
Invited review
Nuclear gene transcription and chromatin in Trypanosoma brucei
David Horn*
London School of Hygiene and Tropical Medicine, Keppel Street, London, WC1E 7HT, UK
Received 2 February 2001; received in revised form 13 February 2001; accepted 25 April 2001
Abstract
As in other eucaryotes, the nuclear genome in Trypanosoma brucei is organised into silent domains and active domains transcribed by
distinct RNA polymerases. The basic mechanisms underlying eucaryotic gene transcription are conserved between humans and yeast, and
understood in some detail in these cells. Meanwhile, relatively little is known about the transcription machinery, the chromatin templates or
their interactions in trypanosomatids. Here, I discuss and compare nuclear gene transcription in T. brucei with transcription in other
eucaryotes focusing in particular on mono-allelic transcription of genes that encode the variant surface glycoproteins. q 2001 Australian
Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
Keywords: Transcription; Trypanosoma brucei; VSG
1. Introduction
Trypanosoma brucei are mono-¯agellated parasitic proto-
zoa. These African trypanosomes cause diseases known as
sleeping sickness in humans and Nagana in domestic live-
stock. They are spread among mammalian hosts by tsetse
¯ies of the genus Glossina, the limited distribution of which
actually prevents T. brucei from spreading beyond Africa. T.
brucei passes through several dividing and non-dividing
stages during its life cycle.
The molecules involved in eucaryotic transcription
include the DNA templates, the RNA polymerases, general
transcription factors, the histones and additional regulatory
molecules (Kornberg, 1999). These molecules and their
interactions are conserved between humans and yeast.
Trypanosoma brucei and several other trypanosomatids
are parasites of signi®cant medical, veterinary and scienti®c
importance but relatively little is known about the transcrip-
tion machinery or the chromatin templates in these diver-
gent protozoa. See Ersfeld et al. (1999) and Belli (2000) for
recent reviews dealing with nuclear organisation in T.
brucei and chromatin remodelling in trypanosomatids,
respectively.
Genome sequencing is underway for T. brucei (El-Sayed
et al., 2000) as well as for the other pathogenic trypanoso-
matids, Trypanosoma cruzi and Leishmania major. The T.
brucei genome size is ,3.5 £ 107 bp with 11 homologous
chromosome pairs and additional intermediate and minichro-
mosomes of uncertain ploidy. Sequence comparisons
between known rRNAs (Sogin et al., 1989), histones
(Thatcher and Gorovsky, 1994) and RNA polymerases
(Jess et al., 1990) suggest that trypanosomes diverged early
from the eucaryotic lineage and several novel molecular and
biochemical features of trypanosomatids also appear to
re¯ect divergence (Donelson et al., 1999). In trypanosoma-
tids almost all protein-coding genes appear to be transcribed
as polycistrons and trans-splicing leads to the addition of a
common sequence to the 5 0 end of every mRNA. Conse-
quently most genes appear to be transcribed constitutively
suggesting the need for signi®cant regulation of mRNA and/
or protein abundance at the post-transcriptional level.
Transcription of genes encoding abundant surface
proteins in T. brucei, the variant surface glycoproteins
(VSGs) and the repetitive EP/GPEET procyclins, differs
from that of most other protein-coding genes in at least
two important ways. Firstly, they are transcribed in a
stage-speci®c manner. The VSG is expressed during the
bloodstream stage of the life cycle and the EP/GPEET
procyclins in the insect midgut stage. Second, VSG and
EP/GPEET genes are almost certainly transcribed by
RNA polymerase I (Lee and Van der Ploeg, 1997), a poly-
merase con®ned to rRNA transcription in other eucaryotes.
In each cell, of the 1000 or so VSG genes, about 20 are
localised in polycistronic, telomeric transcription units
that contain several other `expression site associated
genes' encoding various cell-surface proteins (Pays and
Nolan, 1998). However, gene organisation does not appear
International Journal for Parasitology 31 (2001) 1157±1165
0020-7519/01/$20.00 q 2001 Australian Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
PII: S0020-7519(01)00264-8
www.parasitology-online.com
* Tel.: 144-20-7927-2352; fax: 144-20-7636-8739.
E-mail address: [email protected] (D. Horn).

to be optimal as the telomere-proximal VSG, which gener-
ates the cell's most abundant protein, is furthest from the
promoter. The telomeric VSGs are expressed in a mutually
exclusive manner. Low frequency switches in VSG expres-
sion via VSG gene rearrangements or via switching tran-
scription between telomeres allow this extracellular
parasite to undergo antigenic variation such that some T.
brucei cells continually escape host immune responses
(Rudenko et al., 1998). Thus, determination of the molecu-
lar mechanisms and machinery regulating VSG gene expres-
sion remain major goals in the ®eld of the molecular biology
of T. brucei. In addition to the bloodstream VSGs, ,20
telomeric VSGs can be expressed in the insect vector sali-
vary gland and for a few days following mammalian infec-
tion but these `metacyclic' VSGs are in monocistronic
transcription units. The minichromosomes serve as reposi-
tories of additional telomeric VSG genes.
2. RNA polymerases, promoters and terminators
In most eucaryotes, RNA polymerase I is responsible for
the generation of rRNA, RNA polymerase II for mRNA and
small nuclear RNAs and RNA polymerase III for transfer
RNA and other small RNAs. A similar situation appears to
operate in trypanosomes with the major exception that RNA
polymerase I generates some mRNAs (see above). The
genes for the largest sub-units of all three T. brucei RNA
polymerases (Jess et al., 1990) and promoters that recruit
each of them have now been identi®ed (see below). In other
eucaryotes, phosphorylation of the RNA polymerase II C-
terminal domain occurs prior to transcription elongation and
the C-terminal domain couples transcription to mRNA
capping, splicing and polyadenylation (Proudfoot, 2000).
Despite the absence of a typical C-terminal domain on T.
brucei RNA polymerase II (Evers et al., 1989) this protein is
found to be phosphorylated when isolated from elongating
transcription complexes (Chapman and Agabian, 1994) but
a link to mRNA processing has yet to be established.
Unusually, T. brucei has two loci encoding similar RNA
polymerase II genes but the signi®cance is unclear since
disruption of one of these genes has no detectable effect
(Chung et al., 1993).
In other organisms, promoters are composed of core and
regulatory regions. The core includes a sequence ,25 bp
upstream from the start site that is recognised by TATA-
box-binding protein while positive or negative regulatory
sequences can be located nearby or at great distances.
