
Molecular Biosystems
Cite this: DOI: 10.1039/c0xx00000x
www.rsc.org/xxxxxx
Dynamic Article Links ►
Communication Oligomerization Enhancement and Two Domain Swapping Mode Detection for Thermostable Cytochrome c552 by Elongation of the Major Hinge Loop† Chunguang Ren,a Satoshi Nagao,a Masaru Yamanaka,a Hirofumi Komori,b Yasuhito Shomura,c,d Yoshiki Higuchi,c,d and Shun Hirota*a 5
Received (in XXX, XXX) Xth XXXXXXXXX 20XX, Accepted Xth XXXXXXXXX 20XX DOI: 10.1039/b000000x
High-order oligomers of Hydrogenobacter thermophilus cytochrome c552 increased as inserting more Gly residues between Ala18 and Lys19 at the major hinge loop of the 10
wild-type protein. N-terminal and C-terminal domain swapping were elucidated by X-ray crystallography for the mutant with insertion of three Gly residues at the hinge loop.
Protein self-assemblies have been constructed by various groups utilizing well-established interactions, such as metal 15
coordination,1 chemical cross-linking,2 host−guest interaction,3 hydrophobic interaction,4 and electrostatic interaction.5 Structured protein oligomers may be used as building blocks to create diverse functional nanomaterials.6 For example, well-ordered porphyrin clusters constructed with heme protein 20
nanorings have been reported.7 Proteins also self-assemble by three-dimensional (3D) domain swapping, which has been defined by Eisenberg and co-workers.8 In domain swapping, a protein molecule exchanges its structural element or domain with the corresponding region of another molecule.9 We have 25
shown that heme proteins,10 including horse cytochrome (cyt) c10a and Hydrogenobacter thermophilus (HT) cyt c552,10b domain swap. HT cyt c552 has three long α-helices, and His14 and Met59 are coordinated to the heme iron.11 HT cyt c552 exhibits high stability, where the denaturation temperature of the oxidized 30
protein is higher than 100 °C.12 Different domain swapping modes (N-terminal and C-terminal regions) and different oligomerization orders have been observed for proteins in the same c-type cyt protein family. HT cyt c552 mainly forms small size oligomers by swapping the region containing the N-35
terminal α-helix and heme,10b whereas horse cyt c forms high-order oligomers larger than 50mers by swapping the C-terminal α-helix.10a Since the hinge loop of HT cyt c552
(Ala18Lys19Lys20) is shorter than that of horse cyt c (Thr78‒Ala83), high-order oligomer formation may have been 40
inhibited by the steric hindrance between the protomers in the HT cyt c552 oligomer.10b In this study, the amount of high-order oligomers increased by elongation of the hinge loop in HT cyt c552. We also obtained two different thermostable domain-swapped dimers of the three Gly-inserted HT cyt c552 mutant, 45
and elucidated their structures by X-ray crystallography, showing that domain swapping may occur at both N-terminal and C-terminal regions in HT cyt c552. 50
55
Fig. 1. Schematic view of insertion of Gly residues between Ala18 and 60
Lys19 at the hinge loop of WT cyt c552 for formation of high-order oligomers.
We constructed HT cyt c552 mutants by inserting one (insG1), two (insG2) or three (insG3) Gly residues between Ala18 and 65
Lys19 at the hinge loop of the wild-type (WT) protein to reduce the steric repulsion in the oligomers, so that the mutants may form high-order oligomers (Figure 1). Oxidized WT cyt c552 and its Gly-inserted mutants in 50 mM potassium phosphate buffer, pH 7.0, precipitated by an addition of ethanol (90% (v/v)). After 70
the precipitate was lyophilized and dissolved in the same buffer, the amount of high-order oligomers (larger than decamers) eluting earlier than the elution volume of 13.7 mL in the size exclusion chromatogram obtained using Superdex 200 was ~1% against total protein for the WT protein (Figure 2a). The 75
corresponding area for the mutants increased to ~5%, ~9% and ~10%, for insG1, insG2 and insG3, respectively, indicating formation of more high-order oligomers with increase in insertion of Gly residues at the hinge loop. These results show that the length and flexibility of the hinge loop affect the size 80
aGraduate School of Materials Science, Nara Institute of Science and Technology, 8916-5 Takayama, Ikoma, Nara 630-0192, Japan. E-mail: [email protected]; Fax: (+81)-743-72-6119
bFaculty of Education, Kagawa University, 1-1 Saiwai, Takamatsu, Kagawa 760-8522, Japan.
cDepartment of Life Science, Graduate School of Life Science, University of Hyogo, 3-2-1 Koto, Kamigori-cho, Ako-gun, Hyogo 678-1297, Japan.
dRIKEN SPring-8 Center, 1-1-1 Koto, Sayo-cho, Sayo-gun, Hyogo 679-5148, Japan.
†Electronic supplementary information (ESI) available: Full experimental details, crystallographic data, chromatograms and spectra. See DOI: 10.1039/ b000000x
This journal is © The Royal Society of Chemistry [year] [journal], [year], [vol], 00–00 | 1

