
A quantitative view on mRNA translation: the relative role of initiation and elongation
Luca Ciandrini ([email protected])University of Montpellier, France
13th June 2016Quantitative Laws II, Villa del Grumello, Como
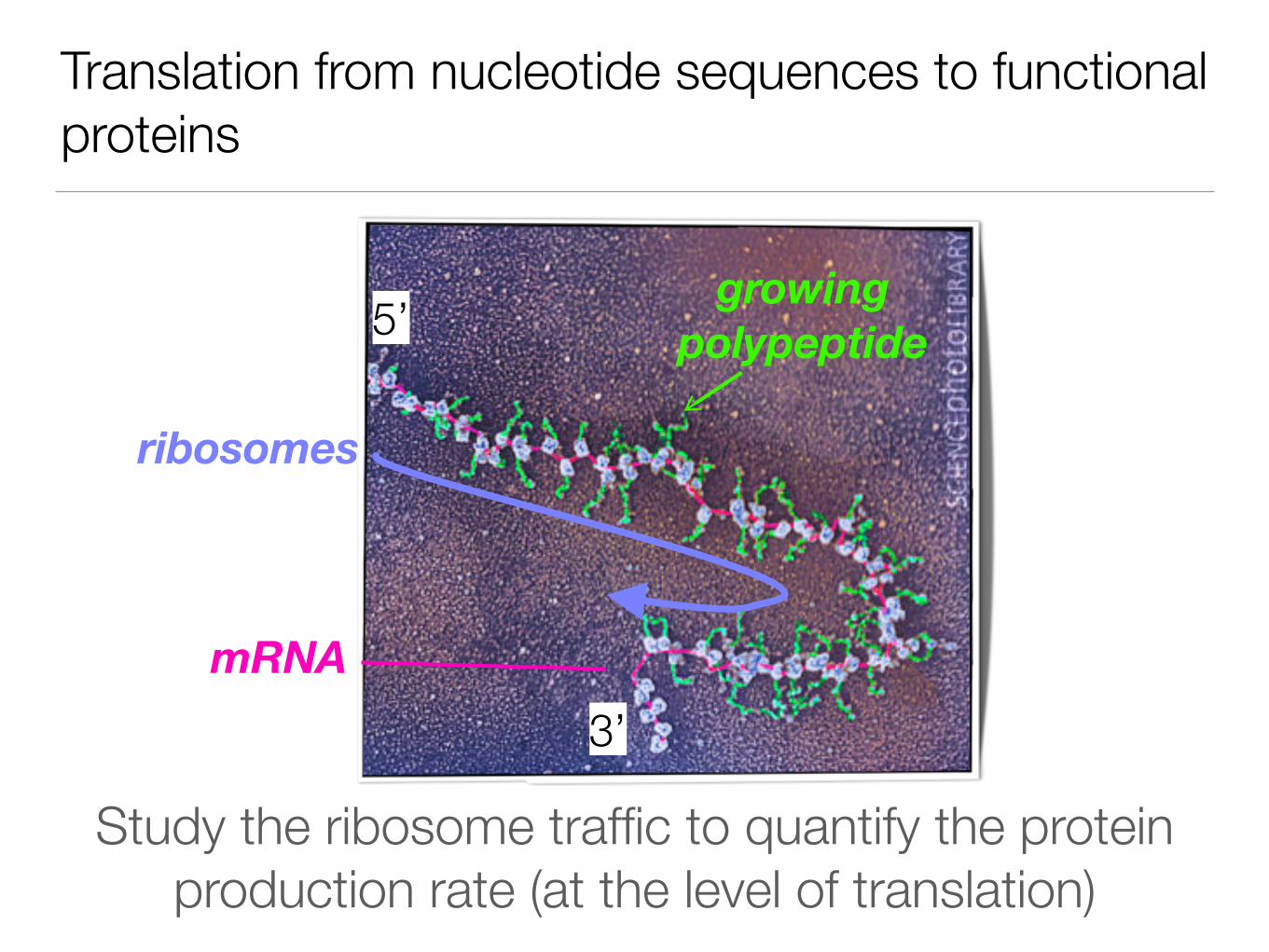
Translation from nucleotide sequences to functional proteins
Study the ribosome traffic to quantify the protein production rate (at the level of translation)
ribosomes
growing polypeptide
mRNA
5’
3’
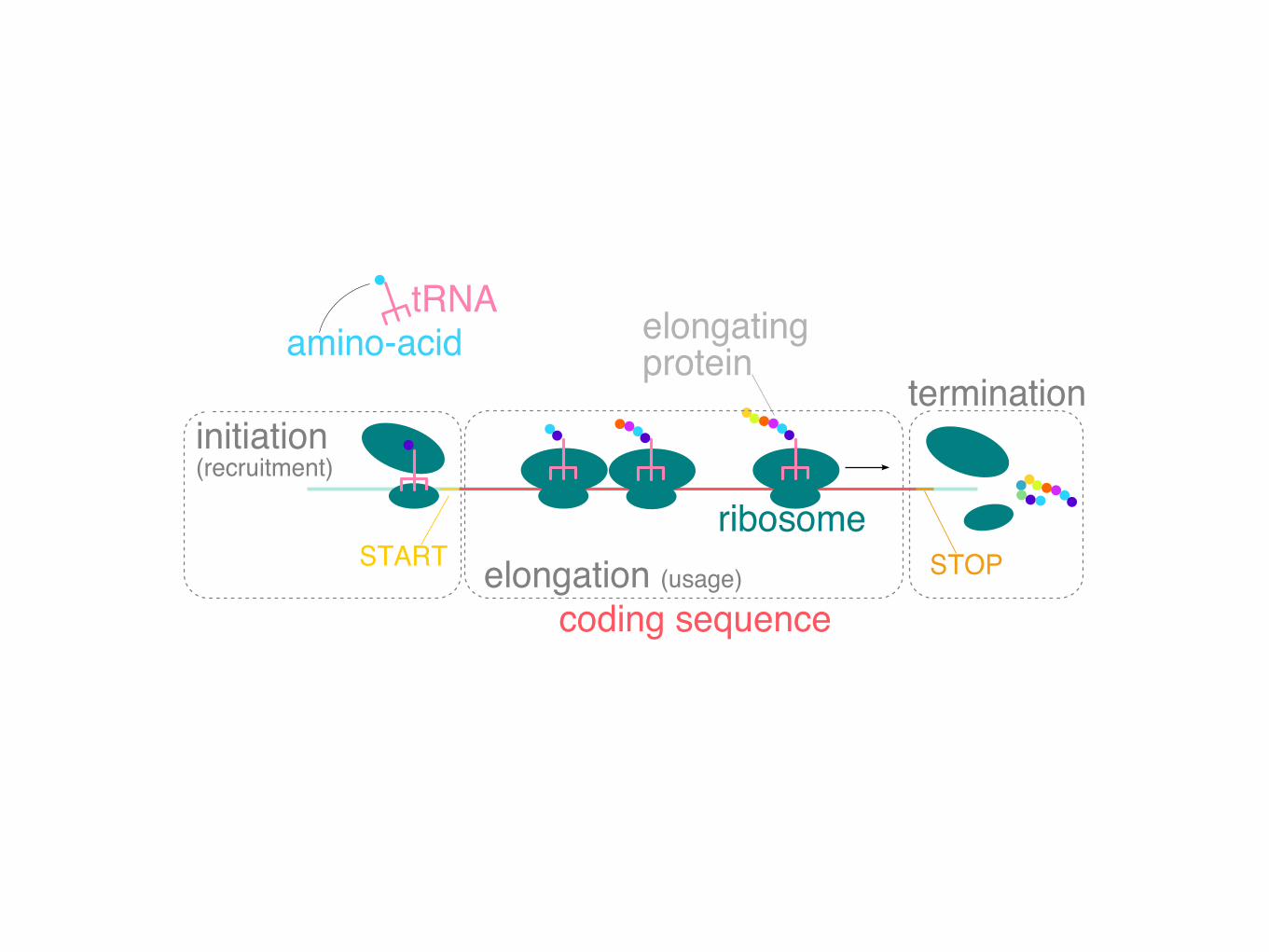
coding sequence
ribosomeSTART
elongatingprotein
initiation(recruitment)
tRNAamino-acid
elongation (usage)
termination
STOP
initiation(recruitment)
elongation (usage)
termination

coding sequence
ribosomeSTART
elongatingprotein
initiation(recruitment)
tRNAamino-acid
elongation (usage)
termination
STOP
initiation(recruitment)
elongation (usage)
termination
How can we predict/vary the strength of initiation?
Ribosome recruitment Ribosome usage
How depletion of ribosomes affects gene expression and growth rates?
Codon usage and bias? Evolution? Does noise depend
on codon sequences, etc etc?
Can we tune and predict, just by varying the sequence, “optimal” protein production rates?P. Geulich, L. Ciandrini, R.J. Allen and M. C. Romano, Phys Rev E (2012)
Shah et al, Cell (2013)

