dna replication - copying the genetic blueprint
TRANSCRIPT

1
Marc S. Wold
Medical Biochemistry September 13, 2010
Readings: Lippencot - Chapter 29 Lehninger - Chapter 24
DNA replication-copying the genetic blueprint
Outline
DNA replication – General mechanism of DNA replication
– Replication enzymes
– Prokaryotic replication – Eukaryotic replication

2
Study Questions and Key Concepts 1. Define stages of replication & semi-discontinuous synthesis 2. What are the two classes of DNA polymerases? What is the
general mechanism and requirements for DNA polymerase function including the function of DNA polymerase accessory complexes?
3. What are the 7 types of proteins needed for needed for progression of replication fork? The function of each class at replication fork?
4. What are the similarities and differences between prokaryotic and eukaryotic DNA replication?
5. Why is a linear chromosome a problem in replication? How do mammalian cells overcome this problem?
6. What is the enzymatic activity of telomerase and what is its role in the cell?
7. What is the general mechanism of initiation of replication?
8. What is the arrangement of origins of replication on eukaryotic chromosomes?
DNA RNA Proteins
Cellular Information Flow
Replication
DNA Repair
Transcription Translation
RNA processing
DNA metabolism
DNA RNA Proteins
Protein degradation
Chromatin Structure - organization of DNA in Cell
Cell Cycle regulation - coordination of cellular information flow with other cellular and tissue processes
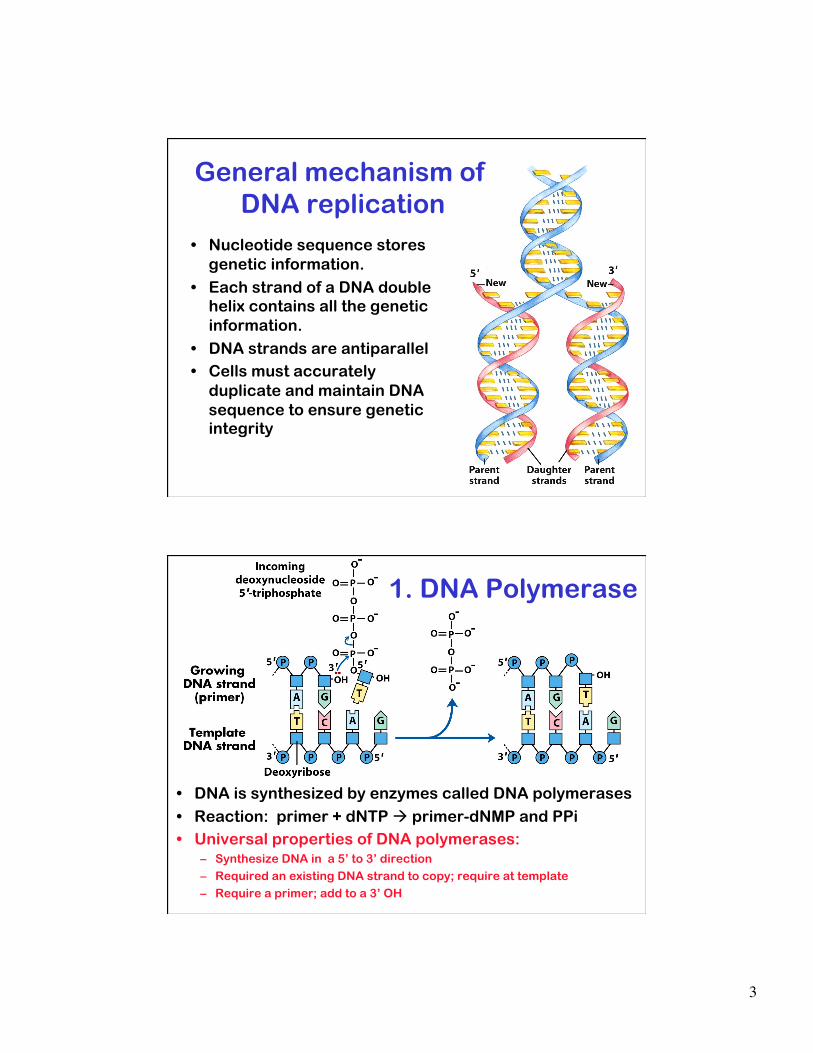
3
• Nucleotide sequence stores genetic information.
• Each strand of a DNA double helix contains all the genetic information.
• DNA strands are antiparallel • Cells must accurately
duplicate and maintain DNA sequence to ensure genetic integrity
General mechanism of DNA replication
• DNA is synthesized by enzymes called DNA polymerases
• Reaction: primer + dNTP primer-dNMP and PPi • Universal properties of DNA polymerases:
– Synthesize DNA in a 5’ to 3’ direction – Required an existing DNA strand to copy; require at template – Require a primer; add to a 3’ OH
1. DNA Polymerase

4
DNA Replication - definitions • Origin of Replication - DNA sequence
at which replication initiates • Replication forks - points of
replication that move through DNA • Unidirectional or bidirectional
replication - replication occurs with either 1 or 2 forks, bidirectional replication most common
Semi-discontinuous DNA replication • Two strands are antiparallel & DNA polymerase synthesizes 5’=>3’ • At a replication fork the two strands in the double helix must be
synthesized by a different mechanisms at the same time. • DNA synthesis is continuous on the leading strand and discontinuous on
the lagging strand. • The short DNA fragments on lagging strand are called Okazaki fragments. • DNA polymerase requires a primers so each Okazaki fragment must begin
with a primer.

