
10/09/08Biochemistry: Nucleic Acid Struct II
Nucleic AcidStructure II
Andy HowardIntroductory Biochemistry
9 October 2008

10/09/08 Biochemistry: Nucleic Acid Struct II p. 2 of 47
What we’ll discuss Folding kinetics Supercoils Nucleosomes Chromatin and chromosomes Lab synthesis of genes tRNA & rRNA structure

10/09/08 Biochemistry: Nucleic Acid Struct II p. 3 of 47
Getting from B to Z
Can be accomplished without breaking bonds
… even though purines have their glycosidic bonds flipped (anti -> syn) and the pyrimidines are flipped altogether!

10/09/08 Biochemistry: Nucleic Acid Struct II p. 4 of 47
Summaries of A, B, Z DNA

10/09/08 Biochemistry: Nucleic Acid Struct II p. 5 of 47
DNA is dynamic Don’t think of these diagrams as static The H-bonds stretch and the torsions
allow some rotations, so the ropes can form roughly spherical shapes when not constrained by histones
Shape is sequence-dependent, which influences protein-DNA interactions

10/09/08 Biochemistry: Nucleic Acid Struct II p. 6 of 47
Intercalating agents Generally: aromatic compounds that can
form -stack interactions with bases Bases must be forced apart to fit them in Results in an almost ladderlike structure
for the sugar-phosphate backbone locally Conclusion: it must be easy to do local
unwinding to get those in!

10/09/08 Biochemistry: Nucleic Acid Struct II p. 7 of 47
Instances of inter-calators

10/09/08 Biochemistry: Nucleic Acid Struct II p. 8 of 47
Denaturing and Renaturing DNA
See Figure 11.17 When DNA is heated to 80+ degrees
Celsius, its UV absorbance increases by 30-40%
This hyperchromic shift reflects the unwinding of the DNA double helix
Stacked base pairs in native DNA absorb less light
When T is lowered, the absorbance drops, reflecting the re-establishment of stacking
10/09/08 Biochemistry: Nucleic Acid Struct II p. 9 of 47
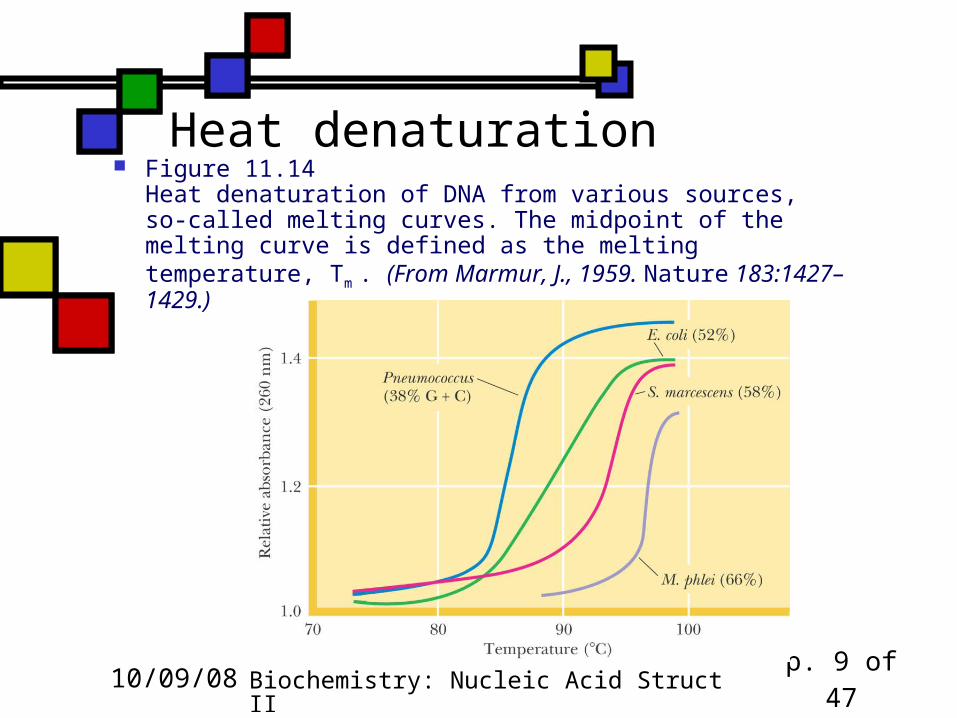
Heat denaturation Figure 11.14
Heat denaturation of DNA from various sources, so-called melting curves. The midpoint of the melting curve is defined as the melting temperature, Tm . (From Marmur, J., 1959. Nature 183:1427–1429.)
10/09/08 Biochemistry: Nucleic Acid Struct II p. 10 of 47
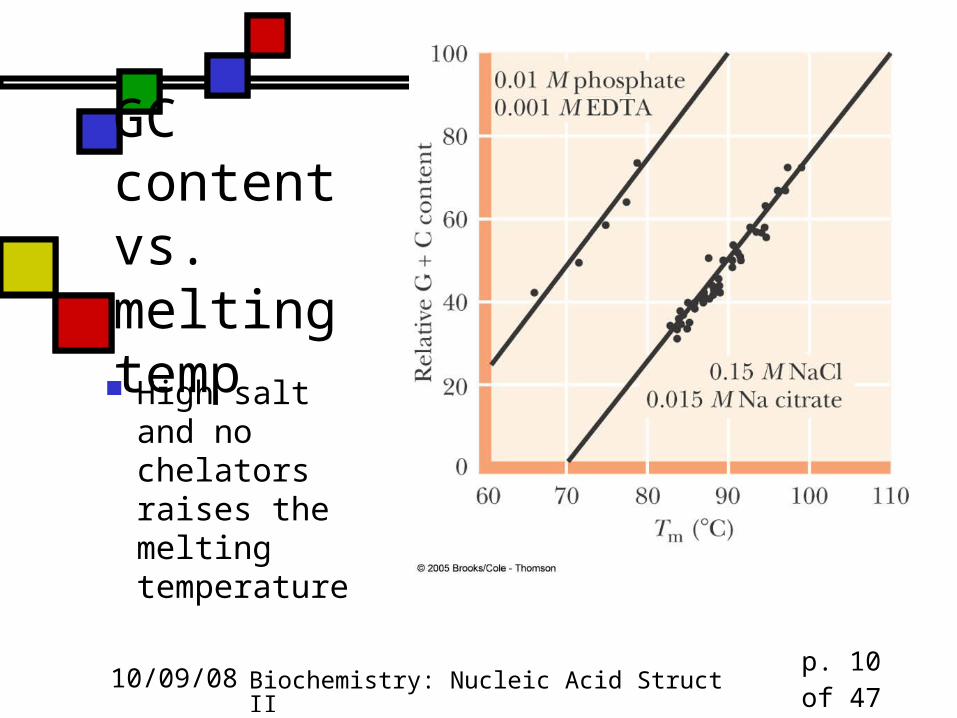
GC content vs. melting temp High salt and
no chelators raises the melting temperature