Flow of ribosomes ~ driven lattice gas
Macdonald et al., Biopolymers (1968)Totally Asymmetric Simple Exclusion Process
coding sequence
ribosomeSTART
elongatingprotein
initiation(recruitment)
tRNAamino-acid
elongation (usage)
termination
STOP
initiation(recruitment)
elongation (usage)
termination

The model can predict translation efficiency depending on initiation and elongation rates
ribosomal current (mRNA efficiency)
initiation rate codon dependent elongation rates
termination rate
L. Ciandrini, I. Stansfield and M. C. Romano, PLoS Comp Bio (2013)
initia
tion
lim
ited elongation
limited

The model can predict ribosome density depending on initiation and elongation rates
ribosomal density
initiation rate codon dependent elongation rates
termination rate
L. Ciandrini, I. Stansfield and M. C. Romano, PLoS Comp Bio (2013)
initia
tion
lim
ited ribosomes
queueing

1. How initiation affects mRNA efficiency
L. Dias Fernandes, A. de Moura, L. Ciandrini, to be submitted J.C. Walter, L. Ciandrini, in preparation
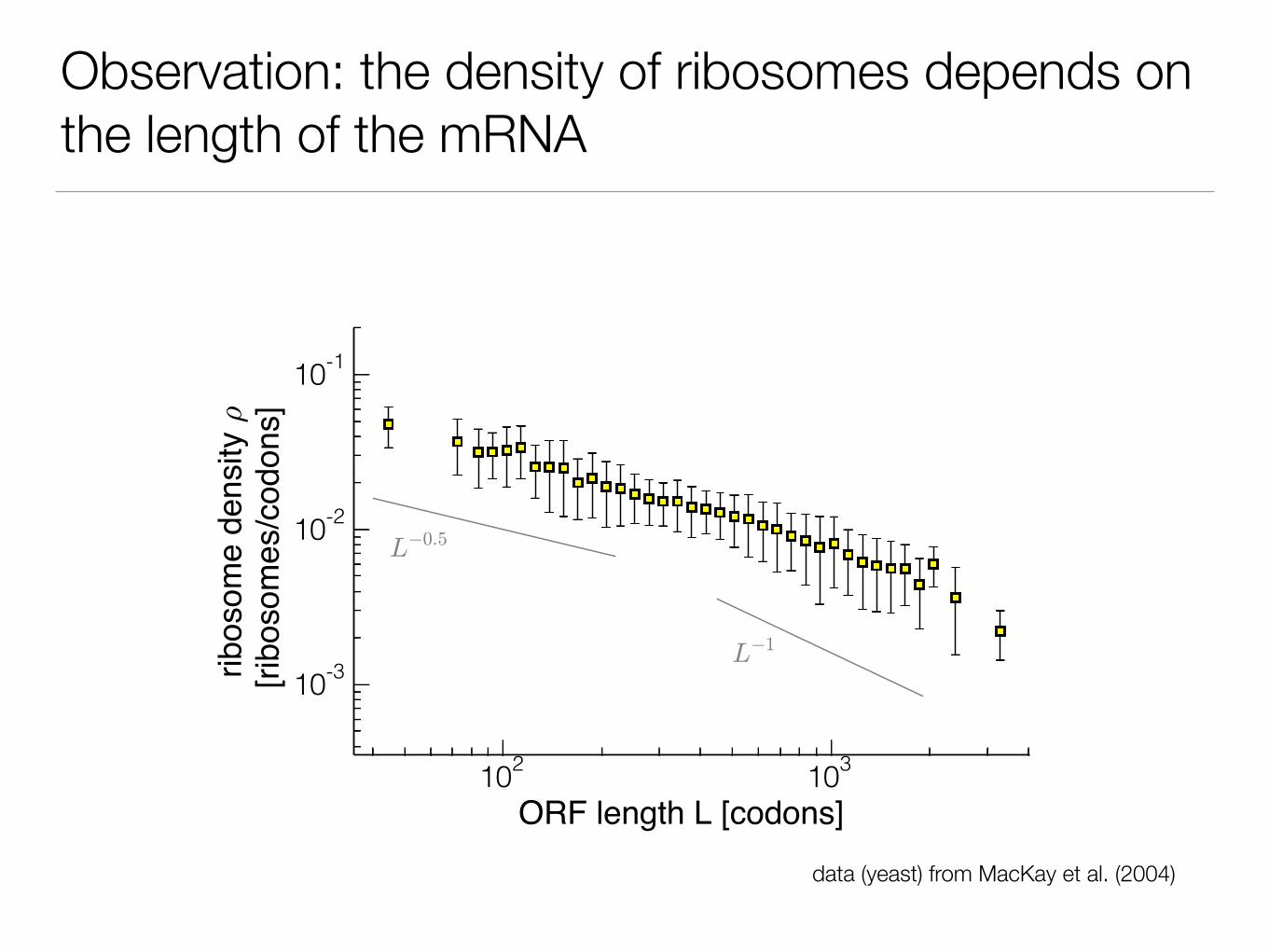
Observation: the density of ribosomes depends on the length of the mRNA
data (yeast) from MacKay et al. (2004)

The local concentration of ribosomes determines translation initiation
↵ = ↵o
c = ↵o
(c1 + cR
)
initiation rate
overall ribosome concentration
contribution due to terminating ribosomes
(feedback)
Initiation depends on local concentrations of ribosomes
R
r
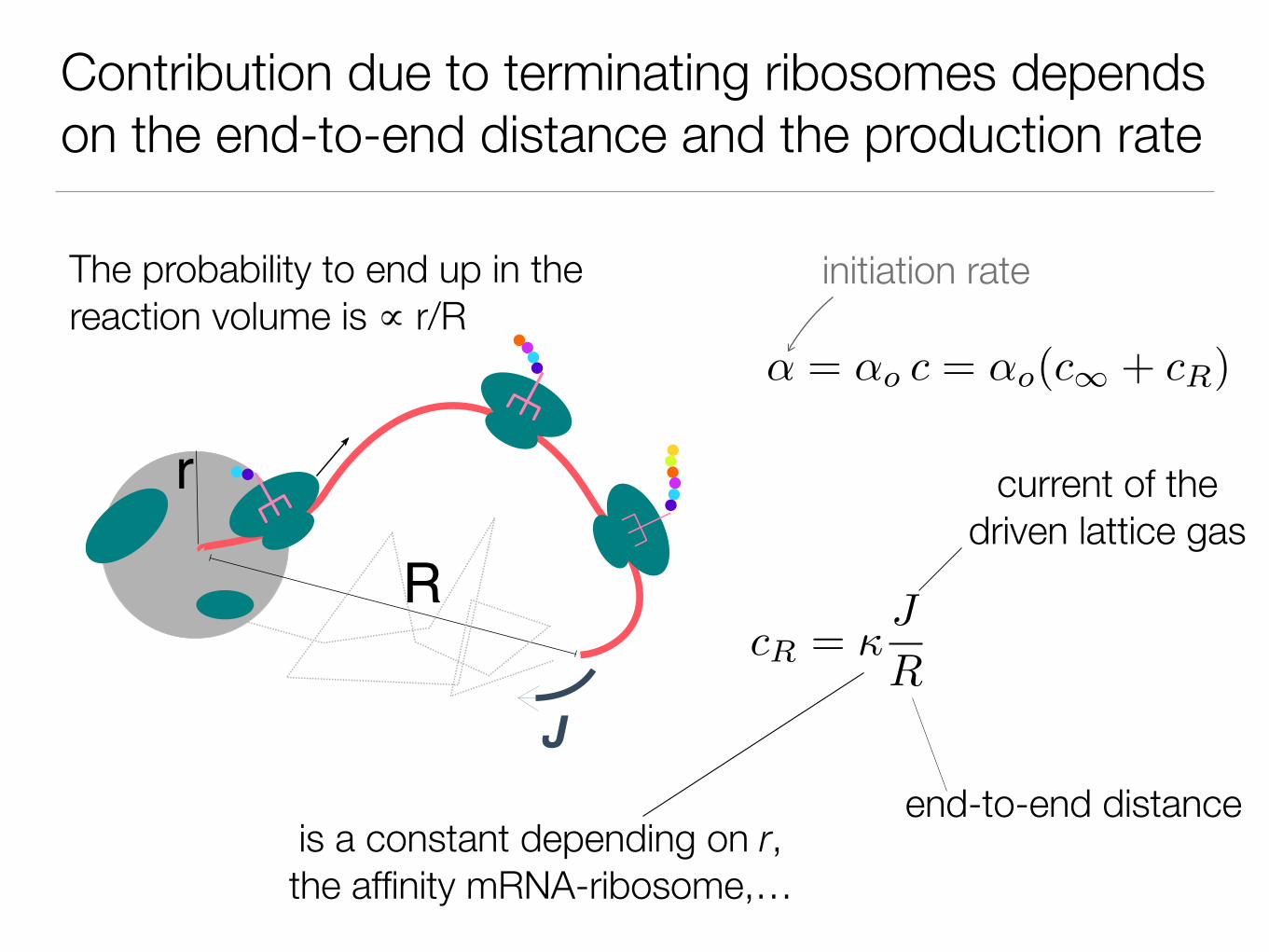
Contribution due to terminating ribosomes depends on the end-to-end distance and the production rate
↵ = ↵o
c = ↵o
(c1 + cR
)
initiation rate
R
r
The probability to end up in the reaction volume is ∝ r/R
J
current of the driven lattice gas
end-to-end distanceis a constant depending on r, the affinity mRNA-ribosome,…

