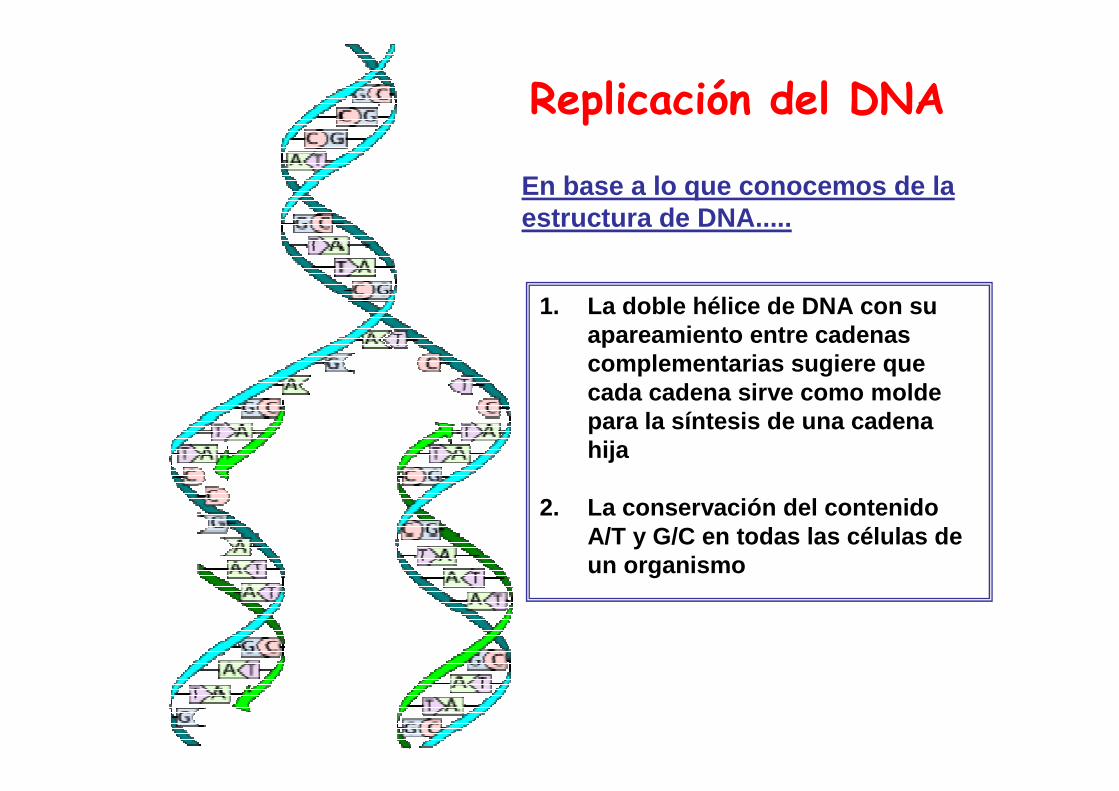
Replicación del DNA
1. La doble hélice de DNA con su apareamiento entre cadenas complementarias sugiere que cada cadena sirve como molde para la síntesis de una cadena hija
2. La conservación del contenido A/T y G/C en todas las células de un organismo
En base a lo que conocemos de la estructura de DNA.....

Posibles mecanismos de replicaciPosibles mecanismos de replicacióón del n del DNADNA
Estructura del DNA
Doble Hélice
Cadenas complementarias
¿Pruebas experimentales?
Hebras parentales Replicación Hebras hijas
Semiconservadora
Conservadora
Dispersa al azar

Experimento de Meselson y Stahl
Separación de DNA por centrifugación en gradiente de CsCl
Alimentar cultivo de E. coli con fuente de 14N o 15N y purificar el DNA
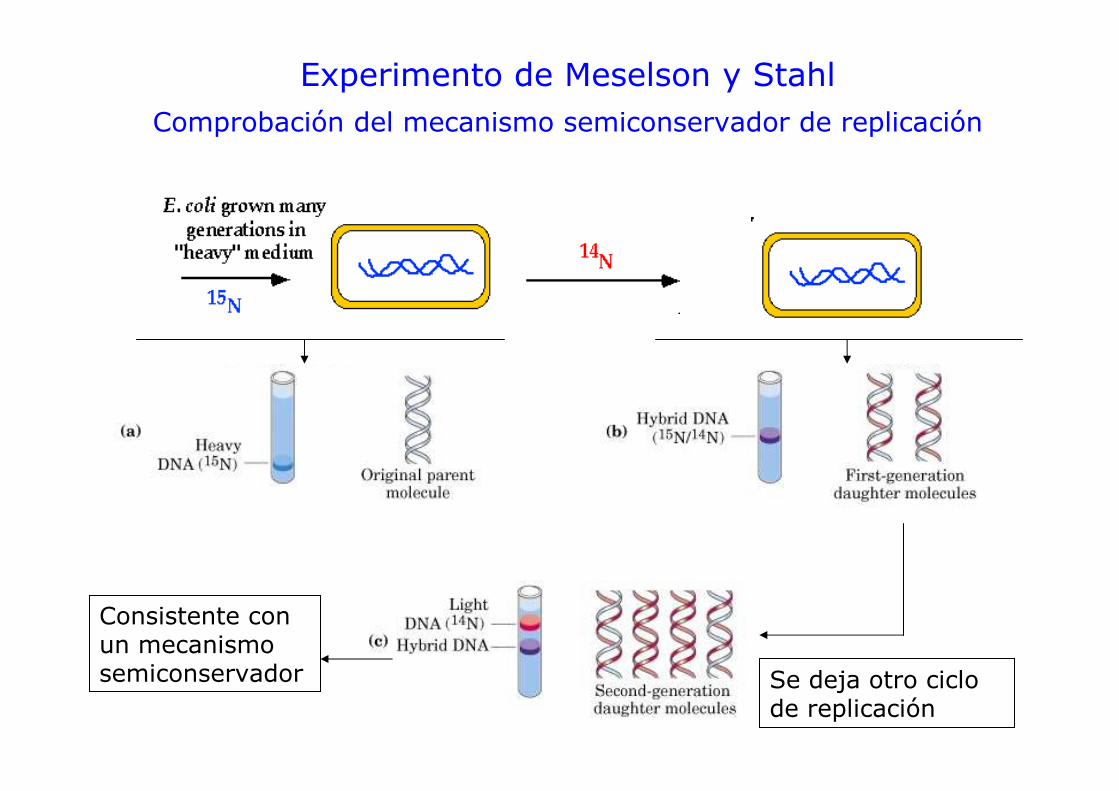
Se deja otro ciclo de replicación
Experimento de Meselson y StahlComprobación del mecanismo semiconservador de replicación
Consistente con un mecanismo semiconservador
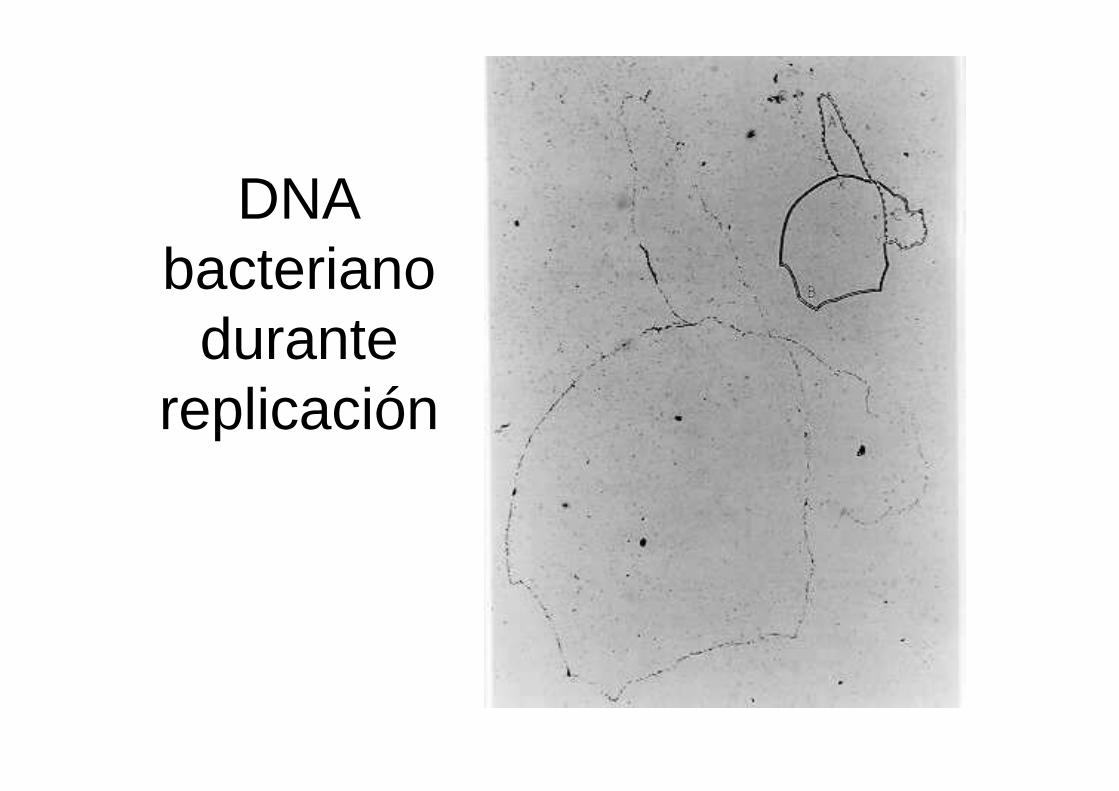
DNA bacteriano
durante replicación