5
Definitions: Stages of DNA Replication • Initiation of Replication - The process
that leads primer synthesis and assembly of replication fork.
• Elongation - The process of DNA synthesis; replication forks move through DNA synthesizing new DNA strands.
• Termination - When replication stops. Usually occurs when replication forks meet.
DNA polymerases • There are two classes of DNA polymerases in cells
• Classical DNA polymerases (Replicative DNA polymerases) - involved in DNA replication of undamaged DNA and some DNA repair processes
• Translesion DNA polymerases - involved in DNA repair and DNA synthesis of damaged DNA
Clinical Correlation: Many antiviral agents target viral DNA polymerases.
• Acyclovir (Acycloguanosine, Zovirax®) - inhibitor of Herpes DNA polymerases
• Entecavir (Baraclude®) - inhibites hepatitis B virus (HBV) DNA polymerase

6
DNA polymerases in E. coli • There are five DNA polymerases in E. coli - 3 classical &
2 translesion • Classical polymerases:
– DNA polymerase and exonuclease activities – Number of copies per cell and activity vary (e.g. processivity)
– Primary role - replication (III) and repair processes (I & II).
Table for illustrative purpose
DNA polymerase III: a replication machine
• DNA Polymerase III holoenzyme is a complex replication machine: 10 subunits, rapid rate of synthesis (1000 nt/sec), accurate (1 error/105 nt) and high processivity (>105 nt before dissociation).
• Key concept: DNA Polymerase III holoenzyme is composed of three subcomplexes
Table for illustrative purpose

7
Polymerase accessory factors: Sliding
clamps & Clamp loaders
• DNA polymerase III achieves high processivity by being topologically linked to DNA.
• Sliding clamp: toroid shaped Beta complex is a sliding clamp. Interacts with core polymerase and holds it on the DNA.
• Clamp loader: beta is loaded onto double-stranded DNA by the gamma complex.
• A similar mechanism is used other processive DNA polymerases
2. Primases - starting nascent DNA chains
• Primases synthesize short RNA oligonucleotide primers
• Can initiate synthesis on ssDNA de novo (no 3’=OH needed).
• Most primases start synthesis at a random sites
• Usually part of protein complex at replication fork
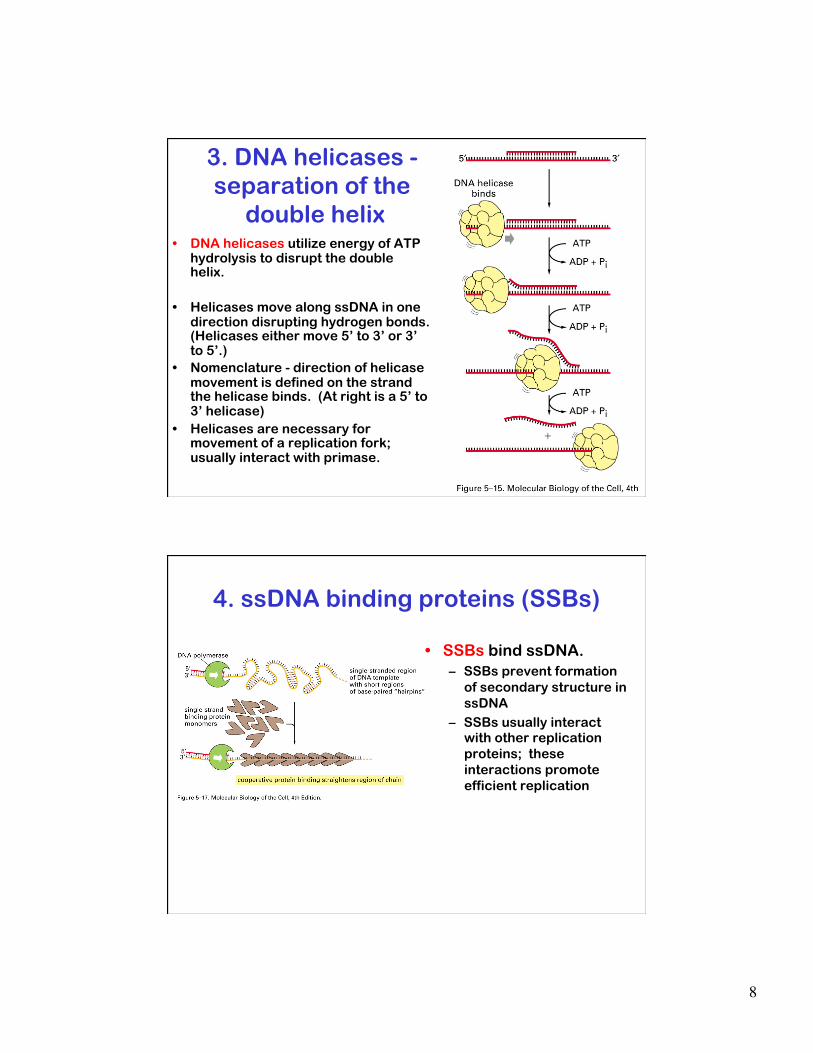
8
3. DNA helicases - separation of the
double helix • DNA helicases utilize energy of ATP
hydrolysis to disrupt the double helix.
• Helicases move along ssDNA in one direction disrupting hydrogen bonds. (Helicases either move 5’ to 3’ or 3’ to 5’.)
• Nomenclature - direction of helicase movement is defined on the strand the helicase binds. (At right is a 5’ to 3’ helicase)
• Helicases are necessary for movement of a replication fork; usually interact with primase.
4. ssDNA binding proteins (SSBs)
• SSBs bind ssDNA. – SSBs prevent formation
of secondary structure in ssDNA
– SSBs usually interact with other replication proteins; these interactions promote efficient replication

9
5. DNA nucleases • Nucleases DNA or RNA
– Nucleases can degrade DNA from its end 5’3’ or 3’5’ (exonucleases) or internally (endonuceases)
– 5’3’ exonuclease required in replication to remove RNA primers to give continuous DNA strand
• DNA ligases form phosphodiester bonds; join strands of DNA – Require high energy
cofactor (ATP or NAD)
6. DNA ligases
• Parental strands start replication as a single helix but end replication as one strand of each of two separate helices
• Topoisomerases release the links between the parental DNA strands
7. Topoisomerases

