
CHAPTER 1
GENERAL INTRODUCTION

1.1 BACTERIAL BIODIVERSITY OF ANTARCTICA
1.1.1 Definition of Biodiversity
Biodiversity in general terms, refers to the variety and abundance of all life forms
that live on earth. The basic unit of biodiversity is the species. However, the most
accepted and comprehensive definition of biodiversity is that biodiversity "encompasses
all life forms, ecosystems and ecological process and acknowledges the hierarchy of
genetic, taxon and ecosystem levels" (The International Union for Conservation of
Nature and Natural Resources).
1.1.2 Importance of Exploration and Conservation of
Biodiversity
Biodiversity which includes all the animals, plants and microorganisms such as
viruses, bacteria, algae, fungi, yeasts, protozoans, etc., has been recognized as a critical
component of medicine, agriculture, forestry and fisheries and also plays an important
role in conserving earth's life supporting systems and in enhancing the quality of human
life. Therefore, the study of biodiversity is necessary to understand the interactions
between and among the species of a given habitat and to group them as a living unit in a
given ecological unit. Table 1.1 shows the extent of biodiversity that has been estimated.
Till now, more than one million plant and animal species (Mayr, 1998 and Wilson,
1992) have been described on the basis of visual observation of their distinguished
morphological characteristics. However, despite the fact that microorganisms comprise
the greatest number of individual organisms on earth (Oiembo, 1991 ), they have not
been described to the same extent as plants and animals as they require an appreciable
amount of laboratory work. About 1% of approximately 5 X 105 viral species (Colwell et

Table 1.1. Known and estimated numbers of biological species*1
Group Known Estimated total Percentage of species Species Known species
Archae a 205762 NK NK
Viruses3 5699L 5.0x10:> 1.14
Bacteria 28784L 3.0 X 10ti 2.8
Fungi4 69000 1.5 X 10ti 5
Algae 40,000 6.0 X 104 67
Bryophytes 17000 25000 28
Gymnosperms 750 NK NK
Angiosperms 250000 2.7x105 93
Protozoa3 40000 2.0x10:> 31
Porifera 5000 NK NK
Cnidaria 9000 NK NK
Nematodes 15000 500000 3
Crustacea 38000 NK NK
Insects 800000 6 to 10 x 106 8 to 13
Arthropods I Invertebrates 132460 NK NK
Molluscs 50000 NK NK
Echinoderms 6100 NK NK
Amphibians 4184 NK NK
Reptiles 6380 NK NK
Fishes 19000 21000 90
Birds 9198 - - 100
Mammals 4170 - -.100
*NK, Not Known
Data from: 1Bull et at. (1992); 21nternet (Till March, 2002). http://www.ncbi.nlm.nih.gov/Taxonomy/txstat.cgi 3Colwell eta/. (1994) 4Rossmann (1993)
2

a/., 1994), 5 to 10% of approximately 1.5 X 106 species of fungi (Rossman, 1993), 31%
of the approximately 2 X 105 species of protozoans (Systematic agenda 2000, 1994) and
3% of the approximately 109 bacterial species (Holt et a/., 1994; Systematic agenda
2000, 1994) have been described (Table 1.1 ). The above estimates emphasize the
potential of biodiversity and the need for exploration and conservation of biological
species such as the bacteria.
1.1.3 Bacterial Diversity : The Conventional Approach
Animals and plants exhibit an immense range of morphological features which
are of taxonomic importance. In contrast, most prokaryotes are either spheres, rods or
spirals and display few taxonomically informative morphological features. Because of
their much lower morphological diversity, the difficulties associated with bacterial
identification are greater than with eukaryotic algae. Consequently, subcellular attributes
(physiological or biochemical) have been the most useful characteristics for taxonomic
and identification purposes. Identification of bacterial species is typically laborious and
often uncertain. In fact, identification requires isolation of pure cultures and enumeration
of multiple physiological and biochemical traits. But, this approach is also not free of
problems, since it is not possible to culture all the microbes from a given sample due to
incomplete knowledge of their nutritional requirements, their association with other
organisms, their optimum conditions of growth, etc.
1.3.1.1 The great plate count anomaly
Microbiologists for a very long time consistently observed that in situ methods of
detection of microbes always exceeded the viable celi counts by severai orders of
magnitude irrespective of whether the sample analysed originated from aquatic habitats,
sediments or soils. In all these studies, culturable bacteria by viable plate count barely
exceeded 3% of the total cell count as detected by in situ methods (Amann eta/., 1995).
3

Staley and Kanopka (1985) referred to this anomaly as the "Great Plate Count Anomaly"
and it was attributed to our inability to culture microbes because conditions suitable for
growth were not known or because the microbes entered a non-culturable state. For
instance, it is well known that many pathogens like Salmonella enteritidis (Roszak eta/.,
1984), Vibrio cholerae (Colwell et at., 1985) and V. vulnificus (Oliver et a/., 1991)
become non-culturable in the presence of salt water, fresh water or low temperatures.
Some of the microorganisms which have been detected but not yet cultured include the
giant bacterium (500 to 800 J.lm) in the intestine of Surgeonfish (Fishelson et a/., 1985),
the symbiotic microbes of cattle intestines (Wolin, 1979), the bacterial endosymbionts of
protozoa (Gertz, 1986), magnetotactic bacteria (Mann et a/., 1990), deep sea vent
tubeworm associated bacteria (Stahl et a/., 1984), bacteria from Antarctica (Priscu,
1998; Priscu eta/., 1998) etc.
The thousand-fold discrepancy in numbers between direct enumeration and agar
plate counts has cast doubts on plating methods. A lack of bacterial density dependence
in heterotrophic activity has been reported in Antarctic (Bird and Karl, 1991; Hagson et
a/., 1983a, b; Karl et a/., 1991) and sub-Antarctic ecosystems (Deli lie and Bouvy, 1989)
and this lack of direct correlation between bacterial abundance and activity may indicate
that a large and variable proportion of the bacterial cells are dead or inactive. On the
other hand, a sub-Antarctic study (Delille and Bouvy, 1989) revealed that the ratio
between total and culturable bacterial abundance were sometimes relatively low when
compared to other reported oceanic values (Horowitz et a/., 1983; Meyer-Reil et a/.,
1980; Sugita et a/., 1982). Thus, there is an urgent need to study the "viable, non
cultivable fraction" of the bacterial assemblage.
4