10/09/08 Biochemistry: Nucleic Acid Struct II p. 11 of 47
How else can we melt DNA?
High pH deprotonates the bases so the H-bonds disappear
Low pH hyper-protonates the bases so the H-bonds disappear
Alkalai is better: it doesn’t break the glycosidic linkages
Urea, formamide make better H-bonds than the DNA itself so they denature DNA

10/09/08 Biochemistry: Nucleic Acid Struct II p. 12 of 47
What happens if we separate the strands?
We can renature the DNA into a double helix
Requires re-association of 2 strands: reannealing
The realignment can go wrong Association is 2nd-order, zippering is first
order and therefore faster

10/09/08 Biochemistry: Nucleic Acid Struct II p. 13 of 47
Steps in denaturation and renaturation

10/09/08 Biochemistry: Nucleic Acid Struct II p. 14 of 47
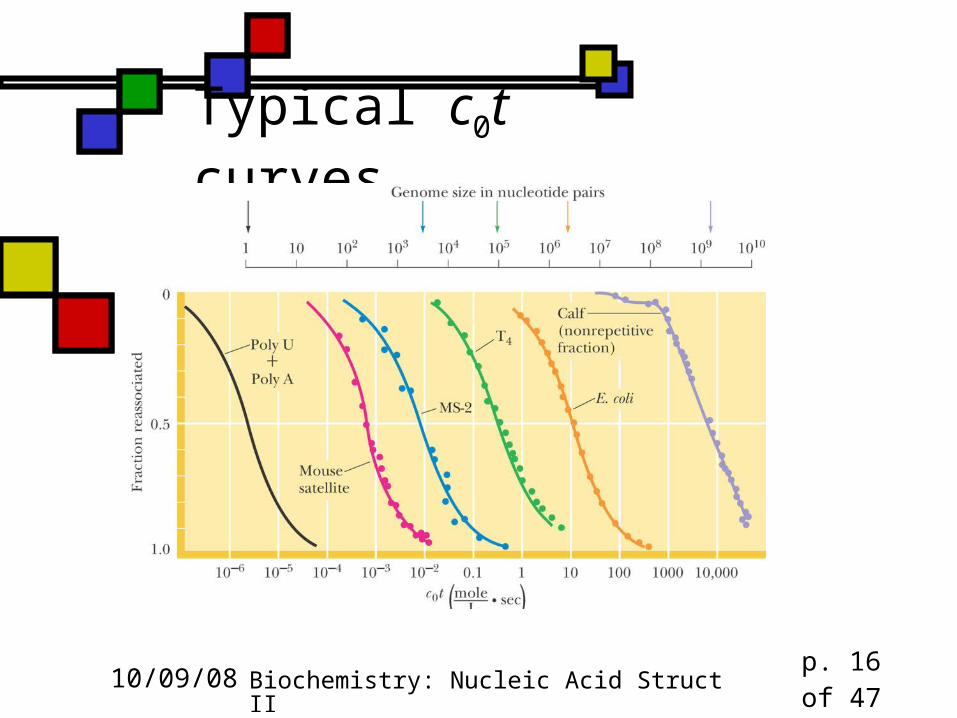
Rate depends on complexity The more complex DNA is, the longer it
takes for nucleation of renaturation to occur
“Complex” can mean “large”, but complexity is influenced by sequence randomness: poly(AT) is faster than a random sequence

