domain structure of synaptotagmin (~65)” · homologous to protein kinase c. sucrose density gra-...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1991 by The American Society for Biochemistry and Molecular Biology, Inc.
Domain Structure of Synaptotagmin (~65)”
Vol. 266, No. 1, Issue of January 5, pp. 623-629,1991 Printed in U. S. A.
(Received for publication, July 13, 1990)
Mark S. Perin$$, Nils BroseT, Reinhard Jahnll, and Thomas C. Sudhof$II** From the $Howard Hughes Medical Institute and the 11 Department of Molecular Genetics, the University of Texas Southwestern Medical Center, Dallas. Texas 75235 and the llMax-Planck-Institute fur Psychiatrie, Planegg-Martinsried, Federal Republic of Germany
Synaptotagmin (p65) is an abundant and evolution- arily conserved protein of synaptic vesicles that con- tains two copies of an internal repeat homologous to the regulatory region of protein kinase C. In the cur- rent study, we have investigated the biochemical prop- erties of synaptotagmin, demonstrating that it contains five protein domains: an intravesicular amino-termi- nal domain that is glycosylated but lacks a cleavable signal sequence; a single transmembrane region; a se- quence separating the transmembrane region from the two repeats homologous to protein kinase C; the two protein kinase C-homologous repeats; and a conserved carboxyl-terminal sequence following the two repeats homologous to protein kinase C. Sucrose density gra- dient centrifugations and gel electrophoresis indicate that synaptotagmin monomers associate into dimers and are part of a larger molecular weight complex. A sequence predicted to form an amphipathic a-helix that may cause the stable dimerization of synaptotagmin is found in its third domain between the transmembrane region and the protein kinase C-homologous repeats. Synaptotagmin contains a single hypersensitive pro- teolytic site that is located immediately amino-termi- nal to the amphipathic a-helix, suggesting that synap- totagmin contains a particularly exposed region as the peptide backbone emerges from the dimer. Finally, subcellular fractionation and antibody bead purifica- tion demonstrate that synaptotagmin co-purifies with synaptophysin and other synaptic vesicle markers in brain. However, in the adrenal medulla, synaptotag- min was found in both synaptophysin-containing mi- crovesicles and in chromaffin granules that are devoid of synaptophysin, suggesting a shared role for synap- totagmin in the exocytosis of small synaptic vesicles and large dense core catecholaminergic vesicles.
Synaptotagmin (p65) is an abundant integral membrane protein of synaptic vesicles whose primary structure contains two copies of a repeat homologous to the regulatory region of protein kinase C (1 ,2) . In the preceding study, synaptotagmin was shown to be structurally and functionally conserved from Drosophila to humans (3). These experiments demonstrated that the protein kinase C-homologous repeats of synaptotag- min are conserved between vertebrates and invertebrates and that these repeats most likely mediate the ability of recom-
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
3 Present address: Division of Neuroscience, Baylor College of Medicine, Houston, TX 77030.
** TO whom correspondence and reprint requests should be ad- dressed.
binant synaptotagmin from humans, rats, and Drosophila to bind to phosphatidylserine (3).
The unique structural and functional properties of synap- totagmin suggest a central role of the protein in the exocytosis of synaptic vesicles. Therefore, the elucidation of its subcel- lular distribution with respect to small synaptic vesicles and large dense core vesicles and the determination of the trans- membrane orientation, glycosylation, and other structural features of synaptotagmin is of great importance.
In order to study the biochemical properties of synaptotag- min, we have now raised antibodies against five different epitopes of the rat protein. In addition, the 5’ end of the synaptotagmin message was mapped to ensure the validity of the proposed amino terminus of synaptotagmin. Sucrose den- sity gradient centrifugations demonstrate that synaptotagmin is part of a high molecular weight complex in the synaptic vesicle membrane that contains synaptotagmin dimers as its basic unit. Our results demonstrate that rat synaptotagmin is a glycoprotein that contains an amino-terminal intravesicular sequence which is translocated without a cleaved signal se- quence. We suggest that synaptotagmin dimerizes via an amphipathic a-helix that is located between the transmem- brane region and the internal repeats homologous to protein kinase C. A single hypersensitive proteolytic site is found in synaptotagmin that maps to the amino-terminal end of the amphipathic a-helix, compatible with the notion that the synaptotagmin peptide backbone is particularly exposed at this point because it emerges from a rigidly interacting do- main. Together, these results lead to a five-domain model of the structure of synaptotagmin in which the last two domains, the carboxyl terminus and the protein kinase C-homologous repeats, are the conserved and presumably functionally active parts of the protein.
EXPERIMENTAL PROCEDURES
Materials-Reverse transcriptase was obtained from Life Sciences, and proteases and endoglycosidase F from Boehringer Mannheim. Restriction enzymes, DNA modifying enzymes, and DNA molecular weight markers were purchased from New England Biolabs. CHAPS’ and Zwittergent 3-10 were obtained from Calbiochem. RNA molecular weight markers were from Bethesda Research Laboratories, and reagents for SDS-PAGE including protein molecular weight markers and protein assay components were from Bio-Rad. Antibodies against synaptophysin, synaptobrevin, rab3, and cytochrome bSG1 were de- scribed previously (4-7). The antibody against the 116,000 subunit of the proton pump was raised against a synthetic peptide corresponding to the carboxyl-terminal 18 amino acids of the subunit.’ Peroxidase- labeled secondary antibodies were obtained from Cappel. Eupergit C1Z methacrylate Microbeads were purchased from Rohm Pharma.
