
Are Domesticated Roots Replacements or Fall-Back Foods
in Hunter-Gatherer Diets?
Karen L. Kramer and Russell D. Greaves
December 2010
Corresponding Author: Karen L. Kramer
Affiliation: Associate Professor
Department of Human Evolutionary Biology
Harvard University
Cambridge, MA 02138
Coauthor: Russell D. Greaves
Affiliation: Research Associate
Peabody Museum of Archaeology and Ethnology
Harvard University
Cambridge, MA 02138
Please direct correspondence to:
(617) 495-1870
Manuscript submitted to Proceedings of the Royal Society December 13, 2010
Please do not cite without authors’ permission
Manuscript information:
Pages: 19
Tables: 4
Figures: 1
Supplementary material
This research was funded by the National Science Foundation (0349963 and DBS-
9123875), L.S.B. Leakey Foundation, and the Milton Fund.

Are Domesticated Roots Replacements or Fall-Back Foods
in Hunter-Gatherer Diets?
Abstract
Roots are key wild food resources for both modern and past hunter-gatherers living in
savanna, steppe and forested environments. Many foragers who rely on these resources,
also cultivate small quantities of domesticated plant foods. Traditional approaches to the
transition from hunting and gathering to agriculture postulate that wild foods drop out of
the diet because of their poor quality or because they require more labor investment
compared to cultigens. However, little is known about the comparative dietary quality or
labor demands of wild and cultivated roots. We use data from Pumé foragers of
Venezuela who combine small amounts of seasonal manioc cultivation within a
predominant reliance on hunting, wild roots, fishing and fruit collection. Results show
that wild roots have a higher nutritional content and require less labor than manioc.
Despite its lower quality and returns, the opportunity cost of adding manioc into the diet
is low and introduces added diversity to the diet during a particularly stressful time of the
year. Some plant cultivation for fall-back foods is both a potentially stable and rational
forager strategy for maintaining a broad subsistence base centered on wild animal and
plant food procurement.
Keywords: hunter-gathers (foragers); diet; nutritional composition; fall-back foods; Pumé
1. INTRODUCTION Hunting and gathering has been a successful human strategy because it incorporates a
broad spectrum of subsistence options. Hunter-gatherers maximize food security through
diverse food targets and an array of behaviors (mobility, technology, re-assortment of
resident populations). Categorical distinctions are often drawn between purely hunting
and gathering economies and those involving domesticated crops or animals. The
underlying assumption is that any plant food cultivation or animal husbandry signifies a
directional shift toward greater reliance on food production. We raise the question
whether this model of linear cultural evolution overlooks an important dimension of
hunter-gatherer strategies. Instead of replacing wild foods, the incorporation of some
horticulture within a foraging economy can be a long-term and stable strategy that
maintains foraging options.
While modern hunter-gatherers primarily live in marginal or underpopulated areas, both
archaeological data and ethnographic examples indicate that foraging economies not
uncommonly occurred in close proximity and interaction with food production.
Archaeologically, transitions from the first emergence of cultigens visible in the record to
systems nearly completely reliant on domesticates occur across several thousands of
years (Piperno & Pearsall 1998; Smith 2001). Prior to widespread intensive farming,
many populations incorporated variable amounts of cultivated foods combined with
primary, or significant, reliance on wild plant gathering and foraging for protein
resources (Harris 1989; Wills 1995; Winterhalder & Kennett 2006). Ethnographic data
also indicate that many modern forager societies periodically use or have historically

used cultivars or domesticated animals (Table 1). Although these patterns are
appreciated, many current views still emphasize a trajectory of replacement of foraging
following the introduction of cultigens (Bettinger et al. 2010; Zeder & Smith 2009).
Table 1 about here
The observed persistence of hunting and gathering despite long periods of association
with small amounts of cultivated food challenges the view that domesticates will
inevitably replace wild foods. To model this problem we propose that to incorporate
cultigens, foragers trade off the food value of new resources against the opportunity cost
of the time required for new activities (planting, weeding, tending animals) that could
otherwise be spent foraging. Given this trade-off, we expect that if cultivated foods are
more nutritious or offer significant labor savings in search or handling costs, they would
replace wild foods. Alternatively, if cultivated foods do not offer these advantages, then
hunters and gatherers may either ignore them, or incorporate them into the diet as
complementary or fall-back foods.
We compare wild and domesticated roots because they share many similar
characteristics, offering controls in modeling the trade-offs involved in adopting a
cultivated resource. Roots are important plant resources for many hunter-gatherers and
also feature in key evolutionary debates on dietary shifts, expansion into novel
environments, food sharing practices, and the division of labor (Marean 1997; O’Connell
et al. 1999; Laden & Wrangham 2005). Underground plant storage organs (including
tubers, corms, roots, and bulbs, collectively referred to as roots throughout), have several
key attributes that make them particularly attractive hunter-gatherer foods. They can be
harvested across a greater proportion of the year than many fruits and seeds, especially in
more seasonally extreme environments. Data from Africa (Tucker & Young 2005;
Yasuoka 2006) Australia (O’Connell & Hawkes 1981), Asia (Eder 1978; Endicott &
Bellwood 1991), and the Americas (Hurtado & Hill 1987; Kelly 1995: table 3-3) indicate
return rates for roots average about 1000-3000 kcal/hr. While return rates can be higher
for resources such as honey or meat, roots are more frequently included as daily dietary
items because of their predictability. Roots provide reliable returns because patch
locations are known and condition is usually monitored prior to foraging trips.
Consequently, search time is negligible and daily variance in return rates is much lower
than for hunted or fished resources (Greaves 1997b; Tucker 2006).
Roots are generally high in carbohydrates and starch, important to human diets
particularly in environments poor in terrestrial fauna, or when downturns occur in the
availability of protein and fat resources. Among the hunter-gathers for whom subsistence
data are available, roots are the second most important class of food beside meat and may
represent as much as 30-37% of the diet (Hadza and G/wi, respectively; Lancaster
2000:58). However, few nutritional composition data exist, and only for various African
and Australia roots habitually used by foragers (Cane 1987; Miller et al. 1993;
Schoeninger et al. 2001; Yasuoka 2006).

