discussion - inflibnetshodhganga.inflibnet.ac.in/bitstream/10603/4494/11/11_chapter 5.pdf ·...
TRANSCRIPT

Discussion
129
The assessment of intraspecific variability existing in the germplasm of a
species is of great interest for conservation of genetic resources as well as for
broadening of genetic base of species to be exploited by plant breeders. The aim of
the present study was to find out whether phenotypic variations in the plants collected
from different locations in North India were merely epigenetic or genetic. Further, if
the morphovariants have any bearing on their medicinally active principles. So a set
of 34 accessions of E. alba belonging to three different morphotypes were assembled
from different locations in North India. Presently, all the 34 accessions of E. alba
were maintained at Botanic Gardens, Punjabi University, Patiala. Investigations were
carried out to characterize and evaluate the collected germplasm of E. alba for
morphological, cytological, biochemical and molecular parameters.
5.1 . MORPHOLOGICAL ANALYSIS
Morphology has been used to distinguish subspecies and varieties.
Morphological variation and geographical isolation among populations are also
prerequisite to the formation of subspecies and species (Losos and Glor, 2003). The
survey of wild plants revealed the existance of intraspecific morphological variations.
Intraspecific morphological variations are considered as one of the important factor in
the process of evolution. Intraspecific morphovariants within a species have been
considered the incipient species at various stages of development of new species
(Fosberg, 1942). Intraspecific variations of morphological characters are quite
common in flowering plants. The morphology of Eclipta genotypes varied, as has
been previously reported by Gupta (1977). The variability can be attributed to some
genetic reasons as the variants within a species posess same chromosome number.
Morphological variations have been reported in medicinal plants like Lasiurus
scindicus (Yadav and Krishna, 1986; Arshad et al., 2009; Nisar et al., 2010),
Hedysarum boreale (Johnson et al., 1989), Eragrostis species (Tefera et al., 1992),
Cymbopogon jawarancusa (Arshad et al., 1995), Sporobolus iocladus (Arshad et al.,
1999), Dicanthium annulatum (Agarwal et al., 1999), Cenchrus ciliaris (Mansoor et

Discussion
130
al., 2002; Arshad et al., 2007), Zizania palustris var. palustris (Lu et al., 2005) and
Panicum antidotale (Sohail et al., 2010).
Taxonomically there exist many synonyms of Eclipta alba as Eclipta
prostrata (L.) L., Eclipta erecta L., Eclipta puncatata L., Verbesina alba L.,
Verbesina prostrata L. In various Floras, Eclipta alba (L.) Hassk., and Eclipta erecta
L., are accepted as synonyms and Eclipta prostrata (L.) L. is considered as a distinct
species. Boissier (1875) described Eclipta marginata Hochst. and Steud. ex Boiss.,
characterizing its cypselas winged differing from Eclipta alba (L.) Hassk., which has
wingless cypselas. He cited Eclipta prostrata under the synonymy of Eclipta
marginata and Eclipta alba. But Vassilczenko (1959) has considered Eclipta alba and
Eclipta prostrata as two distinct species. He kept Eclipta marginata under the
synonymy of Eclipta prostrata and Eclipta erecta under Eclipta alba. He
characterized Eclipta prostrata by having cypselas smooth and fringed and Eclipta
alba with cypselas tuberculate along the ribs or over whole surface. Rechinger (1977)
has considered Eclipta prostrata and Eclipta alba as two valid species but with some
modification in characters different from Vassilczenko. He characterizes Eclipta
prostrata by having cypselas marginate and tuberculate or smooth and Eclipta alba
with cypselas not marginate and smooth. He placed Eclipta marginata under the
synonymy of Eclipta prostrata as suggeated by Vassilczenko (1959). Abedin and
Tajuddin (2009) examined a large number of specimens and found that Eclipta
prostrata (L.) L., is the only distinct species and the various names mentioned above
are all synonyms. Actually the cypselas of the ray florets are smooth and winged
while of disc florets are tuberculose and winged completely or basally in young
condition. It is these incomplete specimens that have resulted in 2-3 different
species. In the light of Article 57 of the code (International Code of Botanical
Nomenclature, 1978) Eclipta alba (Linn.). Hassk. should be followed. Eclipta alba is
commonly known as safed bhangra (Hindi) when in flower and as Kalobhangro when
in fruit. Pila bhangra is the name given to closely related plant Wedelia chinensis

Discussion
131
(Wealth of India, 1952; Bhargava and Seshadri, 1972). Wedelia chinensis is often
confused with Eclipta alba and thus considered as its adulterant.
Eclipta alba is monospecific, but its evolution under natural selection in a
wide range of agro-climatic areas has resulted in wide phenotypic diversity. Thus it is
evolutionary dynamic genera. It is erect, partly erect or prostrate and is described as
variable species by taxonomists. There is intraspecific variation in this polymorphic
species. It is very variable in habit, hairiness, size, color and shape of leaves and stem
and size of flower head. Among North Indian populations, three morphotypes of
Eclipta alba can easily be identified i.e. erect, semierect and prostrate (Table 5.1). The
erect plants are tall and upright while the prostrate types have all creeping branches.
The semierect type plants have lower parts of branches creeping and tips are
ascending. The prostrate type of accessions show highest incidence (Fig.VI).
Table 5.1: Habit variation among different accessions of Eclipta alba germplasm
Morphotype Accessions
Erect EPBP1, EPBG1, EPBG2, EPBL1, EPBJ1, EPBF1, EJKJ1
Semierect SPBP1, SPBG1, SPBG2, SPBG3, SPBA1, SPBT1, SPBT2, SHRA1,SUPM1
Prostrate PPBP1, PPBG1, PPBG2, PPBA1, PPBA2, PPBA3, PPBA4, PPBJ1,PPBN1, PPBR1, PPBM1, PRJS1, PJKJ1, PUTC1, PHRP1, PHRP2,PHPS1, PUPM1
Fig. VI: Frequency distribution of three morphotypes on the basis of plant habit
Discussion
132
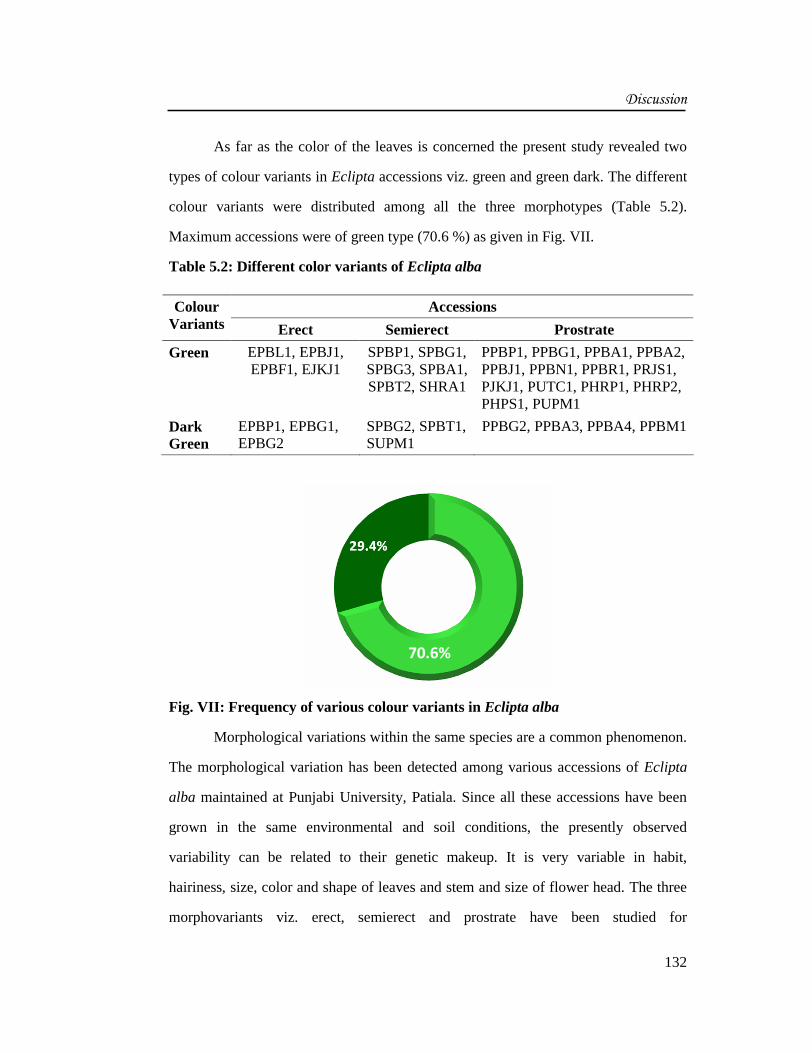
As far as the color of the leaves is concerned the present study revealed two
types of colour variants in Eclipta accessions viz. green and green dark. The different
colour variants were distributed among all the three morphotypes (Table 5.2).
Maximum accessions were of green type (70.6 %) as given in Fig. VII.
Table 5.2: Different color variants of Eclipta alba
AccessionsColourVariants Erect Semierect Prostrate
Green EPBL1, EPBJ1,EPBF1, EJKJ1
SPBP1, SPBG1,SPBG3, SPBA1,SPBT2, SHRA1
PPBP1, PPBG1, PPBA1, PPBA2,PPBJ1, PPBN1, PPBR1, PRJS1,PJKJ1, PUTC1, PHRP1, PHRP2,PHPS1, PUPM1
DarkGreen
EPBP1, EPBG1,EPBG2
SPBG2, SPBT1,SUPM1
PPBG2, PPBA3, PPBA4, PPBM1
Fig. VII: Frequency of various colour variants in Eclipta alba
Morphological variations within the same species are a common phenomenon.
The morphological variation has been detected among various accessions of Eclipta
alba maintained at Punjabi University, Patiala. Since all these accessions have been
grown in the same environmental and soil conditions, the presently observed
variability can be related to their genetic makeup. It is very variable in habit,
hairiness, size, color and shape of leaves and stem and size of flower head. The three
morphovariants viz. erect, semierect and prostrate have been studied for
70.6%
Discussion
133
height/spread, leaf length, number of nodes, branches and inflorescences diameter
(Table 5.3). Among the three types of morphotypes, semierect morphotypes have
highest values of morphological characters as compared to erect morphotypes and the
prostrate morphotypes are the smallest.
Table 5.3: Mean values of various morphological characters of Eclipta alba
Erect type Semierect type Prostrate typeCharacters
Range Mean Range Mean Range Mean
Plant height/Spread (cm)
38.2 – 54.7 45.4 32.6 – 56.8 46.2 21.5 – 38.9 29.3
Nodes/plant 9 -15 11.2 9 - 16 12.4 9 - 13 10.9
Branches/plant 5 -9 7.0 6 - 13 9.4 8 - 11 8.8
Leaf length (cm) 4.2 - 5.8 4.9 3.2 - 5.3 4.5 1.8 - 3.8 2.6
Inflorescence(mm)
5 - 7 5.7 4 - 7 5.4 3 - 5 4.0
5.2 REPRODUCTIVE STUDIES
Studies in the field of reproductive biology can provide important paradigms
for biodiversity conservation, reclamation and restoration. Such studies in
reproductive biology will also help in developing strategies to preserve the genetic
potential of rare species and are crucial for restoration and reintroduction. The
phenological studies are useful in planning out for cultivation of the species on large
scale (Schemske et al., 1994; Delanoe et al., 1996; Wafai et al., 1996; Bernardello, et
al., 2001). There is always a buzz all over the globe to unravel the basic and detailed
information about the reproductive biology of important plant species preferably for
their genetic improvement and cultivation. Phenology in general and reproductive
phenology in particular is a critical and important trait of a plant because it determines
the growth, developmental pattern and number of potential mates thus providing a
mechanism for reproductive isolation and speciation over time (Rathcke, 1983;
Bronstein et al., 1990).