10
Type I Topoisomerases
• Type I topoisomerases make a transient ssDNA break that then allow the DNA to swivel
• Change links between strands (L) in units of one
Clinical Correlation: • Topoisomerase inhibitors (antibiotics) - quinolones, fluroquinolones (e.g.
nalaidixic acid, coumermycin A, and novobiocin).
• Topoisomerase I inhibitors (chemotherapy) - camptothecin & less toxic varients: Topotecan (Hycamtin®), irinotecan (Camptosar®) are used in chemotherapy.
• Type II topoisomerase cause a transient dsDNA break.
• Change links between strands (L) in units of two
• Can link or unlink two molecules of DNA (catenation or decatenation).
• Form a covalent intermediate with the transiently broken end(s) of the DNA.
• Most topoisomerases relax supercoiled DNA.
Clinical Correlation: • Topoisomerase II inhibitors -
Etoposide phosphate (Eposin®, Etopophos®, VP-16®) - used in chemotherapy.
Type II Topoisomerases

11
Replication fork and elongation
• Replication proteins act coordinately at the replication fork to promote efficient DNA synthesis
• Leading and lagging strand synthesis are coordinated • Movie showing E. coli fork at:
http://www.freesciencelectures.com/video/dna-replication-process/
Table for illustrative purpose
Replication Fork
• Helicase separating DNA strands
• Primase synthesizing primers for lagging strand
• Primosome: “helicase/primase complex” synthesizes multiple primers on the lagging strand
• SSB stabilizing ssDNA • Topoisomerase
removing links between parental DNA
• Nuclease removing primers
• Processing of Okazaki fragments

12
Processing of Okazaki fragments • Processing of okazaki
fragments requires: – nuclease to remove primer, – DNA polymerase to fill in gap and – ligase to seal nick
• In E. coli, processing requires – DNA pol I (5’3’ exonuclease and
DNA polymerase activities) and – ligase
Eukaryotic replication: Different proteins, similar functions and mechanism
• Replication proteins have have conserved functions.
• The structures of Pol III beta subunit and PCNA shown a right
Clinical Correlation: • PCNA antibody - used in diagnosis as a
marker for proliferating cells.
Table for illustrative Purpose only

13
Eukaryotic DNA polymerases
• There are at least 15 polymerases in eukaryotic cells
• Designated by Greek letters • Classical DNA polymerases (Replicative polymerases)
– Alpha polymerase & primase - initiation of replication & okazaki fragments
– Beta-DNA repair
– Gamma - mitochondrial DNA replication – Delta & epsilon - chromosomal DNA replication
• Translesion DNA polymerases (DNA repair & DNA synthesis of damaged DNA) – Zeta, eta, theta, … - DNA repair and bypass synthesis
DNA polymerase alpha, delta and epsilon all function at the eukaryotic replication fork
Prokaryotic vs Eukaryotic Replication
Prokaryotes differences
– 20-30 minute cell doubling – Small genomes (105-107) – Single chromosome – Single origin of replication
similarities – Efficient DNA synthesis – Highly regulated
• Replicate once and only once per cell cycle
• Coordinate with nutritional state
• DNA damage and checkpoints can stop replication
Eukaryotes differences
– 8 hour S phase – Large genomes (107-1012) – Multiple chromosomes – Multiple origins of replication
similarities – Efficient DNA synthesis – Highly regulated
• Replicate once and only once per cell cycle
• Coordinate replication with cell cycle
• DNA damage and checkpoints can stop replication
E.coli cell Human metaphase chromosomes
Chromosomal DNA

14
Eukaryotic chromosomes
• Centromere = attachment point for proteins that link the chromosome to the mitotic spindle
• Telomeres = sequences at the end that help to stabilize the chromosome.
• Consequences of linear chromosomes: DNA ends • Check point activation (ends must not activate checkpoints)
• 5’ end problem
Telomeres
• Telomeres: Specialized structure at the ends of eukarkyotic chromosomes
• Telomeres contain thousands of copies of short 6-8 nt, GC-rich repeats. (In humans: (CCCTAA)n)
• Telomere sequences bound by multiple telomeric proteins and form a specialized, looped nucleoprotein complex (T-loop)
• Telomere structure prevents DNA end from being recognized as double strand break
Top: Proposed stucture of Telomere loop (T loop)
Bottom: electron micrograph of Mouse telomere

15
The problem with 5’ ends of
linear genomes) • How to synthesize the 5’
end of the lagging strand.
• The last primer must be removed and filled in or else there will be loss of genetic material.
Lagging strand!
Telomere Synthesis
• 3’ (parental ) strand is extended by specialized RNA protein complex: telomerase
• Telomerase contains a “reverse transcriptase” activity that uses internal RNA as template to synthesize telomere repeats de novo
• 5’ strand is synthesized by lagging strand mechanism • Telomerase extension prevents loss of DNA at the 5’ end