10/09/08 Biochemistry: Nucleic Acid Struct II p. 15 of 47
Second-order kinetics
Rate of association: -dc/dt = k2c2
Boundary condition is fully denatured concentration c0 at time t=0:
c / c0 = (1+k2c0t)-1
Half time is t1/2 = (k2c0)-1
Routine depiction: plot c0t vs. fraction reassociated (c /c0) and find the halfway point.
10/09/08 Biochemistry: Nucleic Acid Struct II p. 16 of 47
Typical c0t curves
10/09/08 Biochemistry: Nucleic Acid Struct II p. 17 of 47
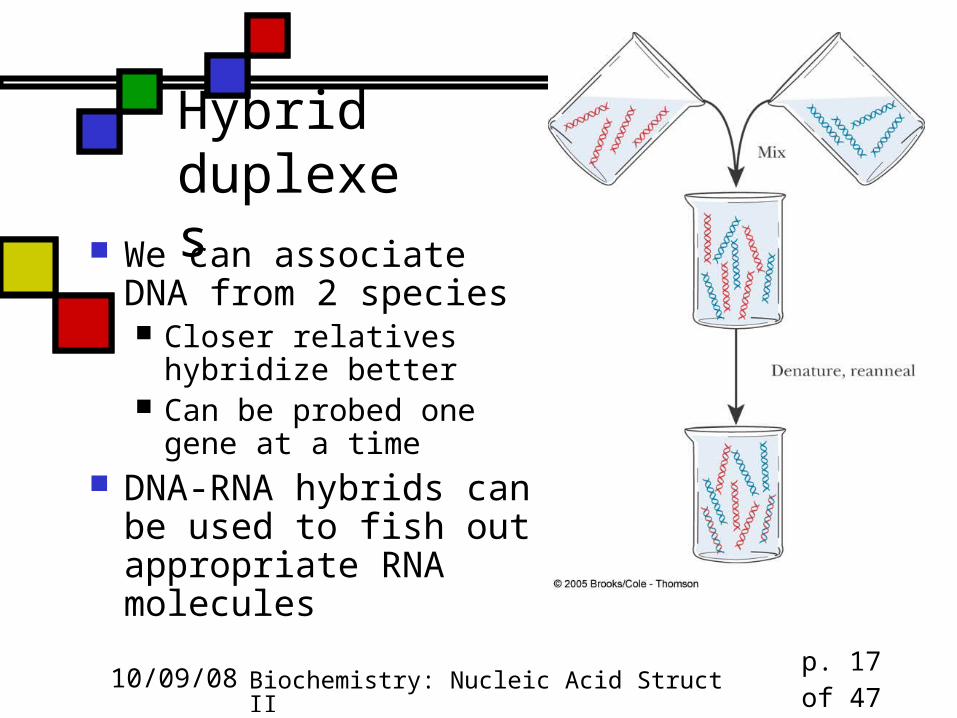
Hybrid duplexes
We can associate DNA from 2 species Closer relatives hybridize
better Can be probed one gene
at a time DNA-RNA hybrids can
be used to fish out appropriate RNA molecules

