
RESEARCH LETTERS Open Access
Downregulation of CDKL1 suppressesneuroblastoma cell proliferation, migrationand invasionWeiyi Li1, Jing Cao2, Jian Liu1, Wenli Chu1, Congqing Zhang1, Shuiling Chen1 and Zefeng Kang1*
* Correspondence: [email protected] Hospital, China Academy ofChinese Medical Sciences, No 33Lugu Road, Shijingshan district,Beijing 100040, ChinaFull list of author information isavailable at the end of the article
Abstract
Background: Cyclin-dependent kinase-like 1 (CDKL1) is a member of the cell divisioncontrol protein 2-related serine–threonine protein kinase family. It is known to occurin various malignant tumors, but its role in neuroblastoma (NB) remains unclear.
Methods: We constructed a CDKL1-silenced NB cell strain (SH-SY5Y) and used real-time PCR and western blotting to confirm the silencing. Functional analyses wereperformed using the MTT, colony-formation, FACS, wound-healing and transwellinvasion assays.
Results: The expression of CDKL1 was significantly upregulated in NB tissue ascompared to the adjacent normal tissue. CDKL1 knockdown significantly suppressedcell viability and colony formation ability. It also induced cell cycle G0/G1 phasearrest and apoptosis, and suppressed the migration and invasion ability of SH-SY5Ycells. CDKL1 knockdown decreased the CDK4, cyclin D1 and vimentin expressionlevels, and increased the caspase-3, PARP and E-cadherin expression levels in SH-SY5Y cells.
Conclusions: Our findings suggest that CDKL1 plays an important role in NBcell proliferation, migration and invasion. It might serve as a potential targetfor NB therapy.
Keywords: Neuroblastoma, SH-SY5Y, CDKL1, Proliferation, Migration, Invasion
BackgroundNeuroblastoma (NB) is a solid carcinoma of the developing sympathetic nervous sys-
tem [1, 2]. It frequently metastasizes to bone with a mortality rate above 93% [3]. It
commonly arises in young children and accounts for 15% of all childhood
cancer-related deaths [4, 5]. Despite remarkable progress in therapeutic strategies, the
prognosis of NB is still poor.
Accumulating evidence suggests that a series of tumor-suppressor genes and onco-
genes are closely associated with the pathogenesis and development of NB [6, 7]. Iden-
tification and characterization of biomarker candidates in NB may provide critical
clues for the development of therapeutic approaches.
Protein kinase pathways are considered to regulate a wide range of cellular physio-
logical processes, including metabolism, cell division and cell death [8, 9]. The CDK
family (including CDK1 through CDK20) is a group of serine–threonine kinases that
Cellular & MolecularBiology Letters
© The Author(s). 2019 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 InternationalLicense (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium,provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, andindicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 https://doi.org/10.1186/s11658-019-0139-z

could modulate G1/S and G2/M cell cycle checkpoints by forming active CDK–cyclin
complexes [10, 11]. For example, CDC2 (CDK1) cooperates with cyclin A to mediate
G2/M transition and with cyclin B to stimulate mitosis in mammalian cells [12]. Based
on their biochemical and genetic structures, cyclin-dependent kinase-like 1–3
(CDKL1–3), CDK10 and PCTAIRE are considered to be CDC2-related kinase family
members [13]. CDKL1, which is also a member of the CDKL kinase family, has the
conserved MAP kinase Thr-Xaa-Tyr (Thr-Asp-Tyr) dual phosphorylation motif [14].
The transition of CDKL1 is distributed in various organs including the brain, lungs,
kidneys and ovaries [14]. Hsu et al. [15] identified zebrafish CDKL1 and showed that
knockdown of CDKL1 decreased neuogenin-1 expression and lead to abnormal devel-
opment of the brain. Recent studies further demonstrated that CDKL1 is associated
with the development and progression of malignant tumors, including gastric cancer
[16], breast cancer [17], melanoma [18] and colorectal cancer [19]. CDKL1 is highly
expressed in gastric cancer tissues and its disruption reduces cell viability and induces
apoptosis in gastric cancer cells [16]. Furthermore, RNAi-mediated knockdown of
CDKL1 suppressed cell growth and metastasis, promoted cell death and caused G1
phase arrest in human melanoma cells [18].
Interestingly, the fetal form of CDKL1 has been shown to exist in cultured astrocytes
and neuroblastoma cells [20]. Moreover, an earlier study found that rat neuroblastoma
cells exhibit elevated CDKL1 expression [20]. However, the biological function of
CDKL1 in NB remains largely unknown.
In this study, we observed overexpression of CDKL1 in NB tissues as compared with
adjacent tissues. Our in vitro experiments indicate that downregulation of CDKL1 at-
tenuated growth and invasion, and induced cell cycle arrest and apoptosis in NB
SH-SY5Y cells. These results indicate that CDKL1 functions as an oncogene in NB.
Our investigation may provide critical starting points for novel therapeutic
interventions.
Materials and methodsClinical specimens
Samples of NB tissue and corresponding adjacent normal nerve tissue were obtained
from 8 patients attending the Eye Hospital of the China Academy of Chinese Medical
Sciences in Beijing. All the clinical specimens were frozen immediately after surgery
and stored at − 80 °C for real-time PCR and western blotting analysis. All patients pro-
vided written consent.
