
IntroductorybiophysicsA.Y.2016-17
5.X-raycrystallographyanditsapplicationstothestructural
problemsofbiologyEdoardoMilotti
Dipartimento diFisica,Università diTrieste

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Theinteratomicdistanceinametalliccrystalcanberoughlyestimatedasfollows.
Take,e.g.,iron
• density:7.874g/cm3
• atomicweight:56• molarvolume:VM =7.1cm3/mole
thentheinteratomicdistanceisroughly
d ≈ VMNA
3 ≈ 2.2nm

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Theatomiclatticecanbeusedasortofdiffractiongratingforshort-wavelengthradiation,about100timesshorterthanvisiblelightwhichisintherange400-750nm.
Since
1nmradiationcorrespondstoabout1keV photonenergy.
Eγ =hcλ
≈ 2·10−25 J mλ
≈ 1.24 eV µmλ

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
!"#$%&'$(")*
S#%/J&T&U2*#<.%&CKET3&VG$GG./"#%G3&W.%-$/;
+(."J&AN&>,%()&CTDB3&S.%)(/3&X.1*&W.%-$/;
Y#<.)&V%(Z.&(/&V=;1(21&(/&CTCL&[G#%&=(1&"(12#5.%;&#G&*=.&"(GG%$2*(#/&#G&\8%$;1&<;&2%;1*$)1]
9/(*($));&=.&1*:"(."&H(*=&^_/F*./3&$/"&*=./&H(*=&'$`&V)$/2I&(/&S.%)(/3&H=.%.&=.&=$<()(*$*."&(/&CTBD&H(*=&$&*=.1(1&[a<.% "(.&9/*.%G.%./Z.%12=.(/:/F./ $/&,)$/,$%$)).)./ V)$**./[?&
7=./&=.&H#%I."&$*&*=.&9/1*(*:*.&#G&7=.#%.*(2$)&V=;1(213&=.$"."&<;&>%/#)"&Q#--.%G.)"3&:/*()&=.&H$1&$,,#(/*."&G:))&,%#G.11#%&$*&*=.&4/(5.%1(*;&#G&0%$/IG:%*&(/&CTCL3&H=./&=.&$)1#&%.2.(5."&=(1&Y#<.)&V%(Z.?&

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
ArnoldSommerfeld(1868-1951)
...FourofSommerfeld'sdoctoralstudents,WernerHeisenberg,WolfgangPauli,PeterDebye,andHansBethewentontowinNobelPrizes,whileothers,mostnotably,WalterHeitler,RudolfPeierls,KarlBechert,HermannBrück,PaulPeterEwald,EugeneFeenberg,HerbertFröhlich,ErwinFues,ErnstGuillemin,HelmutHönl,LudwigHopf,AdolfKratzer,OttoLaporte,WilhelmLenz,KarlMeissner,RudolfSeeliger,ErnstC.Stückelberg,HeinrichWelker,GregorWentzel,AlfredLandé,andLéonBrillouinbecamefamousintheirownright.ThreeofSommerfeld'spostgraduatestudents,LinusPauling,IsidorI.RabiandMaxvonLaue,wonNobelPrizes,andtenothers,WilliamAllis,EdwardCondon,CarlEckart,EdwinC.Kemble,WilliamV.Houston,KarlHerzfeld,WaltherKossel,PhilipM.Morse,HowardRobertson,andWojciechRubinowiczwentontobecomefamousintheirownright.WalterRogowski,anundergraduatestudentofSommerfeldatRWTHAachen,alsowentontobecomefamousinhisownright.
MaxBornbelievedSommerfeld'sabilitiesincludedthe"discoveryanddevelopmentoftalents."AlbertEinsteintoldSommerfeld:"WhatIespeciallyadmireaboutyouisthatyouhave,asitwere,poundedoutofthesoilsuchalargenumberofyoungtalents."Sommerfeld'sstyleasaprofessorandinstitutedirectordidnotputdistancebetweenhimandhiscolleaguesandstudents.Heinvitedcollaborationfromthem,andtheirideasofteninfluencedhisownviewsinphysics.Heentertainedtheminhishomeandmetwiththemincafesbeforeandafterseminarsandcolloquia.Sommerfeldownedanalpineskihuttowhichstudentswereofteninvitedfordiscussionsofphysicsasdemandingasthesport....
fromhttps://en.wikipedia.org/wiki/Arnold_Sommerfeld