10/09/08 Biochemistry: Nucleic Acid Struct II p. 18 of 47
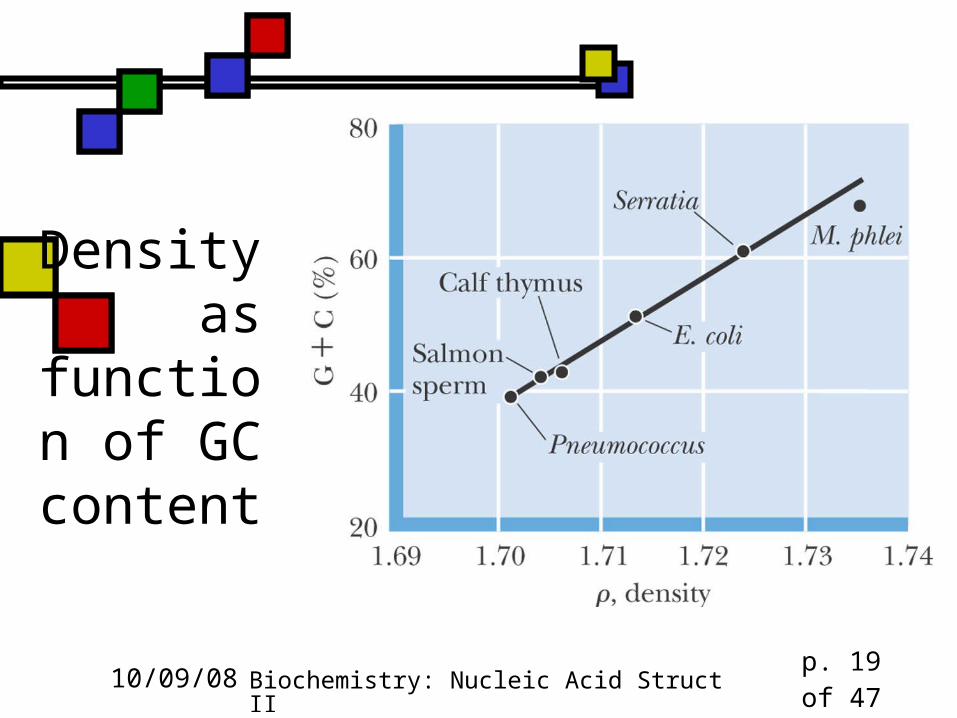
GC-rich DNA is denser DNA is denser than RNA or protein, period,
because it can coil up so compactly Therefore density-gradient centrifugation
separates DNA from other cellular macromolecules
GC-rich DNA is 3% denser than AT-rich Can be used as a quick measure of GC
content
10/09/08 Biochemistry: Nucleic Acid Struct II p. 19 of 47
Density as
function of GC
content

10/09/08 Biochemistry: Nucleic Acid Struct II p. 20 of 47
Tertiary Structure of DNA In duplex DNA, ten bp per turn of helix Circular DNA sometimes has more or less
than 10 bp per turn - a supercoiled state Enzymes called topoisomerases or gyrases
can introduce or remove supercoils Cruciforms occur in palindromic regions of
DNA Negative supercoiling may promote
cruciforms

10/09/08 Biochemistry: Nucleic Acid Struct II p. 21 of 47
DNA is wound Standard is one winding per helical
turn, i.e. 1 winding per 10 bp Fewer coils or more coils can happen: This introduces stresses that favors
unwinding Both underwound and overwound
DNA compact the DNA so it sediments faster than relaxed DNA

10/09/08 Biochemistry: Nucleic Acid Struct II p. 22 of 47
Linking, twists, and writhe
T=Twist=number of helical turns W=Writhe=number of supercoils L=T+W = Linking number is constant
unless you break covalent bonds

10/09/08 Biochemistry: Nucleic Acid Struct II p. 23 of 47
Examples with a tube

10/09/08 Biochemistry: Nucleic Acid Struct II p. 24 of 47
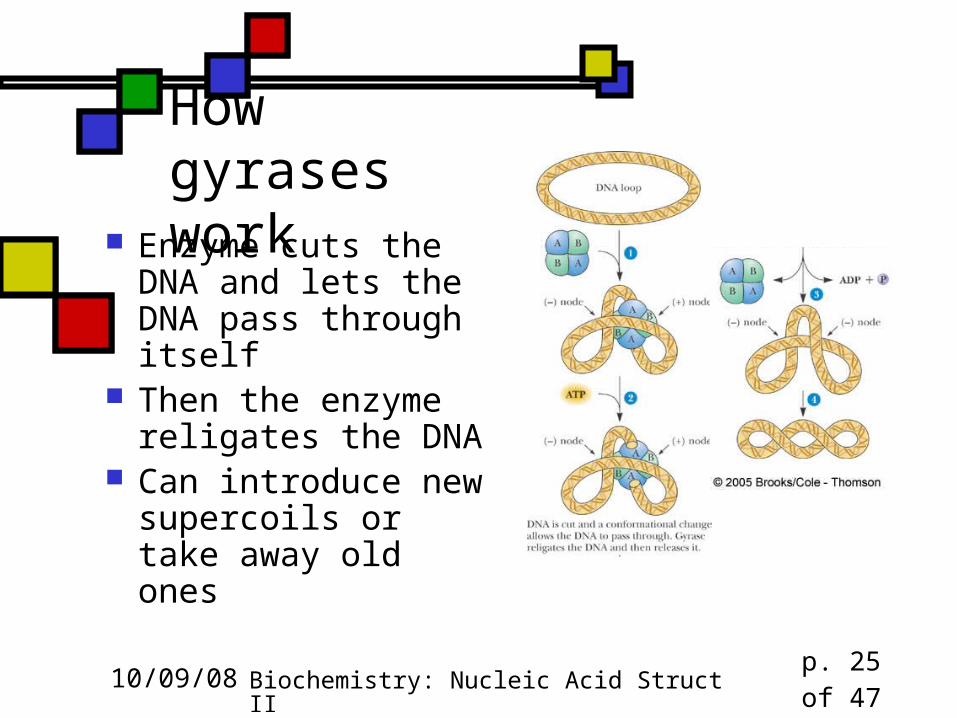
How this works with real DNA
10/09/08 Biochemistry: Nucleic Acid Struct II p. 25 of 47
How gyrases work
Enzyme cuts the DNA and lets the DNA pass through itself
Then the enzyme religates the DNA
Can introduce new supercoils or take away old ones

