
JOURNAL OF BACTERIOLOGY, Nov. 2006, p. 7742–7758 Vol. 188, No. 220021-9193/06/$08.00�0 doi:10.1128/JB.00555-06Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Influence of the Two-Component System SaeRS on Global GeneExpression in Two Different Staphylococcus aureus Strains†
Kathrin Rogasch,1 Vanessa Ruhmling,1 Jan Pane-Farre,1 Dirk Hoper,1 Christin Weinberg,1Stephan Fuchs,1 Mareike Schmudde,2 Barbara M. Broker,2 Christiane Wolz,3
Michael Hecker,1 and Susanne Engelmann1*Institut fur Mikrobiologie, Ernst-Moritz-Arndt-Universitat, D-17487 Greifswald, Germany1; Institut fur Immunologie und
Transfusionsmedizin, Ernst-Moritz-Arndt-Universitat, D-17487 Greifswald, Germany2; and Institut furMedizinische Mikrobiologie und Hygiene, Universitat Tubingen, D-72074 Tubingen, Germany3
Received 19 April 2006/Accepted 2 August 2006
The two-component system SaeRS consisting of the histidin kinase SaeS and the response regulator SaeRis known to act on virulence gene expression in Staphylococcus aureus. In order to get a more comprehensivepicture on SaeR-regulated genes, we studied the contribution of the two-component system on global geneexpression by using both the proteomic and transcriptomic approach. Altogether, a loss of SaeRS resulted ina decreased amount of at least 17 extracellular proteins and two cell surface-associated proteins, among themseveral important virulence factors such as HlgA, HlgB, HlgC, LukF, and LukM. SaeRS activates the expres-sion of these genes at the transcriptional level. The amount of the five proteins Aur, SspA, SsaA, Plc, and GlpQwas negatively influenced by SaeRS. However, the transcription of the corresponding genes was not affected bythe two-component system. SaeRS had also no measurable influence on the transcription of the regulatorygenes agr, sarA, arlRS, and sigB that contribute to the regulation of SaeRS-dependent virulence factorsidentified in this investigation. Our results clearly show that SaeRS is strongly involved in the tight temporalcontrol of virulence factor expression in S. aureus. Its precise role within the regulatory network remains to bedetermined.
Staphylococcus aureus is a gram-positive bacterium that col-onizes the anterior nares of at least one-third of the humanpopulation but also causes a variety of infections ranging fromsuperficial lesions, such as wound infections and abscesses, tosevere systemic infections such as bacteremia, endocarditis,pneumonia, and osteomyelitis. The pathogenicity of this or-ganism largely depends on the successful adaptation to thehuman host and the environmentally coordinated expression ofvirulence factors. The expression of virulence factors in S.aureus is regulated during the growth cycle by a network ofinteracting regulators (for a review, see reference 41). Thebest-characterized virulence-associated regulons thus far arethe agr regulon (accessory gene regulator), the SarA regulon(staphylococcal accessory regulator), the �B regulon (alterna-tive sigma factor), the Rot regulon (regulator of toxins), andthe ArlRS regulon (autolysis-regulated locus) (7, 15, 20, 37, 47,60, 61).
The sae locus was first described by Giraudo et al. (27)following the characterization of a Tn551 insertional mutant ofS. aureus RC161. sae is a regulatory locus that consists of fouropen reading frames, two of them encode the response regu-lator and the sensor kinase, respectively (23). Two additionalopen reading frames coding for hypothetical proteins are prob-ably important for the functionality of the sae operon (42, 56).
The two-component system SaeRS itself activates the expres-sion of several virulence factors such as serine protease SspA,thermonuclease Nuc, coagulase Coa, alpha-hemolysin Hla, be-ta-hemolysin Hlb, extracellular adherence protein Eap, extra-cellular matrix binding protein Emp, protein A, and fibronec-tin binding protein FnbA (24, 27–30, 42). In contrast, theexpression of the cap operon is repressed by SaeRS (56). Thetranscription of the sae operon is influenced by environmentalsignals such as pH, salt, and glucose concentrations or subin-hibitory concentrations of antibiotics (42, 35). Moreover, thetranscription of sae is controlled by other virulence-associatedregulators (24, 42). It has been shown that agr might influencethe transcription of sae, which would explain the concomitantinfluence of both regulators on the synthesis of extracellularproteins (25, 28, 42). In contrast, an sae mutation has no effecton the expression of agr and sarA (24). Therefore, it has beensuggested that SaeRS regulates the synthesis of extracellularproteins downstream of agr and might modify quorum sensing-dependent regulation by sensing of additional signals. Whetheror not SaeR interacts with other regulators is largely unknown.
Furthermore, there are data indicating that the sae locus isessential for virulence gene expression under in vivo conditions(28, 29, 45). Consequently, the virulence of the sae mutant,assayed by the intraperitoneal injection in mice, was signifi-cantly lower than that of the parental strain (45). The impor-tance of SaeRS for the virulence of S. aureus was also empha-sized in two whole-genome screens for the identification ofgenes required for full virulence (1, 4).
To date, only a few genes are known to be regulated by thetwo-component system. Therefore, we used transcriptomic andproteomic approaches to define the SaeRS regulon structure in
* Corresponding author. Mailing address: Institut fur Mikrobiolo-gie, Ernst-Moritz-Arndt-Universitat, F.-L.-Jahn-Str. 15, D-17487,Greifswald, Germany. Phone: 49-3834-864227. Fax: 49-3834-864202.E-mail: [email protected].
† Supplemental material for this article may be found at http://jb.asm.org/.
7742
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

two different S. aureus strains: COL and Newman. Transcrip-tome analyses with a full genome DNA array of S. aureusprovided information on the role of SaeR as a global transcrip-tional regulator, whereas the proteomic approach allows theinvestigation of the influence of SaeRS on the amount ofproteins within and outside the cells. Since the extracellularproteome of S. aureus represents a reservoir of virulence fac-tors, we focused especially on this subproteome.
MATERIALS AND METHODS
Bacterial strains and culture conditions. The bacterial strains used in thepresent study are listed in Table 1. The mutated saeS gene of NCTC8325-4saeS::Tn917 (29) was transduced into the wild-type strain COL by using phage 85(5), resulting in an isogenic saeS mutant strain. The insertion event was con-firmed by PCR using oligonucleotides specific for saeS and Tn917 (saeS andtn917) (Table 2). Bacterial cultures were inoculated with an overnight culture toan optical density at 540 nm (OD540) of 0.05 into tryptic soy broth (TSB),followed by incubation with agitation at 37°C. For all cultures, a flask-to-mediumratio of 5:1 was used. Bacterial growth was monitored by measuring the OD540.
Preparation of protein extracts. For the preparation of extracellular proteinextracts, bacteria were grown in TSB. At different optical densities (OD540), theextracellular proteins from 100 ml of supernatant were precipitated, washed,dried, and resolved as described previously (60).
Cytoplasmic proteins were prepared from bacteria grown in TSB mediumto different optical densities (OD540s of 1, 6, and 10). In each case, cells from50 ml of culture were used to isolate cytoplasmic proteins as described byKohler et al. (34).
The protein concentration was determined by using Roti-Nanoquant accord-ing to the manufacturer’s instructions (Carl Roth GmbH & Co., Karlsruhe,Germany).
Analytic and preparative 2D-PAGE. Preparative two-dimensional polyacryl-amide gel electrophoresis (2D-PAGE) was performed by using the immobilizedpH gradient (IPG) technique described by Bernhardt et al. (6). The proteinsamples were separated on 2D gels using linear IPG strips (GE-Healthcare,Little Chalfont, United Kingdom) in the pH range of 3 to 10. The resultingprotein spots were stained with silver nitrate or with colloidal Coomassie brilliantblue G-250 (8, 10). For protein identification by matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF-MS), 350-�g portionsof protein extracts were separated on 2D gels, and the proteins were stained withcolloidal Coomassie brilliant blue (10). Coomassie blue-stained protein spotswere cut from gels by using the spot cutter Proteome Work (GE-Healthcare)with a picker head of 2 mm and transferred into 96-well microtiter plates.Digestion with trypsin and subsequent spotting of the peptide solutions onto theMALDI targets were performed automatically in the Ettan Spot Handling Work-station (GE-Healthcare) using a protocol described by Eymann et al. (18).MALDI-TOF-MS analyses of spotted peptide solutions were carried out on aProteome-Analyzer 4700 (Applied Biosystems, Foster City, CA). The spectrawere recorded in reflector mode in a mass range from 900 to 3,700 Da with afocus mass of 2,000 Da. For one main spectrum, 25 subspectra with 100 spots persubspectrum were accumulated by using a random search pattern. Automatic ormanual calibration was performed as described by Eymann et al. (18). Aftercalibration, the peak lists were created by using the “peak-to-mascot” script ofthe 4700 Explorer software. The resulting peak lists were analyzed by using themascot search engine (Matrix Science, London, United Kingdom) and the ge-nome sequence of S. aureus COL (22) and Mu50 (36).
Quantitation of protein spots. For quantitation of extracellular proteins, theEttan-fluorescence difference gel electrophoresis (DIGE) technique was used(GE-Healthcare). Protein extracts were labeled with CyDye DIGE Cy2, Cy3, orCy5 (GE-Healthcare) prior to separation on 2D gels as described by Ziebandt et
al. (60). Cy2-, Cy3-, and Cy5-labeled proteins were detected by using a Typhoonlaser scanner 9400 (GE-Healthcare). The unfixed gels were scanned according tothe Ettan DIGE user manual (GE-Healthcare) with 254-dpi resolution. Theresulting images were compared, and spots were quantified by using Delta2DSoftware from Decodon GmbH (Greifswald, Germany). Only volume ratios of�2 or �0.5 and a probability value � of �5% were defined as significant changesbetween the different strains.
Transcriptional analyses. Total RNA from S. aureus was isolated by using theacid-phenol method (21). Digoxigenin-labeled RNA probes were prepared by invitro transcription with T7 RNA polymerase by using the Dig-RNA labelingmixture (Roche, Indianapolis, IN) and appropriate PCR fragments as templates.The PCR fragments were generated by using chromosomal DNA of S. aureusCOL isolated with the chromosomal DNA isolation kit (Promega, Madison, WI)according to the manufacturer’s recommendations and the respective oligonu-cleotides (Table 2). Reverse primers contain the T7 RNA polymerase recogni-tion sequence at the 5� end (33). Northern blot analyses were carried out aspreviously described (57). Before hybridization, each RNA blot was stained withmethylene blue in order to check RNA loading and blotting. Only blots showingequal amounts of 23S rRNA and 16S rRNA for each sample loaded onto therespective gel were used for hybridization experiments. The digoxigenin-labeledRNA marker I (Roche) was used to calculate the sizes of the transcripts. Thehybridization signals were detected by using a Lumi-Imager (Roche) and ana-lyzed by using the software package LumiAnalyst (Roche).
For DNA microarray analyses, sciTRACER S. aureus N315 full genome mi-croarrays (Scienion, Berlin, Germany) containing PCR products correspondingto 2,334 genes derived from the genome sequence of S. aureus N315 were used.The integrity of RNA was ensured by gel electrophoresis and by analysis with theAgilent 2100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany). Fluo-rescent probes were prepared by reverse transcription of 10 �g of total RNAfrom S. aureus COL, S. aureus Newman, and their isogenic saeS mutants. Syn-thesis, purification, and hybridization of fluorescence-labeled cDNAs, as well asthe washing of slides, were carried out as recommended by the manufacturer(Scienion). Each slide was hybridized competitively with cDNAs of the wild typeand its isogenic saeS mutant labeled with Cy3 and Cy5, respectively.
Slides were scanned by using a ScanArray scanner (PE Biosystems, Weiter-stadt, Germany), and the obtained images were quantified with ScanArrayExpresssoftware (version 3.0; PE Biosystems) using “adaptive threshold” as the quanti-tation method. All subsequent calculations were performed using R2.2.1 (44)and Limma 2.4.7 (52, 53). Prior to all further analysis steps, the reproducibilityof the data was determined as follows. The raw spot intensities from the twoidentical adjacent probes (designated by 1 and 2 in the following formulae) werecompared by calculating the log2(G1/G2) in the green channel and log2(R1/R2)in the red channel, respectively. Furthermore, the reproducibility of the ratiobetween the red and green channel intensities of the spot pairs was compared bycalculating the log2[(R1/G1)/(R2/G2)]. For spot pairs with a good reproducibil-ity, all of these values tend toward 0. From the lists of the red, green, andred-green reproducibility scores, respectively, all extremes were removed array-wise by iterative calculation and removal of the outliers according to Chauvenet’scriterion (11) until no further outliers could be detected. The methods imple-mented in limma allow for weighting the impact of every datum on the calcula-tions for normalization and linear modeling by assigning every spot intensity aspotweight. Spotweights for calculations with limma functions were attributed bygiving weights 0 to all spots that were determined to be outliers with respect toat least one of the reproducibility criteria. Furthermore, all control spots wereassigned weight 0 for the subsequent calculations. Moreover, all signals corre-sponding to genes not encoded in the S. aureus COL genome as revealed by anucleotide/nucleotide BLAST (E-value cutoff of 10e-50) with the S. aureus N315open reading frames against the COL genomic sequence were omitted from theCOL/COL�sae data set by assigning a spotweight of 0. The most stringent upperand lower limits of all single arrays were used as the overall spot reproducibilitythreshold for the whole data set. All spots failing these defined limits were given
TABLE 1. Strains used in this study
Strain Genotype Relevant phenotype Source or reference
S. aureus NCTC8325-4 �saeS saeS::Tn917, isogenic to strain NCTC8325-4 SaeR� SaeS� 29S. aureus COL MRSA 50S. aureus COL �saeS saeS::Tn917, isogenic to strain COL MRSA, SaeR� SaeS� This studyS. aureus Newman 16S. aureus Newman �sae saeS::Tn917, isogenic to strain Newman SaeR� SaeS� 29
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7743
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