1.1.4 Bacterial Diversity : A Modern Approach
Due to the great plate count anomaly, estimates of microbial diversity from
various distinct habitats is bound to be inaccurate. Therefore, there is a need to develop
methods which can identify and characterize the non-culturable microorganisms which
constitute the vast majority (> 97%) (Amann et a/., 1995) based on a culture-
independent approach. This problem was circumvented by the development of
ribosomal RNA sequencing by Carl Weese (Fox eta/., 1980; Weese, 1987; Weese and
Fox, 1977). The molecular sequencing of rRNA established not only a molecular
sequence based phylogenetic analysis but also lead to the survey of various phylotypes
(distribution of microorganisms) in the natural communities without the need to culture
microorganisms (Pace eta/., 1985, 1986).
1.1.4.1 rRNA approach to biodiversity
In this approach, the microbial diversity of an environmental sample is
established based on the sequence analysis of conserved genes such as the rRNA
genes. The procedure involves the following steps: (i) extraction of total community DNA
from the environmental sample (ii) amplification of 168 rRNA gene fragments (iii)
construction of gene libraries (iv) identification of unique clones by RFLP of the cloned
168 rRNA gene fragments (v) sequencing of the rRNA clones and (vi) analysis of the
retrieved sequences by comparison with rRNA database (Chandler et a/., 1997;
Hugenholtz and Pace, 1996).
This approach has now been used to enumerate the biodiversity of symbiotic
bacteria of tubeworms (Riftia pachyptila) (Stahl eta!., 1984) and clams (Calyptogena
sp.) of hydrothermal vents, endosymbiotic bacteria of insects (Unterman et a/., 1989),
bacterial and archaei symbionts of protozoa (Amann et a/., 1991 ), bacterial symbionts of
fish and has also helped in identifying many animal and human pathogens, the
5

magentotactic bacteria which orient themselves along the lines of a magnetic field,
marine pice-plankton and bacteria of biofilms and soils (Amann et a/., 1995). More
recently, the rRNA approach has also been used to identify bacteria from the soils, lakes
and ice of Antarctica (Brambilla et a/., 2001; Pace, 1999a,b; Priscu, 1998) and the
results indicated the presence of a number of unidentified bacteria.
These community based studies have lead to the discovery of greater diversity in
the microbial world than the world of higher organisms (Weese, 1994). Further these
studies indicated that most bacteria are cosmopolitan (distributed in more than one
geographical location on the earth) though a few are endemic to a particular area. Most
of the endemic bacteria are extremophiles since they originate from extreme
environments such as hydrothermal vents, deep sea, earth crust, cold habitats (Arctic,
Antarctic etc.) etc., and are comparatively less studied due to various reasons such as
inability to access such samples, inability to culture them easily, inability to sustain them
in culture for long periods, etc. Therefore, there is a need to study the biodiversity of
extremophiles especially the bacteria about which very little is known with respect to
their biodiversity, taxonomic identity and phylogenetic position.
The rRNA approach definitely reflects the greater degree of biodiversity in an
environmental sample compared to the viable count approach but even in this procedure
problems arise due to: (i) inability to extract nucleic acids from all the microorganisms I
(because some bacteria do not lyse well), (ii) selective amplification of only certain rRNA
genes, (iii) formation of chimeric PCR products and (iv) different rRNA genes having
different cloning efficiencies (Amann et at., 1995). Furthermore, using this approach it
has not been easy to identify the bacteria upto the species level thus making
phylogenetic affiliations difficult. In addition, very few strategies exist to culture the non-
culturable bacteria and identify the unidentified bacteria. Therefore, keeping in view
6

these difficulties, the present study was directed towards the identification of the
culturable bacteria using the conventional approach.
1.1.5 Extremophiles
The first use of the term extremophile appeared in 197 4 in a paper by Mac Elory,
and it referred to organisms that are fond of or tolerate extreme environments such as
high or low temperature; acidic or alkaline pH, high salt, poor or rich nutrients, etc.
Depending on their tolerance, extremophiles have been further designated as :
i) Alkaliphilic : an organism with optimal growth at pH above 10
ii) Barophilic an organism that lives optimally at high hydrostatic pressure
iii) Endolithic an organism that lives in rocks
iv) Acidophilic : an organism with optimal growth at or below pH 3
v) Halophilic : an organism requiring at least 2M NaCI
vi) Oligotrophic an organism with optimal growth in nutrient limited conditions
vii) Xerotolerant : an organism capable of growth at low water activity.
viii) Psychrophilic : an organism having a optimum growth temperature below 15°C
and 20°C as the maximum temperature of growth
ix) Hyperthermophilic : an organism having a optimum growth temperature of 80°C
or higher.
These extremophiles due to their novel and unique abilities to survive in extreme
environments could serve as excellent model systems to understand the limits under
which biological systems could survive. Further, the study of cellular components and
their pathways may provide an insight into their biotechnological potential (Table 1.2).
Among the extremophiles, the psychrophiles, which are present in various habitats such
as Antarctica, Arctic and other cold climates, are the most predominant group, but are
also the least studied.
7