Elements important for the function of the various T. brucei
promoters are illustrated in Fig. 1.
In addition to rRNA promoters, there are three additional
promoters that recruit RNA polymerase I in T. brucei
(Vanhamme and Pays, 1995), the VSG expression site,
metacyclic VSG and EP/GPEET promoters. These RNA
polymerase I promoters have no obvious sequence similar-
ity but there are some common structural features (Fig. 1)
and they all display a similar level of activity in transient
transfection assays (Kim and Donelson, 1997; Vanhamme
and Pays, 1995). In these assays, a region of ,70 bp is
suf®cient for fully active VSG expression site and metacyc-
lic VSG promoters, whereas an equivalent activity from the
EP/GPEET and rRNA promoters requires an additional
upstream control element(s). However, a hybrid VSG
expression site/rRNA promoter does remain functional
(Vanhamme et al., 1995). Thus, the combined `core'
elements from VSG promoters and from these other promo-
ters are not functionally equivalent but the elements prox-
imal to the initiation site may recruit the same factors.
Assays with hybrid promoters con®rm the activity of the
upstream control elements and indicate that EP/GPEET
and rRNA promoter upstream control elements are inter-
changeable (Janz and Clayton, 1994). Previously, it was
unclear whether the putative metacyclic VSG promoter
identi®ed in bloodstream trypanosomes (Kim and Donelson,
1997) could function in metacyclic trypanosomes (Graham
et al., 1999). This now seems likely since 5 0 RACE analysis
indicates that transcription initiation sites for metacyclic
VSGs in tsetse salivary gland-derived cells coincide with
those seen in bloodstream-derived cells (J.D. Barry et al.,
personal communication).
The spliced-leader RNA is encoded by ,100 genes
arranged in tandem arrays. These genes are transcribed by
RNA polymerase II (Gilinger and Bellofatto, 2001) and they
provide the 39-nt fragment that is trans-spliced to the 5 0 end
of every mRNA. The spliced-leader RNA gene promoter
(GuÈnzl et al., 1997) is illustrated in Fig. 1. Although no
other con®rmed RNA polymerase II promoter has been
identi®ed in trypanosomatids, RNA polymerase II transcrip-
tion as measured by UV treatment followed by nuclear run-
on does re¯ect the pattern of gene organisation on L. major
chromosome 1 (P.J. Myler et al., personal communication).
This chromosome appears to contain two large divergent
polycistronic transcription units with 29 genes encoded on
one DNA strand and the remaining 50 genes encoded on the
opposite strand (Myler et al., 1999). Thus, these experi-
ments provide tentative evidence that transcription initiates
at the single `strand in¯ection point' on this chromosome
and proceeds bi-directionally towards each telomere. It will
certainly be interesting to see how these results relate to the
situation in T. brucei and T. cruzi and to see whether the
putative promoter fragments can function on plasmids or at
other chromosomal loci.
A number of genes transcribed by RNA polymerase III
have been characterised in trypanosomatids including those
encoding the U2, U3 and U6 snRNA and the 7SL RNA. The
promoters for these genes require extragenic A and B blocks
that are the intragenic control regions for a divergently
oriented tRNA gene. The tRNAThr/U6 snRNA transcription
unit (Nakaar et al., 1997) is illustrated in Fig. 1.
Some T. brucei promoter elements, such as the upstream
control element of the rRNA promoter (Laufer et al., 1999),
domain IV of the EP/GPEET promoter (Laufer and GuÈnzl,
D. Horn / International Journal for Parasitology 31 (2001) 1157±11651158

2001), some portions of the spliced-leader RNA promoter
(GuÈnzl et al., 1997) and the B block of the U6 snRNA gene
(Nakaar et al., 1997) have been shown to be more important
for transcription activity in vivo than in vitro. This suggests
that these elements (Fig. 1) are speci®cally involved in
chromatin organisation or the recruitment of factors in a
chromosomal context.
Due to the relative lack of clearly de®ned transcription
units in trypanosomatids transcription termination has not
been investigated at many loci. Transcription termination
speci®c for RNA polymerase I occurs within the terminal
2 kbp of a ,10-kbp EP/GPEET transcription unit (Berberof
et al., 1996). DNA fragments from this region also terminate
transcription from VSG expression site and rRNA promoters
in an orientation dependent manner and the sequence
involved appears to consist of several attenuator elements.
A 9-bp motif within these elements may function in attenua-
tion by recruiting a protein that also binds promoters and
telomeres (Berberof et al., 2000). RNA polymerase II tran-
scription is terminated at spliced-leader RNA loci by a string
of .six T bases at the 3 0 end of the gene (Sturm et al., 1999).
3. General transcription factors
Synthesis of mRNA requires the combined activities of a
large number of polypeptides. In most eucaryotes, TATA-
box-binding protein is central to complex assembly at core
promoter elements prior to transcription initiation by all three
eucaryotic RNA polymerases (Green, 2000). Although tran-
D. Horn / International Journal for Parasitology 31 (2001) 1157±1165 1159
Fig. 1. T. brucei promoters. Schematic diagram of the RNA polymerase I promoters; VSG expression site (ES), metacyclic (M) VSG, EP/GPEET and rRNA, an
RNA polymerase II promoter for the spliced-leader (SL) RNA and an RNA polymerase III promoter for the tRNAThr/U6 snRNA. Transcription start-sites are
indicated as arrows while boxes indicate elements shown to be important for promoter activity in vitro and/or in vivo. Grey boxes represent regions that appear
to be speci®cally important in vivo (see text). A longer box does not necessarily indicate a larger element, but rather that ®ne mapping within that region has not
been done. The structure of the M VSG promoter was determined by mutagenesis (Kim and Donelson, 1997) but this has not been veri®ed by linker-scanning
analysis. Some promoter elements are labelled and the positions of the VSG promoters relative to telomeres (end) are indicated (see text).

scription can occur in the absence of TATA-box-binding
protein, this protein has an ancient evolutionary origin even
appearing in the archea. Neither TATA-box-binding protein
nor general transcription factors that are well conserved
between human and yeast appear to have orthologues in
trypanosomatids suggesting that these factors are either
poorly conserved or have been replaced by a distinct
complex.