10/09/08 Biochemistry: Nucleic Acid Struct II p. 26 of 47
Typical gyrase action
Takes W=0 circular DNA and supercoils it to W=-4
This then relaxes a little by disrupting some base-pairs to make ssDNA bubbles

10/09/08 Biochemistry: Nucleic Acid Struct II p. 27 of 47
Superhelix density Compare L for real DNA to what it would
be if it were relaxed (W=0): That’s L = L - L0
Sometimes we want = superhelix density= specific linking difference = L / L0
Natural circular DNA always has < 0

10/09/08 Biochemistry: Nucleic Acid Struct II p. 28 of 47
< 0 and spools The strain in < 0 DNA can be alleviated
by wrapping the DNA around protein spool That’s part of what stabilizes nucleosomes

10/09/08 Biochemistry: Nucleic Acid Struct II p. 29 of 47
Cruciform DNA Cross-shaped structures arise from
palindromic structures, including interrupted palindromes like this example
These are less stable than regular duplexes but they are common, and they do create recognition sites for DNA-binding proteins, including restriction enzymes

10/09/08 Biochemistry: Nucleic Acid Struct II p. 30 of 47
Cruciform DNA example

10/09/08 Biochemistry: Nucleic Acid Struct II p. 31 of 47
Eukaryotic chromosome structure
Human DNA’s total length is ~2 meters! This must be packaged into a nucleus that
is about 5 micrometers in diameter This represents a compression of more
than 100,000! It is made possible by wrapping the DNA
around protein spools called nucleosomes and then packing these in helical filaments

10/09/08 Biochemistry: Nucleic Acid Struct II p. 32 of 47
Nucleosome Structure Chromatin, the nucleoprotein
complex, consists of histones and nonhistone chromosomal proteins
Histone octamer structure has been solved (without DNA by Moudrianakis, and with DNA by Richmond)
Nonhistone proteins are regulators of gene expression