If cultivated foods are incorporated into hunter-gatherer diets, then certain characteristics
would make some crops more likely adoptions. Manioc is a cultivated tuber, originally
domesticated in northern South America. Manioc is composed almost exclusively of
carbohydrates, with minimal amounts of other nutrients. It has a long seasonal
availability, is less vulnerable to pest loss than many seed crops, is available in large
package sizes, can be stored underground with no additional investment in structural
facilities, and has similar harvesting requirements to wild roots (Wilson & Dufour 2006).
Bitter manioc is more widely cultivated than sweet manioc because it grows well in poor
tropical soils. However, the toxins (cyanogenic glycosides) in bitter manioc must be
processed through multiple steps to reduce the cyanide content and prepare it for
consumption. In addition to the Pumé we report on here, several mobile South American
foragers historically include some manioc cultivation within their primary reliance on
wild foods, including the Nukak, Hoti and Hiwi (see Table 1).
Pumé foragers of the Venezuelan neotropical savannas combine small amounts of
seasonal manioc cultivation with their primary nutritional reliance on wild resources. To
examine the utility of the model that foragers trade off food value and opportunity costs,
we analyze the nutritional composition, labor investment, and food returns of wild and
cultivated roots used by the Pumé to address several questions. As traditional approaches
suggest, does the use of cultigens indicate the replacement of lower quality or more
labor-intensive wild foods with more economical domesticated foods? Can foragers
incorporate some cultivated plants into their diet in ways that support continued primacy
of hunting and gathering? How can the potential combination of hunting, gathering, and
some incorporation of non-foraged food sources inform us about more dynamic ways to
view human dietary decisions?
2. MATERIAL & METHODS
(a) Study population The Pumé inhabit the low savannas (llanos) of west central Venezuela within the Orinoco
River drainage. Part of the Pumé population are horticulturalists and a smaller number
(~1000 Pumé in ~20 communities) subsist as mobile foragers in the savannas between
these drainages (Mitrani 1988). Referred to as the savanna Pumé, they depend on
hunting, fishing, wild root collection, gathering of mangos from feral groves, and
seasonal manioc cultivation (Gragson 1989; Greaves 1997a, 1997b). The savanna Pumé
move camps five to six times a year in response to hyperseasonal fluctuations in rainfall.
Roots represent the single most important wild plant food, especially in the wet season.
Return rates were determined using focal follow techniques over an 18-month period
(Table 2; Greaves 1997b). During root trips, women target predetermined root patches
and spend almost all foraging trip time engaged in digging and harvesting roots. Most
roots are relatively small (mean=~20 g), but are located in dense patches, and are only a
few centimeters below the ground surface. Unlike foods targeted by men, patch locations
are known and root collection requires no search time. Monitoring the condition of wild
roots is embedded as part of normal travel by women during other mobile foraging
activities. Also unlike male-acquired foods, return rates are directly proportional to the
time and labor invested in root collection (Greaves 1997b). Wild roots are collected in

large quantities and constitute approximately 25% of the total annual Pumé diet,
comparable to that of other savanna foragers (Hurtado & Hill 1987; Marlowe &
Berbesque 2009).
[Table 2 about here]
Bitter manioc is the only cultivated plant successfully grown by the savanna Pumé.
Manioc is only consumed during part of the wet season, when it may constitute up to 35-
40% of the seasonal diet, but only ~10% of the annual diet. Garden locations and size are
limited by the relatively small patches of forest not flooded during the wet season (0.10-
0.12 ha/person [Gragson 1989]). Preparation of new swiddens occurs in brief bouts
during the dry season and weeding and harvesting during the wet season, considerably
shorter than their wild food procurement commitments (Figure 1). While the savanna
Pumé infrequently attempt to plant other crops, soils of the seasonally inundated
Venezuelan low savannas are extremely poor for agriculture (Elizalde et al. 2007).
[Figure 1 about here]
(b)Nutrient composition analyses Four species of commonly eaten roots and one sample of cultivated manioc were
collected and analyzed to obtain macronutritional data on Pumé roots. All samples were
collected by Pumé women during their normal foraging activities. Cooked samples were
prepared by various Pumé households as part of regular foraging, cooking, and
consumption activities. This ensures that the foods analyzed are generated from normal
dietary selection and processing. Nutrient analyses were performed in the Nutritional
Ecology Laboratory at Harvard University using standard laboratory procedures (see
ESM for description of lab methods; Conklin-Brittain et al. 2006).
3. RESULTS Table 3 (Table S1) shows the results from multiple samples of three tubers regularly
collected by the Pumé compared to the nutritional value of their manioc. D. margaretae
is collected in the wet season in the second highest quantities of any wild plant food, and
M. cannifolia is the primary dry season tuber. The total kcal available from manioc, D.
margaretae and M. cannifolia are comparable, but wild roots all are higher in protein, fat,
and mineral content.
[Table 3 about here]
Comparatively, the most important Venezuelan roots have a higher caloric value, 324-
349 kcal/100 g, than those used by other foragers. For the Hadza, wild tubers average
roughly 300 kcal/100 g (Schoeninger et al. 2001). Aka wild yams provide 120 kcal/100 g
(Yasuoka 2006: table IV) and various Australian roots produce between 135-336
kcal/100 g (Miller et al. 1993; Cane 1987: table VI). Pumé roots have relatively low
mineral content, are low in lipids, and exhibit low-medium protein content. However,
these analyses indicate that roots crucial to the Pumé diet have lower fiber content and