¿Es la replicación del DNA unidireccional o bidireccional?
En bacterias, que tienen genoma circular cerrado
Replicación del DNA de E. coli por 1.5 generaciones en presencia de nucleótidos marcados
Autoradiografía
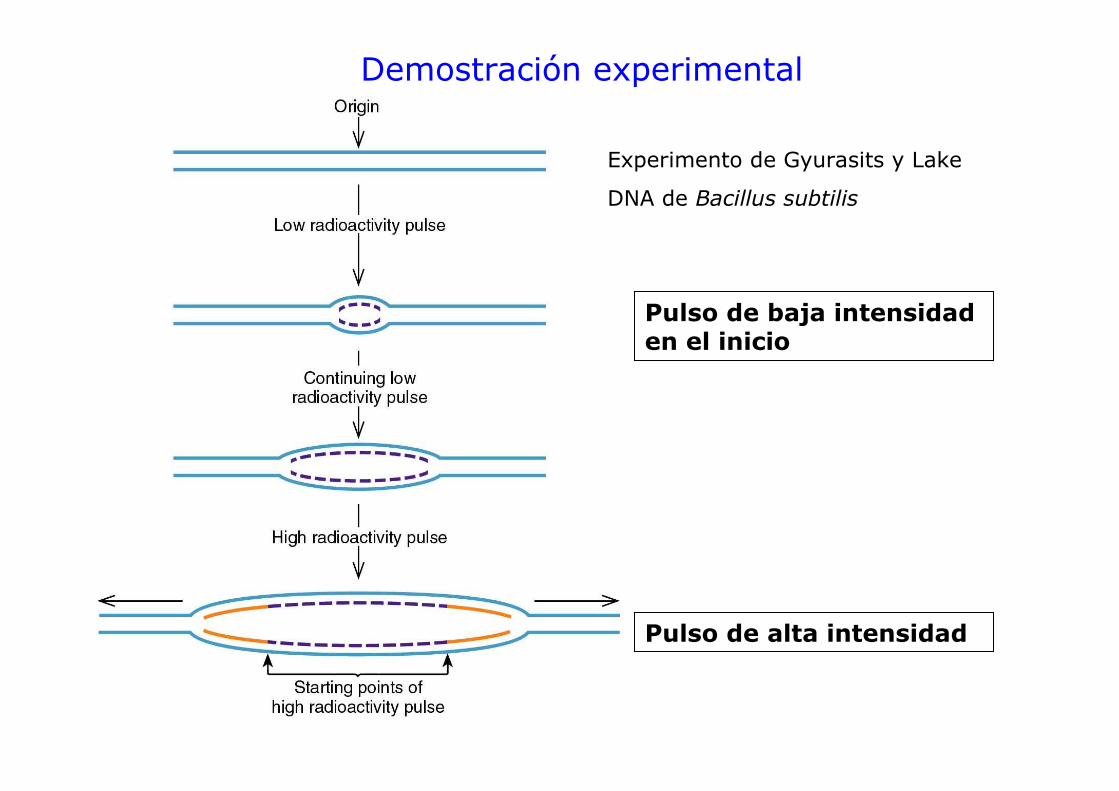
Demostración experimental
Experimento de Gyurasits y Lake
DNA de Bacillus subtilis
Pulso de baja intensidad en el inicio
Pulso de alta intensidad

Resultados del experimento de Gyurasits y Lake

• Horquillas en ambas direcciones muestran marca fuerte, lo que indica que que fueron activas durante el segundo pulso. Ambas horquillas surgen de un punto inicial que es el origen de replicación. Las horquillas se mueven en direcciones opuestas.
• Evidencia de que la replicación del DNA es bidireccional y a partir de un origen de replicación

• Cada cadena de DNA sirve como molde para la síntesis de una cadena nueva
• Los substratos son desoxiribonucleósidos 5’ fosfato (dN TPs: dATP, dTTP, dGTP, dCTP) (nucleótidos)
• Las cadenas de DNA se separan y se sintetiza la heb ra complementaria de cada una, de manera simultánea
• Las DNA polimerasas añaden dNTPs al molde de la cadena madre en dirección 5’ a 3’
DNAn + dNTP (DNA)n+1 + PPi
DNA polimerasa
ReacciReaccióón bn báásica de la replicacisica de la replicacióón de DNAn de DNA

• La DNA polimerasa requiere un 3’OH libre para poder polimerizar
Se produce un ataque Se produce un ataque nucleofnucleofíílicolico del 3del 3’’OH al OH al fosfato fosfato αααααααα del del dNTPdNTP entranteentrante
γγγγ
ββββ
ααααγγγγ
ββββαααα
DNAn
dNTP
molde
(DNA)n+1
PPi
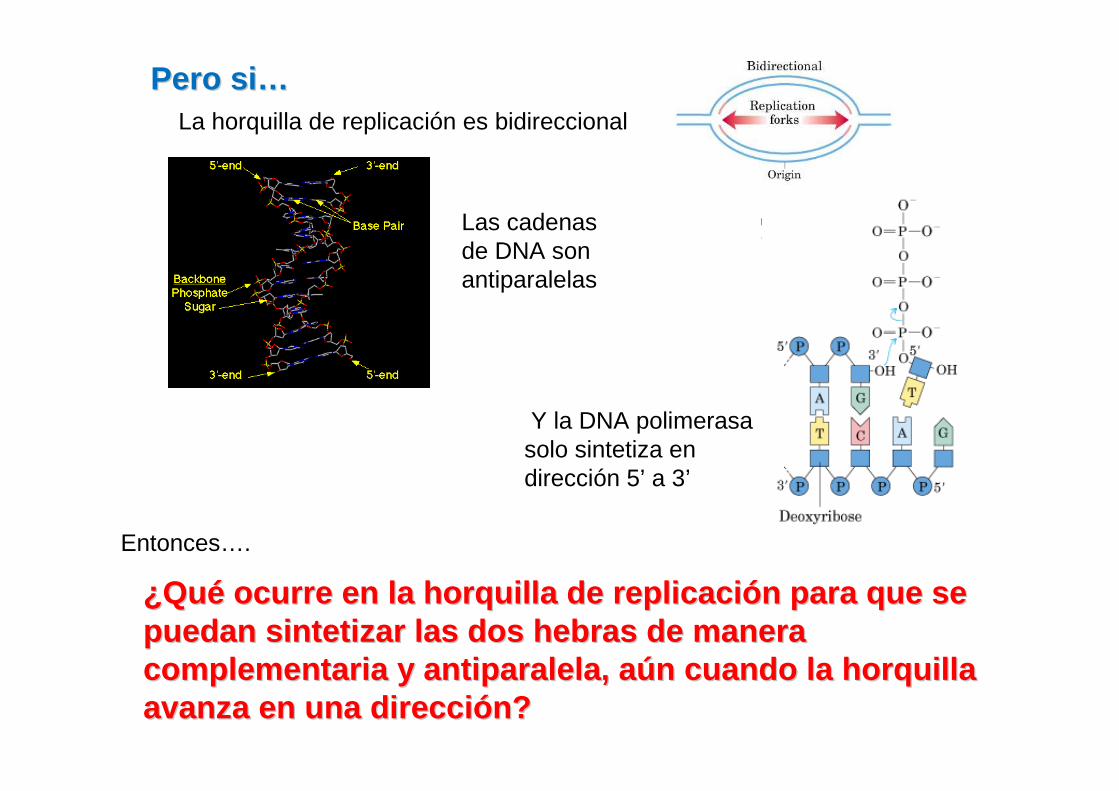
Pero siPero si ……
Y la DNA polimerasa solo sintetiza en dirección 5’ a 3’
Entonces….
¿¿QuQuéé ocurre en la horquilla de replicaciocurre en la horquilla de replicaci óón para que se n para que se puedan sintetizar las dos hebras de manera puedan sintetizar las dos hebras de manera complementaria y antiparalela, acomplementaria y antiparalela, a úún cuando la horquilla n cuando la horquilla avanza en una direcciavanza en una direcci óón?n?
La horquilla de replicación es bidireccional
Las cadenas de DNA son antiparalelas