Discussion
134
The present work summarizes the salient features of the reproductive
strategies of the herb Eclipta alba and this information may be of immense use in
planning its cultivation and long term conservation. The genus has considerable
phenotypic plasticity and has morphological variations viz. erect, semierect and
prostrate. Many aspects of reproductive behavior of plants are plastic and can vary
greatly among individuals of the same species living in different habitats. As all the
morphotypes of Eclipta alba are grown under identical conditions. Considerable
diversity was observed among the three morphotypes which includes number of ray
and disc florets, number of seeds produced per capitulum, pollen fertility and seed
germination. Taylor (1987) stated that intra-population morphological variation was
as great as or greater than inter-population variation and therefore, the morphological
variation was largely due to phenotypic plasticity. The phenotypic variability in E.
alba increases its ability to colonize a wide range of habitats. In cool or dry weather,
or in closely mown lawns, the leaves usually spread flat against the surface of the
ground to form an almost prostrate rosette (Longyear, 1918; Lovell and Rowan,
1991). In warmer weather or in areas where it is crowded by taller vegetation, the
leaves stand in more or less erect tufts (Longyear, 1918). The rosette enables it to
survive mowing, grazing and competition with grasses. Also its ability to reproduce
by sexual and vegetative means accounts for the species colonizing and stabilizing
fast in new habitats. This probably accounts for the rapid spread of Eclipta alba.
Vegetative reproduction allows it to occupy a temporary site quickly while light seeds
produced by sexual reproduction allow distance dispersal to new sites.
5.2.1 Floral Morphology
Studies on biology of the species revealed that the taxon is hermaphrodite. The
Eclipta alba is propagated by seeds and rarely through vegetative cuttings. E. alba is
annual to biennial plant that flowers throughout year. The flowers are showy and are
borne in aggregates of capitular heads for pollinator attraction. Several workers

Discussion
135
contended that the heads of composites represent single flowers (Good, 1931; Leppik,
1960). However, this is true only in terms of pollinator attraction. Presentation of
many single ovuled flowers for pollination over a great period of time results in much
higher time results in much higher rates of out crossing than in many ovuled single
flowers (Brutt, 1961). Onset and duration of flowering, relative maturation of male
and female sex organs and the number and arrangement of flowers in a plant
profoundly influence the pollinator visitation pattern in the taxa under reference
which has a direct bearing on the success of their sexual cycle (Siddique, 1991).
Different reproductive characters of Eclipta alba were quantitatively assessed.
The total number of ray and disc florets in a flower head found to be variable for the
three accessions i.e. in erect (81.34) followed by semierect (79.66) and least in
prostrate (66.67) type. Occasional occurrence of twin capitulum has been observed in
some accessions is also observed in other members of Asteraceae. Ray florets were
larger than disc florets. However, their ovary size and bifid stigma length were almost
of equal size. A very thick mat of dense hairs was present on both upper and lower
surfaces of stigma in disc floret while dense hairs were present only on the upper
stigma surface in ray floret. In the disc floret stigma protruded through the
syngenesious column of anther. Protandry was the common phenomenon. The
characteristics of reproductive apparatus influenced the reproductive fitness. The ray
florets open before the disc florets to enhance the chances of cross pollination. The
flower opening is asynchronous which ensures pollen availability for a longer time to
affect pollination and possibilities of higher fruit and seed set. In Asteraceae the
florets are protandrous when hermaphrodite, and among the species in which the level
and type of self-incompatibility has been measured, the vast majority are
sporophytically self-incompatible (Nettancourt, 1977; Patil et al., 1979; Stuessy et al.,
1986). Consequently vector mediated crossing is extremely important.

Discussion
136
5.2.2 Flowering Phenology and Anthesis
Phenological studies play an important role in planning of conservational
strategies and finalizing the protocol for large scale cultivation of plants (Schemske et
al., 1994; Delano et al., 1996; Wafai et al., 1996; Bernardello et al., 2001; Wani et al.,
2006).
According to Reich (1995), the phenology of vegetative phase is important as
cycles of leaf flush and leaf fall are intimately related to growth, plant water retention
and gas exchange. Floral initiation, development, and anthesis of Asteraceae are
generally described as occurring in an acropetal or centripetal direction on the
capitulum (Cronquist, 1955; Sattler, 1973; Reese and Hilger, 1984). In E. alba,
anthesis proceeds gradually towards the centre of a head. Ray florets present in the
periphery opens as the capitulum start opening. The anthesis starts at morning and
florets remained open till the stigma shrivels. The outer ray florets opens first so as to
enhance the chances of cross pollination from pollens of disc florets of other flowers
than the pollens from disc florets of the same capitulum. Presently, it has been
observed that anthesis in E. alba is asynchronous from floret to floret as well as
capitulum to capitulum. The protracted asynchronous mode of pollen presentation by
the species guarantees the long term availability of the pollen to ensure effective
pollination (Wyatt, 1982). The present study confirms the earlier reports of
asynchronous anthesis in Inula racemosa (Wani et al., 2006)
5.2.3 Pollen Fertility
Pollen represents a critical stage in the life cycle of plants as viable pollen is
crucial for successful plant reproduction. Plant fertility can influence many aspects of
population ecology and evolution. The quantity and quality of pollen produced by a
plant is an important component of reproductive success.
Pollen analysis revealed that Eclipta alba has high pollen fertility. Pollen
fertility was studied throughout the year from January to December to study the effect

Discussion
137
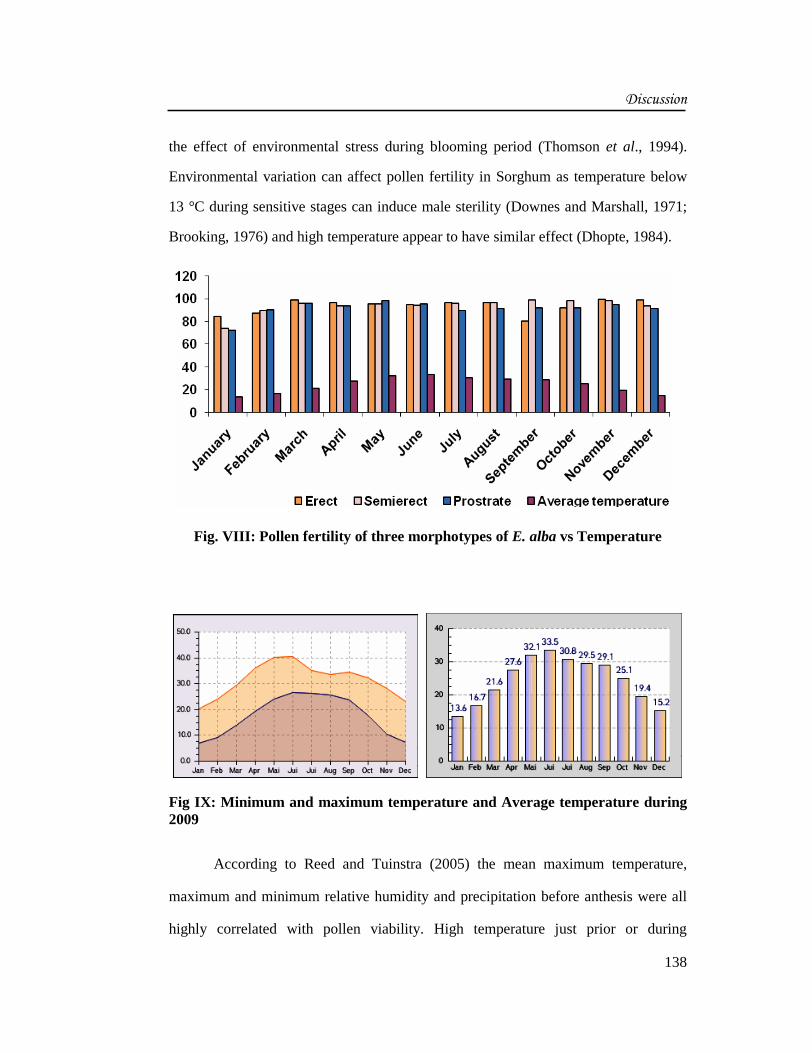
of seasonal variations. The erect morphotypes have maximum percentage of viable
pollens than the semierect and the prostrate morphotypes (Fig. VIII). Pollen fertility
was highest in the month of November in the erect morphotypes (99.34%) In the
semierect plants, it was highest in the month of September (98.41%) and in prostrate
it is highest in the month of May (97.80%). The Eclipta plants did not show any heat
stress as far as pollen fertility was concerned. In erect morphotypes, pollen fertility is
lowest in the month of September (80.04%). In semierect and prostrate morphotypes,
the pollen fertility was lowest in the month of January i.e. 74.04% and 71.96%,
respectively. The pollen is spheroidal, tricolporate and echinate. Polar diameter was
22.45 μm (20-25 μm) and equatorial diameter was 22.5 μm (20 - 25 μm). P/E ratio is
0.99 μm. The length of spine was 2.5 μm (1.5- 4 μm). Exine thickness was 7.4 μm (1 -
2 μm (Zafar et al., 2007).
Pollen fertility depends on the efficiency of the meiotic process. In the
presently analyzed accessions meiotic abnormalities have been found to be
responsible for pollen sterility which could have an important effect on fitness of the
population. However, if meiosis was regular, for example, chromosomes pair and
segregate normally, sterility of the pollen grain was not expected to occur because of
cytological reasons (Boff and Schifino-Wittmann, 2002). There are many non-genetic
causes of decline in pollen viability, including pollen age and exposure to
environmental stresses such as temperature and humidity (Stone et al., 1995; Kelly et
al., 2002). Variations in temperature, humidity, and cloud cover can influence pollen
production and viability (Artschwager and McGuire, 1949; Brooking, 1979; McLaren
and Wehner, 1992). Cold temperature stress prior to flowering appears to reduce
pollen viability during anthesis by disrupting meiosis during early stages of
microsporogenesis (Brooking, 1979). Environmental factors like high temperature,
drought conditions and soil factors may seriously affect pollen performance (Zamir
and Gaddish, 1987). Assessment of pollen viability is very important in understanding
Discussion
138
the effect of environmental stress during blooming period (Thomson et al., 1994).
Environmental variation can affect pollen fertility in Sorghum as temperature below
13 °C during sensitive stages can induce male sterility (Downes and Marshall, 1971;
Brooking, 1976) and high temperature appear to have similar effect (Dhopte, 1984).
Fig. VIII: Pollen fertility of three morphotypes of E. alba vs Temperature
Fig IX: Minimum and maximum temperature and Average temperature during2009
According to Reed and Tuinstra (2005) the mean maximum temperature,
maximum and minimum relative humidity and precipitation before anthesis were all
highly correlated with pollen viability. High temperature just prior or during