Cell lines and transfection
Human NB cell lines, including SH-SY5Y, LAN5 and SH-EP, were obtained from the
American Type Culture Collection (ATCC) and cultured in Dulbecco’s modified Eagle’s
medium (DMEM, Invitrogen) containing 10% fetal bovine serum (FBS, Invitrogen). Hu-
man nerve cell line U343 was cultured in Ham F10/DMEM (1:1, Sigma) medium sup-
plemented with 15% FBS (GIBCO). All cells were maintained in a humidified
atmosphere containing 5% CO2 at 37 °C.
To downregulate the expression of CDKL1, a small interfering RNA (siRNA) target
sequence (5′-CTACTGTGATACCAAGAAA-3′) for the CDKL1 gene (NM_004196)
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 2 of 11

was designed and constructed. The non-sense sequence (5′-TTCTCCGAACGTGT
CACGT-3′) was designed and used as the negative control (NC). These siRNA con-
structs were then cloned into the GV115 plasmid vector (GeneChem), which contains
the green fluorescent protein (GFP) gene as a reporter. Next, lentivirus particles were
constructed by transfecting the siRNA plasmid into HEK293 cells. For cell transfection,
we added lentivirus particles expressing CDKL1-siRNA (siCDKL1) or NC to 6-well
plates containing SH-SY5Y cells grown to confluence of 1 × 104 cells per well. The
multiplicity of infection (MOI) was 20. After 48 h transfection, the transfection effi-
ciency was detected in stably transfected GFP-expressing cells using fluorescence mi-
croscopy (Olympus).
Real-time PCR
Total RNA was isolated from tissues or cells using Trizol reagent (Invitrogen) and re-
verse transcribed to cDNA using PrimeScript RT Master Mix (Perfect Real Time,
Takara) according to the manufacturers’ protocols. The cDNA was amplified via
real-time PCR using a Thermal Cycle Dice Real Time System TP800 (Takara). The fol-
lowing primers were used:
� CDKL1 forward: 5′-CGAATGCTCAAGCAACTCAAGC-3′, CDKL1 reverse: 5′-
GCCAAGTTATGCTCTTCACGAG-3′
� β-actin forward: 5′-TCAGGTCATCACTATCGGCAAT-3′, β-actin reverse: 5′-
AAAGAAAGGGTGTAAAACGCA-3′
The expression of CDKL1 mRNA was normalized to β-actin and calculated using the
2-ΔΔCt method. All samples were prepared in triplicate and analyzed three times.
Western blotting
Total protein lysates were extracted from tissues or cells by lysing in cell lysis buffer
(Sigma-Aldrich). The protein concentration in the lysates was quantified using a BCA
protein assay kit (Thermo Scientific). Equal amounts of proteins were then separated
via SDS-PAGE, followed by transfer to PVDF membrane (Millipore). After blocking
with 5% skim milk, the membrane was incubated with primary antibodies against
CDKL1 (1:1000, Abcam, UK), CDK4 (1:1000, Proteintech, USA), cyclin D1 (1:1000,
Proteintech, USA), caspase-3 (1:1000, Proteintech, USA), PARP (1:1000, Cell signaling,
USA), E-cadherin (1:1000, Proteintech, USA), vimentin (1:1000, Cell signaling, USA) or
anti-GAPDH (1:5000, Proteintech, USA), followed by incubation with horseradish
peroxidase-conjugated secondary antibody (1:10000, China). The immunoreactive
bands were visualized using an enhanced chemiluminescence kit (Thermo Fisher).
MTT assay
Transfected cells were plated in 96-well plates at a density of 2000 cells/well and the
plates were incubated in 5% CO2 at 37 °C. The cell viability was determined via MTT
assay after 24 and 48 h. Briefly, 100 μl MTT (0.5 mg/ml, Sigma-Aldrich) was added into
each well and incubated for 4 h. The supernatant was absorbed and 150 μl DMSO was
added to each well to dissolve the formazan crystals. The optical density (OD) value of
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 3 of 11

each well was measured at a wavelength of 595 nm with a microplate reader
(ThermoFisher).
Colony formation assay
Transfected cells were resuspended in medium and seeded in 6-well plates at a density
of 800 cells per well. After 10 days of culture, the cells were fixed with paraformalde-
hyde (Sangon Biotech) for 15 min and stained with 0.1% crystal violet (BioSharp) for
20 min. The colonies (> 50 cells per colony) were photographed and counted.
Flow cytometry analysis
After transfection, the cells were collected, washed with PBS and seeded in 8-cm steril-
ized culture dishes for 12 h. Then the cells were fixed with 70% ethanol for 2 h and
stained with 5 mg/ml propidium iodide (PI; BD Biosciences). After incubation for 15
min in the dark at room temperature, stained cells were analyzed for DNA content
using FACSCalibur flow cytometry (BD Biosciences). For cell apoptosis analysis, the
cells were stained with an Annexin V–APC/7-AAD Apoptosis Detection kit (KeyGEN)
according to the manufacturer’s instructions. Early apoptotic (annexin V+/7-AAD-)
and late apoptotic (annexin V+/7-AAD+) cells were analyzed and quantitated using
FACSCalibur flow cytometry (BD Biosciences).