The mRNA persistence length depends on the ribosomal density
R ⇠p
`pL R ⇠q`polyp L
`p= persistence length `polyp = f`+ (1� f)`p
f = fraction of transcript occupied by ribosomes

The initiation rate α is therefore length-dependent (via “recycling” and finite-size effects)
• Fix a tentative initiation rate
• Simulate the system and extract J and R
• Compute the initiation rate
• Compare to the tentative oneJ
We develop a self-consistent method to compute the initiation rate

initiation rate:

L. Dias Fernandes, A. De Moura, L. Ciandrini, to be submitted
finite size + local concentration
The theory is consistent with the ribosome density data
initiation rate:
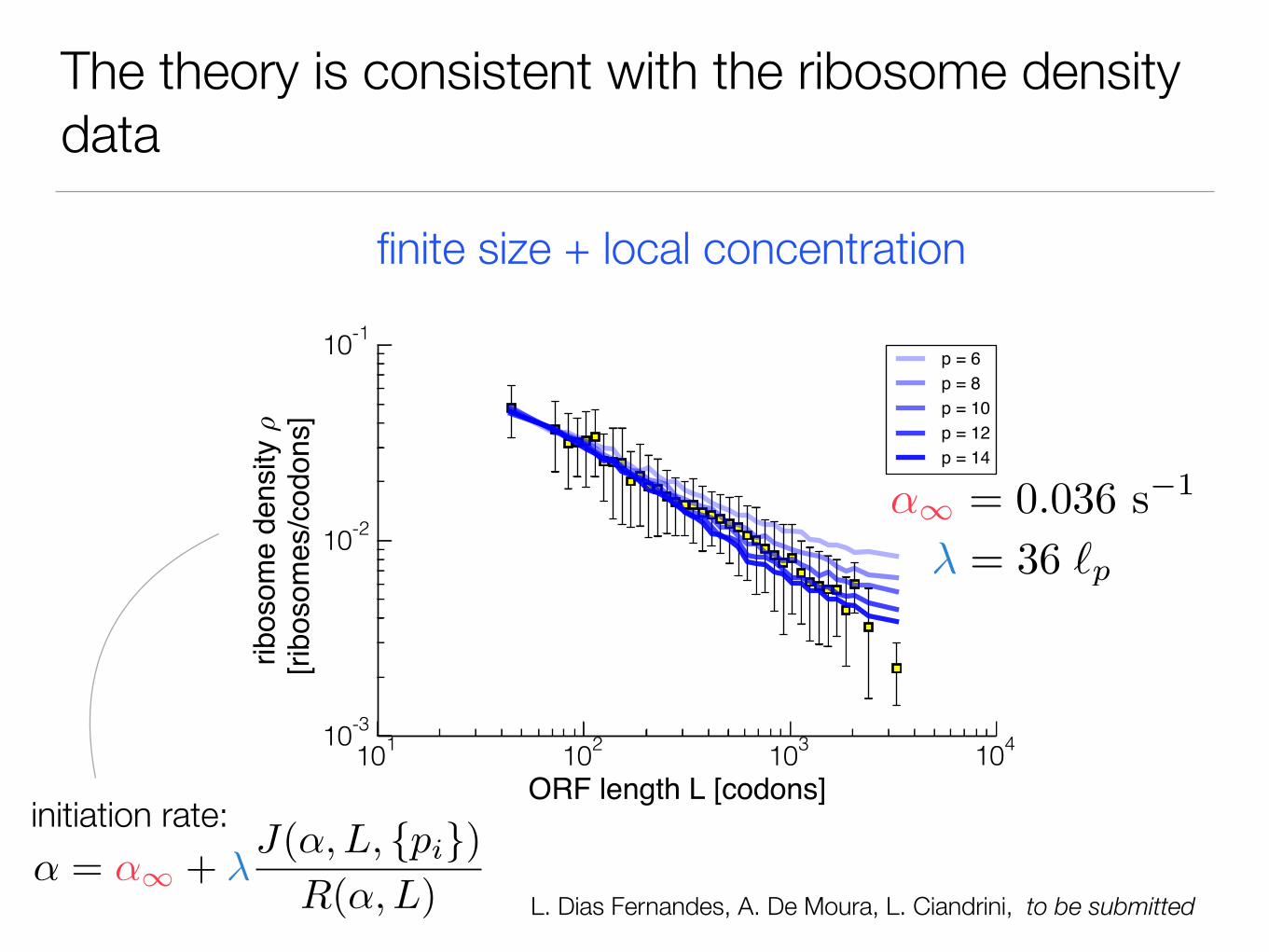
The theory is consistent with the ribosome density data
finite size + local concentration
initiation rate:
L. Dias Fernandes, A. De Moura, L. Ciandrini, to be submitted
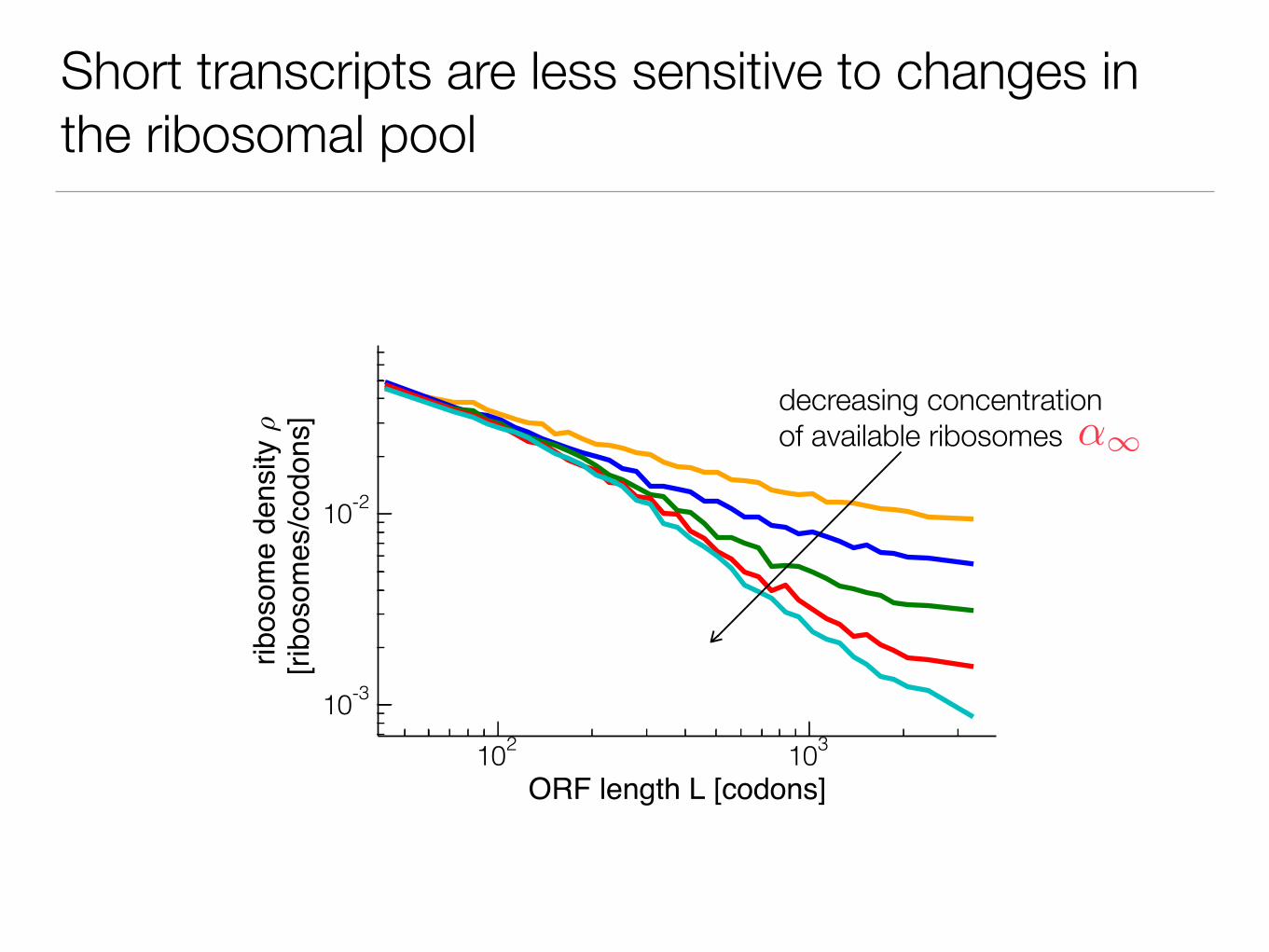
Short transcripts are less sensitive to changes in the ribosomal pool
decreasing concentration of available ribosomes
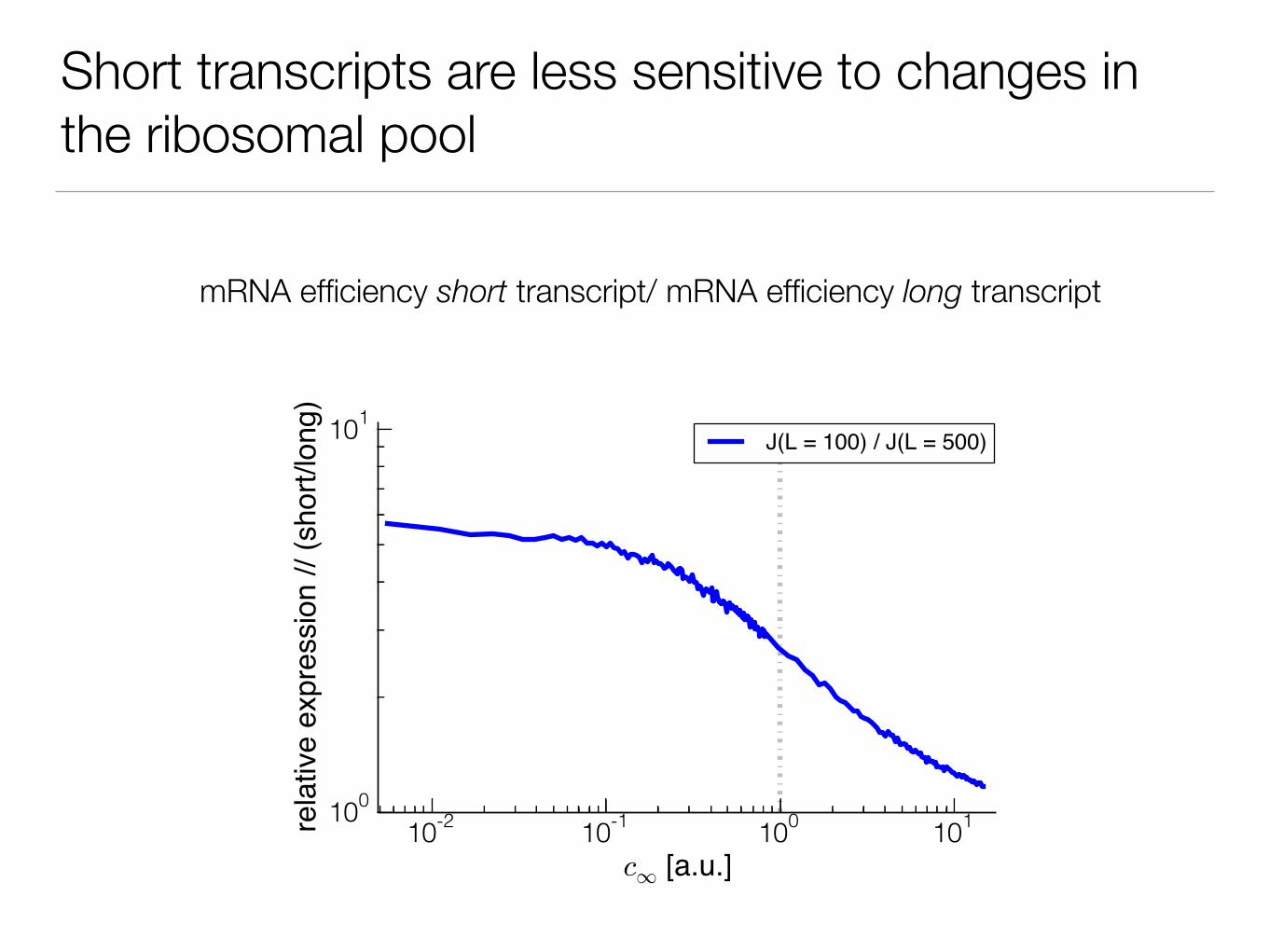
Short transcripts are less sensitive to changes in the ribosomal pool
mRNA efficiency short transcript/ mRNA efficiency long transcript