La replicación del DNA es semidiscontinua
Síntesis continua requiere que las DNA polimerasassintetisen DNA de 5’->3’ y de 3’ -> 5’
Síntesis discontinua las dos cadenas nuevas se sintetizan en fragmentos, en dirección de 5’->3
Síntesis semidiscontinuauna cadena se sintetiza en forma continua y la otra en forma discontinua, en dirección de 5’->3.
Mecanismos para explicar la sMecanismos para explicar la s ííntesis de ntesis de las cadenas complementariaslas cadenas complementarias

Durante la replicación del DNA se sintetizan fragmentos que varían entre 1000 a 2000
nucleótidos
Experimentos de Experimentos de ReijiReiji OkazakiOkazaki
¿Explicación?

3’3’
3’
5’
5’
5’
*
Replicación semidiscontinua:
Una hebra se sintetiza en forma continua y la otra discontinua
Cadena adelantada o guía (leading)/ Cadena retrasada (lagging)

3’3’
3’
5’
5’
5’
5’
5’
5’
3’
3’
3’
*
*
Como la dirección de síntesis de la cadena discontinua es opuesta a la dirección de apertura de la horquilla, se comienza
una nueva cadena.
En cambio, la cadena continua simplemente se sigue alargando

Fragmentos de Okazaki
Como la DNA polimerasa no puede polimerizar una hebra Como la DNA polimerasa no puede polimerizar una hebra nueva de DNA si no cuenta con un cebador que tenga un nueva de DNA si no cuenta con un cebador que tenga un
33’’OH libre, es necesario que estos cebadores sean OH libre, es necesario que estos cebadores sean sintetizados previo a que la DNA polimerasa actsintetizados previo a que la DNA polimerasa actúúe e
Hebra Hebra ““guguííaa””
Hebra Hebra ““retrasadaretrasada””
continua
discontinua
El crecimiento de ambas cadenas es en sentido 5’ 3’

Los cebadores son sintetizados por una DNA primasa
La DNA primasa emplea ribonucleótidos para sintetizar los cebadores

Generalidades de la Replicación de DNA
• Semiconservadora• A partir de un origen de replicación• Bidireccional• Semidiscontinua• Requiere de un cebador de RNA

Origen de ReplicaciOrigen de Replicacióón n OriCOriC
1. Activación por metilación de A en GATC
2. Unión de DnaA “abre el DNA”
3. Unión de DnaB y DnaC- actividad helicasaATP dependiente
4. Unión de SSB para mantener separadas las cadenas de DNA
Cajas reconocidas por DnaA (9 pb) Secuencias repetidas en tandem (13 pb)
En bacteria hay un solo origen de replicación

Proteínas que intervienen en la apertura de la doble hebra de DNA preparándola para ser replicada:
DnaA, DnaB, DnaC, y HUen OriC
El DNA cuando estásuperenrollado no se
puede replicar.

Proteína Dna A• La proteína DnaA es un monómero de 52
kDa. Tiene una muy alta afinidad por ATP (Kd = 0.03 µM) y lo hidroliza lentamente a ADP en una forma dependiente de DNA. Tiene un dominio de unión a ATP.
• Se une con alta afinidad y de forma cooperativa a las cajas dnaA de oriC y se ha calculado una estequiometría de 30 subunidades de dnaA por oriC.
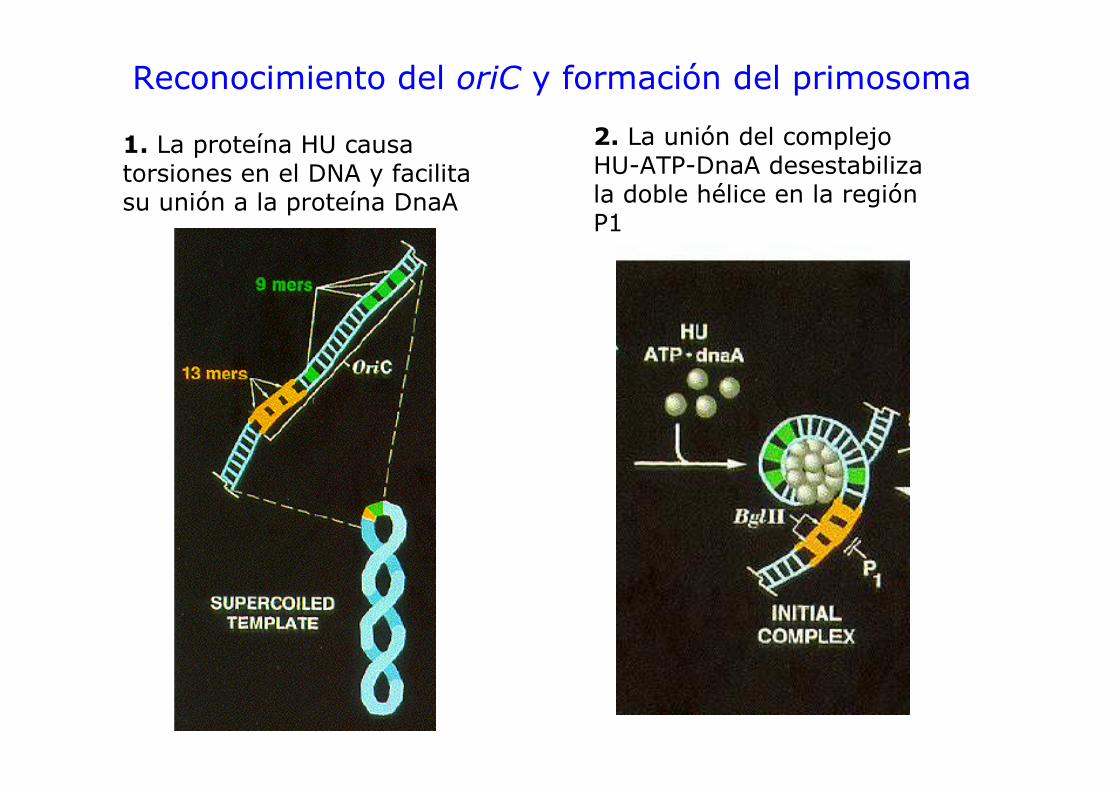
Reconocimiento del oriC y formación del primosoma
1. La proteína HU causa torsiones en el DNA y facilita su unión a la proteína DnaA
2. La unión del complejo HU-ATP-DnaA desestabiliza la doble hélice en la región P1
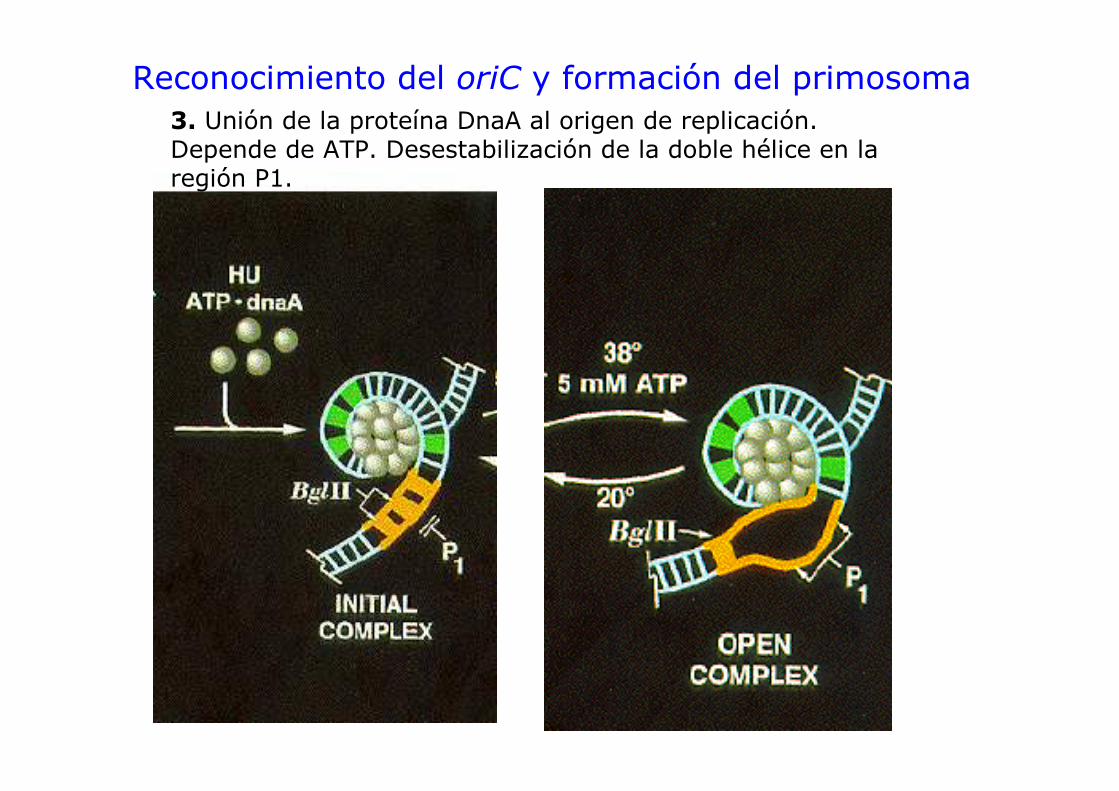
Reconocimiento del oriC y formación del primosoma3. Unión de la proteína DnaA al origen de replicación. Depende de ATP. Desestabilización de la doble hélice en la región P1.