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
...Suchwasthestateofaffairsas,oneeveninginFebruary1912,P.P.Ewald cametovisitme.(...)hewasfacedatthattimewithcertaindifficultiesandcametomewitharequestforadvice.
Nowitwasnot,however,possibleformetoassisthimatthattime.ButduringtheconversationIwassuddenlystruckbytheobviousquestionofthebehaviour ofwaveswhichareshortbycomparisonwiththelattice-constantsofthespacelattice.Anditwasatthatpointthatmyintuitionforopticssuddenlygavemetheanswer:latticespectrawouldhavetoensue.
Thefactthatthelatticeconstantincrystalsisofanorderof10-8 cmwassufficientlyknownfromtheanalogywithotherinteratomicdistancesinsolidandliquidsubstances,and,inaddition,thiscouldeasilybearguedfromthedensity,molecularweightandthemassofthehydrogenatomwhich,justatthattime,hadbeen particularlywelldetermined.
TheorderofX-raywavelengthswasestimatedbyWienandSommerfeld at10-9 cm.Thustheratioofwavelengthsandlatticeconstantswasextremelyfavourable ifX-raysweretobetransmittedthroughacrystal.IimmediatelytoldEwald thatIanticipatedtheoccurrenceofinterferencephenomenawithX-rays....
(fromvonLaue’sNobelLecture)

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
R#/&g$:.n1&.`,.%(-./*$)&)$;#:*

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
9-$F.&*$I./&(/&CTCA&#G&$/&\8%$;&(/*.%G.%./2.&#G&$&Z(/2&<)./".&2%;1*$)?&
o(/2&<)./".&bo/Q3&1,=$).%(*.c&H$1&#/.&#G&*=.&G(%1*&2%;1*$)1&(/5.1*(F$*."&<;&g$:.3&0%(."%(2=&$/"&e/(,,(/F?
bV=#*#F%$,=J&+.:*12=.1 ':1.:-c
!"#$#%&"'()#%*))+,#-.&%#/)%0(-1#
23 %.4*)5#-.$-#%4(&5%#$/)#4/5)/)5#$-#-.)#64()70($/#()8)(93 &-#%)--()5#$((#:0)%-&4"%#4"#-.)#"$-0/)#4;#<=/$>%

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
“Dear Mr. Laue! I cordially salute you on your marvelous success. Your experiment counts among the most glorious that Physics has seen so far.”
Albert Einstein
(onapostcardtovonLaue,datedJune1912)

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
/:,$8:--:"0$9*',?$@,"<<
S#%/J&A&p:);&CKDA3&X(F*#/3&4/(*."&e(/F"#-
+(."J&CA&'$%2=&CTLA3&g#/"#/3&4/(*."&e(/F"#-
Y#<.)&V%(Z.&(/&V=;1(21&(/&CTCO&[G#%&*=.(%&1.%5(2.1&(/&*=.&$/$);1(1&#G&2%;1*$)&1*%:2*:%.&<;&-.$/1&#G&\8%$;1]

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
8:--:"0$("B,*'5*$@,"<<
S#%/J&NC&'$%2=&CKTB3&>".)$(".3&>:1*%$)($
+(."J&C&p:);&CTEC3&9,1H(2=3&4/(*."&e(/F"#-
Y#<.)&V%(Z.&(/&V=;1(21&(/&CTCO&[G#%&*=.(%&1.%5(2.1&(/&*=.&$/$);1(1&#G&2%;1*$)&1*%:2*:%.&<;&-.$/1&#G&\8%$;1]

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
Y$h) 2%;1*$)1

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
Q,=$).%(*.3&+#)#-(*.3&h=$)2#,;%(*.?&g#2$)(*;J&p#,)(/&0(.)"3&7%(8Q*$*.&+(1*%(2*3&p$1,.%&h#:/*;3&'(11#:%(3&4Q>&b=**,JMM./?H(I(,."($?#%FMH(I(MQ,=$).%(*.c

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Crystalstructure
Inordertoproceed,andexplainthecontributionsbyvonLaueandtheBraggs,wemustdescribeorderincrystals
basis
lattice

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
a1
a2
a1 anda2 aretheprimitivelatticevectors,andthetranslationvectors
generatethewholelattice
T = u1a1 + u2a2 u1,2 integers( )

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
a1
a2
Thispairofa1 anda2 isnotprimitivebecausetheydonotgeneratethewholelattice

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
a1
a2
Theprimitivevectorsalsodefinethecrystalaxes
Theassociatedparallelogramistheprimitivecell
a1
a2

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
a1
a2
Theprimitivevectorsalsodefinethecrystalaxes
Theassociatedparallelepipedistheprimitivecell
Volumeofprimitivecell:
a1
a2
a3
a3
3D V = a1 ⋅a2 × a3

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
7=.%.&$%.&-$/;&*;,.1&#G&)$**(2.1?&7=.;&$%.&1;1*.-$*(2$));&2)$11(G(."&<;&"(12%.*.&1,$2.&F%#:,1?&
7=.&2#--#/&/#-./2)$*:%.&(1&*=$*&#G&*=.&S%$5$(1 g$**(2.1?&
!`$-,).1
L10'>030' T.@7.(-?1*> %0@E>-$'&70'

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
7=.&2:<(2&)$**(2.1
Q(-,).&2:<(2&b12c S#";82./*.%."&2:<(2&b<22c&&&0$2.82./*.%."&2:<(2&bG22c