Wound-healing assay
Approximately 1 × 106 cells were cultivated in 6-well culture plates (Corning Inc.) and
grown to 90% confluence overnight. Subsequently, a line was scratched with a 200 μl
pipette tip (time 0) and cells were incubated with serum-free medium for the next 48 h.
After washing with PBS, the migrating cells were photographed at 0 and 48 h. The mi-
gration distance was calculated during closure of the wounded region.
Trans-well invasion assay
For the transwell invasion assay, about 1 × 105 cells/ml transfected cells were seeded in
the upper chamber with Matrigel Basement Membrane Matrix in serum-free DMEM
medium at 37 °C for 24 h. Then, 500 μl DMEM medium containing 10% FBS was added
to the lower chamber. The invasive cells, present on the underside of the upper cham-
ber, were then fixed with 4% paraformaldehyde and stained with 2% crystal violet.
Stained cells were photographed under a light microscope. Each sample was prepared
in triplicate and analyzed three times.
Statistical analysis
Quantitative data were representative of three sets of independent experiments per-
formed in triplicate for each group. The experimental data are expressed as means ±
standard deviation (SD). Statistical analysis was performed using GraphPad Prism soft-
ware. Student’s t test was used to evaluate the difference between two groups and the
level of significance was set at p < 0.05.
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 4 of 11
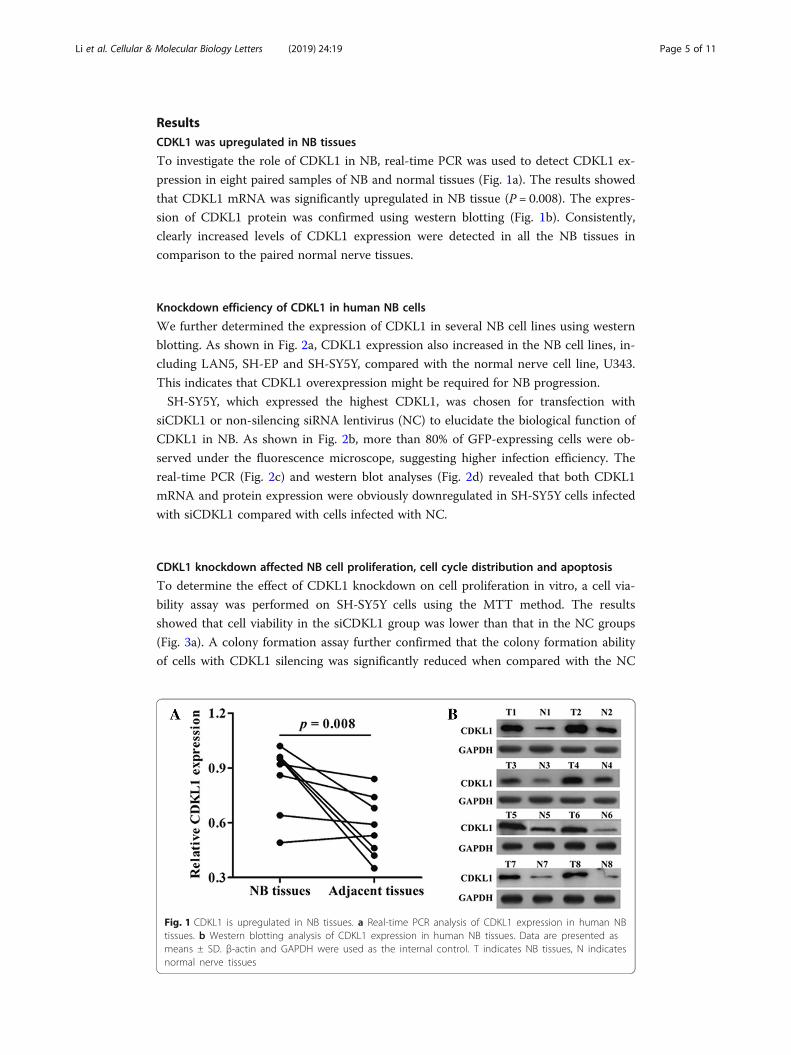
ResultsCDKL1 was upregulated in NB tissues
To investigate the role of CDKL1 in NB, real-time PCR was used to detect CDKL1 ex-
pression in eight paired samples of NB and normal tissues (Fig. 1a). The results showed
that CDKL1 mRNA was significantly upregulated in NB tissue (P = 0.008). The expres-
sion of CDKL1 protein was confirmed using western blotting (Fig. 1b). Consistently,
clearly increased levels of CDKL1 expression were detected in all the NB tissues in
comparison to the paired normal nerve tissues.
Knockdown efficiency of CDKL1 in human NB cells
We further determined the expression of CDKL1 in several NB cell lines using western
blotting. As shown in Fig. 2a, CDKL1 expression also increased in the NB cell lines, in-
cluding LAN5, SH-EP and SH-SY5Y, compared with the normal nerve cell line, U343.
This indicates that CDKL1 overexpression might be required for NB progression.
SH-SY5Y, which expressed the highest CDKL1, was chosen for transfection with
siCDKL1 or non-silencing siRNA lentivirus (NC) to elucidate the biological function of
CDKL1 in NB. As shown in Fig. 2b, more than 80% of GFP-expressing cells were ob-
served under the fluorescence microscope, suggesting higher infection efficiency. The
real-time PCR (Fig. 2c) and western blot analyses (Fig. 2d) revealed that both CDKL1
mRNA and protein expression were obviously downregulated in SH-SY5Y cells infected
with siCDKL1 compared with cells infected with NC.