Production of Synaptotagmin Antibodies-Three antibodies were
’ The abbreviations used are: CHAPS, 3-[(3-~holamidopropyl)- dimethylammonio]-1-propanesulfonic acid; SDS-PAGE, sodium do- decyl sulfate-polyacrylamide gel electrophoresis.
* T. C. Siidhof and M. S. Perin, unpublished observation.
623

624 Structure of Synaptotagmin
raised against synthetic peptides coupled to keyhole limpet hemocy- anin as described (4). The synthetic peptides had the following sequences (residue numbers as in (2), the topography of synaptotag- min is shown in Fig. 1 for identification of the domains): CMVSASH- PEALA (residues 1 to 11, amino terminus), CAINMKDVKDLG- KTMKDQALKD (residues 100 to 120, between transmembrane re- gion and A repeat), and CMDVGGLSDPYVKIHL (residues 302 to 316, B repeat). The amino-terminal cysteine in each peptide is not present in the protein sequence and was introduced to allow efficient coupling. In addition to these three antipeptide antibodies, antibodies were raised against two bacterial recombinant proteins incorporating sequences comprised of residues 78 to 421 (complete sequence car- boxyl-terminal to the transmembrane region) or residues 265 to 381 (B repeat). The production of the recombinant proteins was as described previously (2, 3, 8), and they were used for immunization after partial purification of inclusion bodies containing recombinant protein.
Subcellular Fractionation and Immunobead Purification-Purifi- cation of rat brain cortex synaptic vesicles with controlled pore-glass chromatography as the final step was performed as described previ- ously (9, 10). For most biochemical experiments, vesicles of lesser purity obtained before the controlled pore-glass chromatography were used. Subfractionation of bovine adrenal medulla via centrifugation through a 1.6 M sucrose step gradient for the purification of chro- maffin granules was performed as described (11). Immunobead puri- fication of synaptophysin-containing vesicles was performed as de- scribed using Eupergit C1Z methacrylate Microbeads coated with a purified monoclonal antibody against synaptophysin (12).
Protease and Endoglycosidase F Digestion of Synuptotagmin-syn- aptic vesicles (75 pg of protein) were digested with different concen- trations of pronase or trypsin in 0.13 M NaCl, 20 mM HEPES-NaOH, pH 7.4, for 30 min at 37 "C. Reactions were stopped by boiling in electrophoresis sample buffer (62 mM Tris-HC1, pH 6.8, 5% 2-mer- captoethanol, 2% SDS). Endoglycosidase F digestions were performed in 0.13 M NaCl, 20 mM HEPES-NaOH, pH 7.4, 1% CHAPS, 10 mM 2-mercaptoethanol, and 0.2 unit of endoglycosidase F (13). In the experiments in which synaptic vesicles were subjected both to endo- glycosidase F and protease digestion, synaptic vesicles were first digested with trypsin as described above. The reaction was stopped with 1 mg/ml N-tosyl-L-lysine chloromethyl ketone, and CHAPS and 2-mercaptoethanol were added to 1% and 10 mM, respectively. 0.2 unit of endoglycosidase F was added, and the mixture was incubated overnight at 37 'C.
SDS-PAGE and immunoblotting were performed as described (4, 7, 14, 15). Proteins reactive with antibodies were visualized with peroxidase-labeled secondary antibodies. Protein assays were per- formed according to Bradford (16).
Sucrose Gradient Centrifugations-Synaptic vesicles (1.5 mg of protein) were solubilized in 0.1 M NaCl, 50 mM HEPES-NaOH, pH 7.4, 1.8% CHAPS, or Zwittergent 3-10 by stirring for 1 h on ice, and insoluble residue was removed by centrifugation (150,000 X gmex for 30 min). The supernatant was loaded on 5-20% linear sucrose gra- dients containing 0.9% CHAPS or Zwittergent 3-10 and 50 mM HEPES-NaOH, pH 7.4, and centrifuged for 16 h at 210,000 X gmax. Gradient fractions were analyzed by immunoblotting for synaptophy- sin, synaptobrevin, synaptotagmin, and the M, = 116,000 k proton pump subunit. In separate, identically performed sucrose gradients, bovine serum albumin (Mr = 65,000), alcohol dehydrogenase ( M , = 150,000), catalase (Mr = 250,000), and p-galactosidase (Mr = 520,000) were fractionated as molecular weight standards.
Primer extension analyses were performed as described (17) using three "P-labeled oligonucleotides complementary to the following cDNA sequences: oligonucleotide A, 898 to 918; oligonucleotide B, 615 to 641; oligonucleotide C, 390 to 424 (nucleotide numbers accord- ing to Ref 2). Reaction products were analyzed by PAGE followed by autoradiography of the dried gels.
RNA blotting experiments were performed by hybridizing 32P- labeled synthetic oligonucleotides (B and C as described above) to blots of poly(A')-enriched RNA from rat brain. Hybridization were performed at 42 "C and washed at 50 "C as described (18,19).
RESULTS
Antibodies against Different Domains of Synaptotagmin- The primary structures of the rat, human, and Drosophila synaptotagmins as deduced from their cDNA sequences pre- dict the biosynthesis of a highly conserved protein that con-
tains a single transmembrane region and two copies of a repeat homologous to the regulatory region of protein kinase C (3). The domain structure of rat synaptotagmin is compared to that of protein kinase C in Fig. 1, illustrating the localiza- tion of the internal repeats in synaptotagmin with respect to its transmembrane region and the similarity between repeats and protein kinase C.