Discussion
139
microsporogenesis significantly reduced the pollen fertility. According to Sato et al.
(2002), the most sensitive period for heat stress was found to be 7-15 days before
anthesis. Presently, the pollen fertility has been studied in various morphotypes of E.
alba to see the seasonal variation and it was found that average temperature has no
correlation with pollen fertility (Table 5.4). However, there was positive and
significant correlation between maximum temperature and pollen fertility in the three
morphotypes of Eclipta. The minimum, maximum and average temperature
throughout the year is shown in Fig. IX.
Table 5.4: Correlation coefficient between Pollen fertility in E. alba and Averagemonthly temperature during 2009
MorphotypePollen fertility vs
Avg.temperature
Pollen fertility vsAvg. min.
temperature
Pollen fertility vs Avg.max. temperature
Erect 0.117454 0.555268** 0.540597**
Semierect 0.16533 0.602594** 0.511173**
Prostrate 0.178289 0.178289 0.624462**
** Significant at 0.01
5.2.4 Pollination system
In Asteraceae the basic attraction unit for pollinators is the inflorescence
(capitulum) composed of ray florets, which are petal analogues (Mani and Saravanan,
1999). The presence of ray florets is positively associated with plant fitness when
pollinators are abundant (Stuessy et al., 1986, Nielsen et al., 2002). Flowers
essentially act as a food source for pollinators and other visitors thereby giving the
study of flowering phenology both ecological and evolutionary significance.
Purseglove (1968) postulated that flowers with white corolla, strong perfume,

Discussion
140
abundant nectar and sticky pollen are entomophilous and are visited by bees and
thrips. However, our study showed that white flowers of E. alba were not visited by
many insects except thrips and ants and these may carry out the pollination suggesting
entomophilous pollination. Small Thrips tabaci insects were often seen visiting all
florets of a head one by one going down to the corolla tube and comes out to visit the
next one. Earlier studies have shown that Thrips tabaci and Thrips palmi of order
Thysanoptera are hosts of Eclipta (Salas, 2003).
5.2.5. Seed production and Fertility
The total number of seeds produced per capitulum in the three morphotypes of
Patiala populations of E. alba was also variable i.e. erect morphotypes (83±6.55)
followed by semierect (72±7.54) and least in prostrate (59±3.0). Bagging experiment
revealed that E. alba is self as well as cross compatible which is evident from high
seed set in open pollinated flowers and low seed set in bagged flowers. The seed
production was enhanced by insect pollination. E. alba showed protandrous condition
which favors outbreeding. The species was cross compatible. Open pollinated heads
produced large quantity of seeds while those bagged to exclude the foreign pollen
produced less number of seeds. Self and cross pollination has been recorded in several
species of Compositae. Self pollen can germinate as revealed by the bagging
experiments. Low fruit and seed set in bagged flowers may be due to the existence of
protandry, which tends to prevent self pollination and encourages cross pollination.
There is seasonal variation on seed germination in E. alba as observed in the months
of January and July. It was observed that in the month of January seed germination
was very low in all the three morphotypes and was very high during July in all the
three morphotypes.

Discussion
141
5.3 CYTOLOGICAL STUDIES
The data on chromosome number and meiotic analysis are prerequisites to
overall understanding of a genome and its genetics. Presently detailed cytological
studies have been carried out in different accessions of Eclipta alba (L.) Hassk. All
these accessions have same chromosome number n=11 and is in accordance with
earlier records for the species (Keil and Stuessy 1975; Koul et al., 1976a; Pinkava and
keil, 1977; Powell and Powell, 1978; Sidhu, 1979; Robinson et al., 1981; Morton,
1981; Kumar and Trivedi, 1983; Trivedi and Rajesh, 1984; Razaq et al., 1988; Gill
and Omoigui, 1988; Mathew and Mathew, 1988; Gupta and Gill, 1989; Trivedi and
Trivedi, 1992; Hunziker et al., 1989; Mohamed, 1997). Most of the previous reports
from India and outside also reported the same chromosome number. However Kumar
et al. (1990), Singh (1993) and Kumar and Singh (1989) reported aneuploid cytotypes
(2n=20,22) in the species. Mohan et al. (1962) found somatic variability i.e. 2n =18;
Silvestre (1980) investigated n=11 and 2n=18 and Renard et al. (1983) found 2n=18
for E. alba. Husaini and Iwo (1990) reported its haploid number n=12.
Meiotic studies carried out in various accessions of Eclipta alba revealed that
the majority of the collections showed normal meiotic behavior with 11 bivalents at
metaphase-I and regular chromosome segregation at anaphase-I and II. At diakinesis
one or two bivalents were seen to be associated with darkly stained nucleolus. Ring
and rod bivalents were observed at diakinesis, ring shaped were more frequent than
rod shaped bivalents.
In the life cycle of an organism the meiosis is genetically the most significant
activity consisting of highly coordinated physiological, biochemical, cytogenetic and
phenotypic events which lead to gene recombination, chromosome reduction and
gamete formation (Kaul and Singh, 1991). Mutation in any of the genes governing the

Discussion
142
events disrupts meiosis and culminates in gametic sterility and other abnormalities.
Basic data on meiotic behavior and pollen viability estimations are important for the
germplasm characterization, to determine the genetic variability and evolution
processes of the species analyzed.
The observed precocious chromosome migration to the poles or early
disjunction in accession PHPS1 may have resulted from precocious chiasma
terminalization at diakinesis or metaphase-I. Late disjunction of chromosomes as
observed in accession PHPS1 was generally attributed to the interlocking of
chiasmata. The presence of bridges with or without fragment reflected structural
changes such as heterozygous inversions, chromatid exchanges, deletions and
duplications. The observed subchromatic or side arm bridges could be originated from
an abnormal function at the places where the chiasmata are formed (Brandham, 1969;
Brandham, 1970).
Abnormalities in microsporogenesis become evident by occurrence of dyads,
triads, polyads, tetrads with micronuclei, etc. Dyads, triads with three microspores of
different sizes suggest that restitution nuclei may also occur. Dyads were formed
when there was failure of one meiotic division. Formation of triads could be due to
asynchrony in the second division where one nucleus fails to divide while the other
enters telophase-II. Usually in a triad one 2n microspore and two n microspores are
formed. The consequences of 2n are jumbo pollen formation.
The micronuclei at dyad or tetrad stage of PMC’s might have resulted due to
non-orientation of chromosomes and laggards. Micronuclei lead to the formation of
micro pollen which indicates the loss of genetic material. Their presence, therefore,
suggests that the resultant product of meiotic division is deficient in one or the other
chromosome. This usually leads to the formation of sterile pollen grains. According to

Discussion
143
Singhal et al. (2010) the large sized pollen grains, almost double size, may be of ‘2n’
constitution and play an important role in the origin of intraspecific polyploidy in the
species. The small sized fertile pollen grains which are the products of hypoploid
PMCs could lead to the origin of aneuploids. Presently, different sized pollen grains
have been observed in different accessions E. alba but the cytological status could not
be ascertained. This aberrant meiotic behavior has an influence on the low viability of
pollen. The presence of laggards, bridges, unoriented chromosomes during anaphase-
I/telophase-II and consequently micronuclei and microcytes and polyads during
meiosis was reported in many species such as Capsicum annuum L. (Raja Rao et al.,
1987), Vicia faba and Pisum sativum (Souguir et al., 2008) and Brachiaria hybrid
(Mendes-Bonato et al., 2006) etc.
Chiasmata Frequency Analysis
Estimation of chiasma frequency has been regarded as most easy and direct
method of scoring the total number of crossing over events in the genome. Chiasma
corresponds to the points of physical exchange between homologous non-sister
chromatids (Tease and Jones, 1978) and their frequency is regarded as a good
estimate of the level of genetic recombination in the species (Colombo, 1992). Studies
conducted on a variety of plants revealed the existence of intraspecific variability in
chiasma frequency among different accessions (Sanhez-Moran et al., 2002). It is well
known that chiasma frequency varies in different environments (Barber, 1940; Fedak,
1973; Dowrick, 1957). Rees and Dale (1974) have considered the intraspecific
variability as a mean of generating new forms of recombination which influence
variability within natural populations in an adaptive way.
Eclipta alba plants flower throughout the year. The recombination processes
of the three morphotypes of the species have been studied to know if there is any