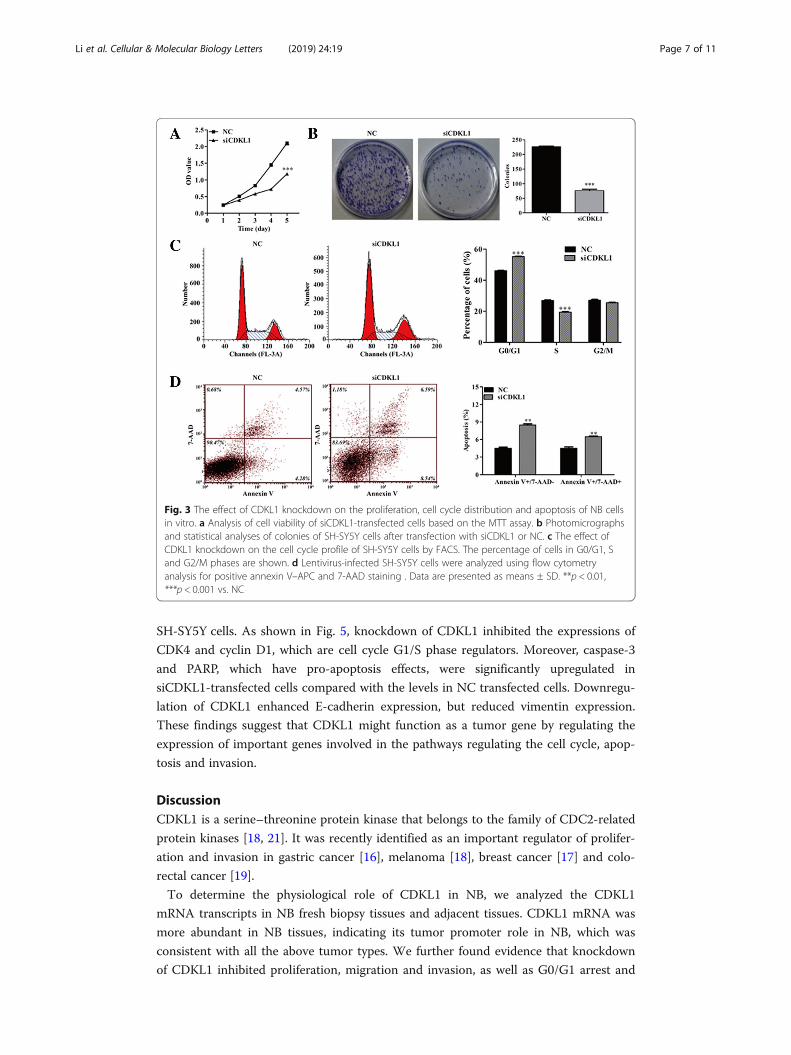
CDKL1 knockdown affected NB cell proliferation, cell cycle distribution and apoptosis
To determine the effect of CDKL1 knockdown on cell proliferation in vitro, a cell via-
bility assay was performed on SH-SY5Y cells using the MTT method. The results
showed that cell viability in the siCDKL1 group was lower than that in the NC groups
(Fig. 3a). A colony formation assay further confirmed that the colony formation ability
of cells with CDKL1 silencing was significantly reduced when compared with the NC
Fig. 1 CDKL1 is upregulated in NB tissues. a Real-time PCR analysis of CDKL1 expression in human NBtissues. b Western blotting analysis of CDKL1 expression in human NB tissues. Data are presented asmeans ± SD. β-actin and GAPDH were used as the internal control. T indicates NB tissues, N indicatesnormal nerve tissues
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 5 of 11

groups (Fig. 3b), suggesting that CDKL1 knockdown results in the inhibition of NB cell
proliferation.
As CDKL1 is related to CDKs, we examined the effect of CDKL1 knockdown on cell
cycle regulation using flow cytometry. As shown in Fig. 3c, there was an increase in the
proportion of cells in the G0/G1 phase (55.18 ± 1.14 vs. 46.06 ± 0.78%, p < 0.001) in the
siCDKL1 group, which also displayed fewer cells in S phase (19.38 ± 1.05 vs. 26.85 ±
0.94%, p < 0.001) compared with the NC group.
We also evaluated the rate of apoptosis, aiming to investigate the effects of CDKL1
knockdown on the cell death of SH-SY5Y cells. As depicted in Fig. 3D, downregulation
of CDKL1 significantly elevated the early apoptotic rate from 4.49% ± 0.21 to 8.49% ±
0.17% and the late apoptotic rate from 4.51% ± 0.31 to 6.48% ± 0.54% (p < 0.01). These
data demonstrate that suppression of cell proliferation might be caused by cell cycle ar-
rest and apoptosis induced by CDKL1 knockdown in NB cells.
CDKL1 knockdown affected NB cell migration and invasion ability
We subsequently investigated the effect of CDKL1 knockdown on cell migration and
invasion. The wound healing assay results showed that SH-SY5Y cells transfected with
siCDKL1 had a lower scratch closure rate (migration suppression) than that for control
cells (Fig. 4a, p < 0.01).