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Reciprocallatticevectors
Thesevectorsdefinethereciprocallatticeandhavetheproperty
andtheydefineareciprocallattice,bymeansofthetranslationvectors
b1 = 2πa2 × a3a1 ⋅a2 × a3
; b2 = 2πa3 × a1a2 ⋅a3 × a1
; b3 = 2πa1 × a2a3 ⋅a1 × a2
;
ai ⋅b j = 2πδ ij
G = v1b1 + v2b2 + v3b3

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Example:reciprocallatticetoasimplecubic(sc)lattice
a1 =a00
⎛
⎝
⎜⎜
⎞
⎠
⎟⎟ a2 =
0a0
⎛
⎝
⎜⎜
⎞
⎠
⎟⎟a3 =
00a
⎛
⎝
⎜⎜
⎞
⎠
⎟⎟
b1 =1 a00
⎛
⎝
⎜⎜⎜
⎞
⎠
⎟⎟⎟b2 =
01 a0
⎛
⎝
⎜⎜⎜
⎞
⎠
⎟⎟⎟b3 =
001 a
⎛
⎝
⎜⎜⎜
⎞
⎠
⎟⎟⎟
Thereciprocallatticeisagainsc;thesclatticeisself-dual.

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Example:body-centeredcubiclattice(bcc)
fromC.Kittel,“Introd
uctio
ntoSolidStatePhysic
s,8t
hed
.(Wiley,20
05)

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
a1 =
a2
−x + y + z( ); a2 =a2x − y + z( ); a3 =
a2x + y − z( );
a2 × a3 =a2
2⌢y + ⌢z( ); a3 × a1 =
a2
2⌢x + ⌢z( ); a1 × a2 =
a2
2⌢x + ⌢y( );
a1 ⋅ a2 × a3( ) = a3
2
b1 =2πa⌢y + ⌢z( ); b2 =
2πa⌢x + ⌢z( ); b3 =
2πa⌢x + ⌢y( );

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
b1 =
2πay + z( ); b2 =
2πax + z( ); b3 =
2πax + y( );
Thereciprocallatticevectorsofthebcclatticecorrespondtothelatticevectorsofthefcclattice.

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Diffractionofwavesbycrystals
crystalsurface
θ θ
θ
a
b
d

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
crystalsurface
θ θ
θ
b
pathdifferencebetweenraysaandb:
constructiveinterferencecondition:(Bragglaw)
a
2d sinθ
2d sinθ = nλ
d

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Remarks:
• thescatteringcross-sectionissmall,thusX-rayspenetratethecrystalandarescatteredbydifferentplanes
• thescatteringcross-sectionissmall,thustheX-raybeamisnotsignificantlyattenuatedbypreviouscrystalplanes
• X-raysarescatteredbyelectrons,andtheyarescatteredmorewheretheelectrondensityishigher
• theBragglawimpliesthat
nλ2d
= sinθ ≤1 ⇒ n ≤ 2dλ

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Crystallinesolids
Incrystallinesolids,theelectrondensityisperiodicwithrespecttothediscretetranslationsthatdefinethecrystallattice
Then,the electrondensitycanthenbeexpressedasaFourierseries
n r( ) = n r +T( ) T = u1a1 + u2a2 + u3a3; ui ∈Z
n r( ) = nG exp iG ⋅r( )G∑

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
n r( ) = nG exp iG ⋅r( )G∑
WestillhavetofindthevectorsGthatleadtothecorrectdefinitionoftheFourierseries.
Nowwenotethat
andweseethatperiodicityworksif
n r( ) = nG exp iG ⋅r( )G∑ = n r +T( ) = nG exp iG ⋅r + iG ⋅T( )
G∑
G ⋅T = 2πm

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Thecondition issatisfiedbythereciprocallatticetranslations.
Indeed ai ⋅b j = 2πδ ij
G = v1b1 + v2b2 + v3b3T = u1a1 + u2a2 + u3a3
G ⋅T = v1b1 + v2b2 + v3b3( ) ⋅ u1a1 + u2a2 + u3a3( )= 2π v1u1+ v2u2 + v3u3( )
G ⋅T = 2πm

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Itisalsoeasytoseethat
whereVC isthevolumeofacellofthecrystal(proveitashomework!).
nG = 1VC
n r( )exp −iG ⋅r( )dVcell∫

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
X-raydiffraction
fromC.Kittel,“Introd
uctio
ntoSolidStatePhysic
s,8t
hed
.(Wiley,20
05)

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
r
k
ϕ
r sinϕ
path length: r sinϕoptical path length: kr sinϕ
scalar product: k·r = kr cos π2−ϕ⎛
⎝⎜⎞⎠⎟ = kr sinϕ
π2−ϕ
incomingwave