a spotweight of 0.1. The remaining spots meeting all three reproducibility criteriawere assigned a spotweight of 1. The overall spot reproducibility measures areshown in Table S1 in the supplemental material. Table S2 in the supplementalmaterial displays the percentage of spots meeting the different criteria (not
included in this percentages are empty and control spots). Before a linear modelwas calculated, the data were normalized within arrays by using the Loessfunction (13) and subsequently normalized between arrays using the methodAquantile (55). Simultaneously with within-array normalization, the backgroundcorrection as proposed by Edwards (17) and implemented in limma (55) wasperformed. A linear model was calculated from the normalized data according tothe guidelines of the limma users guide (17 December 2005 version). The linearmodel fitted with the limma functions accounts for dye effects and considers theduplicate spots as technical replicates (54). All spots with a P value of �0.05 andinduced or repressed at least 2.5-fold were considered to be significantly regu-lated.
RESULTS
Differential transcription of the sae-operon during growth inTSB medium in S. aureus COL and S. aureus Newman. North-ern blot experiments using an saeR-specific RNA probe re-vealed three transcripts of about 3.0, 2.4, and 2.0 kb. As alreadypublished by Steinhuber et al. (56), saeR and saeS are cotrans-cribed with orf3 and orf4 upstream of saeR (Fig. 1). Whereasthe 3.0-kb transcript comprises all four open reading frames,the 2.4- and 2.0-kb transcripts are initiated from internal pro-moters in front of orf3 and saeR, respectively (56). All threetranscripts were differentially expressed during growth. In S.aureus COL, sae transcription was almost nondetectable at lowoptical densities but strongly increased at an OD540 of 4 (Fig.1). In contrast, in exponentially growing cells of S. aureusNewman, the transcriptional level of sae was significantlyhigher than in S. aureus COL. Only a very slight increase of saetranscription at high optical densities could be observed instrain Newman. Very similar observations were done by othergroups comparing the expression of sae in S. aureus Newmanwith that of either SH1000 or 8325-4 (30). Therefore, both S.aureus Newman and S. aureus COL were used to study SaeRS-dependent gene expression in S. aureus. In both strains the2.4-kb transcript starting in front of orf3 was the most abundanttranscript (Fig. 1). When RNA of the saeS mutant strains wasused, no saeR-specific RNA could be detected (data notshown; see also reference 56). These results strongly imply theloss of both the sensor kinase SaeS and the response regulatorSaeR in the saeS mutant strains used in the present study.
Extracellular proteome of S. aureus COL and S. aureus New-man. The differences between the extracellular protein pat-terns of strain COL and strain Newman were striking (Fig. 2).In all, 42 proteins were identified from S. aureus COL and 47were identified from S. aureus Newman (Table 3). The proteinsof both strains were assigned to the open reading frame num-bers defined in the S. aureus COL genome sequencing project(22) or in the Mu50 and N315 genome sequencing project (36).Superantigen proteins were named according to the nomen-clature suggested by Lina et al. (38). At least 49 of the iden-tified proteins both in COL and Newman showed signal se-quences typical for Sec-translocated proteins (Table 3). Theremaining proteins were of cytoplasmic origin. The presence ofthese proteins in the supernatant might be due to cell lysis.
A comparison of the list of potentially Sec-translocated pro-teins identified in S. aureus COL with those identified in S.aureus Newman showed an overlap of 21 proteins (Table 3).Among these are the metalloprotease Aur, the cysteine pro-teases SspA and SspB, and the serine protease SplA. Further-more, we found the gamma-hemolysin components HlgA,HlgB, HlgC, LukF, LukE, and LukM; the alpha-hemolysin
TABLE 2. Oligonucleotides used in this study
Oligonucleotide Sequence (5�–3�)a
agrAfor.....................TTCATTTGCGAAGACGATCCagrArevT7................CTAATACGACTCACTATAGGGAGATTCTC
ACCGATGCATAGCAGarlRfor......................GGTTTAGATAAAGCGCTTAGarlRrevT7.................CTAATACGACTCACTATAGGGAGACGCC
ACGAACTGTTTCAATCaurfor........................CACCAGCAGCATTAGCGATTaurrevT7...................CTAATACGACTCACTATAGGGAGATCTA
GGCTAAATCCACCGTCcoafor........................GCGAGACAAGATTCAATAAGcoarevT7...................CTAATACGACTCACTATAGGGAGACGCA
GTACCATCTGCATGTGearfor ........................GCAAGTGCTTTAGTTTTAACearrevT7 ...................TAATACGACTCACTATAGGGAGA- CCAT
TATAGTTTTCACCAACefbfor ........................GCGGCAATAGGTATTACTACefbrevT7 ...................CTAATACGACTCACTATAGGGAGAGCTT
TTCTGTGTCACTGACfnbAfor.....................TGCAAATACGACAGATACTTfnbArevT7................CTAATACGACTCACTATAGGGAGATTGG
CCACCTTCATAACCTAfnbBfor .....................GGTCAAGTTATGGCGACAGGfnbBrevT7 ................CTAATACGACTCACTATAGGGAGACGAT
TGCTCCTTGCGCTTGAglpQfor .....................GCTGCTTCTGCTGTTTTTACglpQrevT7 ................CTAATACGACTCACTATAGGGAGACAAT
CGCATAAGAGCGTATChlYfor .......................AACACGTATAGTCAGCTCAGhlYrevT7 ..................CTAATACGACTCACTATAGGGAGAAACT
GTAGCGAAGTCTGGTGmapfor ......................GCAGTAAATGGCACATCAACmaprevT7 .................CTAATACGACTCACTATAGGGAGAAAGT
CAAGATCACTGATGCCplcfor ........................GGTTCACATGATAGTGGCTCplcrevT7 ...................CTAATACGACTCACTATAGGGAGACTAT
TAAATGCGCTGCCTCCRNAIIIfor................AGGAAGGAGTGATTTCAATGRNAIIIrevT7...........CTAATACGACTCACTATAGGGAGAACTC
ATCCCTTCTTCATTACsaeRfor.....................CTTAACTGATCGTGGATGATGsaeRrevT7................CTAATACGACTCACTATAGGGAGAGTGT
ATATGGACATTCACGGsaeS...........................ATCATGGGCTAAGTTTTGAATCsarAfor .....................TAGGGAGGTTTTAAACATGGsarArevT7 ................CTAATACGACTCACTATAGGGAGAGTTG
TTTGCTTCAGTGATTCsebfor........................GCAGAGAGTCAACCAGATCCsebrevT7...................CTAATACGACTCACTATAGGGAGATCTC
CTGGTGCAGGCATCATsigBfor ......................AAATAATGGCGAAAGAGTCGsigBrevT7 .................CTAATACGACTCACTATAGGGAGACATA
ATGGTCATCTTGTTGCsplFfor ......................GCAGCATTGACGATTTTAACsplFrevT7 .................TAATACGACTCACTATAGGGAGACAGGA
GAGAAATAAACAGCAssaAfor .....................GCTCATGCTTCTGAGCAAGAssaArevT7 ................CTAATACGACTCACTATAGGGAGACTGG
GCCATAACCATAGTTCsspBfor .....................CAAAGCCGATTCACACTCTAsspBrevT7.................CTAATACGACTCACTATAGGGAGAGATC
TTCTTGTATCGCTTCGtn917.........................ACGCAAGACCAATCACTCTC
a The T7 promoter sequences are underlined.
7744 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