Discussion
144
fluctuation in the process in different months of the year. Natural variation in
recombination frequency in Eclipta alba has been assessed by analyzing variation in
chiasma frequency among three morphotypes. There was no significant variation in
mean chiasma frequency between plants within accessions, but there was significant
variation between accessions. The different prostrate type of accessions of E. alba
were found to have higher recombination values than the values shown by plants of
other morphotypes except during the months of March and September when the erect
type of plants show higher recombination values (Fig. X). In all the accessions
belonging to the three morphotypes of the species a common trend was observed i.e.
in each morphotype the average chiasma frequency and recombination index were
highest during the months of July and lowest during the months of Janauary.
The chiasma frequencies of large number of plant chromosomes have been
determined in the last centaury (Nilson et al., 1993). Studies conducted on a variety of
plants revealed the existence of intraspecific variability in chiasma frequency among
different accessions (Sanhez-Moran et al., 2002). It is well known that chiasma
frequency varies in different environments (Barber, 1940; Fedak, 1973; Dowrick,
1957).
According to Fedak (1973) chiasma frequency varies in different
environments. The seasonal variation in chiasma frequencies has been reported in a
number of animals (Cobror et al., 1986). There is considerable literature documenting
the influence of temperature (Dowrick, 1957), mineral nutrients (Law, 1963; Bennett
and Rees, 1970), chemicals (Barber, 1940) and ionizing radiations (Lawrence, 1961)
on chiasma formation. Since specific environmental factor such as temperature might
have contributed to seasonal variation in the chiasma frequency in the present case an
attempt has been made to correlate minimum temperature with chiasma frequencies in

Discussion
145
the three morphotypes (Fig. XI). One cannot conclude that the average minimum
temperature during the first week of alternate months is more important than the day
temperature in causing changes in pairing and chiasma formation in chromosomes.
Maximum and minimum temperature is not sufficiently accurate criteria of daily
fluctuation in temperature. Nevertheless, the present study has shown the decrease in
the chiasma frequency with increase in minimum night temperature. In all the
morphotypes of E. alba minimum chiasma frequency was observed during the months
of July when average night temperature was 26.9 ºC. However, Dowrick (1957)
recorded increased chiasma frequencies in Tradescantia bracteata and Uvularia
perfoliata as a result of increase in temperature. Similar observations have been made
by Hovin (1958) in amphihaploid of Poa annua. Genetic as well as environmental
factors have generally been considered the reason for differences in chromosome
pairing.
Variation in chiasma frequency and localization has also been shown to be
genetically controlled (Quick, 1993). The existence of significant interaccession
variation for chiasma frequency implies that the accessions concerned differ for
genetic factors or elements which effects chiasma frequency (Sanchez-Moran et al.,
2002). The presently observed cyclic variation in average chiasma frequencies,
recombination index and excess chiasma in the three morphotypes of Eclipta alba
clearly depict the influence of environmental factors on process of recombination.

Discussion
146
Fig. X: Distribution of Recombination index and the number of ExcessChiasmata per bivalent during different months in various morphotypes ofE. alba
Fig. XI: Relationship of mean Chiasma Frequency and mean minimum nighttemperature

Discussion
147
5.4 MOLECULAR ANALYSIS
An enormous amount of genetic diversity is characteristic of natural
populations. The random genetic drift, restricted gene flow and differential selection
pressures jointly influence genetic structure of natural populations. These effects lead
to low within and comparatively high among population genetic variation in species
consisting of small and isolated populations (Nevo, 1983; Holderegger and Schneller,
1994). DNA based molecular markers are versatile tool for the plant genome analysis
and for differentiating different genotypes. A number of molecular techniques like
Random Amplified Polymorphic DNA (RAPD), Amplified Fragment Length
Polymorphic DNA (AFLP), Restricted Length Polymorphic DNA (RFLP) and Inter
Simple Sequence repeat (ISSR) etc are now being successfully used for genetic
analysis of medicinal plants (Joshi et al., 2004). DNA markers are highly stable and
specific. It has immense applications in the standardization of medicinal plants and its
products. Molecular analysis techniques allow the estimation of genetic variability
and divergence between species and populations, and thus they can be used for
phylogenetic studies, conservation programs and control of the genetic origin of
products.
In general, genetic diversity can be measured at any functional level from
blueprint (DNA) to phenotype. Presently, possible genetic variation among E. alba
genotypes collected from different locations was analyzed using analysis to
discriminate molecular variability. Random Amplified Polymorphic DNA markers
were chosen since they are an effective and relatively inexpensive technique and do
not require any prior sequence information (Jones et al., 1997; Chenuil, 2006). These
dominant neutral markers have been applied to a wide range of plant species like
Azadirachta (Farooqui et al., 1998), Juniperus communis (Adams and Pandey, 2003),
Codonopsis pilsula (Fu et al., 1999), Allium schoenoprasum (Friesean et al., 1999),
etc.

Discussion
148
Presently, RAPD based cluster analysis revealed genetic variability in
different populations of E. alba. Accession PHPS1 collected from district Sirmaur
(Himachal Pradesh) was found to be significantly different from the rest of accessions
displaying a similarity coefficient of 0.59 with rest of the accessions. RAPD based
analysis revealed genetic variability among different accessions belonging to three
morphotypes of E. alba. Closeness among prostrate accessions collected from district
Amritsar (PPBA1 and PPBA2), Patiala and Jalandhar (PPBP1 and PPBJ1) and
Panchkula (PHRP1 and PHRP2) was quite evident. Similarly genetic closeness in
semierect accessions SPBP1 and SPBG3 collected from Patiala and Gurdaspur has
been demonstrated. The association between genotypes from contiguous regions may
be the result of similar agro climatic conditions or due to seed movement and gene
flow, however the closeness between distantly located genotypes may be attributed to
the unique and broad genetic base of the species. Thus the accessions collected from
different localities on molecular analysis revealed high variation within the species
indicating their adaptive potential.
Genetic diversity within a species has always enthused evolutionary and
conservation biologists. The capability of a species to adapt to environmental changes
depends upon genetic diversity in the species (Neel and Ellstrand, 2003). The species
with depletion in gene pool and reduced genetic diversity pose challenges in the
selection pressure brought in by the environmental changes (Caro and Laurenson,
1994). The evolution of plants in distinct agro-climatic zones demonstrates significant
levels of variations in response to the selection pressure in the zones (Singh et al.,
1998).
Although the data presented here are not conclusive to infer the genetic
relationship between the various accessions but they reflect the utility of RAPD in the
analysis of genetic variability distribution within this important medicinal herb. The

Discussion
149
present study and similar studies on lotus, sweet potato, oil palm and potato (Campos
et al., 1994; Connoly et al., 1994; Shah et al., 1994; Demeke et al., 1996) suggest that
RAPD is more appropriate for analysis of genetic variability in closely-related
genotypes. Ray and Bhattacharya (2010) used the RAPD technique for confirmation
of genetic fidelity of synseed generated plantlets of Eclipta alba. Molecular analysis
of intraspecific variation in particular may find application in resolving disputes of
taxonomic identities, relations and authentication of the species, developing a
comprehensive database of genetic variability in the species for future reference.
5.5 BIOCHEMICAL ANALYSIS
The curative activity of plant based drugs is attributed to the chemicals present
in them and majority of which are their secondary metabolites which help the plant to
adapt to biotic and abiotic stress. Most of the Ayurvedic herbs are collected from
nature without bothering about their genetic makeup which is bound to affect their
chemical constitution. The major challenge for conservation and improvement of
genetic resources of medicinal plants is the identification of better chemotypes which
have both good quality and quantity of active principle. Plant secondary metabolites
(PMS) play an important role in the survival of the plants in their habitat and are
involved in resistance against pest and diseases, the attraction of pollinators, the
attraction with symbiotic microorganisms, etc (Dixon, 2001; Harborne, 2001). PMS
are also of interest because they determine the quality of food (color, taste and aroma)
and ornamental plants (flower color, smell).
The chemical composition of the Eclipta alba plants has been investigated by
many workers. The herb Eclipta alba contains mainly coumestans i.e. wedelolactone
and demethylwedelolactone, polypeptides, polyacetylenes, thiophene-derivatives,
steroids, triterpenes and flavonoids. Qualitative analysis of the phytochemicals of
aqueous extracts revealed the presence of carbohydrates, saponins, phytosterols,

Discussion
150
phenols, flavonoids and tannins in E. alba plants (Khanna and Kannabiran, 2007).
Study of chemical constituents of E. alba has been carried out by a number of workers
(Govindachari et al., 1956; Bhargava et al., 1972; Karrer, 1977; Wagner et al., 1986;
Singh et al., 1988; Nguyen and Doan, 1989; Han et al., 1998).
The present study conducted on various accessions of Eclipta alba revealed
the presence of inter-accession variation in various phytochemicals. The data is
presented in Tables 4.10 and 4.11.
5.5.1 Carbohydrates
Anthrone assay (Chitra et al., 2009) revealed that the extract has
0.11µg/100mg and 1.37µg/100mg of carbohydrates on dry basis in root and leaf
sample, respectively. Anitha (2002) reported 232 mg/g of carbohydrates in E. alba.
Among various accessions of E. alba maintained at Patiala the carbohydrate content
ranged from 16.13 mg/g to 18.69 mg/g with a mean value of 17.45±0.701 mg/g. The
semierect plants have overall highest carbohydrate content.
5.5.2 Proteins
The protein content determined by Chitra et al. (2009) by using Lowry’s
method was 1.31µg/100mg and 0.74µg/100mg for leaf and root samples, respectively.
Anitha (2002) reported 0.1 mg/g of proteins in E. alba. Rao et al. (2009) reported 14.5
mg/g total protein content in E. prostrata in wet sample. In the present study, the
protein content varied between 12.21 mg/g to 14.85 mg/g with mean value of
13.69±0.678 mg/g. Protein content was highest among erect plants.
5.5.3 Phenols
These nitrogen-containing compounds protect plants from a variety of
herbivorous animals, and many possess pharmacologically important activity. So far
as plant phenolics constitute one of the major groups of compounds acting as primary
antioxidants or free radical terminators, it was reasonable to determine their total