Proteína DnaB. HelicasaHelicasa
Monómeros de 50 kDa que forman un homohexámero.
Tiene dominios que se requieren para:
• Interacción con la proteína DnaC
• Unión a DNA de cadena sencilla
• Unión a DNA de cadena doble
• Activación de la primasa
• Hidrólisis de ATP
La helicasa rodea una de las hebras del DNA duplex y se desplaza logrando la apertura de la doble hélice por exclusión estérica.
Una hebra es excluida del canal interno, mientras que la otra hebra es retenida en el interior del anillo.

DnaBDnaB. . helicasahelicasa
Rompe los puentes de hidrógenoentre las bases

Reconocimiento del oriC y formación del pre-primosoma
3. Unión de DnaB y DnaC a la región abierta del DNA.

Reconocimiento del oriC y formación del primosoma
1. La proteína HU causa torsiones en el DNA y facilita su unión a la proteína DnaA
2. La unión del complejo HU-ATP-DnaAdesestabiliza la doble hélice en la región P1
3. Unión de DnaB a la región abierta del DNA. Unión de DnaC a DnaB

Proteína SSB “Single-strand binding”
La proteína SSB se une al DNA de cadena sencilla con alta afinidad y así previene que se vuelva a formar el híbrido DNA-DNA.
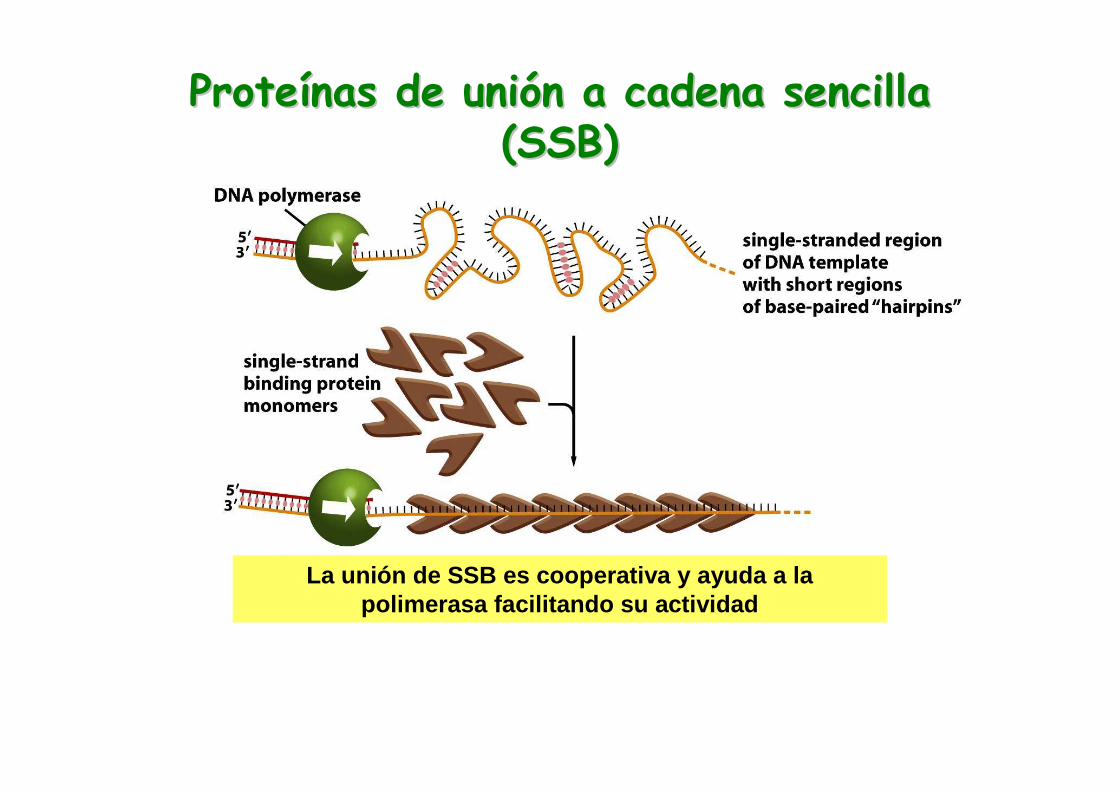
ProteProteíínas de uninas de unióón a cadena sencilla n a cadena sencilla (SSB)(SSB)
La unión de SSB es cooperativa y ayuda a la polimerasa facilitando su actividad

Estructura de las SSBEstructura de las SSB

• Una vez que se ha abierto el DNA y se evita su rehibridación o autohibridación,
comienza la síntesis