and amount of domain-swapped oligomers. In addition, almost no oligomers dissociated by incubation at 50 °C for 1 h, indicating relatively high thermostability of the oligomers (Figure S1). 5
10
15
Fig. 2. Size exclusion chromatograms of WT and mutant cyt c552 after 20
treatment with ethanol: (a) WT, (b) insG1 mutant, (c) insG2 mutant, and (d) insG3 mutant. After addition of ethanol up to 90% (v/v) to WT or mutant cyt c552 (500 μM) at 70 °C, the obtained precipitates were lyophilized, dissolved in buffer at 4 °C, and analysed with a Superdex 200 column. 25
The dimers exhibiting the peak with the second largest
elution volume in the size exclusion chromatogram obtained using Superdex 75 (Figure S2) were further purified with cation exchange chromatography (Figure S3). Two peaks were 30
observed in the cation exchange chromatograms of the WT and Gly-inserted mutant dimers, showing that two types of dimers existed for all the proteins investigated (Figure S3, curves a‒d). However, the two dimers of the WT protein could not be separated by cation exchange chromatography, since the two 35
dimer peaks were overlapped. The peaks of the two dimers separated more in the cation exchange chromatograms for the Gly-inserted mutants (Figure S3), and the two dimers (major and minor) were purified successfully for the insG3 mutant (Figure S3, curves c and d). The peak position of the insG3 40
major dimer in the cation exchange chromatogram was shifted from those of the insG3 minor dimer and WT dimers (Figure S3, curves e and f), presumably due to elongation of the hinge loop and change in the surface charge density of the insG3 major dimer compared to the WT dimers by the insertion of Gly 45
residues. Since significant amount of high-order oligomers were obtained and the two dimers were purified for the insG3 mutant, we investigated the dimers and oligomers of the insG3 mutant in more detail.
Most of the major and minor insG3 dimers did not dissociate 50
by incubation at 70 °C for 30 min (Figure S4), showing that the dimers were thermostable. However, the minor dimer dissociated by incubation at 80 oC for 30 min, whereas the major dimer did not dissociate by incubation at similar conditions (Figure S4). The maximum wavelength of the Soret band of 55
oxidized insG3 major and minor dimers was 410 nm (Figure S5A), which was similar to that of the oxidized WT monomer. The absorption coefficient of the Soret band for both major and minor dimers was obtained as 109,000 ± 1000 M−1 cm−1 (heme unit) by the pyridine hemochrome method.13 This value was also 60
similar to the absorption coefficients obtained for the WT monomer (109,000 ± 2000 M−1cm−1) and insG3 monomer (109,000 ± 1000 M−1cm−1), and the reported value for the WT monomer (105,000 M−1 cm−1 at 409.5 nm).14 In the circular dichroism spectra of oxidized insG3 major and minor dimers, 65
negative Cotton effects were observed at 208 and 222 nm, where the intensities were similar to those of the WT and insG3 monomers (Figure S5B). These results indicated that the active site and secondary structures were similar among the monomers and dimers. 70
The structure of the insG3 major dimer at 1.26 Å resolution exhibited a domain-swapped structure, where the N-terminal region containing the heme (up to Lys17) was exchanged between protomers (PDB ID: 5AUR) (Figure 3B). The swapped region in the insG3 major dimer was similar to that reported for 75
the WT dimer.10b The structure of the insG3 minor dimer at 1.30 Å resolution also exhibited a domain-swapped structure (PDB ID: 5AUS) (Figure 3C). Interestingly, the swapping region of the minor dimer was the C-terminal region (from Pro61), which was different from that of the WT and major dimers. Although 80
multiple domain swapping modes have been reported for several non-heme proteins,9, 15 the present result on cyt c552 is the first example for a heme protein to exhibit different modes of domain swapping, indicating that multiple mode domain swapping may also occur in heme proteins. 85
90
95
Fig. 3. Crystal structures of WT cyt c552 and its insG3 major and minor 100
dimers: (A) WT monomer (PDB ID: 1YNR, red), (B) insG3 major dimer (PDB ID: 5AUR, red and green) and (C) insG3 minor dimer (PDB ID: 5AUS, red and blue). The hemes are shown as stick models in pale colours. Side-chain atoms of His14 and Met62 (Met59 for the WT monomer) are shown as stick models. The inserted residues 105
(Gly19Gly20Gly21) are shown in pink. The pink arrow in the WT monomer structure represents the insertion position. The hinge loops of the major dimer are shown in orange and pink. The hinge loops of the minor dimer are shown in cyan. The dotted lines represent the residues with electron densities not detected. 110
The protein structures of the insG3 dimers corresponded
well to that of the WT monomer (Figure S6). We calculated the root-mean-square deviation (rmsd) values for the Cα atoms between the structures of the insG3 major (minor) dimer and the 115
WT monomer (four molecules in the asymmetric unit) (Table S1). Residues in the N-terminal region before the hinge loop of one protomer and residues in the C-terminal region after the hinge loop in the other protomer in the same insG3 major (minor) dimer were compared with the corresponding structural 120
region of the monomer. The rmsd values of the major and minor
2 | Journal Name, [year], [vol], 00–00 This journal is © The Royal Society of Chemistry [year]

dimers were 0.25−0.35 Å and 0.32−0.45 Å, respectively. These results indicate that the structures were similar between the WT monomer and the protomers of the insG3 major and minor dimers. For the insG3 minor dimer, Gly19‒Lys22 were undetectable in a protomer structure, due to their flexibility, and 5
excluded for the calculations. The heme iron coordination structure (Met−Fe−His) was
similar among the WT monomer and both the insG3 major and minor dimers (Figure 4). Met62 coordinated to the heme in both insG3 dimers, but it originated from the other protomer to which 10
the heme belonged. The Fe−His14 distance of the insG3 major and minor dimers was 2.04−2.07 Å and 2.04−2.05 Å, respectively, whereas the Fe−Met62 distance was 2.25−2.39 Å and 2.34−2.35 Å, respectively. These distances were similar to the corresponding distances in the WT monomer (Fe−His14, 15
2.05−2.09 Å; Fe−Met59, 2.33−2.40 Å) (Table S2). These results were consistent with the absorption spectra (Figure S5A), indicating that the active site structure was similar among the monomer and both dimers. 20
Fig. 4. Active site structures of WT cyt c552 and its insG3 major and minor dimers: (A) WT monomer (red), (B) insG3 major dimer (PDB ID: 5AUR) and (C) insG3 minor dimer (PDB ID: 5AUS). The hemes are 25
shown in pink stick models. The red and green (or blue) strands in the insG3 major (or minor) dimer structure are regions from different protomers. Side-chain atoms of His14 and Met62 (Met59 for the monomer) are shown as stick models.
30
Negative staining transmission electron microscopy (TEM) was performed to investigate the morphology of insG3 oligomers from approximately the octamer to pentadecamer, purified by size exclusion chromatography. Unfortunately, we could not observe TEM images for isolated insG3 oligomers. 35
We speculated that the hydrophobic property of the insG3 mutant inhibited the interaction of the protein with the staining compound, phosphotungstic acid, and thus replaced Trp57 of the insG3 mutant with Lys (insG3/W57K mutant). The insG3/W57K mutant formed high-order oligomers similar to the 40
insG3 mutant by treatment with ethanol (Figure S7). The TEM image of oligomeric insG3/W57K mutant (from octamer to pentadecamer) exhibited well-separated protein ring structures at 7–10 nm in diameter (Figure 5). Although the size of the ring structures was at the resolution limit, observation of the ring 45
images were reproducible (Figure S8). The region containing the N-terminal α-helix and heme is hydrophobic and forms a well-organized structure in cyt c552, and thus this region would be unstable in aqua solution and may bind to the rest of the protein, resulting in formation of a ring-structured oligomer. Linear 50
structures may also exist in the oligomers, but were undetectable, since the size of HT cyt c552 (~2 nm in diameter) was too small. Many double-ring structures were also observed in the TEM image. Major N-terminal and minor C-terminal domain swapping were observed in the insG3 dimer structures (Figure 55
3). By the C-terminal domain swapping, it is difficult to form high-order oligomers, since its hinge loop was constructed with only three residues, similar to the WT protein (Figure 2). Therefore, Gly-inserted cyt c552 molecules may domain swap the N-terminal region successively and form a ring structure, 60
whereas the ring structures may be connected by domain swapping the C-terminal region. Construction of a protein nanoring has been reported by introducing specific interactions, such as metal ion chelation and chemical cross-linking.1d, 2a We constructed protein oligomers with a thermostable protein using 65
domain swapping. Especially, the protein structure, including the active site, of the monomer is maintained in the oligomers produced by domain swapping. These results indicate that relatively strong interactions are necessary to obtain stable protein nanostructures, and domain swapping may be useful to 70
control interaction between protein molecules and construct functional protein oligomers. 75
80
Fig. 5. Negative staining TEM image of insG3/W57K cyt c552 85
oligomers: (A) Overall view, expanded views of (B) single- and (C) double-ring structures.
In summary, we have succeeded in increasing the amount of
high-order domain-swapped oligomers of thermostable HT cyt 90
c552 by inserting Gly residues into the hinge loop. Major and minor insG3 dimers exhibited different domain-swapped structures, where the N-terminal region was exchanged between protomers for the major dimer and the C-terminal region for the minor dimer. Single- and double-ring structures were observed 95
for the oligomers. These results show that domain swapping can be applied to construct protein nanostructures, providing a new design strategy for the construction of protein assemblies.
The authors thank Ms. Sakiko Fujita and Mr. Masahiro Fujihara, Nara Institute of Science and Technology (NAIST), 100
for their technical assistance on TEM observation, and Mr. Leigh McDowell, NAIST, for his advice on manuscript preparation. The synchrotron radiation experiments were performed at the BL38B1 and BL44XU beamlines of SPring-8 with the approval of JASRI (No. 2014B1438 (S.N.) and No. 105
2015A6999 (H.K.)). The authors are grateful to Prof. Yoshihiro Sambongi, Hiroshima University, for a kind gift of pKO2 and pEC86 plasmid DNAs. This work was partially supported by Grants-in-Aid for Scientific Research (Category B, No. 26288080; S.H.) and Challenging Exploratory Research (No. 110
15K13744; S.H.) from JSPS.
Notes and references
This journal is © The Royal Society of Chemistry [year] Journal Name, [year], [vol], 00–00 | 3