Discussion
151
amount in the selected plant extracts (Cook and Samman, 1996). A recent study by
Chitra et al. (2009) reported 0.12 µg/100 mg of phenolic compounds in leaf samples
and 0.06 µg/100mg of phenolic compounds in root samples. In the fresh leaves of
Eclipta alba Rao et al. (2009) observed phenol content as 19.5±0.458 GAE units in
fresh E. alba. According to another study the phenol content in the methanolic extract
was 30.4 mg/g of dry mass in Eclipta alba (Veeru et al., 2009). In the presently
studied accessions of Eclipta alba, the phenol content varied between 17.33 mg/g to
20.97 mg/g with a mean value of 18.83±1.088 mg/g. Among three morphotypes, the
minimum phenol content was observed in prostrate accessions.
5.5.4 Saponins
Saponins are amphipathic glycosides grouped phenomenologically by the
soap-like foaming they produce when shaken in aqueous solutions. Saponins, a
diverse group of secondary metabolites characterized by their structure containing a
steroidal or triterpene aglycone and one or more sugar chains (Hostettmann and
Marston, 1995; Osbourn, 2003; Guclu and Mazza, 2007), occur constitutively in
many plants and, everting various biological activities, may be considered as part of
plant defense system (Potter and Kimmerer, 1989; Osbourn, 1996; Agrell et al.,
2003).
Present study has revealed the variation in saponins content in the leaves of
various accessions. Saponins content ranged between 28.3 mg/g to 48.5 mg/g with
overall mean of 39.4±4.89 mg/g. Crude saponins was the major phytochemical
constituent present in highest percentage in E. prostrata (4.5%). Other
phytochemicals estimated in this plant were present only in very low concentrations
(Khanna and Kannabiran, 2007).

Discussion
152
5.5.5 Wedelolactone
Wedelolactone is a naturally occurring coumestan isolated from aerial parts of
Eclipta alba. The E. alba plants contain wedelolactone (1.6%) as principle coumestan
compound due to which it is valued as hepatic drug. The coumestan Wedelolactone is
found to be present in leaf and stem of natural herb but absent in roots.
Chemoprofiling using HPLC have a great role in quality control of medicinal plants
(raw drugs) and in the finished herbal drugs.
The present HPLC estimation revealed the considerable phytochemical
(wedelolactone) variation in the studied accessions of Eclipta alba. The
phytochemical diversity measured as quantitative difference in the accumulated
wedelolactone ranged from 0.111 mg/g to 1.43 mg/g with a mean value of
0.512±0.385. Plant specific marker compound (wedelolactone) showed quantitative
variations among the genotypes, which could not be correlated with allelic variation
but the active principle content when measured under uniform growing conditions,
variations so observed presumably had a genetic basis.
Variation in morphological characters and active principle constituents of
Eclipta prostrata Linn. under different seasonal and soil conditions has been
determined by Gupta (1977). The concenteration of wedelolactone as estimated by
UV spectrophotometric method at 351 nm is 0.0078±0.000081% in leaf and
0.0042±0.000081% in stem (Zafar and Sagar, 1999). HPLC analysis of Eclipta alba
demonstrated the presence of major proportion of wedelolactone in case of petroleum
ether extract (1.9±0.2% w/w) and very small quantity of around 0.2±0.01% w/w in
case of ethanolic extract (Roy et al., 2008). According to Wagner et al. (1986)
extracts of vegetative tissue of E. alba plants cultivated in India produce 15.9%

Discussion
153
wedelolactone. Methanol extracts of E. alba contain 1.6% wedelolactone (Jayathirtha
and Mishra, 2004).
5.5.6 Phytate Content
Elements are necessary for the healthy development of man, animals and
plants. Their concentrations in the plant differ from part to part and from plant to
plant, depending on the elements level in the soil and presence of allelochemicals.
Phytate is one of the allelochemicals found in plants (Aletor, 1993). It is the most
important antinutritional factor because more than 50% of phosphorus is present in
the form of phytate in plant foods and organic soils serving as phosphorus depot in the
body, which is broken down by phytate to myo-inositol. Myo-inositol increased the
oxygen transporting capacity of hemoglobin in red blood cells, improves and
regulates cellular metabolism. Phytate has been shown to play a role in preventing
colorectal carcinoma, hypercholesterolaemia and renal calculi (Marounek et. al.,
2000). The aquatic weed samples were found to be high in phytate. It is likely that
high phytate intake interferes with mineral absorption. Even though some plants have
a substantial quantity of the minerals, it is felt that a large portion of these minerals
may not be available due to the presence of phytate (Abulude, 2005).
In the present study phytate contents in three morphotypes of E. alba has been
compared and it was found that phytate was highest in the erect type i.e. EPBP1
(467.26 mg/100g) followed by semierect i.e. SPBP1 (463.80 mg/100g) and least in
the prostrate type i.e. PPBP1 (449.96 mg/g). Earlier, Abulude (2005) reported 440
mg/100g DM of phytate in E. alba. The present results suggest that the weed is not
suitable as diet as it has high phytate content as the bioavailability of minerals of the
weed would be low.

Discussion
154
5.6 CORRELATION ANALYSIS
The better understanding of genetic variation at the intraspecific level help in
identifying superior genotypes for crop improvement as well as to evolve strategies
for the effective in situ and ex situ conservation programmes, although such empirical
determination of genetic diversity can be obtained by evaluating morphological and
biochemical traits. The aim of the present study was to find out whether phenotypic
variations in the genotypes grown in different locations are merely epigenetic or
genetic. Furthermore, if the same are genetically different, then their medicinally
active principles have to be estimated to find out possible quantitative variation,
which accord for the variation in medicinal activity of the plants collected from
different locations.
The morphological and biochemical characteristics of the different accessions
of E. alba have been studied. The correlation coefficients among the major
characteristics showed that these are positively associated with each other. The
present correlation study revealed that the various phytochemicals in the leaves are
positively correlated with morphological characters such as plant height and leaf size.
The plant height was found positively and very significantly correlated with
biochemicals i.e. carbohydrates (0.96), proteins (0.86), phenols (0.96), saponins (0.93)
and wedelolactone (0.45). The correlation coefficients among different characteristics
are given in Table 5.5. Medicinal plants are valued for the biochemicals present in the
leaves. The results clearly indicate that the selection of Eclipta alba plants with good
yield depends upon large plants with big sized leaves. Thus the data of morphological
characters showed usefulness of selecting the genotypes with good yield for
commercial cultivation.

Discussion
155
Table 5.5: Correlation between morphological and biochemical characters indifferent morphotypes of Eclipta alba
Characters X1 X2 X3 X4 X5 X6 X7
Erect plants
X1 Plant height 1
X2 Leaf length 0.76881 1
X3 Carbohydrate 0.80712 0.65082 1
X4 Protein 0.80712 0.72277 0.40075 1
X5 Phenol 0.78923 0.68420 0.69454 0.60356 1
X6 Saponins 0.22869 -0.10455 -0.50396 0.99685 -0.78999 1
X7 Wedelolactone 0.21001 0.52017 0.82516 -0.86703 0.45152 -0.90376 1
Semierect plants
X1 Plant height 1
X2 Leaf length 0.66263 1
X3 Carbohydrate 0.95396 0.71477 1
X4 Protein 0.92332 0.73002 0.97984 1
X5 Phenol 0.97159 0.67557 0.98527 0.95723 1
X6 Saponins 0.95835 0.63983 0.91870 0.87730 0.96840 1
Prostrate plants
X1 Plant height 1
X2 Leaf length 0.10312 1
X3 Carbohydrate 0.91224 0.00926 1
X4 Protein 0.85304 0.19734 0.87237 1
X5 Phenol 0.85249 0.11730 0.89982 0.88941 1
X6 Saponins 0.08757 0.45153 -0.12209 -0.05697 -0.14778 1
X7 Wedelolactone 0.27901 0.204803 0.36016 0.26990 0.47040 0.19241 1
Total accessions
X1 Plant height 1
X2 Leaf length 0.8341 1
X3 Carbohydrate 0.9632 0.8023 1
X4 Protein 0.8539 0.7825 0.8348 1
X5 Phenol 0.9652 0.8129 0.9512 0.8705 1
X6 Saponins 0.9334 0.7676 0.9431 0.8263 0.9349 1
X7 Wedelolactone 0.4584 0.2735 0.3587 0.5177 0.2551 0.3497 1