higher nutritional value than those reported for the Hadza, Aka, and Australian
Aboriginals.
Decisions to incorporate cultivated resources in a hunting and gathering diet include not
only nutritional quality, but also the labor costs of cultigens and their complementarity
with the time allocation and mobility demands of foraging. There is no direct means to
compare return rates for wild and cultivated roots because unlike root collection, much of
gardening work involves time expenditures without correlated returns. Additionally,
calculating an accurate individual return rate is not feasible because many individuals
contribute field labor to other families’ plots, and harvesting and processing are often
performed by people without gardens. However, relative time investments in acquisition
and processing are comparable (Table 4). There is considerable asymmetry between time
invested in garden preparation and the lack of any pre-harvest investment in wild root
patches because of labor required to clear and burn forest, prepare, plant, and weed
gardens. Search, travel and harvest investments in wild and cultivated roots are
equivalent. Both wild root patches and gardens are known locations involving no search
effort, and search times within garden or wild root patches do not vary. Access is also
similar. Both wild and cultivated roots are located in the same kinds of soil and are
harvested with similar tools.
[Table 4 about here]
Post-harvest processing, however, involves very different time commitments. While
some wild plant foods that are infrequently collected in small amounts by the Pumé
require leaching or other processing to remove toxins, the most common wild roots
require only boiling to be edible. In contrast, post-harvest manioc processing involves a
considerable time commitment. To remove the cyanide, bitter manioc is grated, the liquid
expressed, then sifted to separate and expose it for additional oxidation, and finally
cooked. For an average savanna Pumé return of manioc (8.3 kg, n=24) this process
minimally requires two hours. Although manioc tubers are larger than wild roots, return
rates are expected to be lower for manioc since cultivation requires additional garden
work and detoxification. Evaluated on these characteristics, manioc appears to provide no
clear nutritional or labor advantage.
4. DISCUSSUION These results raise several points pertinent to hunter-gatherer research. First, why would
hunter-gatherers include cultigens in their subsistence base rather than intensify wild root
collection? Second, is a mix of some cultivation with wild root collection a stable
strategy that supports a continued hunter-gatherer lifeway? Finally, what predictions can
be made from our results?
(a) Opportunity costs If wild roots are nutritionally more valuable than manioc, why invest any time and energy
in swidden production, which involves tasks not part of normal wild food foraging
activities? We suggest that despite additional labor requirements, manioc cultivation does
not conflict with foraging activities and the opportunity cost of allocating time to

cultivation is relatively low. Scheduling of garden labor is flexible, distributed throughout
the year, and timed around other temporally constrained foraging tasks. Garden
preparation, calorically expensive work, occurs during the dry season when food is
relatively more abundant and disease less prevalent, minimizing competing energy
demands. Because manioc can be stored in the ground until needed with no loss in yield,
unlike seed and fruit crops, harvest labor bottlenecks and conflicts with other activities
are avoided. Compared with other crops, vegetative propagation requires less time in
garden preparation and reduces new planting vulnerability to rainfall and pests,
decreasing overall labor cultivation costs. Manioc returns are predictably high, search
costs are low, and crop patches are accessed using the same tools and technology as wild
roots. These combined qualities make manioc an attractive complement to wild
resources.
While investing in cultivation supplies additional calories at a relatively low cost during
the wet season, the savanna Pumé could alternatively increase their foraging range for
wild roots. Because manioc is utilized during the same season of highest inputs from wild
roots, this suggests increasing wild root productivity is not as economical an option as
combined foraging and minimal cultivation. Intensifying root collection requires
additional mobility and energetic costs during the season of greatest nutritional and
epidemiological stress. During the wet season, women annually lose approximately 8%
of their body weight (Kramer & Greaves 2007, 2010). Rather than expanding foraging
ranges, the savanna Pumé promote plant productivity of nearby locations where roots are
not otherwise available and importantly increase dietary diversity during the lean season.
(b) Foraging and cultivation as a stable hunter-gatherer strategy Hunter-gatherers maintain broad dietary options through an array of alternative and fall-
back strategies (Kelly 1995). The incorporation of horticulture within a foraging
economy may be part of this broad set of options. The savanna Pumé are not under land
or other constraints commonly linked with economic shifts toward food intensification.
Manioc is incorporated into the diet situationally in response to the changing availability
of wild foods, or is left unharvested as a source of future calories, and is best
characterized as a seasonal fall-back food. During any particular sample year, a large
proportion of households (~45%) have no gardens under cultivation. Many of these are
young families, but older families with dependents also exhibit fluctuating annual
commitments to any horticultural effort.
Dependence on wild roots in combination with manioc cultivation appears to be a stable
subsistence strategy among the savanna Pumé. Historic accounts indicate the savanna
Pumé practice a long-established mixture of minimal cultivation with primary hunting
and gathering and this is not a novel or recent introduction (Mitrani 1988). Manioc
provides a predictable alternative that can be used in relation to changes in the
availability of wild plants.
Humans and many other animals incorporate fall-back subsistence behaviors directed
toward foods that are not their primary dietary targets (Lambert 2009). Hunter-gatherer
studies generally downplay food production components of foraging societies, or exclude

those groups as not representative of hunter-gatherers. Rather than a classificatory
approach, we emphasize that some cultivation is a backup strategy that enables hunter-
gathers to retain reliance on wild foods, and that this approach better accounts for the
empirical archaeological and ethnographic data on hunter-gatherers. Viewing hunter-
gatherer use of cultivated foods as a potential fall-back strategy is consistent with the
observations that diverse groups of foragers include some horticulture, minor animal
husbandry, periodic wage labor, or trade and exchange as complementary activities to
support a primary dependence on wild foods.
(c) Predictions Given the results from our model expectations, we would predict in cross-cultural
analysis of hunter-gatherers that: 1) If labor and scheduling investments in wild and
cultivated resources do not conflict (low opportunity cost) then mixed hunter-gatherer
strategies will be stable and long term. 2) Under these circumstances we would predict
that investment in cultivated resources, animal domestication or wage labor will be
annually or seasonally variable depending on wild resource harvests. 3) Under other
circumstances, some wild foods may be replaced if gardening, labor exchange, or
procurement of select resources for trade (i.e., bushmeat, fish, forest products) can be
done at a lower cost than continued total dietary reliance on hunting and gathering.
The Pumé and several groups in Table 1 exemplify the first two predictions, where there
is no loss in diversity of wild foods while including some cultivation. Hunters and gathers
also may combine a diet dominated by wild foods if particular crops present investment
conflicts. For example, the Mikea (Tucker 2006) partially commit to maize cultivation
while retaining wild roots in their diet. Future-discounting influences the decision to
maintain investment in wild root collection. While time spent in additional garden labor
has the delayed payoff of potentially higher future returns, time spent in wild root
collection has the immediate payoff of meeting daily food requirements. Foragers who
replace more costly wild foods but continue to use others are an appropriate test of the
third prediction. For example, several southeast Asian foragers who historically depended
on wild roots have reduced their dietary reliance on particular varieties primarily in
response to the availability of rice. Agta (Headland & Bailey 1991; Peterson 1978) and
Batak (Eder 1978; Endicott & Bellwood 1991) foragers live in forests with unusually
high diversities of wild yams. Both groups continue to gather certain wild yams, but have
added some cultivation and obtain rice through exchange of forest foods and products.
The wild root species that have dropped out of their diets have high processing costs or
particularly high toxin content.
In sum, the Pumé results point to two main reasons why horticultural practices are
compatible with stable hunting and gathering economies. First, although wild foods are
often characterized as being of lower value compared to cultigens, our analyses show that
wild roots are nutritionally of higher quality than manioc, require similar search and
travel times, and save time in field preparation, maintenance and post harvesting
processing costs. Manioc’s nutritional poverty makes it an unlikely candidate to replace
reliance on plant foods. However, manioc is a predictable backup up food that smoothes
potential fluctuations in the availability of wild foods during a particularly food-limited