1. (a) I. Medalsy, O. Dgany, M. Sowwan, H. Cohen, A. Yukashevska,
S. G. Wolf, A. Wolf, A. Koster, O. Almog, I. Marton, Y. Pouny, A. Altman, O. Shoseyov and D. Porath, Nano Lett., 2008, 8, 473–477; (b) A. D. Malay, J. G. Heddle, S. Tomita, K. Iwasaki, N. Miyazaki, K. Sumitomo, H. Yanagi, I. Yamashita and Y. Uraoka, Nano Lett., 5
2012, 12, 2056–2059; (c) W. Zhang, Q. Luo, L. Miao, C. Hou, Y. Bai, Z. Dong, J. Xu and J. Liu, Nanoscale, 2012, 4, 5847–5851; (d) Y. Bai, Q. Luo, W. Zhang, L. Miao, J. Xu, H. Li and J. Liu, J. Am. Chem. Soc., 2013, 135, 10966–10969.
2. (a) J. C. T. Carlson, S. S. Jena, M. Flenniken, T.-f. Chou, R. A. 10
Siegel and C. R. Wagner, J. Am. Chem. Soc., 2006, 128, 7630–7638; (b) H. Kitagishi, K. Oohora, H. Yamaguchi, H. Sato, T. Matsuo, A. Harada and T. Hayashi, J. Am. Chem. Soc., 2007, 129, 10326–10327; (c) K. Oohora, S. Burazerovic, A. Onoda, Y. M. Wilson, T. R. Ward and T. Hayashi, Angew. Chem. Int. Ed., 2012, 15
51, 3818–3821. 3. C. Hou, J. Li, L. Zhao, W. Zhang, Q. Luo, Z. Dong, J. Xu and J. Liu,
Angew. Chem., Int. Ed., 2013, 52, 5590–5593. 4. (a) M. J. Boerakker, J. M. Hannink, P. H. H. Bomans, P. M.
Frederik, R. J. M. Nolte, E. M. Meijer and N. A. J. M. Sommerdijk, 20
Angew. Chem. Int. Ed., 2002, 41, 4239–4241; (b) M. J. Boerakker, N. E. Botterhuis, P. H. H. Bomans, P. M. Frederik, E. M. Meijer, R. J. M. Nolte and N. A. J. M. Sommerdijk, Chem. Eur. J., 2006, 12, 6071–6080.
5. D. P. Patterson, M. Su, T. M. Franzmann, A. Sciore, G. Skiniotis 25
and E. N. G. Marsh, Protein Sci., 2014, 23, 190–199. 6. (a) Y.-T. Lai, K.-L. Tsai, M. R. Sawaya, F. J. Asturias and T. O.
Yeates, J. Am. Chem. Soc., 2013, 135, 7738–7743; (b) L. Miao, J. Han, H. Zhang, L. Zhao, C. Si, X. Zhang, C. Hou, Q. Luo, J. Xu and J. Liu, ACS Nano, 2014, 8, 3743–3751; (c) S. Sim, D. Miyajima, T. 30
Niwa, H. Taguchi and T. Aida, J. Am. Chem. Soc., 2015, 137, 4658–4661.
7. J.-H. Jeoung, D. A. Pippig, B. M. Martins, N. Wagener and H. Dobbek, J. Mol. Biol., 2007, 368, 1122–1131.
8. M. J. Bennett, S. Choe and D. Eisenberg, Proc. Natl. Acad. Sci. 35
U.S.A., 1994, 91, 3127–3131. 9. (a) Y. Liu, P. J. Hart, M. P. Schlunegger and D. Eisenberg, Proc.
Natl. Acad. Sci. U.S.A., 1998, 95, 3437–3442; (b) Y. Liu, G. Gotte, M. Libonati and D. Eisenberg, Nat. Struct. Mol. Biol., 2001, 8, 211–214. 40
10. (a) S. Hirota, Y. Hattori, S. Nagao, M. Taketa, H. Komori, H. Kamikubo, Z. Wang, I. Takahashi, S. Negi, Y. Sugiura, M. Kataoka and Y. Higuchi, Proc. Natl. Acad. Sci. U.S.A., 2010, 107, 12854–12859; (b) Y. Hayashi, S. Nagao, H. Osuka, H. Komori, Y. Higuchi and S. Hirota, Biochemistry, 2012, 51, 8608–8616; (c) S. Nagao, H. 45
Osuka, T. Yamada, T. Uni, Y. Shomura, K. Imai, Y. Higuchi and S. Hirota, Dalton Trans., 2012, 41, 11378–11385; (d) Y.-W. Lin, S. Nagao, M. Zhang, Y. Shomura, Y. Higuchi and S. Hirota, Angew. Chem. Int. Ed., 2015, 54, 511–515; (e) S. Nagao, M. Ueda, H. Osuka, H. Komori, H. Kamikubo, M. Kataoka, Y. Higuchi and S. 50
Hirota, PLoS ONE, 2015, 10, e0123653; (f) M. Yamanaka, S. Nagao, H. Komori, Y. Higuchi and S. Hirota, Protein Sci., 2015, 24, 366–375.
11. C. Travaglini-Allocatelli, S. Gianni, V. K. Dubey, A. Borgia, A. Di Matteo, D. Bonivento, F. Cutruzzolà, K. L. Bren and M. Brunori, J. 55
Biol. Chem., 2005, 280, 25729–25734. 12. Y. Yamamoto, N. Terui, N. Tachiiri, K. Minakawa, H. Matsuo, T.
Kameda, J. Hasegawa, Y. Sambongi, S. Uchiyama, Y. Kobayashi and Y. Igarashi, J. Am. Chem. Soc., 2002, 124, 11574–11575.
13. E. A. Berry and B. L. Trumpower, Anal. Biochem., 1987, 161, 1–15. 60
14. X. Wen and K. L. Bren, Biochemistry, 2005, 44, 5225–5233. 15. (a) Y. Liu, G. Gotte, M. Libonati and D. Eisenberg, Protein Sci.,
2002, 11, 371–380; (b) I.-J. L. Byeon, J. M. Louis and A. M. Gronenborn, J. Mol. Biol., 2003, 333, 141–152; (c) M. A. Schumacher, M. Crum and M. C. Miller, Structure, 2004, 12, 849–65
860; (d) K.-E. Chen, A. A. Richards, J. K. Ariffin, I. L. Ross, M. J. Sweet, S. Kellie, B. Kobe and J. L. Martin, Acta Crystallogr. D, 2012, 68, 637–648.
4 | Journal Name, [year], [vol], 00–00 This journal is © The Royal Society of Chemistry [year]