16
Telomerase, cancer, …
• Most human cells do not express telomerase and telomere length decreases as normal cells grow.
• Immortalized cells usually have induced expression of telomerase • Normal cells that have telomerase expression activated do not stop
growing. • Telomerase expression is a critical factor in transformation of cells and the
cancer development. • Telomere length decreases with age (and stress).
Aubert & Lansdorp (2008) Physiol Rev 88, 557 Harley et al (1990) Nature 354, 458 Epel et al (2004) PNAS 101, 17312
“Accelerated telomere shortening in response to life stress”
General Mechanism of
Initiation • Specific complexes form at
origin • Origin is activated causing
local opening of the DNA helix and loading of DNA helicase
• Primase binds and synthesizes initial RNA primers
• DNA polymerase extends primers. Leading and lagging strand synthesis established
• The primary (only) specificity in DNA replication is at initiation.
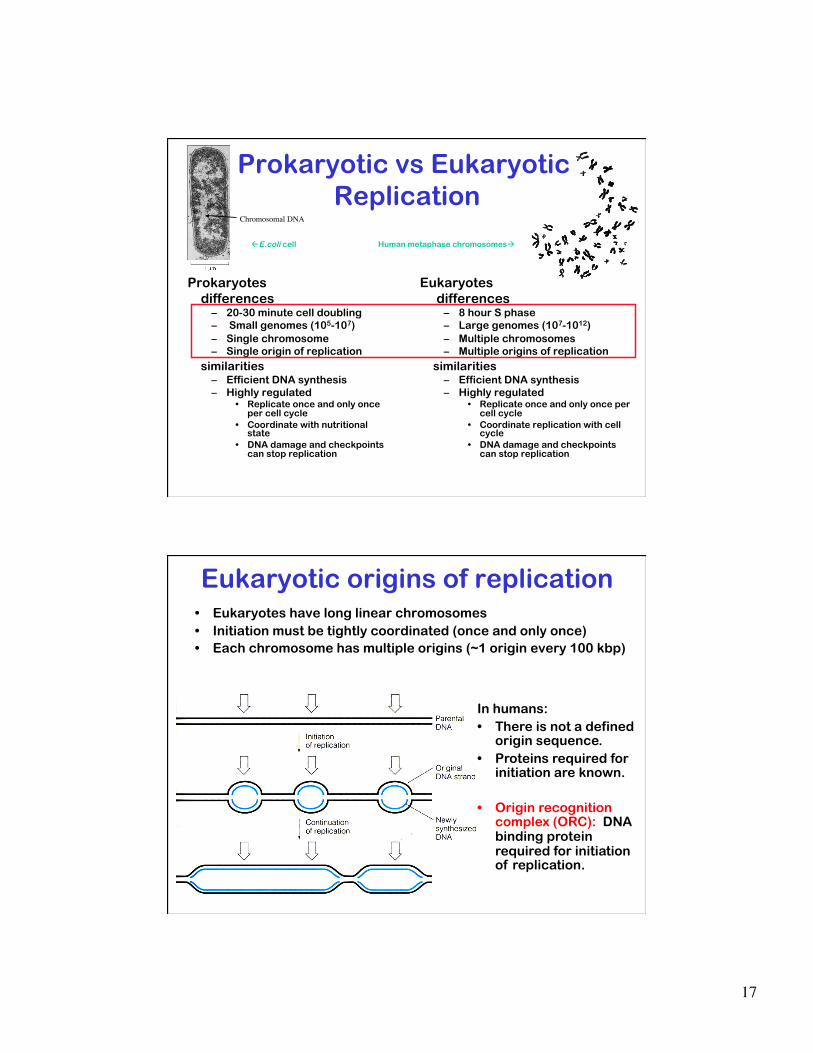
17
Prokaryotic vs Eukaryotic Replication
Prokaryotes differences
– 20-30 minute cell doubling – Small genomes (105-107) – Single chromosome – Single origin of replication
similarities – Efficient DNA synthesis – Highly regulated
• Replicate once and only once per cell cycle
• Coordinate with nutritional state
• DNA damage and checkpoints can stop replication
Eukaryotes differences
– 8 hour S phase – Large genomes (107-1012) – Multiple chromosomes – Multiple origins of replication
similarities – Efficient DNA synthesis – Highly regulated
• Replicate once and only once per cell cycle
• Coordinate replication with cell cycle
• DNA damage and checkpoints can stop replication
E.coli cell Human metaphase chromosomes
Chromosomal DNA
Eukaryotic origins of replication • Eukaryotes have long linear chromosomes • Initiation must be tightly coordinated (once and only once) • Each chromosome has multiple origins (~1 origin every 100 kbp)
In humans: • There is not a defined
origin sequence. • Proteins required for
initiation are known.
• Origin recognition complex (ORC): DNA binding protein required for initiation of replication.
18
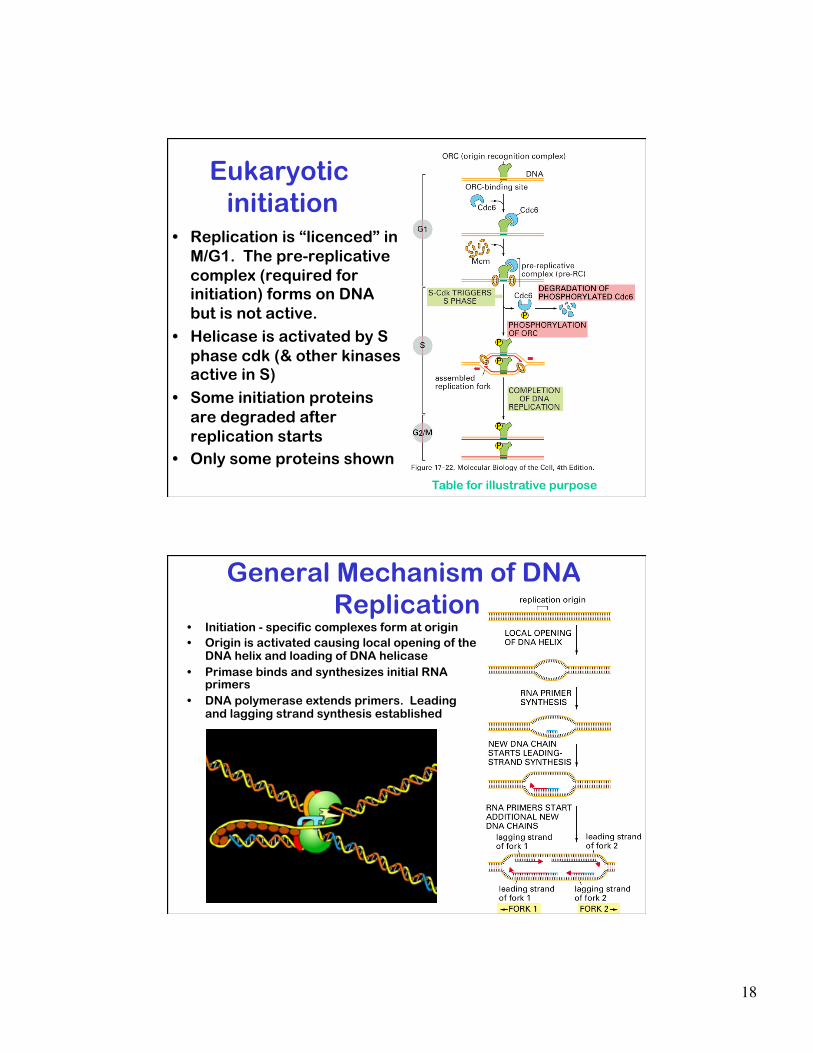
Eukaryotic initiation
• Replication is “licenced” in M/G1. The pre-replicative complex (required for initiation) forms on DNA but is not active.
• Helicase is activated by S phase cdk (& other kinases active in S)
• Some initiation proteins are degraded after replication starts
• Only some proteins shown
Table for illustrative purpose
General Mechanism of DNA Replication
• Initiation - specific complexes form at origin • Origin is activated causing local opening of the
DNA helix and loading of DNA helicase • Primase binds and synthesizes initial RNA
primers • DNA polymerase extends primers. Leading
and lagging strand synthesis established