season. Second, wild roots persist in the diet and have their greatest returns during the
horticultural season. Wild plant foods are commonly thought to remain in the diet
primarily to stabilize fluctuations in the availability of cultivated foods. In contrast, we
argue that manioc adds dietary diversity and a resource that does not conflict with the
scheduling and labor demands of wild-food gathering. The seasonal incorporation of
manioc by savanna Pumé does not signal a directional shift away from hunting and
gathering toward greater reliance on food production. Instead, low-level incorporation of
fall-back horticulture extends the viability of a primarily hunting and gathering economy.
From a perspective that hunting and gathering involves diversified subsistence and
strategic use of fall-back foods, there is no a priori reason why hunter-gatherers would
exclude minimal incorporation of food production or labor exchange for food as outside
of their expected lifeways.
Acknowledgements We thank the Pumé of Doro Aná, who have patiently and enthusiastically supported our
research and endeavored to teach us about the economic complexities of foraging. We are
grateful to Nancy Conklin-Britain, director of the Nutritional Ecology Laboratory at
Harvard University, Christi Butler and Paula Lu for performing the nutritional analyses.
We wish to thank the National Science Foundation (DBS-9123875 and 0349963), the
L.S.B. Leakey Foundation, and the Milton Fund for supporting the Pumé research.
This research was conducted under human subjects approval by the Harvard and the
University of New Mexico. Fieldwork was permitted in Venezuela by the Ministerio de
Educación, Dirección de Asuntos Indígenas (permit no. 02/92) and the Consejo Nacional
de Investigaciones Científicos y Tecnológicos (no. 1622). Field samples were collected
under permits from the Ministerio del Ambiente y de los Recursos Naturales Renovables
(MARNR), Instituto Nacional de Parques, (nos. 92-492 and 92-2946) and from the
MARNR, Servicio Autónomo Forestal Venezolano, (nos. 148 and 1).

Figure 1. Relative proportion of the time that Pumé males and females contribute to wild
resource and food production during the dry and wet seasons (Gragson 1989: table 6.14;
1990: figs 4-5).
M
M
M
M
F
F
F
F
0%
20%
40%
60%
80%
100%
Wild resources Manioc Wild resources Manioc
Dry season Wet season

Table 1. Modern hunter-gatherer societies who periodically use or have historically
combined wild resources and some use of cultivars or domesticated animals.
Africa
Mbuti, Efe, Aka Bailey & Aunger 1989; Hart & Hart 1986
Kalahari foragers Hitchcock & Ebert 1984; Lee 1979; Wilmsen 1989
Mikea Tucker 2006
Asia
Punan Hoffman 1986
Agta Headland & Bailey 1991; Peterson 1978
Mlabri Oota et al. 2005; Pookajorn 1992
South America
Sirionó Stearman 1989
Nukak Politis 2007
Hoti Coppens 1983; Zent & Zent 2007
Hiwi Hurtado & Hill 1990; Metzger & Morey 1983
Pumé Gragson1989; Greaves 1997a, 1997b