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
r ′k′ϕ
r sin ′ϕ
path length: r sin ′ϕoptical path length: ′k r sin ′ϕ
scalar product: ′k ·r = ′k r cos π2+ϕ⎛
⎝⎜⎞⎠⎟ = −kr sin ′ϕ
π2+ϕ
diffractedwave

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
totalopticalpathlengthdifference
scatteringvector
kr sin'+ kr sin'0 = k · r� k0 · r= �(k0 � k) · r= ��k · r
�k = k0 � k
k0 = k+�k

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Theamplitudescatteredindirectionk’isproportionalto
dF = n r( )exp −iΔk ⋅r( )dV
ThetotalscatteredamplitudeisproportionaltotheFouriertransformofthechargedistribution
F = n r( )exp −iΔk ⋅r( )dVV∫
= nG exp iG ⋅r( )G∑⎡⎣⎢
⎤⎦⎥exp −iΔk ⋅r( )dV
V∫
= nG exp i G − Δk( ) ⋅r⎡⎣ ⎤⎦dVV∫
G∑

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
F = nG exp i G − Δk( ) ⋅r⎡⎣ ⎤⎦dVV∫
G∑
ThisscatteredamplitudereachesalocalmaximumforeachgivenG when
otherwiseitisnegligiblysmall.
Takingintoaccountenergyconservation,sothatk =k’,wefind
Δk =G
′k =G + k
⇒ k2 = ′k 2 = G + k( )2 = G2 + 2G ⋅k + k2
⇒ G2 + 2G ⋅k = 0
(vectorequation)

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
G2 + 2G ⋅k = 0
or,equivalently(withtheremarkthatitholdsfor–G asitaswellasG)
whichisarestatementofBragg’slaw ,andthusweexpectpeaksatthesevaluesofk.
(seeKittel forfurtherdetails).
G2 = 2G ⋅k
2d sinθ = nλ
(scalarequation)

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Δk =G
a1·Δk = 2πv1; a2 ·Δk = 2πv2; a3·Δk = 2πv3;
impliesthatthefollowingequationsmustalsohold
Laueequationseachequationdefinesaconeinkspace;
diffractionoccursonlywhereinthosedirectionswhereconesintersect;
thisisaseverelylimitingcondition,realizedonlywithsystematicsweepingor,occasionally,bychance

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Thepowdermethod
Inthismethodtheconeaxisisdefinedbytheincomingbeam
D.K.Chakrabarty,“SolidStateChe
mistry”(New
AgeInternational,
2010
)

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
Q(/F).&2%;1*$) V#);2%;1*$))(/.&,#H".%

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
D.K.Chakrabarty,“SolidStateChe
mistry”(New
AgeInternational,20
10)

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Whenthelocalmaximumconditionissatisfied,i.e.,
thescatteredamplitudebecomes
WhenthisissummedovertheN cellsofacrystal,wefindtheamplitudeofthecorrespondingdiffractionmaximum
F = n r( )exp −iΔk ⋅r( )dVV∫ = n r( )exp −iG ⋅r( )dV
V∫
Δk =G
F = N n r( )exp −iG ⋅r( )dVcell∫ = NSG

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
SG = n r( )exp −iG ⋅r( )dVcell∫
isthestructurefactor.
Nowintroducetheelectrondensityofeachatominthecell(j-thatom)
then
n r( ) = nj r − rj( )j∑
SG = n r( )exp −iG ⋅r( )dVcell∫ = nj r − rj( )exp −iG ⋅r( )dV
cell∫
j∑

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
SG = n r( )exp −iG ⋅r( )dVcell∫ = nj r − rj( )exp −iG ⋅r( )dV
cell∫
j∑
= exp −iG ⋅rj( ) nj r − rj( )exp −iG ⋅ r − rj( )⎡⎣ ⎤⎦dVcell∫
j∑
= exp −iG ⋅rj( ) nj s( )exp −iG ⋅s( )dVcell∫
j∑
= f j exp −iG ⋅rj( )j∑
where
istheatomicformfactor.
f j = nj s( )exp −iG·s( )dVcell∫

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Thedatainversionproblem
SincetheamplitudeisaFouriersum
wecanfind– atleast inprinciple– theelectrondensityandtheatomiccoordinatesbyFourierinversion.
However,theproblemismoredifficult,becausewedonotmeasuretheamplitude,buttheintensity
andthereforephaseislost(thisisthephaseproblem).
FG = NSG = N fj exp −iG ⋅rj( )j∑ = N fj exp −ivxx j − ivyyj − ivzz j( )
j∑
I ∝ FG2

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Atomicpositionsarealsounknown.Thustherearetworelatedproblems
• phaseproblem• atomicpositions
Severalmethodsexist,thatrelyoninitialtrialsolutionswithsuccessiverefinements.