19
DNA mutagenesis and repair 99:163 - Medical Biochemistry
September 15, 2010
Marc S. Wold
Readings: Lippencott - Chapter 29 Lehninger - Chapter 25
Hakem (2008) EMBO J. 27, 589
Outline
• DNA repair – DNA repair proteins and mutations
– Types of damage
– Cellular DNA Repair pathways – Defects in DNA metabolism and human disease

20
Study Questions & Key Concepts 1. What are the types of modifications cause DNA
damage? 2. How is damaged DNA changed to a genetic mutation? 3. What DNA repair pathways/processes occur prior to
DNA replication? During or after DNA replication? 4. What is the general mechanism of (what type of damage
is repaired and what is the general mechanism of: proofreading, mismatch repair, NER, BER, error-prone repair (translesion synthesis), NHEJ, recombinational repair?
5. What pathway is defective in Xeroderma pigmentosum?HNPCC? Fanconi’s Anemia?
6. How do the two pathways for repairing double-stranded DNA breaks differ?
DNA RNA Proteins
Cellular Information Flow
Replication
DNA Repair
Transcription Translation
RNA processing
DNA metabolism
DNA RNA Proteins
Protein degradation
Chromatin Structure - organization of DNA in Cell
Cell Cycle regulation - coordination of cellular information flow with other cellular and tissue processes

21
Dangerous environments
• Many environments are dangerous
• Danger can be mitigated but not always eliminated
• This is also true in cells where the environment contributes to DNA damage
Dangerous environments
• Many environments are dangerous
• Danger can be mitigated but not always eliminated
• This is also true in cells where the environment contributes to DNA damage
• Environmental chemicals
• Radiation • Natural chemistry
• Normal metabolic processes

22
Environmental Chemicals Many chemicals can modify DNA. • Aflatoxin (peanut butter)
• Cyclic hydrocarbons (grilling and cooking)
• Nitrates - Sodium nitrate (preservative in prepared meat products-prevents growth of Clostridium botulinum. Converted to nitrosamines by in stomach)
• Reactive oxygen species (O2-,
HO•) Produced by phagocytes (to kill pathogens), during electron transport in mitochondria metabolism and by oxidizing agents (H2O2)
• Most modified bases are repaired by one or more repair systems.
Types of DNA damage
Thymine dimer (& other intra-base bonds) Caused by UV
Single strand break Caused by reactive chemicals, oxidative damage, radiation
Double strand break Caused by reactive chemicals, oxidative damage, radiation
Depurination (G or A) Spontaneous
Deamination (C, A, G) Spontaneous U!
*
* Chemical modification (alkalyation, oxidative damage, …) Caused by reactive chemicals, reactive oxygen species
C! Mismatchs Caused by replication errors
Intercalation (binding of chemicals between base pairs) Cause by chemical exposure
inter-strand Crosslinks Caused by reactive chemicals, radiation, free radicals

23
Frequency of DNA damage DNA lesions/mammalian cell/day
(from J. Bartek)
• 50,000 single-strand breaks
• 10,000 depurinations • 600 deaminations
• 2,000 oxidative lesions • 5,000 alkylated bases
• 10 inter-strand crosslinks
• 10 double-strand breaks
U!
*
*
C!
DNA repair: Why does DNA damage cause
mutations? • Damaged (chemically
modified) DNA has altered (or no) base pairing
• DNA polymerase can mis-read a damaged template and incorporate the wrong base

24
• Making a mutation: during replication, the wrong base is inserted opposite a damaged base causing a permanent change in genetic information - a mutation.
• Replication fork arrest: the replicative DNA polymerase�cannot copy the damaged base causing the replication fork to stall.
Replication converts damage to a mutation
Types of DNA Repair
• Replication fidelity and Post replicative repair – Proof-reading (during replication) – mismatch repair
• Damage Repair – Nucleotide Excision – Base Excision – Direct reversal – Transcription Coupled – Specialized repair (inter-strand cross links, specialized
damage)
• Translesion Synthesis & Error-prone Repair (SOS) • Double-strand Break repair
– Nonhomologous End Joining (NHEJ)
– Recombinational repair
DNA damage occurs as a result of environmental (chemical modification, radiation, etc.) insult, natural cellular processes (reactive oxygen species) or fundamental chemistry. All damage needs to be repaired.