2. How elongation affects mRNA efficiency
AJ Kemp, R Betney, LC, ACM Schwenger, MC Romano,I Stansfield, Molecular Microbiology (2013) B Gorgoni, L. Ciandrini, M Mc Farland, M C Romano, I Stansfield, submitted Y. Meriguet, S. Guiziou, J. Bonnet, L. Ciandrini, in preparation
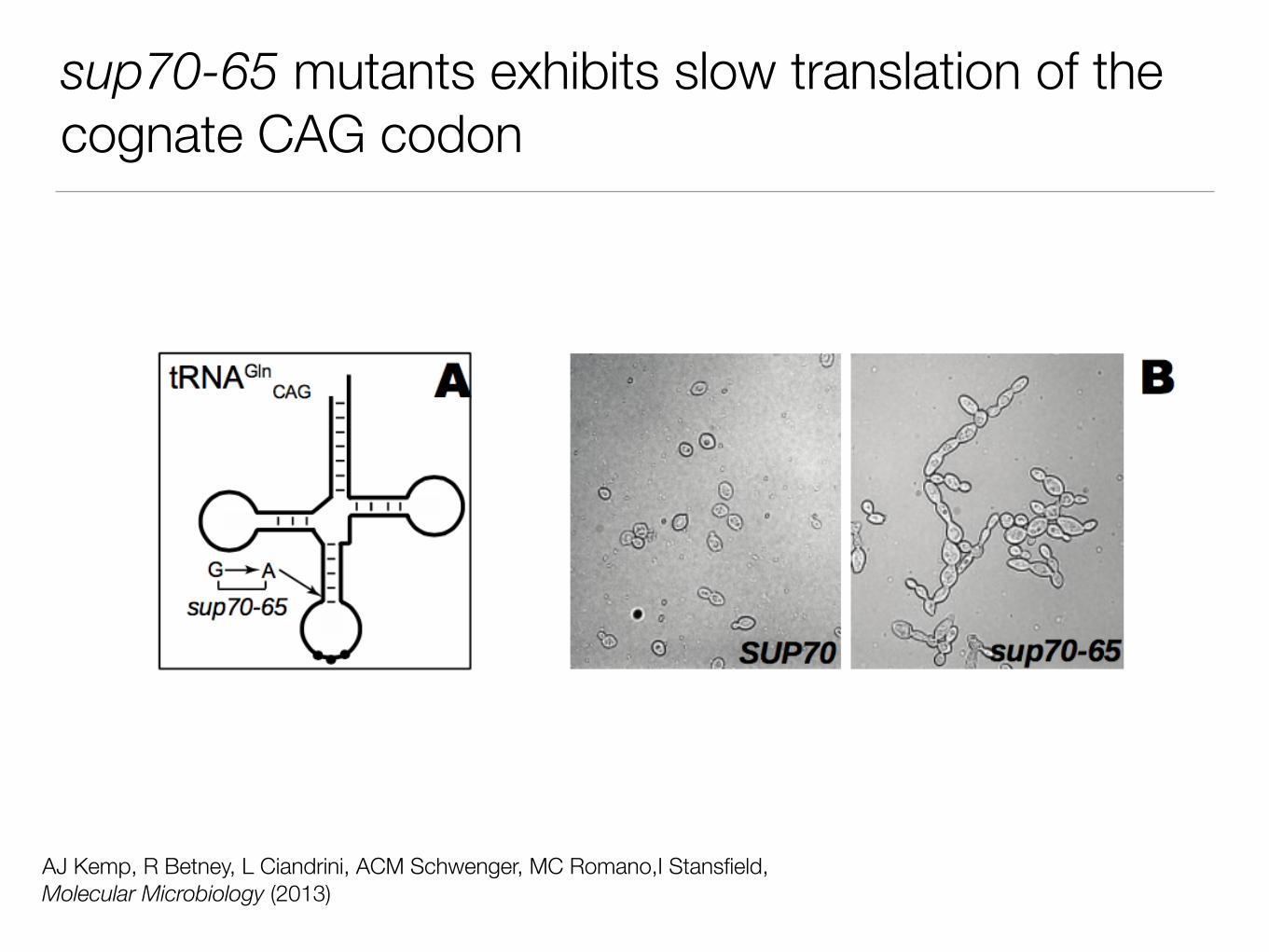
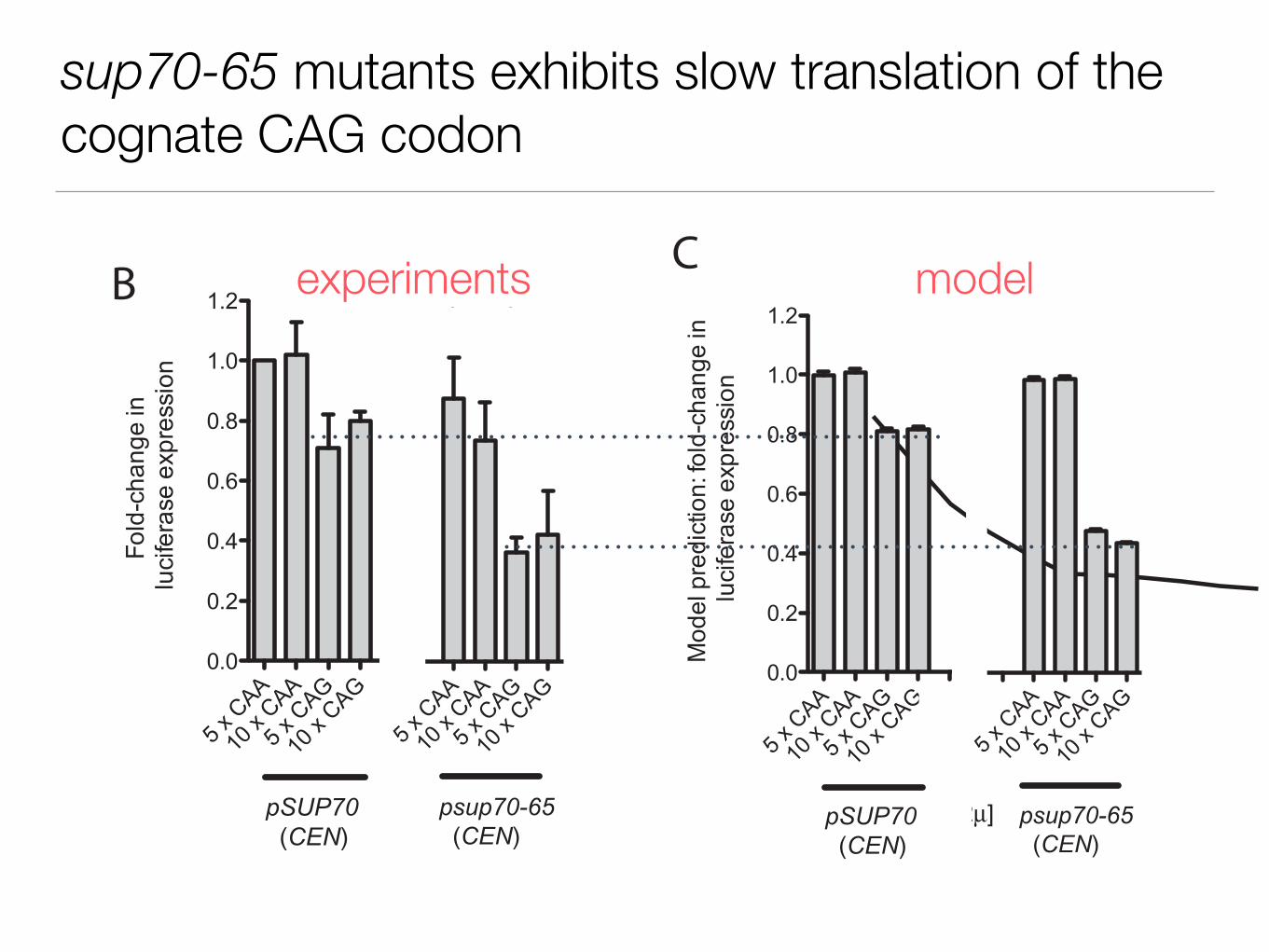
sup70-65 mutants exhibits slow translation of the cognate CAG codon
AJ Kemp, R Betney, L Ciandrini, ACM Schwenger, MC Romano,I Stansfield, Molecular Microbiology (2013)