HlY; the lipases Lip, GlpQ, and Plc; the thermonuclease Nuc;and proteins such as Aly, IsaA, Sbi, and SsaA. Although all ofthese proteins were detectable in both strains, some of themdiffered significantly in amount. For instance, Nuc, Sbi, LukF,LukE, LukM, HlgB, HlgC, and HlgA were present at a higherlevel in strain Newman, whereas the amounts of SspA, Aur,and SplABCF were increased in strain COL.
Twelve Sec-translocated proteins were unique for S. aureusCOL, and 16 were only detectable in S. aureus Newman (Table3 and Fig. 2). There are two possible explanations for this: (i)the proteins are not encoded in the genome of S. aureus COLor Newman, respectively, or (ii) the proteins are synthesized invery low amounts and thus remained below the detection limit.For S. aureus COL, it was possible to discriminate betweenthese two possibilities, since the genome sequence is available.Surprisingly, only the genes for SEA and Ssl7 were missing inS. aureus COL. The reason for the absence of a particularprotein in the S. aureus Newman strain is not as easily deter-
mined because its genome has not been sequenced. However,by using the PCR technique we could demonstrate the pres-ence of the gene coding for Hlb in S. aureus Newman. Theabsence of Hlb on 2D gels of S. aureus Newman might be dueto the presence of an hlb-converting bacteriophage. This no-tion is supported by the observation that the phage-encodedSEA was detectable in strain Newman (Table 3) (14). Genescoding for SEB, SEI (also SElQ), SEK, and Ear could not bedetected in strain Newman (data not shown). Since all of thesegenes are localized on pathogenicity island vSa1, this geneticelement might be absent in strain Newman. Finally, Pls wasfound only in S. aureus COL. The protein is known to beencoded on mec cassette type I (32), which is missing in strainNewman.
Influence of a mutation in saeS on the amount of extracel-lular proteins in S. aureus COL and S. aureus Newman. Thedifferential proteome display can also be used to visualize theentire set of extracellular proteins in the wild type in compar-
FIG. 1. Transcriptional analysis of the sae operon. (A) Northern blot analysis of the sae operon. RNA was isolated from S. aureus COL andS. aureus Newman grown in TSB medium at 37°C. The membrane was hybridized with digoxigenin-labeled RNA probes specific for saeR. Relevanttranscripts are indicated by arrows. (B) Methylene blue-stained blots are shown as indication for equal RNA loading. (C) Growth curve of S. aureusCOL and Newman. The sampling points are indicated in the growth curve by arrows and optical density numbers. (D) Schematic representationsof the gene loci based on the sequence of S. aureus COL and the transcriptional organization of the predicted operons. The dotted line representsthe RNA probe used in the experiments.
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7745
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

ison with a regulatory mutant. In order to quantify changes inthe level of extracellular proteins, we used the Ettan-fluores-cence DIGE technique (GE-Healthcare). To investigate theinfluence of a mutation in saeS on the extracellular proteomeof S. aureus COL, the extracellular protein pattern at OD540sof 6 and 10 of the respective wild-type strain was compared tothose of the isogenic saeS mutant (Fig. 3A). Eight proteinswere found in higher amounts in the wild-type strain than inthe saeS mutant (Fig. 3A and Table 4), among them the he-molysins Hlb and HlY, the thermonuclease Nuc, the leucocidincomponent LukF, the enterotoxin SEB, and the serine pro-teases SplA and SplC, which are described to play a role asvirulence factors. Furthermore, the amount of the pathogenic-ity island protein Ear was also positively influenced by SaeRS.Enterotoxin B, alpha-hemolysin, and beta-hemolysin areamong the most abundant proteins.
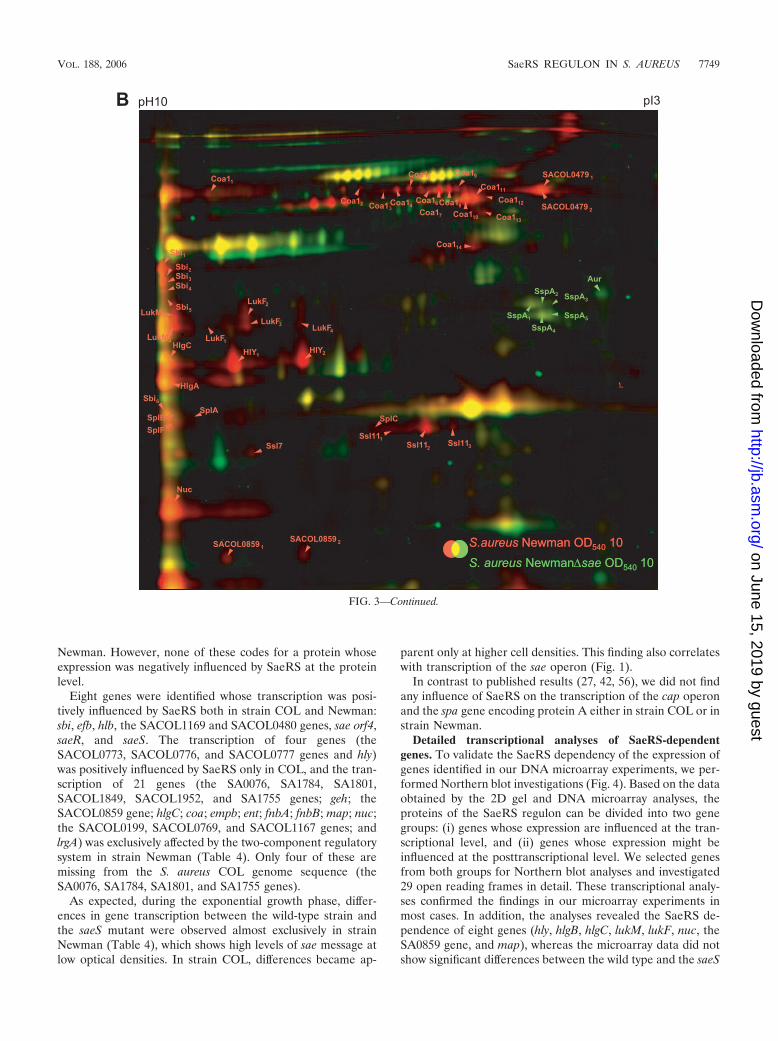
Because of the aforementioned high level of sae-specificmRNA in S. aureus Newman, the influence of the two-compo-nent system on the expression of extracellular proteins was alsoanalyzed in this strain. Here we could show that the amount ofat least 15 proteins was decreased by a mutation in saeS (Fig.3B and Table 4). Five of these proteins (HlY, LukF, SplA,SplC, and Nuc) were positively influenced by SaeRS also in S.aureus COL. The other 10 SaeRS dependently expressed pro-
teins (Coa, SACOL0479, SACOL0859, Sbi, HlgA, HlgB, HlgC,LukM, Ssl11, and Ssl7) could be identified to be influenced bySaeRS only in strain Newman. Three proteins (Hlb, SEB, andEar) were influenced by SaeRS in strain COL but not encodedin strain Newman (Table 4).
Besides the positive effect of SaeRS on the amount of ex-tracellular proteins, the amount of five proteins (Aur, SspA,GlpQ, Plc, and SsaA) was negatively influenced by this regu-latory system: Aur and SspA were influenced by SaeRS instrain Newman, whereas the levels of GlpQ, Plc, and SsaAwere influenced only in strain COL.
To confirm that saeS mutation was responsible for changesin the extracellular protein pattern of S. aureus, we inserted thewild-type sae operon into the geh locus of the saeS mutant ofstrain Newman. In this way the extracellular protein patterncould be restored to the wild-type phenotype, suggesting thatthe observed changes in extracellular protein expression can beattributed to the mutation in saeS (data not shown).
Finally, we compared the cytoplasmic proteome of the wild-type strains COL and Newman with those of their isogenic saeSmutants grown in TSB medium to an OD540 of 1, 6, or 10.There was no significant influence of SaeRS on this subpro-teome (data not shown). This identifies SaeRS as an important
FIG. 2. Comparison of the extracellular protein pattern of S. aureus COL with the extracellular protein pattern of S. aureus Newman.False-colored dual-channel image of 2D gels of extracellular proteins of S. aureus COL (red) and S. aureus Newman (green). Proteins (100 �g)isolated from the supernatant of S. aureus COL and S. aureus Newman grown in TSB medium to an OD540 of 10 were separated on 2D gels. Theidentified proteins are assigned to the open reading frame number as defined in the S. aureus COL, N315 and Mu50 genome sequencing projects(22, 36) and listed in Table 3. Protein spots that are yellow are present in equal amounts in both strains, protein spots that are red are present inhigher amounts in strain COL, and protein spots that are green are present in higher amounts in strain Newman.
7746 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