Discussion
156
5.6.1 Inter Accession variability
Variation among phytoconstituents within and between populations of a
species are not uncommon. Morphological as well as biochemical variability has ben
reported among populations, land races, accessions, etc. in a number of crops (Wilson
et al., 1990; Ouendeba et al., 1995, Zeinali et al., 2004). Such variations have genetic
as well as environmental basis. Variations in medicinal plants is often noticed at
chemical level which is due to synthesis and accumulation of various biochemicals.
Since the presently investigated accessions are growing in same environmental and
edaphic conditions the observed variability may be related to their genetic makeup.
The present study on various accessions of Eclipta alba revealed the presence
of inter-accession variation in various phytochemicals. Analysis of variance
(ANOVA) was carried out in different accessions of Eclipta alba covering various
parameters. The ANOVA revealed significant differences among various accessions
for various characters (Table 5.6). The results indicated the presence of adequate
amount of variability in the germplasm under study.
5.6.2 Phytochemical variability vs Morphotype
Phenotypic variations often give valuable clue to the underlying genetic
variations. The variation of phenotypic characters, especially quantitative ones, differs
greatly among populations rather than within the populations (Schemske et al., 1994).
Variations in the alkaloid content due to differences in the morphological characters
have also been observed in Cymbopogon flaxuosus by Seeni et al. (2003).
Quantitative variation in various phytoconstituents has often been noticed in
medicinal plants. This diversity, may be due to the synthesis and accumulation of
various biochemicals, can be continuous or discrete and may be coded by many or
few genes together with some inputs from the environment (Mallet, 1996).
Discussion
157
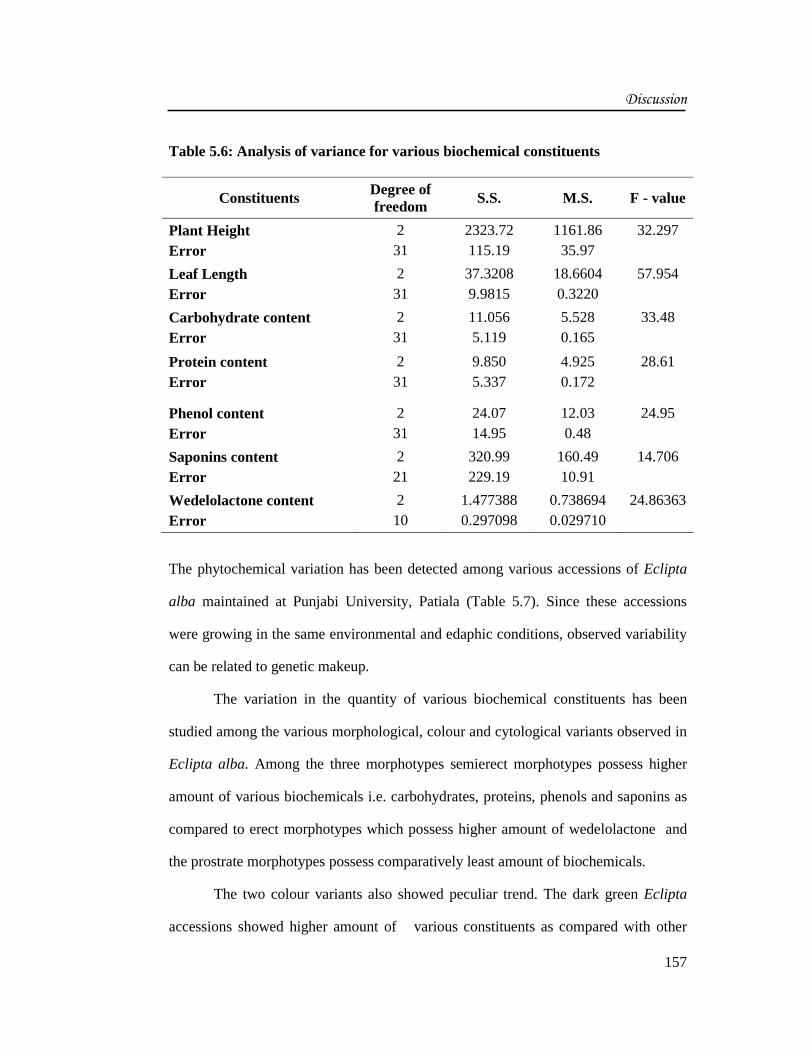
Table 5.6: Analysis of variance for various biochemical constituents
Constituents Degree offreedom
S.S. M.S. F - value
Plant HeightError
231
2323.72115.19
1161.8635.97
32.297
Leaf LengthError
231
37.32089.9815
18.66040.3220
57.954
Carbohydrate contentError
231
11.0565.119
5.5280.165
33.48
Protein contentError
231
9.8505.337
4.9250.172
28.61
Phenol contentError
231
24.0714.95
12.030.48
24.95
Saponins contentError
221
320.99229.19
160.4910.91
14.706
Wedelolactone contentError
210
1.4773880.297098
0.7386940.029710
24.86363
The phytochemical variation has been detected among various accessions of Eclipta
alba maintained at Punjabi University, Patiala (Table 5.7). Since these accessions
were growing in the same environmental and edaphic conditions, observed variability
can be related to genetic makeup.
The variation in the quantity of various biochemical constituents has been
studied among the various morphological, colour and cytological variants observed in
Eclipta alba. Among the three morphotypes semierect morphotypes possess higher
amount of various biochemicals i.e. carbohydrates, proteins, phenols and saponins as
compared to erect morphotypes which possess higher amount of wedelolactone and
the prostrate morphotypes possess comparatively least amount of biochemicals.
The two colour variants also showed peculiar trend. The dark green Eclipta
accessions showed higher amount of various constituents as compared with other

Discussion
158
colour variant (Table 5.8). Among the dark green morphotypes of Eclipta alba,
semierect morphotypes possess higher amount of all the biochemicals followed by
erect morphotypes and prostrate morphotypes.
Table 5.7: Mean performance of three morphotypes of Eclipta alba
Constituents ErectMorphotypes
SemierectMorphotypes
ProstrateMorphotypes
Carbohydrate (mg/g) 17.96±0.321 18.12±0.474 16.91±0.399
Protein (mg/g) 14.20±0.217 14.32±0.441 13.19±0.454
Phenol (mg/g) 19.71±0.587 19.73±1.073 18.03±0.466
Saponins (mg/g) 41.5±1.22 44.3±4.07 36.2±3.24
Wedelolactone (mg/g) 1.11±0.304 0.492±0.137 0.292±0.121
Table 5.8: Mean performance of Eclipta alba accessions with green and darkgreen leaves
Green Eclipta Dark Green EcliptaConstituents
Erect Semierect Prostrate Erect Semierect Prostrate
Carbohydrate
(mg/g)
17.80±0.20 18.04±0.45 16.88±0.37 18.19±0.33 18.28±0.57 17.04±0.53
Protein (mg/g) 14.10±0.23 14.19±0.38 13.16±0.47 14.33±0.10 14.49±0.58 13.3±0.40
Phenol (mg/g) 19.33±0.46 19.58±1.04 17.99±0.41 20.21±0.21 20.04±1.28 18.21±0.65
Saponins(mg/g)
40.8±1.41 44.4±4.14 36.04±3.26 42.3±0.353 44.1±4.86 37.2±4.10
Wedelolactone(mg/g)
-- 0.589±0.13 0.245±0.10 1.11±0.30 -- 0.431±0.03
Analysis of variance (one way ANOVA) revealed highly significant
differences among three morphotypes of E. alba for all the characters observed.
(Table 5.9).
Discussion
159
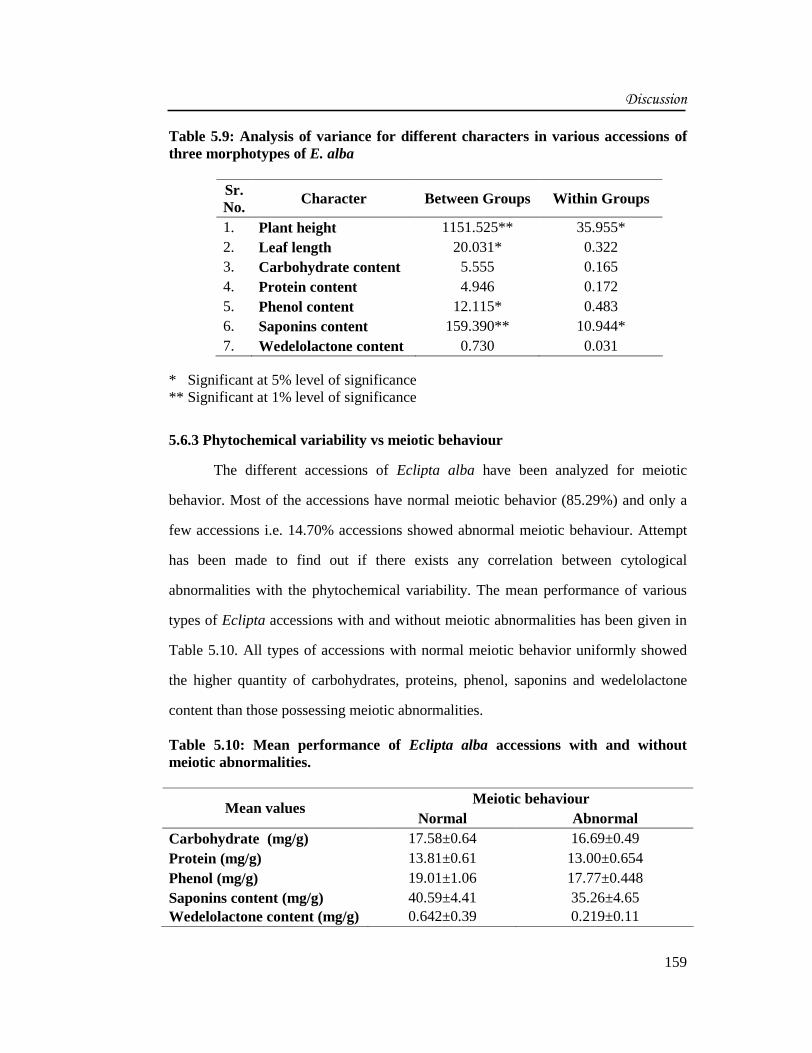
Table 5.9: Analysis of variance for different characters in various accessions ofthree morphotypes of E. alba
Sr.No.
Character Between Groups Within Groups
1. Plant height 1151.525** 35.955*2. Leaf length 20.031* 0.3223. Carbohydrate content 5.555 0.1654. Protein content 4.946 0.1725. Phenol content 12.115* 0.4836. Saponins content 159.390** 10.944*7. Wedelolactone content 0.730 0.031
* Significant at 5% level of significance** Significant at 1% level of significance
5.6.3 Phytochemical variability vs meiotic behaviour
The different accessions of Eclipta alba have been analyzed for meiotic
behavior. Most of the accessions have normal meiotic behavior (85.29%) and only a
few accessions i.e. 14.70% accessions showed abnormal meiotic behaviour. Attempt
has been made to find out if there exists any correlation between cytological
abnormalities with the phytochemical variability. The mean performance of various
types of Eclipta accessions with and without meiotic abnormalities has been given in
Table 5.10. All types of accessions with normal meiotic behavior uniformly showed
the higher quantity of carbohydrates, proteins, phenol, saponins and wedelolactone
content than those possessing meiotic abnormalities.
Table 5.10: Mean performance of Eclipta alba accessions with and withoutmeiotic abnormalities.
Meiotic behaviourMean values
Normal AbnormalCarbohydrate (mg/g) 17.58±0.64 16.69±0.49Protein (mg/g) 13.81±0.61 13.00±0.654Phenol (mg/g) 19.01±1.06 17.77±0.448Saponins content (mg/g)Wedelolactone content (mg/g)
40.59±4.410.642±0.39
35.26±4.650.219±0.11