Table 2. Descriptive characteristics (mean and SD) of Pumé wild root returns for adult
femalesa.
Small Roots*
(n=45 person trips)
Large Roots**
(n=101 person trips)
Average age 36.6 (±11.0) 44.4 (±11.4)
Distance 1.6 km (±.6) 5.6 km (± 2.6)
Hours 4.19 hrs (±2.24) 4.84 hrs (±1.46)
Weight 6.4 k (± 4.7) 13.8 k (± 5.9)
Return rate 1.66 k/hr (±1.06) 2.95 k/hr (±1.14)
Kcal/trip 3340 kcal 10,488 kcal
Distances, times and return rates include round trips from camps.
* <5 cm; primarily Myrossma cannifolia and Dioscoria sp.
** >5 cm; primarily Dracontium margaretae a Women 20 and older. Men seldom forage for roots.
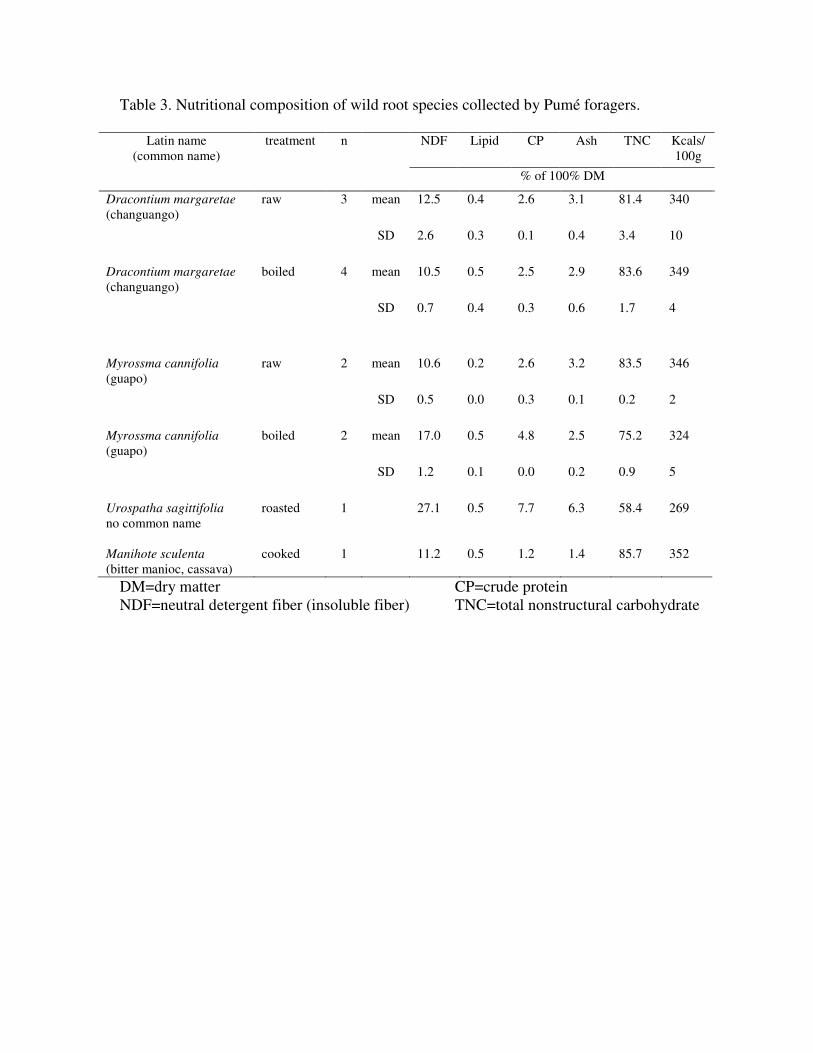
Table 3. Nutritional composition of wild root species collected by Pumé foragers.
Latin name
(common name)
treatment n NDF Lipid CP
Ash
TNC
Kcals/
100g
% of 100% DM
Dracontium margaretae
(changuango)
raw 3 mean 12.5 0.4 2.6 3.1 81.4 340
SD 2.6 0.3 0.1 0.4 3.4 10
Dracontium margaretae
(changuango)
boiled 4 mean 10.5 0.5 2.5 2.9 83.6 349
SD 0.7 0.4 0.3 0.6 1.7 4
Myrossma cannifolia
(guapo)
raw 2 mean 10.6 0.2 2.6 3.2 83.5 346
SD 0.5 0.0 0.3 0.1 0.2 2
Myrossma cannifolia
(guapo)
boiled 2 mean 17.0 0.5 4.8 2.5 75.2 324
SD 1.2 0.1 0.0 0.2 0.9 5
Urospatha sagittifolia
no common name
roasted 1 27.1 0.5 7.7 6.3 58.4 269
Manihote sculenta
(bitter manioc, cassava)
cooked 1 11.2 0.5 1.2 1.4 85.7 352
DM=dry matter CP=crude protein
NDF=neutral detergent fiber (insoluble fiber) TNC=total nonstructural carbohydrate
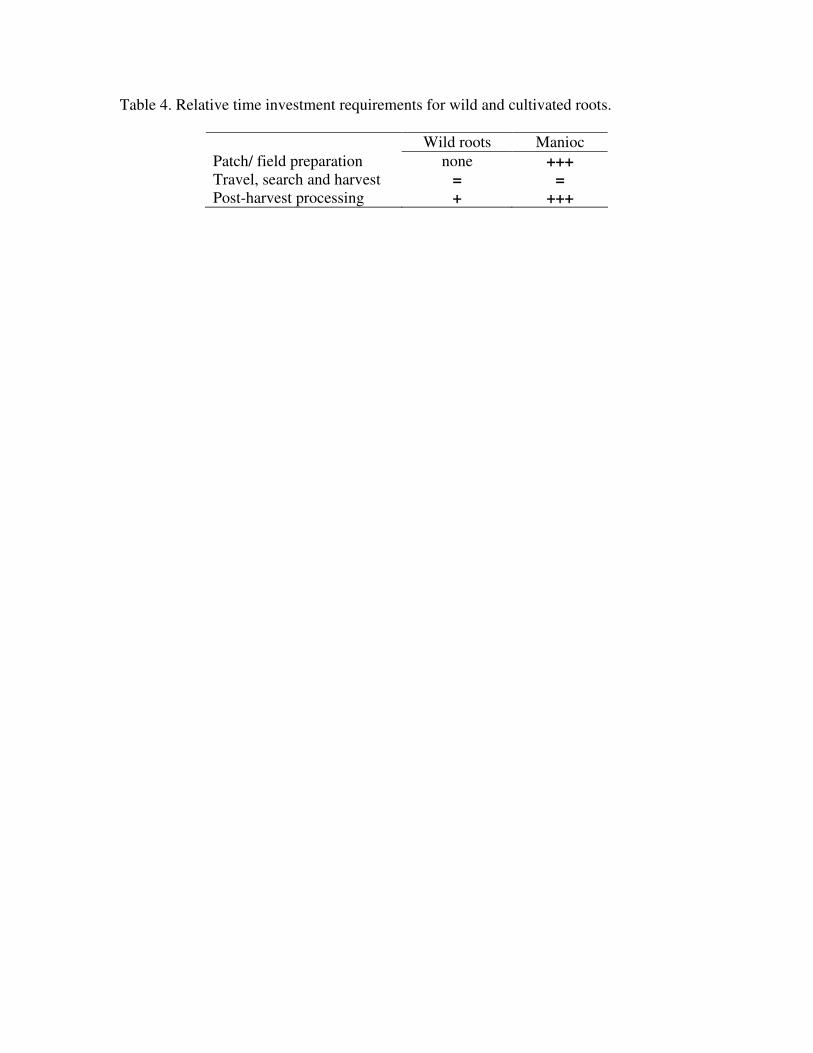
Table 4. Relative time investment requirements for wild and cultivated roots.
Wild roots Manioc
Patch/ field preparation none +++ Travel, search and harvest = = Post-harvest processing + +++