Table 1.2. Extremophiles and their uses in Biotechnology*
Source Use Hyperthermophiles DNA polymerase DNA amplification by PCR Alkaline phosphatase Diagnostics Proteases and lipases Dairy products Lipases, pullulanases, proteases Detergents Proteases, amylases, a-glucosidases Baking and brewing Alcohol dehydrogenase Chemical synthesis Xylanases Paper bleaching Lenthionin Pharmaceuticals Oil degrading microorganisms Surfactants for oil recovery Sulfur oxidizing microorganisms Bioleaching, coal, waste gas desulfurization Hyperthermophilic consortia Waste treatment and methane production Psychrophiles Alkaline phosphatase Molecular biology Proteases, lipases, cellulases, amylases Detergents Lipases, proteases Cheese manufacturing and dairy Production Proteases Contact lens cleaning, meat tenderizing Polyunsaturated fatty acids Food additives, dietary supplements 13-Galactosidases Lactose hydrolysis in milk products Ice nucleating proteins Artificial snow, ice cream, for freezing
Applications Dehydrogenases Biotransformations Oxidases Bioremediation, environmental biosensors Methanogens Methane production
Halophiles Bacteriorhodopsin Optical switches and photocurrent
generators in bioelectronics Polyhydroxyalkanoates Medical plastics Rheological polymers Oil recovery Lipids Liposome for drug delivery and cosmetic
packaging and heating oils Alkaliphiles Proteases, celluloses, xylanases, lipases Detergents Proteases Gelatin removal on X-ray film Elastases, keratinases Hide dehairing Cyclodextrins Food stuffs, chemicals, pharmaceuticals Xylanases and proteases Pulp bleaching Pectinases Acidophiles Sulfur oxidizing microorganisms Recovery of metals, desulfurication of coal
*Cavicchioli and Thomas (2000).
8

1.1.6 Microbiology of Antarctica
Antarctica is the fifth largest continent of the earth's seven continents and is
located entirely south of latitude 66°30'. It is mostly circular in shape with a long arm, the
Antarctic Peninsula reaching out towards South America and with two indentations, the
Ross and Weddell seas and their ice shelves. Antarctica is covered 95% with ice and
contains 90% of the world's fresh water and is considered uninhabitable due to the
extreme cold conditions, high velocity winds, very scanty rainfall, low humidity and non
availability of water. Despite the harsh climatic conditions, certain life forms such as
mites, ticks, seals, penguins, mosses, lichens, bacteria, yeasts, algae, etc. do exist in
Antarctica. Table 1.3 summarizes some physical and climatic features of Antarctica (Fig.
1.1 ).
The distribution and density of life forms in Antarctica is grossly influenced by the
season, availability of water and nutrients. Taking these factors into consideration,
Holdgate (1964) suggested that the continent of Antarctica could be divided into two
zones for the purpose of studying the microorganisms namely, (i) the maritime and (ii)
the continental Antarctica. The temperature on the main continent of Antarctica rarely
exceeds ooc and the terrain is almost permanently covered with frozen ice where as the
maritime Antarctica comprising coastal region, the peninsula and associated islands
have temperatures ranging from ooc to +5°C and the ice coverage is seasonal. The
maritime Antarctic environment can once again be divided into two main areas, the
water column that is subjected to a change due to the weather, radiation, tides etc, and
the benthic sediment that is unaffected by above changes. The water column contains
freely suspended bacteria or bacteria attached to other organisms and the number
decreases with depth. The highest population of bacteria is found where phytoplankton
is abundant. In mud and sand, the number of bacteria again decreases with depth.
9