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
ThePattersonfunction
WefoundearlierthatthescatteredamplitudeisaFouriertransform
sothattheelectrondensitycanberetrievedfromFourierinversion
HoweverwecannotmeasureFdirectly,butonlythescatteredintensity(nophase!),whichisproportionalto|F|2.
F G( ) = n r( )exp −iG ⋅r( )dVV∫
n r( ) = 1V
F G( )exp iG ⋅r( )G∑

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
ThePattersonfunctionP wasintroducedbyArthurLindo Pattersonin1935,anditisessentiallytheinverseFouriertransformofthescatteredintensity
whichissomewhatsimilarto
P u( ) = 1V
F G( ) 2 exp iG ⋅u( )G∑
n r( ) = 1V
F G( )exp iG ⋅r( )G∑

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
NowdenotetheFouriertransformwiththesymbolT andtheinversetransformwithT-1,then
Since ,wefind
P u( ) = T −1 F 2( ) = T −1 F* ⋅F( ) = T −1 F*( )⊗T −1 F( )
applyconvolutiontheorem
n r( ) = T −1 F( )F* G( ) = F −G( )
⎧⎨⎪
⎩⎪
P u( ) = n(r)⊗n(−r) = n(r)n(u+ r)V∫ dr
thisshowsthatthePattersonfunctionistheconvolutionoftheelectrondensitywiththemirrorimageofthedensity

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
AnotherwayoflookingatthePattersonfunction
Peaksoccurat ,i.e.,thePattersonisafunctionofthedistancesbetweenatoms.
P u( ) = 1V
F G( ) 2 exp iG ⋅u( )G∑
= 1V
n ′r( )exp iG ⋅ ′r( )d ′VV∫⎡
⎣⎢
⎤
⎦⎥ n ′′r( )exp −iG ⋅ ′′r( )d ′′V
V∫⎡
⎣⎢
⎤
⎦⎥exp iG ⋅u( )
G∑
= 1V
n ′r( )n ′′r( )V∫ exp iG ⋅ ′r − ′′r + u( )⎡⎣ ⎤⎦
G∑ d ′V d ′′V
thePattersonfunctionhaspeakswherethisparenthesisvanishesandwherethedensitiesarehighest
u = ri − rj

74 Protein Crystallography
Figure 5.1. Building up the Patterson function from the atomic positions. (a) Oneunit cell of a simple two-dimensional crystal containing four atoms. (b) The sameunit cell shown in (a), but showing the vectors connecting the four atoms. (c) Theinteratomic vectors from (b), shown emanating from a common origin. This is howthe vector peaks appear in the Patterson function. (d) Multiple unit cells of thePatterson function. Peaks corresponding to interatomic vectors are shown as dots.The vectors are drawn for only the central unit cell, but all the unit cells are identical.
If we use F as the coe≈cient in a Fourier series we obtain the electron density, r(x).
r(x) has peaks corresponding to the atomic positions, xj. By analogy, we might
expect that using F 2 as the coe≈cient in a Fourier series will give us a function,
P(u), that has peaks corresponding to the interatomic vectors, (xj – xk). This is
indeed the case and is easily demonstrated. Combining equations (5.1) and (5.2)
we obtain
P(u) =1
V !h
"!j
!k
fj fk exp(2pih § [xj – xk])#exp(–2pih § u)
Combining terms leads to the following:
P(u) =1
V !h
!j
!k
fj fk exp(–2pih § [u – (xj – xk)])
74 Protein Crystallography
Figure 5.1. Building up the Patterson function from the atomic positions. (a) Oneunit cell of a simple two-dimensional crystal containing four atoms. (b) The sameunit cell shown in (a), but showing the vectors connecting the four atoms. (c) Theinteratomic vectors from (b), shown emanating from a common origin. This is howthe vector peaks appear in the Patterson function. (d) Multiple unit cells of thePatterson function. Peaks corresponding to interatomic vectors are shown as dots.The vectors are drawn for only the central unit cell, but all the unit cells are identical.
If we use F as the coe≈cient in a Fourier series we obtain the electron density, r(x).
r(x) has peaks corresponding to the atomic positions, xj. By analogy, we might
expect that using F 2 as the coe≈cient in a Fourier series will give us a function,
P(u), that has peaks corresponding to the interatomic vectors, (xj – xk). This is
indeed the case and is easily demonstrated. Combining equations (5.1) and (5.2)
we obtain
P(u) =1
V !h
"!j
!k
fj fk exp(2pih § [xj – xk])#exp(–2pih § u)
Combining terms leads to the following:
P(u) =1
V !h
!j
!k
fj fk exp(–2pih § [u – (xj – xk)])
fromLattm
an&Loll:“Proteincrystallography:acon
ciseguide
”,John
Hop
kinsUniv.Press,20
08
EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
The Patterson Function 77
Figure 5.3. An example of how atomic positions can be inferred from the Pattersonfunction when heavy atoms are present. (a) A simple organic compound—iodoben-zene—containing a single heavy atom. The iodine atom is dark gray and is placed atthe origin of an arbitrary coordinate system. (b) The Patterson function calculatedfrom this molecule. Peaks in the Patterson function are shown as dots. Vectors be-tween the iodine atom and carbon atoms appear as dark peaks, while carbon–carbonvectors are shown as light peaks. There is also a peak at the origin, as is always the casefor the Patterson function. Note how the dark peaks reveal both the structure of themolecule and its mirror image.
ing a moderate number of light atoms plus a few heavy atoms, the heavy atom-
heavy atom peaks can frequently be identified in the Patterson function, allowing
the heavy atom positions to be inferred.
Unfortunately, in large molecules like proteins, the Patterson functions are so
complex that even heavy atom peaks become lost. However, as we have seen in
Chapter 4, the positions of the heavy atoms must be known before we can
calculate phases in the MIR experiment. How do we find them? In this case, we
use what are called di√erence Patterson functions to accentuate the heavy atom
peaks and allow their identification.
Suppose we are searching for the positions of the heavy atoms in a heavy
atom derivative of a protein. The heavy atoms are bound to the protein and
occupy specific sites in the crystal lattice. Now imagine that we could magically
erase all the protein atoms from the crystal, without changing the positions of the
heavy atoms. The di√raction pattern of this imaginary heavy atom-only crystal
would correspond to the structure factors FH(h). The Patterson function calcu-
lated from these data would have coe≈cients F 2H(h) and would be ideally suited
for determining the positions of the heavy atoms. Of course, we can’t erase the
protein atoms, and we can’t measure FH, but we can approximate FH. As men-
tioned in Chapter 4, the isomorphous di√erence qFiso = FPH – FP can be used as
fromLattm
an&Loll:“Proteincrystallography:acon
ciseguide
”,John
Hop
kinsUniv.Press,20
08