How do we affect translation if we insert CAA and CAG repeats at the 5’end?
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
rib
oso
ma
lde
nsi
ty
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
ribo
som
ald
en
sity
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
ribo
som
ald
en
sity
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on5 x
CAA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
rib
oso
ma
lde
nsi
ty
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
rib
oso
ma
lde
nsi
ty
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
modelexperiments
sup70-65 mutants exhibits slow translation of the cognate CAG codon
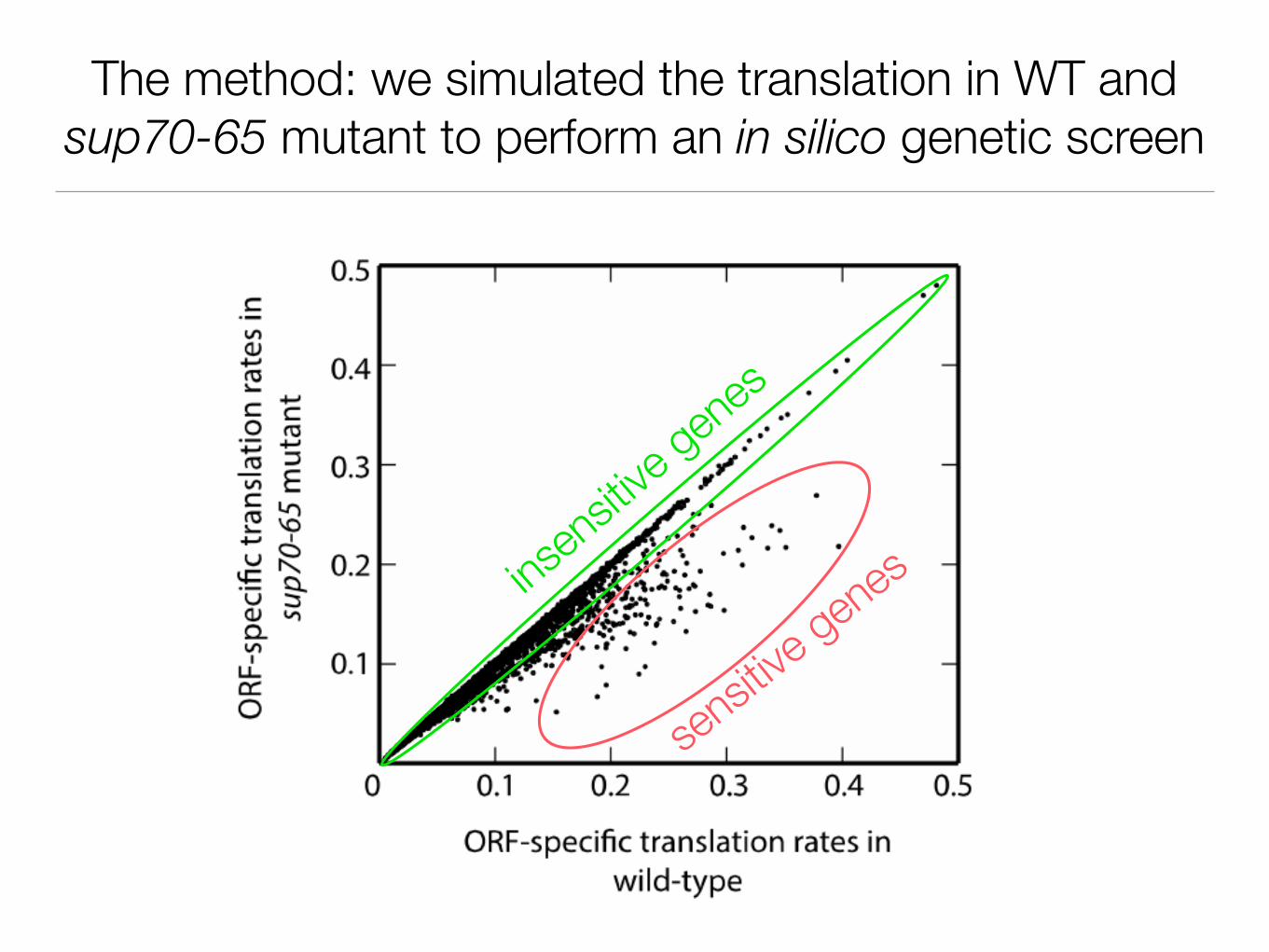
The method: we simulated the translation in WT and sup70-65 mutant to perform an in silico genetic screen
sensitiv
e genesins
ensitiv
e gene
s
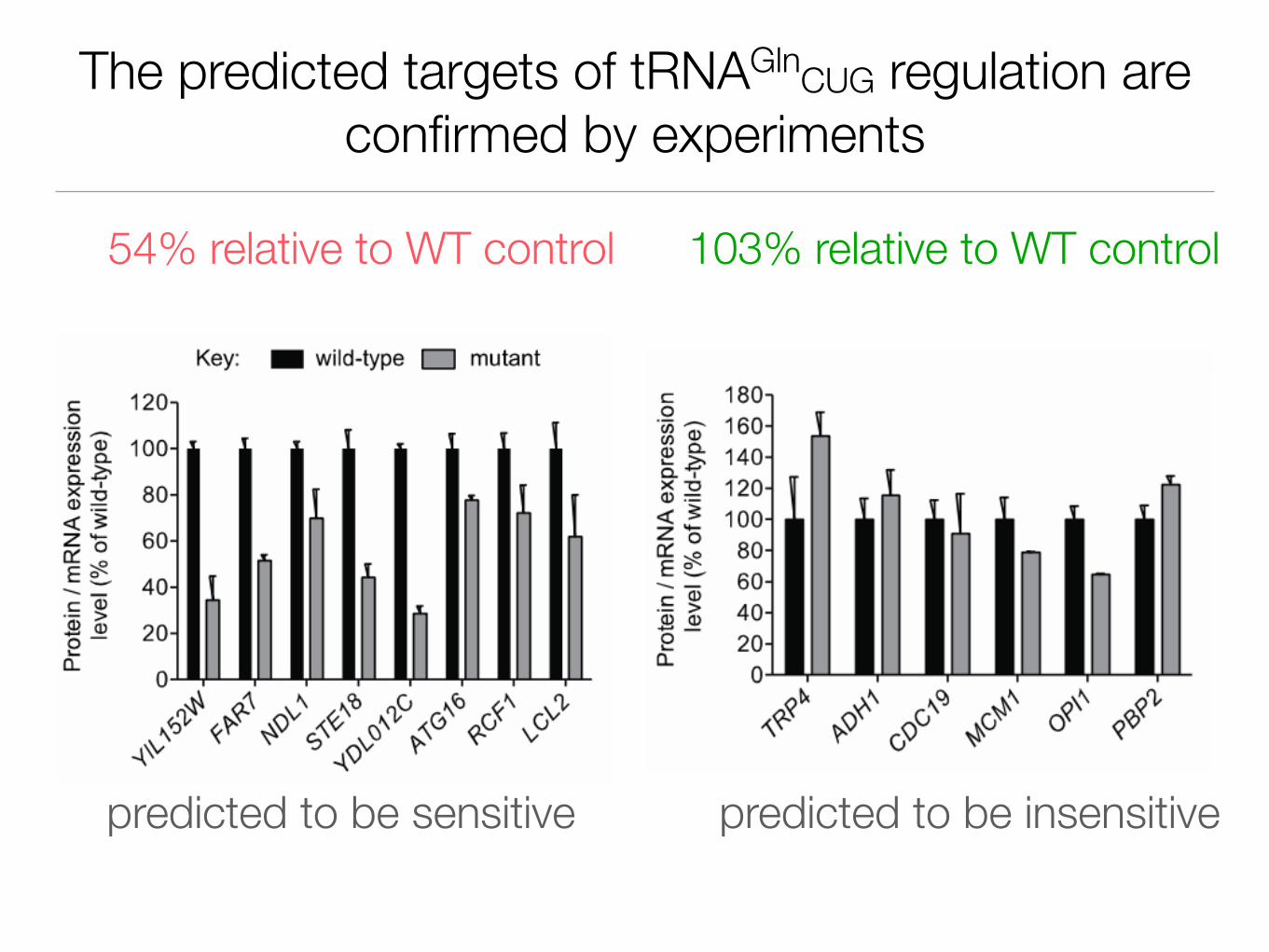
The predicted targets of tRNAGlnCUG regulation are confirmed by experiments
54% relative to WT control 103% relative to WT control
predicted to be sensitive predicted to be insensitive
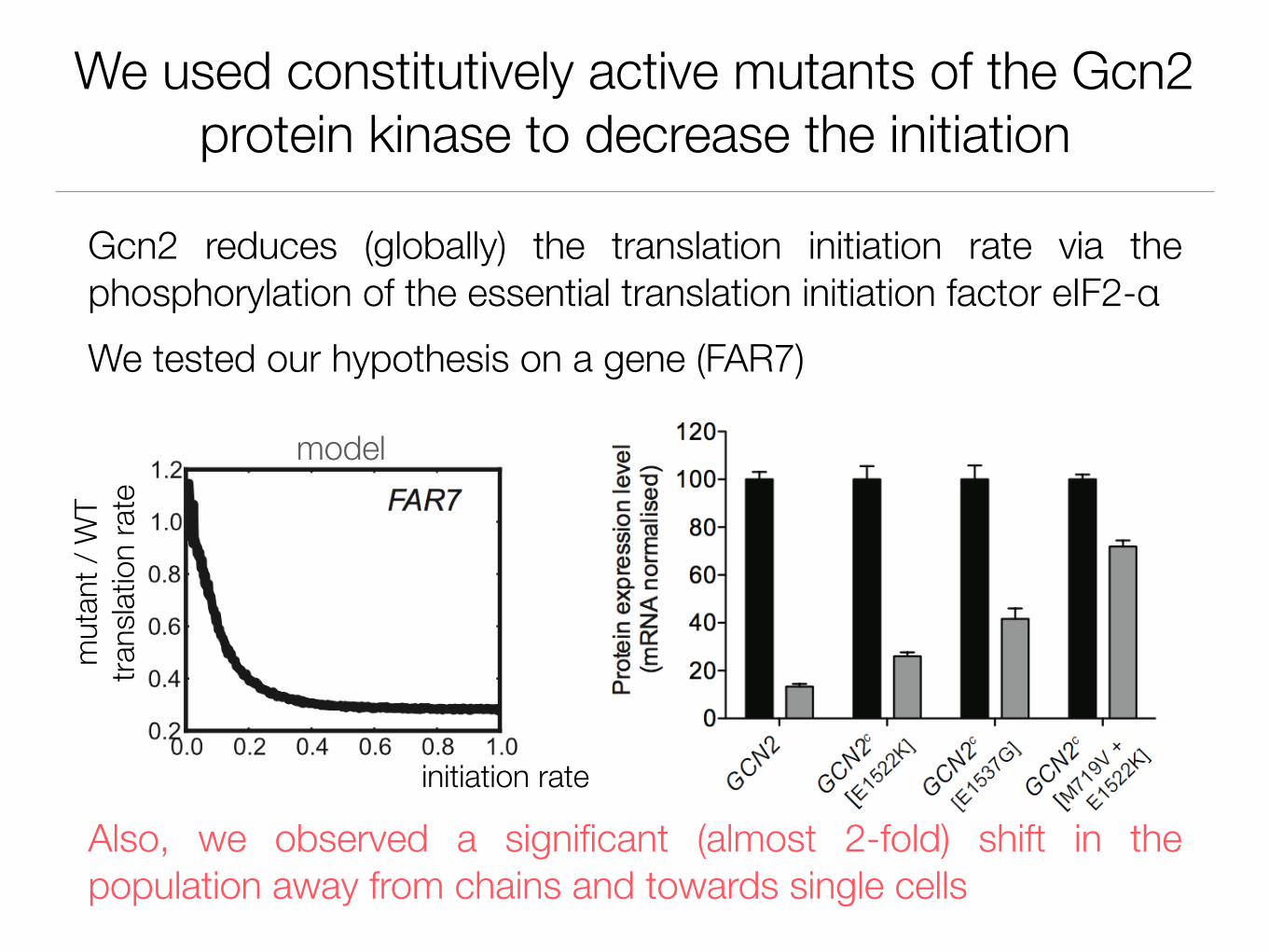
We used constitutively active mutants of the Gcn2 protein kinase to decrease the initiation
Gcn2 reduces (globally) the translation initiation rate via the phosphorylation of the essential translation initiation factor eIF2-αWe tested our hypothesis on a gene (FAR7)
Also, we observed a significant (almost 2-fold) shift in the population away from chains and towards single cells
initiation rate
mut
ant /
WT
trans
latio
n ra
te
model