References
Bailey, R. C. & Aunger, R., Jr. 1989 Net hunters vs. archers: variation in women’s
subsistence strategies in the Ituri forest. Human Ecology 17, 273-297.
Bettinger, R. L., Barton, L. & Morgan, C. 2010 The origins of food production in north
China: a different kind of agricultural revolution. Evolutionary Anthropology 19, 9-21.
Cane, S. 1987 Australian aboriginal subsistence in the Western Desert. Human Ecology
15, 391-434.
Conklin-Brittain, N. L., Knott C. D. & Wrangham, R. W. 2006. Energy intake by wild
chimpanzees and orangutans: methodological considerations and a preliminary
comparison. In Feeding ecology in apes and other primates: ecological, physical and
behavioural aspects (eds. G. Hohmann, M.M. Robbins & C. Boesch), pp. 445-465.
Cambridge, UK: Cambridge University Press.
Coppens, W. 1983 Los Hoti, In Los aborígenes de Venezuela, Vol. II (ed. W. Coppens),
pp. 243-301. Caracas, Venezuela: Fundación La Salle de Ciencias Naturales.
Eder, J. F. 1978 The caloric returns to food collecting: disruption and change among the
Batak of the Philippine tropical forest. Human Ecology 6, 55-69.
Elizalde, G., Viloria, J. & Rosales, A. 2007 Geografía de suelos de Venezuela.
GeoVenezuela, 2, pp. 402-537. Caracas, Venezuela: Fundación Empresas Polar.
Endicott, K. & Bellwood, P. 1991 The possibility of independent foraging in the rain
forest of Peninsular Malaysia. Human Ecology 19, 151-185.
Gragson T. L. 1989 Allocation of Time to Subsistence and Settlement in a Ciri Khonome
Pumé Village of the Llanos of Apure, Venezuela. Ph.D. dissertation, Pitsburgh, PA: The
Pennsylvania State University.
Gragson T. L. 1990 The allocation of time to subsistence in lowland South America:
analogies for the transition from forging to agriculture. Paper presented at the sixth
international conference on hunting and gathering, May 28-June 11, 1990. Fairbanks,
AK: Univ of Alaska.
Greaves, R. D. 1997a Hunting and multifunctional use of bows and arrows:
ethnoarchaeology of technological organization among Pumé hunters of Venezuela. In
Projectile Technology (ed. H. Knecht), pp. 287-320. New York, NY: Plenum Press.
Greaves, R. D. 1997b Ethnoarchaeological investigation of subsistence mobility, resource
targeting, and technological organization among Pumé foragers of Venezuela. Ph.D. dissertation,
Albuquerque, NM: University of New Mexico.

Harris, D. R. 1989 An evolutionary continuum of people-plant interactions. In Foraging
and farming: the evolution of plant exploitation (eds. D. R. Harris & G.C. Hillman), pp.
11-26. London, UK: Unwyn Hyman.
Hart, T. B. & Hart, J. A. 1986 The ecological basis of hunter-gatherer subsistence in
African rain forests: the Mbuti of eastern Zaire. Human Ecology 14, 29-55.
Headland, T. N. & Bailey R. C. 1991 Have hunter-gatherers ever lived in tropical rain
forests independently of agriculture? Human Ecology 19, 115-122.
Hitchcock, R. K. & Ebert, J. I. 1984 Foraging and food production among Kalahari
hunter/gatherers. In From hunters to farmers: the causes and consequences of food
production in Africa (eds. J. D. Clark and S. A. Brandt), pp. 328-348. Berkeley, CA:
University of California Press.
Hoffman, C. 1986 The Punan: hunters and gatherers of Borneo. Ann Arbor, MI: UMI
Research Press.
Hurtado, A, M. & Hill, K. 1987 Early dry season subsistence ecology of Cuiva (Hiwi)
foragers of Venezuela. Human Ecology 15, 163-187.
Hurtado, A, M. & Hill, K. 1990 Seasonality in a foraging society: variation in diet. Work
effort, fertility, and sexual division of labor among the Hiwi of Venezuela. Journal of
Anthropological Research 46, 293-346.
Kelly, R. L. 1995 The foraging spectrum: diversity in hunter-gatherer lifeways.
Washington, DC: Smithsonian Institution Press.
Kramer, K. L. & Greaves, R, D. 2010 Synchrony between growth and reproductive
patterns in human females: early investment in growth among Pumé foragers. American
Journal of Physical Anthropology. 141, 235-244.
Kramer, K. L. & Greaves, R, D. 2007 Changing patterns of infant mortality and fertility
among Pumé foragers and horticulturalists. American Anthropologist 109, 713-726.
Laden G. & Wrangham, R. W. 2005 The rise of hominids as an adaptive shift in fallback
foods: plant underground storage organs (USOs) and australopith origins. Journal of
Human Evolution 49, 482-498.
Lancaster, J. B., Kaplan, H. S., Hill, K. & Hurtado, A.M. 2000 The evolution of life
history, intellgence and diet among chimpanzees and human foragers. In Perspectives in
ethology, Vol 13: evolution culture and behavior (eds. F. Toneau & N. S. Thompson), pp.
47-72. New York, NY: Kluwer Academic/Plenum Publishers.