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
The Patterson Function 79
Figure 5.4. Isomorphous di√erence Patterson map for a mercury derivative of aprotein. The crystals in this example contain hemoglobin from the annelid Glyceradibanchiata. This two-dimensional contour plot shows the v = 0 section of the fullthree-dimensional Patterson function (the u-w plane corresponds to the x-z plane inthe real unit cell). The section plotted extends over the full unit cell in w (across) andhalfway along the unit cell in u (down). The origin is at the upper left, and the largepeak there represents vectors between atoms and themselves. The peak about halfwaydown on the left, lying at approximately u= 0.45 and w = 0.04, represents a vectorbetween two symmetry-related mercury atoms. It occurs at the position 2x, 2z, wherex and z are the mercury atom coordinates in the unit cell. The refined coordinates forthe mercury atom are x = 0.225 and z = 0.007. Reproduced from the dissertation TheStructure of Glycera Hemoglobin by E. A. Padlan.
we may choose the correct arrangement of heavy atoms, or we may choose its
inverse. If the incorrect arrangement of heavy atoms is chosen, it will give rise to
an inverse image—a protein containing d-amino acids and left-handed a-helices.
Changing the sign of every phase angle will invert the handedness of the image,
and most programs contain a switch to do this. (Crystallography is capable of
determining the absolute handedness of molecules through the use of anomalous
scattering, but we won’t describe how in this book. Note that Linus Pauling,
Robert Corey, and Herman Branson, in their landmark 1951 paper describing
the structure of the a-helix, did not attempt to assign absolute handedness and
actually chose arbitrarily to draw the helix as left-handed! Later that same year
Johannes Bijvoet published his account of how the absolute configuration of
chiral molecules can be determined by using anomalous scattering.)
The example we just examined o√ers no information about the y coordinate
of the heavy atom. In this case, we are at liberty to set the y coordinate equal to
fromLattm
an&Loll:“Proteincrystallography:acon
ciseguide
”,John
Hop
kinsUniv.Press,20
08