JO " • o· Gough Island (South AfricafGermany)
So~1th 4Uarwc Ocean
Bird Island (UK) • ~al'1<~e ,,. 1<. r.so· s.:o;'<l S~ii .., VI~
~· ' lA
0 d . ,~ rca as (Argentmal• c-~
Neumayer (Germany) · J<!
Marion Island (South Africa)e lt10' a17 Ocear~
iles Crozet (France)e <to
· .. ~a·tri (India) & Novolazarevskaya (Russia)
HaHey (UK)
iY'owa (JaparJ) Melodezhnaya (Russia,- . •
lies Kerguelen (France) • Maws on '(Australia)
. - Progress (Russia) Zhongshan Chma) • D.avis (.A:ustralia)
~·w • Amundsen-Scott (U. S.) M" 'R . ) DO' E
• vos tok (Russi uny , ussra
• Casey {Australia)
• D umont d'Urville (France)
120"
eJ>almer(U. S . .. • vernads (Ukraine)
• Macquarie Island (Australia)
Rothera (U.KJ• an Martin (ArgentinafGermanv)
150 • Aunnlu
Fig 1.1 Map of Antarctica showing the location of permanent stat ions of different countries
10
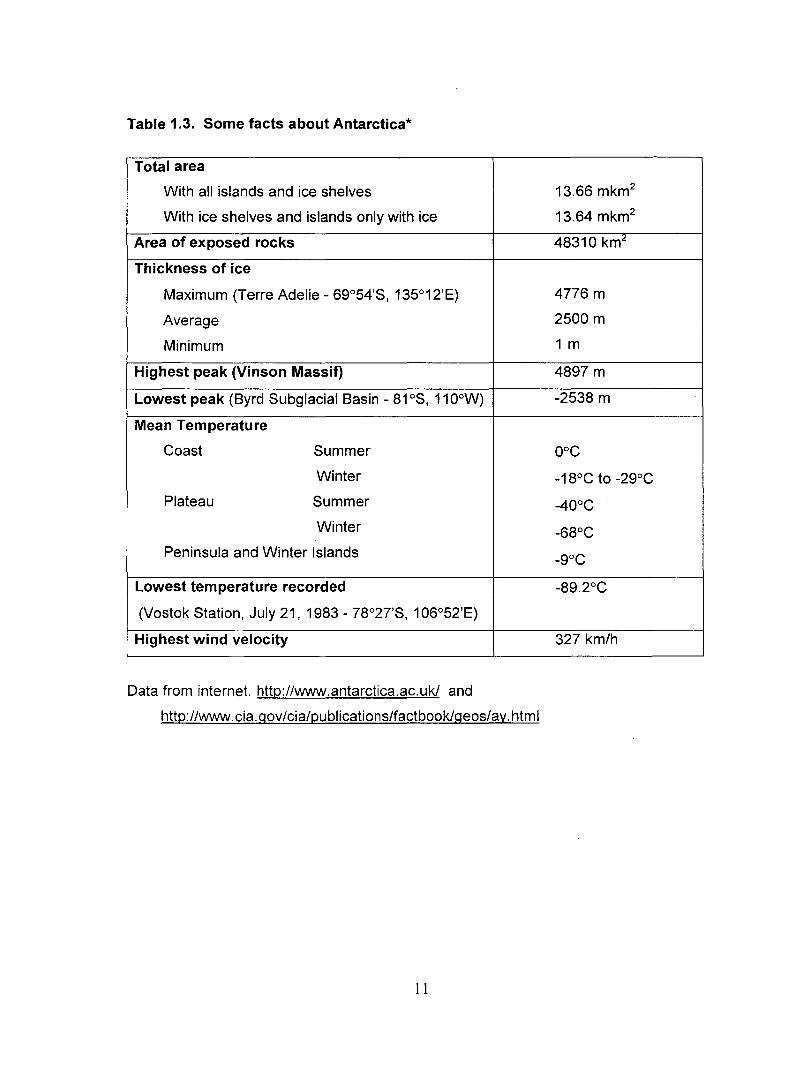
Table 1.3. Some facts about Antarctica*
Total area
With all islands and ice shelves
With ice shelves and islands only with ice
Area of exposed rocks
Thickness of ice
Maximum (Terre Adelie- 69°54'S, 135°12'E)
Average
Minimum
Highest peak (Vinson Massif)
Lowest peak (Byrd Subglacial Basin - 81 as, 11 0°W)
Mean Temperature
Coast Summer
Winter
Plateau Summer
Winter
Peninsula and Winter Islands
Lowest temperature recorded
(Vostok Station, July 21, 1983- 78°27'S, 1 06°52'E)
Highest wind velocity
Data from internet. http://www.antarctica.ac.uk/ and
http://www.cia.gov/cia/publications/factbook/geos/ay.html
11
13.66 mkm2
13.64 mkm2
48310 km2
4776 m
2500 m
1m
4897 m
-2538 m
oac
-18°C to -29°C
-40°C
-68°C
-9aC
-89.2°C
327 km/h

Ekelof, a member of the Swedish National Expedition (1901 to 1903) is regarded
as the father of Antarctic Microbiology. He studied the seasonal variation of microbiota in
the soil and air at Snow Hill Island, Antarctic Peninsula (Ekelof, 1908a,b) and
demonstrated the presence of bacteria, yeasts and fungi for the first time in Antarctica.
These studies of Ekelof were confirmed by Pirie (1904, 1912) of Scottish Antarctic
Expedition, Gazert (1912) of the German Antarctic Expedition (1901-1903) and Tsilinsky
(1908) of the French Antarctic Expedition (1903-1905) who reported the presence of
bacteria in soil and water samples from Laurie Island, South Orkney Island, Wilhelm II
land and Dance Coast, Antarctic Peninsula. Mclean of the Australian expedition (1911-
1914) isolated Gram-positive cocci and Gram-negative and Gram-positive rod shaped
bacteria from Snow and ice at Adelie land near the South pole (McClean 1918a, b). He
also observed that the heterotrophic bacteria not only survive but also multiply at low
temperatures (1 to 2°C). Despite this intensive activity for two decades (1901-1919),
research activity was stalled till 1941, when Darling and Siple isolated close to 180
bacterial cultures from the snow, ice, water, plant debris and soils of Scott Glacier, Ford
Range and Little America and identified them as belonging to the genera
Achromobacter, Flavobacterium and Bacillus. However, the actual impetus to Antarctic
Microbiology was further given by the International Geophysical Year (1957-1958)
following which studies were extended to enumerating bacteria from various habitats of
Antarctica (Flint and Stont, 1960; Straka and Stokes, 1960). Simultaneously, there was
a greater emphasis in trying to identify these unique microorganisms (Boyd, 1962; Boyd
and Boyd, 1962; Friedmann, 1980; Margni and Castrelos, 1963, 1965; Marshall, 1966;
Meyer, 1962; Pfiser, 1965; Tsyganov, 1970). These efforts led to the identification of a
number of new genera and species of bacteria from Antarctica. The various specific
sites in Antarctica from where new taxa have been reported are as follows:
12