S1
Supporting Information Oligomerization Enhancement and Two Domain Swapping Mode Detection for Thermostable Cytochrome c552 by Elongation of the Major Hinge Loop Chunguang Ren,a Satoshi Nagao,a Masaru Yamanaka,a Hirofumi Komori,b Yasuhito Shomura,c,d Yoshiki Higuchi,c,d and Shun Hirota*a aGraduate School of Materials Science, Nara Institute of Science and Technology, 8916-5 Takayama, Ikoma, Nara 630-0192, Japan bFaculty of Education, Kagawa University, 1-1 Saiwai, Takamatsu, Kagawa 760-8522, Japan. cDepartment of Life Science, Graduate School of Life Science, University of Hyogo, 3-2-1 Koto, Kamigori-cho, Ako-gun, Hyogo 678-1297, Japan. dRIKEN SPring-8 Center, 1-1-1 Koto, Sayo-cho, Sayo-gun, Hyogo 679-5148, Japan.
Contents
Materials and Methods p. S2 Table S1. Root-mean-square deviation values between the protomer of p. S5 insG3 cyt c552 dimers and WT cyt c552 monomer. Table S2. Fe−His14 and Fe−Met62 (Met59 for the WT cyt c552 monomer) p. S6
distances of WT cyt c552 monomer and insG3 cyt c552 dimers. Table S3. Nucleotide sequences of the primers. p. S7 Table S4. Statistics of data collection and structure refinement of the insG3 p. S8 cyt c552 major dimer. Table S5. Statistics of data collection and structure refinement of the insG3 p. S9 cyt c552 minor dimer. Figure S1. Size exclusion chromatograms of insG3 cyt c552 oligomers with p. S10 and without incubation. Figure S2. Size exclusion chromatograms of WT and mutant cyt c552. p. S11 Figure S3. Cation exchange chromatograms of WT and mutant cyt c552 dimers. p. S12 Figure S4. Size exclusion chromatograms of insG3 cyt c552 dimers after p. S13 incubation at various temperatures. Figure S5. Optical absorption and CD spectra of oxidized WT cyt c552 monomer, p. S14 insG3 cyt c552 monomer and insG3 cyt c552 dimers. Figure S6. Overlapped view of the structures of the WT cyt c552 monomer p. S15 and insG3 cyt c552 dimers. Figure S7. Size exclusion chromatograms of insG3 and insG3/W57K cyt c552. p. S16 Figure S8. TEM image of insG3/W57K cyt c552 oligomers. p. S17

S2
Materials and Methods
Plasmids. Plasmid pKO2 was used for expression of Hydrogenobacter thermophilus (HT)
cyt c552.1 Escherichia coli strain DH5α was used for routine gene engineering. Gly residues
were inserted into the sequence between Ala18 and Lys19 of HT cyt c552 (insG1, insG2, and
insG3). Trp57 of the insG3 mutant was substituted with Lys (insG3/W57K). KOD-Plus-DNA
Polymerase (Toyobo, Osaka, Japan) with forward and reverse primers (Sigma-Aldrich, St
Louis, MO, USA) (Table S3, Supporting Information) was used for PCR-based mutagenesis
of the pKO2 plasmid. DNA sequencing of mutant plasmids were conducted with a BigDye
Terminator version 3.1 cycle sequencing kit (Applied Biosystems, Inc., Foster City, CA) and
an ABI PRISM 310 genetic analyzer sequencing system (Applied Biosystems, Inc.). The
obtained plasmids were introduced into competent Escherichia coli JCB387 cells.
Protein Purification and Oligomerization. Wild-type (WT) HT cyt c552 and its mutants
were expressed in Escherichia coli and purified as reported previously.2 The oxidized form of
WT and mutant HT cyt c552 were prepared by an addition of potassium ferricyanide.
Oxidized oligomeric WT and mutant HT cyt c552 were prepared by dissolving oxidized WT,
insG3 or inG3/W57K HT cyt c552 (1 mM) in 50 mM potassium phosphate buffer, pH 7.0,
followed by an addition of ethanol (final concentration, 90% (v/v)) at 70 °C. After
centrifugation of the solution at 8,000 g for 1 h, the precipitate was lyophilized to remove
residual ethanol. The lyophilized precipitate was dissolved in 50 mM potassium phosphate
buffer, pH 7.0, at 4 °C.
The oxidized insG3 mutant oligomers of HT cyt c552 were loaded to a gel filtration column
(HiLoad 26/60 Superdex 75, GE Healthcare) with 50 mM potassium phosphate buffer, pH 7.0,
using a fast protein liquid chromatography (FPLC) system (BioLogic DuoFlow 10, Bio-Rad,
CA) at 4 °C to obtain the insG3 dimers. The insG3 dimers were purified with a Mono STM
5/50 GL cation exchange column using the FPLC system (BioLogic DuoFlow 10, Bio-Rad)
equilibrated with 10 mM potassium phosphate buffer, pH 7.0, at a flow rate of 0.5 mL/min.
The insG3 dimers were eluted with a gradient of 0 to 250 mM Na2SO4 over a period of 240
min. High-order oligomers of insG3 and insG3/W57K mutants were purified with a HiLoad
26/600 Superdex 200 column (GE Healthcare) using the FPLC system (BioLogic DuoFlow
10, Bio-Rad).
The absorption coefficients of the oxidized insG3 monomer and dimers were obtained with
the pyridine hemochrome method (ε = 109,000 ± 1000 M−1cm−1 at 410 nm for the monomer
and dimers).3 The concentrations of the proteins were calculated from the absorbance at 410
nm with this absorption coefficient, and adjusted to desired concentrations.
Size Exclusion Chromatography Analysis. The oligomeric HT cyt c552 solutions were
analyzed by size exclusion chromatography (column: Superdex 75 10/300 GL, GE healthcare)
using the FPLC system (BioLogic DuoFlow 10, Bio-Rad) with 50 mM potassium phosphate
buffer, pH 7.0, at 4 °C. The amount of oligomers was estimated by integration of the peak

S3
areas in the elution curves using the program Igor Pro 6.0.
Optical Absorption and Circular Dichroism Measurements. Optical absorption spectra
were measured with a UV-2450 spectrophotometer (Shimadzu, Japan) using a
1-cm-pathlength quartz cell at room temperature. Circular dichroism (CD) spectra were
measured with a J-725 CD spectropolarimeter (Jasco, Japan) using a 0.1-cm-path-length
quartz cell at 25 °C.
X-ray Crystallographic Analysis. Crystallization of HT cyt c552 insG3 major and minor
dimers was carried out at 277 K using the sitting drop vapor diffusion method with crystal
plates (CrystalClear D Strips, Douglas Instruments, Hampton Research, CA). Oxidized insG3
major dimer was dissolved in 10 mM Tris-HCl buffer, pH 7.0, at a protein concentration of
15.4 mg/ml. Oxidized insG3 minor dimer was dissolved in 10 mM HEPES buffer, pH 7.0, at a
protein concentration of 22.1 mg/ml. Droplets prepared by mixing 1 μl (2μl for the insG3
minor dimer) of the insG3 major dimer solution with 1 μl (2 μl for the insG3 minor dimer)
reservoir solution were equilibrated. The best reservoir solution for the insG3 major dimer
was found to be 200 mM potassium iodide, containing 15% w/v PEG 3,350. The best
reservoir solution for the insG3 minor dimer was 100 mM Tris-HCl, pH 8.5, containing 200
mM sodium acetate and 30% w/v PEG 4,000. Crystals were observed after incubation at 277
K for four and three weeks for the insG3 major and minor dimer, respectively.
The diffraction data were collected at BL44XU and BL38B1 beamlines at SPring-8, Japan,
for the insG3 major and minor dimer, respectively. The crystal of the insG3 major dimer
treated with a cryoprotectant (200 mM potassium iodide and 40% w/v PEG 3,350) was
mounted on a cryo-loop, and flash-frozen at 100 K in a nitrogen cryo system. The crystal of
insG3 minor dimer without a treatment of a cryoprotectant was mounted on a cryo-loop and
flash-frozen in liquid nitrogen. The detector was MX300HE and MX225HE (Rayonix) for the
insG3 major and minor dimer, respectively. The crystal-to-detector distance was 150 mm and
95 mm for the insG3 major and minor dimer, respectively. The wavelength was 0.9000 Å and
1.0000 Å for the insG3 major and minor dimer, respectively. The oscillation angle was 1.0 °
and 0.5 °, and the exposure time was 0.5 s and 2.0 s per frame for the insG3 major and minor
dimer, respectively. The total number of frames was 180 and 200 for the insG3 major and
minor dimer, respectively. The diffraction data were processed using the program HKL20004
for the insG3 major dimer, and the program iMosflm5 and Scala6 for the insG3 minor dimer.
The preliminary structure was obtained by a molecular replacement method (MOLREP)
using the atomic coordinates of the structure of monomeric HT cyt c552 (PDB code: 1YNR) as
a starting model for both dimers. There were two and one dimers in the asymmetric unit of the
crystal for the major and minor dimer, respectively. The Gly19–Lys22 residues of one
protomer of the minor dimer were not able to place their model in the structure of the insG3
minor dimer, due to the unclear electron density. The structure refinement was performed
using the program, REFMAC.7 The molecular model was manually corrected, and water