Conclusions
• Initiation might be the origin of the ribosome density/
mRNA length dependence
• Elongation can be used to regulate gene expression
through the tuning of single-codon elongation rates
• Both initiation and elongation can be limiting steps
of translation
Further perspectives
• How to independently study and modulate initiation and elongation?
…undergoing experiments…

Acknowledgements
Thank you
Ian StansfieldBarbara GorgoniIMS, University of Aberdeen
Jean-Charles WalterL2C, University of Montpellier
soutien UM Défi InPhyNiTi (Interfaces physiques Numérique et Théorique)
Sarah GuiziouJerome BonnetCBS, University of Montpellier
Yoann MeriguetDIMNP, University of Montpellier
Lucas Dias FernandesAlessandro de MouraMaria C RomanoICSMB, University of Aberdeen


The relative positioning of codons (elongation) affects translational efficiency
Shuffling synonymous codons does not affect the final product (aa-sequence) and keeps the same codon usage
(codon used in a sequence).
original
synonymous

The relative positioning of codons (elongation) affects translational efficiency
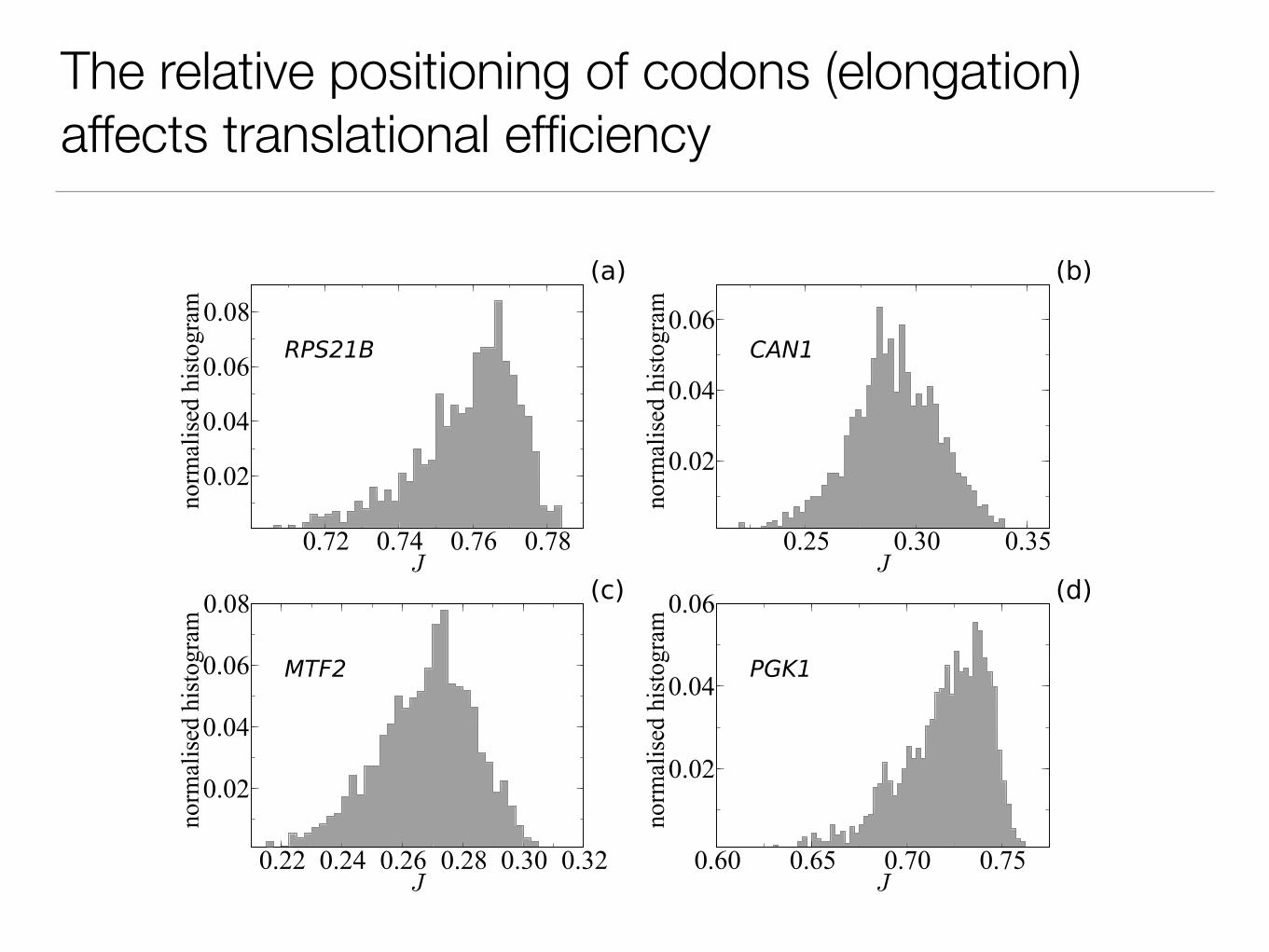
The relative positioning of codons (elongation) affects translational efficiency

We are going to design sequences with high, medium and small translation efficiencies and test them

The limited amount of resources constrains translation efficiency*
*Translation efficiency = overall rate of translation per mRNAScott et al., Science 2010
There is evidence that the amount of ribosomes restrains translation efficiency (see works by Scott, Klumpp, Hwa; Shah et al. from Plotkin-Kudla groups).
Competition for translational resources affects translation

The limited amount of resources constrains translation efficiency*
*Translation efficiency = overall rate of translation per mRNAScott et al., Science 2010
There is evidence that the amount of ribosomes restrains translation efficiency (see works by Scott, Klumpp, Hwa; Shah et al. from Plotkin-Kudla groups).
Competition for translational resources affects translation
Can competition for resources be exploited to regulate gene expression?

Conclusions
• Finite-size effects are relevant in translation
• The three-dimensional conformation of the polysome might enhance initiation (ribosome
“recycling”)
• The selfish mRNA: is recycling a way to build up a
personal pool of ribosomes?