In order to investigate the localization of synaptotagmin to synaptic vesicles and the domain structure of synaptotagmin, antibodies were raised against five different epitopes from rat synaptotagmin. Three of these were directed against synthetic peptides corresponding to the amino-terminal 11 amino acids of synaptotagmin, the sequence in the region separating the transmembrane region from the A repeat, and to a conserved sequence in the B repeat of synaptotagmin. The other two antibodies were raised against recombinant proteins contain- ing either the entire sequence of rat synaptotagmin carboxyl- terminal to the transmembrane region (presumably its cyto- plasmic domains) or the B repeat only. All five antibodies specifically and sensitively reacted with an M, = 65,000 pro- tein in rat brain, although with different affinities, suggesting that they all recognized synaptotagmin (data not shown).
In a comparison of different species, the antibody against the amino-terminal peptide was specific for rat synaptotagmin and did not recognize bovine synaptotagmin (data not shown). The greatest sequence differences between human and rat synaptotagmin are found at the amino terminus (3), suggest- ing that sequence differences between bovine and rat proteins at the amino terminus may also be responsible for a lack of cross-speciescross-reactivity of the amino-terminal antibody. All other antibodies reacted with bovine synaptotagmin, but none reacted with Drosophila synaptotagmin (3).
Subcellular Localization of Synaptotagmin in Rat Brain and Bovine Adrenal Medulla-Using these antibodies, we exam- ined whether synaptotagmin is highly enriched in synaptic vesicles or if it is shared with other organelles. For this purpose, rat brain homogenates were depleted of mitochondria and nuclei by low speed centrifugation (25,000 X gmaX for 20 min). Synaptic vesicles were immunoprecipitated from the resulting supernatant using beads coated with a monoclonal antibody against synaptophysin, a well characterized synaptic vesicle-specific protein in brain (4, 20, 21). Synaptotagmin was quantitatively co-immunoprecipitated with synaptophy- sin from the low speed supernatant by these antibody beads (Fig. 2). No synaptotagmin remained in the supernatant after synaptophysin-containing membranes had been removed, suggesting that in brain little synaptotagmin is localized in membranes other than synaptic vesicles.
In addition to antibody bead precipitation, the distribution of synaptotagmin on synaptic vesicles during the classical synaptic vesicle purification scheme with controlled pore- glass chromatography as the last step (9, 10) was studied and compared with that of synaptophysin, synaptobrevin (5), and rab3A (6). Again we observed complete co-purification of all of these synaptic vesicle markers with synaptotagmin (data not shown). These results suggest that, like synaptophysin, synaptobrevin, and rab3, synaptotagmin is highly enriched on synaptic vesicles in brain.
Several proteins such as synaptophysin and rab3 are exclu- sively localized to synaptic vesicles in brain but are also expressed in the adrenal medulla (6, 21, 22). Here, synapto- physin, synaptobrevin, and rab3 are found on synaptic-like microvesicles that are distinct from the secretory granules in these cells (6, 22). We therefore studied the subcellular local- ization of synaptotagmin in the bovine adrenal medulla using centrifugation and antibody bead precipitation. Total med-
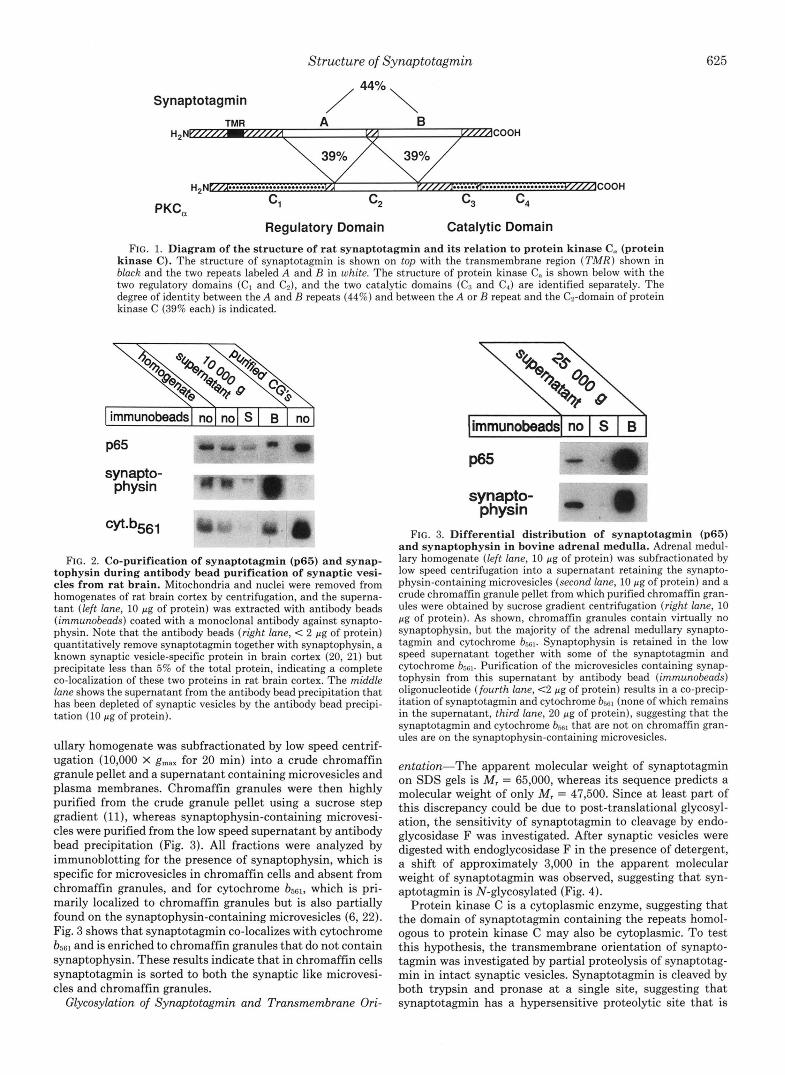
Structure of Synaptotagmin 625
Synaptotagmin / 44%\
TMR A B
COOH
PKC, Regulatory Domain Catalytic Domain
FIG. 1. Diagram of the structure of rat synaptotagmin and its relation to protein kinase C, (protein kinase C). The structure of synaptotagmin is shown on top with the transmembrane region (TMR) shown in black and the two repeats labeled A and B in white. The structure of protein kinase C. is shown below with the two regulatory domains (C1 and C2), and the two catalytic domains (Cs and C,) are identified separately. The degree of identity between the A and B repeats (44%) and between the A or B repeat and the C2-domain of protein kinase C (39% each) is indicated.