Using a combination of in vitro activity assays, gel mobi-
lity shift assays and footprinting assays with extracts from
trypanosomatids including Leptomonas spp., speci®c bind-
ing activities have been characterised for the RNA polymer-
ase I promoters (see Laufer and GuÈnzl, 2001), the spliced-
leader RNA RNA polymerase II promoter (see Gilinger and
Bellofatto, 2001) and RNA polymerase III (Bell and Barry,
1995) promoters. Results regarding the functional signi®-
cance of factors that bind to T. brucei RNA polymerase I
promoters appear to be rather controversial. For example,
factors bind double-stranded DNA from the VSG expression
site promoter (Pham et al., 1997) and a common factor binds
single-stranded DNA from the VSG expression site promoter,
telomeric repeats and the EP/GPEET terminator (Berberof et
al., 2000). However, transcription from the VSG expression
site promoter appears to require only factors that bind
double-stranded DNA (Laufer and GuÈnzl, 2001). The VSG
expression site, EP/GPEET and rRNA promoters recruit at
least one common factor and the rRNA and spliced-leader
RNA promoters, which recruit different RNA polymerases,
also appear to recruit a common factor (Laufer and GuÈnzl,
2001). This second factor does not bind the VSG expression
site or EP/GPEET promoters. In some cases it appears that
promoter elements co-operate in factor recruitment since
altered spacing of the elements disrupts factor binding. In
addition, a trans-activating transcription factor speci®c for
the EP/GPEET promoter appears to be developmentally
regulated, expressed only at the insect midgut stage (Laufer
et al., 1999).
4. The histones
Chromosomal DNA is folded with nucleosomes that
contain two molecules of each core histone, H2A, H2B,
H3 and H4. In other eucaryotes, nucleosomes repress tran-
scription in vitro and in vivo whereas transcription factors
counteract nucleosome-mediated repression. Chromatin
structure is regulated by covalent modi®cation of the
DNA or the histones or by a variety of other factors. For
example, histone N-terminal tails that extend beyond the
nucleosome core can be acetylated, phosphorylated, ubiqui-
tinated, methylated or ADP ribosylated. TATA-box-binding
protein associated factor (TAF) II250 alone can catalyse the
®rst three of these modi®cations (Mizzen and Allis, 2000)
resulting in altered chromatin structure and transcription.
T. brucei core histone sequences (Fig. 2) were identi®ed
by combining partial peptide sequences (Bender et al.,
1992) with expressed sequence tags (El-Sayed et al.,
1995). Although trypanosomatid histones (Galanti et al.,
1998) are among the most divergent known (Thatcher and
Gorovsky, 1994), the level of sequence conservation
suggests that the overall structure of the T. brucei nucleo-
some is probably similar to that in other eucaryotes. Thus,
lysine residues, substrates for acetylation or methylation
(Strahl and Allis, 2000), appear to be conserved within the
N-terminal tails of the core histones (Fig. 2). Phosphoryla-
tion of serine 10 in histone H3 is involved in chromosome
condensation (Wei et al., 1999) in other eucaryotes. The
absence of this residue in the T. brucei histone may be
functionally signi®cant since chromosomes do not condense
during mitosis in trypanosomatids. Genes encoding histone
`variants' that may in¯uence chromatin structure, organisa-
tion or metabolism are also present in the T. brucei genome
but the function of these proteins is currently unknown.
Linker histones contribute to higher order chromatin
structure by binding the linker DNA ¯anking the nucleo-
some core. Although the linker histone H1 is non-essential
D. Horn / International Journal for Parasitology 31 (2001) 1157±11651160
Fig. 2. T. brucei core histones. Predicted protein sequences of T. brucei (Tb) and S. cerevisiae (Sc) core histones were aligned using ClustalW followed by
manual adjustment. Identical residues are white on a black background and dashes indicate gaps introduced to optimise the alignment. Since lysine (K) residues
within the N-terminal tails of histones H3 and H4 are well-characterised substrates for covalent modi®cation (see text), these tails have been speci®cally
adjusted to optimise lysine alignments. Possible substrates for acetylation are indicated (*) on T. brucei histones H3 and H4.
in Tetrahymena, Xenopus or yeast (Patterton et al., 1998) it
does appear to regulate transcription in these organisms
(Wolffe, 1997). For example, histone H1 regulates core
histone acetylation (Gunjan et al., 2001) and in Tetrahy-
mena, phosphorylation of histone H1 affects transcription
by mimicking H1 removal (Dou et al., 1999).
T. brucei histone H1 sequence was identi®ed by compar-
ing peptide sequences (Burri et al., 1995) with expressed
sequence tags (El-Sayed et al., 1995). Like other protozoa,
T. brucei has atypical linker histones lacking a globular
domain. Histone H1 from T. brucei appears to have a rather
simple, repetitive sequence composed almost entirely of ®ve
amino acid repeats, (X3K2)11±14 where X is a neutral amino
acid (Ala, Val, Pro or occasionally Leu or Gly) and K is
lysine. Similar to the situation in other eucaryotes, H1 phos-
phorylation in T. brucei may be inversely related to chromo-
some compaction (Burri et al., 1995).
5. Histone acetylation and chromatin structure
The link between transcription and acetylation of the 1-
amino group of speci®c lysine residues in the N-terminal tails
of the core histones is well-documented (Strahl and Allis,
2000) and conserved from protozoa to mammals. In general,
histone acetylation increases the rate of transcription so
histone acetyltransferases (Brown et al., 2000) and histone
deacetylases (Ng and Bird, 2000) activate and repress tran-
scription, respectively. These enzymes have no observable
preference for a speci®c DNA sequence environment so
targeting is via association with DNA-binding transcription
factors within large multi-component complexes. A protein
module known as a bromodomain binds to acetylated lysines
on histones H3 and H4 and appears to provide a direct link to
transcription activation (Jacobson et al., 2000) as illustrated
by TAFII250 which has two bromodomains. In yeast, acety-
lation of lysine residues 9, 14, 18 and 23 in histone H3 and 5,
8, 12 and 16 in histone H4, can in¯uence transcription
(Waterborg, 2000). The corresponding lysines in T. brucei
may be residues 10, 16, 19 and 23 in histone H3 and 2, 5, 10
and 14 in histone H4 (Fig. 2).
There are several deacetylases, acetyltransferases and
bromodomain proteins that clearly target histones in
human cells and in the yeast Saccharomyces cerevisiae.