2. Use strep-tag at the 5‘end to detect the
queue
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
ribo
som
ald
en
sity
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
ribo
som
ald
en
sity
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on5 x
CAA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
rib
oso
ma
lde
nsi
ty
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
rib
oso
ma
lde
nsi
ty
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
A.J. Kemp, R. Betney, L. Ciandrini, A.C.M. Schwenger, M.C. Romano, I. Stansfield, Molecular Microbiology (2013)
Good agreement with theory
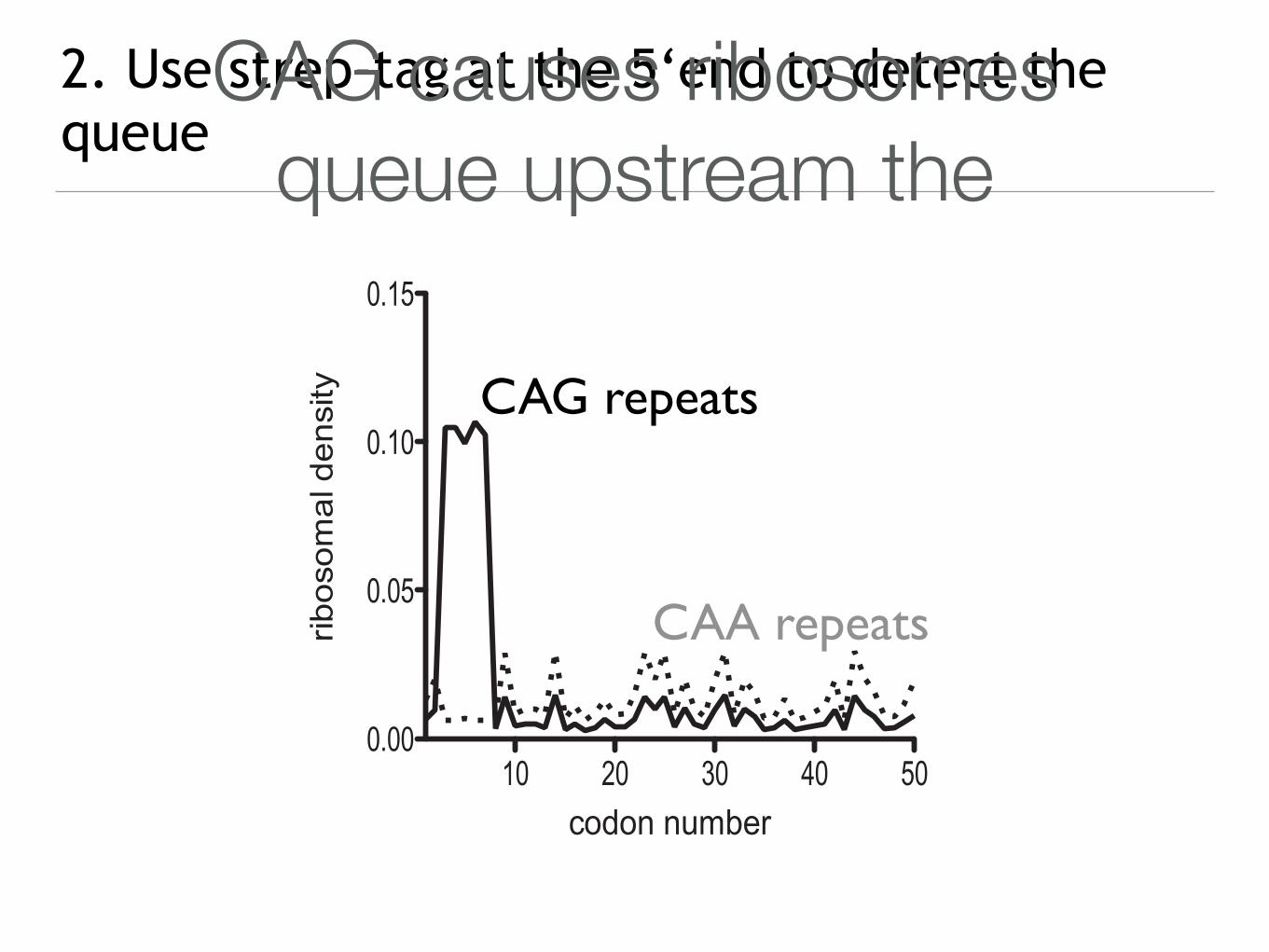
2. Use strep-tag at the 5‘end to detect the
queueCAG causes ribosomes
queue upstream the
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
SUP70wild-type
multicopytRNACUG
sup70-65
Fold
-cha
nge
inlu
cife
rase
expr
essi
on
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG
5 x C
AA
10 x
CAA
5 x C
AG
10 x
CAG0.0
0.2
0.4
0.6
0.8
1.0
1.2
Mod
elpr
edic
tion:
fold
-cha
nge
inlu
cife
rase
expr
essi
on
B
C
A
luc
luc
luc
luc
5 x CAA
10 x CAA
5 x CAG
10 x CAG
*** *********
*** *********
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
pSUP70 (CEN)
pSUP70-[2µ]
psup70-65 (CEN)
0.0 0.5 1.0 1.5 2.0 6 9 12 150.00