Discussion
160
Cell cycle is under the control of many genes. A number of genes have been
identified for completion of meiotic cell cycle in plants and animals (Ohta et al.,
1997; Spellman et al., 1998; Doyle et al., 1999). Any disturbance in these genes and
their control system may result in meiotic abnormalities (Kaul and Murthy, 1985;
Caetana-Pereira and Pagliarini, 2001).
The reduced amount of carbohydrate, proteins, phenols and saponins in the
presently studied Eclipta alba plants with abnormal meiosis is an indication that these
plants possess aberrant metabolism as well. The presence of normal meiosis helps
these plants in reproduction and seed setting. The present analysis clearly revealed
that the plants with normal meiotic behavior should be used for medicinal and
commercial purpose.
5.7 BIOACTIVITY OF EXTRACTS
The leaf extracts in three different solvents viz. acetone, methanol and water
were evaluated for antimicrobial activity against five strains of test organisms
obtained from MTCC collection. The results obtained showed that all kind of leaf
extracts of E. alba exhibit bactericidal effects on the test microorganisms. The results
are presented in Fig. XII. Among the three morphotypes, the leaf extracts of erect type
of plants showed maximum antimicrobial activity against all the bacteria except B.
subtilis where the extracts of prostrate type showed maximum activity. The semierect
type of plants also maintained the intermediate position among the three morphotypes.
The most susceptible bacteria was S. aureus followed by E. coli while the most
resistant bacteria were P. aeruginosa followed by K. pneumoniae and B. subtilis.
The antibacterial response of three types of extracts of Eclipta alba showed
good antibiotic activity against both gram positive and gram negative strains except P.
aeruginosa. The reason for the different sensitivity between gram-positive and gram-
negative bacteria could be ascribed to the morphological differences between these
micro organisms, Gram-negative bacteria having an outer phospholipid membrane
carrying the structural lipo-polysaccharide components. This makes the cell wall

Discussion
161
impermeable to lipophilic solutes, while porins constitutes a selective barrier to the
hydrophilic solutes with an exclusion limit of about 600 Da (Nikaido and Vaara,
1985) The Gram-positive bacteria should be more susceptible having only an outer
peptidoglycan layer which is not an effective permeability barrier (Scherer and
Gerhardt, 1971). Phytochemical constituents such as tannins, flavonoids, alkaloids
and several other aromatic compounds are secondary metabolites of plants that serve
as defense mechanisms against predation by many microorganisms, insects and
herbivores (Lutterodt et. al., 1999; Marjorie, 1999). The demonstration of
antibacterial activity of E. alba leaf extracts against both gram positive and gram
negative bacteria may be indicative of the presence of broad spectrum antibiotic
compounds (Srinivasan et. al., 2001). Out of the three solvents used for extraction, the
acetone extracts showed the highest antibiotic activity against the test organisms,
followed by the methanol extracts and water extracts. Different solvents have been
reported to have the capacity to extract different phytoconstituents depending on their
solubility or polarity in the solvent. Acetone extracts in this study might have had
higher solubility for more phytoconstituents, consequently the highest antibacterial
activity.
The biochemical composition of plants is the most common parameter used
for the characterization of plants. The slight variation in biochemical compositions of
the E. alba studied here, however, could be well attributed to genetic rather than
environmental and seasonal factors. Phytochemical screening of the extracts of E.
alba by earlier workers revealed the presence of tannins, coumestans, saponins,
alkaloids, etc. (Dalal et al., 2010; Upadhyay et al., 2001; Wagner et al., 1986; Singh
et al., 2001). Of various phytoconstituents, wedelolactones (coumestan), present E.
alba is believed to be responsible for almost all the antibacterial activities (Dalal et
al., 2009). In addition, other secondary metabolites such as phenolics could be held
partially responsible for some of these biological activities.

Discussion
162
Fig. XII: Histograms showing antibacterial activity of three differentmorphotypes of E. alba against different organisms
Erect
Semierect
Prostrate

Discussion
163
Presently, the high antimicrobial activity observed in the erect type of
morphotypes can be attributed to the presence of higher quantities of wedelolactone
(1.08 mg/g) saponins (42.6 mg/g) and phenols 19.98 mg/g than other morphotypes.
The prostrate type of E. alba plants exhibited minimum antimicrobial activity and had
lower quantities of phytoconstituents (0.374 mg/g, 39.4 mg/g, 18.26mg/g
respectively) too.
5.8 In vitro STUDIES
Plant tissue culture is a fascinating and useful tool which allows the rapid
multiplication of many genetically identical plants using relatively small amounts of
space, supplies and time. In vitro propagation has been proved as a potential
technology for large scale multiplication of medicinal plant species (Lui and Li, 2001;
Wawrosch et al., 2001; Martin, 2002, 2003, Azad et al., 2005; Faisal et al., 2003;
Hassan and Roy, 2005). Therefore, it was important to develop an efficient
micropropagation technique for Eclipta alba to rapidly disseminate superior clones
once they are identified. Micropropagation via shoot culture, often utilized to
maintain clonal fidelity, would be especially appropriate in this respect (Sen and
Sharma, 1991). Micropropagation of E. alba plants was achieved through nodal
culture as well as callus culture. In vitro micropropagation of Eclipta alba from nodal
segment explants was reported earlier by Franca et al., 1995; Borthakar et al., 2000;
Gawde and Partkar, 2004; Dhaka and Kothari, 2005; Baskaran and Jayabalan, 2005;
Hussain and Anis, 2006; Han et al., 2007; Ray and Bhattacharya, 2008. Plantlet
regeneration via callus formation in E. alba is reported by Zafar and Sagar (1999).
5.8.1 Shoot proliferation
Growth regulators especially cytokinins (Kinetin, BA, 2 ip, Zeatin) are one of
the most important factors affecting shoot proliferation (Lane 1979; Stolz 1979;
Bhojwani 1980; Garland and Stoltz, 1981). Studies conducted by different workers
clearly indicated that BA is more effective, reliable and useful cytokinin for shoot

Discussion
164
proliferation in E. alba (Franca et al., 1995; Gawde and Partkar, 2004; Dhaka and
Kothari, 2005; Baskaran and Jayabalan, 2005; Hussain and Anis, 2006; Han et al.,
2007; Ray and Bhattacharya, 2008). However, Borthakur et al. (2000) reported
maximum number of shoots in MS medium supplemented with 0.05 mg/L kinetin.
Presently, nodal explants of Eclipta alba on MS media supplemented with two
concentrations (0.5 mg/L and 1.0mg/L) of BAP alone and BAP with NAA were
cultured for shoot regeneration. A combination of BAP with NAA was not found
satisfactory for shoot multiplication (Table 4.15). This is in contrast to earlier work by
Hassan et al. (2008) who reported 94% of explants forming 18 shoots on the same
medium. In the present study the highest frequency of shoot production was recorded
in MS medium supplemented with 1.0 mg/L BA which confirms the earlier studies by
Franca et al. (1995), Gawde and Paratkar (2004) and Dhaka and Kothari (2005).
5.8.2 Regeneration from Callus
Zafar and Sagar (1999) are probably the first to report of plantlet regeneration
via callus formation in E. alba. They raised plants from totipotent callus of leaf, stem
and root cultured on MS medium supplemented with various growth regulators and
found 2,4-D (4.52µM); 2,4-D (4.52µM ) + 6-BA (4.43); 2,4-D (4.52µM ) + 6-BA
(4.43) + Kin (4.64µM) + IAA (5.7µM) and 2,4-D (22.62µM) to be the most effective
for callus formation.
In the present study, two media were tried i.e. 2,4-D (1.0 mg/L) and 2,4-D (1.0
mg/L) + 6-BA (1.0 mg/L) and callus response was observed in both media as reported
by Zafar and Sagar (1999). The media which showed regeneration from callus are 6-
BA (2.0 mg/L) + IAA (0.5 mg/L), 6-BA (2.0 mg/L) + NAA (0.5 mg/L), Kin (2.0
mg/L) + NAA (0.5 mg/L) and Kin (1.0 mg/L) + IAA (0.2 mg/L). These combinations
of cytokinin and auxin concentrations have not been reported in earlier investigations
in E. alba.

Discussion
165
5.8.3 Rooting
Rooting of microshoots has been reported to be controlled by various factors
like growth regulators in the medium (Bhojwani and Razdan, 1992; Feng et al., 2000;
Abrie and Staden, 2001), basal salt composition (Skirvin and Chu, 1979; Zimmerman
and Broome, 1981; Garland and Stoltz, 1981), genotype (Rines and McCoy, 1981) as
well as cultural conditions (Murashige, 1977). Auxins were found to stimulate root
growth (Thimann and Went, 1934). Use of auxins singly or in combination for rooting
was reported by different authors (Sahoo and Chand, 1998; Ajithkumar and Seeni,
1998; Rai, 2002, Baskaran and Jayabalan, 2005; Sivakumar and Krishnamurthy,
2000; Hassan and Roy, 2005; Rahman et al., 2006; Baksha et al., 2007).
According to Dhaka and Kothari (2005) rooting was best achieved on MS
medium supplemented with 1.0 mg/L IBA. Baskaran and Jayabalan (2005) observed
that rooting was highest (94.3%) on full strength MS medium containing 9.8 μM IBA.
Hussain and Anis (2006) demonstrated most efficient rooting on half strength MS
medium augmented with 0.5 µM IBA. Hassan et al. (2008) reported in vitro raised
shoots rooted on half strength MS medium with 0.5 mg/L IBA +0.1 mg/L NAA.
In the present study also IBA has been proved to be more effective among all
other auxins. Maximum root length was achieved in half strength MS medium with
1.0 mg/L IBA which supports earlier investigations by Baskaran and Jayabalan
(2005). However Hussain and Anis (2006) reported maximum root length using 0.5
IBA. This difference may be due to the difference in the genotype of the plant used. In
Eclipta alba, Franca et al. (1995) achieved multiple rooting using MS medium
supplemented with 2.4 µM 2-isopentyladenine (2iP). Gawde and Paratkar (2004)
observed best response of rooting in the low amount of BA (0.44 µM). Cent per cent
shoots developed roots directly from shoot base when transferred to growth regulator-
free MS medium (Ray and Bhattacharya, 2008).