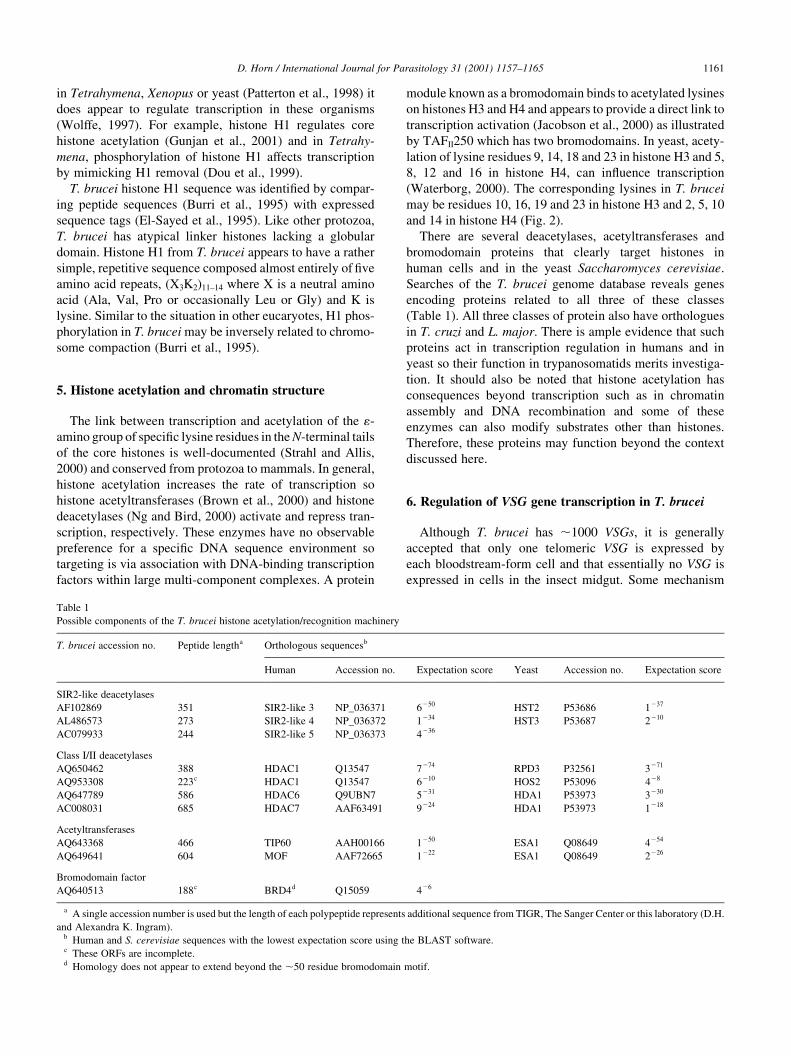
Searches of the T. brucei genome database reveals genes
encoding proteins related to all three of these classes
(Table 1). All three classes of protein also have orthologues
in T. cruzi and L. major. There is ample evidence that such
proteins act in transcription regulation in humans and in
yeast so their function in trypanosomatids merits investiga-
tion. It should also be noted that histone acetylation has
consequences beyond transcription such as in chromatin
assembly and DNA recombination and some of these
enzymes can also modify substrates other than histones.
Therefore, these proteins may function beyond the context
discussed here.
6. Regulation of VSG gene transcription in T. brucei
Although T. brucei has ,1000 VSGs, it is generally
accepted that only one telomeric VSG is expressed by
each bloodstream-form cell and that essentially no VSG is
expressed in cells in the insect midgut. Some mechanism
D. Horn / International Journal for Parasitology 31 (2001) 1157±1165 1161
Table 1
Possible components of the T. brucei histone acetylation/recognition machinery
T. brucei accession no. Peptide lengtha Orthologous sequencesb
Human Accession no. Expectation score Yeast Accession no. Expectation score
SIR2-like deacetylases
AF102869 351 SIR2-like 3 NP_036371 6250 HST2 P53686 1237
AL486573 273 SIR2-like 4 NP_036372 1234 HST3 P53687 2210
AC079933 244 SIR2-like 5 NP_036373 4236
Class I/II deacetylases
AQ650462 388 HDAC1 Q13547 7274 RPD3 P32561 3271
AQ953308 223c HDAC1 Q13547 6210 HOS2 P53096 428
AQ647789 586 HDAC6 Q9UBN7 5231 HDA1 P53973 3230
AC008031 685 HDAC7 AAF63491 9224 HDA1 P53973 1218
Acetyltransferases
AQ643368 466 TIP60 AAH00166 1250 ESA1 Q08649 4254
AQ649641 604 MOF AAF72665 1222 ESA1 Q08649 2226
Bromodomain factor
AQ640513 188c BRD4d Q15059 426
a A single accession number is used but the length of each polypeptide represents additional sequence from TIGR, The Sanger Center or this laboratory (D.H.
and Alexandra K. Ingram).b Human and S. cerevisiae sequences with the lowest expectation score using the BLAST software.c These ORFs are incomplete.d Homology does not appear to extend beyond the ,50 residue bromodomain motif.

operates to prevent transcription (elongation) at VSG loci
that appear to represent ,20% of the genome. This includes
40 or so telomeric VSG expression sites with associated
promoters, several hundred chromosome-internal VSGs
and the minichromosomes. The chromosome-internal
VSGs appear to lack promoters but these genes are not
expressed even if a promoter is inserted immediately
upstream (Horn and Cross, 1997).
Transcription from VSG expression site, EP/GPEET and
rRNA promoters is repressed when they are inserted at an
`inactive' VSG expression site in bloodstream form cells
(Horn and Cross, 1995; Horn and Cross, 1997; Rudenko
et al., 1995). A rRNA promoter replacing a VSG expression
site promoter can be switched on and off (Rudenko et al.,
1995) and rRNA and EP/GPEET promoters show clear
developmental regulation at these loci (Horn and Cross,
1997). These results indicate that VSG expression site
switching is independent of the VSG expression site promo-
ter but dependent upon some other property of the locus in
bloodstream-form cells. In addition, an EP/GPEET promo-
ter upstream control element can overcome repression of a
VSG expression site promoter in chromatin (Qi et al., 1996)
presumably via transcription factor recruitment. These data
are consistent with a model in which chromatin components
regulate accessibility of the DNA template to transcription
(elongation) factors. By demonstrating that promoters are
repressed when inserted into an `inactive' VSG expression
site, other studies also support a role for chromatin in VSG
regulation at the bloodstream-stage (Graham et al., 1998)
and the insect midgut-stage (Navarro et al., 1999; Qi et al.,
1996; Rudenko et al., 1994).