10/09/08 Biochemistry: Nucleic Acid Struct II p. 33 of 47
Histone types H2a, H2b, H3, H4 make up the core
particle: two copies of each, so: octamer
All histones are KR-rich, small proteins
H1 associates with the regions between the nucleosomes
10/09/08 Biochemistry: Nucleic Acid Struct II p. 34 of 47
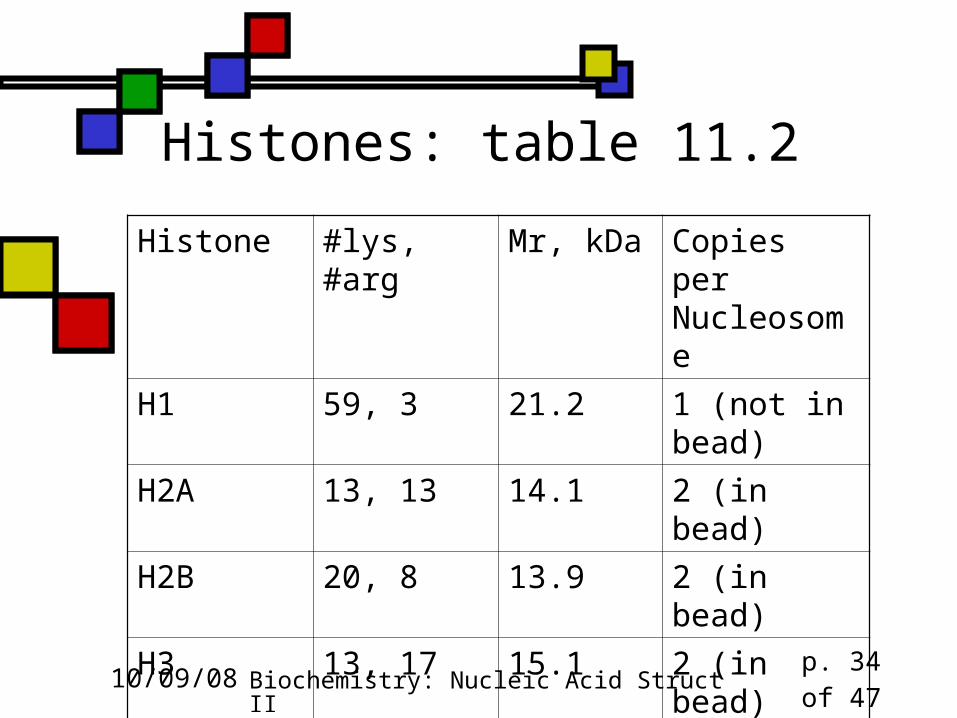
Histones: table 11.2
Histone #lys, #arg Mr, kDa Copies per Nucleosome
H1 59, 3 21.2 1 (not in bead)
H2A 13, 13 14.1 2 (in bead)
H2B 20, 8 13.9 2 (in bead)
H3 13, 17 15.1 2 (in bead)
H4 11, 14 11.4 2 (in bead)
10/09/08 Biochemistry: Nucleic Acid Struct II p. 35 of 47
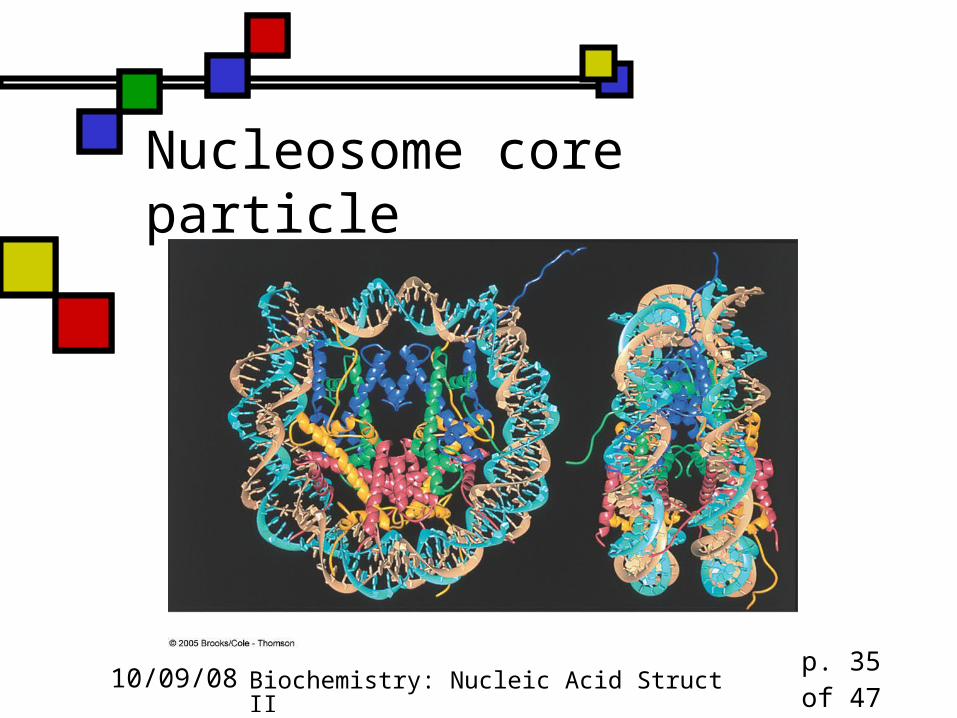
Nucleosome core particle

10/09/08 Biochemistry: Nucleic Acid Struct II p. 36 of 47
Half the core particle
Note that DNA isn’t really circular: it’s a series of straight sections followed by bends

10/09/08 Biochemistry: Nucleic Acid Struct II p. 37 of 47
Histones, continued Individual nucleosomes
attach via histone H1 to seal the ends of the turns on the core and organize 40-60bp of DNA linking consecutive nucleosomes
N-terminal tails of H3 & H4 are accessible
K, S get post-translational modifications, particularly K-acetylation

10/09/08 Biochemistry: Nucleic Acid Struct II p. 38 of 47
Chromosome structure: levels Each of the
first 4 levels compacts DNA by a factor of 6-20; those multiply up to > 104
10/09/08 Biochemistry: Nucleic Acid Struct II p. 39 of 47
Synthesizing nucleic acids
Laboratory synthesis of nucleic acids requires complex strategies
Functional groups on the monomeric units are reactive and must be blocked
Correct phosphodiester linkages must be made
Recovery at each step must high!