i. Fresh water lakes (Franzmann eta/., 1991 and 1997; McCammon eta/., 1998)
ii. Saline and hypersaline lakes and ponds (Bowman eta/., 1997d; Dobson eta/.,
1993; Labrenz et a/., 1998; Mountfort et a/., 1988)
iii. Soil and Sand stone (Bowman eta/., 1996; Friedmann, 1982; Friedmann eta/.,
1993; Hirsch eta/., 1998; Schumann eta/., 1997; Shivaji et at., 1988, 1989a,
1989b, 1991, 1992; Suzuki et at., 1997;)
iv. Sea-ice (Bowman, 1998; Bowman et at., 1997a, e; Junge et at., 1998)
v. Coastal region (Bozal eta/., 1997) and the
vi. Marine environment (Bowman et at., 1997c; Gosink et at., 1998; lrgens et at.,
1996).
Apart from describing new species of bacteria, the spatial distribution and
variability of prokaryotes in various environmental sites such as coastal waters and sea
ice (Oelille, 1996; Franzmann, 1996; Gosink and Staley, 1995; Maugeri et al, 1996;
Murray, 1998; Bowman et a/., 1997a, 1997d, 1997c; Wynn-Williams, 1996), soil
(Friedmann, 1980), glaciers (Abyzov et al, 1998) and lakes (Kriss, 1976; Osnitskaia,
1978, Priscu, 1995, 1998) have been well-documented. The introduction of culture
independent identification of microorganisms by molecular cloning of 16S rONA led not
only to the discovery of unique organisms but also indicated the greater diversity among
Antarctic bacteria (Brambilla et at., 2001; Pace, 1999a, b) most of which have not yet
been cultured. Internet browsing indicated that close to 700, 16S rONA sequences of
bacteria from Antarctica have been deposited in the EMBL data-base and interestingly a
vast majority of the sequences (500 sequences) belonged to the non-culturable Gram
negative and Gram-positive bacteria. The culturable bacteria that have been isolated
and described from various habitats of Antarctica are listed in Tables 1.4 and 1.5. These
bacteria play an important role in influencing nutrient cycling in Antarctica.
13

T bl 14 G a e .. f b t . . I t d t 2002 f ram-nega 1ve ac er.a 1so a e up o rom var1ous h b"tats fA ta f a 1 0 n rc 1ca
Bacterium 165 rRNA gene Habitat Reference
Accession No.*
Achromobacter ruhlandii AF205370 Quartz stone Smith et al. (2000)
Aequorivita crocea AY027806 Quartz stone Smith eta/. (2000),
Bowman and Nicholes (2002)
Aequorivita ferruginea AY027802-4 Quartz stone Smith et al. (2000),
Bowman and Nicholes (2002)
Aequorivita lipolytica AY027805 Quartz stone Smith et al. (2000),
Bowman and Nicholes (2002)
Alteromonas halplanktis X67024 Sea-ice Feller eta/. (1992)
Antarctobacter heliothermus AJ012704 Ekho Lake, Vestfold Hills Labrenz et a/. ( 1998)
Chromobacterium lividum NS Water, maritime Wynn-Williams (1983)
Co/wel/ia demingiae U85845 Sea-ice Bowman eta/. (1998a)
Colwellia hornerae U85847 Sea-ice Bowman eta/. (1998a)
Co/wel/ia psychroerythraea AF001375 Sea-ice Bowman eta/. (1998a)
Co/wel/ia psychrotropica U85846 Sea-ice Bowman eta/. (1998a)
Colwellia rossensis U14581 Sea-ice Bowman eta/. (1998a)
Desulfovibrio sp. NS Lake water Rees eta/. (1986)
Flavobacterium frigidarium AF162266 Marine sediment, Adelaide Island Humphry eta/. (2001)
Flavobacterium gil/isiae U85889 Sea-ice, Pyrdz Bay McCammon and Bowman (2000)
Flavobacterium hibernum L39067 Fresh water McCammon and Bowman (1998)
Flavobacterium tagetincola U85887 Ace Lake, Vestfold Hills McCammon and Bowman (2000)
Flavobacterium xanthum AF030380 Mud pool McCammon and Bowman (2000)
Gelidibacter a/gens U62914 Sea-ice, Vestfold Hills Bowman eta/. (1997a)
Glacieco/a pallidula U85854 Sea-ice Bowman eta/. (1998b)
Glacieco/a punicea U85853 Sea-ice Bowman et a/. ( 1998b)
14