25
Types of DNA Repair
• Replication fidelity and Post replicative repair – Proof-reading (during replication) – mismatch repair
• Damage Repair – Nucleotide Excision – Base Excision – Direct reversal – Transcription Coupled – Specialized repair (inter-strand cross links, specialized
damage)
• Translesion Synthesis & Error-prone Repair (SOS) • Double-strand Break repair
– Nonhomologous End Joining (NHEJ)
– Recombinational repair
DNA damage occurs as a result of environmental (chemical modification, radiation, etc.) insult, natural cellular processes (reactive oxygen species) or fundamental chemistry. All damage needs to be repaired.
• All DNA polymerases: – Synthesize DNA in a 5’ to 3’ direction – Required an existing DNA strand to copy; require at template – Require a primer; add to a 3’ OH
• DNA polymerases can have several intrinsic enzyme activities: – DNA polymerase (all) – 3’ to 5’ exonuclease (many) – 5’ to 3’ exonuclease (a few - E.coli Pol I)
DNA Polymerase
26
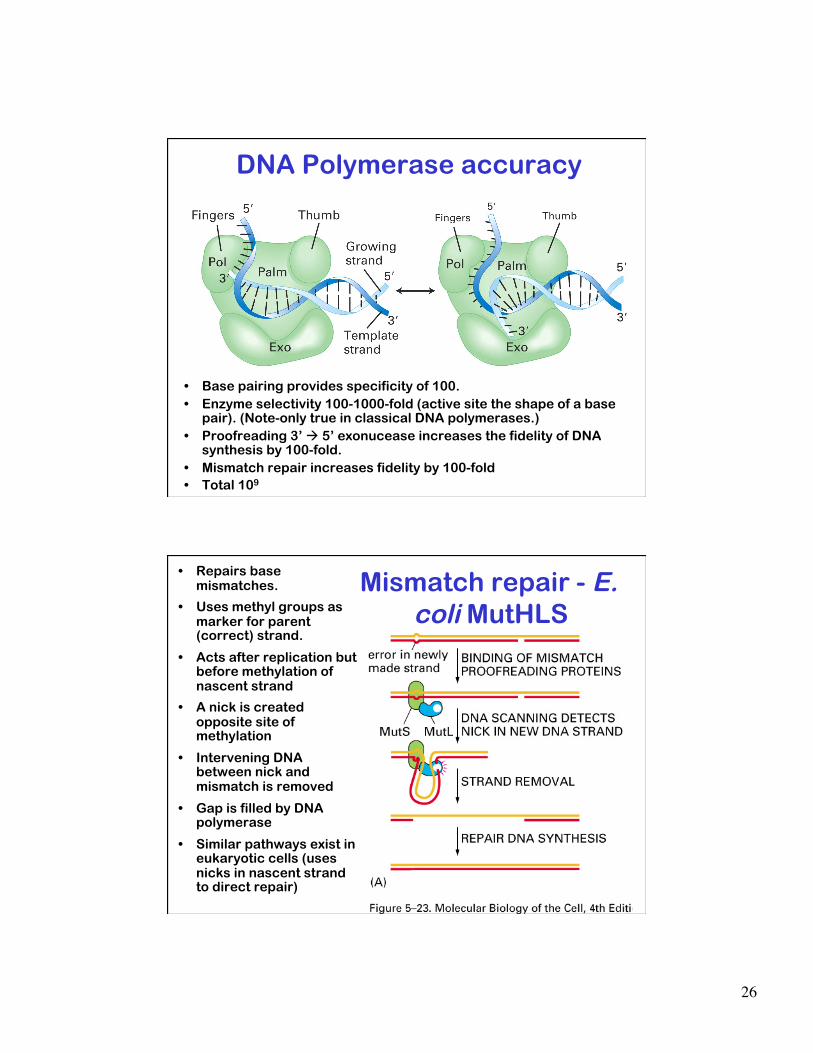
DNA Polymerase accuracy
• Base pairing provides specificity of 100. • Enzyme selectivity 100-1000-fold (active site the shape of a base
pair). (Note-only true in classical DNA polymerases.) • Proofreading 3’ 5’ exonucease increases the fidelity of DNA
synthesis by 100-fold. • Mismatch repair increases fidelity by 100-fold • Total 109
Mismatch repair - E. coli MutHLS
• Repairs base mismatches.
• Uses methyl groups as marker for parent (correct) strand.
• Acts after replication but before methylation of nascent strand
• A nick is created opposite site of methylation
• Intervening DNA between nick and mismatch is removed
• Gap is filled by DNA polymerase
• Similar pathways exist in eukaryotic cells (uses nicks in nascent strand to direct repair)

27
DNA metabolism and disease II: Human mismatch repair
Hereditary nonpolyposis colon cancer (HNPCC) – Accounts for 5% of colon cancer – Affected individuals have an 80% chance of developing cancer
during their life time – Caused by mutations in one of genes involved in mismatch repair.
(e.g hMLH1, hMSH2 or one of other mismatch repair proteins). – Autosomal dominant!
Defects in mismatch repair genes lead to human disease w
ww.
nitis
urgi
cal.c
om
Types of DNA Repair
• Replication fidelity and Post replicative repair – Proof-reading (during replication) – mismatch repair
• Damage Repair (anytime) – Nucleotide Excision – Base Excision – Direct reversal – Transcription Coupled – Specialized repair (inter-strand cross links, specialized
damage)
• Translesion Synthesis & Error-prone Repair (SOS) • Double-strand Break repair
– Nonhomologous End Joining (NHEJ) – Recombinational repair
DNA damage occurs as a result of environmental (chemical modification, radiation, etc.) insult, natural cellular processes (reactive oxygen species) or fundamental chemistry. All damage needs to be repaired.

28
Bulky adducts Modifications that disrupt
the DNA duplex are called bulky adducts.
Caused by:
• Ultraviolet light • Chemical modifications
(alkalytion) • Some types of oxidative
damage
UV- causes interbase crosslinks: pyrimidine (cyclobutane) dimers.
Nucleotide Excision Repair • Nucleotide excison repair (NER) is the major pathway for
repairing damaged bases (pyrimidine dimers, alkylated DNA and other bulky adducts).
• NER protein complex (called Excinuclease) binds to site of damage and cuts damaged strand on either side creating a small gap
• Defects in this pathway result in the disease Xeroderma Pigmentosum