10/09/08 Biochemistry: Nucleic Acid Struct II p. 40 of 47
Solid Phase Oligonucleotide Synthesis
Dimethoxytrityl group blocks the 5'-OH of the first nucleoside while it is linked to a solid support by the 3'-OH
Step 1: Detritylation by trichloroacetic acid exposes the 5'-OH
Step 2: In coupling reaction, second base is added as a nucleoside phosphoramidate

Figure 11.29Solid phase oligonucleotide synthesis. The four-step cycle starts with the first base in nucleoside form (N-1) attached by its 3'-OH group to an insoluble, inert resin or matrix, typically either controlled pore glass (CPG) or silica beads. Its 5'-OH is blocked with a dimethoxytrityl (DMTr) group (a). If the base has reactive NH2 functions, as in A, G, or C, then N-benzoyl or N-isobutyryl derivatives are used to prevent their reaction (b). In step 1, the DMTr protecting group is removed by trichloroacetic acid treatment. Step 2 is the coupling step: the second base (N-2) is added in the form of a nucleoside phosphoramidite derivative whose 5'-OH bears a DMTr blocking group so it cannot polymerize with itself (c).

10/09/08 Biochemistry: Nucleic Acid Struct II p. 42 of 47
Solid Phase Synthesis
Step 3: capping with acetic anhydride blocks unreacted 5’-OHs of N-1 from further reaction
Step 4: Phosphite linkage between N-1 and N-2 is reactive and is oxidized by aqueous iodine to form the desired, and more stable, phosphate group