We then observed NB cell invasion through transwell matrigel. As shown in Fig. 4b,
reduced CDKL1 expression levels significantly hindered the invasion of SH-SY5Y cells
compared with controls (p < 0.001). These findings demonstrate that CDKL1 might be
closely associated with the migration and invasion of NB cells.
Molecular targets of CDKL1 in NB cells
To gain insights into the molecular mechanisms of CDKL1 knockdown on the regula-
tion of the cell cycle, apoptosis and invasion, western blot analysis was performed on
Fig. 2 The efficiency of CDKL1 using an siRNA lentivirus. a Western blotting analysis of the CDKL1expression in human NB cell lines. b Lentivirus infection in the SH-SY5Y cell line. Bright and fluorescentphotomicrographs of SH-SY5Y cells were taken 48 h after lentivirus infection. c and d Real-time PCR andwestern blotting were used to determine the CDKL1 expression in SH-SY5Y cells infected with siCDKL1 orthe negative control (NC) siRNA lentivirus. Data are presented as means ± SD. **p < 0.01 vs. NC
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 6 of 11
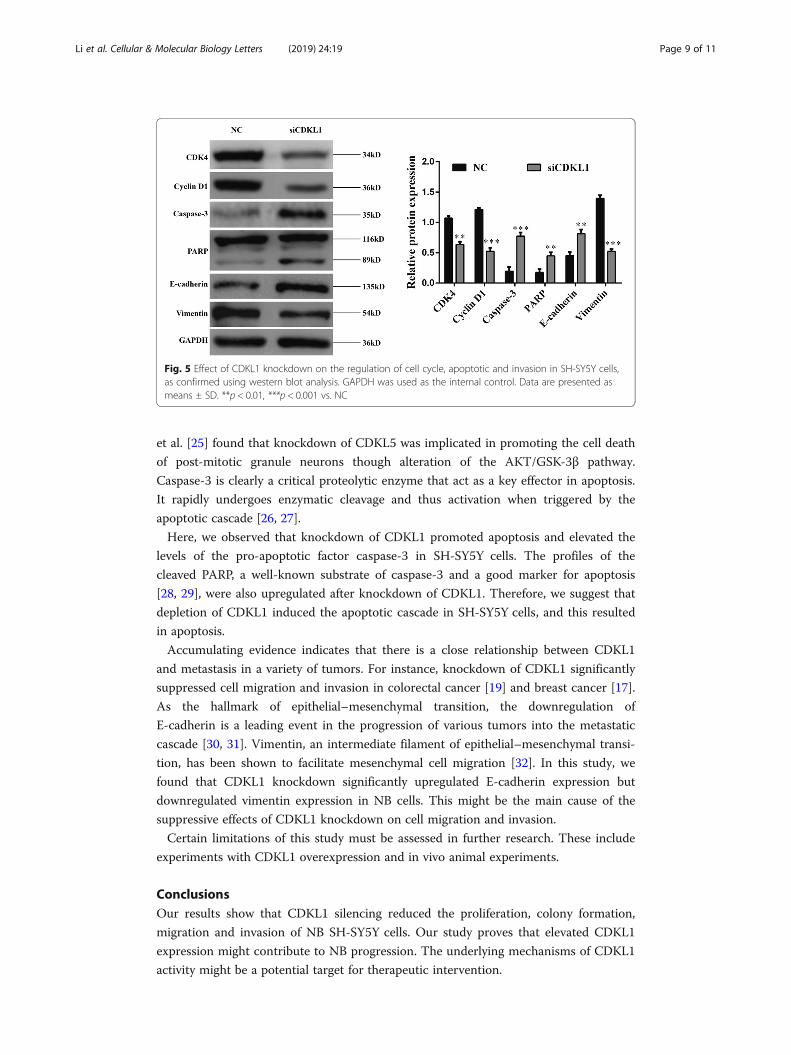
SH-SY5Y cells. As shown in Fig. 5, knockdown of CDKL1 inhibited the expressions of
CDK4 and cyclin D1, which are cell cycle G1/S phase regulators. Moreover, caspase-3
and PARP, which have pro-apoptosis effects, were significantly upregulated in
siCDKL1-transfected cells compared with the levels in NC transfected cells. Downregu-
lation of CDKL1 enhanced E-cadherin expression, but reduced vimentin expression.
These findings suggest that CDKL1 might function as a tumor gene by regulating the
expression of important genes involved in the pathways regulating the cell cycle, apop-
tosis and invasion.
DiscussionCDKL1 is a serine–threonine protein kinase that belongs to the family of CDC2-related
protein kinases [18, 21]. It was recently identified as an important regulator of prolifer-
ation and invasion in gastric cancer [16], melanoma [18], breast cancer [17] and colo-
rectal cancer [19].