29
Clinical Correlation Xeroderma pigmentosum (Groups
A-G) • Rare genetic disorder that is
caused by mutations in proteins involved in NER
• Patients have thin, unevenly pigmented skin. Also causes spidery blood vessels in the skin (telangiectasia). Very high sensitivity to sunlight and high incidence of cancer.
• Patients often develop skin cancer before age of five
• Very rare: ~2/million live births in Western Europe (Kleijer et al 2008, DNA Repair 7,744).
Base Excision Repair • Base excision repair (BER) removes modified or damaged bases
including alkylated DNA, oxidative damage, some bulky adducts. • Glycosydic bonds linking the damaged base to the DNA are cleaved
by specific glycosylases. • The Abasic site (deoxyribose with no base attached) is recognized
by an AP-endonuclease which nicks the damaged strand.
• Nuclease & polymerase synthesize across AP site.
• Nick sealed by DNA ligase.
30
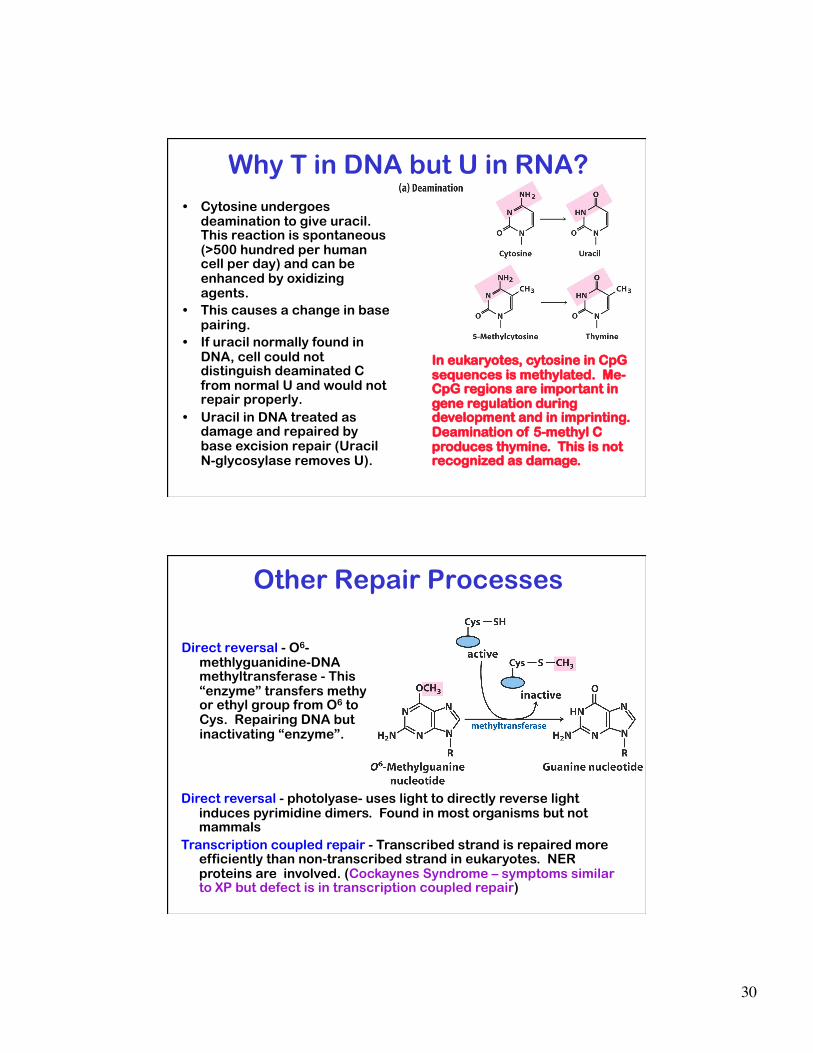
In eukaryotes, cytosine in CpG sequences is methylated. Me-CpG regions are important in gene regulation during development and in imprinting. Deamination of 5-methyl C produces thymine. This is not recognized as damage.
Why T in DNA but U in RNA? • Cytosine undergoes
deamination to give uracil. This reaction is spontaneous (>500 hundred per human cell per day) and can be enhanced by oxidizing agents.
• This causes a change in base pairing.
• If uracil normally found in DNA, cell could not distinguish deaminated C from normal U and would not repair properly.
• Uracil in DNA treated as damage and repaired by base excision repair (Uracil N-glycosylase removes U).
Other Repair Processes
Direct reversal - O6-methlyguanidine-DNA methyltransferase - This “enzyme” transfers methy or ethyl group from O6 to Cys. Repairing DNA but inactivating “enzyme”.
Direct reversal - photolyase- uses light to directly reverse light induces pyrimidine dimers. Found in most organisms but not mammals
Transcription coupled repair - Transcribed strand is repaired more efficiently than non-transcribed strand in eukaryotes. NER proteins are involved. (Cockaynes Syndrome – symptoms similar to XP but defect is in transcription coupled repair)

31
Inter-strand cross-links • Repair of cross-linked DNA remains poorly understood.
– Repair requires other repair repair proteins (NER, HR) and an additional set of proteins
• Fanconi Anemia is caused by a defect in proteins involved in repair of inter-strand cross-links. (Fanconi patients have normal sensitivity to most DNA damaging agents but are very sensitive to cross-linking agents.)
• Fanconi Anemia is a form of aplastic anemia. Patients also develop acute myeloid leukemia (AML) and carcinomas at elevated rates
• Recessive mutation in one of ~15 genes (includes Fanconi genes and BRCA2).
• Estimated 1-300 in US is a carrier. Incidence 1~360,000
Clinical correlation • Cisplatin (Platinol-AQ®) –
widely used in chemotherapy. Forms DNA cross-link. Dr. Carol Wynman, Erasmus Medical Center
Types of DNA Repair
• Replication fidelity and Post replicative repair – Proof-reading (during replication) – mismatch repair
• Damage Repair (anytime) – Nucleotide Excision – Base Excision – Direct reversal – Transcription Coupled – Specialized repair (interstrand cross links, specialized
damage)
• Tranlesion Synthesis & Error-prone Repair (SOS) • Double-strand Break repair (during replication)
– Nonhomologous End Joining (NHEJ) – Recombinational repair
DNA damage occurs as a result of environmental (chemical modification, radiation, etc.) insult, natural cellular processes (reactive oxygen species) or fundamental chemistry. All damage needs to be repaired.

32
Error Prone Repair • When the replication fork encounters a unrepaired lesion or a
strand break, the fork stalls. Specialized repair processes (translesion synthesis, recombinational repair, fork restart) allow synthesis to continue.
• Pathway requires translesion DNA polymerases.
• Translesion synthesis - Translesion DNA polymerases incorporate nucleotides opposite damage allowing classical polymerases to continue synthesis.
Error-prone repair - Translesion synthesis
Strand break
Future of Medicine: Repair vs. mutagenesis
• Mutations in translesion polymerases affect mutation rate
• XP-variant (XP-V) have mutations in DNA polymerase eta (η) and increased rate of mutations. This is caused by cells inability to carry out DNA replication in the presence of DNA damage
• Could cells be made less sensitive to mutagens by modulating the activities of classical and translesion DNA polymerases?
M.T. Washington, U of Iowa