10/09/08 Biochemistry: Nucleic Acid Struct II p. 43 of 47
Activation of the phosphoramidate
10/09/08 Biochemistry: Nucleic Acid Struct II p. 44 of 47
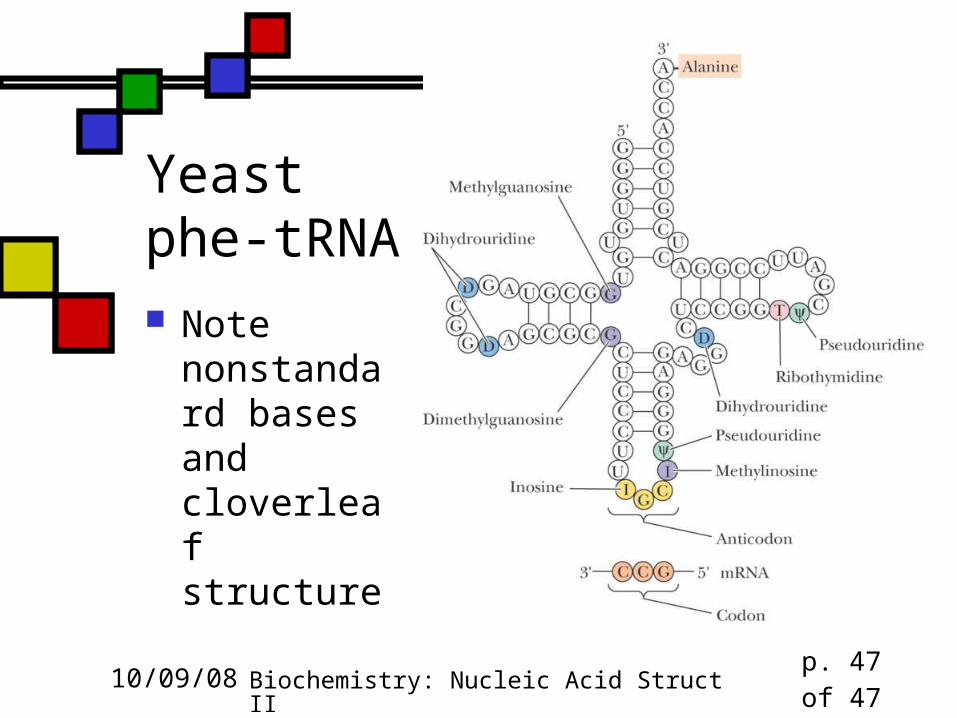
Secondary and Tertiary Structure of RNA
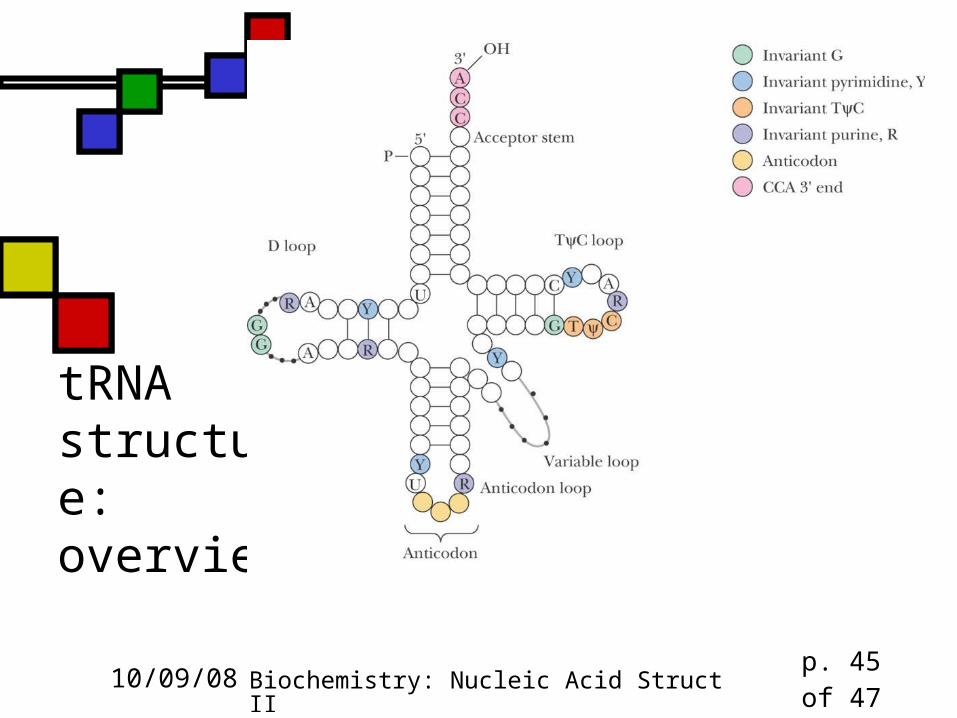
Transfer RNA Extensive H-bonding creates four double
helical domains, three capped by loops, one by a stem
Only one tRNA structure (alone) is known Phenylalanine tRNA is "L-shaped" Many non-canonical base pairs found in tRNA
10/09/08 Biochemistry: Nucleic Acid Struct II p. 45 of 47
tRNA structure: overview
10/09/08 Biochemistry: Nucleic Acid Struct II p. 46 of 47
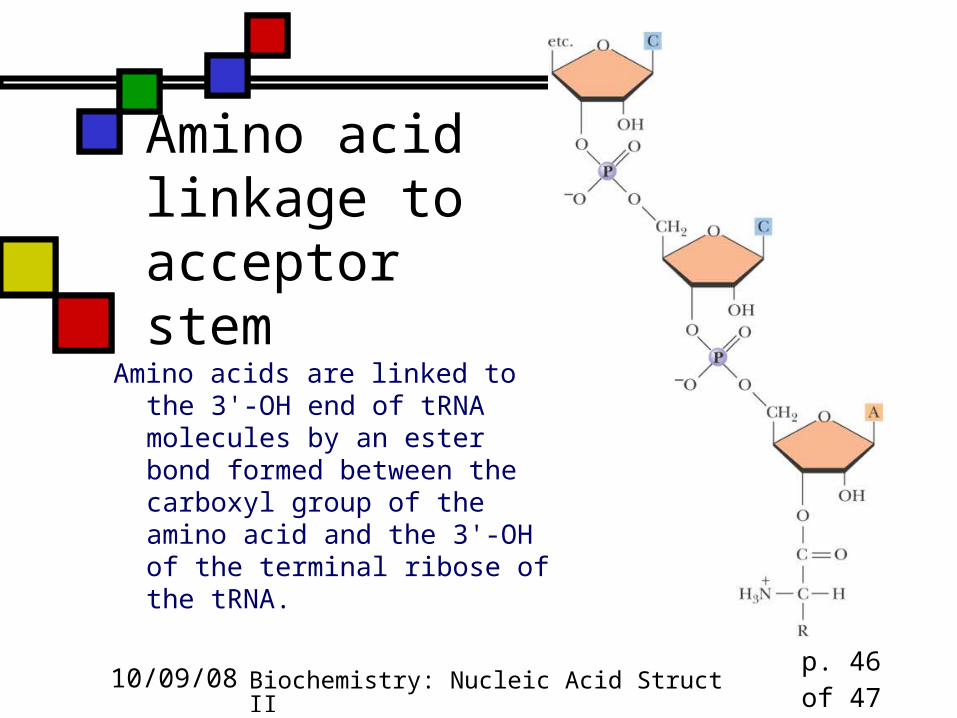
Amino acid linkage to acceptor stem
Amino acids are linked to the 3'-OH end of tRNA molecules by an ester bond formed between the carboxyl group of the amino acid and the 3'-OH of the terminal ribose of the tRNA.
10/09/08 Biochemistry: Nucleic Acid Struct II p. 47 of 47
Yeast phe-tRNA Note
nonstandard bases and cloverleaf structure







