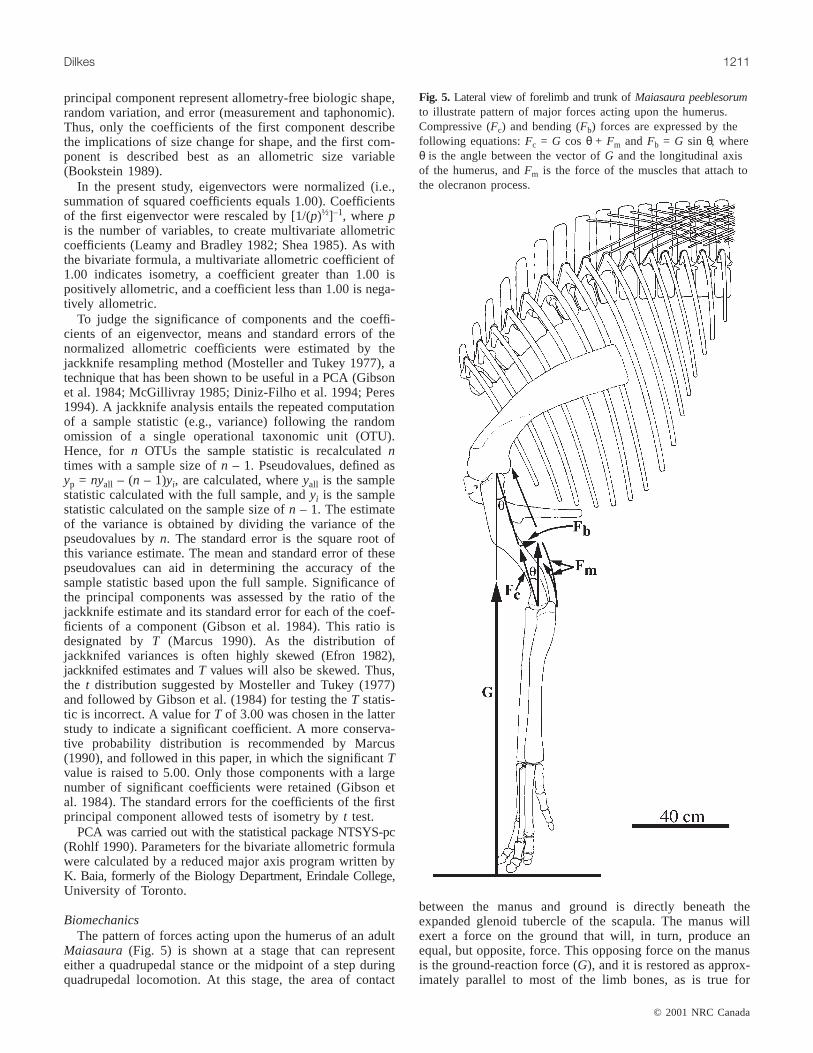
BiomechanicsThe pattern of forces acting upon the humerus of an adult
Maiasaura (Fig. 5) is shown at a stage that can representeither a quadrupedal stance or the midpoint of a step duringquadrupedal locomotion. At this stage, the area of contact
between the manus and ground is directly beneath theexpanded glenoid tubercle of the scapula. The manus willexert a force on the ground that will, in turn, produce anequal, but opposite, force. This opposing force on the manusis the ground-reaction force (G), and it is restored as approx-imately parallel to most of the limb bones, as is true for
© 2001 NRC Canada
Dilkes 1211
Fig. 5. Lateral view of forelimb and trunk ofMaiasaura peeblesorumto illustrate pattern of major forces acting upon the humerus.Compressive (Fc) and bending (Fb) forces are expressed by thefollowing equations:Fc = G cos θ + Fm and Fb = G sin θ, whereθ is the angle between the vector ofG and the longitudinal axisof the humerus, andFm is the force of the muscles that attach tothe olecranon process.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:07 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
1212 Can. J. Earth Sci. Vol. 38, 2001
large quadrupedal mammals such as horses (Biewener et al.1983), though not for smaller mammals that tend to movewith flexed limbs (Biewener 1983). When the head of thehumerus is articulated with the glenoid, the long axis of thehumerus is clearly articulated at an angle to the direction ofG (Fig. 5). Consequently, as the manus was fixed upon theground, the humerus was subject to torque that attempted torotate it counterclockwise (when viewed left laterally) andflex the elbow. Collapse of the forelimb was prevented bythe contraction of muscles, primarily M. triceps, thatattached to the olecranon process. SinceG acted at an angleto the humerus, not only was the whole humerus subject totorque about the glenoid, but also the shaft of the humerusexperienced compressive and bending components ofG(Fig. 5). Bone is stronger in compression than tension(Currey 1984), but bending is the major type of loadingexperienced by limb bones in tetrapods with an uprightstance such as in mammals (Rubin and Lanyon 1982;Biewener and Taylor 1986), in contrast to the predominantlytorsional strains in the limb bones of sprawling tetrapods(Blob and Biewener 1999). Curvature of the bone (Bertram
and Biewener 1992) and loading that is at an angle to thelong axis of the bone are responsible for these bendingstresses. In the subsequent biomechanical analyses, it isassumed that bending was the primary source of stress onthe limb bones ofMaiasaura.
Analysis of the bending of the humeral and tibial shaftsunder loading is based upon the equations of beam theory asoutlined in a number of texts (e.g., Roark and Young 1975;Wainwright et al. 1976). Stresses produced in a beam that isbent about its neutral axis (an axis along which the materialexperiences neither compressive nor tensile stress) areexpressed by the basic beam formula
sMyI
=
whereM is the torque (moment) of the force that is produc-ing the bending,y is the distance from the neutral axis to apoint within the cross section of the beam (typically theouter edge to determine the maximum stress), andI is thesecond moment of area. When comparing two beams subjectto the same torque, the stress can be minimized by maximizingthe ratio I/y.
Torque is the product of the force component (Fb) appliedat the free end of the beam and the distance (L) from the freeend to the plane of the cross section, at which the secondmoment of area is determined. Hence, the standard beamformula can also be expressed as
sF Ly
I= b
Stress can be minimized by diminishingL in addition tomaximizing I/y.
In this study, the humerus and tibia are modelled asstraight beams that are subject to stresses produced fromground-reaction forces during stance and locomotion.Although bending could occur at any point along the lengthof the humerus, it is assumed that the complex pattern ofstresses produced by the attachment of the variousappendicular muscles between the humeral head and the cornerof the deltopectoral crest would contribute to the overallmorphology of the bone. Since the relative proportions ofstresses produced by muscular activity is unknown, it is notpossible to separate the different influences of muscularstresses and bending stresses. Hence, beam theory is notapplicable to this region of the humerus, and only the shaftfrom the distal condyles to the deltopectoral crest will bemodelled as a beam. There is a slight curvature to the shaftof the humerus (Fig. 5), which implies that one should usethe equations for the bending of a curved beam. However,the radius of this curvature is at least a magnitude largerthan the depth of the beam, and thus in this situation anyerrors incurred by using the standard equations for a straightbeam will be small (Roark and Young 1975). There is nodetectable curvature to the shaft of the tibia at any growthstage.
Cross-sectional outlines of the humerus and tibia werereconstructed from measurements of cortical bone thicknessand circumference for the purpose of calculatingI. Theshafts of those few humeri that are undistorted have a virtuallycircular cross section. Others were restored with a circular
Fig. 6. Methods used for determination of the second moment ofarea for the shafts of the humerus and tibia ofMaiasaurapeeblesorum. The second moment of area is calculated for bendingof the humerus about the neutral axesX and Y. Other variablesare A (cross-sectional area),r (radius of medullary cavity), andR(radius of shaft).
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:09 AM
Color profile: DisabledComposite Default screen

cross section. A definite asymmetry between the relativethicknesses of the caudal and cranial sections of corticalbone suggested that bending along the shaft of the humerusshould be studied along two neutral axes: one that is di-rected lateromedially (X), and a second that is directedcraniocaudally (Y; Fig. 6A). The totalI for the humeral shaftabout each neutral axis was determined by dividing the crosssection into 1 mm thick slices, for which the area was calcu-lated graphically and then summing theI of each slice(Fig. 6A). A circular cross-sectional outline of the tibia wasalso reconstructed based upon undistorted specimens ofdifferent sizes. As there is little variation among the corticalbone thicknesses in any direction in the tibial shaft,I wascalculated according to the formula for a thick-walled tube(Fig. 6B). The value ofr (radius of medullary cavity) foreach section was calculated based upon the average of themeasured cortical bone thicknesses.
Calculation of second moment of areas from reconstructionsof cross sections rather than cross sections taken directlyfrom specimens will introduce some error due to deviationsfrom the assumption of circular cross sections. However,even the slight distortion evident in many specimens wouldintroduce a larger error if not corrected. Furthermore, anyerror created through the use of reconstructions is probablysmall compared with the actual changes in cortical bonethicknesses and second moment of areas across the sizerange studied forMaiasaura.
Units of I are linear dimensions raised to the power offour. Therefore, when regressed against length, the line ofisometry will have a slope of 4.00. Positive and negativeallometry are defined as greater than or less than 4.00,respectively.
As an additional measure of the strength of the bone, surfacearea of the cross section was also calculated and regressedagainst greatest length. Since the surface area is proportionalto the square of a linear dimension, isometry is present whenthe regression coefficient is 2.00, positive allometry existswhen the coefficient is greater than 2.00, and negativeallometry exists when the coefficient is less than 2.00.