\ \ 1 immunobeadsl no] no] S
synapto- physin
'yfSb561
FIG. 2. Co-purification of synaptotagmin (p65) and synap- tophysin during antibody bead purification of synaptic vesi- cles from rat brain. Mitochondria and nuclei were removed from homogenates of rat brain cortex by centrifugation, and the superna- tant (left lane, 10 pg of protein) was extracted with antibody beads (immunobeads) coated with a monoclonal antibody against synapto- physin. Note that the antibody beads (right lane, < 2 pg of protein) quantitatively remove synaptotagmin together with synaptophysin, a known synaptic vesicle-specific protein in brain cortex (20, 21) but precipitate less than 5% of the total protein, indicating a complete co-localization of these two proteins in rat brain cortex. The middle lane shows the supernatant from the antibody bead precipitation that has been depleted of synaptic vesicles by the antibody bead precipi- tation (10 pg of protein).
ullary homogenate was subfractionated by low speed centrif- ugation (10,000 X gmaX for 20 min) into a crude chromaffin granule pellet and a supernatant containing microvesicles and plasma membranes. Chromaffin granules were then highly purified from the crude granule pellet using a sucrose step gradient (ll), whereas synaptophysin-containing microvesi- cles were purified from the low speed supernatant by antibody bead precipitation (Fig. 3). All fractions were analyzed by immunoblotting for the presence of synaptophysin, which is specific for microvesicles in chromaffin cells and absent from chromaffin granules, and for cytochrome b561, which is pri- marily localized to chromaffin granules but is also partially found on the synaptophysin-containing microvesicles (6, 22). Fig. 3 shows that synaptotagmin co-localizes with cytochrome bs61 and is enriched to chromaffin granules that do not contain synaptophysin. These results indicate that in chromaffin cells synaptotagmin is sorted to both the synaptic like microvesi- cles and chromaffin granules.
Glycosylation of Synaptotagmin and Transmembrane Ori-
limmunobeadsl no I S I B I
synapto- physin
FIG. 3. Differential distribution of synaptotagmin (p65) and synaptophysin in bovine adrenal medulla. Adrenal medul- lary homogenate (left lane, 10 pg of protein) was subfractionated by low speed centrifugation into a supernatant retaining the synapto- physin-containing microvesicles (second lane, 10 pg of protein) and a crude chromaffin granule pellet from which purified chromaffin gran- ules were obtained by sucrose gradient centrifugation (right lane, 10 pg of protein). As shown, chromaffin granules contain virtually no synaptophysin, but the majority of the adrenal medullary synapto- tagmin and cytochrome b561. Synaptophysin is retained in the low speed supernatant together with some of the synaptotagmin and cytochrome b561. Purification of the microvesicles containing synap- tophysin from this supernatant by antibody bead (immunobeads) oligonucleotide (fourth lane, <2 pg of protein) results in a co-precip- itation of synaptotagmin and cytochrome bSs1 (none of which remains in the supernatant, third lane, 20 pg of protein), suggesting that the synaptotagmin and cytochrome b561 that are not on chromaffin gran- ules are on the synaptophysin-containing microvesicles.
entution-The apparent molecular weight of synaptotagmin on SDS gels is M , = 65,000, whereas its sequence predicts a molecular weight of only M, = 47,500. Since a t least part of this discrepancy could be due to post-translational glycosyl- ation, the sensitivity of synaptotagmin to cleavage by endo- glycosidase F was investigated. After synaptic vesicles were digested with endoglycosidase F in the presence of detergent, a shift of approximately 3,000 in the apparent molecular weight of synaptotagmin was observed, suggesting that syn- aptotagmin is N-glycosylated (Fig. 4).
Protein kinase C is a cytoplasmic enzyme, suggesting that the domain of synaptotagmin containing the repeats homol- ogous to protein kinase C may also be cytoplasmic. To test this hypothesis, the transmembrane orientation of synapto- tagmin was investigated by partial proteolysis of synaptotag- min in intact synaptic vesicles. Synaptotagmin is cleaved by both trypsin and pronase a t a single site, suggesting that synaptotagmin has a hypersensitive proteolytic site that is

626 Structure of Synaptotagmin
Endo F - + dimer- --
monomer- \ -84 y
-33 L
-24 - 16
-47 9 x
FIG. 4. Endoglycosidase F digestion of synaptotagmin. The apparent molecular weight of synaptotagmin was analyzed by SDS- PAGE and immunoblotting (75 pg of protein/lane) before or after endoglycosidase F digestion of synaptic vesicles. Numbers on the right indicate positions of molecular weight markers. Both the lower and upper immunoreactive band of M, = 65,000 and 130,000 are sensitive to endoglycosidase F and both are recognized by five different anti- bodies to synaptotagmin (data not shown), suggesting that they represent synaptotagmin monomers and dimers.