S4
molecules were picked up in the electron density map using the program, COOT.8 The data
collection and refinement statistics of the major and minor dimers are summarized in Tables
S4 and S5, Supporting Information, respectively.
Transmission Electron Microscopy. A 2 µl aliquot of oxidized oligomeric HT cyt c552
insG3/W57K mutant (2 µM, heme unit) dissolved in water was spread onto a carbon-coated
200-mesh copper grid (1606, JEOL) and incubated at room temperature for 10 min. After
removing the solution with a filter paper, the oligomers were stained by an addition of 5%
(w/w) phosphotungstic acid, pH 7.0 (adjusted by 1 M NaOH), at room temperature for 10 min.
The phosphotungstic acid solution was removed with a filter paper. After repeating the
addition and removal of phosphotungstic acid twice more, the sample was vacuumed 1 h in a
desiccator. The transmission electron microscope (TEM) image of oligomeric HT cyt c552
insG3/W57K mutant was taken at 18 °C with a JEM-3100FEF microscope (JEOL) at 300 kV.
References
1. K. Oikawa, S. Nakamura, T. Sonoyama, A. Ohshima, Y. Kobayashi, S.-I. J. Takayama, Y.
Yamamoto, S. Uchiyama, J. Hasegawa and Y. Sambongi, J. Biol. Chem., 2005, 280, 5527‒
5532.
2. (a) J. Hasegawa, S. Uchiyama, Y. Tanimoto, M. Mizutani, Y. Kobayashi, Y. Sambongi and
Y. Igarashi, J. Biol. Chem., 2000, 275, 37824‒37828; (b) J. Hasegawa, H. Shimahara, M.
Mizutani, S. Uchiyama, H. Arai, M. Ishii, Y. Kobayashi, S. J. Ferguson, Y. Sambongi and Y.
Igarashi, J. Biol. Chem., 1999, 274, 37533‒37537.
3. E. A. Berry and B. L. Trumpower, Anal. Biochem., 1987, 161, 1‒15.
4. Z. Otwinowski and W. Minor, Methods Enzymol., 1997, 276, 307-326.
5. A. W. Leslie and H. Powell, Evolving Methods for Macromolecular Crystallography, 2007,
245, 41‒51.
6. M. D. Winn, C. C. Ballard, K. D. Cowtan, E. J. Dodson, P. Emsley, P. R. Evans, R. M.
Keegan, E. B. Krissinel, A. G. W. Leslie, A. McCoy, S. J. McNicholas, G. N. Murshudov,
N. S. Pannu, E. A. Potterton, H. R. Powell, R. J. Read, A. Vagin and K. S. Wilson, Acta
Crystallogr D, 2011, 67, 235‒242.
7. A. T. Brünger, P. D. Adams, G. M. Clore, W. L. DeLano, P. Gros, R. W. Grosse-Kunstleve,
J.-S. Jiang, J. Kuszewski, M. Nilges, N. S. Pannu, R. J. Read, L. M. Rice, T. Simonson and
G. L. Warren, Acta Crystallogr D, 1998, 54, 905‒921.
8. P. Emsley and K. Cowtan, Acta Crystallogr D, 2004, 60, 2126‒2132.

S5
Table S1. Root-mean-square deviation (rmsd) values between the Cα atoms of the protomers
of insG3 cyt c552 dimers and WT cyt c552 monomer.a
RMSD (Å) Major dimerb 0.25−0.35 Minor dimerc 0.32−0.45
aThere are four independent cyt c552 molecules in the asymmetric unit of the WT monomeric
cyt c552 crystal, whereas there are four and two protomers in the asymmetric unit of the insG3
major and minor dimeric cyt c552 crystal, respectively. bPDB ID: 5AUR. Residues 1−17 of one protomer and residues 24−83 in the other protomer in
the same dimer were compared with the corresponding structural region of the monomer. The
hinge loop (Ala18−Lys23) was excluded from the calculation. cPDB ID: 5AUS. Residues 1−57 of one protomer and residues 61−83 in the other protomer in
the same dimer were compared with the corresponding structural region of the monomer. The
hinge loop (Gly58−Val60) and Gly19−Gly21 were excluded from the calculation. Lys22 with
unclear electron density of one protomer was also excluded.

S6
Table S2. Fe−His14 and Fe−Met62 (Met59 for the WT cyt c552 monomer) distances of the
WT cyt c552 monomer and insG3 cyt c552 dimers.a
aThere are four independent cyt c552 molecules in the asymmetric unit of the WT monomeric cyt c552 crystal. There are four and two protomers in the asymmetric unit of the insG3 major and minor dimeric cyt c552 crystal, respectively. bPDB ID: 1YNR. There are four independent HT cyt c552 molecules in the asymmetric unit of the monomeric cyt c552 crystal. cPDB ID: 3VYM. There is one HT cyt c552 molecule in the asymmetric unit of the WT dimeric cyt c552 crystal. dPDB ID: 5AUR. There are four independent HT cyt c552 molecules in the asymmetric unit of the insG3 major dimeric cyt c552 crystal. ePDB ID: 5AUS. There are two independent HT cyt c552 molecules in the asymmetric unit of
the insG3 minor dimeric cyt c552 crystal.
Fe−His14 (Å) Fe−Met62(Met59) (Å) Wild-type monomerb 2.05‒2.09 2.33‒2.40
Wild-type dimerc 1.89 2.28 insG3 major dimerd 2.04‒2.07 2.25‒2.39 insG3 minor dimere 2.04‒2.05 2.34‒2.35