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
14 Protein Crystallography
Figure 1.8. Illustration of how a crystal is built up by symmetric repetition of simpleelements. (a) The asymmetric unit is the smallest entity that is necessary to build upthe entire crystal. In this example, the asymmetric unit corresponds to a singlemolecule. (b) Identical copies of this molecule are generated by the space groupsymmetry operations. In the example shown, each of the four molecules in the unitcell is related to the other three by twofold (180\) rotations about one of threesymmetry axes. The three rotational symmetry axes are parallel to the unit cell edges.This type of packing arrangement is known as 222 symmetry. These four moleculescomprise the contents of the unit cell, which is shown in (c). The unit cell is a box thatencloses the various symmetry-related copies of the asymmetric unit. The edges ofthe unit cell are defined by three vectors, a, b, and c. Finally, as shown in (d), multiplecopies of the unit cells are stacked together to form the crystal, much as bricks arestacked to form a wall. Each unit cell is related to all of its neighbors by a puretranslation that constitutes an integer number of steps in a, b, and c. Kindly providedby Alexander McPherson.
means that molecules in the crystal are superimposed on copies of themselves
when reflected through a particular plane or point. Mirror planes and inversions
change the hand of objects and can therefore not be present in protein crystals,
since the amino acids comprising proteins are chiral. Finally, translations can be
combined with rotations or mirror planes to give screw axes or glide planes,
respectively.
14 Protein Crystallography
Figure 1.8. Illustration of how a crystal is built up by symmetric repetition of simpleelements. (a) The asymmetric unit is the smallest entity that is necessary to build upthe entire crystal. In this example, the asymmetric unit corresponds to a singlemolecule. (b) Identical copies of this molecule are generated by the space groupsymmetry operations. In the example shown, each of the four molecules in the unitcell is related to the other three by twofold (180\) rotations about one of threesymmetry axes. The three rotational symmetry axes are parallel to the unit cell edges.This type of packing arrangement is known as 222 symmetry. These four moleculescomprise the contents of the unit cell, which is shown in (c). The unit cell is a box thatencloses the various symmetry-related copies of the asymmetric unit. The edges ofthe unit cell are defined by three vectors, a, b, and c. Finally, as shown in (d), multiplecopies of the unit cells are stacked together to form the crystal, much as bricks arestacked to form a wall. Each unit cell is related to all of its neighbors by a puretranslation that constitutes an integer number of steps in a, b, and c. Kindly providedby Alexander McPherson.
means that molecules in the crystal are superimposed on copies of themselves
when reflected through a particular plane or point. Mirror planes and inversions
change the hand of objects and can therefore not be present in protein crystals,
since the amino acids comprising proteins are chiral. Finally, translations can be
combined with rotations or mirror planes to give screw axes or glide planes,
respectively.
fromLattman&Loll:“Proteincrystallography:aconciseguide”,JohnHopkinsUniv.Press,2008

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
W):2#1.&91#-.%$1. h%;1*$)1
W):2#1.&(1#-.%$1. bW9c&2$*$);Z.1&*=.&%.5.%1(<).&(1#-.%(Z$*(#/&#G&+8F):2#1.&$/"&+8`;)#1.&*#&+8G%:2*#1.&$/"&+8`;):)#1.3&%.1,.2*(5.);?9/&*=.&(/":1*%;3&F):2#1.&(1#-.%$1. (1&:1."&$)-#1*&.`2):1(5.);&(/&*=.&2#/5.%1(#/&#G&1*$%2=.1&*#&1:F$%1?&W9&-$;&<.&*=.&-#1*&(-,#%*$/*&#G&$))&(/":1*%($)&./Z;-.1&#G&*=.&G:*:%.?

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
g;1#Z;-.&h%;1*$)1
7=.&);1#Z;-.1&$%.&./Z;-.1&*=$*&"$-$F.&<$2*.%($)&2.))&H$))1&<;&2$*$);Z(/F&=;"%#);1(1&#G&C3L8<.*$8)(/I$F.1&<.*H../&Y8$2.*;)-:%$-(2 $2("&$/"&Y8$2.*;)8+8F):2#1$-(/.&%.1(":.1&(/&$&,.,*("#F);2$/&$/"&<.*H../&Y8$2.*;)8+8F):2#1$-(/.&%.1(":.1&(/&2=(*#".`*%(/1?&g;1#Z;-.&(1&$<:/"$/*&(/&$&/:-<.%&#G&1.2%.*(#/13&1:2=&$1&*.$%13&1$)(5$3&=:-$/&-()I3&$/"&-:2:1?

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
!`2.)1(/ h%;1*$)1
!`.)2(/ (1&$&F)#<:)$%&,%#*.(/&G#:/"&(/&1#-.&,)$/*1?&9*&(1&$/&(/1.2*&,#(1#/&$/"&,%.5./*1&(/1.2*&G.."(/F?

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Introduction 13
Figure 1.7. Gallery of pictures illustrating a variety of protein crystals. Note the widevariety of shapes. Note also the variation in the perfection of the external appearanceof the crystals, which is not necessarily related to perfection in internal order. Thenames of the proteins appear in the individual frames. Kindly provided by AlexanderMcPherson.
inversions, and translations. In a crystal possessing rotational symmetry, every
molecule in the crystal is superimposed on an identical copy of itself when
rotated by a specific angle (for example, 180\) about a particular axis. Allowed
rotational symmetries are twofold (180\), threefold (120\), fourfold (90\), and
sixfold (60\). Note that fivefold symmetry is not allowed in crystals, nor is
sevenfold symmetry or higher. When we say they are not allowed, we mean that it
is physically impossible to build up a repeating three-dimensional array that is
based on fivefold or sevenfold symmetry. Mirror symmetry or inversion symmetry
agalleryofp
roteins,fro
mLattm
an&Loll:“Proteincrystallography:a
concise
guide
”,John
Hop
kinsUniv.Press,20
08

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
Q*%:2*:%.&#G&=./&.FF8H=(*.&);1#Z;-.