Results
In each of the PCAs, the first principal componentaccounted for at least 95% of the variance. Eigenvalues ofeach succeeding component attained a maximum in a fewinstances of only 2% and were usually less than 1%. In addi-tion, there is a gradual decline in the eigenvalues beyond thefirst component. Both results suggest that those axes beyondthe first component are not stable. Jackknife resampling pro-vides additional support for this interpretation. AllT valuesfor the first principal component of each analysis are greaterthan 5.00. In contrast, few coefficients of the other compo-nents are greater than 5.00. Only two of the 10 coefficientsfor principal component 2 of the humerus haveT values thatare greater than 5.00, and two of the 13 coefficients for principalcomponent 2 and one of the 13 coefficients for principalcomponent 3 of metatarsal IV haveT values that are greaterthan 5.00. The small and gradually declining eigenvaluesand small number of significant coefficients for those com-ponents beyond principal component 1 demonstrate that theorientation of these axes within multidimensional space is
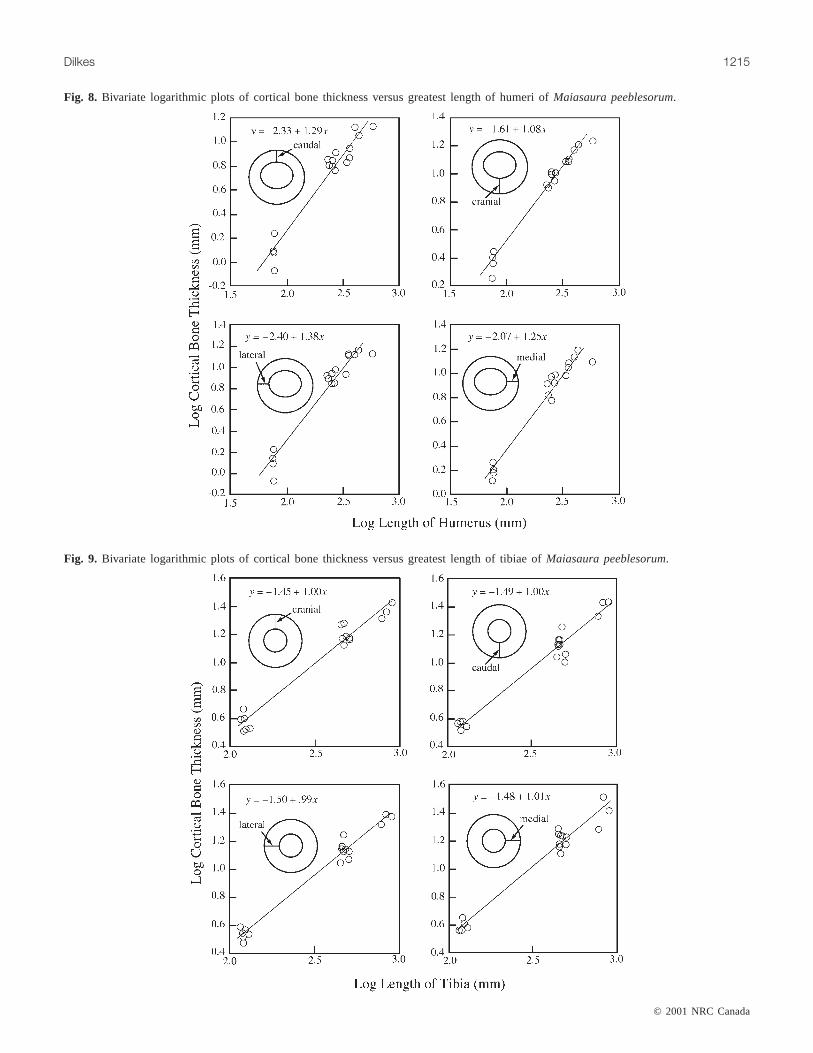
largely arbitrary, and any biological interpretations would bemeaningless. Accordingly, only the first principal compo-nent is discussed. Results of multivariate and bivariate ana-lyzes are summarized in Tables 1–4. Patterns of allometry ofexternal dimensions are also presented in Fig. 7. Graphs ofregressions of cortical bone thickness, surface area of crosssection, and second moment of area versus size are given inFigs. 8–13.
HumerusThere is a distal and lateral displacement of the
deltopectoral crest during growth as shown by positiveallometry of variables 4, 5, and 8 and negative allometry ofvariable 6 (Table 1; Fig. 7A). Minimum circumference ofthe shaft (variable 11) is negatively allometric (Table 2).Negative allometry of the length of the shaft is shown byvariables 6 and 7. Caudal, lateral, and medial dimensions ofcortical bone for the humerus exhibit positive allometry,whereas the cranial dimension is isometric (Table 3; Fig. 8).The second moment of areas for the humeral shaft along theX (lateromedial) andY (craniocaudal) axes are isometricduring growth (Table 4; Fig. 11). Surface area of the shaft ispositively allometric (Table 4; Fig. 13A). Thus, the resultsindicate that the humeral shaft of an adult is stouter than thatin a juvenile, but this stoutness has been achieved by adecrease in the relative width of the marrow cavity (Fig. 14).Nonetheless, there is evidence that the cortical bone of thehumeri of large adults (7 m or greater in body length) is notmarkedly greater than that of smaller adults, indicative of alater ontogenetic expansion of the medullary cavity (Horneret al. 2000).
UlnaResults are suggestive of a positively allometric increase
in the craniocaudal diameter of the shaft (variable 1). How-ever, none of the coefficients are significantly different fromisometry (Table 1).
FemurThere is an allometric increase in the size of the fourth
trochanter as shown by negative allometry of variable 2 andpositive allometry of variable 3 (Table 2; Fig. 7C). Thecircumference of the shaft (variable 4) is negativelyallometric. As the shaft is virtually cylindrical, negativeallometry of its circumference implies negative allometry ofthe diameter.
TibiaThe circumference of the tibia is negatively allometric; all
other measured external dimensions are isometric (Table 2;Fig. 7D). Given the cylindrical shape of the shaft, the nega-tive allometry of the circumference demonstrates negativeallometry of the diameter. All dimensions of cortical bone(Table 3) and the cross-sectional area for the tibial shaft(Table 4) are isometric. In contrast, the second moment ofarea of the tibial shaft is negatively allometric (Table 4).
MetatarsalsAll measured variables for the second metatarsal are iso-
metric (Table 1). The third and fourth metatarsals exhibitsimilar patterns of allometric growth (Tables 1, 2). Greatest
© 2001 NRC Canada
Dilkes 1213
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:10 AM
Color profile: DisabledComposite Default screen

length and some dimensions that are approximately parallelto the longitudinal axis are positively allometric, and only asingle dimension of the distal condyle of the third and fourthmetatarsals is negatively allometric. Circumference of theshaft of the third metatarsal is isometric (Table 2), and thecaudal dimension of cortical bone thickness is positivelyallometric (Table 3).
Discussion
The results demonstrate that the pattern of allometricgrowth for the forelimb was clearly different from that forthe hindlimb. The distinctive patterns of morphometricchange in the limbs are most likely due to stresses experi-enced by the limb bones during growth. Stresses on limbbones of terrestrial vertebrates are produced by gravity andthe muscular forces required for moving the body. Bone canadjust to these stresses by (i) changing the mechanical prop-erties of the bone material, (ii ) changing the angulation ofthe long bones with respect to the direction of the vector ofthe ground-reaction forces, and (iii ) changes in skeletalallometry. Any combination of these changes would maintaina similar safety margin (ratio of failure stress to functionalstress) throughout a size range.
Mechanical properties such as bending strength, compressivestrength, and various elastic moduli (e.g., Young’s modulus)of mature, compact bone in long bones are relatively invariantbetween species of terrestrial mammals and birds of differentsizes (Biewener 1982; Currey 1984) and are not importantfactors in explaining the maintenance of skeletal stresses inanimals of different sizes (Biewener 1990). However, themechanical properties of growing bone in juveniles of mammals(Currey and Pond 1989) and birds (Carrier and Leon
1990) exhibit definite changes that are explained best asadaptive responses to changing functions. The rapidly growingyoung of mammals and birds are characterized by thepresence offibrolamellar bone, which has lower stiffness(i.e., lower Young’s modulus) and lower strength due to itslower content of calcium and greater porosity (Currey andPond 1989; Brear et al. 1990) than adult bone. Limb bonesthat are subject to locomotory stresses at an early age canadjust to these stresses either by increasing the mineralizationof the bone (Currey and Pond 1989; Brear et al. 1990) or byincreasing the amount of bone (Carrier and Leon 1990).Fibrolamellar bone is present in the cortical bone of the limbbones ofMaiasauraat several stages of growth and becomesincreasingly laminar in organization as the individualmatures (Horner et al. 2000).
Limb posture is an important means by which safety factorsare maintained in quadrupedal mammals of greatly differentsizes that have similar proportions (Biewener 1989, 1990;Bertram and Biewener 1990). Small mammals run in asemierect position, whereas larger mammals move withmore upright limbs. Birds also show this size-dependentshift in limb orientation (Gatesy and Biewener 1991). Thereis a closer alignment between the axis of a limb and theground-reaction force in the more vertically oriented limbsof larger tetrapods, which decreases the size of the momentarm of this force and, hence, both the torque about a jointand the bending stress imposed upon the long bone(Biewener 1989). Furthermore, the muscular force requiredto counter the torque is also diminished, which, in turn,decreases the compressive stress on the bone. Posturalchanges may have occurred during growth inMaiasaura, solarge adults walked with more upright limbs. Unfortunately,it cannot be determined if the young ofMaiasaurawalked
© 2001 NRC Canada
1214 Can. J. Earth Sci. Vol. 38, 2001
Fig. 7. Summary of results of bivariate and multivariate morphometric analyses of external dimensions of limb bones ofMaiasaura peeblesorum.Not drawn to scale.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:12 AM
Color profile: DisabledComposite Default screen
© 2001 NRC Canada
Dilkes 1215
Fig. 8. Bivariate logarithmic plots of cortical bone thickness versus greatest length of humeri ofMaiasaura peeblesorum.