33
Types of DNA Repair
• Replication fidelity and Post replicative repair – Proof-reading (during replication) – mismatch repair
• Damage Repair (anytime) – Nucleotide Excision – Base Excision – Direct reversal – Transcription Coupled – Specialized repair (interstrand cross links, specialized
damage)
• Tranlesion Synthesis & Error-prone Repair (SOS) • Double-strand Break repair
– Nonhomologous End Joining (NHEJ) (any time) – Recombinational repair (after replication)
DNA damage occurs as a result of environmental (chemical modification, radiation, etc.) insult, natural cellular processes (reactive oxygen species) or fundamental chemistry. All damage needs to be repaired.
There are two general pathways for repairing ds-DNA breaks: • Non-homologous end-joining: repair with loss of several bases.
(Predominant pathway in mammalian cells) • Homologous end-joining or recombination: completely restores
sequence from second copy of DNA. (Predominant pathway in bacteria and unicellular eukaryotes.)
• Both processes involve MRN complex (MRE11, RAD50, NBS1)
Double-strand break
repair

34
Non-homologous End Joining
• Proteins required non-homologous end joining include: end recognition (Ku, DNA-PK ) and processing nucleases (Rad50, MRE11, NBS1, and ligase).
• Some DNA is lost at the junction
Recombinational ds-break repair • Recombinational repair
requires resection of the DNA ends and then general recombination
• Note involvement of ATM, NBS1, BRCA and MRE proteins
General recombination

35
• Recombination initiates from either a double-stranded or single-stranded break.
• Strand invasion leads to the formation of joint molecule - Holliday junction.
Homologous recombination
• Strand invasion leads to the formation of joint molecule - Holliday junction.
• Migration and resolution of Holliday junction leads to exchange of DNA sequences.
• Catalyzed by recA in bacteria, Rad51 in eukaryotes
• Resolution of Holliday junction results in: – Repaired break – Recombined DNA
Homologous recombination
2 Holliday Junctions
36
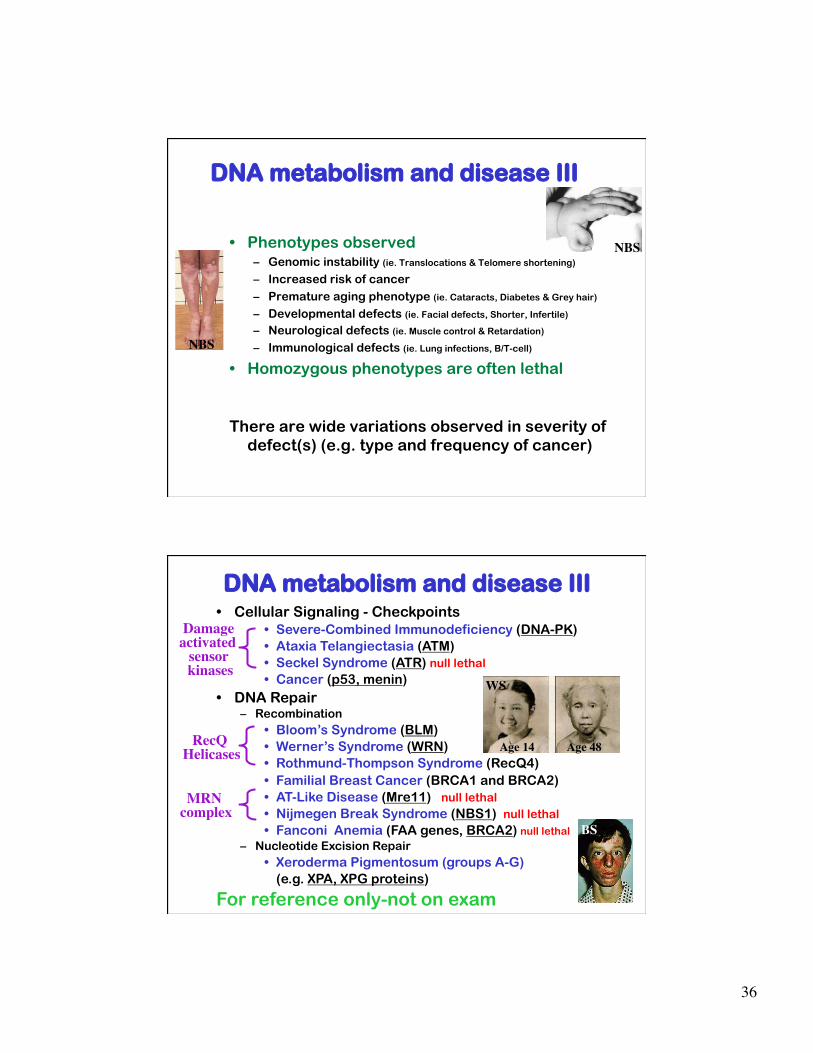
DNA metabolism and disease III
• Phenotypes observed – Genomic instability (ie. Translocations & Telomere shortening) – Increased risk of cancer – Premature aging phenotype (ie. Cataracts, Diabetes & Grey hair)
– Developmental defects (ie. Facial defects, Shorter, Infertile)
– Neurological defects (ie. Muscle control & Retardation) – Immunological defects (ie. Lung infections, B/T-cell)
• Homozygous phenotypes are often lethal
There are wide variations observed in severity of defect(s) (e.g. type and frequency of cancer)
NBS
NBS
DNA metabolism and disease III • Cellular Signaling - Checkpoints
• Severe-Combined Immunodeficiency (DNA-PK) • Ataxia Telangiectasia (ATM) • Seckel Syndrome (ATR) null lethal • Cancer (p53, menin)
• DNA Repair – Recombination
• Bloom’s Syndrome (BLM) • Werner’s Syndrome (WRN) • Rothmund-Thompson Syndrome (RecQ4) • Familial Breast Cancer (BRCA1 and BRCA2)
• AT-Like Disease (Mre11) null lethal • Nijmegen Break Syndrome (NBS1) null lethal • Fanconi Anemia (FAA genes, BRCA2) null lethal
– Nucleotide Excision Repair • Xeroderma Pigmentosum (groups A-G)
(e.g. XPA, XPG proteins)
For reference only-not on exam
Age 14 Age 48
WS
BS MRN
complex
RecQ Helicases
Damage activated
sensor kinases