regulatory system for classical virulence factors, which directlyinteracts with the host.
Genomewide expression profiling of SaeRS-dependentgenes using DNA microarrays. For a global view on the role of
SaeRS in gene regulation/expression, DNA microarray studieswere carried out. Since the proteomic approach seemed toshow that SaeRS affects the amounts of both early and latevirulence factors, transcriptional analyses were performed at
TABLE 3. Extracellular proteins identified from 2D gels of S. aureus COL and S. aureus Newman
Protein Function Mr/pI matureORFID S. aureus
COL NCBI(NC002951)
GeneIDSignal peptide
prediction(SignalP)
IdentificationS. aureus
COL Newman
AhpC Alkyl hydroperoxide reductase, C subunit 21.0/4.7 SACOL0452 57286825 � �Aly N-Acetylmuramoyl-L-alanine amidase domain protein 69.3/6.3 SACOL2666 57286570 � � �Asp23 Alkaline shock protein 23 19.1/4.8 SACOL1984 57284860 � �Aur Zinc metalloproteinase aureolysin 56.4/4.9 SACOL2659 57286563 � � �Coa1 Staphylocoagulase precursor 71.7/8.9 SACOL0209 57286673 � �Coa2 Staphylocoagulase precursor 59.2/8.1 SACOL0857 57284318 � �Ear Pathogenicity island protein 20.3/9.9 SACOL0908 57285784 � �EF-G Translation elongation factor G 76.6/4.5 SACOL0593 57285609 �Eno Enolase 47.1/4.3 SACOL0842 57284304 �Exo3 Exotoxin 3, putative 27.8/9.7 SACOL1180 57284465 � �FbaA Fructose-bisphosphate aldolase, class II 30.8/4.7 SACOL2117 57286333 � �Fhs Formate-tetrahydrofolate ligase 61.2/6.8 SACOL1782 57286216 � �GapA1 Glyceraldehyde-3-phosphate dehydrogenase 36.3/4.6 SACOL0838 57284300 � �Geh Lipase precursor 71.3/9.6 SACOL0317 57285430 � �GlpQ Glycerophosphoryl diester phosphodiesterase
GlpQ, putative35.3/9.1 SACOL0962 57285836 � � �
GuaB Inosine-5-monophosphate dehydrogenase 52.9/5.7 SACOL0460 57286833 �Hlb Phospholipase C 37.2/9.1 SACOL2003 57284878 � �HlgA Gamma-hemolysin, component A 35.0/10.1 SACOL2419 57285149 � �HlgB Gamma-hemolysin, component B 36.7/9.8 SACOL2422 57285152 � � �HlgC Gamma-hemolysin, component C 35.6/9.4 SACOL2421 57285151 � � �HlY Alpha-hemolysin precursor 35.9/9.1 SACOL1173 57284458 � � �IsaA Immunodominant antigen A 24.2/6.6 SACOL2584 57286492 � � �Lip Lipase 76.7/7.7 SACOL2694 57285248 � � �LukE Leukotoxin LukE 34.8/10.0 SACOL1881 57284801 � �LukF Leukocidin F subunit precursor, putative 33.4/9.0 SACOL2004 57284879 � � �LukM Aerolysin/leukocidin family protein 40.4/10.0 SACOL2006 57284880 � �LytM Peptidoglycan hydrolase 35.1/6.4 SACOL0263 57286723 � �Nuc Thermonuclease precursor 25.1/9.7 SACOL0860 57284321 � � �Pbp2 Penicillin-binding protein 2 80.4/9.1 SACOL1490 57284591 � �Pbp3 Penicillin-binding protein 3 77.2/9.7 SACOL1609 57286131 �PdhD Pyruvate dehydrogenase complex E3 component,
lipoamide dehydrogenase49.4/4.6 SACOL1105 57285891 �
Plc 1-Phosphatidylinositol phosphodiesterase 37.1/8.6 SACOL0078 57286628 � � �Pls Methicillin-resistant surface protein 16.5/3.8 SACOL0050 57286605 � �RplM Ribosomal protein L13 16.3/9.7 SACOL2207 57284988 �SACOL0444 Conserved hypothetical protein 21.3/5.5 SACOL0444 57286817 � �SACOL0479 Surface protein, putative 56.5/4.5 SACOL0479 57285506 � �SACOL0723 LysM domain protein 28.2/6.6 SACOL0723 57285692 � �SACOL0859 Hypothetical protein 17.7/9.7 SACOL0859 57284320 � �SACOL0973 Fumarylacetoacetate hydrolase family protein 33.1/4.6 SACOL0973 57285846 �SACOL2197 Surface protein, putative 15.4/9.2 SACOL2197 57284979 � �SACOL2295 Hypothetical protein, similar to secretory antigen precursor 17.4/6.0 SACOL2295 57286421 � � �Sbi Immunoglobulin-binding protein Sbi 50.0/9.9 SACOL2418 57285148 � � �SceD SceD protein, putative 24.1/5.8 SACOL2088 57284955 � �SEA Enterotoxin type A precursor 29.7/8.1 SAV1948a 14247721 � �SEB Staphylococcal enterotoxin B 31.4/8.9 SACOL0907 57285783 � �SEI (SElQ) Staphylococcal enterotoxin type I 28.1/8.7 SACOL0887 57284346 � �SEK Staphylococcal enterotoxin 27.7/8.3 SACOL0886 57284345 � �Spa Immunoglobulin G binding protein A precursor 55.6/5.4 SACOL0095 57286644 � �SplA Serine protease SplA 25.9/9.5 SACOL1869 57284790 � � �SplB Serine protease SplB 26.1/6.7 SACOL1868 57284789 � � �SplC Serine protease SplC 26.1/6.8 SACOL1867 57284788 � � �SplF Serine protease SplF 25.7/9.7 SACOL1864 57284785 � � �SsaA Secretory antigen precursor 29.3/9.1 SACOL2291 57286417 � � �Ssl1 Staphylococcal enterotoxin-like gene 1 25.6/9.0 SACOL0468 57285495 � �Ssl11 Staphylococcal enterotoxin-like gene 11 25.4/9.1 SACOL0478 57285505 � �Ssl2 Staphylococcal enterotoxin-like gene 2 26.5/9.6 SACOL0469 57285496 � �Ssl7 Staphylococcal superantigen-like gene 7 26.2/9.2 SAV0426* 14246194 � �SspA V8 protease 36.3/4.8 SACOL1057 57284428 � � �SspB Cysteine protease precursor SspB 44.5/6.7 SACOL1056 57284427 � � �Stp Staphopain 44.2/10.1 SACOL1970 57284846 � �Tkt Transketolase 72.2/4.7 SACOL1377 57284532 �TrxB Thioredoxin-disulfide reductase 33.6/5.9 SACOL0829 57284291 �YfnI Sulfatase family protein 74.4/9.4 SACOL0778 57285741 � � �
a ORRFID of S. aureus Mu50 (36).
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7747
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

OD540s of 1 and 6. In S. aureus COL, the transcription of 12genes was positively influenced by SaeRS (Table 4). Amongthese were six genes encoding extracellular proteins and fivegenes encoding cytoplasmic proteins. The transcription of onegene belonging to the sae operon itself that probably codes fora lipoprotein seems to be also affected by SaeRS. Whether thetranscription of the sae operon is influenced by SaeRS itself orthe loss of the sae transcript is only due to the mutation eventis not entirely clear. Three of six of the extracellular proteinswere detected in the supernatant of S. aureus COL, and theabundance of two of them (HlY and Hlb) was affected bySaeRS. Interestingly, we observed no differences in the tran-scription of the serine protease genes splA and splC and of nucbetween the S. aureus COL wild type and its saeS mutant,although there was a strong negative influence at the proteinlevel (Table 4 and Fig. 3). SEB, Ssl11, SACOL0479, and thepathogenicity island protein Ear were also strongly induced at
the protein level in the saeS mutant (Fig. 3 and Table 4).Since these genes were not included in the microarray, notranscriptional data are available. Surprisingly, no geneswere found whose transcription was upregulated in the saeSmutant. Obviously, SaeRS does not inhibit gene transcrip-tion in S. aureus COL.
In strain Newman, the transcriptional level of 29 genes wasreduced in the saeS mutant (Table 4). Thirteen genes probablycode for extracellular proteins, four code for membrane pro-teins, three code for cell wall associated proteins, one codesfor a lipoprotein, and eight code for cytoplasmic proteins.Among the 13 genes encoding extracellular proteins, 5 werealso shown to be affected by SaeRS at the protein level (Ssl11,SACOL0859, Nuc, Sbi, and HlgC). The remaining eight geneproducts have not been identified in the supernatant of S.aureus Newman to date. In contrast to S. aureus COL, thetranscription of four genes was inhibited by SaeRS in strain
FIG. 3. Extracellular proteome of S. aureus COL and Newman in comparison to the extracellular proteome of their isogenic saeS mutants.False-colored dual-channel images of 2D gels illustrate the differences in the protein pattern of the wild type (red) and the respective saeS mutant(green) grown in TSB medium to an OD540 of 10. Prior to separation by 2D-PAGE, protein extracts of the respective strains were labeled withCyDye DIGE fluors (Cy3 and Cy5) (GE-Healthcare). A mixture of 50 �g of the wild type and of the respective saeS mutant was separated on bothgels. Extracellular proteins whose amounts were higher in the wild-type strain are red; proteins present only in the mutant are green. Proteinswhose levels did not change after mutation in saeS are yellow. (A) Extracellular proteome of S. aureus COL and its isogenic saeS mutant;(B) extracellular proteome of S. aureus Newman and its isogenic saeS mutant.
7748 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
Newman. However, none of these codes for a protein whoseexpression was negatively influenced by SaeRS at the proteinlevel.
Eight genes were identified whose transcription was posi-tively influenced by SaeRS both in strain COL and Newman:sbi, efb, hlb, the SACOL1169 and SACOL0480 genes, sae orf4,saeR, and saeS. The transcription of four genes (theSACOL0773, SACOL0776, and SACOL0777 genes and hly)was positively influenced by SaeRS only in COL, and the tran-scription of 21 genes (the SA0076, SA1784, SA1801,SACOL1849, SACOL1952, and SA1755 genes; geh; theSACOL0859 gene; hlgC; coa; empb; ent; fnbA; fnbB; map; nuc;the SACOL0199, SACOL0769, and SACOL1167 genes; andlrgA) was exclusively affected by the two-component regulatorysystem in strain Newman (Table 4). Only four of these aremissing from the S. aureus COL genome sequence (theSA0076, SA1784, SA1801, and SA1755 genes).
As expected, during the exponential growth phase, differ-ences in gene transcription between the wild-type strain andthe saeS mutant were observed almost exclusively in strainNewman (Table 4), which shows high levels of sae message atlow optical densities. In strain COL, differences became ap-
parent only at higher cell densities. This finding also correlateswith transcription of the sae operon (Fig. 1).
In contrast to published results (27, 42, 56), we did not findany influence of SaeRS on the transcription of the cap operonand the spa gene encoding protein A either in strain COL or instrain Newman.
Detailed transcriptional analyses of SaeRS-dependentgenes. To validate the SaeRS dependency of the expression ofgenes identified in our DNA microarray experiments, we per-formed Northern blot investigations (Fig. 4). Based on the dataobtained by the 2D gel and DNA microarray analyses, theproteins of the SaeRS regulon can be divided into two genegroups: (i) genes whose expression are influenced at the tran-scriptional level, and (ii) genes whose expression might beinfluenced at the posttranscriptional level. We selected genesfrom both groups for Northern blot analyses and investigated29 open reading frames in detail. These transcriptional analy-ses confirmed the findings in our microarray experiments inmost cases. In addition, the analyses revealed the SaeRS de-pendence of eight genes (hly, hlgB, hlgC, lukM, lukF, nuc, theSA0859 gene, and map), whereas the microarray data did notshow significant differences between the wild type and the saeS
FIG. 3—Continued.
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7749
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