Fragmentos de Okazaki
Como la DNA polimerasa no puede polimerizar una hebra Como la DNA polimerasa no puede polimerizar una hebra nueva de DNA si no cuenta con un cebador que tenga un nueva de DNA si no cuenta con un cebador que tenga un
33’’OH libre, es necesario que estos cebadores sean OH libre, es necesario que estos cebadores sean sintetizados previo a que la DNA polimerasa actsintetizados previo a que la DNA polimerasa actúúe e
Hebra Hebra ““guguííaa””
Hebra Hebra ““retrasadaretrasada””
continua
discontinua
El crecimiento de ambas cadenas es en sentido 5’ 3’
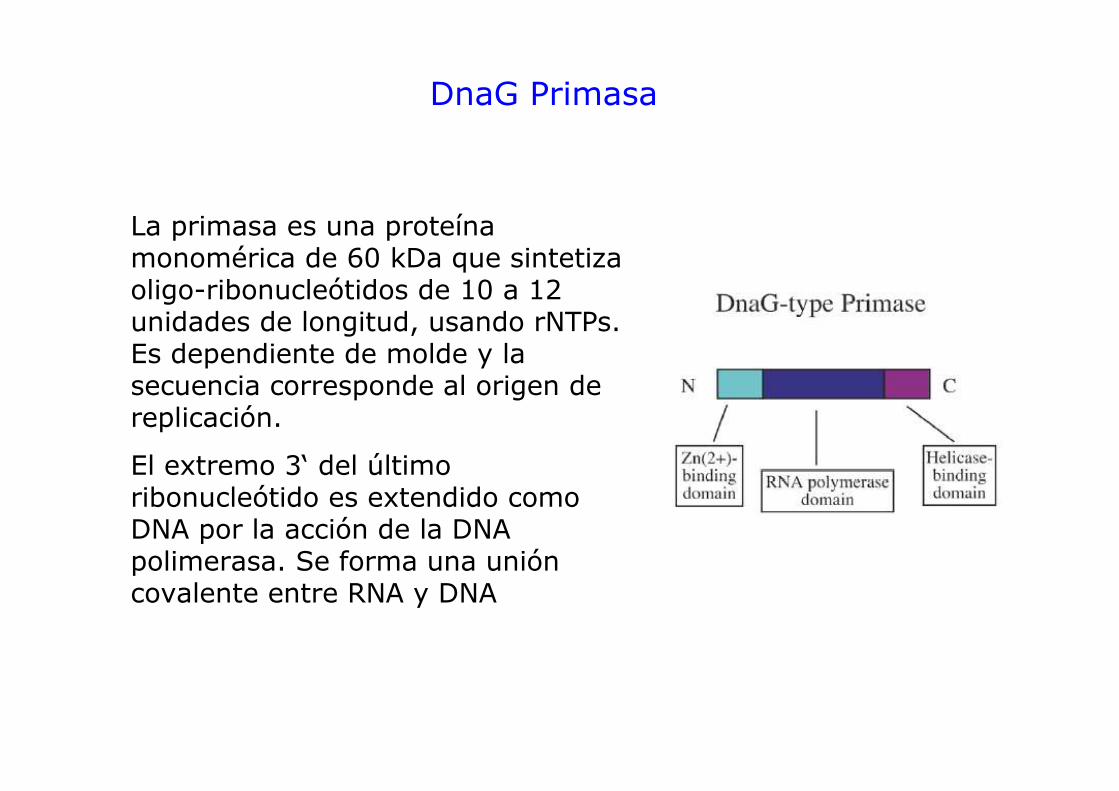
DnaG Primasa
La primasa es una proteína monomérica de 60 kDa que sintetiza oligo-ribonucleótidos de 10 a 12 unidades de longitud, usando rNTPs. Es dependiente de molde y la secuencia corresponde al origen de replicación.
El extremo 3‘ del último ribonucleótido es extendido como DNA por la acción de la DNA polimerasa. Se forma una unión covalente entre RNA y DNA

DNA primasa
RNA polimerasa que sintetiza los cebadores de RNA

La La DNA polimerasa IIIDNA polimerasa III cataliza la reaccicataliza la reaccióón de n de polimerizacipolimerizacióón durante la replicacin durante la replicacióón n
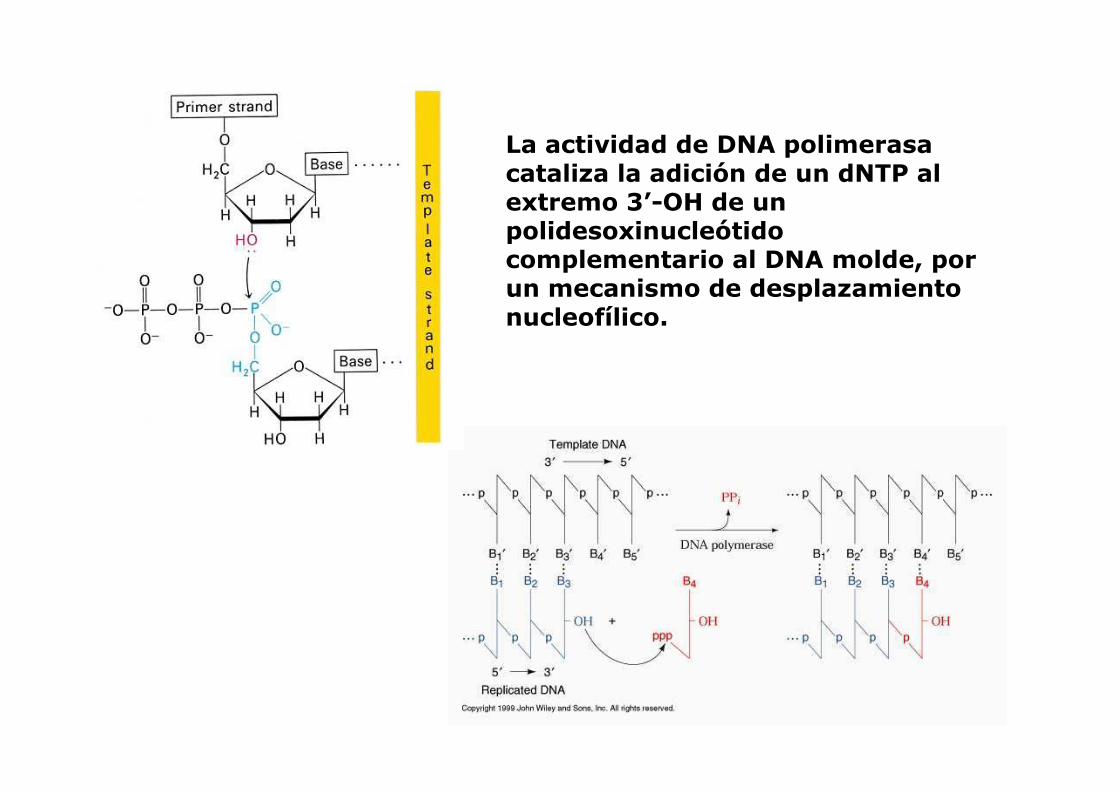
La actividad de DNA polimerasa cataliza la adición de un dNTP al extremo 3’-OH de un polidesoxinucleótidocomplementario al DNA molde, por un mecanismo de desplazamiento nucleofílico.

Con un alto grado de fidelidad gracias a su Con un alto grado de fidelidad gracias a su actividad actividad exonucleasaexonucleasa

La FIDELIDAD se refiere al seguimiento exacto de la secuencia que sirve como molde. En promedio, las DNA pols, 1 error por cada 108 nts
La actividad de actividad de exonucleasaexonucleasa 3’� 5’de la DNA polimerasa contribuye a la fidelidad pues tiene actividad correctora (proofreading).
Hay dos propiedades importantes de las DNA POLIMERASAS:
FIDELIDAD Y PROCESIVIDAD
La PROCESIVIDAD se refiere a la capacidad de una DNA polimerasa de elongar una cadena de DNA por muchos nucleótidos antes de disociarse del complejo que forma con el sustrato.
La DNA polimerasa I tiene una procesividad baja. Es distributiva.
La DNA polimerasa III tiene una procesividad alta.
Competencia cinética entre la actividad de Polimerasa y de Exonucleasa

Actividad de Actividad de exonucleasaexonucleasa 33’’ 55’’

Si la síntesis no fuera de
5´-3´ la Polimerasa no podría efectuar el
proofreading
Mismatch