To determine the physiological role of CDKL1 in NB, we analyzed the CDKL1
mRNA transcripts in NB fresh biopsy tissues and adjacent tissues. CDKL1 mRNA was
more abundant in NB tissues, indicating its tumor promoter role in NB, which was
consistent with all the above tumor types. We further found evidence that knockdown
of CDKL1 inhibited proliferation, migration and invasion, as well as G0/G1 arrest and
Fig. 3 The effect of CDKL1 knockdown on the proliferation, cell cycle distribution and apoptosis of NB cellsin vitro. a Analysis of cell viability of siCDKL1-transfected cells based on the MTT assay. b Photomicrographsand statistical analyses of colonies of SH-SY5Y cells after transfection with siCDKL1 or NC. c The effect ofCDKL1 knockdown on the cell cycle profile of SH-SY5Y cells by FACS. The percentage of cells in G0/G1, Sand G2/M phases are shown. d Lentivirus-infected SH-SY5Y cells were analyzed using flow cytometryanalysis for positive annexin V–APC and 7-AAD staining . Data are presented as means ± SD. **p < 0.01,***p < 0.001 vs. NC
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 7 of 11

apoptosis. These results indicate that CDKL1 might play an important role in the oc-
currence and development of NB.
Recent studies confirmed that knockdown of CDKL1 lead to a blockage of cell
growth and arrest of G0/G1 phase in colorectal cancer [19] and melanoma cells [18].
By contrast, CDKL1 is associated with G2/M phase cell cycle arrest in breast cancer
[17]. In our study, there was a significant increase in the G0/G1 fraction and accom-
panying decrease in the S fraction.
This suggests that CDKL1 may exert various functions in cell cycle progression pos-
sibly based on the tumor type. As is known to all, tumor initiation and progression are
associated with cell cycle defects, which are regulated by CDK–cyclin complexes [22].
In mammalian cells mitogens impact development by triggering three D-type cyclins
(D1, D2, and D3) which function coordinately with CDKs (CDK4 or CDK6) to promote
cell cycle transition from G1 to S [23]. In our study, a significant reduction in CDK4
and cyclin D1 protein levels was observed in the CDKL1 knockdown group compared
with the control group. We thus concluded that downregulation of CDKL1 is involved
in the blockage of G0-G1/S transition of NB cells caused by altered expression of
CDK4 and cyclin D1.
Apoptosis is critical for successful organism development and the maintenance of tis-
sue homeostasis [24]. It is clear that genetic alteration of the oncogene could destroy
the process of apoptosis and affect tumorigenesis and development [22]. Increasing evi-
dence shows that disruption of CDKL1 is implicated in cancer cell apoptosis, including
gastric cancer [16] and melanoma [18]. Sun et al. [16] revealed that downregulation of
CDKL1 triggers activation of pro-apoptotic protein BIK (BCL2-interacting killer). Fuchs
Fig. 4 The effect of CDKL1 knockdown on cell migration and invasion of NB cells in vitro. a Wound-healingassays were performed to investigate the migratory ability of SH-SY5Y cells. b Representative images areshown for siCDKL1-transfected SH-SY5Y cells in a transwell assay. The invasion was quantified by countingthe cells in 3 random microscopic fields. Data are presented as means ± SD. **P < 0.01, ***P < 0.001 vs. NC
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 8 of 11
et al. [25] found that knockdown of CDKL5 was implicated in promoting the cell death
of post-mitotic granule neurons though alteration of the AKT/GSK-3β pathway.
Caspase-3 is clearly a critical proteolytic enzyme that act as a key effector in apoptosis.
It rapidly undergoes enzymatic cleavage and thus activation when triggered by the
apoptotic cascade [26, 27].
Here, we observed that knockdown of CDKL1 promoted apoptosis and elevated the
levels of the pro-apoptotic factor caspase-3 in SH-SY5Y cells. The profiles of the
cleaved PARP, a well-known substrate of caspase-3 and a good marker for apoptosis
[28, 29], were also upregulated after knockdown of CDKL1. Therefore, we suggest that
depletion of CDKL1 induced the apoptotic cascade in SH-SY5Y cells, and this resulted
in apoptosis.
Accumulating evidence indicates that there is a close relationship between CDKL1
and metastasis in a variety of tumors. For instance, knockdown of CDKL1 significantly
suppressed cell migration and invasion in colorectal cancer [19] and breast cancer [17].
As the hallmark of epithelial–mesenchymal transition, the downregulation of
E-cadherin is a leading event in the progression of various tumors into the metastatic
cascade [30, 31]. Vimentin, an intermediate filament of epithelial–mesenchymal transi-
tion, has been shown to facilitate mesenchymal cell migration [32]. In this study, we
found that CDKL1 knockdown significantly upregulated E-cadherin expression but
downregulated vimentin expression in NB cells. This might be the main cause of the
suppressive effects of CDKL1 knockdown on cell migration and invasion.
Certain limitations of this study must be assessed in further research. These include
experiments with CDKL1 overexpression and in vivo animal experiments.
ConclusionsOur results show that CDKL1 silencing reduced the proliferation, colony formation,
migration and invasion of NB SH-SY5Y cells. Our study proves that elevated CDKL1
expression might contribute to NB progression. The underlying mechanisms of CDKL1
activity might be a potential target for therapeutic intervention.
Fig. 5 Effect of CDKL1 knockdown on the regulation of cell cycle, apoptotic and invasion in SH-SY5Y cells,as confirmed using western blot analysis. GAPDH was used as the internal control. Data are presented asmeans ± SD. **p < 0.01, ***p < 0.001 vs. NC
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 9 of 11

AbbreviationsATCC: American Type Culture Collection; CDKL1: cyclin-dependent kinase-like 1; DMEM: Dulbecco’s modified Eagle’smedium; FBS: fetal bovine serum; NB: neuroblastoma
AcknowledgementsThis study was supported by the China Academy of Chinese Medical Sciences in Beijing.