TA
BL
E4.
Gen
esth
eex
pres
sion
ofw
hich
isin
fluen
ced
bySa
eRS
inS.
aure
usC
OL
and
S.au
reus
New
man
Cat
egor
yan
dge
neF
unct
ion
DN
Am
icro
arra
y2D
-PA
GE
OR
FID
S.au
reus
CO
LG
eneI
D
Spot
volr
atio
f
Spot
IDh
Spot
volr
atio
g
CO
L/
CO
L�
sae
OD
1
CO
L/
CO
L�
sae
OD
6
New
man
/N
ewm
an�
sae
OD
1
New
man
/N
ewm
an�
sae
OD
6
CO
L/
CO
L�
sae
OD
6
CO
L/
CO
L�
sae
OD
10
New
man
/N
ewm
an�
sae
OD
6
New
man
/N
ewm
an�
sae
OD
10
Gen
esw
hose
expr
essi
onis
posi
tivel
yin
fluen
ced
bySa
eRSa
Cyt
opla
smic
prot
eins
SA00
76ge
neH
ypot
hetic
alpr
otei
nSA
0076
(N31
5)15
9257
84N
CN
Cd
2.51
ND
eN
DN
DN
DSA
1784
gene
Hyp
othe
tical
prot
ein
SA17
84(N
315)
1592
7550
NC
NC
2.51
ND
ND
ND
ND
SA18
01ge
neA
ntir
epre
ssor
SA18
01(N
315)
1592
7569
NC
NC
2.67
ND
ND
ND
ND
SAC
OL
0212
gene
3-H
ydro
xyac
yl-c
oenz
yme
Ade
hydr
ogen
ase
prot
ein
SAC
OL
0212
5728
6676
3.57
ND
ND
ND
ND
saeS
Sens
orhi
stid
ine
kina
seSa
eSSA
CO
L07
6557
2857
307.
195.
0015
.38
9.43
ND
ND
ND
ND
saeR
DN
A-b
indi
ngre
spon
sere
gula
tor
SaeR
SAC
OL
0766
5728
5731
2.83
14.0
811
.62
ND
ND
ND
ND
SAC
OL
0773
gene
Para
-am
inob
enzo
ate
synt
hase
,gl
utam
ine
amid
otra
nsfe
rase
,co
mpo
nent
II
SAC
OL
0773
5728
5738
4.15
3.42
ND
ND
ND
ND
SAC
OL
0776
gene
Con
serv
edhy
poth
etic
alpr
otei
nSA
CO
L07
7657
2857
396.
493.
81N
DN
DN
DN
D
SAC
OL
0777
gene
Ure
a-am
idol
yase
-rel
ated
prot
ein
SAC
OL
0777
5728
5740
5.13
4.18
ND
ND
ND
ND
SAC
OL
1849
gene
Hyp
othe
tical
prot
ein
SAC
OL
1849
5728
4773
3.05
ND
ND
ND
ND
SAC
OL
1952
gene
Fer
ritin
fam
ilypr
otei
nSA
CO
L19
5257
2862
992.
54N
DN
DN
DN
D
Ext
race
llula
rpr
otei
nsSA
1755
gene
Hyp
othe
tical
prot
ein
SA17
55(N
315)
1592
7517
NC
NC
12.5
04.
81N
DN
DN
DN
Dge
hG
lyce
role
ster
hydr
olas
eSA
CO
L03
1757
2854
302.
71G
ehN
DN
DN
DN
Den
tSt
aphy
loco
ccal
ente
roto
xin,
puta
tive
SAC
OL
0442
5728
6815
2.65
ND
ND
ND
ND
ssl1
1St
aphy
loco
ccal
supe
rant
igen
-lik
ege
ne11
SAC
OL
0478
5728
5505
—c
——
—Ss
l11 1
ND
ND
6.13
24.3
7Ss
l11 2
ND
ND
12.6
310
.41
Ssl1
1 3N
DN
D7.
40SA
CO
L04
79ge
neSu
rfac
epr
otei
n,pu
tativ
eSA
CO
L04
7957
2855
06—
——
—SA
CO
L04
791
ND
ND
21.5
115
.07
SAC
OL
0479
2N
DN
D8.
2911
.75
SAC
OL
0480
gene
Hyp
othe
tical
prot
ein
SAC
OL
0480
5728
5507
3.79
5.75
14.7
16.
99N
DN
DN
DN
Dem
pbp
Secr
etor
yex
trac
ellu
lar
mat
rix
and
plas
ma
bind
ing
prot
ein
SAC
OL
0858
5728
4319
2.64
ND
ND
ND
ND
SAC
OL
0859
gene
Hyp
othe
tical
prot
ein
SAC
OL
0859
5728
4320
7.29
SAC
OL
0859
1N
DN
D2.
409.
13SA
CO
L05
892
ND
ND
11.9
116
.22
nuc
The
rmon
ucle
ase
prec
urso
rSA
CO
L08
6057
2843
213.
04N
uc2.
634.
058.
458.
41se
bSt
aphy
loco
ccal
ente
roto
xin
BSA
CO
L09
0757
2857
83—
——
—SE
B1
6.33
4.33
NC
NC
SEB
218
.87
35.2
1N
CN
CSE
B3
13.5
133
.83
NC
NC
SEB
47.
529.
29N
CN
CSE
B5
10.7
514
.57
NC
NC
SEB
618
.18
ND
NC
NC
ear
Path
ogen
icity
isla
ndpr
otei
nSA
CO
L09
0857
2857
84—
——
—E
ar1
3.85
3.56
NC
NC
Ear
26.
294.
39N
CN
CE
ar3
3.45
4.07
NC
NC
Ear
43.
563.
96N
CN
CE
ar5
5.05
4.75
NC
NC
efb
Fib
rino
gen-
bind
ing
prot
ein
SAC
OL
1168
5728
4453
4.21
3.90
19.2
34.
76N
DN
DN
DN
D
7750 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

SAC
OL
1169
gene
Fib
rino
gen-
bind
ing
prot
ein
prec
urso
r-re
late
dpr
otei
nSA
CO
L11
6957
2844
542.
9213
.51
3.74
ND
ND
ND
ND
hlY
Alp
ha-h
emol
ysin
prec
urso
rSA
CO
L11
7357
2844
588.
8515
.15
HlY
1N
D11
.29
11.5
429
.94
HlY
232
.26
11.7
712
.29
14.4
2H
lY3
9.80
14.8
7N
DN
DH
lY4
12.9
921
.48
ND
ND
HlY
576
.92
2.55
ND
ND
HlY
6N
D2.
68N
DN
Dsp
lCSe
rine
prot
ease
SplC
SAC
OL
1867
5728
4788
SplC
2.48
5.05
6.13
10
0sp
lASe
rine
prot
ease
SplA
SAC
OL
1869
5728
4790
SplA
ND
4.77
ND
2.45
map
Map
prot
ein
SAC
OL
2002
5728
4877
16.1
28.
47N
DN
DN
DN
Dhl
bPh
osph
olip
ase
CSA
CO
L20
0357
2848
782.
684.
5611
.11
11.1
1H
lb1
22.2
28.
50N
DN
DH
lb2
21.7
414
.94
ND
ND
Hlb
315
.15
21.3
6N
DN
DH
lb4
ND
10.3
0N
DN
DH
lb5
5.65
1.04
3N
DN
Dlu
kFL
euko
cidi
nF
subu
nit
prec
urso
r,pu
tativ
eSA
CO
L20
0457
2848
79L
ukF
15.
214.
3110
.44
17.4
7L
ukF
2N
DN
D3.
156.
67lu
kMA
erol
ysin
/leuk
ocid
infa
mily
prot
ein
SAC
OL
2006
5728
4880
Luk
MN
DN
D8.
127.
34
sbi
Imm
unog
lobu
lin-b
indi
ngpr
otei
nSB
ISA
CO
L24
1857
2851
482.
5320
.40
3.54
Sbi 1
10.6
63.
60Sb
i 2N
DN
D14
.06
7.11
Sbi 3
ND
ND
6.59
4.19
Sbi 4
ND
ND
7.33
4.48
Sbi 5
ND
ND
4.14
2.47
hlgA
Gam
ma-
hem
olys
in,
com
pone
ntA
SAC
OL
2419
5728
5149
Hlg
AN
DN
DN
D3.
28
hlgC
Gam
ma-
hem
olys
in,
com
pone
ntC
SAC
OL
2421
5728
5151
2.59
Hlg
C2.
693.
59
hlgB
Gam
ma-
hem
olys
in,
com
pone
ntB
SAC
OL
2422
5728
5152
Hlg
BN
DN
D3.
312.
78
ssl7
Stap
hylo
cocc
alsu
pera
ntig
en-
like
gene
7SA
V04
2614
2461
94N
CN
CSs
l7N
CN
C2.
68
Lip
opro
tein
ssa
eor
f4C
onse
rved
hypo
thet
ical
prot
ein
SAC
OL
0768
5728
5733
6.33
37.0
316
.12
ND
ND
ND
ND
Cel
lwal
l-ass
ocia
ted
prot
eins
coa1
Stap
hylo
coag
ulas
epr
ecur
sor
SAC
OL
0209
5728
6673
8.40
Coa
1 1N
DN
D4.
075.
56C
oa1 1
0N
DN
D4.
5C
oa1 1
1N
DN
D53
.02
17.1
4C
oa1 1
2N
DN
D53
.10
20.6
7C
oa1 1
3N
DN
D14
.80
11.3
7C
oa1 1
4N
DN
D10
.64
8.86
Coa
1 15
ND
ND
4.29
Coa
1 16
ND
ND
4.32
3.95
Coa
1 2N
DN
D5.
82C
oa1 3
ND
ND
7.49
Coa
1 4N
DN
D13
.42
31.9
3C
oa1 5
ND
ND
18.2
3C
oa1 6
ND
ND
19.4
817
.02
Coa
1 7N
DN
D18
.74
11.3
8C
oa1 8
ND
ND
42.6
115
.29
Coa
1 9N
DN
D
100
27.1
2fn
bBF
ibro
nect
inbi
ndin
gpr
otei
nB
SAC
OL
2509
5728
5194
5.29
ND
ND
ND
ND
fnbA
Fib
rone
ctin
bind
ing
prot
ein
ASA
CO
L25
1157
2851
963.
73N
DN
DN
DN
D
Con
tinue
don
follo
win
gpa
ge
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7751
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