S7
Table S3. Nucleotide sequences of the primers.
Primer Sequencea insG1-F GGCAAGAAGGTGGGACCTGCTTACGCAG insG1-R AGCTTTCAGATCGTGGCAAGCCATA insG2-F GGAGGCAAGAAGGTGGGACCTGCTTACGCAG insG2-R AGCTTTCAGATCGTGGCAAGCCATA insG3-F GGTGGAGGCAAGAAGGTGGGACCTGCTTACGCAG insG3-R AGCTTTCAGATCGTGGCAAGCCATA
insG3/W57K-F AAGGGTTCTGTTCCCATGCCTCCTCAAAAT insG3/W57K-R CACACCAGAACCGCCCTTCTTTATCTT
aUnderlines indicate the nucleotides for the inserted or modified amino acids.
S8
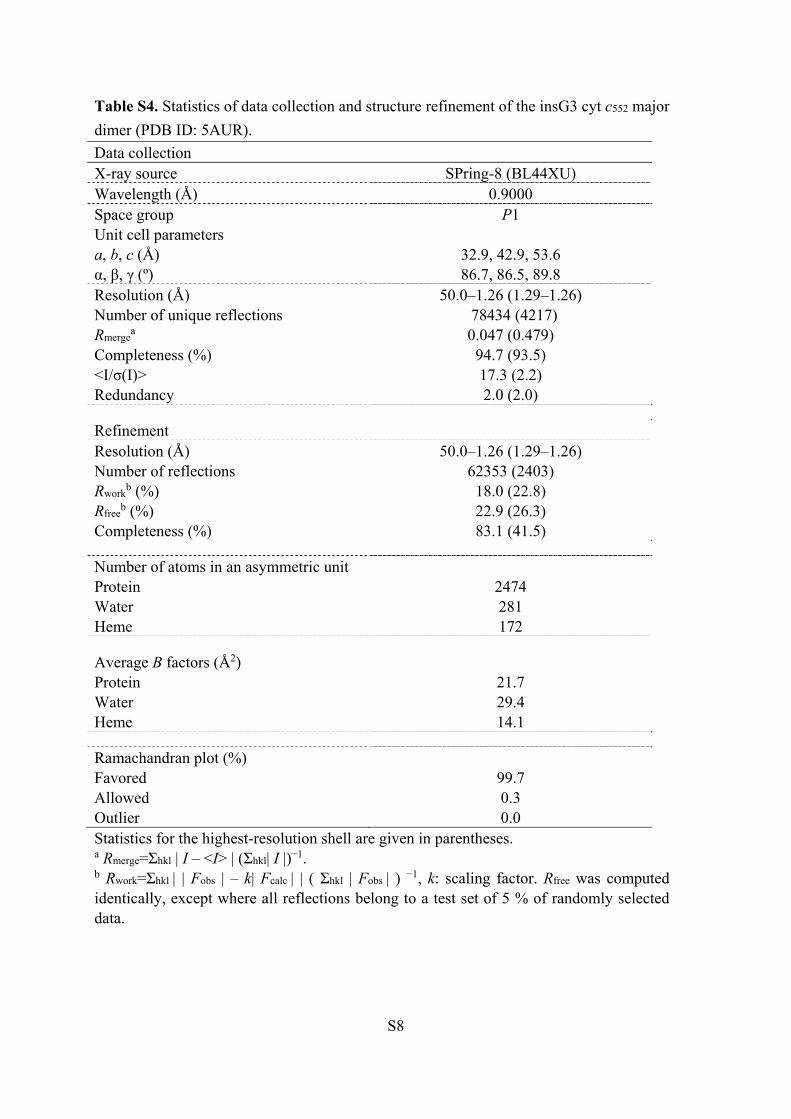
Table S4. Statistics of data collection and structure refinement of the insG3 cyt c552 major
dimer (PDB ID: 5AUR).
Data collection X-ray source SPring-8 (BL44XU) Wavelength (Å) 0.9000 Space group P1 Unit cell parameters a, b, c (Å) 32.9, 42.9, 53.6 α, β, γ (º) 86.7, 86.5, 89.8 Resolution (Å) 50.0–1.26 (1.29–1.26) Number of unique reflections 78434 (4217) Rmerge
a 0.047 (0.479) Completeness (%) 94.7 (93.5) <I/σ(I)> 17.3 (2.2) Redundancy 2.0 (2.0)
Refinement Resolution (Å) 50.0–1.26 (1.29–1.26) Number of reflections 62353 (2403) Rwork
b (%) 18.0 (22.8) Rfree
b (%) 22.9 (26.3) Completeness (%) 83.1 (41.5)
Number of atoms in an asymmetric unit Protein 2474 Water 281 Heme 172
Average B factors (Å2) Protein 21.7 Water 29.4 Heme 14.1
Ramachandran plot (%) Favored 99.7 Allowed 0.3 Outlier 0.0 Statistics for the highest-resolution shell are given in parentheses. a Rmerge=Σhkl | I – <I> | (Σhkl| I |)−1. b Rwork=Σhkl | | Fobs | – k| Fcalc | | ( Σhkl | Fobs | ) −1, k: scaling factor. Rfree was computed identically, except where all reflections belong to a test set of 5 % of randomly selected data.

S9
Table S5. Statistics of data collection and structure refinement of the insG3 cyt c552 minor dimer (PDB ID: 5AUS). Data collection X-ray source SPring-8 (BL38B1) Wavelength (Å) 1.0000 Space group C2221 Unit cell parameters a, b, c (Å) 71.0, 71.0, 62.1 α, β, γ (º) 90.0, 90.0, 90.0 Resolution (Å) 30.0–1.30 (1.37–1.30) Number of unique reflections 38911 (5621) Rmerge
a 0.113 (0.369) Completeness (%) 100.0 (100.0) <I/σ(I)> 6.8 (2.9) Redundancy 3.9 (3.9)
Refinement Resolution (Å) 30.0–1.30 (1.33–1.30) Number of reflections 37017 (2843) Rwork
b (%) 17.1 (20.0) Rfree
b (%) 20.2 (21.6) Completeness (%) 99.9 (99.9)
Number of atoms in an asymmetric unit Protein 1251 Water 216 Heme 86
Average B factors (Å2) Protein 9.9 Water 18.6 Heme 5.6
Ramachandran plot (%) Favored 99.3 Allowed 0.7 Outlier 0.0 Statistics for the highest-resolution shell are given in parentheses. a Rmerge=Σhkl | I – <I> | (Σhkl| I |)−1. b Rwork=Σhkl | | Fobs | – k| Fcalc | | ( Σhkl | Fobs | ) −1, k: scaling factor. Rfree was computed identically, except where all reflections belong to a test set of 5 % of randomly selected data.