Fig. 9. Bivariate logarithmic plots of cortical bone thickness versus greatest length of tibiae ofMaiasaura peeblesorum.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:19 AM
Color profile: DisabledComposite Default screen

with more flexed limbs because there are no clearosteological constraints on the degree of flexion of the hipor knee.
Allometry is a means of compensating for the weaknessof fibrolamellar bone in order for limbs of young birds tofunction at a very early age (Carrier and Leon 1990). It is
also important in the phylogenetic size increase of terrestrialmammals. Geometric similarity (i.e., proportions remainsimilar throughout the size range) has been reported formammals across a size range from a shrew to an elephant(Alexander et al. 1979) and within Carnivora (Bertram andBiewener 1990). A number of studies (Prothero and Sereno
© 2001 NRC Canada
1216 Can. J. Earth Sci. Vol. 38, 2001
Fig. 10. Bivariate logarithmic plots of cortical bone thickness versus score along principal component 1 of metatarsal III ofMaiasaurapeeblesorum.
Fig. 11. Bivariate logarithmic plots of second moment of area versus greatest length of humeri ofMaiasaura peeblesorum.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:24 AM
Color profile: DisabledComposite Default screen

1982; Economos 1983; Bertram and Biewener 1990; Cole1992) have shown that very large mammalian species suchas the ceratomorphs (extant tapirs, rhinoceroses, and theextinct members of this clade) and large bovids depart fromgeometric scaling and often show pronounced positiveallometry of long bone dimensions. Skeletal allometry invery large species produces more robust bones for dealingwith stresses beyond those that can be accommodated by achange in posture (Biewener 1990). Although the aboveallometric patterns in large mammals are phylogenetic ratherthan ontogenetic, the principle is the same in both instances.Gravity imposes far greater stresses on larger individuals,whether that size increase is phylogenetic within a lineage orontogenetic.
The positively allometric growth of the deltopectoral crestof the humerus can be related to ontogenetic changes ineither the probable size or attachment sites of pectoralmuscles. Scars that have been associated phylogeneticallywith a common insertion of M. deltoides clavicularis and M.scapulohumeralis anterior (Dilkes 2000) are concentratedalong the caudal surface of the deltopectoral crest nearest thedistal corner in all observed growth stages. Thus, the distaland lateral ontogenetic shift of the distal corner of thedeltopectoral crest suggests a distal shift in the attachmentsite of these muscles. Contraction of these muscles willelevate and protract the humerus in conjunction with othermuscles with similar actions such as M. deltoides scapularisand M. latissimus dorsi. In addition, a more distally posi-tioned attachment site may act to offset bending momentsacting upon the humeral shaft as the pectoral limb iselevated by decreasing the distance between thedeltopectoral crest and the distal condyle (Currey 1984).
Musculus latissimus dorsi is a particularly interestingmuscle ofMaiasaura, because there is a clear change in themorphology of its insertional scar on the humerus, whichsuggests an ontogenetically increasing usage of the muscle.
The humerus of an embryonicMaiasaurahas an ill-definedshallow pit at the caudal end of a groove on the caudal surfaceof the deltopectoral crest. This pit and groove have muchsharper boundaries in a nestling and are accordingly moreeasily identified (Figs. 15A, 15B). During growth, this scarfor M. latissimus dorsi changes from a pit to a prominentbump composed of highly fibrous bone (Figs. 15C, 15D).According to a customary assumption of proportionalitybetween the rugosity of a muscle scar and the force pro-duced by its muscle, an adult ofMaiasaura used M.latissimus dorsi more consistently than a nestling. The tensionproduced by M. latissimus dorsi would be greater in individualsolder than a nestling, and additional bone was laid down forbetter anchorage of the tendon. However, empirical supportfor this assumption is limited, and it is not valid in thoseinstances where muscle scars are faint or absent (Bryant andSeymour 1990). ForMaiasaura, the scar for M. latissimusdorsi is one of the most readily identifiable scars at allstages of growth and is the only one that exhibits morpho-logical changes during growth. Depending upon thecraniocaudal extent of the likely origin of M. latissimusdorsi from the fascia of the dorsal vertebrae, middle andposterior fibres may have acted to propel the trunk forwardsif an individual moved quadrupedally. Elevation of theforelimb and propulsion of the trunk are likely plesiomorphicactions of M. latissimus dorsi (Jenkins and Goslow 1983).
The entire cranial surface of the deltopectoral crest has acontinuous muscle scar from the probable insertion of M.supracoracoideus and M. pectoralis (Dilkes 2000). A positivelyallometric increase in the size of this scar suggests a probablecorresponding allometric increase in the size of at least oneof these appendicular muscles. When a terrestrial tetrapodmoves quadrupedally, M. supracoracoideus and M. pectoralisact primarily during the propulsive stage (Jenkins andGoslow 1983). Musculus supracoracoideus is a stabilizer ofthe shoulder joint in tetrapods. Musculus pectoralis is one ofa group of postural muscles that helps to support terrestrialvertebrates with erect posture by preventing collapse of thelimbs. Possible ontogenetic enlargement of either M.supracoracoideus or M. pectoralis inMaiasaura may becorrelated with quadrupedal locomotion through increasedmuscular moments needed for postural support and stabilizationof the shoulder joint. In addition, relatively larger attachmentareas for one or both of these muscles suggest a greaterrange of lever arms for the muscle fibres. Those fibres withlever arms significantly different from the distance from theglenoid to the origin of the muscles will tend to generatemaximum torque later during flexion and thus continueflexion beyond the point at which other fibres reach theirmaximum torque (Mameren and Drukker 1979). Since theforelimb remains in contact with the ground during quadru-pedal movement, the trunk is moving forward relative to theforelimb during propulsion and the angle between the trunkand forelimb is changing continually. Hence, the positivelyallometric increase in the attachment area for this pair ofpostural muscles suggests that in the adults different parts ofthe muscles will generate maximum torque at different timesthroughout the propulsive stage and serve to stabilize theshoulder.
For the femur, positive allometry of the fourth trochanterindicates a corresponding increase in the relative size of M.
© 2001 NRC Canada
Dilkes 1217
Fig. 12. Bivariate plot of second moment of area versus greatestlength of tibiae ofMaiasaura peeblesorum.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:25 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
1218 Can. J. Earth Sci. Vol. 38, 2001
caudifemoralis (Dilkes 2000) during growth. As the primaryfemoral retractor muscle, allometric enlargement of M.caudifemoralis would increase the moment generated by thismuscle and would, in turn, correlate with the significantlygreater mass of the adult.
Positive allometry of several longitudinal dimensions andthe isometry or negative allometry of the proximal and distalends of the third and fourth metatarsals indicate that the pesof an adult ofMaiasaurais more slender than that of a juvenile.A more slender pes in a heavier animal might suggest thatthe bending stresses experienced by the metatarsals wouldbe disproportionately larger. However, it is likely that thelarge heel pad that is shown clearly in tracks of hadrosaurs(Langston 1960; Currie et al. 1991) would have dissipatedmuch of the force generated by the pes striking the ground(Alexander et al. 1986). This pad likely reduced the peakmagnitude of bending stresses.
According to beam theory, the main factors that determinethe ability of a beam to resist bending areI and L. As onewould conclude intuitively, a short (smallL) and wide (largeI) cylinder is less easily bent than one that is long andnarrow. Growth ofI for the humeral shaft ofMaiasaura isisometric, which is unexpected because if there is a dispro-portionate increase in stresses on the forelimb due to anontogenetic delay in its use, thenI should show a similarallometric increase. However, positive allometry of corticalbone width and cross-sectional area implies that the humeralshaft of an adult is more resistant to bending stresses thanthat in a juvenile, even though its value ofI maintains thesame proportion to size. Negative allometry ofL (variable 6for the humerus in Table 1) will reduce stress further. In con-trast, the nearly consistent pattern of isometry for corticalbone dimensions and cross-sectional surface area of the tibialshaft and metatarsal III and the negative allometry ofI forthe tibia suggest that isometric growth was sufficient in thehindlimb to withstand ontogenetically increasing locomotorystresses.
Two explanations may be offered for the dramaticallydifferent patterns of allometry in the forelimb and hindlimb.One explanation is that individuals ofMaiasaura were
quadrupedal throughout life, but the percentage of masscarried by the forelimbs increased during growth. Analternative explanation is that juveniles may have walkedpredominantly as bipeds, but adopted an increasinglyquadrupedal stance as they matured. Both explanationsrequire the use of the forelimb for quadrupedal locomotion.The primary difference resides in the stage of growth atwhich quadrupedal locomotion is introduced. It is importantto note that neither explanation restricts the forelimb tolocomotion; each states merely that the strains encounteredduring locomotion are most likely to be the primary factorresponsible for the observed allometric growth pattern.Hadrosaurs probably used their forelimbs for other behav-iors such as manipulation of vegetation while foraging andthe construction of nests. These activities would certainlyplace stresses upon the bones, but any strains experiencedwould be transitory and in the case of nest building wouldlikely apply solely to the sexually mature adults.
The first explanation for the different growth patterns ofthe forelimb and hindlimb requires a cranial shift of the centreof mass during growth so that the forelimb carries a steadilyincreasing proportion of the body mass. Although no pub-lished calculation of the centre of mass for a hadrosaur isavailable, it has been estimated for the iguanodontianIguanodon(Alexander 1985), which has a body shape similarto that of a large hadrosaur. It was found that the centre ofmass was situated virtually above the pelvis, and only asmall percentage of the body mass was likely supported bythe forelimbs. Presumably the forelimbs of adult hadrosaursalso supported only a small percentage of the body mass.Without a reasonably accurate estimate of the relative loca-tions of the centres of mass of a juvenile and an adult ofMaiasauraand the magnitude of the increase in mechanicalloading on the forelimbs, it is unknown whether anyontogenetic shift in the centre of mass occurred and if itcould reasonably account for the differing allometricresponses of the forelimb and hindlimb to mechanical loads.