Sub # por holoenzima
Mr Función
αααα 2 132,000 Actividad polimerasaNúcleo de la polimerasa
εεεε 2 27,000 Exonucleasa 3’→→→→5’
θθθθ 2 10,000Se requiere para la union de DnaB
ττττ 2 71,000Unión estable al molde, dimerización del núcleo
γγγγ 2 52,000
Abrazadera que carga las subunidades ββββ al DNA
δδδδ 1 35,000
δδδδ’ 1 33,000
χχχχ 1 15,000
ψψψψ 1 12,000
ββββ 4 37,000
Pinzas que forman una rueda sobre el DNA y aseguran óptima procesividad
Subunidades de la DNA polimerasa III de Subunidades de la DNA polimerasa III de E. E. colicoli

proteínas ββββ
complejo γγγγ
Pinza rodante
Abrazadera

La DNA pol III es altamente procesiva gracias a las subunidades ββββ

Sub # por holoenzima
Mr Función
αααα 2 132,000 Actividad polimerasaNúcleo de la polimerasa
εεεε 2 27,000 Exonucleasa 3’→→→→5’
θθθθ 2 10,000Se requiere para la union de DnaB
ττττ 2 71,000Unión estable al molde, dimerización del núcleo
γγγγ 2 52,000
Abrazadera que carga las subunidades ββββ al DNA
δδδδ 1 35,000
δδδδ’ 1 33,000
χχχχ 1 15,000
ψψψψ 1 12,000
ββββ 4 37,000
Pinzas que forman una rueda sobre el DNA y aseguran óptima procesividad
Subunidades de la DNA polimerasa III de Subunidades de la DNA polimerasa III de E. E. colicoli

Modelo del dímero

FunciFuncióón en la horquilla de replicacin en la horquilla de replicacióónn

La DNA pol III es la que replica las dos hebras a la vez
Corrige errores con actividad exonucleasa 3’→→→→5’
La replicación de la hebra retrasada se interrumpe cada 1000 nt approx
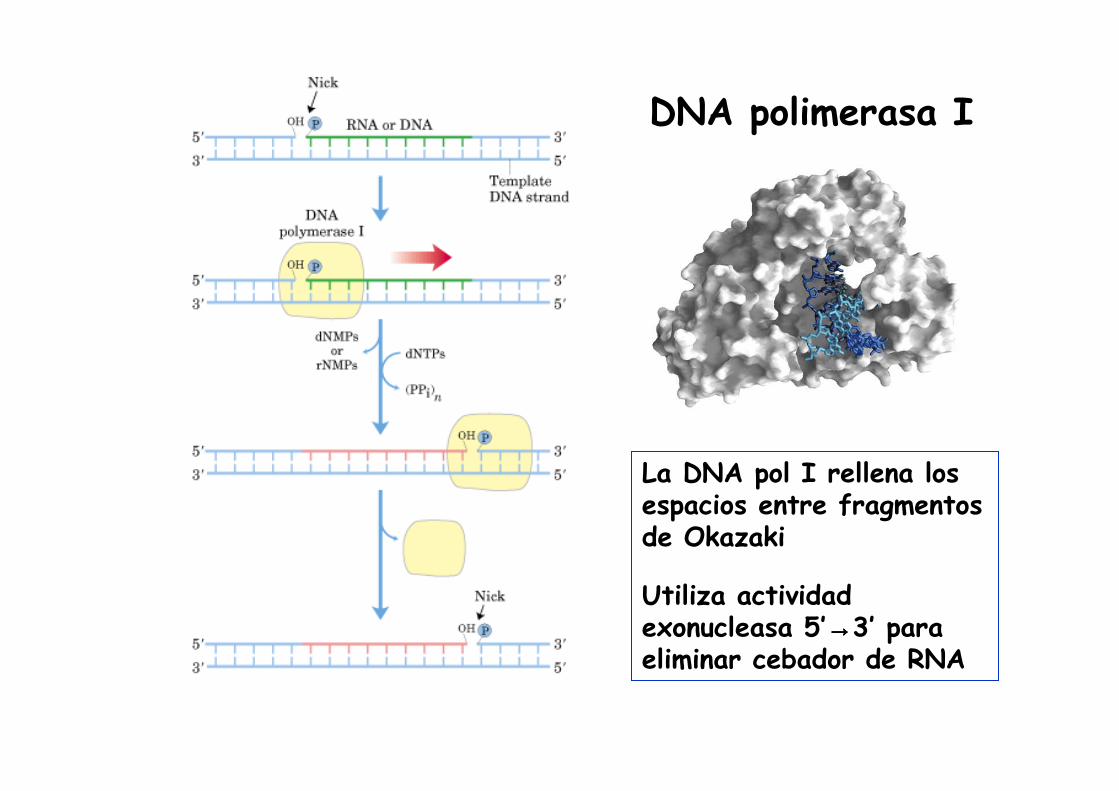
DNA polimerasa I
La DNA pol I rellena los espacios entre fragmentos de Okazaki
Utiliza actividad exonucleasa 5’→→→→3’ para eliminar cebador de RNA

Función de la DNA polimerasa I en la replicación

Función de la DNA ligasa en la replicación
1. Formación del intermediario Enzima ATP
2. Transferencia del adenilo al 5’-P
3. Formación del enlace fosfodiéster
La DNA ligasa de E. coli es una enzima de 75 kDa. Es muy lábil

TERMINACIÓN DE LA REPLICACIÓN EN PROCARIONTES
Las dos horquillas de replicación se aproximan a la misma región que contiene las cajas Ter. Son secuencias de 22 pb, tambien llamados sitios de terminación. Están presentes en tandem (seis) en forma invertida.
A estas secuencias se unen las proteínas TUS (TBP).
La presencia de estas proteínas de unión a DNA causa que se detenga el avance de las horquillas.
Tus: termination utilizationsubstance TBP: Terminationbinding protein. Proteína de 36 kDa que afecta la actividad de la DNA helicasa (DnaB).

TERMINACIÓN. Desenrrollamiento y Síntesis reparativa.
Las dos hebras duplex, productos de la replicación están enrrolladas
La topoisomerasa IV contribuye a la desnaturalización y descatenación de las hebras. Mutantes en el gene de TopoIVexhiben cromosomas que no se han separado totalmente.
Ocurre síntesis reparativa para llenar los huecos

Dna B
Topoisomerasa
SSBFragmentos de
okasaki
DNA pol III
Cadena retrasada
Cadena liderDNA pol III
Helicasa
RNA Primasa
Cebador

Cómo se asegura la célula que su DNA se replica solo una vez?

Las enzimas Dam metilasas regulan el inicio en el origen de replicación

Los cromosomas eucariontes tienen m últiples origenes de replicación

No todos los orígenes de replicaciónse activan al mismo tiempo

REPLICACIÓN DE DNA EN EUCARIONTES
En eucariontes, la replicación comienza en muchos sitios a lo largo de los cromosomas.

Los orígenes de replicación de metazoarios no están definidos por una secuencia específica, como el oriC.
Sino que consisten en:
• Sitios de inicio de alta frecuencia
• Sitios de inicio de baja frecuencia
Los orígenes de replicación se establecen durante la fase G1 del ciclo celular y dependen de mucho parámetros:
Estructura nuclear Estructura de la cromatina
Secuencia de DNA Modificaciones en el DNA
Permite modificar el número y localización de los orígenes de replicación