TA
BL
E4—
Con
tinue
d
Cat
egor
yan
dge
neF
unct
ion
DN
Am
icro
arra
y2D
-PA
GE
OR
FID
S.au
reus
CO
LG
eneI
D
Spot
volr
atio
f
Spot
IDh
Spot
volr
atio
g
CO
L/
CO
L�
sae
OD
1
CO
L/
CO
L�
sae
OD
6
New
man
/N
ewm
an�
sae
OD
1
New
man
/N
ewm
an�
sae
OD
6
CO
L/
CO
L�
sae
OD
6
CO
L/
CO
L�
sae
OD
10
New
man
/N
ewm
an�
sae
OD
6
New
man
/N
ewm
an�
sae
OD
10
Mem
bran
epr
otei
nsSA
CO
L01
99ge
neC
onse
rved
hypo
thet
ical
prot
ein
SAC
OL
0199
5728
5401
3.02
ND
ND
ND
ND
lrgA
Hol
in-li
kepr
otei
nL
rgA
SAC
OL
0247
5728
6708
13.3
3N
DN
DN
DN
DSA
CO
L07
69ge
neC
onse
rved
hypo
thet
ical
prot
ein
SAC
OL
0769
5728
5734
3.11
ND
ND
ND
ND
SAC
OL
1167
gene
Hyp
othe
tical
prot
ein
SAC
OL
1167
5728
4452
3.05
ND
ND
ND
ND
Gen
esw
hose
expr
essi
onis
nega
tivel
yin
fluen
ced
bySa
eRSb
Cyt
opla
smic
prot
eins
SA00
42ge
neC
onse
rved
hypo
thet
ical
prot
ein
SA00
42(N
315)
1592
5749
NC
NC
0.32
ND
ND
ND
ND
SA16
36ge
neH
ypot
hetic
alpr
otei
n,pa
thog
enic
ityis
land
saPI
3SA
1636
(N31
5)15
9273
92N
CN
C0.
28N
DN
DN
DN
D
SA18
09ge
neH
ypot
hetic
alpr
otei
nSA
1809
(N31
5)15
9275
77N
CN
C0.
23N
DN
DN
DN
D
Ext
race
llula
rpr
otei
nspl
c1-
Phos
phat
idyl
inos
itol
phos
phod
iest
eras
eSA
CO
L00
7857
2866
28Pl
c 10.
260.
34Pl
c 20.
37ly
sML
ysM
dom
ain
prot
ein
SAC
OL
0507
5728
5532
0.32
ND
ND
ND
ND
glpQ
Gly
cero
phos
phor
yldi
este
rSA
CO
L09
6257
2858
36G
lpQ
10.
390.
14ph
osph
odie
ster
ase
Glp
Q,
puta
tive
Glp
Q2
0.31
sspA
V8
prot
ease
SAC
OL
1057
5728
4428
SspA
10.
280.
45Ss
pA2
0.23
0.39
SspA
30.
220.
33Ss
pA4
0.41
0.37
SspA
50.
290.
31ss
aASe
cret
ory
antig
enpr
ecur
sor
SAC
OL
2291
5728
6417
SsaA
10.
440.
20Ss
aA2
0.41
0.38
aur
Zin
cm
etal
lopr
otei
nase
aure
olys
inSA
CO
L26
5957
2865
63A
ur0.
340.
36
aT
hat
is,g
enes
who
seex
pres
sion
ispo
sitiv
ely
influ
ence
dby
SaeR
S.b
Tha
tis
,gen
esw
hose
expr
essi
onis
nega
tivel
yin
fluen
ced
bySa
eRS.
c—
,Gen
esno
tpr
esen
ton
the
DN
Am
icro
arra
yus
edin
this
stud
y(S
cien
ion)
.d
NC
,pro
tein
sno
ten
code
din
geno
me
sequ
ence
ofth
ere
spec
tive
stra
in.
eN
D,p
rote
ins
not
iden
tified
on2D
gelo
fse
cret
edpr
otei
ns.
fR
atio
sof
�2.
5or
�0.
4ar
esh
own
that
repr
esen
tdi
ffere
nces
betw
een
the
resp
ectiv
est
rain
sat
asi
gnifi
canc
ele
velo
f0.
05.
gR
atio
sof
�2
or�
0.5
are
show
nth
atre
pres
ent
diffe
renc
esbe
twee
nth
ere
spec
tive
stra
ins
ata
sign
ifica
nce
leve
lof
0.05
.h
See
Fig
.3.
7752 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

mutant. A common theme of these genes is a relatively lowtranscription level that might account for the failure to bedetected as significantly influenced by SaeRS in the array ap-proach.
Moreover, the transcription of four genes (seb, ssl11, theSACOL0479 gene, and ear) that were not present on the mi-croarray was also analyzed by Northern blot experiments. Theobtained data revealed that their transcription was positivelyinfluenced by SaeRS (Fig. 4).
Taken together, our data show that SaeRS seems to activatetranscription of the respective genes rather than to inhibittranscription. The negative effect of SaeRS on the amount ofAur, GlpQ, Plc, SsaA, and SspA probably occurs at the post-transcriptional level, since the transcription of the correspond-ing genes was not influenced by the mutation in saeS duringgrowth in TSB medium (shown for OD540s of 1 and 6 in Fig. 4).
Influence of SaeRS on the transcription of regulatory genesinvolved in virulence gene expression. Most of the genes whoseexpression appears to be influenced by SaeRS are also affectedby agr, SarA, and �B (7, 15, 60, 61). To determine whether theobserved effects of the mutation in saeS on the expression ofthese genes might be mediated by these regulators, the poten-tial influence of SaeRS on the transcription of sigB, sarA, theagrA and RNAIII genes, and arlRS was investigated by North-ern blotting. Similar to sae transcription, differences in thegrowth phase dependence of the transcription of sigB, the agrAand RNAIII genes, and sarA between both wild-type strainscould be observed. Whereas in S. aureus Newman the tran-script levels of all of these genes were largely constant, theirtranscription in S. aureus COL was strongly dependent on thegrowth phase. Here the transcription of the agrA and RNAIIIgenes was upregulated at higher optical densities (OD540 of 6),whereas the transcripts of sigB, arlRS, and sarA disappearedsimultaneously. Importantly, SaeRS did not influence the reg-ulation of these genes (Fig. 5).
DISCUSSION
In addition to host factors (e.g., the immune response ofhealthy or immunocompromised individuals), the pronounceddiversity of the species S. aureus in its equipment with virulencefactors might be responsible for the wide variety of clinicalsymptoms that are characteristic for infections with this micro-organism. Most genes encoding virulence factors are locatedon highly variable regions of the staphylococcal genome, suchas pathogenicity islands, lysogenic bacteriophages, or even onplasmids (2, 39, 40). Interestingly, some virulence-associatedgenes, such as spa, aur, hla, lip, clfAB, map/eap, fnbA, and coa,also belong to the core genome (31, 43). The expression ofvirulence factors in S. aureus is regulated by a very complexnetwork of regulators such as DNA-binding proteins, two-component systems, and the alternative sigma factor �B. How-ever, the contribution of each regulator on virulence geneexpression, their regulatory mechanisms, and their mutual in-terference within this regulatory network are not well under-stood. To elucidate the role of the two-component systemSaeRS in S. aureus virulence gene expression, we combinedproteomic and transcriptomic approaches. Both techniques areexcellent tools to reveal whether individual virulence genes (i)are expressed at all and, if so, (ii) in what quantities and (iii)
under which environmental conditions. The study was initiatedin strain COL, where the genome sequence became availablein 2005 (22). S. aureus Newman was later included in thepresent study because this strain is characterized by an unusu-ally high level of sae-specific mRNA.
Extracellular proteins constitute a reservoir of virulence fac-tors and have important roles in the pathogenicity of bacteria.Therefore, besides the elucidation of virulence factor regula-tion, the comprehensive analysis of the extracellular proteomeof S. aureus may lead to the discovery of new virulence factors.In our study, the amount of eight extracellular proteins in S.aureus COL and 15 in S. aureus Newman was influenced by thetwo-component system SaeRS. Among these were knownSaeRS-dependent proteins such as Nuc, HlY, Hlb, Eap (Map),and Emp (Empbp) (27–30, 42), which confirms the validity ofour experimental approach. In addition, 13 new extracellularproteins which likely are under SaeRS control could be iden-tified. Interestingly, most of the proteins whose amount wasinfluenced by SaeRS play a role in immune evasion (for areview, see reference 19). These are Map, Eap, Efb, HlgA,HlgB, HlgC, LukF, LukM, Ent, SEB, Sbi, and Aur. Others areespecially involved in adhesion to host cells (FnbA and FnbB),or they can damage host cell membranes (Hlb and HlY). Wealso discovered proteins of unknown function in the SaeRSregulon. These are SACOL0479, SACOL0480, SACOL0859,and SACOL1169. Their presence in virulence-associated regu-lons makes it very likely that they play a role in the interactionof S. aureus with its host, probably in immune evasion oradhesion. Interestingly, SACOL1169 shows similarity to fi-brinogen-binding proteins. The elucidation of the precisefunction of the putative virulence factors will be a challengingtask for the future. The amounts of four virulence factors weresimilarly influenced by SaeRS in strain COL and in strainNewman. Interestingly, in strain Newman, more extracellularproteins were positively influenced by the two-component sys-tem, in particular proteins expressed at the early phase ofgrowth.
To distinguish between the effects of SaeRS at the transcrip-tional level and those due to changes at the protein level, wecomplemented our proteome data with transcriptome analysesusing a full genome DNA microarray corresponding to thegenome sequence of S. aureus N315. In this way we couldclearly show that the positive control of genes by SaeRS oc-curred at the transcriptional level. In contrast, the expressionof genes coding for proteins, the amount of which was nega-tively influenced by SaeRS (SsaA, Plc, SspA, and GlpQ) wasnot affected by the two-component system at the mRNA level.Although the aforementioned proteins were identified in bothstrains, the proteases Aur and SspA accumulated only in thesaeS mutant of strain Newman, whereas the lipases GlpQ andPlc, as well as SsaA, accumulated solely in the saeS mutant ofstrain COL.
The transcriptomic approach further allowed the investiga-tion of the transcriptional regulation of genes encoding sur-face-associated and membrane proteins that escape proteomeanalyses focusing on extracellular and cytoplasmic proteins. S.aureus Newman produces significant amounts of surface-asso-ciated proteins leading to exceptionally strong adhesion of thisstrain to host cells and extracellular matrix (59). Interestingly,the expression of genes encoding Coa, FnbB, and FnbA was
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7753
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

FIG. 4. Northern blot analyses of SaeRS-dependent genes. RNA was isolated from S. aureus COL and S. aureus Newman and their respective saeSmutants (�) grown in TSB medium at 37°C (OD540 1 [lanes 1] and OD540 6 [lanes 6]). The membranes were hybridized with digoxigenin-labeledRNA probes specific for the respective genes. Relevant transcripts are indicated by arrows. Schematic representations of the gene loci based on thesequence of S. aureus COL are shown. The dotted lines represent the RNA probe used in the respective experiment. (A) Genes encoding proteins whoseamounts were positively regulated by SaeRS; (B) genes encoding proteins whose stability might be influenced by SaeRS.
7754 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

FIG. 4—Continued.
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7755
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

shown to be positively influenced by SaeRS at low opticaldensities. The same phenomenon was observed for theSACOL0199, SACOL0769, and SACOL1167 genes and lrgA,which code for proteins with membrane-spanning domains. Athigh optical densities, however, the transcription of all of thesegenes was repressed despite the presence of SaeRS. In contrastto the positive effect of SaeRS, the agr quorum-sensing systeminhibits the expression of coa, fnbB, and fnbA, indicating thatRNAIII might act as an antagonist of SaeRS (24, 48, 58, 59).
In contrast to the genes encoding FnbA, FnbB, and Coa,most other genes of the SaeRS regulon, however, are positivelycoregulated both by RNAIII and by SaeRS (9, 15, 24, 26, 30,46, 51, 60). RNAIII appears to synergize with the positive
effect of SaeRS on gene expression at higher optical densitieseither directly or indirectly by enhancing SaeR activity (25, 28,42). Furthermore, the expression of most SaeRS-dependentgenes appears to be positively affected by SarA and negativelyby the alternative sigma factor �B (7, 9, 12, 15, 28–30, 46, 49,60, 61). For nine SaeRS-dependent genes (ear; efb; ent; theSACOL0479, SACOL0480, SACOL0859, and SACOL1169genes; ssl11; and ssl7) identified in the present study, an influ-ence of agr, SarA, �B, ArlSR, or Rot on their transcription hasnot yet been observed (7, 15, 37, 47). However, the publisheddata were obtained with other S. aureus strains and under verydifferent conditions. Nevertheless, the effect of SaeRS seems tobe restricted to virulence gene regulation, which is only a
FIG. 5. Transcriptional analysis of virulence-associated regulatory genes. RNA was isolated from S. aureus COL and S. aureus Newman andtheir respective saeS mutants (�) grown in TSB medium at 37°C (OD540 1 [lanes 1] and OD540 6 [lanes 6]). The membranes were hybridizedwith digoxigenin-labeled RNA probes specific for the respective genes. The relevant transcripts are indicated. Schematic representations of thegene loci based on the sequence of S. aureus COL are shown. The dotted lines represent the RNA probes used in the respective experiments.
7756 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