The second and more likely explanation is that theforelimbs were not used for locomotion until later in life.Given the likelihood that the forelimb only supported a
Fig. 13. Bivariate plots of surface area of shaft versus greatest length of humeri and tibiae ofMaiasaura peeblesorum.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:28 AM
Color profile: DisabledComposite Default screen
© 2001 NRC Canada
Dilkes 1219
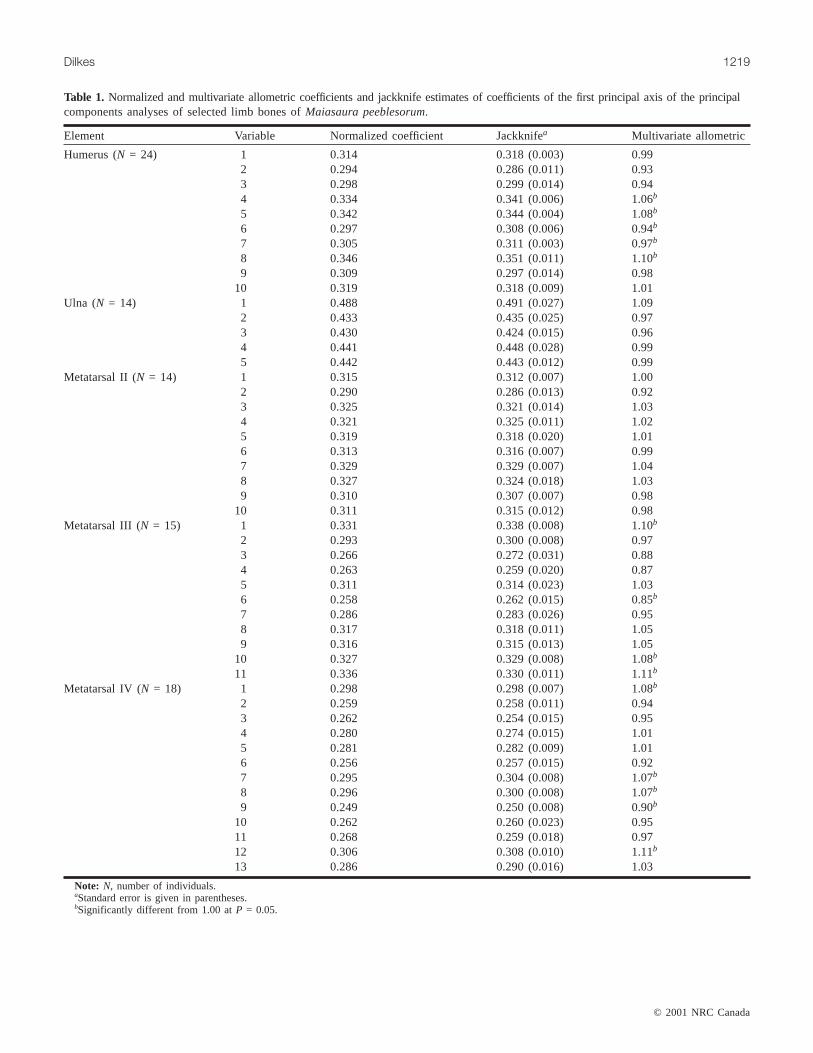
Element Variable Normalized coefficient Jackknifea Multivariate allometric
Humerus (N = 24) 1 0.314 0.318 (0.003) 0.992 0.294 0.286 (0.011) 0.933 0.298 0.299 (0.014) 0.944 0.334 0.341 (0.006) 1.06b
5 0.342 0.344 (0.004) 1.08b
6 0.297 0.308 (0.006) 0.94b
7 0.305 0.311 (0.003) 0.97b
8 0.346 0.351 (0.011) 1.10b
9 0.309 0.297 (0.014) 0.9810 0.319 0.318 (0.009) 1.01
Ulna (N = 14) 1 0.488 0.491 (0.027) 1.092 0.433 0.435 (0.025) 0.973 0.430 0.424 (0.015) 0.964 0.441 0.448 (0.028) 0.995 0.442 0.443 (0.012) 0.99
Metatarsal II (N = 14) 1 0.315 0.312 (0.007) 1.002 0.290 0.286 (0.013) 0.923 0.325 0.321 (0.014) 1.034 0.321 0.325 (0.011) 1.025 0.319 0.318 (0.020) 1.016 0.313 0.316 (0.007) 0.997 0.329 0.329 (0.007) 1.048 0.327 0.324 (0.018) 1.039 0.310 0.307 (0.007) 0.98
10 0.311 0.315 (0.012) 0.98Metatarsal III (N = 15) 1 0.331 0.338 (0.008) 1.10b
2 0.293 0.300 (0.008) 0.973 0.266 0.272 (0.031) 0.884 0.263 0.259 (0.020) 0.875 0.311 0.314 (0.023) 1.036 0.258 0.262 (0.015) 0.85b
7 0.286 0.283 (0.026) 0.958 0.317 0.318 (0.011) 1.059 0.316 0.315 (0.013) 1.05
10 0.327 0.329 (0.008) 1.08b
11 0.336 0.330 (0.011) 1.11b
Metatarsal IV (N = 18) 1 0.298 0.298 (0.007) 1.08b
2 0.259 0.258 (0.011) 0.943 0.262 0.254 (0.015) 0.954 0.280 0.274 (0.015) 1.015 0.281 0.282 (0.009) 1.016 0.256 0.257 (0.015) 0.927 0.295 0.304 (0.008) 1.07b
8 0.296 0.300 (0.008) 1.07b
9 0.249 0.250 (0.008) 0.90b
10 0.262 0.260 (0.023) 0.9511 0.268 0.259 (0.018) 0.9712 0.306 0.308 (0.010) 1.11b
13 0.286 0.290 (0.016) 1.03
Note: N, number of individuals.aStandard error is given in parentheses.bSignificantly different from 1.00 atP = 0.05.
Table 1. Normalized and multivariate allometric coefficients and jackknife estimates of coefficients of the first principal axis of the principalcomponents analyses of selected limb bones ofMaiasaura peeblesorum.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:28 AM
Color profile: DisabledComposite Default screen
minor amount of the body mass, any delay in the adoptionof quadrupedal locomotion would generate a pattern ofstrains greatly different and significantly greater in magni-tude from the earlier pattern. It is generally accepted that dy-namic rather than static mechanical loading will stimulatethe formation of new bone (Lanyon and Rubin 1984;Lanyon 1992), and even a brief period of dynamic loading issufficient (Forwood et al. 1996). Transitory or static loadswill not affect bone mass. Although there is evidence forboth a threshold response to dynamic strains and a linear in-crease in bone formation with the magnitude of the strain(Rubin and Lanyon 1985; Turner et al. 1994), it appears thatbone will respond more to a change in the strain rather thansimply an increase in the absolute magnitude of strain. Fur-thermore, the amount of bone formation is directly propor-tional to the rate of change in strain: a large change in therate of strain will stimulate more formation of bone than a
small change (O’Connor et al. 1982; Turner et al. 1995).Bones of mature and growing terrestrial animals will appar-ently maintain their architecture over a range of daily dy-namic strains, but will change their shape and mass whensubjected to unusual dynamic loading that is different fromthese normal daily strains (Mosley et al. 1997). This adap-tive modelling and remodelling is necessary to maintain thefunctional loading within specific limits to prevent damage(Keller and Spengler 1989; Biewener and Bertram 1993).Thus, a sharp increase in the use of the forelimbs ofMaiasaura for locomotion as an individual grew in sizewould lead to dramatic increases in stresses, and the shaft ofthe humerus apparently adjusted to these new stresses. Theprobable allometric increases in the size of postural musclesand the lever arms of protractor muscles that attach to thehumerus are also consistent with this explanation. The re-sults presented herein of an age-dependent preference forstance agree with those based upon ontogenetic changes inlimb ratios for I. bernissartensis(Norman 1980).