FundingThis study was funded by the Basic Research on the Application of Independent Innovation Plan in Qingdao (No. 15–9–1-59-jch) and Sanming Project of Medicine in Shenzhen (SZSM201812090).
Availability of data and materialsAll data generated or analyzed during this study are included in this published article.
Authors’ contributionsLWY, as the guarantor of integrity of the entire study, designed the study. LWY, LJ, CWL and ZCQ performed theexperiment. CJ and LJ collected and analyzed the data. CJ, LJ, CWL, ZCQ, CSL and KZF were involved in literatureresearch and manuscript preparation. KZF critically reviewed the manuscript. All authors read and approved the finalmanuscript.
Ethics approval and consent to participateAll the clinical specimens were collected in accordance to the Declaration of Helsinki. This study was approved by theResearch Ethics Committee of the Eye Hospital of the China Academy of Chinese Medical Sciences (EHCAC-20170635,date: 2017-6-12).
Consent for publicationNot applicable.
Competing interestsThe authors declare that they have no competing interests.
Publisher’s NoteSpringer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author details1Eye Hospital, China Academy of Chinese Medical Sciences, No 33 Lugu Road, Shijingshan district, Beijing 100040,China. 2Yinan Branch of Qilu Hospital of Shandong University, Linyi, Shandong, China.
Received: 2 September 2018 Accepted: 11 February 2019
References1. Cheung NK, Dyer MA. Neuroblastoma: developmental biology, cancer genomics and immunotherapy. Nat Rev Cancer.
2013;13:397–411. https://doi.org/10.1038/nrc3526.2. Tolbert VP, Coggins GE, Maris JM. Genetic susceptibility to neuroblastoma. Curr Opin Genet Dev. 2017;42:81–90. https://
doi.org/10.1016/j.gde.2017.03.008.3. van Golen CM, Schwab TS, Kim B, Soules ME, Su Oh S, Fung K, van Golen KL, Feldman EL. Insulin-like growth factor-I
receptor expression regulates neuroblastoma metastasis to bone. Cancer Res. 2006;66:6570–8. https://doi.org/10.1158/0008-5472.CAN-05-1448.
4. Sausen M, Leary RJ, Jones S, Wu J, Reynolds CP, Liu X, Blackford A, Parmigiani G, Diaz LA Jr, Papadopoulos N, et al.Integrated genomic analyses identify ARID1A and ARID1B alterations in the childhood cancer neuroblastoma. NatGenet. 2013;45:12–7. https://doi.org/10.1038/ng.2493.
5. Mohammadi M, Goodarzi M, Jaafari MR, Mirzaei HR, Mirzaei H. Circulating microRNA: a new candidate for diagnosticbiomarker in neuroblastoma. Cancer Gene Ther. 2016;23:371–2. https://doi.org/10.1038/cgt.2016.45.
6. Amente S, Milazzo G, Sorrentino MC, Ambrosio S, Di Palo G, Lania L, Perini G, Majello B. Lysine-specific demethylase(LSD1/KDM1A) and MYCN cooperatively repress tumor suppressor genes in neuroblastoma. Oncotarget. 2015;6:14572–83. https://doi.org/10.18632/oncotarget.3990.
7. Durinck K, Speleman F. Epigenetic regulation of neuroblastoma development. Cell Tissue Res. 2018. https://doi.org/10.1007/s00441-017-2773-y.
8. Aouadi M, Binetruy B, Caron L, Le Marchand-Brustel Y, Bost F. Role of MAPKs in development and differentiation: lessonsfrom knockout mice. Biochimie. 2006;88:1091–8. https://doi.org/10.1016/j.biochi.2006.06.003.
9. Deacon EM, Pettitt TR, Webb P, Cross T, Chahal H, Wakelam MJ, Lord JM. Generation of diacylglycerol molecular speciesthrough the cell cycle: a role for 1-stearoyl, 2-arachidonyl glycerol in the activation of nuclear protein kinase C-betaII atG2/M. J Cell Sci. 2002;115:983–9.
10. Morgan DO. Principles of CDK regulation. Nature. 1995;374:131–4. https://doi.org/10.1038/374131a0.11. Wang Q, Ma J, Lu Y, Zhang S, Huang J, Chen J, Bei JX, Yang K, Wu G, Huang K, et al. CDK20 interacts with KEAP1 to
activate NRF2 and promotes radiochemoresistance in lung cancer cells. Oncogene. 2017;36:5321–30. https://doi.org/10.1038/onc.2017.161.
12. Porter LA, Donoghue DJ. Cyclin B1 and CDK1: nuclear localization and upstream regulators. Prog Cell Cycle Res. 2003;5:335–47.
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 10 of 11

13. Yeh CW, Kao SH, Cheng YC, Hsu LS. Knockdown of cyclin-dependent kinase 10 (cdk10) gene impairs neural progenitorsurvival via modulation of raf1a gene expression. J Biol Chem. 2013;288:27927–39. https://doi.org/10.1074/jbc.M112.420265.
14. Taglienti CA, Wysk M, Davis RJ. Molecular cloning of the epidermal growth factor-stimulated protein kinase p56KKIAMRE. Oncogene. 1996;13:2563–74.