At yeast telomeres, histone deacetylases (Imai et al.,
2000) and the histones themselves (Wyrick et al., 1999)
in¯uence gene silencing (Grunstein, 1998) extending at
least 20 kbp from the telomere. In T. brucei, VSG silencing
appears to depend upon epigenetic events that are reversible
and locus-dependent (Horn and Cross, 1997). Thus, telo-
meric silencing in yeast may share parallels with silencing
of telomeric VSG genes. Micrococcal nuclease treatment
indicated that T. brucei DNA, including sequences encoding
the expressed and non-expressed VSG genes (Greaves and
Borst, 1987) is organised into nucleosomal arrays (Hecker et
al., 1989). Therefore, chromatin-modifying enzymes in T.
brucei, those that acetylate core histones and those that
phosphorylate histone H1 for example, may be involved
in developmental regulation and VSG transcription regula-
tion. Indeed, isoforms of histone H1 vary, electron dense
chromatin is more prominent and chromatin ®bres are more
condensed in bloodstream-stage cells relative to the insect
midgut-stage (Ersfeld et al., 1999; Hecker et al., 1995).
Similarly, in T. cruzi a decrease in the rate of transcription
in non-proliferative cells correlates with an increase in the
amount of electron-dense heterochromatin (Elias et al.,
2001). Nucleases and heterologous RNA polymerases are
commonly used to probe the accessibility of DNA within
chromatin. Such studies in bloodstream-stage T. brucei cells
show increased nuclease sensitivity at the telomeric end of
an active VSG expression site (Greaves and Borst, 1987) but
show no differential sensitivity between active and
repressed transcription states in the regions surrounding
VSG expression site promoters (Navarro and Cross, 1998).
In contrast T7 RNA polymerase accessibility is increased at
both ends of the expression site at the insect-stage relative to
the bloodstream-stage (Navarro et al., 1999). These results
certainly suggest a role for developmentally regulated chro-
matin remodelling in VSG regulation but it is not clear if
speci®c chromatin structures are required for VSG gene
regulation at the bloodstream-stage.
Since native VSG promoters appear to lack proximal
upstream control elements it seems likely that mutually
exclusive activation of one of these promoters in vivo is
achieved via a more distal positive 'control element'. The
telomere itself which is composed of TTAGGG repeats is a
candidate for such an element since all VSG expression site
and metacyclic VSG promoters (,40 in total) appear to be
located proximal to telomeres and telomeric loci are the
exclusive expression sites for VSG genes. Thus, T. brucei
promoters may be unable to participate in the generation of
processive transcription in a chromosomal context unless a
control element recruits additional factors. Developmen-
tally regulated expression of factors speci®c for the meta-
cyclic VSG and VSG expression site promoter sequences,
the presence of 50-bp repeats upstream of VSG expression
site promoters and/or differences in distance from the telo-
mere (Fig. 1) could explain the activity of these promoters at
different life cycle stages.
Several other factors, apart from those discussed above,
could be involved in mutually exclusive VSG expression at
the bloodstream-stage and VSG inactivation in the insect-
stage. Although DNA methylation has not been observed in
T. brucei (Gommers-Ampt and Borst, 1995), nucleotide
base glucosylation forms the J-base speci®cally in the
bloodstream-stage. This J base may strengthen silencing
and/or help to stabilise repeats in the genome by acting as
a `¯ag' to target speci®c factors to repetitive regions (van
Leeuwen et al., 2000). Sub-nuclear organisation may be
important although VSG activation does not appear to be
based upon RNA polymerase I recruitment through locali-
sation in the nucleolus. The active VSG actually appears to
be transcribed at a discreet site outside the nucleolus
(Chaves et al., 1998). Productive VSG transcription may
depend upon speci®c recruitment of a transcription elonga-
tion factor or RNA processing machinery (Vanhamme et al.,
2000). This is supported by the observation of truncated
transcription at repressed VSG expression sites. Since the
VSG lies ,50 kbp from its promoter transcription
complexes with low processivity will not produce VSG
mRNA. However, at least two observations appear to favour
a model in which productive transcription depends upon the
speci®c recruitment of an elongation factor rather than the
RNA processing machinery. First, inhibition of trans-spli-
cing does not affect transcription of the a-tubulin genes
D. Horn / International Journal for Parasitology 31 (2001) 1157±11651162

(Tschudi and Ullu, 1990) suggesting that the two processes
are not coupled at least in the case of genes transcribed by
RNA polymerase II. Second, repressed rRNA promoters
inserted at VSG loci generate processive transcription
complexes in bloodstream form cells (Horn and Cross,
1997) suggesting that reduced processivity is speci®cally
associated with the VSG expression site promoter in this
life cycle stage.
In summary, T. brucei telomeres may recruit relatively
abundant negative regulatory factors that can repress tran-
scription (elongation) over long distances, possibly via
DNA folding or looping while a single telomere may bind
a dominant positive regulatory factor that would counteract
silencing at that locus and allow switching between telo-
meres. The proposed positive factor could be a specialised
nuclear pore. Such an association could facilitate processive
transcription and allow rapid export of VSG mRNA explain-
ing why mature VSG transcripts are not detected in isolated
nuclei (Vanhamme et al., 2000). A precedent for such a
model exists in the yeast S. cerevisiae where telomeres
interact with nuclear pore complexes (Galy et al., 2000)
and telomere folding can facilitate gene silencing (de
Bruin et al., 2000) or gene activation (de Bruin et al., 2001).
7. Concluding remarks
As expected for an early-branching eucaryote, the tran-
scription machinery and certain aspects of gene expression
appear divergent in T. brucei relative to those observed in
later-branching eucaryotes. However, as in other eucaryotes,
T. brucei chromosomes are organised into distinct transcrip-
tion domains, which involves the speci®c targeting of tran-
scription factors. It is interesting that there are no clear
examples of regulated transcription in T. cruzi or Leishma-
nia. Distinct from the situation in T. brucei, transcription
occurs on transiently transfected plasmids containing a
trans-splice acceptor site and an adjacent polypyrimidine
tract but no additional trypanosomatid sequence (Curotto
de Lafaille et al., 1992). These same plasmid DNA sequences
may be unable to recruit RNA polymerase II on chromo-
somes while the `promoters' that can, may fail to act as
promoters when moved from their native context and placed
on plasmids. Thus, it is unclear how or to what extent RNA
polymerase II is targeted to speci®c DNA sequences or chro-
mosomal domains in trypanosomatids. Since RNA polymer-
ase I promoter regulation in T. brucei appears to be dependent
upon chromosomal context it may be necessary to dissect
promoter control elements at their native loci. However,
such experiments with VSG expression site promoters did
not reveal speci®c regulatory sequences proximal to the
core promoter (Blundell and Borst, 1998; Navarro and
Cross, 1998).