- 39 x - 27 $ - 17
FIG. 5. Presence of a hypersensitive proteolytic site in syn- aptotagmin. Intact synaptic vesicles (75 pg of protein) were digested with the indicated amounts of pronase or trypsin for 1 h at 37 "C. Reaction products were analyzed by SDS-PAGE and immunoblotting using the antibody against the complete cytoplasmic domain contain- ing both C2-repeats. Numbers on the right show positions of molecular weight markers.
equally cleaved by these two proteases despite their different substrate specificities (Fig. 5). Using the antibody directed against the carboxyl-terminal domain of synaptotagmin, the same proteolytic product of approximately M , = 39,000 was detected over a 30-fold concentration range of both proteases.
The proteolytic fragment produced by mild trypsin or pro- nase treatment of synaptotagmin was soluble, whereas intact synaptotagmin was quantitatively pelleted with synaptic ves- icles (data not shown). Higher protease concentrations led to a complete digestion of the carboxyl-terminal synaptotagmin epitope in intact synaptic vesicles. When proteolysis of syn- aptophysin was investigated under similar conditions, it was left intact except at the highest protease concentrations, and then only the cytoplasmic sequences were affected (4).
The release of a soluble carboxyl-terminal synaptotagmin fragment from intact synaptic vesicles after mild proteolysis suggests that the carboxyl-terminal sequences of synaptotag- min containing the homologies to protein kinase C are indeed cytoplasmic, and that a membrane-bound amino-terminal fragment should remain. To test this hypothesis, untreated and mildly proteolyzed synaptic vesicles were analyzed by immunoblotting using both the antibody against the carboxyl- terminal Cn-domains and an antibody raised against the 11 amino-terminal residues of synaptotagmin. As shown in Fig. 6, both antibodies recognized the same protein in intact synaptic vesicles. However, after partial proteolysis, the amino-terminal antipeptide antibody now labeled an M , = 28,000 band as opposed to the M , = 39,000 fragment detected by the antibody against the cytoplasmic epitope. Furthermore, when synaptic vesicles were subjected to endoglycosidase F digestion after proteolysis, only the smaller amino-terminal fragment changed in electrophoretic mobility (Fig. 6). These results suggest that synaptotagmin contains a small intrave- sicular amino-terminal and large cytoplasmic carboxyl-ter-
NH,-Terminus C, -Domains - I + I + - I r l - l I - I L I - 1 1
-1 30
- 50 9 - 39 L
- 27
- 17
- 75?
X
FIG. 6. Analysis of the endoglycosidase F sensitivity of the amino- and carboxyl-terminal proteolytic fragments of syn- aptotagmin. Synaptotagmin was cleaved at its hypersensitive pro- teolytic site by trypsin. Both intact and cleaved synaptotagmin were then subjected to endoglycosidase F digestion, and the different samples were analyzed by SDS-PAGE (75 pg of protein/lane) and immunoblotting using an antibody against the 11 amino-terminal amino acids (left panel) or against a recombinant protein consisting of the complete cytoplasmic domains of synaptotagmin (right panel). Whereas intact synaptotagmin is endoglycosidase F-sensitive, only the amino-terminal fragment produced by proteolytic cleavage and not the carboxyl-terminal fragment is endoglycosidase F-sensitive.
A B C A B C -,,o - ~p I - "84,
9 -47 ' -33 x - 24 z~ -16
FIG. 7. Mapping of the hypersensitive proteolytic site in synaptotagmin by an analysis of the size and immunoreactiv- i ty of its carboxyl-terminal proteolytic fragment. Untreated rat brain synaptic vesicles (lane A, 75 pg of protein/lane), trypsinized synaptic vesicles (lane B, 75 pg of protein/lane), and the recombinant cytoplasmic domain of synaptotagmin (residues 78 to 421, lane C; 3 pg of protein/lane) were subjected to SDS-PAGE and immunoblot- ting. The left blot was probed with the antibody against the recombi- nant protein in lane C, and the right blot with the antibody against a peptide corresponding to residues 100 to 120. Note that the latter antibody only weakly recognizes the proteolytic fragment and that the proteolytic fragment is approximately 5 kDa smaller than the recombinant protein, suggesting that its amino terminus is approxi- mately localized between residues 90 and 110.
minal sequence. The amino-terminal fragment is N-glycosy- late& inspection of the amino terminus of the synaptotagmin sequence reveals the presence of a single N-linked glycosyla- tion site a t residue 24 (2).
To localize the hypersensitive proteolytic site in the syn- aptotagmin sequence, we studied which of the four antibodies that recognize epitopes carboxyl-terminal to the transmem- brane region reacted with the cytoplasmic proteolytic frag- ment of synaptotagmin. In addition, its size was compared to that of recombinant synaptotagmin containing all sequences carboxyl-terminal to the transmembrane region (Fig. 7). The proteolytic fragment was found to be slightly smaller than the recombinant fragment whose amino terminus corresponds to residue 78 and to react with all antibodies against cytoplasmic sequences of synaptotagmin. The weakest response was ob- tained with the antibody against a synthetic peptide corre- sponding to residues 100 to 120, suggesting that the proteol- ysis affects this epitope. These results indicate that the hy-

Structure of Synaptotagmin 627
persensitive proteolytic site is localized between residues 90 and 100 of synaptotagmin.