Lambert, J. E. 2009 Summary to the symposium issue: primate fallback strategies as
adaptive phenotypic plasticity-scale, pattern, and process. American Journal of Physical
Anthropology 140, 759-766.
Lee, R. B 1979 The !Kung San: men, women and work in a foraging society. Cambridge,
UK: Cambridge University Press.
Marean, C. W. 1997 Hunter-gatherer forging strategies in tropical grasslands: model
building and testing in the east African Middle and Later Stone Age. Journal of
Anthropological Archaeology 16, 189-225.
Marlowe, F. W. & Berbesque, J. C. 2009 Tubers as fallback foods and their impact on
Hadza hunter-gatherers. American Journal of Physical Anthropology 140 (4):751-758.
Metzger D. J. & Morey, R. V. 1983 Los Hiwi (Guahibo). In Los aborígenes de
Venezuela, Vol. II (eds. R. Lizarralde & H. Seijas), pp. 125-216. Caracas, Venezuela:
Fundación La Salle de Ciencias Naturales.
Miller, J. B., James, K. W. &. Maggiore, P. M. A. 1993 Tables of composition of
Australian aboriginal foods. Canberra, Australia: Aboriginal Studies Press.
Mitrani, P. 1988 Los Pumé (Yaruro). In Los aborígenes de Venezuela, Vol. III (ed. J.
Lizot), pp. 147-213. Caracas, Venezuela: Fundación La Salle de Ciencias Naturales.
O’Connell, J. F. & Hawkes, K. 1981 Alyawara plant use and optimal foraging theory. In
Hunter-gatherer foraging strategies: ethnographic and archaeological analyses (eds. B.
Winterhalder & E. A. Smith, pp. 99-125. Chicago, IL: University of Chicago Press.
O’Connell, J. F., Hawkes, K. F. & Blurton Jones, N. G. 1999 Grandmothering and the
evolution of Homo erectus. Journal of Human Evolution 36, 461-485.
Oota, H., Pakendorf, B., Weiss, G., von Haeseler, A., Pookajorn, S., Settheetham-Ishida,
W., Tiwawech, D., Takafumi, I. & Stoneking, M. 2005 Recent origin and cultural
reversion of a hunter-gatherer group. PLOS Biology 3, 536-542.
Piperno, D. R. & Pearsall, D. M. 1998 The origins of agriculture in the lowland
neotropics. San Diego, CA: Academic Press.
Peterson, J. T. 1978 The ecology of social boundaries: Agta foragers of the Philippines.
Urbana, IL: University of Illinois Press.
Politis, G. G. 2007 Nukak: Ethnoarchaeology of an Amazonian People. Walnut Creek,
CA: Left Coast Press.
Pookajorn, S. 1992 The Phi Tong Luang (Mlabri): A hunter-gatherer group in Thailand.
Bangkok, Thailand: Odeon Store Printing House.

Schoeninger, M. J., Bunn, H. T., Murray, S. S. & Marlett, J. A. 2001 Composition of
tubers used by the Hadza foragers of Tanzania. Journal of Food Composition and
Analysis 14, 15-25.
Smith, B. D. 2001 Low-level food production. Journal of Archaeological Science 9, 1-43.
Stearman, A. M. 1989 Yuquí: forest nomads in a changing world. New York, NY: Holt
Reinhart & Winston.
Tucker, B. 2006 A future discounting explanation for the persistence of a mixed
foraging-horticulture strategy among the Mikea of Madagascar. In Behavioral ecology
and the transition to agriculture (eds. D. J. Kennett & B. Winterhalder), pp. 22-40.
Berkeley, CA: University of California Press.
Tucker, B. & Young, A. G. 2005 Growing up Mikea: children’s time allocation and tuber
foraging in southwestern Madagascar. In Hunter-gatherer childhoods: evolutionary
developments and cultural perspectives (ed. B. S. Hewitt), pp. 147-171. Brunswick, NJ:
Aldine Transaction, New.
Wills, W. H. 1995 Archaic foraging and the beginning of food production in the
American Southwest. In Last hunters-first famers: new perspectives in the prehistoric
transition to agriculture (eds. T. D. Price & A. B. Gebauer), pp. 215-242. Santa Fe, NM:
School of American Research Press.
Wilmsen, E. N. 1989 Land filled with flies: a political economy of the Kalahari. Chicago,
IL: University of Chicago Press.
Wilson, W. M. & Dufour, D. F. 2006 Ethnobotanical evidence for cultivar selection
among Tukanoans: manioc (Manihot esculenta Crantz) in the northwest Amazon. Culture
& Agriculture 28, 122-130.
Winterhalder, B. & Kennett, D. J. 2006 Behavioral ecology and the transition from
hunting and gathering to agriculture. In Behavioral ecology and the transition to
agriculture (eds. D. J. Kennett & B. Winterhalder), pp. 1-21. Berkeley, CA: University of
California Press.
Yasuoka, H.
2006 Long term foraging expeditions (molongo) among Baka hunter-gatherers in the
northwestern Congo Basin, with special reference to the “wild yam question”. Human
Ecology 34, 275-296.
Zeder, M. A. & Smith, B. D. 2009 A conversation on agricultural origins: talking past
each other in a crowded room. Current Anthropology 50, 681-691.

Zent, E. L. & Zent, S. 2007 Los Jodï (Hotï). In Salud indígena en Venezuela, Vol 1 (eds.
G. Freire & A. Tillet), pp. 77-130. Caracas, Venezuela: Ministerio del Poder Popular para
la Salud, Editorial Arte.