subset of the many genes described to be regulated by theother regulators such as agr or SarA.
The present study provides clear evidence that althoughsome virulence-associated genes are present in strain COL andin strain Newman the expression of these genes varied betweenthese strains. Whereas some of these virulence factors weresynthesized in different amounts, other virulence factors arenot expressed at all in one of the two strains (Fig. 2). Thereare two possible explanations for this: (i) the activity of viru-lence-associated regulators differs between the strains and/or(ii) there are differences between the regulatory regions of therespective genes. The comparison of the two S. aureus strainsrevealed a correlation between the transcription of regulatorygenes and the expression of virulence-associated genes. Weobserved differences in the amount of transcripts of regulatorygenes such as agr, sarA, sigB, and saeRS between COL andNewman. However, the differences in the transcriptional reg-ulation were most at the sae and the agr loci. Very similarobservations for the sae operon were reported by other groups(30, 56) that observed reduced sae transcription in derivativesof 8325 compared to strain Newman. Moreover, the fact that S.aureus Newman shows an extraordinary behavior in regulationof gene expression is also supported by the finding that muta-tions in sarA and agrA did not affect biofilm formation, asobserved for many other strains (3). Analysis of the SaeRSregulon structure in strain COL and strain Newman grownunder nearly identical conditions reveals remarkable differ-ences. Consequently, for a thorough understanding of the com-plex interaction of S. aureus regulators and their function in thevirulence of S. aureus, the global role of these regulators ingene expression must be analyzed systematically in variousstrains with different clinical behaviors and under strictly de-fined conditions. However, the present study is limited sincethe genome sequence of S. aureus Newman has not been avail-able to date. Therefore, for a more comprehensive analysis ofdifferences in global virulence gene expression in various S.aureus strains, only sequenced strains should be used in orderto determine whether the respective genes are present andwhether there are sequence variations particularly in the reg-ulatory regions of these genes or within regulatory genes.
The genomewide expression profiling of SaeRS-dependentgenes revealed that the regulator SaeR might act as an activa-tor of transcription. SaeR is a typical response regulator that isexpected to be activated by phosphorylation and then able tobind to a specific region of the DNA in front of its target genes.Clearly, SaeRS is involved in the tight control of the temporallycoordinated expression of virulence factors in S. aureus. How-ever, its role within the regulatory network is not yet entirelyclear. SaeRS did not affect the transcription of other regulatorygenes, suggesting that SaeR might be an essential downstreameffector molecule within the regulatory network. WhetherSaeR directly regulates its target genes remains to be deter-mined. A comparison of the upstream sequences of all SaeRS-dependent genes, however, did not reveal a consensus se-quence that might serve as a binding site of SaeR. Gel shiftexperiments are currently under way to determine whetherSaeR directly binds to the promoter region of the SaeRS-dependent genes.
In conclusion, we have identified SaeRS as an importantregulatory system of staphylococcal virulence gene expression.
Many SaeRS-dependent genes are also regulated by agr, SarA,and/or �B. According to the function of SaeRS-dependentgenes, the two-component system might be crucial for thecomplex interactions of S. aureus with the eukaryotic immunesystem via expression of proteins involved in adhesion andimmune evasion. The signals that are responsible for the acti-vation of SaeRS under in vivo conditions remain to be eluci-dated.
ACKNOWLEDGMENTS
We thank Silva Holtfreter for helpful discussion on SAg nomencla-ture and Birgit Voigt, Haike Henkel, and Dirk Albrecht for support inprotein digestion and identification. We are grateful to Thomas Meierand Anita Harang for excellent technical assistance. We also thankDecodon GmbH (Greifswald, Germany) for providing Delta2D soft-ware.
This study was supported by grants of the BMBF (031U107A/-207A,031U213B), the DFG (GK212/3-00, WO578/5-1), the Land MV, andthe Fonds der Chemischen Industrie to M.H., C.W., and S.E.
REFERENCES
1. Bae, T., A. K. Banger, A. Wallace, E. M. Glass, F. Aslund, O. Schneewind,and D. M. Missiakas. 2004. Staphylococcus aureus virulence genes identifiedby bursa aurealis mutagenesis and nematode killing. Proc. Natl. Acad. Sci.USA 101:12312–12317.
2. Bayles, K. W., and J. J. Iandolo. 1989. Genetic and molecular analyses of thegene encoding staphylococcal enterotoxin D. J. Bacteriol. 171:4799–4806.
3. Beenken, K. E., J. S. Blevins, and M. S. Smeltzer. 2003. Mutation of sarA inStaphylococcus aureus limits biofilm formation. Infect. Immun. 71:4206–4211.
4. Benton, B. M., J. P. Zhang, S. Bond, C. Pope, T. Christian, L. Lee, K. M.Winterberg, M. B. Schmid, and J. M. Buysse. 2004. Large-scale identificationof genes required for full virulence of Staphylococcus aureus. J. Bacteriol.186:8478–8489.
5. Berger-Bachi, B. 1983. Increase in transduction efficiency of Tn551 mediatedby the methicillin resistance marker. J. Bacteriol. 154:533–535.
6. Bernhardt, J., K. Buttner, C. Scharf, and M. Hecker. 1999. Dual channelimaging of two-dimensional electropherograms in Bacillus subtilis. Electro-phoresis 20:2225–2240.
7. Bischoff, M., P. Dunman, J. Kormanec, D. Macapagal, E. Murphy, W.Mounts, B. Berger-Bachi, and S. Projan. 2004. Microarray-based analysis ofthe Staphylococcus aureus �B regulon. J. Bacteriol. 186:4085–4099.
8. Blum, H., H. Beier, and H. S. Gross. 1987. Improved silver staining of plantprotein, RNA and DNA in polyacrylamide gels. Electrophoresis 8:93–99.
9. Bronner, S., P. Stoessel, A. Gravet, H. Monteil, and G. Prevost. 2000. Vari-able expressions of Staphylococcus aureus bicomponent leucotoxins semi-quantified by competitive reverse transcription-PCR. Appl. Environ. Micro-biol. 66:3931–3938.
10. Candiano, G., M. Bruschi, L. Musante, L. Santucci, G. M. Ghiggeri, B.Carnemolla, P. Orecchia, L. Zardi, and P. G. Righetti. 2004. Blue silver: avery sensitive colloidal Coomassie G-250 staining for proteome analysis.Electrophoresis 25:1327–1333.
11. Chauvenet, W. 1863. A manual of spherical and practical astronomy. J. B.Lippincott, Philadelphia, Pa.
12. Cheung, A. L., Y. T. Chien, and A. S. Bayer. 1999. Hyperproduction ofalpha-hemolysin in a sigB mutant is associated with elevated SarA expressionin Staphylococcus aureus. Infect. Immun. 67:1331–1337.
13. Cleveland, W. S., E. Grosse, and W. M. Shyu. 1992. Local regression models,p. 309–376. In J. M. Chambers and T. J. Hastie (ed.), Statistical models.Wadsworth & Brooks, Pacific Grove, Calif.
14. Coleman, D. C., D. J. Sullivan, R. J. Russell, J. P. Arbuthnott, B. F. Carey,and H. M. Pomeroy. 1989. Staphylococcus aureus bacteriophages mediatingthe simultaneous lysogenic conversion of beta-lysin, staphylokinase and en-terotoxin A: molecular mechanism of triple conversion. J. Gen. Microbiol.135:1679–1697.
15. Dunman, P. M., E. Murphy, S. Haney, D. Palacios, G. Tucker-Kellogg, S.Wu, E. L. Brown, R. J. Zagursky, D. Shlaes, and S. J. Projan. 2001. Tran-scription profiling-based identification of Staphylococcus aureus genes regu-lated by the agr and/or sarA loci. J. Bacteriol. 183:7341–7353.
16. Duthie, E. S., and L. L. Lorenz. 1952. Staphylococcal coagulase; mode ofaction and antigenicity. J. Gen. Microbiol. 6:95–107.
17. Edwards, D. E. 2003. Non-linear normalization and background correctionin one channel cDNA microarray studies. Bioinformatics 19:825–833.
18. Eymann, C., A. Dreisbach, D. Albrecht, J. Bernhardt, D. Becher, S. Gentner,L. T. Tam, K. Buttner, G. Buurman, C. Scharf, S. Venz, U. Volker, and M.Hecker. 2004. A comprehensive proteome map of growing Bacillus subtiliscells. Proteomics 4:2849–2876.
VOL. 188, 2006 SaeRS REGULON IN S. AUREUS 7757
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