S10
Figure S1. Size exclusion chromatograms of insG3 cyt c552 oligomers with and without
incubation. (a) Without incubation. (b) After incubation at 50 °C for 1 h. Measurement
conditions: column, Superdex 75 10/300 GL; flow rate, 0.5 mL/min; monitoring wavelength,
410 nm; solvent, 50 mM potassium phosphate buffer; pH, 7.0; temperature, 4 °C.
Abs
orb
ance
161284Elution volume (mL)
b
a
S11
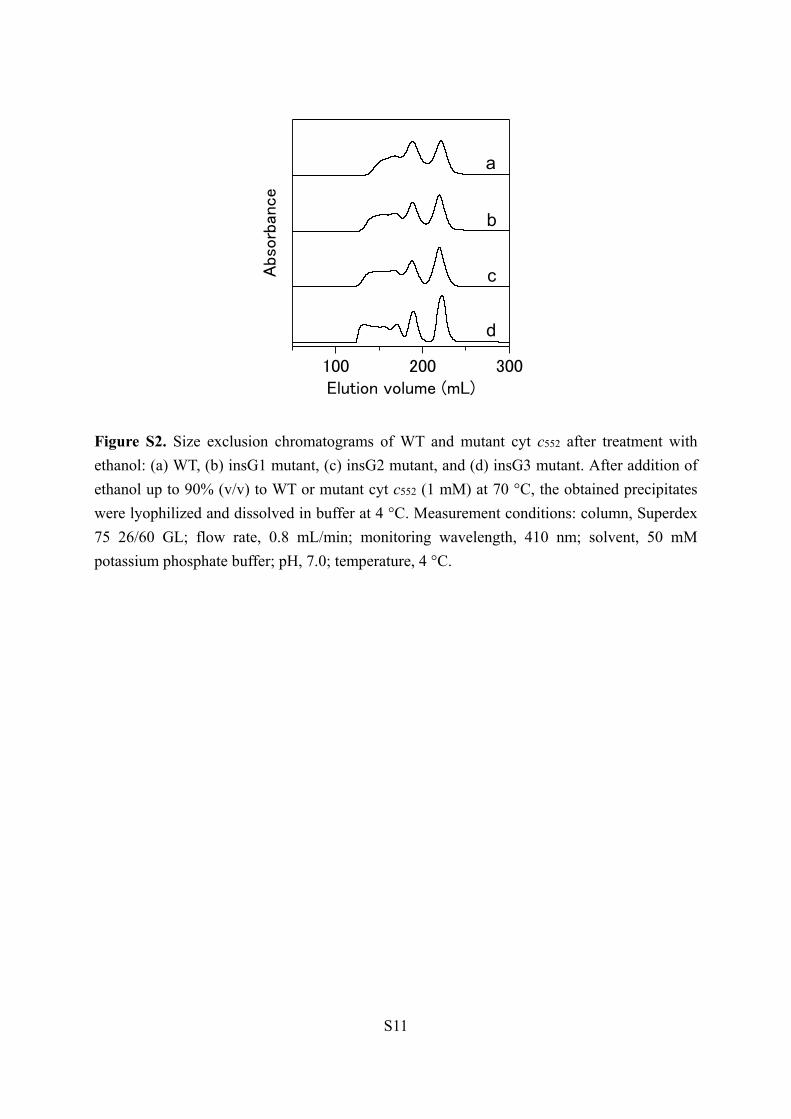
Figure S2. Size exclusion chromatograms of WT and mutant cyt c552 after treatment with
ethanol: (a) WT, (b) insG1 mutant, (c) insG2 mutant, and (d) insG3 mutant. After addition of
ethanol up to 90% (v/v) to WT or mutant cyt c552 (1 mM) at 70 °C, the obtained precipitates
were lyophilized and dissolved in buffer at 4 °C. Measurement conditions: column, Superdex
75 26/60 GL; flow rate, 0.8 mL/min; monitoring wavelength, 410 nm; solvent, 50 mM
potassium phosphate buffer; pH, 7.0; temperature, 4 °C.
Abs
orb
ance
300200100Elution volume (mL)
a
b
c
d
S12
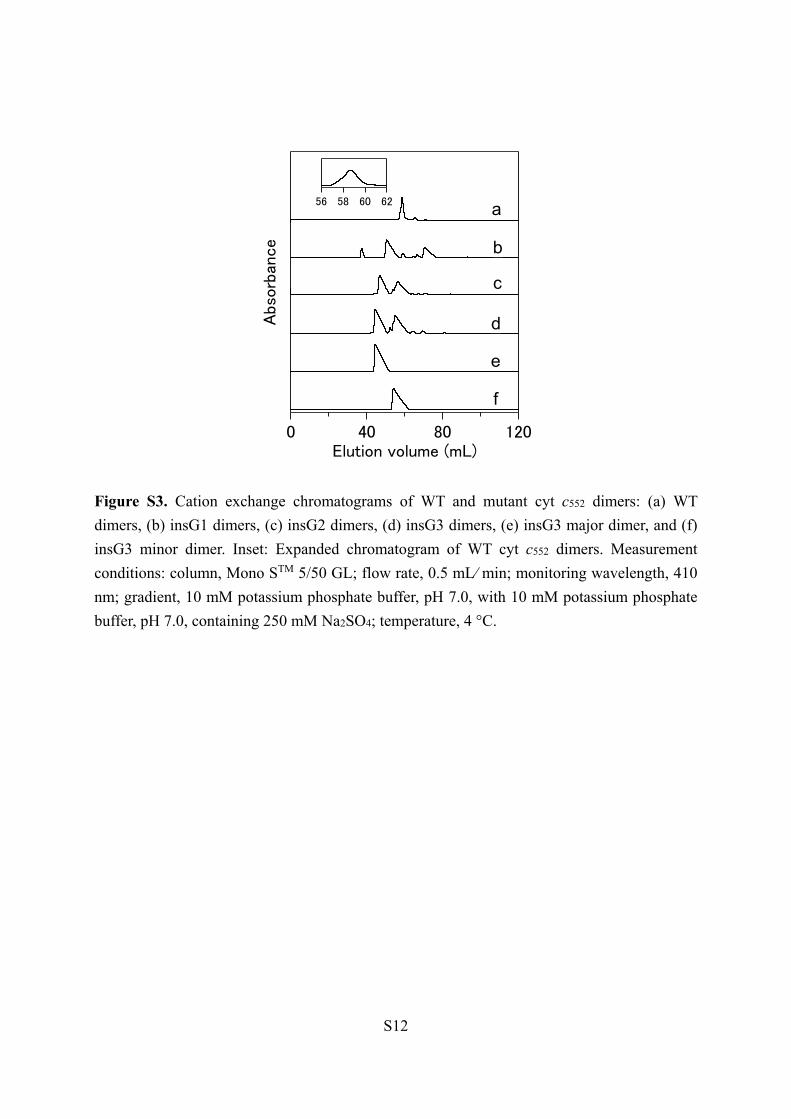
Figure S3. Cation exchange chromatograms of WT and mutant cyt c552 dimers: (a) WT
dimers, (b) insG1 dimers, (c) insG2 dimers, (d) insG3 dimers, (e) insG3 major dimer, and (f)
insG3 minor dimer. Inset: Expanded chromatogram of WT cyt c552 dimers. Measurement
conditions: column, Mono STM 5/50 GL; flow rate, 0.5 mL⁄ min; monitoring wavelength, 410
nm; gradient, 10 mM potassium phosphate buffer, pH 7.0, with 10 mM potassium phosphate
buffer, pH 7.0, containing 250 mM Na2SO4; temperature, 4 °C.
Abso
rban
ce
12080400Elution volume (mL)
62605856 a
b
c
d
e
f
S13
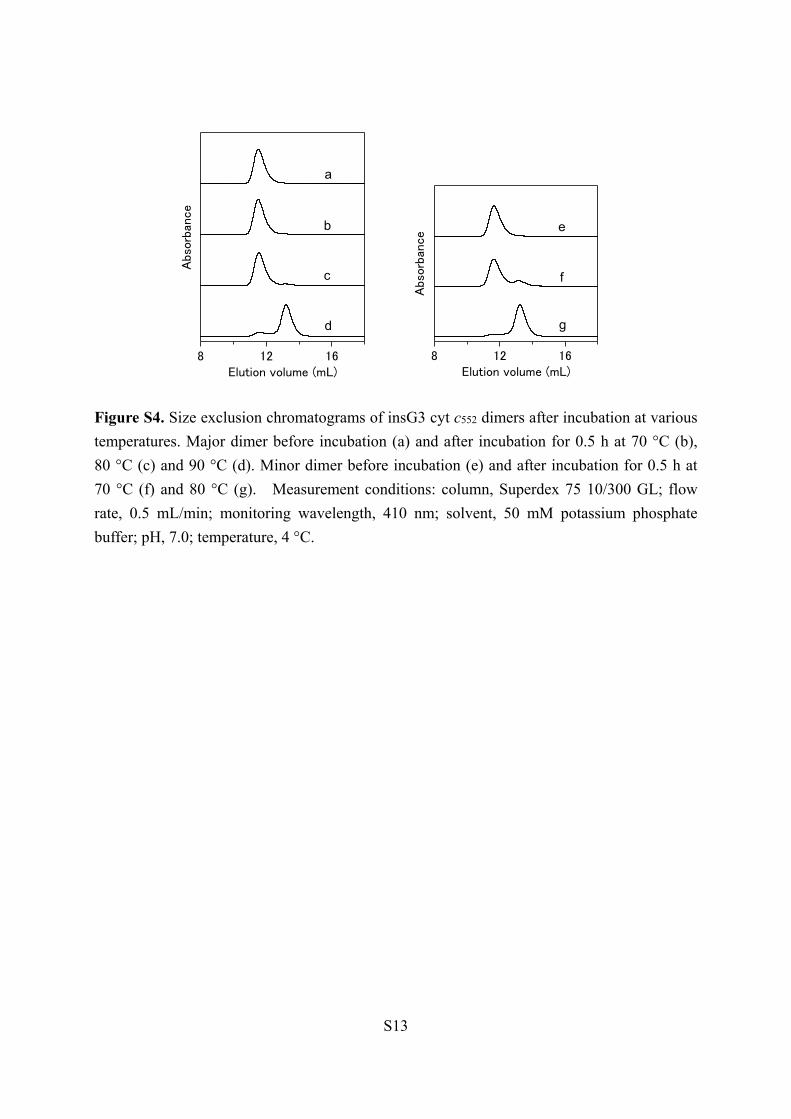
Figure S4. Size exclusion chromatograms of insG3 cyt c552 dimers after incubation at various
temperatures. Major dimer before incubation (a) and after incubation for 0.5 h at 70 °C (b),
80 °C (c) and 90 °C (d). Minor dimer before incubation (e) and after incubation for 0.5 h at
70 °C (f) and 80 °C (g). Measurement conditions: column, Superdex 75 10/300 GL; flow
rate, 0.5 mL/min; monitoring wavelength, 410 nm; solvent, 50 mM potassium phosphate
buffer; pH, 7.0; temperature, 4 °C.
Abs
orb
ance
16128Elution volume (mL)
a
b
c
d
Abs
orb
ance
16128Elution volume (mL)
e
f
g