Table 1.4 contd ... Bacterium 165 rRNA gene Habitat Reference
Accession No.
Halobacterium Jacusprofundi X82170 Deep lake water Franzmann eta/. (1988a)
Halomonas subg/caiesco/a L42614 Sea-ice Franzmann eta/. (1987)
Halomonas variabilis U85873 Sea-ice Bowman eta/. (1997e)
Janthinobacterium lividum NS Soils, Scotia Ridge Shivaji eta/. (1991)
Marinomonas protea AJ238597 Water, Ace Lake Mills et a/. (2002)
Methanococcoides burtonii X65537 Water, Ace Lake Franzmann eta/ .. (1992)
Methylosphaera hansonii U67929 Lake Sediment, Vestfold Hills Bowman eta/. (1997c)
Myxococcus stipitatus NS Soil sample Ruckert (1985)
Myxococcus virescens NS Soil sample Ruckert (1985)
Octadecabacter antarcticus U14583 Sea-ice Gosink eta/. (1997)
Octadecabacter sp. AF001377 Sea-ice Bowman eta/. (1997e)
0/eispira antarctica AJ426422 Sea water Yakimov eta/. (2002)
Polaribacter filamentus U73726 Sea water Gosink eta/. (1998)
Polaribacter franzmannii U14586 Sea water Gosink eta/. (1998)
Polaribacter glomeratus M58775 Sea water Gosink et a/. (1998)
Polaribacter irgensii M61002 Sea-ice Gosink eta/. (1998)
Polaromonas vacuolata U14585 Marine water lrgens eta/. (1996)
Pseudoalteromonas antarctica X98336 Marine water Bozal eta/. (1997)
Pseudoa/teromonas clarkii AF283570 Nudibranch Tritoniella belli Frodyma eta/. (2002)
Pseudoa/teromonas prydzensis U85855 Sea-ice Bowman (1998c)
Pseudoalteromonas sp. ANT6 AF025576 Water Hagstrom et a/. (2000)
Pseudoa/teromonas tetraodonis AF025578 Water Ivan ova et al. (2001)
Pseudomonas azotoformans AF025573 Water Hagstrom et a/. (2000)
Pseudomonas fluorescens NS Soils, Schirmacher Oasis Shivaji eta/. (1989)"
Pseudomonas putida NS Soils, Schirmacher Oasis Shivaji eta/. (1989a)
Pseudomonas syringae NS Soils, Schirmacher Oasis Shivaji eta/. (1989a)
15
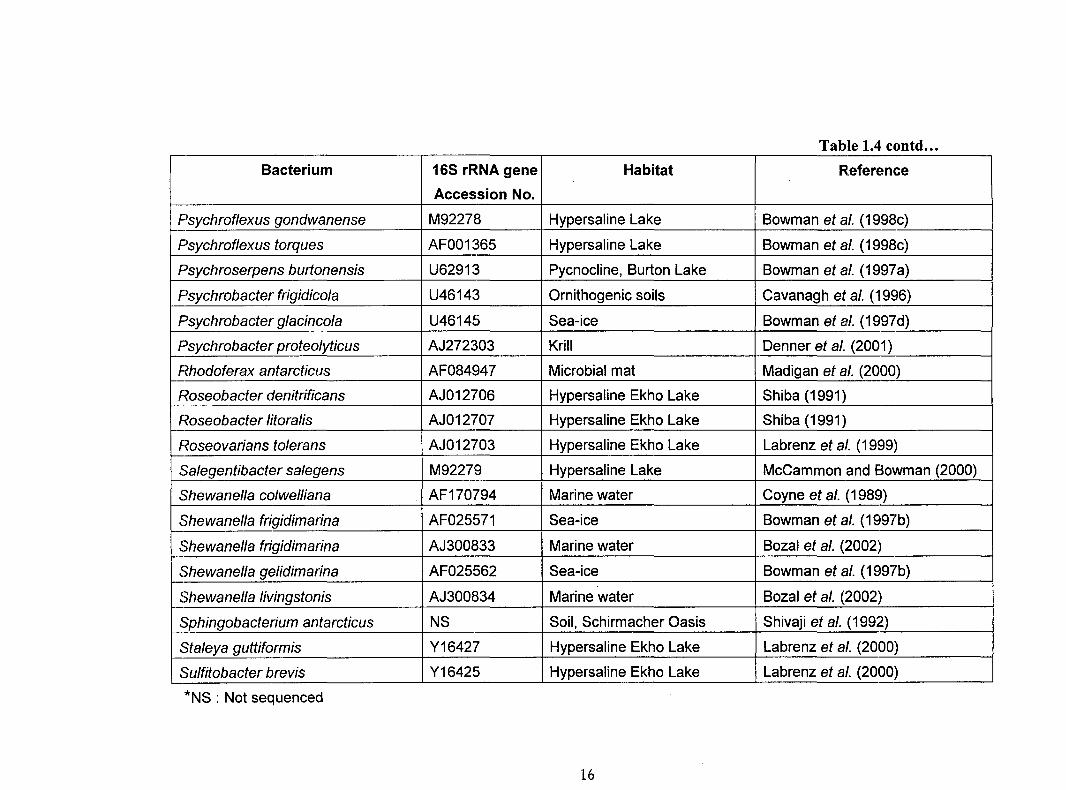
Table 1.4 contd ...
Bacterium 165 rRNA gene Habitat R.eference
Accession No.
Psychroflexus gondwanense M92278 Hypersaline Lake Bowman eta/. (1998c) ~-
Psychroflexustorques AF001365 Hypersaline Lake Bowman eta/. (1998c)
Psychroserpens burlonensis U62913 Pycnocline, Burton Lake Bowman eta/. (1997a)
Psychrobacter frigidico/a U46143 Ornithogenic soils Cavanagh eta/. (1996)
Psychrobacter glacinco/a U46145 Sea-ice Bowman eta/. (1997d)
Psychrobacter proteolyticus AJ272303 Krill Denner eta/. (2001)
Rhodoferax antarcticus AF084947 Microbial mat Madigan et a/. (2000) r--Roseobacter denitrificans AJ012706 Hypersaline Ekho Lake Shiba (1991)
Roseobacter Jitoralis AJ012707 Hypersaline Ekho Lake Shiba (1991)
Roseovarians tolerans AJ012703 Hypersaline Ekho Lake Labrenz eta/. (1999)
Salegentibacter salegens M92279 Hypersaline Lake McCammon and Bowman (2000)
Shewanella colwelliana AF170794 Marine water Coyne eta/. (1989)
Shewanella frigidimarina AF025571 Sea-ice Bowman eta/. (1997b)
Shewanella frigidimarina AJ300833 Marine water Bozal et a/. (2002)
Shewanella gelidimarina AF025562 Sea-ice Bowman eta/. (1997b)
Shewanella livingstonis AJ300834 Marine water Bozal et a/. (2002)
Sphingobacterium antarcticus NS Soil, Schirmacher Oasis Shivaji eta/. (1992)
Sta/eya guttiformis Y16427 Hypersaline Ekho Lake Labrenz et a/. (2000)
Sulfitobacter brevis Y16425 Hypersaline Ekho Lake Labrenz et a/. (2000)
*NS : Not sequenced
16
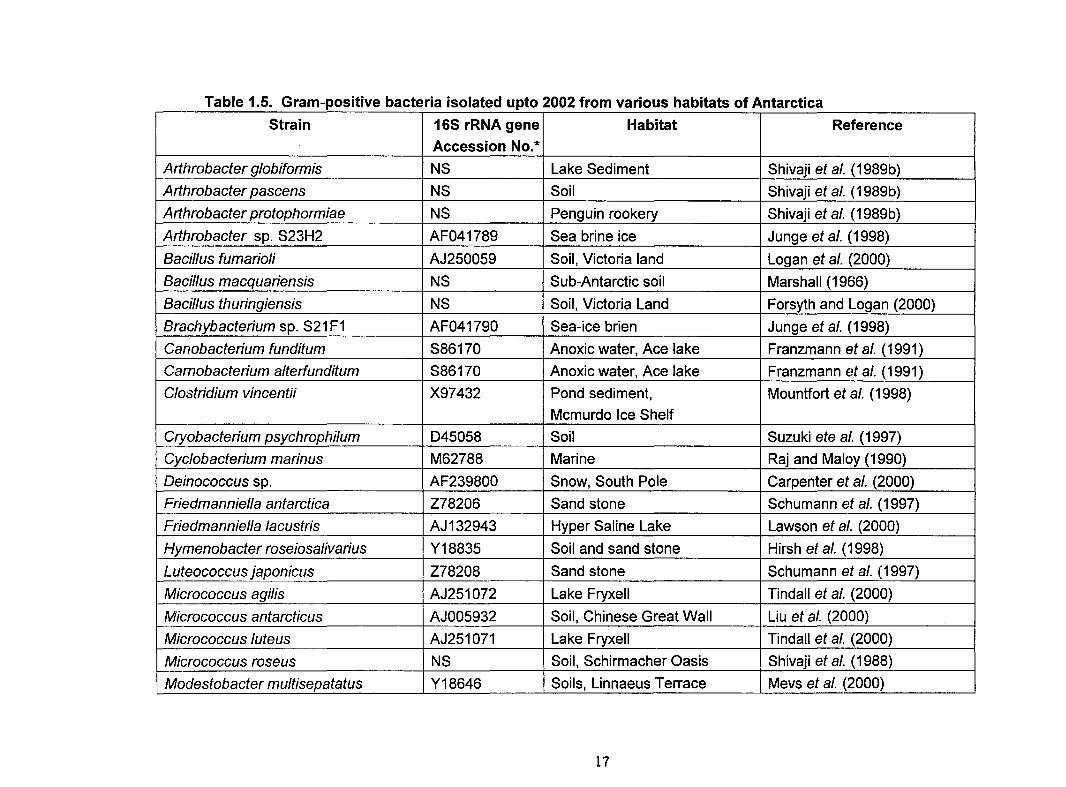
Table 1.5. Gram-positive bacteria isolated upto 2002 from various habitats of Antarctica
Strain 165 rRNA gene Habitat Reference
Accession No.*
Arthrobacter globiformis NS Lake Sediment Shivaji eta/. (1989b)
Arthrobacter pascens NS Soil Shivaji eta/. (1989b)
Arthrobacter protophormiae NS Penguin rookery Shivaji eta/. (1989b)
Arthrobacter sp. S23H2 AF041789 Sea brine ice Junge eta/. (1998)
Bacillus fumarioli AJ250059 Soil, Victoria land Logan et a/. (2000)
Bacillus macquariensis NS Sub-Antarctic soil Marshall (1966)
Bacillus thuringiensis NS Soil, Victoria Land Forsyth and Logan (2000)
Brachybacterium sp. S21 F1 AF041790 Sea-ice brien Junge eta/. (1998)
Canobacterium funditum S86170 Anoxic water, Ace lake Franzmann eta/. (1991)
Carnobacterium alterfunditum S86170 Anoxic water, Ace lake Franzmann eta/. (1991)
Clostridium vincentii X97432 Pond sediment, Mountfort eta/. (1998)
Mcmurdo Ice Shelf
Cryobacterium psychrophilum 045058 Soil Suzuki ete a/. (1997)
Cyc/obacterium marinus M62788 Marine Raj and Maloy (1990)
Deinococcus sp. AF239800 Snow, South Pole Carpenter et a/. (2000)
Friedmanniel/a antarctica Z78206 Sand stone Schumann eta/. (1997)
Friedmanniella lacustris AJ132943 Hyper Saline Lake Lawson et al. (2000)
Hymenobacter roseiosalivarius Y18835 Soil and sand stone Hirsh eta/. (1998)
Luteococcus japonicus Z78208 Sand stone Schumann eta/. (1997)
Micrococcus agi/is AJ251072 Lake Fryxell Tindall et a/. (2000)
Micrococcus antarcticus AJ005932 Soil, Chinese Great Wall Liu et a/. (2000)
Micrococcus luteus AJ251071 Lake Fryxell Tindall et a/. (2000)
Micrococcus roseus NS Soil, Schirmacher Oasis Shivaji eta/.. (1988)
Modestobacter multisepatatus Y18646 Soils, Linnaeus Terrace Mevs et a/. (2000)
17