Mapping of the 5' End of the Synaptotagmin Message-The discrepancy between the apparent molecular weight of syn- aptotagmin as determined by SDS-PAGE and its predicted size cannot be entirely accounted for by N-linked glycosyla- tion since endoglycosidase F treatment of synaptotagmin causes only a small molecular weight shift (Figs. 4 and 6). There are several plausible explanations for the apparent high molecular weight of synaptotagmin. I t may migrate a t a higher molecular weight on SDS-PAGE because it contains addi- tional modifications such as 0-linked sugars, or it may have unusual secondary structures, or the predicted amino termi- nus of synaptotagmin maybe incorrect due to cloning artifacts. We therefore performed a series of experiments to rule out the possibility of cloning artifacts.
The finding that an antibody against the 11 amino-terminal residues of synaptotagmin reacts with the protein in synaptic vesicles demonstrates that these 11 amino acids are present in the mature protein. This suggests that a possible cloning artifact must be localized to the 5' untranslated region. In order to test if the long 5' untranslated region found by cDNA cloning is correct, blots of poly(A+)-enriched RNA from rat brain were hybridized with oligonucleotides from the 5' un- translated region and the coding region. Single messages of similar size (approximately 4.8 kilobases) were specifically hybridized with both probes (Fig. 8).
To investigate the validity of the cDNA cloning further, the 5' end of the rat synaptotagmin message was analyzed by primer extensions. Three different oligonucleotides were used that were complementary to either the 5' untranslated region or the 5' end of the coding region. With all three primers, specific extension products were observed with sizes as pre- dicted by the cDNA sequence (Fig. 9). This result suggests that the 5' untranslated region of the cDNAs is correct and virtually full length and that no 5' coding region was missed due to a cloning artifact. Since the 5' untranslated region of synaptotagmin contains multiple stop codons in all three reading frames, it seems likely that the anomalously high apparent molecular weight of synaptotagmin is due to an intrinsic property of the protein.
Synaptotagmin Is Part of a High Molecular Weight Com- plex-Analysis of synaptotagmin by SDS-PAGE often re- sulted in the specific immunolabeling of a M, = 130,000 protein in addition to the monomeric M, = 65,000 protein. Both the M, = 65,000 and 130,000 bands were similarly sensitive to endoglycosidase F digestion (Fig. 4). Both bands were also recognized by all five synaptotagmin antibodies
A B Vr- -v
-9.5 kb -7.5 kb
'q -4.4kb
-2.4 kb
- 1.4 kb
FIG. 8. RNA blot analysis of the synaptotagmin message in rat brain using an oligonucleotide complementary to the 5' untranslated region (lane A ) and to the coding region of the message predicted from the cDNA sequence (lane B ) . Oligo- nucleotides are the same as in Fig. 9. The numbers on the right indicate positions of molecular weight standards.
A. E. C.
- -404 -309 . - .
-309 c t
0
FIG. 9. Primer extension analysis of the 5' end of the rat brain synaptotagmin message using three different oligonu- cleotides. The positions of the three oligonucleotides complementary to different parts of the cDNA are diagrammed at the bottom. On top, the primer extension products obtained with the three different oligonucleotides were resolved on three gels containing 4.5% ( A ) , 10% ( B ) , and 10% ( C ) acrylamide. Positions of co-electrophoresed radiolabeled molecular weight markers (MspI-cut-pBR322) are shown on the right of each extension product, and the size of each product is given on the left. The size of the longest 920-base pair (bp) product obtained with oligonucleotide A was determined by larger molecular weight markers (HueIII-cut 6x174 RF DNA) in addition to those shown here.
(data not shown), suggesting that the M, = 130,000 band represents a synaptotagmin dimer that is partially resistant to SDS denaturation. To further investigate the possible association of synaptotagmin into dimers and high molecular weight complexes, synaptic vesicle proteins solubilized in CHAPS or in Zwittergent 3-10 were fractionated by sucrose density gradient centrifugation. The sucrose gradient frac- tions were then analyzed for synaptotagmin, synaptophysin, and the 116,000 subunit of the synaptic vesicle proton pump by immunoblotting (Fig. 10).
On sucrose density gradients, synaptotagmin migrates in a single peak as a high molecular weight complex of approxi- mately M, = 220,000 in the presence of CHAPS, a nondena- turing detergent. Its molecular weight is higher than that of synaptophysin, which migrates as a broad peak comprised of dimers, trimers, and tetramers (23), and smaller than the proton pump complex (approximately M, = 530,000) (24). In Zwittergent 3-10, the proton pump is dissociated into its subunits and synaptotagmin co-migrates with the 116,000 subunit of the proton pump on the sucrose gradients. This suggests that Zwittergent 3-10 also partially dissociates the synaptotagmin complex but leaves synaptotagmin dimers in- tact (Fig. 10).
What is the mechanism of synaptotagmin dimerization? Analysis of the primary structure of synaptotagmin indicates the presence of a sequence that has a high potential of forming an amphipathic a-helix (shown in an a-helical wheel presen- tation in Fig. 11). Parallel strands of such an a-helix would not only be held together by their hydrophobic surface but would also form oppositely charged amino acid pairs along the helix, suggesting a mechanism by which stable dimers could be formed.
DISCUSSION
Synaptotagmin is an abundant synaptic vesicle membrane protein that contains two copies of an internal repeat homol- ogous to protein kinase C (2). Synaptotagmin is highly con- served evolutionarily from Drosophila to humans, and it ap-

628 Structure of Synaptotagmin
Fraction: 2 4 6 8 10 12 14 16 18 20
116 K CHAPS P 65
P 38
Fraction: 2 4 6 8 10 12 14 16 18 20 116 K
ZW. 3-10 P65
P 38 4
FIG. 10. Sucrose gradient fractionation of synaptic vesicle proteins solubilized in CHAPS (toppanel) or Zwittergent 3-10 (Zw. 3-10, bottom panel). Gradient fractions were analyzed by SDS-PAGE and immunoblotting, with the same blot being probed simultaneously for three synaptic vesicle proteins, the 116,000 proton pump subunit (116K), synaptotagmin (p65), and synaptophysin (p38). Synaptophysin migrates as a broad peak on sucrose gradients in both CHAPS and Zwittergent 3-10 representing an equilibrium of dimers, trimers, and tetramers (23). The high molecular weight proton pump complex (24) is maintained in CHAPS but dissociated into subunits in Zwittergent 3-10.