DNA polimerasas en células eucariotas
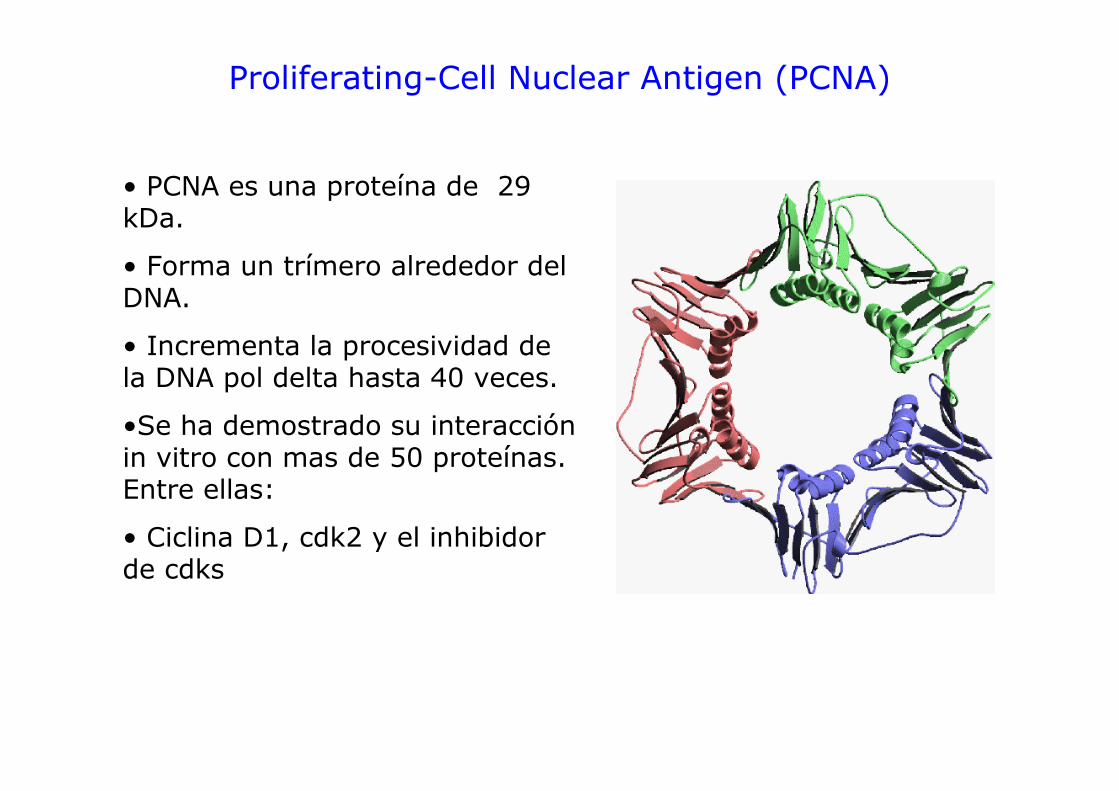
Proliferating-Cell Nuclear Antigen (PCNA)
• PCNA es una proteína de 29 kDa.
• Forma un trímero alrededor del DNA.
• Incrementa la procesividad de la DNA pol delta hasta 40 veces.
•Se ha demostrado su interacción in vitro con mas de 50 proteínas. Entre ellas:
• Ciclina D1, cdk2 y el inhibidor de cdks

Cantidad de DNA, pb/ cél. 3.9 106 109
Velocidad avance horquilla µm/min 30 3
Velocidad de replicación, nt / seg 850 60-90
Número de orígenes de replicación/ célula 1 103-104
Tiempo 1 replicación genómica (hs) 0.27 8
Tiempo 1 división celular (hs) 0.33 24
E. coli Cél humanas
Replicación del ADN:en E. coli vs. en células humanas

Los orígenes de replicación más
estudiados son de levadura

Complejo prereplicativo
Origen de replicaciOrigen de replicaciOrigen de replicaciOrigen de replicacióóóón en eucariotasn en eucariotasn en eucariotasn en eucariotas


TERMINACIÓN EN EUCARIONTES
El dilema de los cromosomas lineales
Durante la terminación en procariontes, hay hidrólisis del cebador pero el extremo 3’ de la cadena funciona para cebar la síntesis que asícompleta la cadena.
Sin embargo, en los cromosomas lineales de eucariontes, después de eliminar al cebador no hay forma de completar la síntesis.
Esto implica que los cromosomas se irían acortando después de cada ronda de duplicación
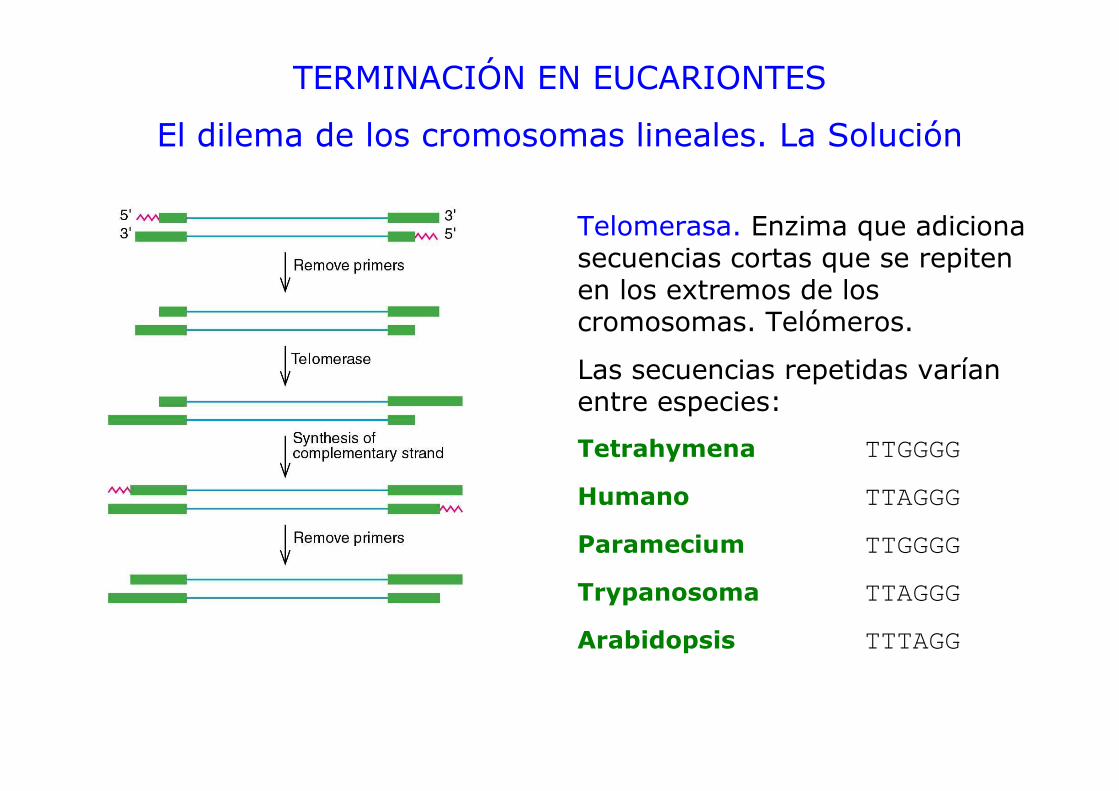
TERMINACIÓN EN EUCARIONTES
El dilema de los cromosomas lineales. La Solución
Telomerasa. Enzima que adiciona secuencias cortas que se repiten en los extremos de los cromosomas. Telómeros.
Las secuencias repetidas varían entre especies:
Tetrahymena TTGGGG
Humano TTAGGG
Paramecium TTGGGG
Trypanosoma TTAGGG
Arabidopsis TTTAGG

La telomerasa es una DNA polimerasa que utiliza RNA como molde
La telomerasa estácompuesta por dos subunidades:
• Subunidad catalítica (proteína)
• Subunidad de RNA asociada
Funciona como molde para elongación de una de las cadenas.

Mecanismo de acción de la telomerasa

Mecanismo de acción de la telomerasa

El resultado:
La telomerasa está presente en células embrionarias, pero en células somáticas su actividad es muy baja.