Table 1.5 contd ••.
Strain 16S rRNA gene Habitat Reference
Accession No
Nocardioides aquaticus AJ132943 Hypersaline Ekho Lake Lawson et a/. (2000)
Nocardiopsis antarcticus X 97885 Yassin eta/. (1997)
Planococcus mcmeekinii AF 041791 Sea-ice Brine Junge eta/. (1998)
Planococcus sp. MB6-16 u 85898 Sea-ice Bowman eta/. (1997e)
Planococcus sp. AF 242541 Hypersaline Pond, Sheridan and Brenchley (2000)
Mcmurdo Ice Shelf
P/anococcus spp. NS Soil, Schirmacher Oasis Shivaji eta/. (1988)
Rhodococcus fascians AF 205371 Quartz stone subliths Smith et a/. (2000)
Kocuria polaris AJ 278868 Cyanobacterial mat sample Reddy eta/. (2002c)
Arthrobacter flavus AJ 242532 Cyanobacterial mat sample Reddy et a/. (2000)
Arthrobacter roseus (CMS 90r) AJ 278870 Cyanobacterial mat sample Reddy et a/. (2002a)
Arthrobacter roseus (CMS 92r) AJ 278869 Cyanobacterial mat sample Reddy eta/. (2002a)
Planococcus antarcticus (CMS 3or) AJ 314744 Cyanobacterial mat sample Reddy eta/. (2002b)
Planococcus antarcticus (CMS 26or) AJ 314745 Cyanobacterial mat sample Reddy eta/. (2002b)
P/anococcus antarcticus (CMS 84or) AJ 314747 Cyanobacterial mat sample Reddy et a/. (2002b)
Planococcus psychrophi/us AJ 314746 Cyanobacterial mat sample Reddy eta/. (2002b)
*NS, Not Sequenced.
18