In other organisms, transcription regulation is understood
in some detail and histone modi®cation plays a major role.
In T. brucei, histone and DNA modi®cation, nuclear orga-
nisation and interactions between the RNA polymerases and
RNA processing machinery all have the potential to regulate
transcription initiation and/or elongation events. Other
important regulatory sequences are those involved in tran-
scription termination (Berberof et al., 1996; Sturm et al.,
1999) and the `insulators' (Bell et al., 2001) that form
boundaries between transcription domains.
Trypanosomatid genome sequence allows us to clarify
and improve our understanding of the relationship between
a variety of processes in these organisms and other eucar-
yotes. As the T. brucei genome sequencing project
approaches completion it will be interesting to see how
VSGs and other genes, known promoters and repetitive
elements are organised and what additional candidate tran-
scription regulatory molecules will be identi®ed. Since the
transcription machinery in trypanosomatids appears to be
divergent, direct characterisation of promoter-binding
factors and other interacting factors will be necessary but
genome sequence data certainly offers new approaches to
experimental design. Combined with established and emer-
ging tools, such as genetic manipulation (Clayton, 1999),
RNA interference (Shi et al., 2000), analysis of transcription
in vitro (Laufer and GuÈnzl, 2001) and `functional genomic'
analysis, these data will continue to facilitate progress in
characterising transcription regulation in the trypanosoma-
tids.
Acknowledgements
I would like to thank John Kelly and Martin Taylor for
critical reading of the manuscript, Vivian Bellofatto, Dave
Barry, Arthur GuÈnzl and Peter Myler for communicating
unpublished data and Alexandra K. Ingram for providing
sequence data. Preliminary sequence data was obtained
from The Insitute for Genomic Research website (www.ti-
gr.org.) and The Sanger Centre website (www.sanger.a-
c.uk). Research in D.H.'s laboratory is supported by a
Research Career Development Fellowship from The Well-
come Trust (052323).
References
Bell, S., Barry, J., 1995. Trypanosome nuclear factors which bind to inter-
nal promoter elements of tRNA genes. Nucleic Acids Res. 23, 3103±10.
Bell, A., West, A., Felsenfeld, G., 2001. Insulators and boundaries: versatile
regulatory elements in the eucaryotic genome. Science 291, 447±50.
Belli, S., 2000. Chromatin remodelling during the life cycle of trypanos-
matids. Int. J. Parasitol. 30, 679±87.
Bender, K., Betschart, B., Schaller, J., Kampfer, U., Hecker, H., 1992.
Sequence differences between histones of procyclic Trypanosoma
brucei brucei and higher eucaryotes. Parasitology 105, 97±104.
Berberof, M., Pays, A., Lips, S., Tebabi, P., Pays, E., 1996. Characterization
of a transcription terminator of the procyclin PARP A unit of Trypano-
soma brucei. Mol. Cell. Biol. 16, 914±24.
Berberof, M., Vanhamme, L., Alexandre, S., Lips, S., Tebabi, P., Pays, E.,
2000. A single-stranded DNA-binding protein shared by telomeric
repeats, the variant surface glycoprotein transcription promoter and
D. Horn / International Journal for Parasitology 31 (2001) 1157±1165 1163

the procyclin transcription terminator of Trypanosoma brucei. Nucleic
Acids Res. 28, 597±604.
Blundell, P.A., Borst, P., 1998. Analysis of a variant surface glycoprotein
gene expression site promoter of Trypanosoma brucei by remodelling
the promoter region. Mol. Biochem. Parasitol. 94, 67±85.
Brown, C., Lechner, T., Howe, L., Workman, J., 2000. The many HATs of
transcriptional coactivators. Trends Biochem. Sci 25, 15±19.
de Bruin, D., Kantrow, S., Liberatore, R., Zakian, V., 2000. Telomere
folding is required for the stable maintenance of telomere position
effects in yeast. Mol. Cell. Biol. 20, 7991±8000.
de Bruin, D., Zaman, Z., Liberatore, R., Ptashne, M., 2001. Telomere loop-
ing permits gene activation by a downstream UAS in yeast. Nature 409,
109±13.
Burri, M., Schlimme, W., Betschart, B., Lindner, H., KaÈmpfer, U., Schaller,
J., Hecker, H., 1995. Partial amino acid sequence and functional aspects
of histone H1 proteins in Trypanosoma brucei brucei. Biol. Cell 83, 23±
31.
Chapman, A.B., Agabian, N., 1994. Trypanosoma brucei RNA polymerase
II is phosphorylated in the absence of carboxyl-terminal domain hepta-
peptide repeats. J. Biol. Chem. 269, 4754±60.
Chaves, I., Zomerdijk, J., Dirks Mulder, A., Dirks, R.W., Raap, A.K., Borst,
P., 1998. Subnuclear localization of the active variant surface glyco-
protein gene expression site in Trypanosoma brucei. Proc. Natl Acad.
Sci. USA 95, 12328±33.
Chung, H.M.M., Lee, M.G.-S., Dietrich, P., Huang, J., Van der Ploeg,
L.H.T., 1993. Disruption of largest subunit RNA polymerase II genes
in Trypanosoma brucei. Mol. Cell. Biol. 13, 3734±43.
Clayton, C.E., 1999. Genetic manipulation of kinetoplastida. Parasitol.
Today 15, 372±8.
Curotto de Lafaille, M.A., Laban, A., Wirth, D.F., 1992. Gene expression in
Leishmania: analysis of essential 5 0 DNA sequences. Proc. Natl Acad.
Sci. USA 89, 2703±7.
Donelson, J.E., Gardner, M.J., El-Sayed, N.M., 1999. More surprises from
Kinetoplastida. Proc. Natl Acad. Sci. USA 96, 2579±81.
Dou, Y., Mizzen, C.A., Abrams, M., Allis, C.D., Gorovsky, M.A., 1999.
Phosphorylation of linker histone H1 regulates gene expression in vivo
by mimicking H1 removal. Mol. Cell 4, 641±7.