0.02
0.04
0.06
0.08
0.10
Fold concentration of tRNACUGGln
relative to wild type
luci
fera
seex
pres
sion
rate
10 20 30 40 500.00
0.05
0.10
0.15
codon number
rib
oso
ma
lde
nsi
ty
0 2 4 6 8 100.020
0.040
0.060
0.080
Number of CAG codons in tandem repeats
Luci
fera
seex
pres
sion
rate
D
F
E
Slow CAG translation in yeast pseudohyphal growth mutants 293
© 2012 Blackwell Publishing Ltd, Molecular Microbiology, 87, 284–300
CAG repeats
CAA repeats
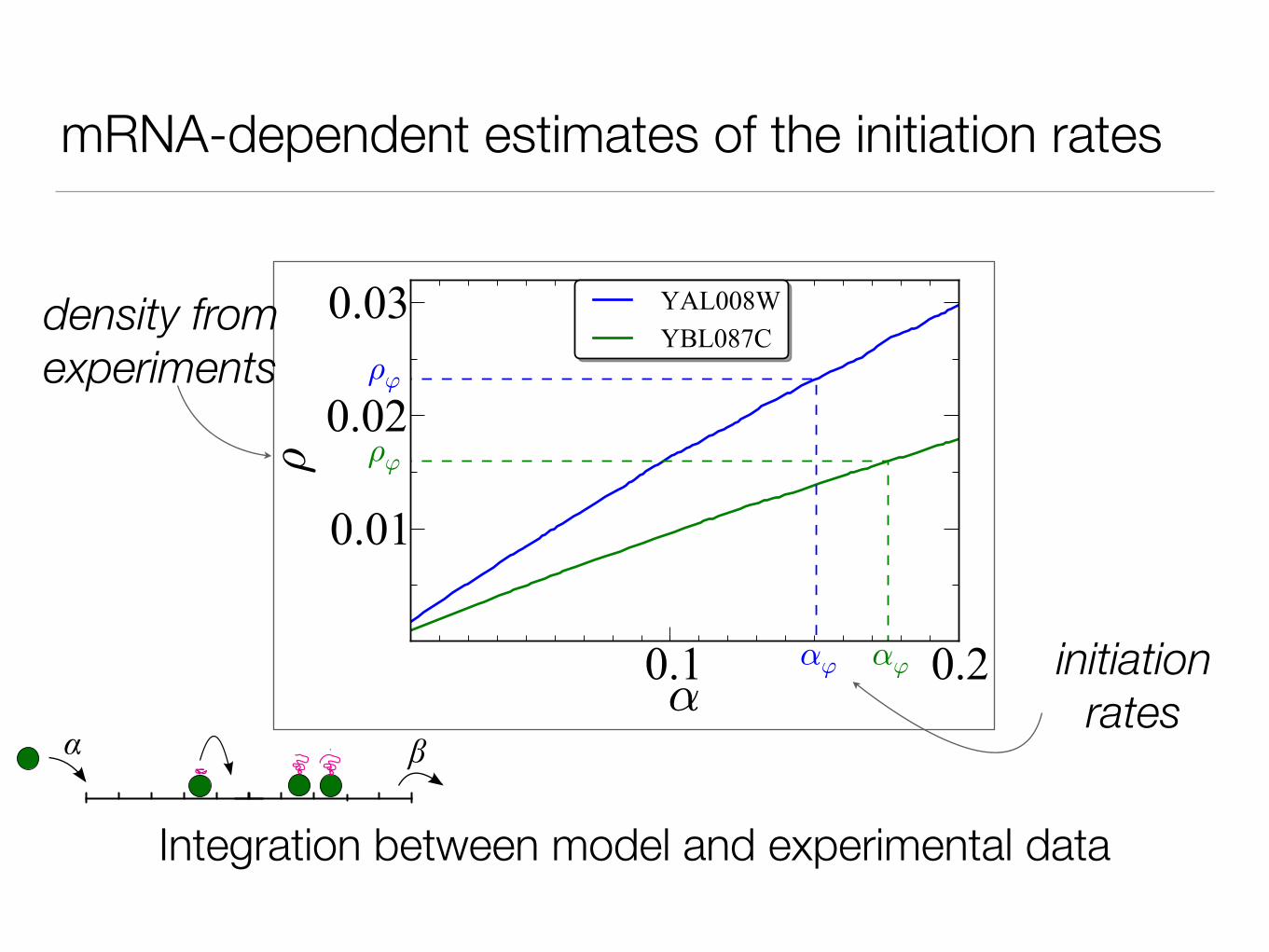
mRNA-dependent estimates of the initiation rates
density from experiments
initiation rates
Integration between model and experimental data
=
=
1 site = 1 codon
mRNA 1D-lattice
ribosomeparticle
initiation elongationtermination
α βγ
γ
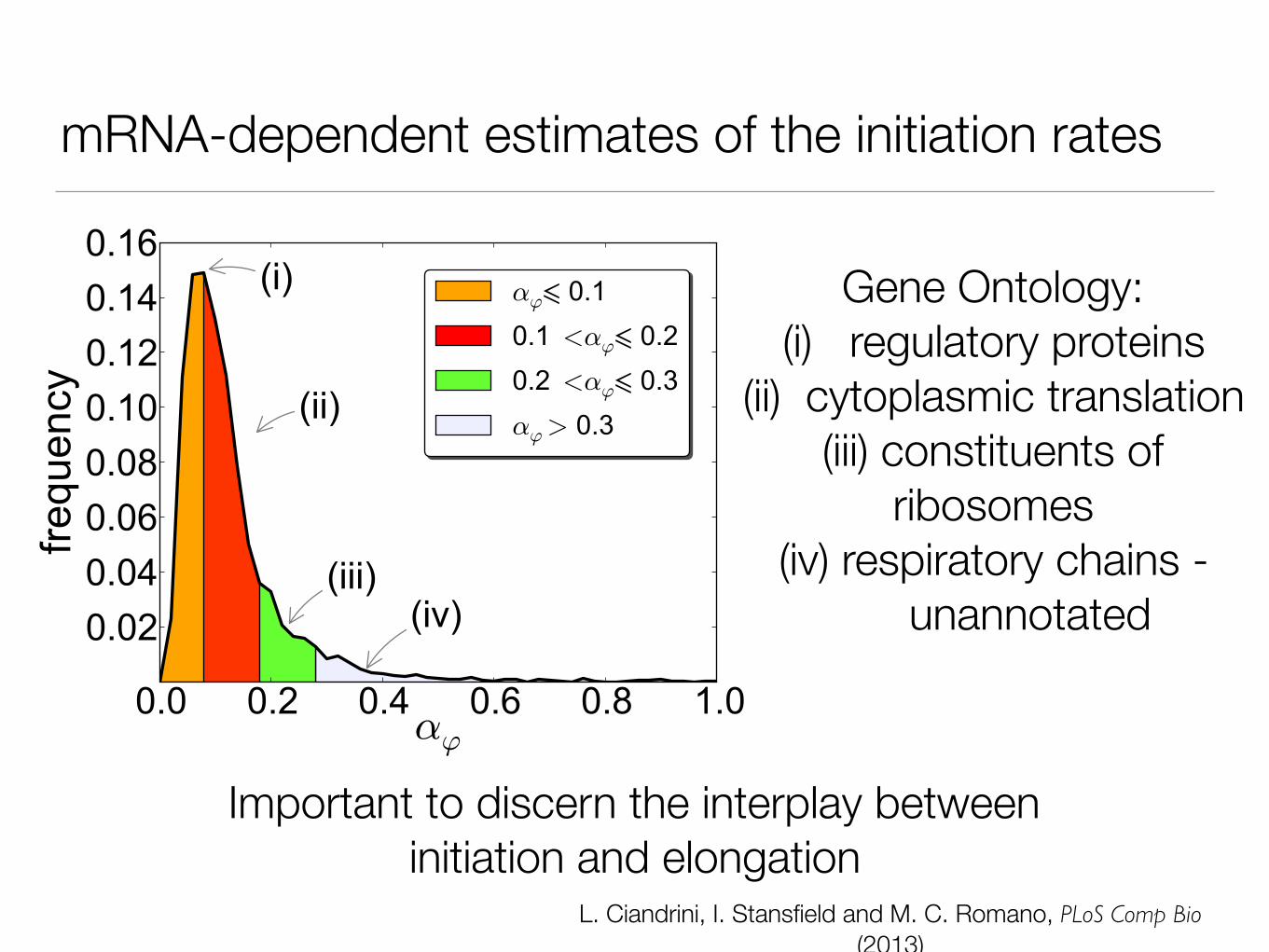
mRNA-dependent estimates of the initiation rates
Important to discern the interplay between initiation and elongation
Gene Ontology: (i) regulatory proteins
(ii) cytoplasmic translation (iii) constituents of
ribosomes (iv) respiratory chains -
unannotated
L. Ciandrini, I. Stansfield and M. C. Romano, PLoS Comp Bio (2013)

�: mRNA-dependent estimates of the initiation ratesProtein production rates
L. Ciandrini, I. Stansfield and M. C. Romano, PLoS Comp Bio (2013)