Topología del DNA
• Tiene implicaciones en los procesos de Replicación, Recombinación y Reparación
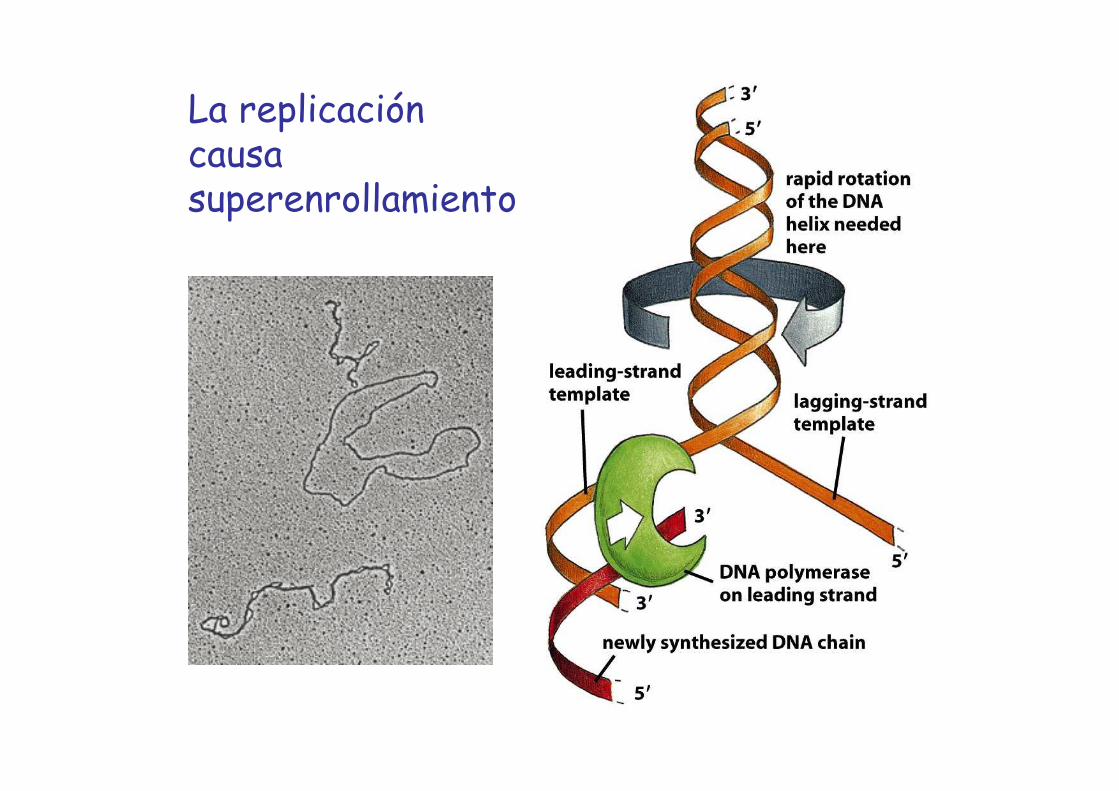
La replicación causa superenrollamiento

La replicación causa superenrollamiento
Región superenrollada
DNA circular
DNA duplexparental relajado
Replicación
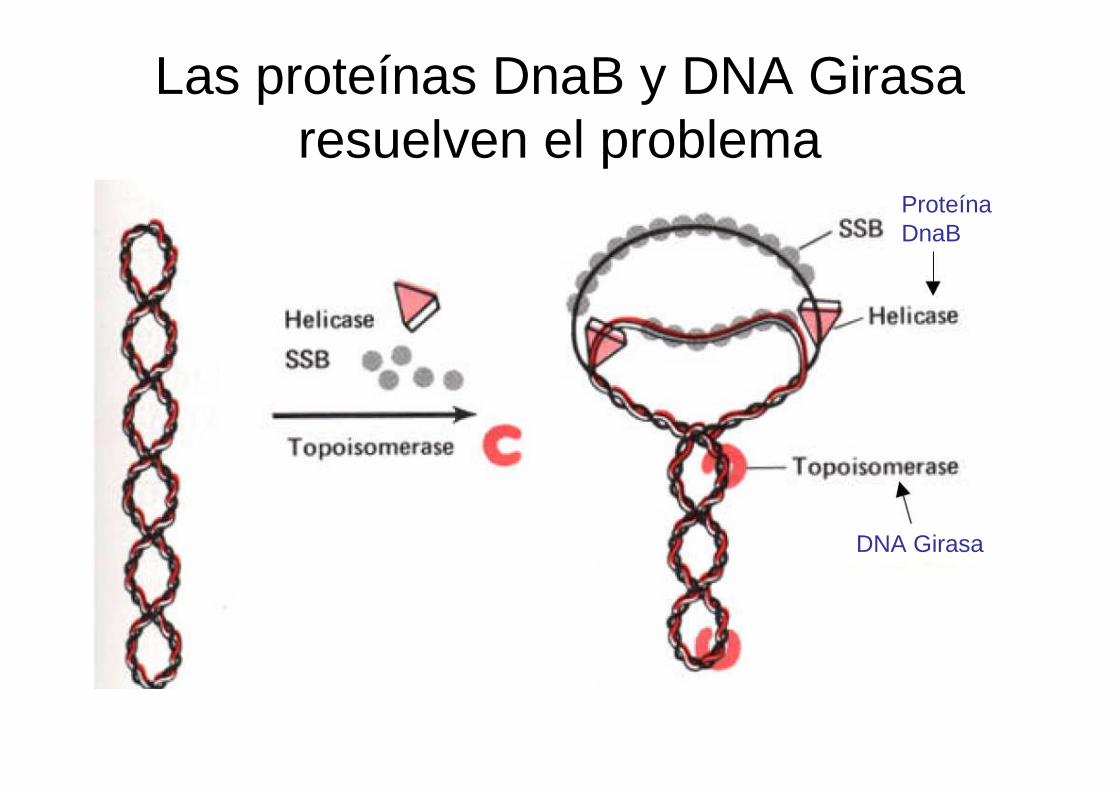
Las proteínas DnaB y DNA Girasaresuelven el problema
DNA Girasa
Proteína DnaB

Las topoisomerasas

Las topoisomerasasTipo I cortan 1 sola hebra

Las topoisomerasas
Tipo II cortan las 2 hebras
Topoisomerasa IVGirasa
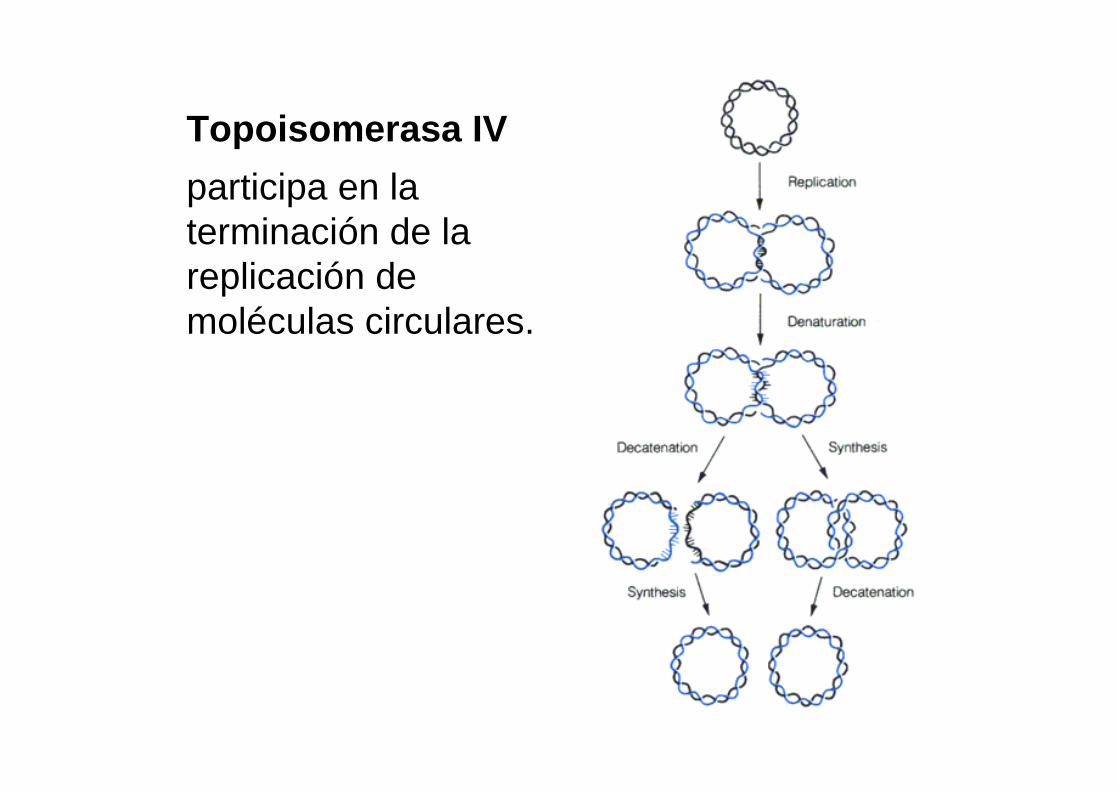
Topoisomerasa IV
participa en la terminación de la replicación de moléculas circulares.

Decatenación

Acción de las Topoisomerasas durante la recombinación
Topoisomerasa I

En Eucariotes, durante la replicación hay modificaciones en los octámeros de histona


Las Histonas parentales se distribuyen
proporcionalmente entre las nuevas cadenas.

El patrón de nucleosomas se hereda a las cadenas hijas