There are some intriguing changes to the cross-sectionalshape of the shaft of the humerus that merit comment. Anestling has a humerus with an elliptical and offset marrowcavity, whereas the marrow cavity of an adult humerus betterapproximates a hollow cylinder (Fig. 14). In contrast, thecross-sectional shape of the tibial shaft remains essentially asymmetrical hollow cylinder from nestling to adult. A beamthat has a more cylindrical cross-sectional shape is strongerand stiffer and thus more resistant to torsional stresses(Roark and Young 1975). The change for the humerus shaftmight then indicate an enhanced resistance to torsion or thatboth craniocaudal and mediolateral bending generatedsignificant stresses. Either way, the change in cross-sectionalshape of the shaft is consistent with adaptive remodelling toa pattern of stresses that a forelimb would experience if theanimal was a quadruped. In addition, the variability in thecross-sectional shape of the bipedal nestling is seeminglygreater than that of the quadrupedal adult (Fig. 14). Thisobservation is similar to the findings of Biewener andBertram (1994) on the effect of use and disuse on growingbone. They noted increased variation in cross-sectionalshape of the tibia in those birds that were denervated. Theirconclusion was that in the absence of normal functionalloading, the usual degree of relative resorption and depositionrequired to maintain a specific bone shape is disrupted. As a
© 2001 NRC Canada
1220 Can. J. Earth Sci. Vol. 38, 2001
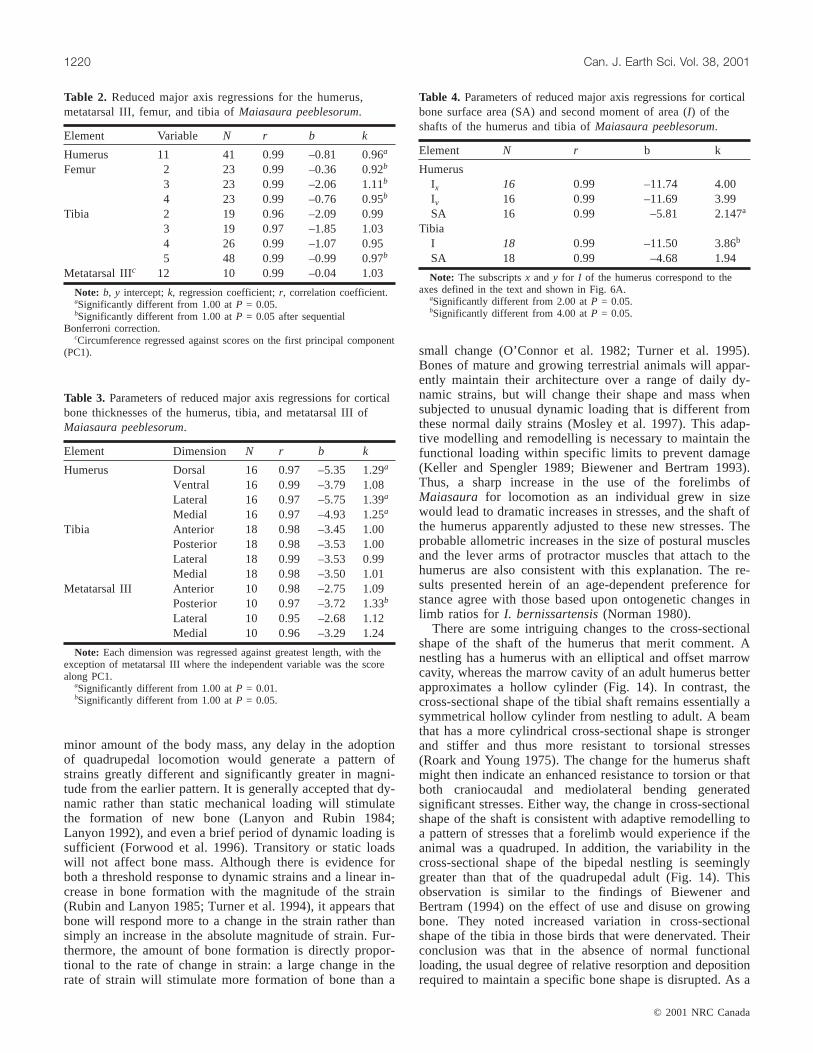
Element Variable N r b k
Humerus 11 41 0.99 –0.81 0.96a
Femur 2 23 0.99 –0.36 0.92b
3 23 0.99 –2.06 1.11b
4 23 0.99 –0.76 0.95b
Tibia 2 19 0.96 –2.09 0.993 19 0.97 –1.85 1.034 26 0.99 –1.07 0.955 48 0.99 –0.99 0.97b
Metatarsal IIIc 12 10 0.99 –0.04 1.03
Note: b, y intercept;k, regression coefficient;r, correlation coefficient.aSignificantly different from 1.00 atP = 0.05.bSignificantly different from 1.00 atP = 0.05 after sequential
Bonferroni correction.cCircumference regressed against scores on the first principal component
(PC1).
Table 2. Reduced major axis regressions for the humerus,metatarsal III, femur, and tibia ofMaiasaura peeblesorum.
Element Dimension N r b k
Humerus Dorsal 16 0.97 –5.35 1.29a
Ventral 16 0.99 –3.79 1.08Lateral 16 0.97 –5.75 1.39a
Medial 16 0.97 –4.93 1.25a
Tibia Anterior 18 0.98 –3.45 1.00Posterior 18 0.98 –3.53 1.00Lateral 18 0.99 –3.53 0.99Medial 18 0.98 –3.50 1.01
Metatarsal III Anterior 10 0.98 –2.75 1.09Posterior 10 0.97 –3.72 1.33b
Lateral 10 0.95 –2.68 1.12Medial 10 0.96 –3.29 1.24
Note: Each dimension was regressed against greatest length, with theexception of metatarsal III where the independent variable was the scorealong PC1.
aSignificantly different from 1.00 atP = 0.01.bSignificantly different from 1.00 atP = 0.05.
Table 3. Parameters of reduced major axis regressions for corticalbone thicknesses of the humerus, tibia, and metatarsal III ofMaiasaura peeblesorum.
Element N r b k
HumerusIx 16 0.99 –11.74 4.00Iy 16 0.99 –11.69 3.99SA 16 0.99 –5.81 2.147a
TibiaI 18 0.99 –11.50 3.86b
SA 18 0.99 –4.68 1.94
Note: The subscriptsx and y for I of the humerus correspond to theaxes defined in the text and shown in Fig. 6A.
aSignificantly different from 2.00 atP = 0.05.bSignificantly different from 4.00 atP = 0.05.
Table 4. Parameters of reduced major axis regressions for corticalbone surface area (SA) and second moment of area (I) of theshafts of the humerus and tibia ofMaiasaura peeblesorum.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:28 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
Dilkes 1221
result, the variability of the cross section is greater. Thedifferences in the variability of cross-sectional geometries ofjuvenile and adult humeri ofMaiasauramay have a similarexplanation. The decrease in the variability of the cross-sectional shape of the shaft of the humerus and its moresymmetrical geometry supports the hypothesis that theforelimb of Maiasauradid not experience a consistent patternof strains until some time after the individual began to moveabout on its hindlimbs.
The hypothesis of an age-dependent change from predom-inantly bipedality to quadrupedality inMaiasaura is con-trasted by the results of a biomechanical study of femoralgrowth in the basal iguanodontianD. lettowvorbecki(Heinrichet al. 1993). It was proposed that juveniles ofDryosauruswere quadrupedal and underwent a shift to bipedality earlyin life, perhaps within the first several months after hatching.This transition was thought to be due to changes in the pro-portions of the body during growth. Although no hatchlingsof D. lettowvorbeckiare known, juveniles of the relatedornithopodsOrodromeus makelaiand M. peeblesorumhaveheads that appear to be larger relative to their bodies than inthe adults. It was concluded that the juveniles ofD. lettowvorbeckiprobably had similar proportions. Withtheir relatively larger heads, the centre of mass was far cranialto the hips, and quadrupedality was necessary to reducestresses on the femur. As the individual matured, its head be-came relatively smaller and the tail relatively larger and thecentre of mass moved closer to the pelvis. Bipedality wasnow the normal stance, as the heavier tail could counterbal-ance the trunk and head.
Data presented in this paper and additional factors suggestthat any change in stance forD. lettowvorbeckimay nothave been from quadrupedal to bipedal. It is now possible tomake more accurate statements about the relative propor-
tions of a juvenileD. lettowvorbeckibecause a juvenile ofDryosaurus altushas been described recently (Carpenter1994). Although a different species, the postcrania of adultsof D. lettowvorbeckiand D. altus are very similar (Galton1981) and it is unlikely that the juveniles would differgreatly. The skull of a juvenile ofD. altus is not nearly aslarge relative to the femur (Carpenter and Alf 1994), as hadbeen suggested forD. lettowvorbeckiby Heinrich et al.(1993, their Fig. 9). Consequently, the centre of mass maynot have been located a great distance cranial to the pelvis injuveniles.
The femur of D. lettowvorbecki has a pronouncedcraniocaudal curvature. Curvature of a long bone increasesbending stresses for a bone under axial compression in thedirection of the curvature (Bertram and Biewener 1988).Femoral curvature is present in all known growth stages ofD. lettowvorbecki(Galton 1981) and the juvenileD. altus(Carpenter 1994); hence, it is reasonable to propose that thecraniocaudal femoral curvature ofD. lettowvorbeckiwouldhave enhanced stresses in this direction relative to otherdirections throughout ontogeny. Heinrich et al. (1993) founda significant increase in the ratio of the maximumI and theminimum I between the small- and medium-sized groups ofD. lettowvorbecki, which they interpreted as evidence for adramatic increase in bending stresses that accompanied ashift from quadrupedality to bipedality. As the direction ofthis ratio is nearly the same as the femoral curvature, it ispossible that the observed changes in biomechanics may rep-resent an adjustment to the craniocaudal stresses created bythe curvature of the femur.
Weight-bearing and the forelimb of hadrosaursThe hypothesis of age-dependent selection of stance in
Maiasauraadvanced in this paper and trackways of quadrupedal
Fig. 14. Representative cross sections of the shafts of humeri ofMaiasaura peeblesorum. (A) Two nestlings. (B) Two adults.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:31 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
1222 Can. J. Earth Sci. Vol. 38, 2001
ornithopods attributed to hadrosaurs (Currie et al. 1991) areseemingly at odds with the general consensus that theforelimb of hadrosaurs shows no clear adaptations forweight-bearing. Thus, there is an apparent mismatchbetween the combined evidence from morphometrics,biomechanics, and ichnology and the morphology of theskeleton that should be addressed.
At the heart of all discussions of the possible functions ofthe forelimb in hadrosaurs has been the manus. Earlier inter-
pretations (e.g., Leidy 1858; Cope 1883) of hadrosaurs asstrictly aquatic swamp dwellers received a significant boostfollowing the discoveries by Charles M. Sternberg in 1908and 1910 of two “mummified” hadrosaurs that have amitten-like manus. Strictly aquatic habits for hadrosaurswere supposedly demonstrated by the presence of skin thatcompletely covered the manus and extended between thedigits to create webbing and the absence of tubercles or cal-luses on the skin impressions (Osborn 1912). Additional
Fig. 15. Development of the muscle scar on the humerus ofMaiasaura peeblesorumthat is attributed to M. latissimus dorsi (Dilkes2000). (A, B) From a nestling. (C, D) From an adult. l.d., M. latissimus dorsi.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:43 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
Dilkes 1223
osteological evidence for aquatic hadrosaurs included thesmall size of the forelimb relative to the hindlimb, theslenderness of the metacarpals, poor articulation of the phalan-ges, and the presence of only two hoof-like unguals (Brown1908, 1912). Lull and Wright (1942) interpreted hadrosaursas bipedal, aquatic herbivores that used their forelimbs forsupport only when resting along a shore, and Sternberg(1965) stated that hadrosaurs were capable of some quadrupedalwalking along the shores of swamps or rivers.