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
???&G#))#H&*=.&)(/I&=**,JMMHHH?%21<?#%FM,"<M=#-.M=#-.?"#

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
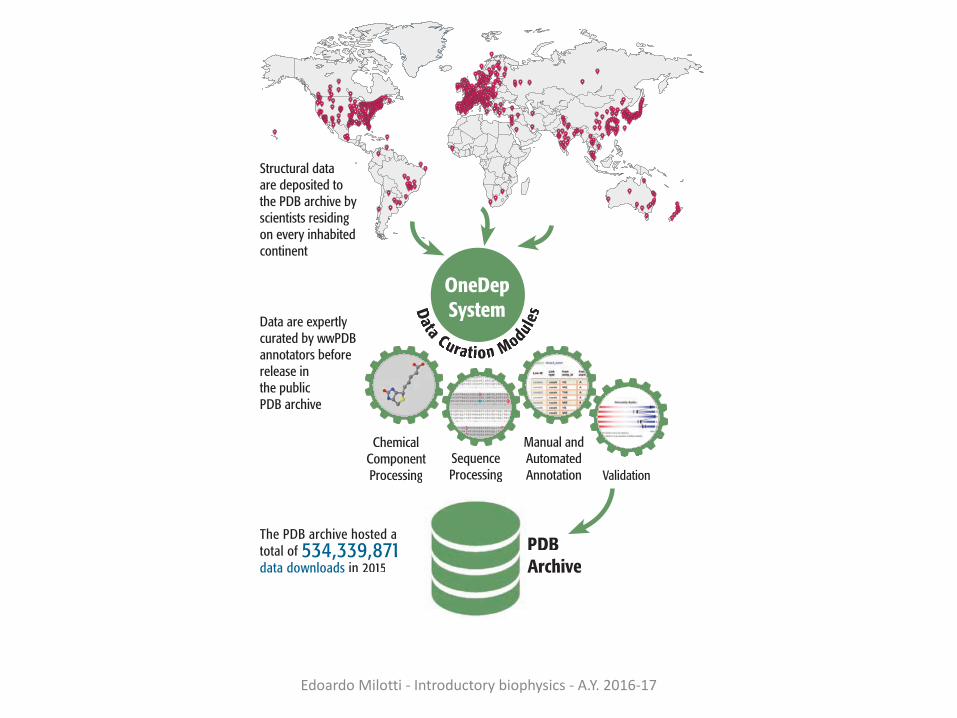
The PDB archive hosted a total of 534,339,871 data downloads in 2015
Structural data are deposited to the PDB archive by scientists residing on every inhabited continent
Data are expertly curated by wwPDB annotators before release in the public PDB archive
OneDepSystemData Curarar tion Modul
esPDB Archive
DATA DEPOSITION AND ANNOTATION
In 2015, the wwPDB curated 10,956 deposited structures. The PDB archive is on track to receive ~12,000 new structures in 2016
ChemicComponentProcessing
SequenceProcessing
Manual and Automated Annotation Validation
al
Ligand Validation Workshop Participants

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
=**,JMMH
HH?%21<?#%FM,"<M1*$*(2?"#w,PF./.%$)m(/G#%-
$*(#/M,"<m1*$*(1*(21M(/".`?=*-)

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
ABCNJ&KENK&1*%:2*:%.1ABCLJ&KKEN&1*%:2*:%.1ABCOJ&KDEN&1*%:2*:%.1ABCDJ&CBBCD&1*%:2*:%.1ABCE&bNcJ&NCNT&1*%:2*:%.1
!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
=**,JMMH
HH?%21<?#%FM,"<M1*$*(2?"#w,PF./.%$)m(/G#%-
$*(#/M,"<m1*$*(1*(21M(/".`?=*-)

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE
ABCNJ&CCK&1*%:2*:%.1ABCLJ&OAD&1*%:2*:%.1ABCOJ&ECO&1*%:2*:%.1ABCDJ&TAK&1*%:2*:%.1ABCE&bNcJ CAC&1*%:2*:%.1
=**,JMMH
HH?%21<?#%FM,"<M1*$*(2?"#w,PF./.%$)m(/G#%-
$*(#/M,"<m1*$*(1*(21M(/".`?=*-)
!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE

!"#$%"#&'()#**(&8 9/*%#":2*#%;&<(#,=;1(21&8 >?@?&ABCD8CE

EdoardoMilotti- Introductorybiophysics- A.Y.2016-17
Homework:
1. Accessthewebsitehttp://www.ks.uiuc.edu anddownloadVMD,themoleculardynamicsvisualizer(lookfordocumentation,quickstart,instructions,etc.onthesamesite)
2. Gotohttp://www.rcsb.org anddownloadPDBentry1fbb (bacteriorhodopsin)
3. RunVMD,loadfile1fbb,andfindawaytodisplayastereoviewofthemoleculeintheball-and-stickrepresentation
4. Printtheresultandbaskinthelightofyoursuccess