Elias, M.C.Q.B., Marques-Porto, R., Freymuller, E., Schenkman, S., 2001.
Transcription rate modulation through the Trypanosoma cruzi life cycle
occurs in parallel with changes in nuclear organisation. Mol. Biochem.
Parasitol. 112, 79±90.
El-Sayed, N.M., Alarcon, C.M., Beck, J.C., Shef®eld, V.C., Donelson, J.E.,
1995. cDNA expressed sequence tags of Trypanosoma brucei rhode-
siense provide new insights into the biology of the parasite. Mol.
Biochem. Parasitol. 73, 75±90.
El-Sayed, N.M., Hegde, P., Quackenbush, J., Melville, S., Donelson, J.,
2000. The African trypanosome genome. Int. J. Parasitol. 30, 329±45.
Ersfeld, K., Melville, S.E., Gull, K., 1999. Nuclear and genome organiza-
tion of Trypanosoma brucei. Parasitol. Today 15, 58±63.
Evers, R., Hammer, A., Kock, J., Waldemar, J., Borst, P., Memet, S.,
Cornelissen, A.W.C.A., 1989. Trypanosoma brucei contains two
RNA Polymerase II largest subunit genes with an altered C-terminal
domain. Cell 56, 585±97.
Galanti, N., Galindo, M., Sabaj, V., Espinoza, I., Toro, G.C., 1998. Histone
genes in trypanosomatids. Parasitol. Today 14, 64±70.
Galy, V., Olivo Marin, J.C., Scherthan, H., Doye, V., Rascalou, N., Nehr-
bass, U., 2000. Nuclear pore complexes in the organization of silent
telomeric chromatin. Nature 403, 108±12.
Gilinger, G., Bellofatto, V., 2001. Trypanosome spliced leader RNA genes
contain the ®rst identi®ed RNA polymerase II gene promoter in these
organisms. Nucleic Acids Res. 29, 1556±64.
Gommers-Ampt, J.H., Borst, P., 1995. Hypermodi®ed bases in DNA.
FASEB J. 9, 1034±42.
Graham, S.V., Wymer, B., Barry, J.D., 1998. Activity of a trypanosome
metacyclic variant surface glycoprotein gene promoter is dependent
upon life cycle stage and chromosomal context. Mol. Cell. Biol. 18,
1137±46.
Graham, S.V., Terry, S., Barry, J.D., 1999. A structural and transcription
pattern for variant surface glycoprotein gene expression sites used in
metacyclic stage Trypanosoma brucei. Mol. Biochem. Parasitol. 103,
141±54.
Greaves, D.R., Borst, P., 1987. Trypanosoma brucei variant-speci®c glyco-
protein gene chromatin is sensitive to single-strand-speci®c endonu-
clease digestion. J. Mol. Biol. 197, 471±83.
Green, M.R., 2000. TBP-associated factors (TAFIIs): multiple, selective
transcriptional mediators in common complexes. Trends Biochem.
Sci. 25, 59±63.
Grunstein, M., 1998. Yeast heterochromatin: regulation of its assembly and
inheritance by histones. Cell 93, 325±8.
Gunjan, A., Sittman, D., Brown, D., 2001. Core histone acetylation is
regulated by linker histone stoichiometry in vivo. J. Biol. Chem. 276,
3635±40.
GuÈnzl, A., Ullu, E., Dorner, M., Fragoso, S.P., Hoffmann, K.F., Milner,
J.D., Morita, Y., Nguu, E.K., Vanacova, S., Wunsch, S., Dare, A.O.,
Kwon, H., Tschudi, C., 1997. Transcription of the Trypanosoma brucei
spliced leader RNA gene is dependent only on the presence of upstream
regulatory elements. Mol. Biochem. Parasitol. 85, 67±76.
Hecker, H., Bender, K., Betschart, B., Modespacher, U.P., 1989. Instability
of the nuclear chromatin of procyclic Trypanosoma brucei brucei. Mol.
Biochem. Parasitol. 37, 225±34.
Hecker, H., Betschart, B., Burri, M., Schlimme, W., 1995. Functional
morphology of trypanosome chromatin. Parasitol. Today 11, 79±83.
Horn, D., Cross, G.A.M., 1995. A developmentally regulated position effect
at a telomeric locus in Trypanosoma brucei. Cell 83, 555±61.
Horn, D., Cross, G.A.M., 1997. Position-dependent and promoter-speci®c
regulation of gene expression in Trypanosoma brucei. EMBO J. 16,
7422±31.
Imai, S., Armstrong, C.M., Kaeberlein, M., Guarente, L., 2000. Transcrip-
tional silencing and longevity protein Sir2 is an NAD-dependent
histone deacetylase. Nature 403, 795±800.
Jacobson, R.H., Ladurner, A.G., King, D.S., Tijan, R., 2000. Structure and
function of a human TAFII250 double bromodomain module. Science
288, 1422±5.
Janz, L., Clayton, C., 1994. The PARP and rRNA promoters of Trypano-
soma brucei are composed of dissimilar sequence elements that are
functionally interchangeable. Mol. Cell. Biol. 14, 5804±11.
Jess, W., Palm, P., Evers, R., Kock, J., Cornelissen, A.W., 1990. Phyloge-
netic analysis of the RNA polymerases of Trypanosoma brucei, with
special reference to class-speci®c transcription. Curr. Genet. 18, 547±
51.
Kim, K.S., Donelson, J.E., 1997. Co-duplication of a variant surface glyco-
protein gene and its promoter to an expression site in African trypano-
somes. J. Biol. Chem. 272, 24637±45.
Kornberg, R., 1999. Eucaryotic transcription control. Trends series (millen-
nium issue), M46±9.
Laufer, G., GuÈnzl, A., 2001. In-vitro competition analysis of procyclin gene
and variant surface glycoprotein gene expression site transcription in
Trypanosoma brucei. Mol. Biochem. Parasitol. 113, 55±65.
Laufer, G., Schaaf, G., Bollgonn, S., GuÈnzl, A., 1999. In vitro analysis of a-
amanitin-resistant transcription from the rRNA, procyclic acidic repe-
titive protein, and variant surface glycoprotein gene promoters in
Trypanosoma brucei. Mol. Cell. Biol. 19, 5466±73.
Lee, M.G., Van der Ploeg, L.H.T., 1997. Transcription of protein-coding
genes in trypanosomes by RNA polymerase I. Annu. Rev. Microbiol.