S14
Figure S5. Optical absorption and CD spectra of oxidized WT cyt c552 monomer (black),
insG3 cyt c552 monomer (red), insG3 cyt c552 major dimer (blue) and insG3 cyt c552 minor
dimer (green): (A) absorption and (B) CD spectra. The concentration of the protein was
calculated from the intensity of its Soret band. Measurement conditions: sample concentration
(heme unit), (A) 16−17 μM and (B) 10 μM; buffer, 50 mM potassium phosphate buffer; pH,
7.0; temperature, (A) room temperature and (B) 25 °C.
120
80
40
0
10
-3 (M
-1 c
m-1
)
700600500400300Wavelength (nm)
A
40
20
0
-20
[]×
10
-3 (de
g・cm
2・dm
ol-
1)
240220200Wavelength (nm)
B

S15
Figure S6. Overlapped view of the structures WT monomer (PDB ID: 1YNR) (gray) and (A)
insG3 cyt c552 major dimer (PDB ID: 5AUR) (red and green) or (B) insG3 cyt c552 minor
dimer (PDB ID: 5AUS) (red and blue). His14, Met62 (Met59 for the monomer) and the
hemes are shown as stick models
S16
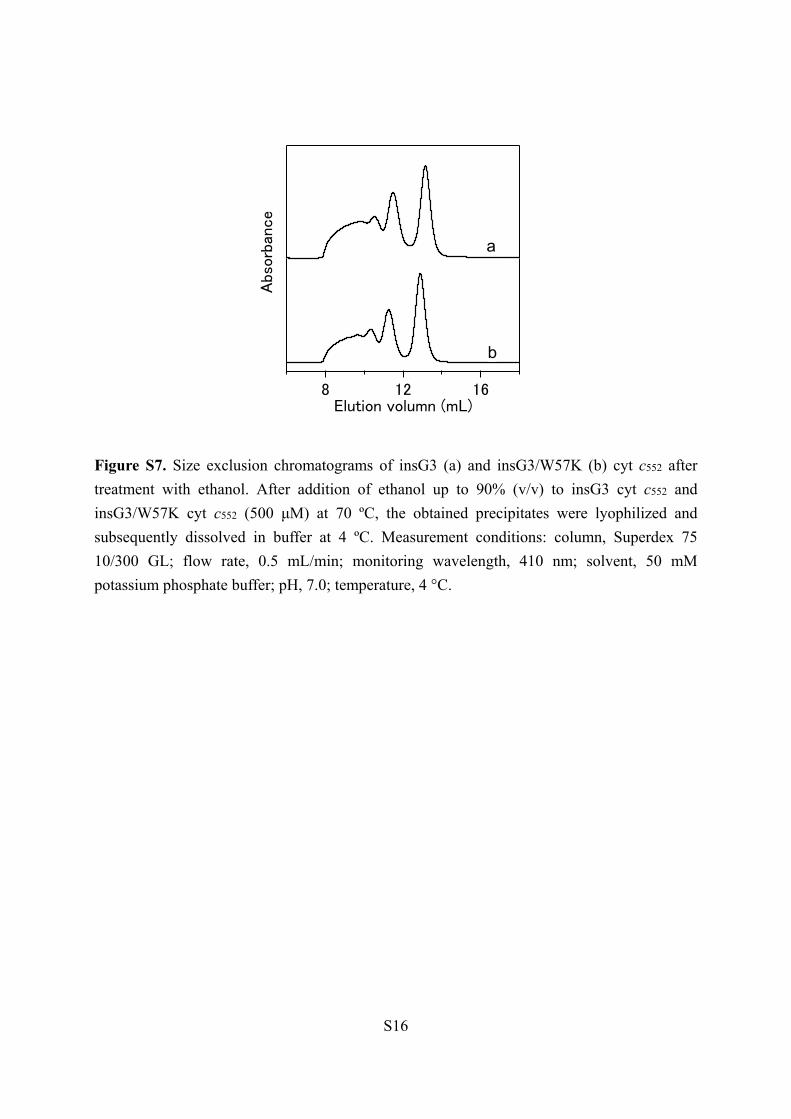
Figure S7. Size exclusion chromatograms of insG3 (a) and insG3/W57K (b) cyt c552 after
treatment with ethanol. After addition of ethanol up to 90% (v/v) to insG3 cyt c552 and
insG3/W57K cyt c552 (500 μM) at 70 ºC, the obtained precipitates were lyophilized and
subsequently dissolved in buffer at 4 ºC. Measurement conditions: column, Superdex 75
10/300 GL; flow rate, 0.5 mL/min; monitoring wavelength, 410 nm; solvent, 50 mM
potassium phosphate buffer; pH, 7.0; temperature, 4 °C.
16128Elution volumn (mL)
Abs
orb
ance
a
b

S17
Figure S8. TEM image of insG3/W57K cyt c552 oligomers. Measurement conditions:
protein, 2 μM (heme); staining reagent, 5% (w/w) phosphotungstic acid (pH 7.0); grid,
carbon-coated 200-mesh copper grid; voltage, 300 kV; temperature, 18 oC.