The prevalent opinion of hadrosaurs as aquatic dinosaurswas overturned by Ostrom (1964) and Galton (1970), whoamassed a wide array of evidence to support a new interpre-tation of hadrosaurs as bipedal, terrestrial herbivores.Ostrom pointed out that the evidence presented by Osborn(1912) showed only that hadrosaurs were not quadrupedal; itdid not show that they were aquatic. As further evidence infavor of only bipedal walking, Ostrom argued that the re-duction of carpal ossification to only a pair of small bonesimplied a weak carpus that was incapable of supporting asignificant portion of body mass. Galton agreed thathadrosaurs were bipedal except when resting, but noted thatreduction of the carpus did not necessarily indicate a weakjoint, since the ceratopsidMonocloniusalso has only twocarpals. A poorly ossified carpus, slender metacarpals, andpoorly articulated phalanges are certainly valid observationsfor the hadrosaurian manus, but other features of the manusthat can be interpreted as adaptations for weight-bearinghave been neglected in the descriptions by Brown (1912)and Parks (1920) and more recently Nelms (1992).
The ground-reaction force would exert a torque about thecarpus if that force was inclined relative to the metacarpals.Without the presence of a complex of ossified carpals tovirtually lock the manus and antebrachium to the carpus asdescribed inI. bernissartensis(Norman 1980), or at leastlimit mobility, even a slight misalignment of the axis of themanus with the ground-reaction force could lead to itscollapse. Thus, the manus of hadrosaurs would be at a con-tinual risk of failure as the vector of the ground-reactionforce changed during locomotion. Use of the forelimb at anypoint during locomotion would then be excluded or, at best,restricted to a quadrupedal stance. However, the presence ofprominent muscle scars on metacarpals III and IV identifiedphylogenetically as likely attachment sites for extensor andflexor muscles (Dilkes 2000) suggests that these musclescould have acted to constrain flexibility of the carpus and,
hence, replaced functionally the carpals. Contraction ofthese muscles would help to counter any rotation of themetacarpals about the carpus by stretching the tendons anddecelerating the manus rapidly as it contacted the ground.Although it is not possible to estimate the relative propor-tions of tendon and muscle fibres in the flexor muscles, it islikely that the length of the tendon was significant becausetendons typically transmit the force of muscles in the manusand pes of terrestrial vertebrates. An alternative hypothesisthat the extensor and flexor muscles served to manipulatethe manus is less likely because the slight curvature of thearticular surfaces on the metacarpals and the antebrachiumimplies a limited range of flexion and extension. A largerrange of flexibility was possible between the metacarpalsand phalanges and between the individual phalanges, but thesoft tissue impressions of the mummified hadrosaur andtrackways show that digits II–IV were held together andonly digit V could move independently.
The three primary metacarpals (II–IV) were united firmlyin life as shown by the close apposition between themetacarpals along their entire lengths. In addition to theirtight connection, the metacarpals join along a narrow arc,with metacarpal III situated cranial to its neighbours to producea semicircular cross section (Fig. 16A). The significance ofthis arrangement can be seen readily when one models themetacarpals as a beam that is bent along theX–X axis. Thisaxis lies within a transverse plane and is the axis alongwhich bending is most likely to occur while the animal isstanding or moving. The second moment of area for a crosssection through the middle of the three metacarpals can becompared to the second moment of area of a hypotheticalmanus in which the metacarpals are aligned transversely asan ellipse (Fig. 16B). The method that was used for calculatingthe second moment of area of the shaft of the humerus(Fig. 6) was also used to calculate the second moment ofarea for the actual cross section of the metacarpals. A formulawas used to calculate the second moment of area for thehypothetical elliptical cross section (Fig. 16B).
The second moment of area along theX–X axis for theactual cross section is approximately 39% greater than thatof an ellipse with the same length and width. Furthermore,the ratioI/y is approximately 15% larger for the actual crosssection than for the elliptical cross section. Thus, the articu-lation of the metacarpals along an arc produces a structurethat is stiffer and subject to less bending strain than one with
Fig. 16. (A) Midpoint cross-sectional outline of articulated metacarpals II–IV of an adult ofMaiasaura peeblesorum. (B) Hypotheticalelliptical outline with length and width equivalent to that of (A). Second moment of area of (A) was calculated by the same method asfor the humeral shaft of Fig. 6, whereas the second moment of area for the elliptical outline was determined by the given formula.Bending is assumed to occur along the lateromedial axisX–X. Variablesa and b are the maximum and minimum radii of the ellipse,respectively.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:44 AM
Color profile: DisabledComposite Default screen

an elliptical cross section. The strong union between meta-carpals II–IV and their articulation along a curved ratherthan a transverse plane are interpreted herein as adaptationsfor weight-bearing.
The statement by Brown (1912) that the phalanges of thehadrosaurian manus are articulated loosely was probably areference to the absence of the type of interphalangeal artic-ulation found in the pes, where rounded distal condyles fitrelatively snugly into proximal concavities on the succeedingphalanx. Interphalangeal joints inMaiasaura and otherhadrosaurs consist of contact between flat or roundedsurfaces with little evidence for constraints on direction ofmobility. However, as digits II–IV of the manus wereenclosed in skin, interphalangeal movement was probablylimited. The greatest degree of flexibility probably occurredbetween the metacarpals and the first phalanges. The articularsurfaces on the distal condyles of the metacarpals are extensiveand notably are larger on the cranial surface than on the caudalsurface (Dilkes 1993), contrary to the statements of Nelms(1992) that the distal ends of hadrosaurid metacarpals areflat. The greater cranial portion of the articular surfaceimplies that the manus could be hyperextended, which iscompatible with a role of support. Curvature of the distalcondyle of metacarpal III is modest, but more pronouncedfor the distal condyles of metacarpals II and IV (Dilkes1993). Furthermore, the distal condyles of metacarpals IIand IV face outwards from the middle metacarpal III as aconsequence of the curved structure of the manus. Hence,the digits spread apart to create a larger surface area for contactwith the ground. The webbing between the digits that earlierresearchers thought was an adaptation for paddling throughthe water are probably displaced digital pads (Bakker 1986).Well-preserved manus prints (Currie et al. 1991; Currie1995) show that the pads of digits II–IV were substantialrelative to the unguals and spread out as the manuscontacted the ground. As with the large heel pad of the pes,the digital pads of the manus would absorb and dissipateimpact forces. Tendons within the manus would also stretchand further dampen the peak stress.
Summary
Since the first description of an articulated forelimb of ahadrosaur, several different lines of evidence have been usedto argue that the forelimb was at best a paddle for swimmingor could support only a small fraction of weight when onland. Quadrupedal walking was virtually excluded. Althoughpresent opinion is that hadrosaurs were facultative bipeds,possible ontogenetic variability in the relative importance ofbipedality and quadrupedality has been considered onlyrarely for ornithopods. Furthermore, the contradictionbetween an assumption of quadrupedality (even if limited)as indicated by trackways and the apparent lack of any clearadaptations for weight-bearing in the forelimb has not beenaddressed. Results from the present study have revealed adichotomy in the morphometrics of growth in the forelimband hindlimb of M. peeblesorumthat is consistent with ahypothesis that the preferred stance changed from bipedalityto quadrupedality during growth. There are numerousfeatures in the manus of hadrosaurs that can be interpretedas adaptations for weight-bearing. The carpus with its
reduced number of carpals was apparently reinforced againstcollapse by the strong attachment of flexor tendons onto thecaudal faces of metacarpals III and IV, metacarpals II–IVwere united firmly and articulated in an arc configurationthat was more resistant to bending than if the metacarpalswere aligned in a transverse row, hyperextension was possibleat the metacarpal–phalangeal joints, the phalanges werebound together by skin and probably ligaments, and themanus had a broad contact with the ground.
Acknowledgments
This paper is a revised portion of a doctorate thesissubmitted to the Department of Zoology at the University ofToronto. I wish to thank my thesis supervisor Dr. RobertReisz for his support throughout my research and Dr. JohnHorner for the opportunity to studyM. peeblesorum.Commentsfrom Drs. R. Reisz, J. Horner, C.S. Churcher, C.McGowan, and D.H. Collins are greatly appreciated. Numerousdiscussions with Dr. J. Horner and Mr. Matt Smith onhadrosaurian anatomy, myology, and biomechanics helped toclarify my ideas. Comments and suggestions by Dr. Hans-DieterSues (Department of Palaeobiology, Royal Ontario Museum)were very helpful. I also thank Dr. N.C. Collins (Departmentof Zoology, Erindale College, University of Toronto) for hiscritical reading of the statistics and alerting me to theBonferroni test. I am grateful to Dr. John Rensberger(Thomas Burke Memorial Museum, University of Washington)for providing a copy of the photogrammetry programGeoInter 3.2 and his advice on its operation. Many of thespecimens incorporated into this study were found duringfieldwork at the Camposaur site in the summers of 1988–1990.This fieldwork was aided greatly by the many individualswho participated in the Museum of the Rockies PaleontologySummer Field School, and I wish to thank them for theirefforts. Partial support for my research was provided by apostgraduate scholarship from the Natural Sciences andEngineering Research Council of Canada and two researchgrants from the Geological Society of America.
References
Alexander, R.M. 1985. Mechanics of posture and gait of some largedinosaurs. Zoological Journal of the Linnean Society,83: 1–25.
Alexander, R.M., Jayes, A.S., Maloiy, G.M.O., and Wathuta, E.M.1979. Allometry of the limb bones of mammals from shrews(Sorex) to elephant (Loxodonta). Journal of Zoology (London),189: 305–314.