51, 463±89.
van Leeuwen, F., Kieft, R., Cross, M., Borst, P., 2000. Tandemly repeated
DNA is a target for the partial replacement of thymine by b-d-glucosyl-
hydroxymethyluracil in Trypanosoma brucei. Mol. Biochem. Parasitol.
109, 133±45.
Mizzen, A., Allis, D., 2000. New insights into an old modi®cation. Science
289, 2290±1.
Myler, P.J., Audleman, L., deVos, T., Hixson, G., Kiser, P., Lemley, C.,
Magness, C., Rickel, E., Sisk, E., Sunkin, S., Swartzell, S., Westlake, T.,
Bastien, P., Fu, G., Ivens, A., Stuart, K., 1999. Leishmania major Frie-
D. Horn / International Journal for Parasitology 31 (2001) 1157±11651164

dlin chromosome 1 has an unusual distribution of protein-coding genes.
Proc. Natl Acad. Sci. USA 96, 2902±6.
Nakaar, V., GuÈnzl, A., Ullu, E., Tschudi, C., 1997. Structure of the Trypa-
nosoma brucei U6 snRNA gene promoter. Mol. Biochem. Parasitol. 88,
13±23.
Navarro, M., Cross, G.A.M., 1998. In situ analysis of a variant surface
glycoprotein expression-site promoter region in Trypanosoma brucei.
Mol. Biochem. Parasitol. 94, 53±66.
Navarro, M., Cross, G.A.M., Wirtz, E., 1999. Trypanosoma brucei variant
surface glycoprotein regulation involves coupled activation/inactiva-
tion and chromatin remodeling of expression sites. EMBO J. 18,
2265±72.
Ng, H., Bird, A., 2000. Histone deacetylases: silencers for hire. Trends
Biochem. Sci. 25, 121±6.
Patterton, H.G., Landel, C.C., Landsman, D., Peterson, C.L., Simpson,
R.T., 1998. The biochemical and phenotypic characterization of
Hho1p, the putative linker histone H1 of Saccharomyces cerevisiae.
J. Biol. Chem. 273, 7268±76.
Pays, E., Nolan, D.P., 1998. Expression and function of surface proteins in
Trypanosoma brucei. Mol. Biochem. Parasitol. 91, 3±36.
Pham, V.P., Rothman, P.B., Gottesdiener, K.M., 1997. Binding of trans-
acting factors to the double-stranded variant surface glycoprotein
(VSG) expression site promoter of Trypanosoma brucei. Mol. Biochem.
Parasitol. 89, 11±23.
Proudfoot, N., 2000. Connecting transcription to messenger RNA proces-
sing. Trends Biochem. Sci. 25, 290±3.
Qi, C.C., Urmenyi, T., Gottesdiener, K.M., 1996. Analysis of a hybrid
PARP/VSG ES promoter in procyclic trypanosomes. Mol. Biochem.
Parasitol. 77, 147±59.
Rudenko, G., Blundell, P.A., Taylor, M.C., Kieft, R., Borst, P., 1994. VSG
gene expression site control in insect form Trypanosoma brucei. EMBO
J. 13, 5470±82.
Rudenko, G., Blundell, P.A., Dirks-Mulder, A., Kieft, R., Borst, P., 1995. A
ribosomal DNA promoter replacing the promoter of a telomeric VSG
gene expression site can be ef®ciently switched on and off in T. brucei.
Cell 83, 547±53.
Rudenko, G., Cross, M., Borst, P., 1998. Changing the end: antigenic varia-
tion orchestrated at the telomeres of African trypanosomes. Trends
Microbiol. 6, 113±6.
Shi, H., Djikeng, A., Mark, T., Wirtz, E., Tschudi, C., Ullu, E., 2000.
Genetic interference in Trypanosoma brucei by heritable and inducible
double-stranded RNA. RNA 6, 1069±76.
Sogin, M.L., Gunderson, J.H., Elwood, H.J., Alonso, R.A., Peattie, D.A.,
1989. Phylogenetic meaning of the kingdom concept: an unusual ribo-
somal RNA from Giardia lamblia. Science 243, 75±77.
Strahl, B., Allis, C., 2000. The language of covalent histone modi®cations.
Nature 403, 41±45.
Sturm, N.R., Yu, M.C., Campbell, D.A., 1999. Transcription termination
and 3 0-end processing of the spliced leader RNA in kinetoplastids. Mol.
Cell. Biol. 19, 1595±604.
Thatcher, T.H., Gorovsky, M.A., 1994. Phylogenetic analysis of the core
histones H2A, H2B, H3, and H4. Nucleic Acids Res. 22, 174±9.
Tschudi, C., Ullu, E., 1990. Destruction of U2, U4, or U6 small nuclear
RNA blocks trans splicing in trypanosome cells. Cell 61, 459±66.
Vanhamme, L., Pays, E., 1995. Control of gene expression in trypano-
somes. Microbiol. Rev. 59, 223±40.
Vanhamme, L., Pays, A., Tebabi, A., Alexandre, S., Pays, E., 1995. Speci®c
binding of proteins to the noncoding strand of a crucial element of the
variant surface glycoprotein, procyclin and ribosomal promoters of
Trypanosoma brucei. Mol. Cell. Biol. 15, 5598±606.
Vanhamme, L., Poelvoorde, P., Pays, A., Tebabi, P., Van Xong, H., Pays,
E., 2000. Differential RNA elongation controls the variant surface
glycoprotein gene expression sites of Trypanosma brucei. Mol. Micro-
biol. 36, 328±40.
Waterborg, J., 2000. Steady-state levels of histone acetylation in Sacchar-
omyces cerevisiae. J. Biol. Chem. 275, 13007±11.
Wei, Y., Yu, L., Bowen, J., Gorovsky, M.A., Allis, C.D., 1999. Phosphor-
ylation of histone H3 is required for proper chromosome condensation
and segregation. Cell 97, 99±109.
Wolffe, A.P., 1997. Histone H1. Int. J. Biochem. Cell Biol. 29, 1463±6.
Wyrick, J.J., Holstege, F.C., Jennings, E.G., Causton, H.C., Shore, D.,
Grunstein, M., Lander, E.S., Young, R.A., 1999. Chromosomal land-
scape of nucleosome-dependent gene expression and silencing in yeast.
Nature 402, 418±21.
D. Horn / International Journal for Parasitology 31 (2001) 1157±1165 1165