Discussion
166
5.8.4 Hardening
For tissue culture generated plants hardening is the most crucial step. The
tissue culture plants are very soft to face ambient environmental conditions (Bhojwani
and Razdan, 1992). In the present study, rooted plantlets were transferred from culture
bottles to plastic pots containing garden soil: farmyard manure (1:1). Borthakur et al.
(2000) transferred the plantlets to pots containing 1:1 mixture of autoclaved sand and
soil. Earlier Gawde and Paratkar (2004) tried a variety of combinations of the
hardening media like soil, sand: soil (1:1), soil: vermicompost (1:1) and knops
hydroponic solutions and achieved 4-5%, 2%, 95% and 100% survival, respectively.
Baskaran and Jayabalan (2005) suggested a mixture of garden soil, farmyard manure
and sand (2:1:1) as hardening media for E. alba plantlets.
Priming of culture regenerated propagules has been recommended for
obtaining better acclimatized plants (Nowak and Shulaev, 2003; Hazarika, 2003). For
priming E. alba microshoots 6.3 µM of chlorocholine chloride (CCCl) was found
most effective (Ray and Bhattacharya, 2008). The authors observed increased number
of roots, elevation of chlorophyll level in leaves and increase in plant biomass in 30
day old treated shoots of E. alba. The primed micropropagated plants were healthy
with 100% survival in soil as compared to 84% survival in non treated plants.
The technique described here appears to be readily adaptable for large scale
clonal propagation and plantation for sustainable use in the industry. Moreover, by
standardizing the protocols for clonal propagation of selected elite plants, it is
possible to achieve a tenfold increase in the products per unit area of cultivation
(Hassan and Roy, 2005).
5.8.5 Somaclonal Variations
The generation of phenotypic variation (quantitative or qualitative) during in
vitro micropropagation has been described as somaclonal variations. These variations

Discussion
167
are most likely to arise as a result of chromosome structural changes (e.g. small
deletions or duplications), gene mutation (both dominant and recessive have been
recovered as the latter being more common), gene amplification, small chromosome
re-arrangement, transposable elements (these may be induced to transpose due to
stress during in vitro culture) and mitotic crossing over during culture induction.
Chromosomal aberrations like laggards, bridges in meiosis I and meiosis II,
micronuclei and change in ploidy level have been observed in tissue regenerants
(Larkin and Scowcroft, 1981; Gupta, 1998; Pontaroli and Camadro, 2005).
Somaclonal variation can be assessed by analysis of phenotype, chromosome number
and structure, proteins, or direct DNA evaluation of plants (De Klerk, 1990).
Micropropagated plants have exhibited various morphological and
biochemical variations due to mutations believed as somaclonal variations (Larkin
and Scowcroft, 1981). Such variations are the case of changes in morphological
features like plant height and leaf size among micropropagated plants (Ravindran et
al., 2004).
In the present study, attempt has been made to induce genetic variability
among three morphotypes of Eclipta alba. The present results exhibit the occurrence
of variation in morphological and biochemical characteristics when we compare the
mother plant and their tissue culture generated plants. All the accessions showed
increased leaf width, carbohydrate and protein contents. The extent of variation
however, differs from accession to accession. This could be attributed to the
differences in genotypes of various accessions. The clonal plants were shorter in size
with decreased branches as compared to their mother plants (control). However, there
was marginal increase in leaf breadth and quantity of basic constituents carbohydrates
and proteins. Somaclonal variations observed in the present investigations may be
because of hormonal effects as suggested by Yipeng et al. (2005) and Ducos et al.
(2003).

Discussion
168
5.9 CULTIVATION TRIAL
Cultivation of medicinal plants has become almost essential due to unplanned
collection of herbs from wild sources. There is a need for a scientific approach for
propagation of medicinal plants and to collect relevant information regarding agro
technology, genuine planting material, economics of field cultivation, high yielding
varieties, etc. One has to explore wild medicinal plant species and to bring them under
cultivation. There is enormous potentiality for commercial cultivation of useful herbs
and production of safe green medicines by establishing plant based industries tract for
the socio-economic development and conservation of genetic diversity. There is need
to standardize cultivation practices to ensure the sustainable supply of standard plant
material, and to reduce the load on wild forms, there is need to mark out better
chemotypes and to standardize their cultivation practices.
The demand of Indian medicinal plants has increased over the years in the
international market. In current years, E. alba has been harvested in huge quantities
from the wild to meet the pharmaceutical demand. For this purpose raw material from
cultivation is always preferred to the collection from wild, however, systematic
cultivation of Eclipta alba plant is not adopted till date. Though Eclipta alba has
acquired great commercial importance for its medicinal and cosmetic preparations but
information is scarce about agronomic management of this crop.
In the present study, Eclipta shows good response treatment to NPK as
compared to NPK + FYM, FYM treatment and control plants. Carbohydrates and
protein content was highest in the plants treated with NPK (5 g/m2) followed by NPK
(2.5 g/m2) and NPK (5 g/m2) + FYM (2 kg/m2). Combination of NPK and FYM
showed modest productivity. In the absence of FYM there were large interactions
between each of the fertilizer nutrients, but with FYM the interactions were small.

Discussion
169
FYM increased the response to N applied alone, but decreased the N response where
basal P and K fertilizers were also applied (Boyd, 1961).
Effects of fertilizers on production of medicinal plants has been studied in
Rheum habarbarum and Beta vulgaris (Ombidi and Saigusa, 2000), Echinacea
purpurea (El- Gengaihi et al., 1998), Echinacea angustifolia (Berti et al., 2002),
Tanacetum parthenium (Default et al., 2003), etc. Increased amount of active
principle with application of fertilizers has been reported in Datura spp. (Esendal et
al., 2000), Echinacea angustifolia (Berti et al., 2002) Atropa belladonna (Baricevic et
al., 2002) and Artemesia spp. (Usha and Swamy, 2002). The data were analyzed
following ANOVA technique. The increase in plant height was significantly affected
by fertilizer treatment (Table 5.11).
Discussion
170
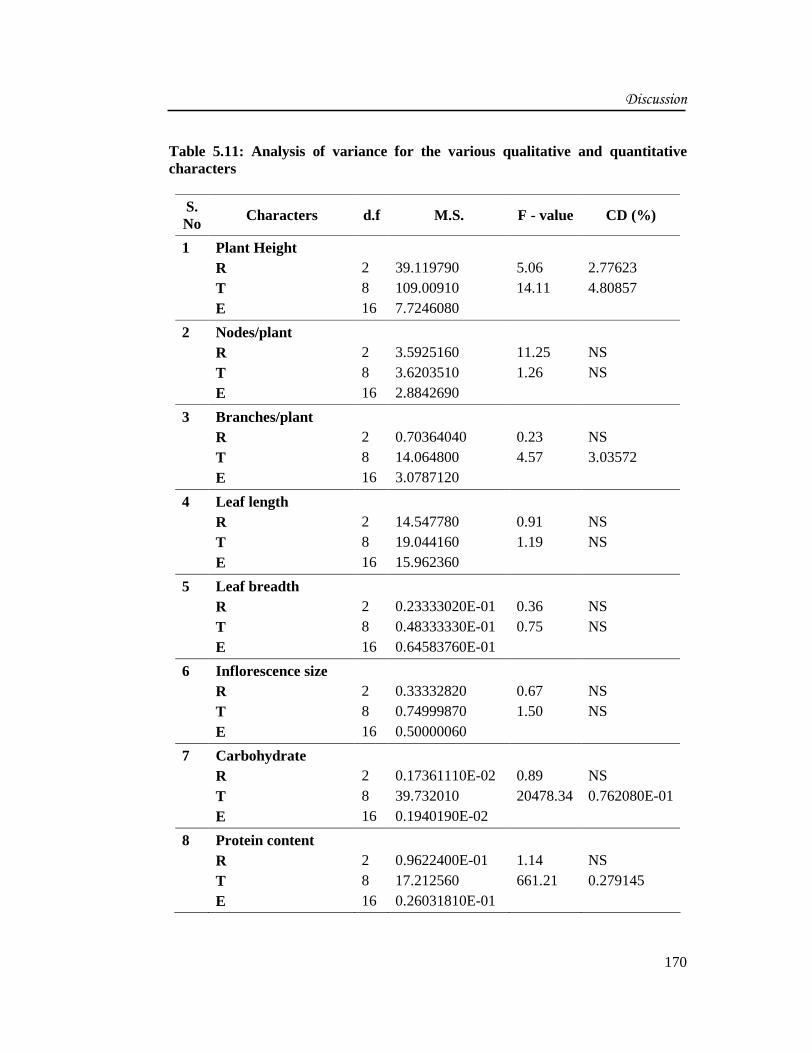
Table 5.11: Analysis of variance for the various qualitative and quantitativecharacters
S.No
Characters d.f M.S. F - value CD (%)
1 Plant HeightRTE
2816
39.119790109.009107.7246080
5.0614.11
2.776234.80857
2 Nodes/plantRTE
2816
3.59251603.62035102.8842690
11.251.26
NSNS
3 Branches/plantRTE
2816
0.7036404014.0648003.0787120
0.234.57
NS3.03572
4 Leaf lengthRTE
2816
14.54778019.04416015.962360
0.911.19
NSNS
5 Leaf breadthRTE
2816
0.23333020E-010.48333330E-010.64583760E-01
0.360.75
NSNS
6 Inflorescence sizeRTE
2816
0.333328200.749998700.50000060
0.671.50
NSNS
7 CarbohydrateRTE
2816
0.17361110E-0239.7320100.1940190E-02
0.8920478.34
NS0.762080E-01
8 Protein contentRTE
2816
0.9622400E-0117.2125600.26031810E-01
1.14661.21
NS0.279145