Alexander, R.M., Bennett, M.B., and Ker, R.F. 1986. Mechanicalproperties and function of the paw pads of some mammals.Journal of Zoology (London),209: 405–419.
Bakker, R.T. 1986. The dinosaur heresies. William Morrow andCompany Inc., New York.
Benton, M.J., Juul, L., Storrs, G.W., and Galton, P.M. 2000. Anatomyand systematics of the prosauropod dinosaurThecodontosaurusantiquusfrom the upper Triassic of southwest England. Journalof Vertebrate Paleontology,20: 77–108.
Bertram, J.E.A., and Biewener, A.A. 1988. Bone curvature: sacri-ficing strength for load predictability? Journal of TheoreticalBiology, 131: 75–92.
© 2001 NRC Canada
1224 Can. J. Earth Sci. Vol. 38, 2001
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:45 AM
Color profile: DisabledComposite Default screen

Bertram, J.E.A., and Biewener, A.A. 1990. Differential scaling ofthe long bones in the terrestrial Carnivora and other mammals.Journal of Morphology,204: 157–169.
Bertram, J.E.A., and Biewener, A.A. 1992. Allometry and curvaturein the long bones of quadrupedal mammals. Journal of Zoology(London),226: 455–467.
Biewener, A.A. 1982. Bone strength in small mammals and bipedalbirds: do safety factors change with body size? Journal of Ex-perimental Biology,98: 289–301.
Biewener, A.A. 1983. Allometry of quadrupedal locomotion: thescaling of duty factor, bone curvature and limb orientation tobody size. Journal of Experimental Biology,105: 147–171.
Biewener, A.A. 1989. Scaling body support in mammals: limb postureand muscle mechanics. Science (Washington, D.C.),245: 45–48.
Biewener, A.A. 1990. Biomechanics of mammalian terrestriallocomotion. Science (Washington, D.C.),250: 1097–1103.
Biewener, A.A., and Bertram, J.A. 1993. Skeletal strain patterns inrelation to exercise training during growth. Journal of Experi-mental Biology,185: 51–69.
Biewener, A.A., and Bertram, J.A. 1994. Structural response ofgrowing bone to exercise and disuse. Journal of Applied Physi-ology, 76: 946–955.
Biewener, A.A., and Taylor, C.R. 1986. Bone strain: a determinantof gait and speed? Journal of Experimental Biology,123: 383–400.
Biewener, A.A., Thomason, J.J., Goodship, A.E., and Lanyon, L.E.1983. Bone stress in the horse forelimb during locomotion atdifferent gaits: a comparison of two experimental methods. Journalof Biomechanics,16: 565–576.
Blob, R.W., and Biewener, A.A. 1999.In vivo locomotor strain inthe hindlimb bones ofAlligator mississippiensisand Iguanaiguana: implications for the evolution of limb bone safety factorand non-sprawling limb posture. Journal of Experimental Biology,202: 1023–1046.
Bookstein, F.L. 1989. “Size and shape”: a comment on semantics.Systematic Zoology,38: 173–180.
Bookstein, F.L. 1990. Introduction to methods for landmark data.In Proceedings of the Michigan Morphometrics Workshop.Edited by F.J. Rohlf and F.L. Bookstein. The University ofMichigan, Ann Arbor, Mich., pp. 215–225.
Bookstein, F., Chernoff, B., Elder, R., Humphries, J., and Smith, G.1985. Morphometrics in evolutionary biology. Academy ofNatural Sciences, Philadelphia, Special Publication 15.
Brear, K., Currey, J.D., and Pond, C.M. 1990. Ontogenetic changesin the mechanical properties of the femur of the polar bearUrsus martimus. Journal of Zoology (London),222: 49–58.
Brown, B. 1908. The trachodon group. The American MuseumJournal,8: 51–56.
Brown, B. 1912. The osteology of the manus in the familyTrachodontidae. Bulletin of the American Museum of NaturalHistory, 31: 105–108.
Bryant, H.N., and Seymour, K.L. 1990. Observations and com-ments on the reliability of muscle reconstruction in fossil verte-brates. Journal of Morphology,206: 109–117.
Carpenter, K. 1994. BabyDryosaurus from the Upper JurassicMorrison Formation of Dinosaur National Monument.In Dinosaureggs and babies.Edited byK. Carpenter, K.F. Hirsch, and J.R.Horner. Cambridge University Press, New York, pp. 288–297.
Carpenter, K., and Alf, K. 1994. Global distribution of dinosaureggs, nests, and babies.In Dinosaur eggs and babies.Edited byK. Carpenter, K.F. Hirsch, and J.R. Horner. Cambridge UniversityPress, New York, pp. 15–30.
Carrier, D., and Leon, L.R. 1990. Skeletal strength and function inthe California gull (Larus californicus). Journal of Zoology(London),222: 375–389.
Christian, A., and Preuschoft, H. 1996. Deducing the body postureof extinct large vertebrates from the shape of the vertebral column.Palaeontology,39: 801–812.
Cole, M. 1992. Postcranial scaling in the extant and fossilCeratomorpha. Journal of Vertebrate Paleontology,12(Suppl.): 25A.
Cope, E.D. 1883. On the characters in the skull of theHadrosauridae. Proceedings of the Academy of the Natural Sci-ences of Philadelphia,35: 97–107.
Currey, J. 1984. The mechanical adaptations of bones. PrincetonUniversity Press, Princeton, N.J.
Currey, J.D., and Pond, C.M. 1989. Mechanical properties of veryyoung bone in the axis deer (Axis axis) and humans. Journal ofZoology (London),218: 59–67.
Currie, P.J. 1995. Ornithopod trackways from the Lower Cretaceousof Canada.In Vertebrate fossils and the evolution of scientificconcepts.Edited byW.A.S. Sarjeant. Gordon and Breach Publishers,Singapore, pp. 431–443.
Currie, P.J., Nadon, G.C, and Lockley, M.G. 1991. Dinosaur foot-prints with skin impressions from the Cretaceous of Alberta andColorado. Canadian Journal of Earth Sciences,28: 102–115.
Dal Sasso, C. and Signore, M. 1998. Exceptional soft-tissue preser-vation in a theropod dinosaur from Italy. Nature,392: 383–387.
Dilkes, D.W. 1993. Growth and locomotion in the hadrosauriandinosaurMaiasaura peeblesorumfrom the Upper Cretaceous ofMontana. Ph.D. thesis, The University of Toronto, Toronto.
Dilkes, D.W. 2000. Appendicular myology of the hadrosauriandinosaur Maiasaura peeblesorumfrom the Late Cretaceous(Campanian) of Montana. Transactions of the Royal Society ofEdinburgh: Earth Sciences,90: 87–125.
Diniz-Filho, J.A.F., Zuben, C.J., Fowler, H.G., Sclindwein, M.N.,and Bueno, O.C. 1994. Multivariate morphometrics and allometryin a polymorphic ant. Insectes Sociaux,41: 153–163.
Dodson, P. 1971. Sedimentology and taphonomy of the OldmanFormation (Campanian), Dinosaur Provincial Park, Alberta(Canada). Palaeogeography, Palaeoclimatology, Palaeoecology,10: 21–74.
Dodson, P. 1980. Comparative osteology of the American ornithopodsCamptosaurusand Tenontosaurus. Memoires de la sociétégéologique de France, ns,139: 81–85.
Economos, A.C. 1983. Elastic and/or geometric similarity in mam-malian design? Journal of Theoretical Biology,103: 167–172.
Efron, B. 1982. The jackknife, the bootstrap and other resamplingplans. SIAM (Society for Industrial and Applied Mathematics)Monograph 38.
Forwood, M.R., Owan, I., Takano, Y., and Turner, C.H. 1996.Increased bone formation in rat tibiae after a single short periodof dynamic loading in vivo. American Journal of Physiology,270: E419–E423.
Galton, P.M. 1970. The posture of hadrosaurian dinosaurs. Journalof Paleontology,44: 464–473.
Galton, P.M. 1981.Dryosaurus, a hypsilophodontid dinosaur fromthe Upper Jurassic of North America and Africa. Postcranialskeleton. Paläontologische Zeitschrift,55: 271–312.
Galton, P.M. 1990. Basal sauropodomorpha–prosauropoda.In Thedinosauria.Edited by D.B. Weishampel, P. Dodson, and H.Osmólska. University of California Press, Berkeley, Calif.,pp. 320–344.
Gatesy, S.M., and Biewener, A.A. 1991. Bipedal locomotion:effects of speed, size and limb posture in birds and humans.Journal of Zoology (London),224: 127–147.
Gibson, A.R., Baker, A.J., and Moeed, A. 1984. Morphometricvariation in introduced populations of the common myna(Acridotheres tristis): an application of the jackknife to principlecomponents analysis. Systematic Zoology,33: 408–421.
© 2001 NRC Canada
Dilkes 1225
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:45 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
1226 Can. J. Earth Sci. Vol. 38, 2001
Heerden, J., and Galton, P.M. 1997. The affinities ofMelanorosaurus— a Late Triassic prosauropod dinosaur fromSouth Africa. Neues Jahrbuch für Geologie und Paläontologie,Monatshefte,1997: 39–55.
Heinrich, R.E., Ruff, C.B., and Weishampel, D.B. 1993. Femoralontogeny and locomotor biomechanics ofDryosauruslittowvorbecki(Dinosauria, Iguanodontia). Zoological Journal ofthe Linnean Society,108: 179–196.
Horner, J.R. 1982. Evidence of colonial nesting and ‘site fidelity’among ornithischian dinosaurs. Nature (London),297: 675–676.
Horner, J.R. 1994. Comparative taphonomy of some dinosaur andextant bird colonial nesting grounds.In Dinosaur eggs andbabies.Edited byK. Carpenter, K.F. Hirsch, and J.R. Horner.Cambridge University Press, New York, pp. 116–123.