19. Foster, T. J. 2005. Immune evasion by staphylococci. Nat. Rev. Microbiol.3:948–958.
20. Fournier, B., A. Klier, and G. Rapoport. 2001. The two-component systemArlS-ArlR is a regulator of virulence gene expression in Staphylococcusaureus. Mol. Microbiol. 41:247–261.
21. Gertz, S., S. Engelmann, R. Schmid, K. Ohlsen, J. Hacker, and M. Hecker.1999. Regulation of �B-dependent transcription of sigB and asp23 in twodifferent Staphylococcus aureus strains. Mol. Gen. Genet. 261:558–566.
22. Gill, S. R., D. E. Fouts, G. L. Archer, E. F. Mongodin, R. T. Deboy, J. Ravel,I. T. Paulsen, J. F. Kolonay, L. Brinkac, M. Beanan, R. J. Dodson, S. C.Daugherty, R. Madupu, S. V. Angiuoli, A. S. Durkin, D. H. Haft, J.Vamathevan, H. Khouri, T. Utterback, C. Lee, G. Dimitrov, L. Jiang, H. Qin,J. Weidman, K. Tran, K. Kang, I. R. Hance, K. E. Nelson, and C. M. Fraser.2005. Insights on evolution of virulence and resistance from the completegenome analysis of an early methicillin-resistant Staphylococcus aureus strainand a biofilm-producing methicillin-resistant Staphylococcus epidermidisstrain. J. Bacteriol. 187:2426–2438.
23. Giraudo, A. T., A. Calzolari, A. A. Cataldi, C. Bogni, and R. Nagel. 1999. Thesae locus of Staphylococcus aureus encodes a two-component regulatorysystem. FEMS Microbiol. Lett. 177:15–22.
24. Giraudo, A. T., A. L. Cheung, and R. Nagel. 1997. The sae locus of Staphy-lococcus aureus controls exoprotein synthesis at the transcriptional level.Arch. Microbiol. 168:53–58.
25. Giraudo, A. T., C. Mansilla, A. Chan, C. Raspanti, and R. Nagel. 2003.Studies on the expression of regulatory locus sae in Staphylococcus aureus.Curr. Microbiol. 46:246–250.
26. Giraudo, A. T., H. Rampone, A. Calzolari, and R. Nagel. 1996. Phenotypiccharacterization and virulence of a sae� agr� mutant of Staphylococcusaureus. Can. J. Microbiol. 42:120–123.
27. Giraudo, A. T., C. G. Raspanti, A. Calzolari, and R. Nagel. 1994. Charac-terization of a Tn551-mutant of Staphylococcus aureus defective in the pro-duction of several exoproteins. Can. J. Microbiol. 40:677–681.
28. Goerke, C., U. Fluckiger, A. Steinhuber, V. Bisanzio, M. Ulrich, M. Bischoff,J. M. Patti, and C. Wolz. 2005. Role of Staphylococcus aureus global regu-lators sae and �B in virulence gene expression during device-related infec-tion. Infect. Immun. 73:3415–3421.
29. Goerke, C., U. Fluckiger, A. Steinhuber, W. Zimmerli, and C. Wolz. 2001.Impact of the regulatory loci agr, sarA, and sae of Staphylococcus aureus onthe induction of alpha-toxin during device-related infection resolved by di-rect quantitative transcript analysis. Mol. Microbiol. 40:1439–1447.
30. Harraghy, N., J. Kormanec, C. Wolz, D. Homerova, C. Goerke, K. Ohlsen, S.Qazi, P. Hill, and M. Herrmann. 2005. sae is essential for expression of thestaphylococcal adhesins Eap and Emp. Microbiology 151:1789–1800.
31. Holden, M. T., E. J. Feil, J. A. Lindsay, S. J. Peacock, N. P. Day, M. C.Enright, T. J. Foster, C. E. Moore, L. Hurst, R. Atkin, A. Barron, N. Bason,S. D. Bentley, C. Chillingworth, T. Chillingworth, C. Churcher, L. Clark, C.Corton, A. Cronin, J. Doggett, L. Dowd, T. Feltwell, Z. Hance, B. Harris, H.Hauser, S. Holroyd, K. Jagels, K. D. James, N. Lennard, A. Line, R. Mayes,S. Moule, K. Mungall, D. Ormond, M. A. Quail, E. Rabbinowitsch, K.Rutherford, M. Sanders, S. Sharp, M. Simmonds, K. Stevens, S. Whitehead,B. G. Barrell, B. G. Spratt, and J. Parkhill. 2004. Complete genomes of twoclinical Staphylococcus aureus strains: evidence for the rapid evolution ofvirulence and drug resistance. Proc. Natl. Acad. Sci. USA 101:9786–9791.
32. Ito, T., Y. Katayama, K. Asada, N. Mori, K. Tsutsumimoto, C. Tiensasitorn,and K. Hiramatsu. 2001. Structural comparison of three types of staphylo-coccal cassette chromosome mec integrated in the chromosome in methicil-lin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother.45:1323–1336.
33. Jorgensen, E. D., R. K. Durbin, S. S. Risman, and W. T. McAllister. 1991.Specific contacts between the bacteriophage T3, T7, and SP6 RNA poly-merases and their promoters. J. Biol. Chem. 266:645–651.
34. Kohler, C., S. Wolff, D. Albrecht, S. Fuchs, D. Becher, K. Buttner, S.Engelmann, and M. Hecker. 2005. Proteome analyses of Staphylococcusaureus in growing and non-growing cells: a physiological approach. Int.J. Med. Microbiol. 295:547–565.
35. Kuroda, M., H. Kuroda, T. Oshima, F. Takeuchi, H. Mori, and K. Hira-matsu. 2003. Two-component system VraSR positively modulates the regu-lation of cell-wall biosynthesis pathway in Staphylococcus aureus. Mol. Mi-crobiol. 49:807–821.
36. Kuroda, M., T. Ohta, I. Uchiyama, T. Baba, H. Yuzawa, I. Kobayashi, L. Cui,A. Oguchi, K. Aoki, Y. Nagai, J. Lian, T. Ito, M. Kanamori, H. Matsumaru,A. Maruyama, H. Murakami, A. Hosoyama, Y. Mizutani-Ui, N. K. Taka-hashi, T. Sawano, R. Inoue, C. Kaito, K. Sekimizu, H. Hirakawa, S. Kuhara,S. Goto, J. Yabuzaki, M. Kanehisa, A. Yamashita, K. Oshima, K. Furuya, C.Yoshino, T. Shiba, M. Hattori, N. Ogasawara, H. Hayashi, and K. Hira-matsu. 2001. Whole genome sequencing of methicillin-resistant Staphylococ-cus aureus. Lancet 357:1225–1240.
37. Liang, X., L. Zheng, C. Landwehr, D. Lunsford, D. Holmes, and Y. Ji. 2005.
Global regulation of gene expression by ArlRS, a two-component signaltransduction regulatory system of Staphylococcus aureus. J. Bacteriol. 187:5486–5492.
38. Lina, G., G. A. Bohach, S. P. Nair, K. Hiramatsu, E. Jouvin-Marche, and R.Mariuzza. 2004. Standard nomenclature for the superantigens expressed byStaphylococcus. J. Infect. Dis. 189:2334–2336.
39. Narita, S., J. Kaneko, J. Chiba, Y. Piemont, S. Jarraud, J. Etienne, and Y.Kamio. 2001. Phage conversion of Panton-Valentine leukocidin in Staphy-lococcus aureus: molecular analysis of a PVL-converting phage, phiSLT.Gene 268:195–206.
40. Novick, R. P. 2003. Mobile genetic elements and bacterial toxinoses: thesuperantigen-encoding pathogenicity islands of Staphylococcus aureus. Plas-mid 49:93–105.
41. Novick, R. P. 2003. Autoinduction and signal transduction in the regulationof staphylococcal virulence. Mol. Microbiol. 48:1429–1449.
42. Novick, R. P., and D. Jiang. 2003. The staphylococcal saeRS system coordi-nates environmental signals with agr quorum sensing. Microbiology 149:2709–2717.
43. Peacock, S. J., C. E. Moore, A. Justice, M. Kantzanou, L. Story, K. Mackie,G. O’Neill, and N. P. Day. 2002. Virulent combinations of adhesin and toxingenes in natural populations of Staphylococcus aureus. Infect. Immun. 70:4987–4996.
44. R Development Core Team. 2005. R: a language and environment for sta-tistical computing. Foundation for Statistical Computing, Vienna, Austria.
45. Rampone, H., G. L. Martinez, A. T. Giraudo, A. Calzolari, and R. Nagel.1996. In vivo expression of exoprotein synthesis with a Sae mutant of Staph-ylococcus aureus. Can. J. Vet. Res. 60:237–240.
46. Reed, S. B., C. A. Wesson, L. E. Liou, W. R. Trumble, P. M. Schlievert, G. A.Bohach, and K. W. Bayles. 2001. Molecular characterization of a novelStaphylococcus aureus serine protease operon. Infect. Immun. 69:1521–1527.
47. Said-Salim, B., P. M. Dunman, F. M. McAleese, D. Macapagal, E. Murphy,P. J. McNamara, S. Arvidson, T. J. Foster, S. J. Projan, and B. N.Kreiswirth. 2003. Global regulation of Staphylococcus aureus genes by Rot.J. Bacteriol. 185:610–619.
48. Saravia-Otten, P., H. P. Muller, and S. Arvidson. 1997. Transcription ofStaphylococcus aureus fibronectin binding protein genes is negatively regu-lated by agr and an agr-independent mechanism. J. Bacteriol. 179:5259–5263.
49. Schmidt, K. A., N. P. Donegan, W. A. Kwan, Jr., and A. Cheung. 2004.Influences of �B and agr on expression of staphylococcal enterotoxin B (seb)in Staphylococcus aureus. Can. J. Microbiol. 50:351–360.
50. Shafer, W. M., and J. J. Iandolo. 1979. Genetics of staphylococcal entero-toxin B in methicillin-resistant isolates of Staphylococcus aureus. Infect. Im-mun. 25:902–911.
51. Shaw, L., E. Golonka, J. Potempa, and S. J. Foster. 2004. The role andregulation of the extracellular proteases of Staphylococcus aureus. Microbi-ology 150:217–228.
52. Smyth, G. K. 2004. Linear models and empirical Bayes methods for assessingdifferential expression in microarray experiments. Stat. Appl. Genet. Mol.Biol. 3:Article3.
53. Smyth, G. K. 2005. Limma: linear models for microarray data, p. 397–420. InR. Gentleman, V. Carey, S. Dudoit, R. Irizarry, and W. Huber (ed.), Bioin-formatics and computational biology solutions using R and bioconductor.Springer, New York, N.Y.
54. Smyth, G. K., J. Michaud, and H. S. Scott. 2005. Use of within-array repli-cate spots for assessing differential expression in microarray experiments.Bioinformatics 21:2067–2075.
55. Smyth, G. K., and T. Speed. 2003. Normalization of cDNA microarray data.Methods 31:265–273.
56. Steinhuber, A., C. Goerke, M. G. Bayer, G. Doring, and C. Wolz. 2003.Molecular architecture of the regulatory locus sae of Staphylococcus aureusand its impact on expression of virulence factors. J. Bacteriol. 185:6278–6286.
57. Wetzstein, M., U. Volker, J. Dedio, S. Lobau, U. Zuber, M. Schiesswohl, C.Herget, M. Hecker, and W. Schumann. 1992. Cloning, sequencing, andmolecular analysis of the dnaK locus from Bacillus subtilis. J. Bacteriol.174:3300–3310.
58. Wolz, C., D. McDevitt, T. J. Foster, and A. L. Cheung. 1996. Influence of agron fibrinogen binding in Staphylococcus aureus Newman. Infect. Immun.64:3142–3147.
59. Wolz, C., P. Pohlmann-Dietze, A. Steinhuber, Y. T. Chien, A. Manna, W. vanWamel, and A. Cheung. 2000. Agr-independent regulation of fibronectin-binding protein(s) by the regulatory locus sar in Staphylococcus aureus. Mol.Microbiol. 36:230–243.
60. Ziebandt, A. K., D. Becher, K. Ohlsen, J. Hacker, M. Hecker, and S. Engelmann.2004. The influence of agr and �B in growth phase-dependent regulation of viru-lence factors in Staphylococcus aureus. Proteomics 4:3034–3047.
61. Ziebandt, A. K., H. Weber, J. Rudolph, R. Schmid, D. Hoper, S. Engelmann,and M. Hecker. 2001. Extracellular proteins of Staphylococcus aureus and therole of SarA and �B. Proteomics 1:480–493.
7758 ROGASCH ET AL. J. BACTERIOL.
on June 15, 2019 by guesthttp://jb.asm
.org/D
ownloaded from