A. E.
FIG. 11. Helical wheel representation of rat synaptotagmin sequence (residues 102-119) with a high potential to form an amphipathic a-helix. In A , the respective amino acids are shown in single-letter code a t their predicted positions. In R, amino acids are represented by their physical characteristics to demonstrate the ex- posure of hydrophobic and charged amino acids on defined sides of the wheel. Closed circles are hydrophobic residues, open circles, neutral residues, and + and - are positively and negatively charged residues, respectively.
pears to bind negatively charged phospholipids with a speci- ficity dependent on both the acidic head group and hydrophobic diacyl backbone (2,3). These properties suggest a central role for synaptotagmin in synaptic vesicle function. Therefore, information on the biochemical properties of syn- aptotagmin in the synaptic vesicle membrane beyond its sequence is of great importance. In this study, we have raised a panel of polyclonal antibodies against different regions of synaptotagmin and used these to study the biochemical prop- erties and subcellular localization of synaptotagmin.
Our results demonstrate that synaptotagmin is a glycopro- tein that contains N-linked sugars. Synaptotagmin exhibits a single proteolytic hypersensitive site that was mapped be- tween residues 90 and 110 based on the sizes of the fragments produced after proteolytic cleavage and on their reactivity with different antibodies. Cleavage at the hypersensitive site results in the release of a soluble M , = 39,000 fragment that corresponds to the carboxyl-terminal two-thirds of the protein and contains the protein kinase C-homologous repeats. A smaller amino-terminal fragment remains associated with the membrane. The amino-terminal proteolytic fragment is sen- sitive to endoglycosidase F digestion, whereas the carboxyl- terminal fragment is not.
Together, these results suggest the carboxyl-terminal two- thirds of synaptotagmin including its protein kinase C-ho- mologous repeats are cytoplasmic and that its 52 amino-
terminal residues are intravesicular and N-glycosylated. This raises the question if synaptotagmin contains a cleavable signal sequence. The amino-terminal sequence of synaptotag- min is only remotely similar to a signal sequence (25) but exhibits sequence similarity to the amino terminus of the yeast a-1 mating factor precursor, a secreted eukaryotic pro- tein that also lacks a cleavable signal sequence (26) (sequences are given in single letter code with the residue numbers shown on the left and right):
* * * * * * * * Synaptotagmin: 9 A L A A P V - T T 16 Mating factor: 17 A L A A P V N T T 25
Since mature synaptotagmin reacts with an antibody raised against the 11 amino-terminal residues of the amino acid sequence predicted from the cDNA clones, the amino termi- nus of synaptotagmin does not contain a cleavable signal sequence.
Prompted by the observation that synaptotagmin often appeared to migrate as a dimer on SDS-PAGE, sucrose den- sity gradient centrifugations were performed in the presence of two different detergents to analyze the subunit structure of synaptotagmin. In the presence of CHAPS, a nondenatur- ing detergent, synaptotagmin migrated as a high molecular weight complex of approximately M , = 220,000. In Zwitter- gent 3-10, a detergent that leads to the dissociation of several multisubunit membrane proteins including the synaptic ves- icle proton pump and the inositol 1,4,5-trisphosphate receptor (27), synaptotagmin migrated as a dimer at the same position as the monomeric 116,000 proton pump subunit (Fig. 10). These results suggest that synaptotagmin is part of a high molecular weight complex in the synaptic vesicle membrane whose basic component is a synaptotagmin dimer.
The primary structure of rat synaptotagmin contains a sequence with a strong potential of forming an amphipathic a-helix. This sequence could dimerize synaptotagmin via its hydrophobic face and form salt bridges between the two parallel helices (Fig. 11). Interestingly, the hypersensitive proteolytic site of synaptotagmin was mapped to the amino- terminal end of this amphipathic a-helix, compatible with the notion that the protein is particularly exposed to proteolytic attack at the point where its peptide backbone emerges from a leucine zipper-like dimer.
These biochemical data together with a sequence compari-

Structure of Synaptotagmin 629
son between different species lead to a five-domain model of synaptotagmin. The first domain consists of the intravesicular glycosylated 52 amino acids, the second domain of the trans- membrane region, and the third domain is formed by the dimerizing amphipathic a-helix. The internal repeats homol- ogous to protein kinase C constitute the fourth domain, and the fifth domain is made up of the last 40 amino acids following the internal repeats. All five domains are present in the rat, human, and Drosophila sequences of synaptotagmin, but only the last two are conserved evolutionarily, suggesting that they form the functional parts of the protein whereas the other domains may have a primarily structural role.
Finally, the subcellular localization of synaptotagmin was investigated biochemically. Together with synaptophysin, synaptotagmin was found to be highly enriched in synaptic vesicles of brain cortex. In contrast, the distributions of synaptophysin and synaptotagmin partially segregate in the adrenal medulla. Here, synaptotagmin has the same distri- bution as cytochrome b561 and is found both on chromaffin granules and on synaptic-like microvesicles, whereas synap- tophysin is exclusively localized to the latter (Figs. 2 and 3).