Horner, J.R., and Makela, R. 1979. Nest of juveniles providesevidence of family structure among dinosaurs. Nature (London),282: 296–298.
Horner, J.R., de Ricqlès, and Padian, K. 2000. Long bone histologyof the hadrosaurid dinosaurMaiasaure pebblesorum: growth dy-namics and physiology based on an ontogenetic series of skele-tal elements. Journal of Vertebrate Paleontology,20: 115–129.
Huxley, J.S. 1932. Problems of relative growth. Methuen, London,U.K.
Jenkins, F.A., Jr., and Goslow, G.E., Jr. 1983. The functional anatomyof the shoulder of the savannah monitor lizard (Varanusexanthematicus). Journal of Morphology,175: 195–216.
Jolicoeur, P. 1963. The multivariate generalization of the allometryequation. Biometrics,18: 497–499.
Keller, T.S., and Spengler, D.M. 1989. Regulation of bone stressand strain in the immature and mature rat femur. Journal ofBiomechanics,22: 1115–1127.
Langston, W.L., Jr. 1960. A hadrosaurian ichnite. NationalMuseum of Canada, Natural History Papers, No. 4, pp. 1–9.
Lanyon, L.E. 1992. Control of bone architecture by functional loadbearing. Journal of Bone and Mineral Research,7: S369–S375.
Lanyon, L.E., and Rubin, C.T. 1984. Static vs. dynamic loads as aninfluence on bone remodelling. Journal of Biomechanics,17: 897–905.
Leamy, L., and Bradley, D. 1982. Static and growth allometry ofmorphometric traits in randombred house mice. Evolution,36: 1200–1212.
Lehman, T.M. 1990. The ceratopsian subfamily Chasmosaurinae:sexual dimorphism and systematics.In Dinosaur systematics:perspectives and approaches.Edited by K. Carpenter and P.J.Currie. Cambridge University Press, Cambridge, pp. 211–229.
Leidy, J. 1858. Remarks concerningHadrosaurus. Academy ofNatural Science of Philadelphia, Proceedings,10: 215–218.
Lull, R.S., and Wright, N.E. 1942. Hadrosaurian dinosaurs ofNorth America. Geological Society of America, Special Paper40, pp. 1–242.
Mameren, H. van, and Drukker, J. 1979. Attachment and compositionof skeletal muscles in relation to their function. Journal ofBiomechanics,12: 859–867.
Marcus, L. 1990. Traditional morphometrics.In Proceedings of theMichigan Morphometrics Workshop.Edited byF. Rohlf and F.L.Bookstein. University of Michigan, Ann Arbor, Mich., pp. 77–122.
McCardle, B.H. 1988. The structural relationship: regression inbiology. Canadian Journal of Zoology,66: 2329–2339.
McGillivray, W.B. 1985. Size, sexual size dimorphism, and theirmeasurement in Great Horned Owls in Alberta. Canadian Journalof Zoology, 63: 2364–2372.
Moratalla, J.J., Sanz, J.L., Jimenez, S., and Lockley, M.G. 1992. Aquadrupedal ornithopod trackway from the Lower Cretaceous of
La Rioja (Spain): inferences on gait and hand structure. Journalof Vertebrate Paleontology,12: 150–157.
Moratalla, J.J., Sanz, J.L., and Jiménez, S. 1994. Dinosaur tracksfrom the Lower Cretaceous of Regumiel de la Sierra (provinceof Burgos, Spain): inferences on a new quadrupedal ornithopodtrackway. Ichnos,3: 89–97.
Mosley, J.R., March, B.M., Lynch, J., and Lanyon, L.E. 1997.Strain magnitude related changes in whole bone architecture ingrowing rats. Bone,20: 191–198.
Mosteller, F., and Tukey, J.W. 1977. Data analysis and regression:a second course in statistics. Addison-Wesley Publishing Company,Reading, Mass.
Nelms, L.G. 1989. Late Cretaceous dinosaurs from the North Slopeof Alaska. Journal of Vertebrate Paleontology,9(Suppl.): 34A.
Nelms, L.G. 1992. On the osteology of the hadrosaurian manus.Journal of Vertebrate Paleontology,12(Suppl.): 45A.
Norman, D.B. 1980. On the ornithischian dinosaurIguanodonbernissartensisfrom the Lower Cretaceous of Bernissart (Bel-gium). Institut Royal des Sciences Naturelles de Belgique,Memoires,178: 1–103.
Norman, D.B. 1986. On the anatomy ofIguanodon atherfieldensis(Ornithischia: Ornithopoda). Bulletin de l’Institut Royal desSciences Naturelles de Belgique,56: 281–372.
O’Connor, J.A., Lanyon, L.E., and MacFie, H. 1982. The influenceof strain rate on adaptive bone remodelling. Journal ofBiomechanics,15: 767–781.
Osborn, H.F. 1912. Integument of the iguanodont dinosaurTrachodon.American Museum of Natural History Memoirs,1: 33–54.
Ostrom, J.H. 1964. A reconstruction of the paleoecology ofhadrosaurian dinosaurs. American Journal of Science,262: 975–997.
Parks,W.A. 1920. The osteology of the trachodont dinosaurKritosaurus incurvimanus. University of Toronto GeologicalSeries,11: 5–74.
Peres, N.P.R. 1994. The jackknifing of multivariate allometric coef-ficients (Jolicoeur 1963): a case study on allometry andmorphometric variation inCorydoras barbatus(Quoy andGaimand, 1824) (Siluriformes, Callichthyes). Arquivos deBiologia e Tecnologia (Curitiba),37: 449–454.
Pérez-Lorente, F., Cuenca-Bescós, G., Aurell, M., Canudo, J.I.,Soria, A.R., and Ruiz-Omeñaca, J.I. 1997. Las Cerradicastracksite (Berriasian, Galve, Spain): growing evidence forquadrupedal ornithopods. Ichnos,5: 109–120.
Prothero, D.R., and Sereno, P.C. 1982. Allometry andpaleoecology of medial Miocene dwarf rhinoceroses from theTexas Gulf Coastal Plain. Paleobiology,8: 16–30.
Rayner, J.M.V. 1985. Linear relations in biomechanics: the statistics ofscaling functions. Journal of Zoology (London),206: 415–439.
Rensberger, J. 1988. Archival recording of spatial relationships offossils using photogrammetric methods. Journal of VertebratePaleontology,8(Suppl.): 24A.
Rice, W.R. 1989. Analyzing tables of statistical tests. Evolution,43: 223–224.
Roark, R.J., and Young, W.C. 1975. Formulas for stress and strain.5th ed. McGraw-Hill, New York.
Rogers, R.R., Swisher, C.C., III, and Horner, J.R. 1993.40Ar/39Arage and correlation of the nonmarine Two Medicine Formation(Upper Cretaceous), northwestern Montana, U.S.A. CanadianJournal of Earth Sciences,30: 1066–1075.
Rohlf, F.J. 1990. NTSYS-pc: numerical taxonomy and multivariateanalysis system. Version 1.60. Exeter Publishing Ltd., Setauket,N.Y.
Rubin, C.T., and Lanyon, L.E. 1982. Limb mechanics as a functionof speed and gait. Journal of Experimental Biology,101: 187–211.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:45 AM
Color profile: DisabledComposite Default screen

© 2001 NRC Canada
Dilkes 1227
Rubin, C.T., and Lanyon, L.E. 1985. Regulation of bone mass by me-chanical strain magnitude. Calcified Tissue Research,37: 411–417.
Russell, D.A. 1970. Tyrannosaurs from the late Cretaceous ofWestern Canada. National Museum of Natural Sciences (Ottawa):Publications in Palaeontology, No. 1.
Sampson, S.D., Ryan, M.J., and Tanke, D.H. 1997. Craniofacialontogeny in centrosaurine dinosaur (Ornithischia: Ceratopsidae):taxonomic and behavioral implications. Zoological Journal ofthe Linnean Society,121: 293–337.
Schmitt, J.G., Horner, J.R., Laws, R.R., and Jackson, F. 1998.Debris-flow deposition of a hadrosaur-bearing bone bed, UpperCretaceous Two Medicine Formation, northwest Montana. Jour-nal of Vertebrate Paleontology,18(Suppl.): 76A.
Shea, B.T. 1985. Bivariate and multivariate growth allometry:statistical and biological considerations. Journal of Zoology(London),206: 367–390.
Smith, D. 1998. A morphometric analysis ofAllosaurus. Journal ofVertebrate Paleontology,18: 126–142.
Sternberg, C.M. 1965. New restoration of hadrosaurian dinosaur.National Museum of Canada, Natural History Paper 30.
Turner, C.H., Forwood, M.R., Rho, J-Y., and Yoshikawa, T. 1994.Mechanical loading thresholds for lamellar andwovenbone for-mation. Journal of Bone and Mineral Research,9: 87–97.
Turner, C.H., Owan, I., and Takano, Y. 1995. Mechanotransductionin bone: role of strain rate. American Journal of Physiology,269: E438–E442.
Varricchio, D.J., and Horner, J.R. 1993. Hadrosaurid andlambeosaurid bone beds from the Upper Cretaceous Two Medi-cine Formation of Montana: taphonomic and biologic implica-tions. Canadian Journal of Earth Sciences,30: 997–1006.
Wainwright, S.A., Briggs, W.D., Currey, J.D., and Gosline, J.M.1976. Mechanical design in organisms. Princeton UniversityPress, Princeton, N.J.
Weishampel, D.B., and Heinrich, R.E. 1992. Systematics ofHypsilophodontidae and basal Iguanodontia (Dinosauria:Ornithopoda). Historical Biology,6: 159–184.
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
0
5
25
75
95
100
J:\cjes\cjes38\cjes-07\E01-003.vpMonday, August 13, 2001 11:48:46 AM
Color profile: DisabledComposite Default screen