15. Hsu LS, Liang CJ, Tseng CY, Yeh CW, Tsai JN. Zebrafish cyclin-dependent protein kinase-like 1 (zcdkl1): identification andfunctional characterization. Int J Mol Sci. 2011;12:3606–17. https://doi.org/10.3390/ijms12063606.
16. Sun W, Yao L, Jiang B, Shao H, Zhao Y, Wang Q. A role for Cdkl1 in the development of gastric cancer. Acta Oncol.2012;51:790–6. https://doi.org/10.3109/0284186X.2012.665611.
17. Tang L, Gao Y, Yan F, Tang J. Evaluation of cyclin-dependent kinase-like 1 expression in breast cancer tissues and itsregulation in cancer cell growth. Cancer Biother Radiopharm. 2012;27:392–8. https://doi.org/10.1089/cbr.2012.1198.
18. Song Z, Lin J, Sun Z, Ni J, Sha Y. RNAi-mediated downregulation of CDKL1 inhibits growth and colony-formation ability,promotes apoptosis of human melanoma cells. J Dermatol Sci. 2015;79:57–63. https://doi.org/10.1016/j.jdermsci.2015.03.020.
19. Qin C, Ren L, Ji M, Lv S, Wei Y, Zhu D, Lin Q, Xu P, Chang W, Xu J. CDKL1 promotes tumor proliferation and invasion incolorectal cancer. Onco Targets Ther. 2017;10:1613–24. https://doi.org/10.2147/OTT.S133014.
20. Yen SH, Kenessey A, Lee SC, Dickson DW. The distribution and biochemical properties of a Cdc2-related kinase, KKIALREin normal and Alzheimer brains. J Neurochem. 1995;65:2577–84.
21. Canning P, Park K, Goncalves J, Li C, Howard CJ, Sharpe TD, Holt LJ, Pelletier L, Bullock AN, Leroux MR. CDKL familykinases have evolved distinct structural features and ciliary function. Cell Rep. 2018;22:885–94. https://doi.org/10.1016/j.celrep.2017.12.083.
22. Evan GI, Vousden KH. Proliferation, cell cycle and apoptosis in cancer. Nature. 2001;411:342–8. https://doi.org/10.1038/35077213.
23. Wainberg ZA, Yufa A, Anghel A, Rogers AM, Manivong T, Adhami S, Hamidi H, Conklin D, Finn RS, Slamon DJ. Abstract4557: expression of p16 in colon cancer and cyclin D1 in gastric cancer predicts response to CDK4/6 inhibition in vitro.Cancer Res. 2014;74:4557.
24. Henson PM, Hume DA. Apoptotic cell removal in development and tissue homeostasis. Trends Immunol. 2006;27:244–50. https://doi.org/10.1016/j.it.2006.03.005.
25. Fuchs C, Trazzi S, Torricella R, Viggiano R, De Franceschi M, Amendola E, Gross C, Calza L, Bartesaghi R, Ciani E. Loss ofCDKL5 impairs survival and dendritic growth of newborn neurons by altering AKT/GSK-3beta signaling. Neurobiol Dis.2014;70:53–68. https://doi.org/10.1016/j.nbd.2014.06.006.
26. Buschhaus JM, Gibbons AE, Luker KE, Luker GD: Fluorescence lifetime imaging of a caspase GD: D: ranceschi M, A JohnWiley & Sons, Inc.; 2017.
27. Lossi L, Cocito C, Alasia S, Merighi A. Ex vivo imaging of active caspase 3 by a FRET-based molecular probedemonstrates the cellular dynamics and localization of the protease in cerebellar granule cells and its regulation by theapoptosis-inhibiting protein survivin. Mol Neurodegener. 2016;11:34. https://doi.org/10.1186/s13024-016-0101-8.
28. Bressenot A, Marchal S, Bezdetnaya L, Garrier J, Guillemin F, Plenat F. Assessment of apoptosis byimmunohistochemistry to active caspase-3, active caspase-7, or cleaved PARP in monolayer cells and spheroid andsubcutaneous xenografts of human carcinoma. J Histochem Cytochem. 2009;57:289–300. https://doi.org/10.1369/jhc.2008.952044.
29. Miyashita T, Okamura-Oho Y, Mito Y, Nagafuchi S, Yamada M. Dentatorubral pallidoluysian atrophy (DRPLA) protein iscleaved by caspase-3 during apoptosis. J Biol Chem. 1997;272:29238.
30. Cheng CW, Wu PE, Yu JC, Huang CS, Yue CT, Wu CW, Shen CY. Mechanisms of inactivation of E-cadherin in breastcarcinoma: modification of the two-hit hypothesis of tumor suppressor gene. Oncogene. 2001;20:3814–23. https://doi.org/10.1038/sj.onc.1204505.
31. Christofori G, Semb H. The role of the cell-adhesion molecule E-cadherin as a tumour-suppressor gene. Trends BiochemSci. 1999;24:73–6.
32. Schoumacher M, Goldman RD, Louvard D, Vignjevic DM. Actin, microtubules, and vimentin intermediate filamentscooperate for elongation of invadopodia. J Cell Biol. 2010;189:541–56. https://doi.org/10.1083/jcb.200909113.
Li et al. Cellular & Molecular Biology Letters (2019) 24:19 Page 11 of 11