Several studies on the distribution of synaptophysin in the adrenal medulla have been reported with originally conflicting results (e.g. see Refs. 12, 22, and 28-30). However, more recently, a consensus seems to have evolved that most if not all synaptophysin in chromaffin cells is not on the chromaffin granules but on unidentified “synaptic-like microvesicles” in agreement with our results (e.g. see Refs. 6 and 31). Two studies have also previously investigated the distribution of both synaptophysin and synaptotagmin (p65) in the adrenal medulla (28, 29). In both studies, synaptotagmin was found to be on chromaffin granules in agreement with our findings, although synaptophysin in both studies was found to have more general distribution different from what is now recog- nized to be its restricted presence in synaptic-like microvesi- cles. The presence of synaptotagmin on chromaffin granules, which are similar to large dense core vesicles in brain, raises the possibility that this particular synaptic vesicle protein may have a general function as a docking or fusion protein in regulated secretion by neurons.
REFERENCES
1. Matthew, W. D., Tsavaler, L., and Reichardt, L. F. (1981) J. Cell
2. Perin, M. S., Fried, V. A., Mignery, G. A., Jahn, R., and Sudhof,
3. Perin, M. S., Johnston, P. A., Ozcelik, T., Jahn, R., Francke, U.,
Biol. 9 1 , 257-269
T. C. (1990) Nature 345 , 260-261
and Sudhof, T. C. (1991) J. Biol. Chem. 2 6 6 , 615-622
4. Johnston, P. A., Jahn, R., and Sudhof, T. C. (1989) J. Biol. Chem.
5. Baumert, M., Maycox, P. R., Navone, F., DeCamilli, P., and Reinhard, J. (1989) EMBO J. 8, 379-384
6. Fischer v. Mollard, G., Mignery, G. A., Baumert, M., Perin, M. S., Hanson, T. J., Burger, P. M., Jahn, R., and Sudhof, T. C. (1990) Proc. Natl. Acad. Sci. U. S. A. 8 7 , 1988-1992
7. Perin, M. S., Fried, V. A., Slaughter, C. A,, and Sudhof, T. C.
8. Studier, F. W., Rosenberg, A. H., and Dunn, J. J. (1989) Methods
9. Nagy, A., Baker, R. R., Morris, S. J., and Whittaker, V. P. (1976)
10. Huttner, W. G., Schiebler, W., Greengard, P., and De Camilli, P.
11. Smith, A. D., and Winkler, H. (1967) Biochem. J. 103, 480-482 12. Burger, P. M., Mehl, E., Cameron, P. L., Maycox, P. R., Baumert,
M., Lottspeich, F., DeCamilli, P., and Jahn, R. (1989) Neuron
13. Johnston, P. A., Camerson, P. L., Stukenbrok, H., Jahn, R., DeCamilli, P., and Sudhof, T . C. (1989) EMBO J. 8 , 2863- 2872
264,1268-1273
(1988) EMEO J. 7, 2697-2703
Enzymol., in press
Brain Res. 109, 285-309
(1983) J. Cell Biol. 96, 1374-1388
3 , 715-720
14. Laemmli, U. K. (1970) Nature 227 , 680-685 15. Towbin, H., Staehelin, T., and Gordon, J . (1979) Proc. Natl. Acad.
16. Bradford, M. M. (1976) Anal. Biochem. 7 2 , 248-254 17. Sudhof, T. C., Russell, D. W., Brown, M. S., and Goldstein, J. L.
(1987) Cell 48,1061-1069 18. Sudhof, T. C., Lottspeich, F., Greengard, P., Mehl, E., and Jahn,
R. (1987) Science 238, 1142-1144 19. Sudhof, T. C., Czernik, A. J., Kao, H.-T., Takei, K., Johnston, P.
A., Horiuchi, A., Kanazir, S. D., Wagner, M. A., Perin, M. S.. De Camilli, P., and Greengard, P. (1989) Science 245, 1474- 1480
20. Jahn, R., Schiebler, W., Ouimet, C., and Greengard, P. (1985) Proc. Natl. Acad. Sci. U. S. A. 8 2 , 4137-4141
21. Wiedenmann, B., and Franke, W. W. (1985) Cell 4 1 , 1017-1028 22. Navone, F., Jahn, R., Di Gioia, G., Stukenbrok, H., Greengard,
23. Johnston, P. A., and Sudhof, T. C. (1990) J. Biol. Chem. 265 ,
24. Xie, X.-S., and Stone, D. K. (1988) J. Biol. Chem. 263 , 9859-
25. von Hiejne, G. (1986) Nucleic Acids Res. 14, 4683-4690 26. Astell, C. R., Ahlstrom-Jonasson, L., and Smith, M. (1981) Cell
27. Mignery, G. A., Newton, C. L., Archer, B. T., 111, and Sudhof, T.
28. Lowe, A. W., Madeddu, L., and Kelly, R. B. (1988) J . Cell Biol.
29. Fournier, S., Novas, M. L., and Trifaro, J.-M. (1989) J. Neuro- chem. 53,1043-1049
30. Obendorf, D., Schwarzenbrunner, U., Fischer-colbrie, R., Laslop, A., and Winkler, H. (1988) J. Neurochem. 5 1 , 1573-1580
31. Cutler, D. F., and Cramer, L. P. (1990) J. Cell Biol. 110, 721- 730
Sci. U. S. A. 7 6 , 4350-4354
P., and De Camilli, P . (1986) J. Cell Biol. 103, 2511-2527
8869-8873
9867
27,15-23
C. (1990) J. Biol. Chem. 2 6 5 , 12679-12685
106,51-59