1.1.7 Nutrient Recycling
Johannes (1968) has defined nutrient recycling as the release of soluble organic
or inorganic nutrients by or from living cells or their remains. The major nutrients, which
are required in abundance are carbon, nitrogen and phosphorous. Oxygen, hydrogen
and sulfur are also required but are readily available and therefore, are not considered
as nutrients. The other elements, such as iron, which are required in small quantity are
called "trace elements". As of now, very little is known about the recycling of nutrients by
bacteria in Antarctica (Stokes and Redmond, 1966). However, it is well-established that
nutrient regeneration is very high in Antarctic waters, which are rich in phytoplankton
production (Herbert and Bell, 1974).
Tanner and Herbert (1981) were the first to estimate simultaneous changes in
phytoplankton populations, inorganic nutrient loads, total aerobic and heterotrophic
bacteria, denitrifying bacteria, proteolytic bacteria, sulphate-reducing bacteria and
nitrogen-fixing bacteria in soils and water and concluded that a symbiotic association
exists between heterotrophic bacteria and phytoplankton and thus, they combinedly
influence nutrient cycling. Further, Deli lie and Legarde (197 4), while investigating the
bacterial activity in Kerguelan Island observed seasonal variations in the cycling of
sulphur. Subsequent studies on psychrophilic bacteria from Antarctica demonstrated the
presence of heterotrophic bacteria capable of fixing nitrogen (Herbert and Bell, 197 4;
Tanner, 1981, 1985; Tanner and Herbert, 1981 ). Voytek and Ward (1995) detected
ammonia-oxidizing bacteria belonging to the beta-subclass of the proteobacteria from
Lake Bonney, Antarctica and further supported the occurrence of nitrification and the
presence of nitrifiers. Nutrient recycling is also dependent on the ability of bacteria to
produce extracellular enzymes such as proteases, lipases, amylases, etc., which break
19

down complex organic molecules to simple carbon compounds which can be utilized as
carbon sources. Several bacteria belonging to various different genera such as
Pseudomonas, Flavobacterium, Moraxella, Alcaligenes (Chessa et at., 2000; Kelly et at.,
1978), Psychrobacter (Cavanagh eta/., 1996; Denner et at., 2001), Clostridium (Dube et
a/., 2001), Alteromonas (Aghajari et a/., 1998a, b), Sphingobacterium antarcticus
(Chattopadhyay et a/., 1995), Shewanel/a sp. (Tsuruta et a/., 1998), Vibrio sp.
(Asgeirsson and Andresson, 2001) etc. and capable of producing the above enzymes
have been identified and studied. In addition, Antarctic yeasts with abilities to produce
extracellular enzymes such as proteases have also been implicated in nutrient recycling
(Demot and Verachtert, 1987; Ray et at., 1992).
1.1.8 Objectives of the Present Study
The extreme climatic conditions such as low temperature, permanently covered
ice, strong winds, low humidity, non-availability of water and minimal rain fall supports
very few life forms on terrestrial regions of Antarctica. However, most of the lakes and
ponds, though they are covered with a thick ice sheet, are dominated by cyanobacteria
present as visible mats underneath the ice shelf. In association with these mats, there
exist many heterotrophic bacteria that dominate the cyanobacteria by number (3 to 5
times) and the concentration ranges from 3.2 X 104 to 4.4 X 107 cells/ml of water. Though
cyanobacteria play a significant role in maintaining the biological activity in Antarctica,
the role of heterotrophic bacteria associated with it and the type of heterotrophs present
in lakes and ponds of Antarctica are not thoroughly studied. Therefore, the present
investigation was focused at characterizing the heterotrophic b3cteria associated with
20

the cyanobacterial mats collected from various ponds and lakes of the McMurdo Dry
Valley region.
The objectives of the present study are summarized as follows :
i. To isolate and identify the culturable bacteria present in association
with cyanobacterial mats up to the species level.
ii. To determine the phylogenetic position of the unique species isolated
from the cyanobacterial mat samples.
iii. To establish physiological groups on the basis of enzymes involved in
iv.
biodegradation.
To evaluate the biotechnological potential of these microorganisms.
·~
F
21