Electronic Supplementary Material
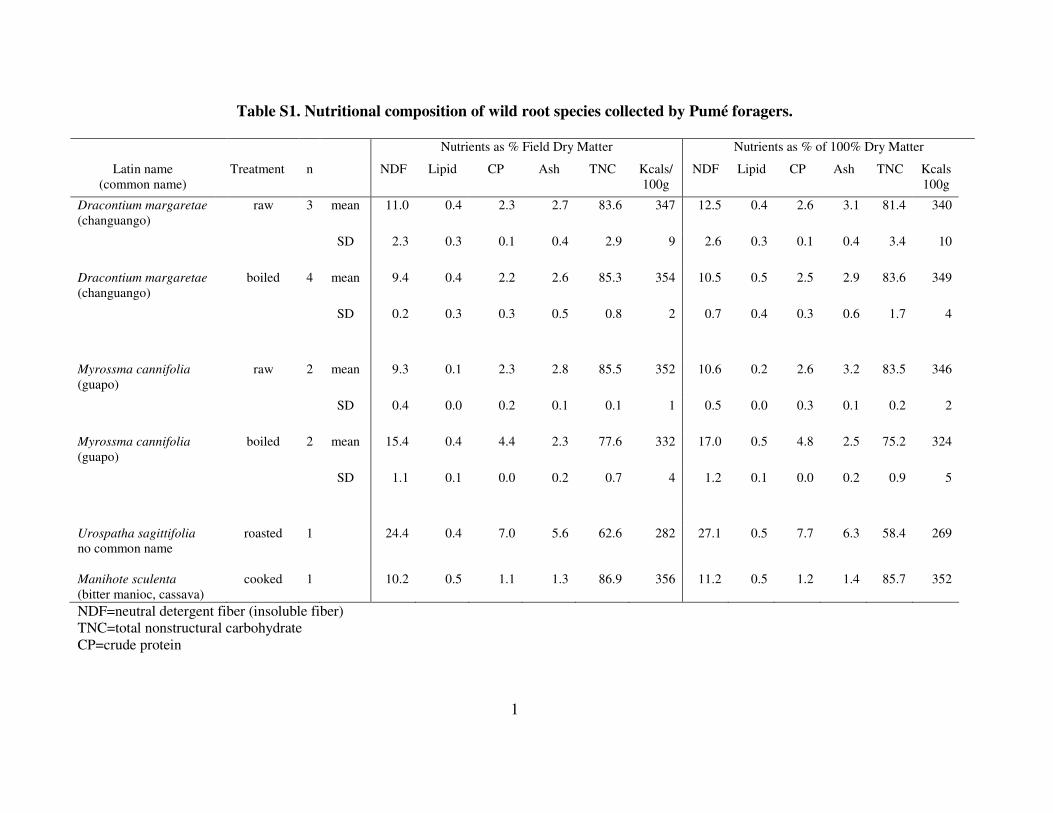
Field sampling and processing Eleven samples of two wild tubers commonly collected by the Pumé
(Dracontiummargaretae and Myrosmacannifolia) and one less frequently gathered
taproot (Urospathasagittifolia) were submitted for analysis (Table S1). Both raw and
cooked samples were processed. One sample of manioc (Manihotesculenta) processed
into cassava also was analyzed. All of these samples represent foods that Pumé women
collected during normal foraging trips. Cooked samples came from various Pumé
households’ preparation of these foods for their own consumption and sharing with other
families, that also were given to Greaves as meals. Each sample was ground through a
hand-cranked meat grinder prior to weighing (a minimum of 40 g was collected for each
sample), transferred to Nalgene sample bottles, mixed with 70% isopropyl alcohol, and
covered with Parafilm prior to securing the lids. Field weights were taken with an O-
Haus model 1010-10 precision balance.
Laboratory processing Nutrient analyses were performed in the Nutritional Ecology Laboratory at Harvard
University using standard laboratory procedures (Conklin-Brittain et al. 2006). Preserved
samples were oven dried at 40º C, then ground through a #20 (1mm) mesh screen using a
Wiley Mill electric grinder. Multiple samples were analyzed to provide checks on results.
Crude protein (CP) was determined from two 0.5 g subsamples, using the Kjedahl
procedure to measure nitrogen content, then multiplying by 6.25. Digestion was
performed with Na2SO4 and CuSO4 in a semi-automated Tecator digestor unit. A Tecator
distillation unit was used to collect the released nitrogen in 4% boric acid, then titrated
with 0.1N HCL.
Lipid was analyzed from two 0.5 g subsamples using a modified AOAC method.
Petroleum ether was added to each sample and separation occurred during seven days at
room temperature. Total lipid content was measured by hot weighing at 60º C.
Fiber was analyzed by a modified Detergent System of Fiber Analysis to identify total
insoluble fiber (NDF, neutral detergent fiber). Six samples were analyzed to determine
other fiber components, but these data are unavailable for all samples. Total NDF was
measured with traditional refluxing apparatus, then total NDF was measured by hot
weighing at 100º C.
Dry matter and ash were analyzed for each of two 0.3 g subsamples. Reported values
represent percent of 100% dry matter (100% DM) and was measured by drying each
subsample at 100º C for eight hours then hot weighing the remainder. The dry matter
sample was then ashed at 520º C for eight hours and hot weighed to obtain the total ash
amounts.
Total non-structural carbohydrates (TNC) were calculated as: TNC=100-%NDF-%lipid-
%CP-%ash. The total energy per sample was measured as kcal per 100g of dry matter,
calculated as: Energy =4 kcal/g*TNC+4kcal/g*CP+9kcal/g*lipid.
1
Table S1. Nutritional composition of wild root species collected by Pumé foragers.
Nutrients as % Field Dry Matter Nutrients as % of 100% Dry Matter
Latin name
(common name)
Treatment n
NDF
Lipid
CP
Ash
TNC
Kcals/
100g
NDF
Lipid
CP
Ash
TNC
Kcals
100g
Dracontium margaretae
(changuango)
raw 3 mean 11.0 0.4 2.3 2.7 83.6 347 12.5 0.4 2.6 3.1 81.4 340
SD 2.3 0.3 0.1 0.4 2.9 9 2.6 0.3 0.1 0.4 3.4 10
Dracontium margaretae
(changuango)
boiled 4 mean 9.4 0.4 2.2 2.6 85.3 354 10.5 0.5 2.5 2.9 83.6 349
SD 0.2 0.3 0.3 0.5 0.8 2 0.7 0.4 0.3 0.6 1.7 4
Myrossma cannifolia
(guapo)
raw 2 mean 9.3 0.1 2.3 2.8 85.5 352 10.6 0.2 2.6 3.2 83.5 346
SD 0.4 0.0 0.2 0.1 0.1 1 0.5 0.0 0.3 0.1 0.2 2
Myrossma cannifolia
(guapo)
boiled 2 mean 15.4 0.4 4.4 2.3 77.6 332 17.0 0.5 4.8 2.5 75.2 324
SD 1.1 0.1 0.0 0.2 0.7 4 1.2 0.1 0.0 0.2 0.9 5
Urospatha sagittifolia
no common name
roasted 1 24.4 0.4 7.0 5.6 62.6 282 27.1 0.5 7.7 6.3 58.4 269
Manihote sculenta
(bitter manioc, cassava)
cooked 1 10.2 0.5 1.1 1.3 86.9 356 11.2 0.5 1.2 1.4 85.7 352
NDF=neutral detergent fiber (insoluble fiber)
TNC=total nonstructural carbohydrate
CP=crude protein







